insulin stimulates the translation of ribosomal proteins ... stimulates the translation of ribosomal...
TRANSCRIPT

THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1988 by The American Society for Biochemistry and Molecular Biology, Inc
Vol. 263, No. 33, Issue of November 25, PP. 17785-17791.1988 Printed in U. S. A.
Insulin Stimulates the Translation of Ribosomal Proteins and the Transcription of rDNA in Mouse Myoblasts*
(Received for publication, June 6, 19888)
Mark L. Hammond and Lewis H. Bowman From the Department of Biology, University of South Carolina, Columbia, South Carolina 29208
Insulin stimulates the translation of ribosomal pro- tein (r-protein) mRNAs and the transcription of rDNA in mouse MM14DZ myoblasts. Analysis of the distri- bution of S16, LlS, and L32 r-protein mRNAs in pol- ysome gradients indicates that the increased transla- tion of these mRNAs in insulin-treated myoblasts is due to the recruitment of mRNAs that were not previ- ously being translated. In contrast, the translational efficiencies of &actin, c-myc, and p31 mRNAs are not affected by insulin. Hybridization analysis of RNA transcribed in nuclear run-on reactions indicates that insulin also stimulates the transcription of rDNA. Both the increases in r-protein translation and rDNA tran- scription occur coordinately and are maximal within 15 min of insulin treatment of myoblasts. However, insulin has no effect on the rate of cell division or the steady state levels of r-protein mRNAs. Surprisingly, after myoblasts differentiate into fibers, insulin does not affect the r-protein mRNA translation or rDNA transcription. These experiments indicate that the syn- thesis of the macromolecular components of ribosomes is tightly and coordinately controlled in myoblasts.
Eukaryotic ribosomes are comprised of equimolar amounts of 70-80 different ribosomal proteins (r-proteins)’ and 4 rRNAs. To understand the mechanisms regulating the bio- synthesis of eukaryotic ribosomes it must be determined how the equimolar accumulation of rRNA and r-protein is coor- dinated. In some cases either rRNA or r-protein is produced in excess, and the accumulation of rRNA and r-protein is coordinated by the turnover of the overproduced species (Craig and Perry, 1971; Clissold and Cole, 1973; Cooper, 1973; Abelson et al., 1974; Bowman and Emerson, 1977; Warner, 1977; Krauter et al., 1980; Wolf et al., 1980; LaMarca and Wasserman, 1984; Jacobs et al., 1985; Pierandrei-Amaldi et al., 1985; ElBaradi et al., 1986; Bell et al., 1988). In many eukaryotic systems equimolar accumulation is accomplished by the synthesis of rRNA and r-protein in approximately equal amounts. However, the mechanisms involved in coor- dinating their synthesis are not known (DePhilip et al., 1980; Faliks and Meyuhas, 1982; Pierandrei-Amaldi et al., 1982; Bozzoni et al., 1984; Al-Atia et al., 1985; Schmidt et al., 1985; Agrawal and Bowman, 1987; Meyuhas et al., 1987).
The possibility that the synthesis of eukaryotic r-proteins is regulated by feedback inhibition as in bacteria (Nomura et
* This work was supported by United States Public Health Service Grant AR 32221 from the National Institutes of Health. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked ‘‘uduer- tisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
The abbreviation used is: r-protein, ribosomal protein.
al., 1984) has been examined by introducing extra copies of r- protein genes or mRNAs into cells (Pearson et al., 1982; Bozzoni et al., 1984; Abovich et al., 1985; Warner et al., 1985; Bowman, 1987b; Rhoads and Roufa, 1987; Pierandrei-Amaldi et al., 1988; Baum et al., 1988). In these cases, the r-proteins are overproduced in proportion to their gene copy number and rapidly turn over. However, in two notable cases the overproduction of an r-protein inhibits the processing of its own mRNA, but the synthesis of other r-proteins is not affected (Bozzoni et al., 1984; Dabeva et al., 1986). Despite these examples, autogenous regulation appears not to be a general mechanism for regulating r-protein synthesis. Only a fraction of the r-proteins has been examined in this manner, and it is still possible that the overproduction of some unstu- died r-proteins may inhibit the synthesis of other r-proteins.
The differentiation of mouse MM14DZ myoblasts in cell culture provides an excellent system for studying the mecha- nisms coordinating rRNA and r-protein formation. Myoblasts divide rapidly in culture until the medium is depleted of growth-promoting factors. Then they withdraw from the cell cycle and fuse into fibers. As the cells withdraw, the synthesis of the muscle-specific contractile proteins initiates (Hauschka et al., 1979; Linkhart et al., 1981; Gospodarowicz et al., 1984). We have previously shown that after mouse myoblasts differ- entiate into fibers the rate of ribosome formation decreases 2-3-fold due to a decrease in the transcription of rDNA and to a decrease in the translation and steady state levels of r- protein mRNAs (Agrawal and Bowman, 1987; Bowman, 1987a). However, other laboratories obtain different results when studying ribosome formation during the differentiation of rat myoblasts (Krauter et al., 1980; Jacobs et al., 1985). In rat only the transcription of rDNA decreases following myo- blast differentiation (Krauter et al., 1979; Jacobs et al., 1985); neither the steady state levels nor the translational efficien- cies of r-protein mRNAs decrease (Jacobs et al., 1985).
In this paper we show that insulin stimulates the transla- tion of r-protein mRNAs and the transcription of rDNA in mouse myoblasts but not in fibers. Maximal stimulation oc- curs within 15 min, making this an excellent system for studying the coordination of r-protein and rRNA metabolism.
EXPERIMENTAL PROCEDURES
Cell Culture”MM14DZ mouse myoblasts (Hauschka et al., 1979) were grown as described previously using partially purified fibroblast growth factor (Bowman, 1987b). Cultures were fed fresh media 24 h after passage. Myoblast fusion initiates after 72 h, and by 120 h greater than 95% of the nuclei are within fibers. Myoblasts were harvested at 48 h and fibers a t 120 h after passage.
Bovine insulin (Collaborative Research, Inc.) was dissolved in water and added to myoblast cultures to a final concentration of 4 Fg/ml 24 h after passage. Experimental fiber cultures received an additional insulin dosage added directly to the plates 96 h after passage unless otherwise indicated.
17785
17786 Insulin Stimulates r-Protein and rRNA Synthesis in Myoblasts Polysome and RNA Isolation-Myoblast and fiber cultures were
made 90 Fg/ml in cycloheximide immediately prior to harvesting. Cultures were harvested in 0.3 M NaCl, 10 mM Tris-HC1, pH 7.4, 10 mM MgC12, 1% Triton X-100, 90 pg/ml cycloheximide and gently homogenized with four strokes in a loose Dounce homogenizer. Ly- sates were centrifuged at 15,000 X g for 5 min, and the supernatant was layered onto an 11-m10.5-1.5 M sucrose gradient with a 1-ml pad of 2 M sucrose made in the above buffer without Triton X-100. Gradients were centrifuged at 40,000 rpm for 165 min in an SW 41 rotor, fractionated, and the absorbance at 254 nm monitored (Agrawal and Bowman, 1987).
Sucrose gradient fractions were made 0.7% in sodium dodecyl sulfate. 20 pg of bacterial tRNA and 20,000 cpm of tritium-labeled bacterial RNA of a high specific activity was added to each fraction to act as carrier and recovery control RNA, respectively. RNA was isolated by phenol-chloroform extraction and ethanol-precipitated twice prior to gel fractionation.
RNA was isolated from myoblast and fiber cultures by phenol- chloroform and LiCl extraction as described (Agrawal and Bowman, 1987; Bowman, 1987a).
Hybridization Analysis-RNAs isolated from both polysome gra- dients and cultures were denatured with glyoxal (McMaster and Carmichael, 1977). A small aliquot from the gradient fractions was measured by liquid scintillation counting to determine the recovery of RNA. Parallel gels were generated by loading equal volumes of the denatured RNA onto different 1% agarose gels. The agarose gels were blotted and hybridized (Agrawal and Bowman, 1987) to nick-trans- lated isolated cDNA inserts for S16, L18, L32 (Meyuhas and Perry, 1980; Agrawal and Bowman, 1987), p31 (Theodor et al., 1985), and p- actin (Ponte et al., 1983) mRNA, pM c-myc54 plasmid for c-myc mRNA (Stanton et al., 1983), and p5B plasmid for 18 S rRNA (Bowman et al., 1981). Filters were rinsed as described (Agrawal and Bowman, 1987) and exposed to Kodak X-Omat AR x-ray film with an intensifying screen at -70 "C. The exposed autoradiographs were scanned in a Helena Quick Scan densitometer to determine the relative hybridized signal in each lane.
Nuclear Transcriptions-Nuclei from myoblast and fiber cultures were isolated and incubated in transcription reaction buffer contain- ing heparin as described (Bowman, 1987a). RNA was isolated and hybridized to the external transcribed spacer probe (p3) (Bowman et al., 1981) as described (Bowman, 1987a).
Filters were rinsed in 0.3 M NaC1, 10 mM Tris-HC1, pH 7.4, 2 mM EDTA, pH 7.4, and 0.1% sodium dodecyl sulfate for 24 h at 42 "C. Hybridized RNA was eluted from the filters and quantitated by scintillation counting (McKnight and Palmiter, 1979).
DNA Quantitation-Cell cultures were harvested in a buffer con- taining 0.5 M NaCl and 2 mM EDTA, pH 7.4, homogenized with a syringe and a 26-gauge needle, and frozen at -20 "C. Thawed samples were made 2.0 M NaC1,50 mM NaP04, pH 7.4, and 0.1 pg/ml Hoechst 33258 dye (Labarca and Paigen, 1980). The fluorescence was deter- mined using a Turner model 111 fluorometer. The DNA content of each sample was then determined from a standard curve derived from dilutions of a calf thymus DNA standard.
RESULTS
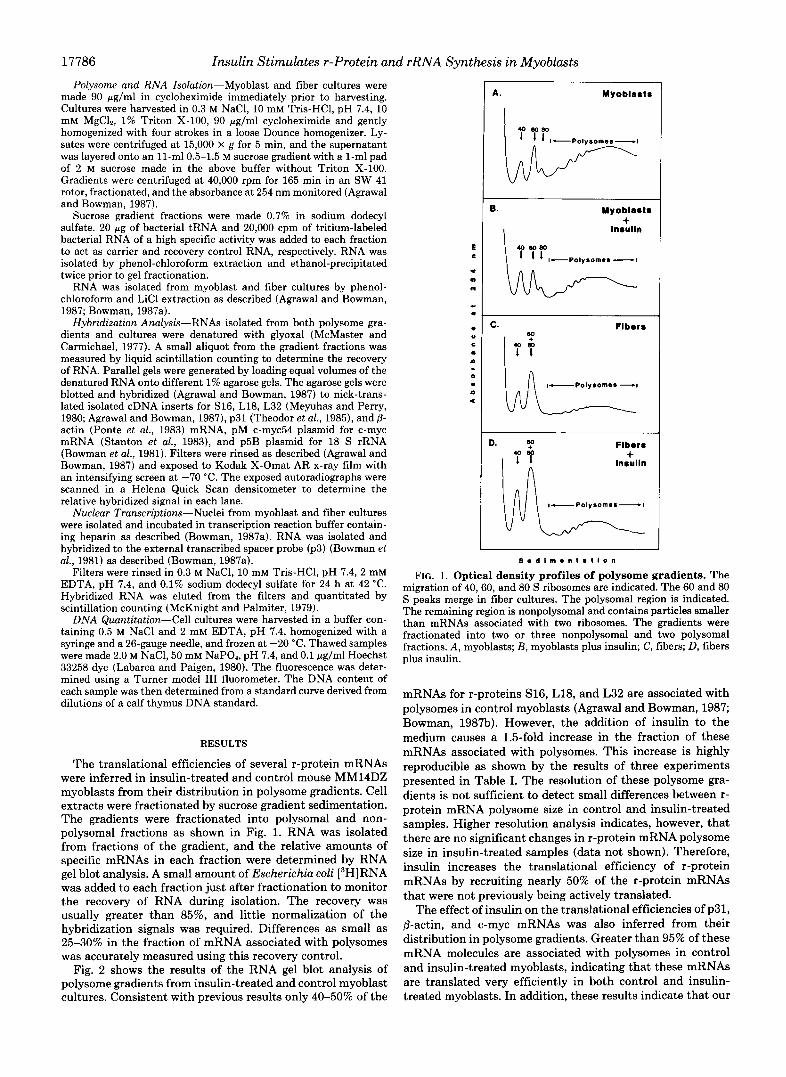
The translational efficiencies of several r-protein mRNAs were inferred in insulin-treated and control mouse MM14DZ myoblasts from their distribution in polysome gradients. Cell extracts were fractionated by sucrose gradient sedimentation. The gradients were fractionated into polysomal and non- polysomal fractions as shown in Fig. 1. RNA was isolated from fractions of the gradient, and the relative amounts of specific mRNAs in each fraction were determined by RNA gel blot analysis. A small amount of Escherichia coli [3H]RNA was added to each fraction just after fractionation to monitor the recovery of RNA during isolation. The recovery was usually greater than 85%, and little normalization of the hybridization signals was required. Differences as small as 25-30% in the fraction of mRNA associated with polysomes was accurately measured using this recovery control.
Fig. 2 shows the results of the RNA gel blot analysis of polysome gradients from insulin-treated and control myoblast cultures. Consistent with previous results only 40-50% of the
D. eo + Fibor8 + In8ulln
-\ ~~
S e d l m e n t a t l o n
FIG. 1. Optical density profiles of polysome gradients. The migration of 40,60, and 80 S ribosomes are indicated. The 60 and 80 S peaks merge in fiber cultures. The polysomal region is indicated. The remaining region is nonpolysomal and contains particles smaller than mRNAs associated with two ribosomes. The gradients were fractionated into two or three nonpolysomal and two polysomal fractions. A, myoblasts; B , myoblasts plus insulin; C, fibers; D, fibers plus insulin.
mRNAs for r-proteins S16, L18, and L32 are associated with polysomes in control myoblasts (Agrawal and Bowman, 1987; Bowman, 1987b). However, the addition of insulin to the medium causes a 1.5-fold increase in the fraction of these mRNAs associated with polysomes. This increase is highly reproducible as shown by the results of three experiments presented in Table I. The resolution of these polysome gra- dients is not sufficient to detect small differences between r- protein mRNA polysome size in control and insulin-treated samples. Higher resolution analysis indicates, however, that there are no significant changes in r-protein mRNA polysome size in insulin-treated samples (data not shown). Therefore, insulin increases the translational efficiency of r-protein mRNAs by recruiting nearly 50% of the r-protein mRNAs that were not previously being actively translated.
The effect of insulin on the translational efficiencies of p31, /3-actin, and c-myc mRNAs was also inferred from their distribution in polysome gradients. Greater than 95% of these mRNA molecules are associated with polysomes in control and insulin-treated myoblasts, indicating that these mRNAs are translated very efficiently in both control and insulin- treated myoblasts. In addition, these results indicate that our

probe:
S16
L18
L32
P31
Insulin Stimulates r-Protein and rRNA Synthesis in Myoblasts
control + insulin
polysomal polysomal polysomal I polysomal non- non -
I
c-myc
I O-actin
I 18 S -" - .
I -a
FIG. 2. RNA gel blot analyses of myoblast polysome density gradients from insulin-treated and control myoblasts. The myoblast cultures were treated with insulin 60 min before harvesting. Myoblast polysome gradients were fractionated into nonpolysomal and polysomal fractions as shown in Fig. 1. RNA was isolated from gradient fractions and separated on parallel agarose gels. The result- ing duplicate RNA gel blots were hybridized to the indicated probes. Representative gel profiles are shown; these are not all derived from the same experiment. This figure does not reflect the relative concen- trations of r-protein mRNA in myoblasts and insulin-treated myo- blasts (see Fig. 5) as different amounts of cells and/or probe are used in some cases. This figure only shows the distribution of mRNA in polysome gradients.
polysome preparations from both control and insulin-treated cultures are not degraded.
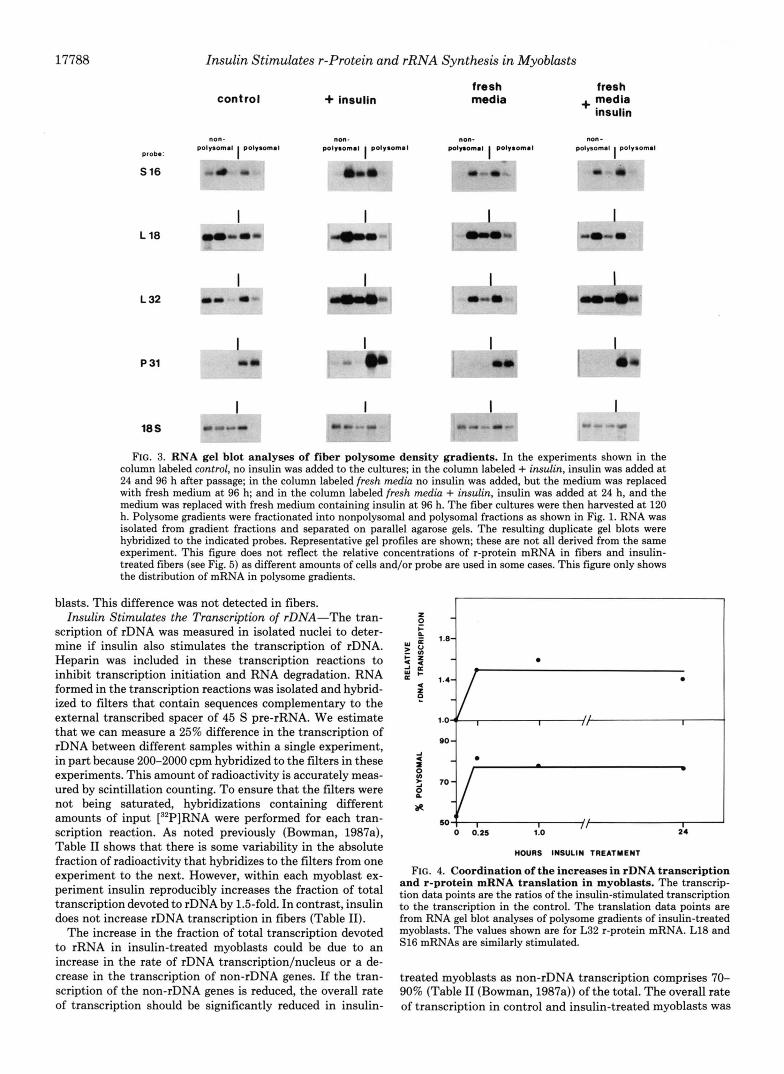
Insulin Does Not Affect the Translation of r-Protein mRNA in Fibers-Fig. 3 and Table I show that about 30-40% of the r-protein mRNAs are associated with polysomes in both con- trol fibers and insulin-treated fibers. In contrast greater than 95% of p31 mRNA is associated with polysomes in both control and insulin-treated fibers. Thus, there is no significant change in the translational efficiency of r-protein or p31 mRNAs in fibers.
One possible explanation for the lack of increase in the translation of r-protein mRNAs in fibers is that component(s) of the medium necessary for mediating the insulin response are metabolized, inactivated, or degraded during the 4 days the medium is left on the culture to allow the fibers to mature.
17787
TABLE I Polysome distribution
Autoradiographs, such as those shown in Figs. 2 and 3, were analyzed by densitometry. The percentage of each RNA located in the polysomal fractions is indicated below. The increase in the frac- tion of r-protein mRNAs associated with polysomes in insulin- treated myoblasts is statistically significant a t 0.05 level for S16 and L18 mRNAs and the 0.01 level for L32 mRNA as determined using the t test.
Probe Experiment Ratio of
1 2 3 to control ~~~$~~ insulin-treated
S16 Myoblasts Myoblasts
Fibers Fibers + insulin
Myoblasts Myoblasts
Fibers Fibers + insulin
Myoblasts Myoblasts
Fibers Fibers + insulin
Myoblasts Myoblasts
Fibers Fibers + insulin
Myoblasts Myoblasts
+ insulin
L18
+ insulin
L32
+ insulin
P3 1
+ insulin
c-myc
+ insulin &actin
Myoblasts Myoblasts
+ insulin 18 S rRNA
Myoblasts Myoblasts
Fibers + insulin
58 54 47 82 69 77
35 35 33 38 39 33
36 37 50 47 47 73
30 40 28 32 39 23
55 43 56 78 69 79
49 54 41 48 42 36
>95 >95 >95 >95 >95 >95
>95 >95 >95 >95 >95 >95
>95 >95 >95 >95 >95 >95
>95 >95 >95 >95 >95 >95
66 44 72 51 53 65
46 44 32 Fibers + insulin ND" 41 30
53 76
1.4 -
34 1.1 37
41 56
1.3 -
33 1.0 31
51 1.5 75
-
48 0.9 42
>95 1.0 >95
>95 1.0 >95
>95 1.0 >95
>95 1.0 >95
61 0.9 57
41 0.9 36
a Not determined.
This possibility was examined by adding fresh medium with and without insulin to fiber cultures 24 h prior to harvesting. Fig. 3 shows that insulin in the presence of fresh media still does not stimulate the translation of r-protein mRNAs. Con- sequently, the lack of response in fiber cultures is not due to a deficiency in the culture medium; rather, fibers and myo- blasts respond differently to insulin.
The Effect of Insulin on the Distribution of Ribosomes in Polysome Gradients-The fraction of ribosomes associated with polysomes was measured by hybridizing gradient frac- tions to a probe complementary to 18 S mRNA (Bowman et al., 1981). Figs. 2 and 3, and Table I show that the percentage of ribosomes in polysomes is higher in myoblasts than in fibers. This overall distribution is not affected by insulin in either myoblasts or fibers, suggesting that insulin does not stimulate a general recruitment of mRNA into polysomes. Furthermore, examination of polysome profiles (Fig. 1) indi- cates a slight difference in the ratio of 40 S to 60 S subunits in insulin-treated myoblasts as compared to control myo-
17788
probe:
S 16
L 18
L 32
P 31
18 S
Insulin Stimulates r-Protein and rRNA Synthesis in Myoblasts
control + insulin fresh media
fresh + media
insulin
I I r - .. . @b
I
FIG. 3. RNA gel blot analyses of fiber polysome density gradients. In the experiments shown in the column labeled control, no insulin was added to the cultures; in the column labeled + insulin, insulin was added a t 24 and 96 h after passage; in the column labeled fresh media no insulin was added, but the medium was replaced with fresh medium at 96 h; and in the column labeled fresh media + insulin, insulin was added a t 24 h, and the medium was replaced with fresh medium containing insulin a t 96 h. The fiber cultures were then harvested at 120 h. Polysome gradients were fractionated into nonpolysomal and polysomal fractions as shown in Fig. 1. RNA was isolated from gradient fractions and separated on parallel agarose gels. The resulting duplicate gel blots were hybridized to the indicated probes. Representative gel profiles are shown; these are not all derived from the same experiment. This figure does not reflect the relative concentrations of r-protein mRNA in fibers and insulin- treated fibers (see Fig. 5) as different amounts of cells and/or probe are used in some cases. This figure only shows the distribution of mRNA in polysome gradients.
blasts. This difference was not detected in fibers. Insulin Stimulates the Transcription of rDNA-The tran-
scription of rDNA was measured in isolated nuclei to deter- mine if insulin also stimulates the transcription of rDNA. Heparin was included in these transcription reactions to inhibit transcription initiation and RNA degradation. RNA formed in the transcription reactions was isolated and hybrid- ized to filters that contain sequences complementary to the external transcribed spacer of 45 S pre-rRNA. We estimate that we can measure a 25% difference in the transcription of rDNA between different samples within a single experiment, in part because 200-2000 cpm hybridized to the filters in these experiments. This amount of radioactivity is accurately meas- ured by scintillation counting. To ensure that the filters were not being saturated, hybridizations containing different amounts of input [32P]RNA were performed for each tran- scription reaction. As noted previously (Bowman, 1987a), Table I1 shows that there is some variability in the absolute fraction of radioactivity that hybridizes to the filters from one experiment to the next. However, within each myoblast ex- periment insulin reproducibly increases the fraction of total transcription devoted to rDNA by 1.5-fold. In contrast, insulin does not increase rDNA transcription in fibers (Table 11).
The increase in the fraction of total transcription devoted to rRNA in insulin-treated myoblasts could be due to an increase in the rate of rDNA transcription/nucleus or a de- crease in the transcription of non-rDNA genes. If the tran- scription of the non-rDNA genes is reduced, the overall rate of transcription should be significantly reduced in insulin-
MOURS INSULIN TREATMENT
FIG. 4. Coordination of the increases in rDNA transcription and r-protein mRNA translation in myoblasts. The transcrip- tion data points are the ratios of the insulin-stimulated transcription to the transcription in the control. The translation data points are from RNA gel blot analyses of polysome gradients of insulin-treated myoblasts. The values shown are for L32 r-protein mRNA. L18 and S16 mRNAs are similarly stimulated.
treated myoblasts as non-rDNA transcription comprises 70- 90% (Table I1 (Bowman, 1987a)) of the total. The overall rate of transcription in control and insulin-treated myoblasts was

Insulin Stimulates r-Protein and rRNA Synthesis in Myoblasts 17789
TABLE I1 rDNA transcription rates
The relative rate of rDNA transcription is expressed as the percentage of total input counts/min that hybridized to the p3 external transcribed spacer probe (Bowman et al., 1981). In each experiment shown below the transcription reactions were performed side by side using the same batch of [3ZP]UTP, filters, and hybridization and rinse solutions. Each hybridization reaction contained a pBR322 filter to monitor background hybridization which was always less than 10 cpm. It is estimated that rDNA transcription comprises 10-30% of the total based on the percent counts/min hybridized here and in Bowman, 1987a, the size of the probe and the size of 45 S pre-rRNA. The insulin-stimulated increase in rDNA transcription in myoblasts is statistically significant at the 0.01 level as determined using the t test.
Input Countsfmin Percent counts fmin hybridized hybridized
Ratio of insulin-treated to control
Experiment 1 Myoblasts 14,800
52,900
Myoblasts + insulin (15 min) 18,700 53,900
Myoblasts + insulin (60 min) 13,500 41,300
Myoblasts 9,230 25,300
Myoblasts + insulin (24 h) 7,140 24,400
Myoblasts 14,700 14,700
Myoblasts + insulin (15 min) 14,500 14,500
A. Fibers 17,100 30,200
Fibers + insulin 15,000 30,700
B. Fibers 15,700 30,600
Fibers + insulin 15,300 30,700
Fibers 17,800 43,800
Fibers + insulin 14,600 51,300
Experiment 2
Experiment 3
Experiment 4
Experiment 5
estimated from the incorporation of [32P]UTP into RNA/pg of DNA in isolated nuclei. These reactions contained 100 PM added unlabeled UTP which should be in excess of the endog- enous UTP, thereby negating any potential differences in the UTP pool of control and insulin-treated myoblasts. Under these conditions the total incorporation of [32P]UTP into RNA/pg of DNA is the same +15% (n = 2) in control and insulin-treated myoblasts suggesting non-rDNA transcription is not decreased (data not shown). Therefore, these results suggest that insulin does in fact stimulate the rate of rDNA transcription in myoblasts.
Temporal Coordination of rDNA Transcription and r-Pro- tein mRNA Translation-The transcription of rDNA and the translation of r-protein mRNA were measured at various times after the addition of insulin to myoblast cultures to determine if these increases are temporally coordinated. Fig. 4 shows that the maximal increase in both the transcription and translation is reached within 15 min of insulin addition. Furthermore, the magnitude of the increases is similar.
Insulin Does Not Increase the Steady State Levels of r- Protein mRNAs or the Growth Rate of Myoblasts-RNA gel blot analysis (Fig. 5) shows that the concentrations of the r-
259 1.8 1,120 2.1
545 2.9 1.7 1,940 3.6
-
425 3.1 1.6 1,260 3.0
216 2.3 549 2.2 238 3.3 735 3.0
327 2.2 409 2.8
431 3.0 503 3.5
453 2.7 714 2.4
373 2.5 916 3.0
346 68 1
2.2 2.2
382 704
2.5 1.1 2.3
325 1.8 2.5
2.3 1.6
-
1.1
1,080
331 810
0.9
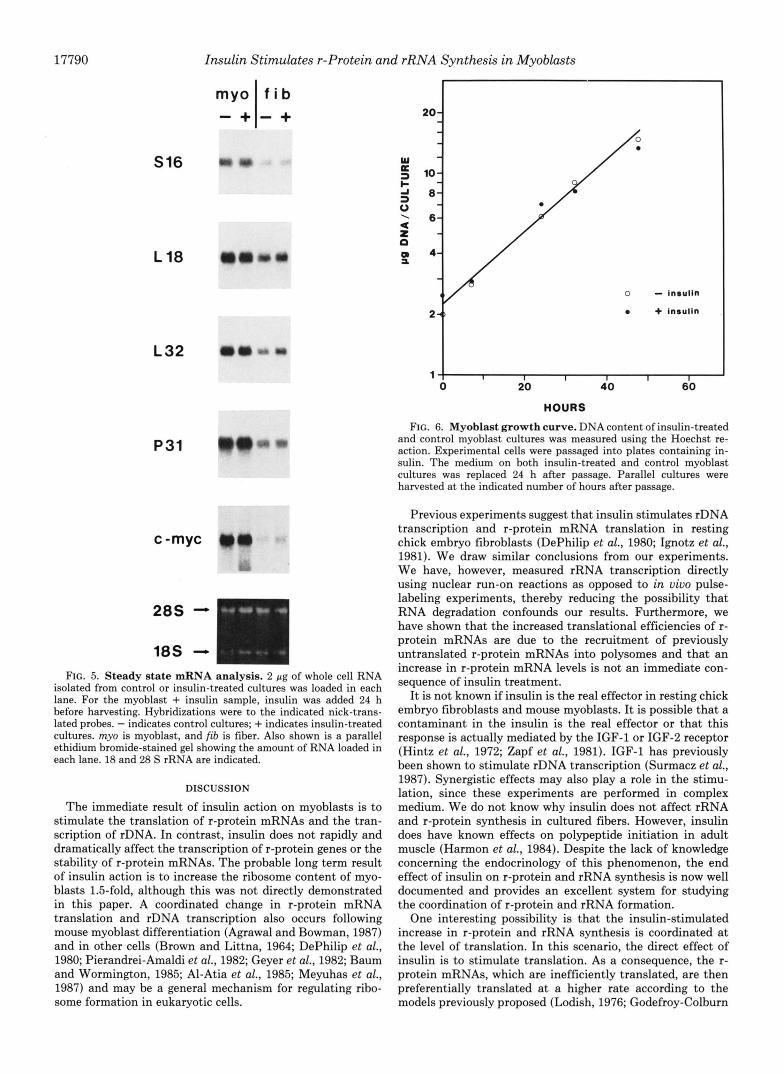
protein mRNAs as well as p31 and c-myc RNAs are unaffected by insulin treatment in both myoblasts and fibers. With respect to myoblasts, similar results were obtained regardless of whether RNA was isolated 2 or 24 h after insulin treatment. Fig. 5 also shows that the steady state levels of r-protein mRNAs decrease after myoblasts differentiate into fibers as observed previously (Agrawal and Bowman, 1987). To ensure that equal amounts of RNA were loaded in each lane, a parallel gel was stained with ethidium bromide (Fig. 5). These experiments indicate that insulin does not rapidly and dra- matically alter the transcription of r-protein genes or the stability of r-protein mRNAs because the steady state levels of r-protein mRNA are unchanged after 2 h of insulin treat- ment. Our experiments are, however, not sensitive enough to eliminate the possibility that small changes in r-protein mRNA levels occur slowly over time due to a small increase or decrease in the transcription of r-protein genes or the stability of r-protein mRNAs.
To determine if insulin stimulates myoblast cell division, the amount of DNA/culture was determined for control and insulin-treated cultures. Fig. 6 shows that insulin does not affect the 15-h myoblast cell division time.
- 1.4
- 1.3
17790
S16
L 18
L 3 2
P31
c -myc
Insulin Stimulates r-Protein and rRNA Synthesis in Myoblasts
myo - + - +
f i b
FIG. 5. Steady state mRNA analysis. 2 pg of whole cell RNA isolated from control or insulin-treated cultures was loaded in each lane. For the myoblast + insulin sample, insulin was added 24 h before harvesting. Hybridizations were to the indicated nick-trans- lated probes. - indicates control cultures; + indicates insulin-treated cultures. my0 is myoblast, and fib is fiber. Also shown is a parallel ethidium bromide-stained gel showing the amount of RNA loaded in each lane. 18 and 28 S rRNA are indicated.
DISCUSSION
The immediate result of insulin action on myoblasts is to stimulate the translation of r-protein mRNAs and the tran- scription of rDNA. In contrast, insulin does not rapidly and dramatically affect the transcription of r-protein genes or the stability of r-protein mRNAs. The probable long term result of insulin action is to increase the ribosome content of myo- blasts 1.5-fold, although this was not directly demonstrated in this paper. A coordinated change in r-protein mRNA translation and rDNA transcription also occurs following mouse myoblast differentiation (Agrawal and Bowman, 1987) and in other cells (Brown and Littna, 1964; DePhilip et al., 1980; Pierandrei-Amaldi et al., 1982; Geyer et al., 1982; Baum and Wormington, 1985; Al-Atia et al., 1985; Meyuhas et al., 1987) and may be a general mechanism for regulating ribo- some formation in eukaryotic cells.
*m - Insulln
1 0 20 40 60
HOURS
FIG. 6. Myoblast growth curve. DNA content of insulin-treated and control myoblast cultures was measured using the Hoechst re- action. Experimental cells were passaged into plates containing in- sulin. The medium on both insulin-treated and control myoblast cultures was replaced 24 h after passage. Parallel cultures were harvested at the indicated number of hours after passage.
Previous experiments suggest that insulin stimulates rDNA transcription and r-protein mRNA translation in resting chick embryo fibroblasts (DePhilip et al., 1980; Ignotz et al., 1981). We draw similar conclusions from our experiments. We have, however, measured rRNA transcription directly using nuclear run-on reactions as opposed to in vivo pulse- labeling experiments, thereby reducing the possibility that RNA degradation confounds our results. Furthermore, we have shown that the increased translational efficiencies of r- protein mRNAs are due to the recruitment of previously untranslated r-protein mRNAs into polysomes and that an increase in r-protein mRNA levels is not an immediate con- sequence of insulin treatment.
It is not known if insulin is the real effector in resting chick embryo fibroblasts and mouse myoblasts. It is possible that a contaminant in the insulin is the real effector or that this response is actually mediated by the IGF-1 or IGF-2 receptor (Hintz et al., 1972; Zapf et al., 1981). IGF-1 has previously been shown to stimulate rDNA transcription (Surmacz et al., 1987). Synergistic effects may also play a role in the stimu- lation, since these experiments are performed in complex medium. We do not know why insulin does not affect rRNA and r-protein synthesis in cultured fibers. However, insulin does have known effects on polypeptide initiation in adult muscle (Harmon et al., 1984). Despite the lack of knowledge concerning the endocrinology of this phenomenon, the end effect of insulin on r-protein and rRNA synthesis is now well documented and provides an excellent system for studying the coordination of r-protein and rRNA formation.
One interesting possibility is that the insulin-stimulated increase in r-protein and rRNA synthesis is coordinated at the level of translation. In this scenario, the direct effect of insulin is to stimulate translation. As a consequence, the r- protein mRNAs, which are inefficiently translated, are then preferentially translated at a higher rate according to the models previously proposed (Lodish, 1976; Godefroy-Colburn

Insulin Stimulates r-Protein and rRNA Synthesis in Myoblasts 17791
and Thach, 1981). If rDNA transcription is controlled by a short-lived factor whose mRNA is also inefficiently trans- lated, a general increase in protein synthesis would also preferentially increase the synthesis of this factor, thereby stimulating rDNA transcription. Previous experiments indi- cate that rDNA transcription in some cases is regulated by a short-lived factor (Mishima et al., 1979; Gokal et al., 1986; Tower and Sollner-Webb, 1987). However, it is clear that this is not the only mechanism for controlling rDNA transcription. For instance, in rat fibers the r-proteins are translated quite efficiently yet rDNA transcription is depressed (Jacobs et al., 1985).
Acknowledgment-We thank Marga Costan for her excellent tech- nical assistance.
REFERENCES
Abelson. H. T.. Johnson, L. F., Penman, S., and Green, H. (1974) Cell 1, 161-165
. .
Abovich. N.. Gritz. L.. Tune. L.. and Rosbach. M. (1985) Mol. Cell. I ", I
Biol. 6,3429-3435
4875
, . .
Agrawal, M. G., and Bowman, L. H. (1987) J. Biol. Chem. 262,4868-
Al-Atia, G. R., Fmscoloni, P., and Jacobs-Lorena, M. (1985) Biochem-
Baum, E. Z., and Wormington, W. M. (1985) Dew. Biol. 111, 488-
Baum, E. Z., Hyman, L. E., and Wormington, W. M. (1988) Deu.
Bell, J., Neilson, L., and Pellegrini, M. (1988) Mol. Cell. Biol. 8, 91-
Bowman, L. H. (1987a) Dew. Biol. 119,152-163 Bowman, L. H. (198713) Mol. Cell. Biol. 7, 4464-4471 Bowman, L. H., and Emerson, C. P., Jr. (1977) Cell 10,587-596 Bowman, L. H., Rabin, B., and Schlessinger, D. (1981) Nucleic Acids
Bozzoni, I., Fragapane, P., Annesi, F., Pierandrei-Amaldi, P., Amaldi,
Brown, D. D., and Littna, E. (1964) J. Mol. Biol. 8,669-672 Clissold, R., and Cole, R. J. (1973) Exp. Cell Res. 80, 159-169 Cooper, H. L. (1973) J. Cell Biol. 59,250-254 Craig, N., and Perry, R. P. (1971) Nut. New Biol. 2 2 9 , 75-80 Dabeva, M. D., Post-Beittenmiller, M. A., and Warner, J. R. (1986)
DePhilip, R. M., Rudert, W. A., and Lieberman, I. (1980) Biochem-
ElBaradi, T. T. A. L., van der Sande, C. A. F. M., Mager, W. H., Raue, H. A., and Planta, R. J. (1986) Curr. Genet. 10,733-739
Faliks, D., and Meyuhas, 0. (1982) Nucleic Acids Res. 10 , 789-801 Geyer, P. K., Meyuhas, O., Perry, R. P., and Johnson, L. F. (1982)
Godefroy-Colburn, T., and Thach, R. E. (1981) J. Biol. Chem. 2 5 6 ,
Gokal, P. K., Cavanaugh, A. H., and Thompson, E. A. (1986) J. Biol.
Gospodarowicz, D., Chang, J., Lui, G., Baird, A., and Bohlent, P.
istry 24,5798-5803
498
Biol. 126,141-149
95
Res. 9,4951-4966
F., and Becarri, E. (1984) J. Mol. Biol. 180 , 987-1005
Proc. Natl. Acad. Sci. U. S. A. 8 3 , 5854-5857
istry 19,1662-1669
Mol. Cell. Biol. 2 , 685-693
11762-11773
Chem. 161 , 2536-2541
(1984) Proc. Natl. Acad. Sci. U. S. A. 8 1 , 6963-6967 Harmon. C. S.. Proud, C. G., and Pain, V. M. (1984) Biochem. J.
223,687-696 Hauschka, S. D., Linkhart, T. A., Clegg, C. H., and Merrill, G. M.
(1979) in Muscle Regeneration (Mauro, A., ed) pp. 211-322, Raven . . - Press, New York
..
Hintz. R. L.. Clemmons. D. R.. Underwood. L. E.. and Van Wvk. J. J. (1972) Proc. Natl. Acad. Sei. U. S. A. 69, 2351-2353
man, I. (1981) Biochemistry 20,2550-2558
- .
Ignotz, G. G., Hokari, S., DePhilip, R. M., Tsukada, K., and Lieber-
Jacobs, F. A,, Bird, R. C., and Sells, B. H. (1985) Eur. J. Biochem.
Krauter, K. S., Soeiro, R., and Nadal-Ginard, R. (1979) J. Mol. Biol.
Krauter. K. S., Soeiro. R., and Nadal-Ginard, R. (1980) J. Mol. Biol.
150,255-263
134,727-741
142 , i45-159 . .
Labarca. C.. and Paitzen. K. (1980) Anal. Biochem. 102. 344-352 LaMarca, M. J., and-Wassarman,'P. M. (1984) Dew. Biol. 102 , 525-
Linkhart, T. A.. Clem C. H.. and Hauschka, S. D. (1981) Deu. Biol. 530
. ". 86,19130
Lodish. H. F. (1976) Annu. Reu. Biochem. 45,39-72 McKnight, G. S., and Palmiter, R. D. (1979) J. Biol. Chem. 2 5 4 ,
McMaster, G. K., and Carmichael, G. G. (1977) Proc. Natl. Acad. Sci.
Meyuhas, O., and Perry, R. P. (1980) Gene (Anst . ) 10,113-129 Meyuhas, O., Thompson, E. A., Jr., and Perry, R. P. (1987) Mol. Cell.
Mishima, Y., Matsui, T., and Muramatsu, M. (1979) J. Biochem.
Nomura, M., Gourse, R., and Baughman, G. (1984) Annu. Rev.
Pearson, N. J., Fried, H. M., and Warner, J. R. (1982) Cell 29,347-
Pierandrei-Amaldi, P., Campioni, N., Beccari, E., Bozzoni, I., and
Pierandrei-Amaldi, P., Becarri, E., Bozzoni, I., and Amaldi, F. (1985)
Pierandrei-Amaldi, P., Bozzoni, I., and Cardinali, B. (1988) Genes
Ponte, P., Gunning, P.. Blau. H.. and Kedes, L. (1983) Mol. Cell. Biol.
, ,
9050-9058
U. S. A. 74,4835-4838
Biol. 7 , 2691-2699
(Tokyo) 85,807-818
Biochem. 53, 75-117
355
Amaldi, F. (1982) Cell 3 0 , 163-171
Cell 42,317-323
Dew. 2,23-31 -. .
3, im-1791 . . . . .
Rhoads. D. G.. and Roufa. D. J. (1987) Mol. Cell. Biol. 7. 3767-3774 Schmidt, T., Chen, P. S.; and Pellegkni, M. (1985) J. 'Biol. Chm.
Stanton, L. W., Watt, R., and Marcu, K. B. (1983) Nature 303,401-
Surmacz, E., Kaczmarek, L., Ronning, O., and Baserga, R. (1987)
Theodor, L., Peleg, D., and Meyuhas, 0. (1985) Biochim. Biophys.
Tower, J., and Sollner-Webb, B. (1987) Cell 50,873-883 Warner, J. T. (1977) J. Mol. Biol. 115 , 315-333 Warner, J. R., Mitra, G., Schwindinger, W. F., Studeny, M., and
Wolf, S., Sameshima, M., Liebhaber, S. A., and Schlessinger, D.
Zapf, J., Froesch, E. R., and Humbel, R. E. (1981) Curr. Top. Cell.
160,7645-7650
406
Mol. Cell. Biol. 7, 657-663
Acta 826,137-146
Fried, H. M. (1985) Mol. Cell. Biol. 5 , 1512-1521
(1980) Biochemistry 19 , 3484-3490
Regul. 19,257-309