intracellular electrical potential measurements in...
TRANSCRIPT

J. Cell Sci. 81, 207-221 (1986) 207Printed in Great Britain © The Company of Biologists Limited 1986
INTRACELLULAR ELECTRICAL POTENTIAL
MEASUREMENTS IN DROSOPHILA FOLLICLES
JOHANNES BOHRMANN1, ERWIN HUEBNER2, KLAUS SANDER1
AND HERWIG GUTZEIT1'*lBiologisches Institut I (Zoologie), Universitdt Freiburg, Albertstr. 21a, D-7800 Freiburg,West Germanydepartment ofZoology, University of Manitoba, Winnipeg, Canada R3T2N2
SUMMARYWe measured the intracellular electrical potential in oocyte and nurse cells of Drosophila follicles
at different developmental stages (6-14) and determined the intrafollicular potential difference.During stages 8—10B, when intrafollicular transport is known to occur, no significant potentialdifference was found. During late vitellogenic stages the nurse cells assume a more positivepotential than the oocyte. This result contrasts with the published data on Hyalophora follicles, inwhich intercellular electrophoresis of negatively charged proteins occurs from nurse cells to oocyteas a result of an intrafollicular potential difference (nurse cells more negative than the oocyte). Sucha potential difference was not observed in Drosophila follicles at any stage, not even afterapplication of juvenile hormone. The extrafollicular electrical field is described with a dipolemodel. The hypothetical dipole is located in the long axis of the follicle and changes its calculatedlength stage-specifically.
INTRODUCTION
In meroistic insect ovaries the nurse cells are intensely engaged in biosynthesis andcontribute much to the rapid growth of the oocyte, whose nucleus is generallyinactive, or nearly so (Bier, 1963; Telfer, 1975). The molecules synthesized in thenurse cells reach the oocyte via cytoplasmic bridges through which the cytoplasmiccontinuity of these germ-line sister cells is maintained. Because of the syntheticdifferences between nurse cells and oocyte in polytrophic follicles (like Drosophila)there is apparently one-way traffic of material from the nurse cells to the oocyte.Despite the apparent simplicity of the system the mechanism of molecular transportis still not satisfactorily explained.
Recently, several studies on the electrophysiological properties of meroisticovarioles showed that electrical currents traverse the follicles in a number of analysedspecies (Jaffe & Woodruff, 1979; Dittmann, Ehni & Engels, 1981; Huebner, 1984;Sigurdson, 1984; Overall & Jaffe, 1985; Bohrmann et al. 1984; Bohrmann, Dorn,Sander & Gutzeit, 1986). A functional role for these currents was suggested by theintriguing observation that in Hyalophora follicles (polytrophic type) and inRhodnius and Oncopeltus ovarioles (telotrophic type) microinjected fluorescentlylabelled proteins migrate according to their electrical charge by way of 'intercellular
• Author to whom reprint requests should be addressed.
Key words: intracellular electrical potential, intercellular electrophoresis, Drosophila oogenesis.

208 J. Bohrmann, E. Huebner, K. Sander and H. Gutzeit
1A vfl B"** Ooc Ooc&NC
10-
-20-
-30
4 min
Fig. 1. A. Photograph of a Drosophila follicle stage 10B impaled with a microelectrodeeach in the oocyte (Ooc) and in a nurse cell (NC). The follicle is held in place by slightlypressing it against the edge of a Scotch tape (arrow) with the rounded tip of a glass rod(GR). FC, follicle cell epithelium covering the oocyte. Bar, 100 ftfn. B. Two-channelrecording from an X/Y chart recorder of the potentials in the oocyte and a nurse cell of astage 10B follicle. The two arrows indicate the moments at which the electrode in theoocyte (1) and the electrode in the nurse cell (2) were withdrawn. There is no potentialdifference between oocyte and nurse cell.
electrophoresis' (Woodruff & Telfer, 1973, 1980; Telfer, Woodruff & Huebner,1981; Woodruff & Anderson, 1984). This concept has been widely accepted andgeneralized, although it has not yet been shown that normal constituents of the nursecell cytoplasm (as opposed to large quantities of microinjected heterologous mol-ecules) are also subjected to intercellular electrophoresis.
Since intracellular electrical potential measurements have been carried out in onlythree species {Hyalophora: Woodruff & Telfer, 1973; Rhodnius: Telfer et al. 1981;Oncopeltus: Woodruff & Anderson, 1984), we studied Drosophila follicles with thesame methods to see if the electrical properties are compatible with the notion ofintercellular electrophoresis in this species.
MATERIALS AND METHODS
Intracellular electrical potential measurementsFollicles of Drosophila melanogaster (strain Oregon R) were carefully isolated in Robb's medium
(R-14), in which they are able to develop from stage 10 up to stage 14 (Petri, Mindrinos, Lombard& Margaritis, 1979; Bohrmann, 1981), or in Robb's balanced saline (DPBS; Robb, 1969). Themeasuring chamber used was a slight modification of the one described by Kiehart (1982).Measuring electrodes and a ground electrode (Ag/AgCl wire) passed through a thin filmof paraffin oil covering the saline-filled chamber. A thin glass rod with a rounded tip usedfor holding the follicles in place was also inserted horizontally into the chamber (Fig. 1A).

Electrical potential in follicles 209
Intracellular recording pipettes were made using 1 mm Kwik-Fil filament glass capillaries pulled ona David Kopf model 700 D pipette puller. The measurements were carried out with a pair of 3 M-KCl-filled electrodes (5-15 M£2 resistance) using a W. P. Instruments S-7000 A system with twoS-7071 A electrometers and a digital oscilloscope (Nicolet Instruments Corp., model 2090 with the207 module). Measurements were made by a cursor on the screen and representative recordingsmade via the X/Y output using a Hewlett Packard 7015 B X/Y recorder.
We measured the intracellular electrical potentials and the potential differences between theoocyte and one or more different nurse cells (e.g., see Fig. 1B) in 243 wild-type follicles (stages6—14; for stages, see King, 1970). They were cultured in R-14 or in some cases in DPBS, with nodifferences noted in the results. About half of the analysed follicles were cultured for 10—60 min in amedium to which the juvenile hormone (JH) analogue Altosid ZR-515 (a gift from Dr Staal,Zoecon Corp., Palo Alto, CA) was added to a final concentration of 10~7M (see Sigurdson, 1984) or5X 10~5 M (see Giorgi, 1979). Both concentrations gave the same results. The solutions were mixedvigorously before use.
Further measurements (without JH) were carried out on 17 dicephalic (die) follicles (Lohs-Schardin, 1982) with nurse cell groups at either pole of the follicle. Also 12 follicles of the mutantbicaudal D (bicD) were investigated (kindly provided by Dr T. Schupbach, Princeton, NJ); inthis mutant no oocyte differentiates among the 16 germ-line sister cells (cystocytes).
For statistical evaluation, we used the F test (to compare variances) and the t test (to comparemean values) according to Sachs (1978).
Resistance measurements
In 20 wild-type follicles (stages 10A-11) we measured the intercellular resistance between theoocyte and one nurse cell with the voltage-clamp method (see Woodruff & Telfer, 1974). In 15 ofthese follicles we also determined the intercellular resistance as well as the membrane resistance ofoocyte and nurse cells (plus follicular epithelium) according to another method described byWoodruff & Telfer (1974). Current was injected either into the oocyte or into a nurse cell and theresulting changes in steady-state potentials of the injected and the coupled cell were monitored.Because of the balanced bridge-compensation circuitry of the W.P.I. S-7071 A electrometers used,only two electrodes were needed in order to inject current and measure the voltage changessimultaneously. The intercellular resistance (R{) between oocyte and a nurse cell was thencalculated as:
/, xU2./l2)-(U2/l2)2
(1)
where U\ is the voltage change in the stimulated cell, I\ the injected current, U2 the voltagechange in the coupled 'cell, and Ur, the voltage change in the latter cell when it is stimulatedby a current I2.
In a similar way and in the same notation, the membrane resistance of the oocyte (i?ooc) or a nursecell (Rnc) w a s calculated as:
or
Extrafollicular current measurementsFor estimation of total current, extracellular measurements were carried out using a vibrating
probe (based on the method of Jaffe & Nuccitelli, 1974; improved by Dorn & Weisenseel, 1982) asdescribed by Bohrmannef al. (1986).
RESULTSChanges of intracellular potential in oocyte and nurse cells during vitellogenesis
The intracellular potential varied considerably between different follicles of thesame developmental stage. However, after we had recorded the potentials of a large

210 J. Bohrmann, E. Huebner, K. Sander and H. Gutzeit
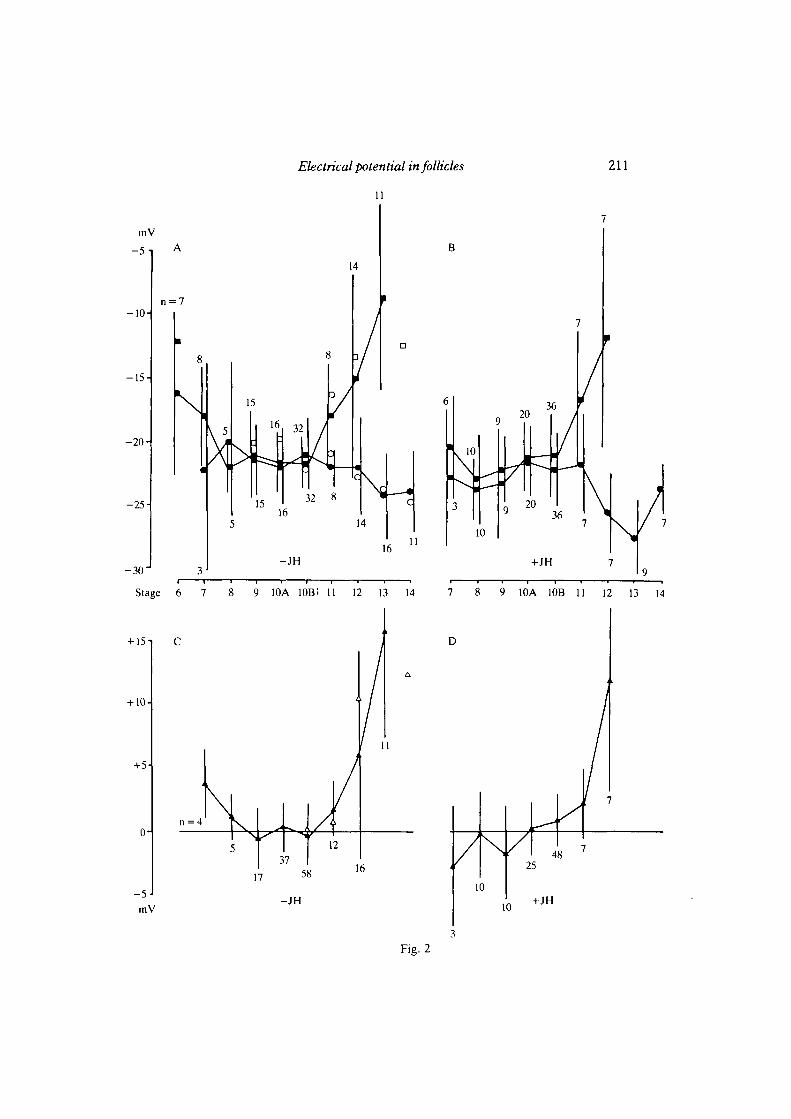
number of follicles (n = 243), a consistent pattern of potential changes duringdifferent phases of oogenesis emerged. For most of vitellogenesis (stages 8-1 OB) themean intracellular potentials recorded in nurse cells and oocyte ranged between —20and — 22 mV (Fig. 2A) . However, some significant changes were apparent bothbefore and after this phase. At stages 6 and 7 (just before and at the beginning ofvitellogenesis) the nurse cell potential was less negative than at mid-vitellogenicstages 9—10. A similar observation was made during late vitellogenesis, when thenurse cells became increasingly depolarized up to about — 9mV at stage 13. At thesame time the oocyte became hyperpolarized, reaching about — 24 mV (Fig. 2A) . Allthese potential changes between stages 6 and 9 as well as between stages 9 and 13 arestatistically significant (P<0 - 05 ) .
Potential differences between oocyte and nurse cells
The potential differences were recorded simultaneously with one electrode in theoocyte and a second electrode in a nurse cell (Fig. 1A,B). Before stage 8 and afterstage 10B the potential differences are significantly different from zero (P < 0-05). Atthese stages the differences are always positive, i.e. the nurse cells represent theanode of the intrafollicular electrophoresis unit. The reverse relationship has beenfound in follicles of Hyalophora (Woodruff & Telfer, 1973) as well as in ovarioles ofRhodnius (Telfer et al. 1981) and Oncopeltus (Woodruff & Anderson, 1984).
From the Hyalophora data, the maximal potential differences in Drosophila shouldbe expected to occur during stages 8-10B. However, we found no significantdifferences (P>0-10) between nurse cells and oocyte in follicles at these stages(Fig. 1B, 2A,C) . Thus Drosophila lacks a pre-condition for efficient transport ofcharged molecules and organelles by intercellular electrophoresis — a sufficientpotential difference between nurse cells and oocyte - while such differences havebeen reported in Hyalophora, Rhodnius and Oncopeltus (Woodruff & Telfer, 1973;Telfer et al. 1981; Woodruff & Anderson, 1984).
Effect of juvenile hormone
Juvenile hormone (JH) was reported to increase the electrical potential differ-ence between tropharium and oocyte in Rhodnius by depolarizing the oocyte
Fig. 2. A,B. Diagram of the mean intracellular electrical potentials and their stage-specific changes in wild-type nurse cells (•) and oocytes (•) of stage 6—14 follicles inR-14 medium (A) and in medium with juvenile hormone analogue (B). Bars indicatestandard deviation; n, numbers of investigated follicles. At stage 6 the accurate deter-mination of the potential of the oocyte was impossible due to its small size. Some meanvalues of potential measurements in follicles of the mutants die and bicD are included in Aand c. (•) die nurse cells; (O) die oocytes; (H) bicD nurse cells. C,D. Diagram of meanpotential differences (nurse cell minus oocyte) during stages 7-13 in normal (c) and JH-containing culture medium (D) determined by simultaneously recording in a nurse celland the oocyte. In many cases the potentials of several nurse cells in one follicle weremeasured one after the other relative to the oocyte. (A) wild-type; (A) die.
Electrical potential in follicles 211
+JH
Stage 6 7 8 9 10A lOBi 11 12 13 14 7 8 9 1OA 1OB 11 12 13 14
+ 15
+ 10-1
+5
(H
- 5mV
n = 4
3712
D
17 58 16
- J H10
4825
10+JH
Fig. 2

212 J. Bohrmann, E. Huebner, K. Sander and H. Gutzeit
(Telfer et al. 1981). In Drosophila follicles JH produced a minor but noticeable effectin the opposite direction. The oocyte was significantly hyperpolarized at stages 10B,12 and 13 (P<0-05; Fig. 2B), while there was no effect on the nurse cell potential(P>0-10). As a result, a small but significant positive potential difference(P< 0-025; Fig. 2D) already became apparent at stage 10B, in contrast to non-JH-treated follicles, where it appeared at stage 11. JH also abolished significantly(P > 0-10) the positive potential difference (P < 0-05) during stage 7 of oogenesis byhyperpolarizing the nurse cells (P<0-05). However, at no stage did JH causesignificant negative potential differences as reported for Rhodnius (Teller et al.1981).
Mutants dicephalic and bicaudal D
Some follicles of the mutant dicephalic (die) possess nurse cell clusters at bothpoles of the follicle, so that the growing oocyte is nursed from both ends (Lohs-Schardin, 1982). The intracellular potential measurements (17 follicles) of the nursecells (at both ends) and the oocyte gave no indication of obvious differences fromwild-type follicles (Fig. 2A,c). In some cases we were able to measure the intra-cellular potential in individual nurse cells that did not participate in the process ofregression, from stage 10B onward, of the nurse cell chamber (Bohrmann, 1981).Presumably, these cells lack an intact cytoplasmic bridge(s) through which cyto-plasm is transported (see Frey, Sander & Gutzeit, 1984). Our measurements indicatethat these 'intact' nurse cells lose their negative potential along with the regressingnurse cells (Fig. 2A,c).
In follicles of the mutant bicaudalD (bicD; T. Schupbach, unpublished data) all16 cystocytes appear to differentiate into nurse cells, so that these follicles do notpossess a functional oocyte capable of entering vitellogenesis; the follicles are blockedat stage 6. The nurse cell potential of these follicles (—12-1 ± 3-2mV (S.D.), n = 12)did not differ significantly (P>0-10) from that of wild-type follicles at stage 6(-16-1 ±6-4mV (s.D.), n = 7).
Determination of resistance
The intercellular resistance between oocyte and nurse cells at stages 10A—11was determined using three different methods as described by Woodruff & Telfer(1974). The voltage-clamp method gave consistently the highest values (Table 1A).Alternatively, current was injected into one cell (either oocyte or nurse cell) and thechange in the steady-state potential of both cells was recorded simultaneously. Thecalculated resistance (see Materials and Methods) did not differ significantly whencurrent was injected into either the oocyte or a nurse cell (P>0-05; Table 1A).
On the basis of the latter measurements the membrane resistances of oocyte andnurse cells (including the follicular epithelium) were also calculated (see Materialsand Methods and Table IB). The resistance of the nurse cell membranes was foundto be significantly higher than that of the oocyte membrane (P<0-05; Table IB).Stage-specific differences were not detected.

Electrical potential in follicles 213
Estimate of intrafollicular current
Although we have been unable to detect significant intercellular potentialdifferences between oocyte and nurse cells during mid-vitellogenic stages (seeabove), the follicles still possess an extracellular electrical field that has beencharacterized previously with a highly sensitive vibrating probe (Bohrmann et al.1984, 1986; Overall & Jaffe, 1985).
The current flux through the follicle can be estimated by integrating the currentflowing through the plane that bisects the extrafollicular electrical field, as shown inFig. 3A (see also Weisenseel, Nuccitelli & Jaffe, 1975). If the current is considered asoriginating from a dipole consisting of a point source and a point sink located insidethe follicle (which is suggested by the geometry of the extrafollicular electrical fieldwith its lateral maxima; see Bohrmann et al. 1986) the field (E) must fall with thedistance (r) from the dipole axis according to:
E<x (r2 + Hz)-3/1. (3)
where H is the dipole half-length (see Weisenseel et al. 1975, and Fig. 3D).When the actual decrease in current density (which is proportional to E) with the
distance from the follicle axis was determined for 21 wild-type and two die follicles(stages 8-11) with the vibrating probe and the experimental data were fitted to thecurves of different dipole half-lengths (e.g., see Fig. 3A-C) a surprising result wasobtained: the estimated length of the hypothetical dipole in the follicle axis is onlyabout 2 jim (H — 1 /im) in wild-type follicles at stages 8 and 9 as well as in both diefollicles at stages 10A and 11 (Table 2), but in wild-type follicles it increases up toabout 140-180 /im (H about 70-90 /im) at stages 10A-11 (Table 2). Although we donot know the actual intrafollicular location of current source and sink, these datasuggest that at stage 10A some fundamental electrophysiological properties of thefollicle change. In the small number oldie follicles analysed, this change could eitherbe hidden by the extreme differences in the extrafollicular electrical field comparedto wild-type follicles (see Bohrmann et al. 1986), or such changes might not occur.
By means of the best-fitting curve (for examples, see Fig. 3A—c) we extrapolatedfor each follicle the current density (i8) directly at the follicle surface and thencalculated, with the corresponding half-length (H) and the follicle radius (a), thetotal current (/) flowing outside the follicle through the bisecting plane (Fig. 3A):
, (4)
(see Weisenseel et al. 1975). In individual follicles this current varies between 0-2and 12nA and has a stage-specific maximum at stage 10A/B (Fig. 4). The currentscalculated in a different way by Overall & Jaffe (1985) lie in a similar range.
If current source and sink are located near the oocyte/nurse cell border (alsoassumed in the model of Jaffe & Woodruff (1979) for Hyalophora), the voltage (U)

214 J. Bohrmann, E. Huebner, K. Sander and H. Gutzeit
Table 1A. Intercellular resistance in vitellogenic follicles
Voltage clamp
Method after Woodruff & Telfer (1974),current applied to:
method Oocyte Nurse cell
Intercellular resistance 24-8 ±7-8 9-0 ± 6-9 6-4 ±5-4x±S.D. (kQ)»
B. Membrane resistance of oocyte and nurse cell
Oocyte membrane Nurse cell membrane
Membrane resistance 2-5 ± 1 2 4-4 ±3-4 6-8 ±2-8X ± S . D .
*n = 15 Drosophila wild-type follicles stages 10A-11 (always same sample); five more follicleswere measured by the voltage clamp method only.
that would produce this extrafollicular current (/) can be calculated, since themembrane resistances of oocyte (Rooc) and nurse cells (Rnc)
a r e known (Table IB):
U = IX(ROOC+Rnc). (5)
This calculated potential difference (—1 to — 84/iV) between oocyte and nurse cellsis compatible with our intracellular measurements, since such low voltage differencesare well below the sensitivity of glass capillary electrodes. From the voltage and theknown intercellular resistance (Table 1A) we calculated the intrafollicular currentbetween the oocyte and one nurse cell to amount to only 0 04—13nA in differentfollicles (stages 8-11).
DISCUSSION
In this and the preceding paper (Bohrmann et al. 1986) we have shown that inDrosophila follicles stage-specific electrical phenomena can be recorded withintracellular and extracellular electrodes. The results reported here are qualitativelyand quantitatively different from the results obtained in other insect species,although we applied the same methods and worked with the same underlyingassumptions. Particularly surprising to us was the absence of a statistically
Fig. 3. A, inset. The extrafollicular current is integrated over the stippled plane thatbisects the follicle and its electrical field. The long axis of the follicle is considered to bethe axis of the hypothetical dipole. With the vibrating probe ( 11 ) we measured thecurrent density parallel to this axis at various distances from the follicle surface. A-C.Examples of current density decrease with the distance (r) from the follicle's axis in wild-type follicles (stages 9—11). Each follicle is shown with the same symbols at differentmeasuring positions. The current density (i) is given in relative units (»'/i), where (T) is thestandard current density at 10 /an from the follicle's surface. The data of differentdevelopmental stages fit to theoretical curves calculated for different dipole half-lengths(H) with eqn (3) (see the text and Table 2). D. Examples of the curves calculated fordifferent dipole half-lengths (H), shown in fim. Field strength (E) is given in arbitraryunits with a proportional factor of 106, which resulted in best-fitting curves.

Electrical potential in follicles
Direction ofcurrent flow - "•
Measuring \position
1-0
0-8-
0-6 •
0-4-
0-2
0-8
Bisecting plane
Long axis 0-6"
Stage 9
H = 1 um
I(X) 2(K) 300
r (/mi)
0-4-
0-2
215
8
100 200 300
r (
D
X0-6-
0-4
0-2
Stages 10B and 11
100 200 300 300
Fig. 3

216 jf. Bohrmann, E. Huebner, K. Sander and H. Gutzeit
Table 2. Stage-specific change in half-length of electrical dipole
Stage
Estimated half-length H (/im)Number of (x±S.D.)of
follicles hypothetical electrical dipole
Wild-type
dicephalic
89
10A10B11
10A11
226ftI11
11
70 ± 2 274 ±1685 ± 7
11
significant intercellular potential difference between oocyte and nurse cells duringmid-vitellogenic stages. Could, perhaps, a small voltage difference of —1 to —84/xV(the range suggested by our calculations), which we would not pick up withintracellular recordings, still result in intercellular migration of charged molecules?
If the electrophoretic mobility is about lfims"1 per Vcm"1 (Jaffe, 1977;Nuccitelli, 1983) the distance, for example, between an apical nurse cell and theooplasm bordering the nurse cells at stage 10A (approx. 150/im, which is also aboutthe length of the hypothetical dipole) would be traversed by a charged molecule in20—150 h. Such slow migration of molecules appears unreasonable in view of the factthat the oogenesis in Drosophila (stage 1-14) takes only about 80 h altogether(Mahowald & Kambysellis, 1980). However, electrophoretic transport over shortdistances (from the proximal nurse cells to the oocyte) is more likely to occur: if one
101
I 6
U 4-
S t a g e X l) IDA IDA IMA H 1MB 11
Fig. 4. Diagram of stage-specific differences in mean total current through 21 wild-typefollicles (stages 8-11), calculated using eqn (4) (see the text). Bars indicate 1/2 standarddeviation; n, numbers of investigated follicles.

Electrical potential in follicles 217
assumes that the calculated potential difference acts only over a small distance of, forexample, 5 fim through the cytoplasmic bridge, this distance would be traversed in1-8 min.
The potential difference observed during late stages of oogenesis is opposite to theexpected value and the results from other insects: in Drosophila the bulk ofmacromolecules and organelles (mostly negatively charged) if moved electro-phoretically would migrate instead from the oocyte into the nurse cells 1 However,during the phase of nurse cell regression the nurse cell cytoplasm streams into theoocyte, presumably as a result of microfilament contraction (Gutzeit, 1986) and,therefore, an electrophoretic transport mechanism would be superfluous (andprobably ineffective) at this stage of oogenesis. Yet in other species with polytrophicovaries (including Hyalophora) cytoplasmic streaming by contractile mechanismsdoes not seem to occur (H. Gutzeit, unpublished data).
We have compiled the available data for Drosophila and Hyalophora in Table 3 topermit a direct comparison. For Hyalophora intrafollicular potential differences ofseveral mV were recorded during mid-vitellogenesis, and consequently the estimatedelectrophoretic velocity of molecules between an apical nurse cell and the oocyte(approx. 500/jm) was about 50 times higher than that calculated for Drosophilafollicles at comparable stages (9-10). Also the calculated current between a singlenurse celland the oocyte was 100-200 times higher in Hyalophora than in Drosophilafollicles. Another difference between the two species is that in Drosophila themembrane resistance of the nurse cell chamber and the oocyte is nearly the same asthe intercellular resistance between nurse cells and oocyte. Therefore, comparativelymore current should be shunted via the extrafollicular current loop in Drosophilathan in Hyalophora, in which the membrane resistance is 10 times higher than theintercellular resistance. The differences beween both species in intrafollicularpotential difference cannot be attributed to juvenile hormone, which in Rhodnius wasreported to increase the potential difference between oocyte and tropharium (Telferet al. 1981). But we cannot exclude the possibility that Drosophila follicles, in orderto establish or maintain an electrical potential difference, need some unknown fac-tors, lacking in our culture medium. However, transport processes are maintained inthis medium, as shown by time-lapse filming and in vitro autoradiography (Gutzeit& Koppa, 1982; H. Gutzeit, unpublished data), and by the fact that the folliclesdevelop in vitro from stage 10 up to stage 14 (Petri et al. 1979; Bohrmann, 1981).
JH was found to affect the oocyte and the nurse cell potential independently indifferent developmental stages. Whereas the oocyte was hyperpolarized significantlyonly at stages 10B, 12 and 13, the nurse cells were hyperpolarized only at stage 7.Probably, at stage 14 the oocyte is not affected by JH because the chorion forms abarrier. The low nurse cell potential of stages 6—7 (see also bicD follicles) is raised byJH applied during stage 7, to reach levels that are normally reached later duringoogenesis (stages 8-10).
We often found that different nurse cells within the same follicle had differentintracellular electrical potentials (also some of them may be positive while others arenegative compared to the oocyte potential). A potential gradient related to the

Tab
le 3
. C
ompa
riso
n be
twee
n D
roso
phil
a an
d H
yalo
phor
a
Intr
acel
lula
r po
tent
ial
of
oocy
te a
nd n
urse
cel
ls
Hya
loph
ora
hso
ph
ila
(c
ompa
rabl
e m
id-
(sta
ges
9- 1
0)
vite
llog
enic
sta
ges)
R
efer
ence
s (f
or H
yalo
phor
a)
U,
=-2
1mV
U
nc =
-21
mV
U
, -3
0 m
V
Un
c -40
mV
W
oodr
uff
& T
elfe
r (1
973)
9
-40
mV
-4
6 m
V
Tel
fer
et a
l. (
1981
) -4
7 m
V
-50
mV
W
oodr
uff,
Hue
bner
&
5? T
elfe
r, p
erso
nal
z D M
embr
ane
resi
stan
ce
Intr
afol
llcu
lar
resi
stan
ce
Tot
al e
xtra
foll
icul
ar c
urre
nt
Intr
afol
licu
lar
curr
ent
betw
een
one
nurs
e ce
ll an
d cm
yte
Mea
sure
d po
tent
ial
diff
eren
ce
betw
een
one
nurs
e ce
ll an
d oo
cyte
Cal
cula
ted
pote
ntia
l di
ffer
ence
ba
sed
on e
xtra
foll
icul
ar
curr
ent
Cal
cula
ted
elec
trop
hore
tic
velo
city
of
nega
tive
ly c
harg
ed
mol
ecul
es f
rom
nur
se c
ells
to
oocy
te
com
mun
icat
ion
Woo
druf
f &
Tel
fer
(197
4)
Woo
druf
f &
Tel
fer
(197
4)
Jaff
e &
Woo
druf
f (1
979)
Woo
druf
f &
Tel
fer
(197
4)
(our
cal
cula
tion
, ba
sed
on
extr
afol
licu
lar
curr
ent)
Woo
druf
f &
Tel
fer
(197
3)
Tel
fer
et a
l. (
1981
) W
oodr
uff,
Hue
bner
&
Tel
fer,
per
sona
l co
mm
unic
atio
n
(our
cal
cula
tion
)
v,,,
1-7
ph
-'
v,,,
, 36
-468
p h
-' (o
ur c
alcu
lati
on)

Electrical potential in follicles 219
distance of the respective nurse cell from the oocyte was not found. It appears thatnurse cells can individually regulate their membrane potential.
The nurse cells become depolarized during late vitellogenesis but this is not onlydue to their regression caused by cytoplasmic streaming into the oocyte, since non-regressing nurse cells, which are often found in die follicles of stages 13 and 14(Bohrmann, 1981), also lose their negative intracellular potential in a stage-specificmanner. The depolarization of the nurse cells during late stages of vitellogenesis(observed with JH already from stage 10B on) is not reflected in the extrafollicularcurrent pattern (Bohrmann et al. 1986; Overall & Jaffe, 1985). These observationsindicate that at least part of the extrafollicular current pattern is produced by thefollicular epithelium, not exclusively by the germ-line cells.
The observed increase in oocyte potential during oogenesis correlates well withthe intracellular potential of fertilized eggs (—27 mV) and early embryonic stages(Miyazaki & Hagiwara, 1976). Extracellular electrical currents (Bohrmann et al.1984, 1986; Overall & Jaffe, 1985) may reflect the necessity of maintaining this nega-tive intracellular potential in the developing oocyte as well as in the young embryo.
Unfortunately, we know very little about the role of the follicular epithelium forthe intra- and extrafollicular current pattern. The follicle cells that migrate betweenthe oocyte and nurse cells have been thought to play a role in the intrafollicularcurrent loop of Hyalophora follicles (Jaffe & Woodruff, 1979). In Drosophila thesecells migrate centripetally at the time when a drastic increase in the hypotheticaldipole length has just appeared (late stage 10A to early 10B) and when theextrafollicular current is at the maximum level. Furthermore, groups of Ca2+-richcells were detected amongst this population of follicle cells (Heinrich & Gutzeit,1985) but their function remains to be elucidated.
We thank Drs M. Weisenseel and A. Dorn for helpful discussions and for making it possible tocarry out the vibrating probe work in their laboratory. The Deutsche Forschungsgemeinschaftgave financial support. Facilities in the laboratory of E.H. are maintained with Canadian NSERCfunds.
REFERENCESBIER, K. (1963). Autoradiographische Untersuchungen fiber die Leistungen des Follikelepithels
und der Nghrzellen bei der Dotterbildung und EisweiBsynthese im Fliegenovar. Wilhelm RouxArch. EntivMech. Org. 154, 552-575.
BOHRMANN, J. (1981). Enrwicklung der Follikel der Drosophila-Mutante dicephalic in vitro.Staatsexamensarbeit, Universitat Freiburg.
BOHRMANN, J., DORN, A., SANDER, K. & GUTZETT, H. (1986). The extracellular electrical currentpattern and its variability in vitellogenic Drosophila follicles. J . Cell Sci. 81, 189-206.
BOHRMANN, J., HEINRICH, U.-R., DORN, A., SANDER, K. & GUTZEIT, H. (1984). Electricalphenomena and their possible significance in vitellogenic follicles of Dmsophila melanogaster.J. Embryol. exp. Morph. 82 Supplement, 151.
DITTMANN, F., EHNI, R. & ENGELS, W. (1981). Bioelectric aspects of the hemipteran telotrophicovariole (Dysdercus intermedius). Wilhelm Roux Arch. EntwMech. Org. 190, 221-225.
DORN, A. & WEISENSEEL, M. H. (1982). Advances in vibrating probe techniques. Protoplasma113, 89-96.

220 J. Bohrmann, E. Huebner, K. Sander and H. Gutzeit
FREY, A., SANDER, K. & GUTZEIT, H. (1984). The spatial arrangement of germ line cells inovarian follicles of the mutant dicephalic in Drosophila melanogaster. Roux Arch, devl Biol. 193,388-393.
GlORGI, F. (1979). In vitro induced pinocytotic activity by a juvenile hormone analogue in oocytesolDrosopkila melanogaster. Cell Tiss. Res. 203, 241-247.
GUTZEIT, H. O. (1986). The role of microfilaments in cytoplasmic streaming in Drosophilafollicles. J . Cell Sri. 80, 159-169.
GUTZEIT, H. O. & KOPPA, R. (1982). Time-lapse film analysis of cytoplasmic streaming duringlate oogenesis in Drosophila melanogaster. J. Embryol. exp. Morph. 67, 101-111.
HETNRICH, U.-R. & GUTZEIT, H. O. (1985). Characterization of cation-rich follicle cells invitellogenic follicles of Drosophila melanogaster. Differentiation 28, 237-243.
HUEBNER, E. (1984). Developmental cell interactions in female insect reproductive organs. InAdvances in Invertebrate Reproduction, vol. 3 (ed. W. Engels, W. H. Clark, Jr, A. Fischer,P. J. W. Olive & D. F. Went), pp. 97-105. Amsterdam: Elsevier.
JAFFE, L. F. (1977)-. Electrophoresis along cell membranes. Nature, Land. 265, 600-602.JAFFE, L. F. & NUCCTTELLI, R. (1974). An ultrasensitive vibrating probe for measuring steady
extracellular currents. 7. Cell Biol. 63, 614-628.JAFFE, L. F. & WOODRUFF, R. I. (1979). Large electrical currents traverse growing cecropia
follicles. Pmc. natn. Acad. Sri. U.SA. 76, 1328-1332.KIEHART, D. P. (1982). Microinjection of echinoderm eggs. inMethods in Cell Biology, vol. 25B,
(ed. L. Wilson), pp. 13-31. New York: Academic Press.KING, R. C. (1970). Ovarian Development in Drosophila melanogaster. New York: Academic
Press.LOHS-SCHARDIN, M. (1982). Dicephalic - a Drosophila mutant affecting polarity in oogenesis and
embryogenesis. Wilhelm Roux Arch. EntwMech. Org. 191, 28-36.MAHOWALD, A. P. & KAMBYSELUS, M. P. (1980). Oogenesis. In Genetics and Biology of
Drosophila, vol. 2d (ed. M. Ashburner & T. R. F. Wright), pp. 141-224. New York: AcademicPress.
MIYAZAKI, S. & HAGIWARA, S. (1976). Electrical properties of the Drosophila egg membrane. DeviBiol. 53, 91-100.
NUCCTTELLI, R. (1983). Transcellular ion currents: Signals and effectors of cell polarity. InModern Cell Biology, vol. 2 (ed. B. H. Satir), pp. 451-481. New York: Liss.
OVERALL, R. & JAFFE, L. F. (1985). Patterns of ionic current through Drosophila follicles and eggs.Devi Biol. 108, 102-119.
PETRI, W. H., MINDRINOS, M. N., LOMBARD, M. F. & MARGARITA, L. H. (1979). In vitro
development of the Drosophila chorion in a chemically defined organ culture medium. WilhelmRoux Arch. EntwMech. Org. 186, 351-362.
ROBB, J. H. (1969). Maintenance of imaginal discs of Drosophila melanogaster in chemicallydefined media. J . Cell Biol. 41, 876-885.
SACHS, L. (1978). Angewandte Statistik. Berlin: Springer Verlag.SlGURDSON, W. J. (1984). Bioelectrical aspects of the Rhodnius pwlixus ovariole: Extracellular
current mapping during oogenesis. Master thesis, University of Manitoba.TELFER, W. H. (1975). Development and physiology of the oocyte-nurse cell syncytium. Adv.
Insect Physiol. 11, 223-319.TELFER, W. H., WOODRUFF, R. I. & HUEBNER, E. (1981). Electrical polarity and cellular
differentiation in meroistic ovarioles. Am. Zool. 21, 675-686.WEISENSEEL, M. H., NUCCITELU, R. & JAFFE, L. F. (1975). Large electrical currents traverse
growing pollen tubes. J . Cell Biol. 66, 556-567.WOODRUFF, R. I. & ANDERSON, K. L. (1984). Nutritive cord connection and dye-coupling of the
follicular epithelium to the growing oocytes in the telotrophic ovarioles in Oncopeltus fasriatus,the milkweed bug. Roux Arch, devl Biol. 193, 158-163.
WOODRUFF, R. I. & TELFER, W. H. (1973). Polarized intercellular bridges in ovarian follicles ofthe cecropia moth.J. Cell Biol. 58, 172-188.

Electrical potential in follicles 221
WOODRUFF, R. I. & TELFER, W. H. (1974). Electrical properties of ovarian cells linked byintercellular bridges. Ann. N.Y. Acad. Set. 238, 408-419.
WOODRUFF, R. I. & TELFER, W. H. (1980). Electrophoresis of proteins in intercellular bridges.Nature, Land. 286, 84-86.
(Received30July 1985 -Accepted 10 September 1985)
