isolation and characterization of five erwinia amylovora ...rba4, -8, -10, and -e; and rkk2, -3, -4,...
TRANSCRIPT

APPLIED AND ENVIRONMENTAL MICROBIOLOGY,0099-2240/01/$04.0010 DOI: 10.1128/AEM.67.1.59–64.2001
Jan. 2001, p. 59–64 Vol. 67, No. 1
Copyright © 2001, American Society for Microbiology. All Rights Reserved.
Isolation and Characterization of Five Erwinia amylovoraBacteriophages and Assessment of Phage Resistance in Strains
of Erwinia amylovoraELISE L. SCHNABEL AND ALAN L. JONES*
Department of Botany and Plant Pathology, Michigan State University, East Lansing, Michigan 48824-1312
Received 12 May 2000/Accepted 16 October 2000
Phages able to infect the fire blight pathogen Erwinia amylovora were isolated from apple, pear, andraspberry tissues and from soil samples collected at sites displaying fire blight symptoms. Among a collectionof 50 phage isolates, 5 distinct phages, including relatives of the previously described phages fEa1 and fEa7and 3 novel phages named fEa100, fEa125, and fEa116C, were identified based on differences in genome sizeand restriction fragment pattern. fEa1, the phage distributed most widely, had an approximately 46-kbgenome which exhibited some restriction site variability between isolates. Phages fEa100, fEa7, and fEa125each had genomes of approximately 35 kb and could be distinguished by their EcoRI restriction fragmentpatterns. fEa116C contained an approximately 75-kb genome. fEa1, fEa7, fEa100, fEa125, and fEa116Cwere able to infect 39, 36, 16, 20, and 40, respectively, of 40 E. amylovora strains isolated from apple orchardsin Michigan and 8, 12, 10, 10, and 12, respectively, of 12 E. amylovora strains isolated from raspberry fields(Rubus spp.) in Michigan. Only 22 of 52 strains were sensitive to all five phages, and 23 strains exhibitedresistance to more than one phage. fEa116C was more effective than the other phages at lysing E. amylovorastrain Ea110 in liquid culture, reducing the final titer of Ea110 by >95% when added at a ratio of 1 PFU per10 CFU and by 58 to 90% at 1 PFU per 105 CFU.
Fire blight, caused by the bacterium Erwinia amylovora, is adevastating disease of apple and pear trees in North America,Europe, the Mediterranean region, and New Zealand. Mostpear and apple cultivars currently in commercial productionare moderately to highly susceptible to fire blight. The patho-gen is also able to infect a few other members of the Rosaceae,including pyracantha, hawthorn, and cotoneaster. A distinctsubtype of E. amylovora is able to cause fire blight on Rubusspp., especially raspberry and blackberry, but not on apple andpear shoots or seedlings (14, 19). Except for differences ingenetic fingerprints (11, 12), strains isolated from Rubus spp.are indistinguishable from tree fruit E. amylovora strains.
Preventing the buildup of epiphytic populations of E. amy-lovora on nutrient-rich stigmatic surfaces of blossoms in thespring is the main strategy for controlling fire blight (6, 22).Streptomycin is no longer effective for controlling epiphytic E.amylovora on blossoms in many fruit-growing regions becauseof the emergence of streptomycin-resistant strains of thepathogen (7). A new strategy for hindering the establishmentof epiphytic E. amylovora on stigmas is the use of bacteria suchas Pseudomonas fluorescens strain A506 (9), Pantoea agglom-erans strain C9-1 (13, 20), and a few other species (8, 13), butso far the level and consistency of control with microbial agentsare lower than those with antibiotics (6). Phages of E. amylo-vora have been proposed as possible control agents for fireblight. The control potential of an E. amylovora phage was firstdemonstrated by Erskine in 1973 (3). Subsequently, symptomsof fire blight were attenuated in apple seedlings inoculated
with fEa1 in conjunction with E. amylovora (15) and in pearfruit inoculated with E. amylovora in the presence of a fEa1-encoded polysaccharide depolymerase (5). Phages of E. amy-lovora have commonly been found on aerial parts and in soilsassociated with fire blight-infected apple trees (3, 15, 16), butthe association of phage with fire blight-infected Rubus spp.has not been investigated. Except for cloning a polysaccharidedepolymerase gene from fEa1 (5), genetic methods have notbeen used to study E. amylovora-specific phages.
In this study the diversity of E. amylovora phages recoveredfrom soils and shoots of fire blight-infected plants was evalu-ated. Phages were characterized using PCR, restriction frag-ment length polymorphism analysis, and pulsed-field gel elec-trophoresis (PFGE). The phage sensitivity of a panel of E.amylovora strains and the ability of the phage to lyse brothcultures of E. amylovora were examined. The ability to defin-itively identify phages is prerequisite to future epidemiologicaland control studies of fire blight using phage.
MATERIALS AND METHODS
Bacterial strains, phages, and media. E. amylovora strain Ea110, isolated in1975 as strain MSU110 from a canker on Jonathan apple in an experimentalorchard near East Lansing, Mich. (15), and phage fEa1, isolated in 1975 asphage PEa1 from blighted Jonathan apple shoots collected near Paw Paw, Mich.(15), were obtained from John S. Hartung, Agricultural Research Service, U.S.Department of Agriculture, Beltsville, Md. (5, 16). Phage fEa7, isolated in 1976as phage PEa7 from blighted apple shoots collected near Berrien Springs, Mich.(15), was obtained from the American Type Culture Collection (ATCC 29780-B2). Phages fEa100, fEa104, fEa125, and fEa116C were isolated in this studyand deposited at the American Type Culture Collection as ATCC 29780-B4,29780-B5, 29780-B3, and 29780-B6, respectively. Forty strains of E. amylovorawere collected from 14 apple orchards located in six counties of Michigan during1997 and 1998. Their identity was confirmed based on their colony color andmorphology on MM2Cu and Luria-Bertani (LB) media (2) and by PCR assaywith the AJ75-AJ76 primer pair (10). E. amylovora strains MR1, -2, -3, and -4;RBA4, -8, -10, and -E; and RKK2, -3, -4, and -5 were collected from three
* Corresponding author. Mailing address: Department of Botanyand Plant Pathology, Michigan State University, East Lansing, MI48824-1312. Phone: (517) 355-4573. Fax: (517) 353-5598. E-mail:[email protected].
59
on June 3, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

raspberry farms in Michigan (11). Other bacteria included seven nonfluorescentand nine fluorescent strains of Pseudomonas spp. and 12 strains of Pantoeaagglomerans previously isolated from Michigan apple orchards (18). Bacteriawere routinely cultured on LB agar and in LB broth. The double-layer agartechnique (1) was used to produce phage plaques on bacterial hosts, using a topagar consisting of 11.5 g of nutrient agar, 5 g of glucose, and 5 g of yeast extractper liter on a bottom agar of 23 g of nutrient agar and 5 g of glucose per liter.
Isolation of phage. Samples were collected from an asymptomatic apple or-chard at Michigan State University, several fire blight-infected commercial appleand pear orchards in southern Michigan, a fire blight-infected apple orchard innorthern California, and a raspberry farm in northern Michigan in 1996 and1997. Aerial tissues (i.e., branches, leaves, and fruit) and/or soil from within thetree dripline were mixed with sterile water. The collected washes were treatedwith chloroform at a final concentration of 5%. Aliquots (100 ml) were mixedwith 100 ml of Ea110 culture at an optical density at 600 nm of 1.0 to 3.0, plated,and assessed for the presence of plaques following 18 h of incubation. Phagewere recovered from individual plaques by soaking isolated agar plugs in 1 ml of100 mM NaCl–50 mM Tris-HCl(pH 7.5)–8 mM MgSO4 for at least 1 h. Phagewere purified using successive rounds of single-plaque isolation.
Isolation of phage DNA. Ea110 was grown overnight in LB broth with agitationat 28°C and diluted to an optical density at 600 nm of 0.14 (approximately 2 3108 CFU/ml). Bacteriophage (2 3 107 PFU) were mixed with 1 ml of dilutedbacteria. Following a 10-min incubation, 9 ml of LB broth was added and thecultures were grown overnight at 28°C with agitation. Chloroform (30 ml) wasadded, and bacterial debris was removed by centrifugation at 4,000 3 g for 15min. DNA was purified either by using the Wizard Lambda DNA Preps DNAPurification System (Promega, Madison, Wis.) or by phenol extraction of con-centrated phage. In the latter case, the cleared bacterial lysate was incubated for30 min at 37°C following addition of 40 ml of nuclease mix (0.25 mg each ofDNase and RNase per ml in 150 mM NaCl–50% [wt/vol] glycerol). Four milli-liters of 33% polyethylene glycol 8000–3.3 M NaCl was then added, and themixture was incubated on ice for at least 30 min followed by centrifugation in15-ml Corex tubes (Corning, Corning, N.Y.) at 10,000 3 g for 10 min. The pelletwas resuspended in 400 ml of 150 mM NaCl–40 mM Tris-HCl (pH 7.4)–10 mMMgSO4 and clarified by centrifugation in a microcentrifuge for 2 min. Thesupernatant fluid was extracted twice with chloroform. Phage DNA was thenreleased by gentle mixing in an equal volume of Tris-buffered phenol (pH 7.9) for5 min. After centrifugation for 5 min, the upper layer was extracted again withphenol and then with chloroform. DNA was precipitated by the addition of 1 mlof 95% ethanol and 50 ml of 3 M sodium acetate (pH 5.2), collected by centrif-ugation for 15 min, rinsed with 300 ml of 70% ethanol, allowed to dry, and gentlyresuspended in 50 ml of 10 mM Tris-HCl (pH 8.0)–0.1 mM EDTA.
fEa1 PCR assay. BglII restriction fragments of fEa1 were ligated intoBamHI-digested, dephosphorylated pGEM3zf(1); ligation products were intro-duced into Escherichia coli strain JM109 by electroporation, and selected trans-formants were analyzed. A clone containing a 1.8-kb BglII fragment was partiallysequenced (GenBank accession no. AF222715). Based on this sequence, PCRprimers were designed to amplify a 304-bp fragment from fEa1 (PEa1A 59-AATGGGCACCGTAAGCAGT and PEa1B 59-TAATGGGTATGATAGAAGGCAGAC). PCR reaction mixtures (20 ml) consisted of 20 mM Tris-HCl (pH 8.4),50 mM KCl, 1.5 mM MgCl2, 0.5 mM (each) primer, 0.16 mM deoxynucleosidetriphosphates (Gibco BRL, Grand Island, N.Y.), 0.5 U of Taq polymerase(Gibco BRL) and 1 ml of phage lysate containing 104 to 107 PFU of phage.Reactions were performed in a PTC-150 minicycler (MJ Research, Watertown,Mass.) with cycling parameters of 95°C for 2 min followed by 35 cycles of 95°Cfor 30 s, 53°C for 30 s, and 72°C for 30 s. Reaction products were analyzed on1.5% (wt/vol) agarose gels in 0.53 Tris-borate-EDTA (TBE) buffer run at 10V/cm, followed by ethidium bromide staining.
Restriction analysis of phage DNA. Purified phage DNA was subjected torestriction analysis according to the manufacturers’ protocols (Gibco BRL andRoche Molecular Biochemicals, Indianapolis, Ind.). The samples were electro-phoresed either through 0.8% agarose in 0.53 TBE buffer at 6 V/cm for 1 to 2 hor through 0.4% agarose in 0.53 TBE buffer at 1 V/cm for 18 to 24 h. The sizesof the fragments were estimated by comparison to HindIII-digested l DNA,high-molecular-weight DNA standards, or the 1-kb DNA Plus ladder (GibcoBRL). Restriction maps were constructed by compiling the data from single- anddouble-enzyme digestions of intact and cloned BglII fragments of phage DNA.
Contour-clamped homogeneous electric field gel analysis. Pulsed-field gelswere run using a CHEF-DR II PFGE system (Bio-Rad Laboratories, Hercules,Calif.). The 1% gels were made with Seakem (Rockland, Maine) Gold agarosein 0.53 TBE buffer. The pulsed-field parameters were as follows: 0.53 TBErunning buffer, 0.1-s initial switch time, 10-s final switch time, 6.0 V/cm, 15-h run
time, and 14°C buffer temperature. The gels were stained with ethidium bromideand visualized using a Foto/Eclipse system (Fotodyne, Inc., Harland, Ws.).
Infection and lysis experiments. Aliquots (50 ml) of overnight cultures ofvarious strains were mixed with 103 PFU of phage (as determined by plaqueformation using strain Ea110 as a standard), incubated for 10 min, mixed with 3ml of top agar, and plated onto bottom agar. Following incubation at 22°C for 18to 42 h, the plates were evaluated for the presence of plaques. Strain and phagecombinations which yielded no plaques were retested at least twice.
Overnight cultures of Ea110 were diluted to an optical density at 600 nm of0.007 (approximately 107 CFU/ml), and 1-ml aliquots were infected with 1.0 3102, 1.0 3 104, and 1.0 3 106 PFU of a single phage; 0.5 3 102, 0.5 3 104, and0.5 3 106 PFU of each of two phages; and 0.33 3 102, 0.33 3 104, and 0.33 3 106
PFU of each of three phages. Three 1-ml aliquots were grown without phage asa control. The cultures were incubated overnight at 28°C with agitation. Bacterialdensities were assessed by measuring the optical density of the cultures at 600nm. Percent growth was calculated for each experiment by dividing the opticaldensities by the average optical density of the cultures grown without phage. Thepercent growth values for three experiments were pooled and analyzed by anal-ysis of variance.
RESULTS
Identification of phage types. The collection of phages thatformed plaques on the E. amylovora Ea110 host consisted ofreference phages fEa1 and fEa7 and 48 new isolates collectedin 1996 and 1997. Forty-four of the new isolates were fromMichigan, including 41 from soil and plant material of fireblight-infected apple orchards, 2 from blighted raspberrycanes, and 1 from a blighted pear shoot. Four additional iso-lates came from soil collected in a fire blight-infected appleorchard in California.
The phage isolates were characterized by plaque morphol-ogy and tested for the presence of a fEa1 sequence by PCR.Forty-two isolates, including fEa1 and isolates from Michiganand California tree fruit orchards, produced large plaques sur-rounded by an expanding translucent halo similar to thosepreviously described for fEa1 and yielded a 0.3-kb PCR frag-ment when tested with fEa1 primers (data not shown). Theseisolates were categorized as presumptive fEa1 isolates.
The remaining eight isolates generated small plaques andproduced no PCR product with the fEa1 primers. EcoRIrestriction analysis of DNAs from these isolates yielded fourpatterns distinct from the EcoRI profile of fEa1 representedby fEa7, soil sample isolates from Michigan (fEa100) andCalifornia (fEa125), and a Michigan apple orchard isolate(fEa116C) (Fig. 1A). The two isolates from raspberry and twoadditional isolates from Michigan apple orchards showed pat-terns identical to that of fEa7 (data not shown). DNAs fromisolates representing the five EcoRI restriction patterns wereanalyzed by PFGE (Fig. 1B). The genome sizes of these phageswere estimated to be 46 kb (fEa1), 35 kb (fEa7), 35 kb(fEa100), 35 kb (fEa125), and 75 kb (fEa116C).
For fEa1, fEa125, and fEa116C the sums of the lengths ofthe observed EcoRI restriction fragments were in agreementwith the genome size estimates from the PFGE analysis. Incontrast, the sums of the lengths of the EcoRI restrictionfragments of fEa7 and fEa100 greatly exceeded the genomesizes observed by PFGE, suggesting that the restriction diges-tions may have been incomplete. To test whether the presenceof impurities was causing partial digestion, the DNA sampleswere subjected to additional phenol and chloroform extrac-tions and ethanol precipitation followed by restriction diges-tion with high levels of EcoRI for 16 h. The resulting digestion
60 SCHNABEL AND JONES APPL. ENVIRON. MICROBIOL.
on June 3, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from
patterns were identical to those originally observed (data notshown), suggesting that impurities were not responsible for theobserved patterns.
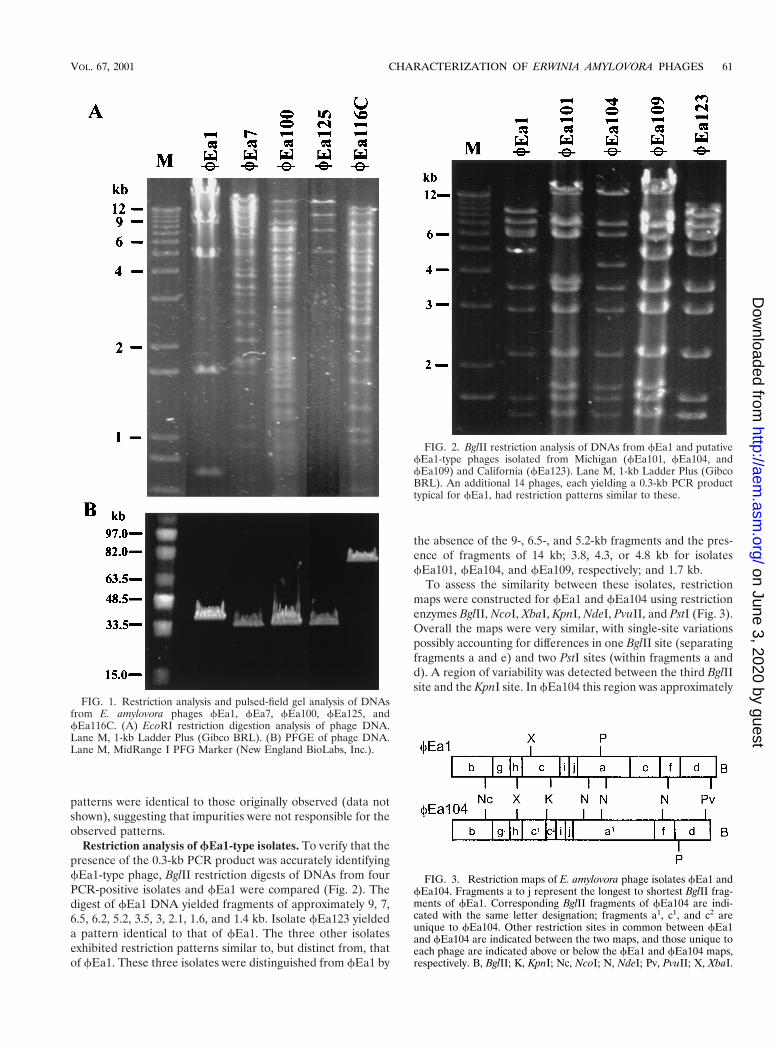
Restriction analysis of fEa1-type isolates. To verify that thepresence of the 0.3-kb PCR product was accurately identifyingfEa1-type phage, BglII restriction digests of DNAs from fourPCR-positive isolates and fEa1 were compared (Fig. 2). Thedigest of fEa1 DNA yielded fragments of approximately 9, 7,6.5, 6.2, 5.2, 3.5, 3, 2.1, 1.6, and 1.4 kb. Isolate fEa123 yieldeda pattern identical to that of fEa1. The three other isolatesexhibited restriction patterns similar to, but distinct from, thatof fEa1. These three isolates were distinguished from fEa1 by
the absence of the 9-, 6.5-, and 5.2-kb fragments and the pres-ence of fragments of 14 kb; 3.8, 4.3, or 4.8 kb for isolatesfEa101, fEa104, and fEa109, respectively; and 1.7 kb.
To assess the similarity between these isolates, restrictionmaps were constructed for fEa1 and fEa104 using restrictionenzymes BglII, NcoI, XbaI, KpnI, NdeI, PvuII, and PstI (Fig. 3).Overall the maps were very similar, with single-site variationspossibly accounting for differences in one BglII site (separatingfragments a and e) and two PstI sites (within fragments a andd). A region of variability was detected between the third BglIIsite and the KpnI site. In fEa104 this region was approximately
FIG. 1. Restriction analysis and pulsed-field gel analysis of DNAsfrom E. amylovora phages fEa1, fEa7, fEa100, fEa125, andfEa116C. (A) EcoRI restriction digestion analysis of phage DNA.Lane M, 1-kb Ladder Plus (Gibco BRL). (B) PFGE of phage DNA.Lane M, MidRange I PFG Marker (New England BioLabs, Inc.).
FIG. 2. BglII restriction analysis of DNAs from fEa1 and putativefEa1-type phages isolated from Michigan (fEa101, fEa104, andfEa109) and California (fEa123). Lane M, 1-kb Ladder Plus (GibcoBRL). An additional 14 phages, each yielding a 0.3-kb PCR producttypical for fEa1, had restriction patterns similar to these.
FIG. 3. Restriction maps of E. amylovora phage isolates fEa1 andfEa104. Fragments a to j represent the longest to shortest BglII frag-ments of fEa1. Corresponding BglII fragments of fEa104 are indi-cated with the same letter designation; fragments a1, c1, and c2 areunique to fEa104. Other restriction sites in common between fEa1and fEa104 are indicated between the two maps, and those unique toeach phage are indicated above or below the fEa1 and fEa104 maps,respectively. B, BglII; K, KpnI; Nc, NcoI; N, NdeI; Pv, PvuII; X, XbaI.
VOL. 67, 2001 CHARACTERIZATION OF ERWINIA AMYLOVORA PHAGES 61
on June 3, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

0.5 kb shorter than in fEa1, it lacked the second XbaI sitefound in fEa1, and it contained a BglII site not found in fEa1,suggesting that an alternate sequence may be present in thisregion.
An additional 13 PCR-positive isolates from Michigan ana-lyzed by BglII digestion yielded patterns similar to those offEa101, fEa104, and fEa109, and one additional isolate fromCalifornia yielded a BglII pattern identical to that of fEa1(data not show). These data support the grouping of all PCR-positive isolates as fEa1-type phages.
Host range of E. amylovora phages. To assess the prevalenceof phage resistance in natural populations of E. amylovora, theability of phages fEa1, fEa7, fEa100, fEa125, and fEa116Cto infect 40 strains of E. amylovora isolated from apple wastested in a plaque formation assay (Table 1). Resistance to atleast one phage was detected in 65% of the strains. Resistanceto fEa100 and fEa125 was found in 50 and 60% of the strains,respectively; resistance to both of these phages was found in45% of the strains. Resistance to the other three phages wasless common. fEa116C formed plaques on all 40 strains, asingle strain was resistant to fEa1, and four strains were re-sistant to fEa7. Ninety percent of the strains were sensitive tophages fEa1, fEa7, and fEa116C.
Strains of E. amylovora which are capable of causing diseaseon Rubus spp. are genetically distinct from strains which causedisease on other rosaceous plants such as apple and pear (11,12). To assess differences in phage sensitivity between the twodistinct types of E. amylovora strains, 12 strains of E. amylovoraisolated from Rubus hosts were also tested for their sensitivityto the phages. Eight strains were sensitive to all five phages.Four strains (all RKK strains) were resistant to fEa1; two ofthese strains (RKK2 and RKK5) were also resistant to fEa100and fEa125.
fEa1, fEa7, and fEa116C were screened for ability toinfect other common orchard bacterial isolates. A collection ofnonfluorescent (7 strains) and fluorescent (9 strains) Pseudo-monas strains and Pantoea agglomerans (12 strains) were chal-
lenged with 103 PFU of each phage. No plaques formed on anyof the strains.
Lysis of E. amylovora strain Ea110 in liquid culture. Theability of the phages to control populations of E. amylovora wastested by infecting liquid cultures of E. amylovora with individ-ual phages or with combinations of phages fEa1, fEa7, andfEa116C (Fig. 4). Data for the seven phage treatments andthree phage concentrations were analyzed by two-way analysisof variance. F values for treatments and phage concentrationswere highly significant (P 5 0.00001), and interaction effectswere not significant. When fEa1, fEa7, fEa100, fEa125, andfEa116C were added individually to cultures of E. amylovorastrain Ea110 at 1 PFU per 10 CFU or 1 PFU per 103 CFU, onlyfEa116C was able to effectively control bacterial populations,reducing optical densities to 96% 6 4% below that of bacteriagrown without phage. A mixture of phages fEa1 and fEa7 wasnot significantly better than either phage alone in controllingbacterial populations. A mixture of phages fEa116C, fEa1,and fEa7 was as effective as fEa116C alone.
DISCUSSION
Five distinct E. amylovora phages were identified from acollection of 50 phage isolates. By cloning and sequencing a1.8-kb BglII fragment of fEa1, a set of PCR primers specificfor detection of fEa1-type phages was developed and used toscreen the collection; others have also found these primersuseful for rapidly differentiating fEa1 from other phages (4).Phages fEa7, fEa100, fEa125, and fEa116C, which were
FIG. 4. Control of growth of E. amylovora strain Ea110 in thepresence of phage(s). Single phage types (fEa1, fEa7, fEa100,fEa125, or fEa116C) or combinations of equal proportions of two orthree phages (fEa1 plus fEa7 or fEa1 plus fEa7 plus fEa116C)were added at 106) (open bars), 104 (hatched bars), and 102 (solid bars)total PFU to 1-ml aliquots of LB broth containing 107 CFU of E.amylovora strain Ea110. The optical densities of the cultures following18 h of growth at 28°C were compared to the optical densities ofcultures grown without phage. The data shown are the averages ofthree replications, with the standard deviation of each mean shown.
TABLE 1. Sensitivity of each of 52 strains of E. amylovorato phages
No. of E. amylovora strainsfrom the following hosta: Plaques formed with the following phageb:
Malus spp.(n 5 40 strains)
Rubus spp.(n 5 12strains)
fEa1 fEa7 fEa100 fEa125 fEa116C
17 0 1 1 2 2 114 8 1 1 1 1 13 0 1 1 2 1 12 0 1 1 1 2 12 0 1 2 2 1 11 0 1 2 2 2 11 0 2 2 2 1 10 2 2 1 1 1 10 2 2 1 2 2 1
a Phages were tested on strains of E. amylovora from Malus spp. collected in sixMichigan counties in 1998 (2 to 14 per county) and on strains from Rubus spp.collected at three sites in Michigan in 1994. Each strain was mixed with 103 PFUof each phage and plated using the double-layer agar technique.
b Phage sensitivity plates were evaluated following 18 to 36 h of incubation. 1,plaques present; 2, no plaques. The total numbers of strains infected were asfollows: fEa1, 47; fEa7, 48; fEa100, 26; fEa125, 30; fEa116C, 52.
62 SCHNABEL AND JONES APPL. ENVIRON. MICROBIOL.
on June 3, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

distinct from fEa1, were differentiated based on genome sizeand restriction fragment pattern. The analysis of these phagesat the DNA level provides a basis for the characterization offuture phage isolates.
Phages specific for E. amylovora were prevalent in Michiganorchard sites with active fire blight infections, and the detec-tion of phages during the later stages of fire blight epidemics inapple orchards is consistent with the results of an earlier Mich-igan study (15). Nearly every site surveyed yielded fEa1-typeisolates, indicating that this phage is commonly associated withE. amylovora in nature. Two major subtypes of fEa1-typephages were observed among the isolates based on restrictionanalysis. The first group included the original fEa1 isolate andthe fEa1-type isolates from California; all of these isolates hadidentical EcoRI restriction patterns. The second group in-cluded the more recent isolates from Michigan, which pro-duced somewhat variable EcoRI restriction patterns whichwere quite distinct from that of the original fEa1 isolate.
For the other four phages, our collection contained eithersingle isolates (fEa100, fEa125, and fEa116C) or five isolates(fEa7). The five isolates of fEa7 displayed no obvious differ-ences in restriction pattern even though two of the isolateswere associated with E. amylovora from raspberry canes, whichrepresent a genetically distinct and often spatially isolatedhost. It is not clear if the paucity of these phages in ourcollection was due to their scarcity in the collected samples orto difficulties in identifying the small plaques formed by thesephages. Because our discovery of phages distinct from fEa1was somewhat fortuitous, it may be reasonable to suppose thatadditional E. amylovora phage types exist.
One explanation for the insensitivity of some strains of E.amylovora to phages may be that they harbor temperate phagesthat render them resistant to other lytic phages. However, thisappears unlikely, because Ritchie (15) tested strains of E. amy-lovora resistant to fEa1 or fEa7 for lysogeny using UV lightand mitomycin C as induction agents; all tests were negativefor temperate phage. Another indication of the presence oftemperate phages is the formation of hazy plaques; none of thephages were observed to form plaques that became hazy in thecenter with age. As reported previously (15, 16), fEa1-typephages produced plaques with a distinct halo after about 18 h.This halo was shown to be the result of the production of apolysaccharide depolymerase, capable of hydrolyzing the cap-sular polysaccharide and not of lysogeny (5, 16).
Ritchie and Klos (15, 16) reported that fEa1 and fEa7infected each of 20 E. amylovora isolates; however, our datafrom additional strains indicate that some strains differ in theirphage sensitivity. All 52 strains of E. amylovora from Malus andRubus hosts were infected by phage fEa116C. Although amore exhaustive screen might yield strains with resistance tofEa116C, it appears that resistance to this phage is rare.Among Malus strains of E. amylovora, 40 to 98% were infectedby the other four phages. Apparently, despite the high degreeof homogeneity among tree fruit strains of E. amylovora, somestrains possess differences that prevent adherence, uptake, orreplication by certain phages. The specific nature of the mech-anisms responsible for phage resistance in the E. amylovorastrains is unknown. The majority of the E. amylovora strainsfrom Rubus were sensitive to all five phages, although resis-tance to fEa1, fEa100, and fEa125 was detected. This is the
first study to show that Rubus and Malus strains of E. amylo-vora were infected by the same phages.
Despite the ability of each of the phages to infect E. amy-lovora strain Ea110, only fEa116C was able to drastically re-duce the final density of the bacteria grown in liquid culture. Inthe case of fEa1, it has been shown that as phage titers in-crease, a phage-encoded polysaccharide depolymerase, whichis able to degrade the capsule of E. amylovora, accumulates inthe medium. When levels of the enzyme are high enough,fEa1 can no longer infect the bacteria, presumably due todestruction of fEa1 binding sites (5, 15). For fEa7, fEa100,and fEa125, the reasons for ineffective growth control areunknown.
The phages isolated in this study may be useful for biocon-trol of E. amylovora, in particular fEa116C and, to a lesserextent, fEa1 and fEa7, because of the infrequent occurrenceof resistant strains in E. amylovora populations. They might beeffective either in orchards or for eliminating E. amylovorafrom the surface of contaminated budwood and fresh fruit.Control of the blossom-infection stage of fire blight is criticalto the overall control of fire blight (6). E. amylovora multiplieson the stigmatic surfaces of blossoms prior to blossom infec-tion (22). If E. amylovora-specific phages were present on theblossoms, they might suppress the growth of E. amylovora onthe stigmatic surfaces. However, natural phage populations arebelow detectable levels during the bloom period (reference 16and this study). Therefore, any control strategy based on phagewould require that blossoms be treated with phage in much thesame way as blossoms are treated with antagonistic bacteria.Prior colonization of the blossom by a suitable host, such as anavirulent strain of E. amylovora (21), has been shown to beimportant for the establishment and maintenance of phagepopulations (17). Without such a host the population of phagerapidly declined, presumably due to UV light or desiccationeffects.
ACKNOWLEDGMENTS
We wish to thank Monica Meyer and Katalin Kasa for their assis-tance with experiments reported here and W. G. D. Fernando for hishelp in the isolation of phages in 1997.
This research was supported in part by the Rackham EndowmentFund, the Michigan Agricultural Experiment Station, and USDA/CS-REES Agreement 97-34367-3967.
REFERENCES
1. Adams, M. H. 1959. Bacteriophages. Interscience Publishers, New York,N.Y.
2. Bereswill, S., S. Jock, P. Bellemann, and K. Geider. 1998. Identification ofErwinia amylovora by growth morphology on agar containing copper sulfateand by capsule staining with lectin. Plant Dis. 82:158–164.
3. Erskine, J. M. 1973. Characteristics of Erwinia amylovora bacteriophage andits possible role in the epidemiology of fire blight. Can. J. Microbiol. 19:837–845.
4. Gill, J. J., A. M. Svircev, A. L. Myers, and A. J. Castle. 1999. Biocontrol ofErwinia amylovora using bacteriophage. Phytopathology 89:S27.
5. Hartung, J. S., D. W. Fulbright, and E. J. Klos. 1988. Cloning of a bacte-riophage polysaccharide depolymerase gene and its expression in Erwiniaamylovora. Mol. Plant-Microbe Interact. 1:87–93.
6. Johnson, K. B., and V. O. Stockwell. 1998. Management of fire blight: a casestudy in microbial ecology. Annu. Rev. Phytopathol. 36:227–248.
7. Jones, A. L., and E. L. Schnabel. 2000. The development of streptomycinresistant strains of Erwinia amylovora, p. 235–251. In J. L. Vanneste (ed.),Fire blight: the disease and its causative agent Erwinia amylovora. CABInternational, Wallingford, Oxon, United Kingdom.
8. Kearns, L. P., and C. N. Hale. 1996. Partial characterization of an inhibitory
VOL. 67, 2001 CHARACTERIZATION OF ERWINIA AMYLOVORA PHAGES 63
on June 3, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

strain of Erwinia herbicola with potential as a biocontrol agent for Erwiniaamylovora, the fire blight pathogen. J. Appl. Bacteriol. 81:369–374.
9. Lindow, S. E., G. McGourty, and R. Elkins. 1996. Interactions of antibioticswith Pseudomonas fluorescens A506 in the control of fire blight and frostinjury of pear. Phytopathology 86:841–848.
10. McManus, P. S., and A. L. Jones. 1995. Detection of Erwinia amylovora bynested PCR and PCR-dot-blot and reverse-blot hybridizations. Phytopathol-ogy 85:618–623.
11. McManus, P. S., and A. L. Jones. 1995. Genetic fingerprinting of Erwiniaamylovora strains isolated from tree-fruit crops and Rubus spp. Phytopathol-ogy 85:1547–1553.
12. Momol, M. T., E. A. Momol, W. F. Lamboy, J. L. Norelli, S. V. Beer, and H. S.Aldwinckle. 1997. Characterization of Erwinia amylovora strains using ran-dom amplified polymorphic DNA fragments (RAPDs). J. Appl. Microbiol.82:389–398.
13. Pusey, P. L. 1997. Crab apple blossoms as a model for research on biologicalcontrol of fire blight. Phytopathology 87:1096–1102.
14. Ries, S. M., and A. G. Otterbacher. 1977. Occurence of fire blight on thorn-less blackberry in Illinois. Plant Dis. Rep. 61:232–235.
15. Ritchie, D. 1978. Bacteriophages of Erwinia amylovora: their isolation, dis-tribution, characterization, and possible involvement in the etiology and
epidemiology of fire blight. Ph. D. thesis. Michigan State University, EastLansing.
16. Ritchie, D. F., and E. J. Klos. 1977. Isolation of Erwinia amylovora bacte-riophage from the aerial parts of apple trees. Phytopathology 67:101–104.
17. Schnabel, E. L., W. G. D. Fernando, L. E. Jackson, M. P. Meyer, and A. L.Jones. 1998. Bacteriophage of Erwinia amylovora and their potential forbiocontrol. Acta Hortic. 489:649–654.
18. Schnabel, E. L., and A. L. Jones. 1999. Distribution of tetracycline resistancegenes and transposons among phylloplane bacteria in Michigan apple or-chards. Appl. Environ. Microbiol. 65:4898–4907.
19. Starr, M. P., C. Cardona, and D. Folsom. 1951. Bacterial fire blight ofraspberry. Phytopathology 41:915.
20. Stockwell, V. O., K. B. Johnson, and J. E. Loper. 1998. Establishment ofbacterial antagonists of Erwinia amylovora on pear and apple blossoms asinfluenced by inoculum preparation. Phytopathology 88:506–513.
21. Tharaud, M., J. Laurent, M. Faize, and J.-P. Paulin. 1997. Fire blightprotection with avirulent mutants of Erwinia amylovora. Microbiology 143:625–632.
22. Thomson, S. V. 1986. The role of stigma in fire blight infections. Phytopa-thology 76:476–482.
64 SCHNABEL AND JONES APPL. ENVIRON. MICROBIOL.
on June 3, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from