insertional mutation on mouse chromosome 18 with vestibular and
TRANSCRIPT

Copyright 0 1994 by the Genetics Society of America
Insertional Mutation on Mouse Chromosome 18 With Vestibular and Craniofacial Abnormalities
Chao-Nan Ting,**’.‘ David Kohrman,**‘ Daniel L. Burgess,* Ann Boyle? Richard A. Altschuler,* G. Gholizadeh? Linda C. Samuel~on,*’~ Wonhee Jang* and Miriam H. Meisler*
*Department of Human Genetics, University of Michagan, Ann Arbor, Michigan 48109-0618, TKresge Hearing Research Institute, University of Michigan, Ann Arbor, Michigan 48109-0506 and *Department of Genetics, Yale University,
New Haven, Connecticut 06510 Manuscript received June 25, 1993
Accepted for publication September 28, 1993
ABSTRACT A dominant mutation was generated in transgenic mice as a consequence of insertional mutation.
Heterozygous mice from transgenic line 9257 (Tg?257) are hyperactive with bidirectional circling behavior and have a distinctive facial appearance due to hypoplasia of the nasal bone. Morphological analysis of the inner ear revealed asymmetric abnormalities of the horizontal canal and flattening or invagination of the crista ampullaris, which can account for the circling behavior. The sensory epithelium appeared to be normal. The transgene insertion site was localized by in situ hybridization to the B 1 band of mouse chromosome 18. Genetic mapping in an interspecific backcross demonstrated the gene order centrome~e-Tg?~’~-8.8 f 3.4-Grl-1, Egr-1, Fgf-1, Apc--14.7 f 4.3-Pdgfr. The phenotype and the mapping data suggest that the transgene may be inserted at the Twirler locus. Homozygosity for the transgene results in prenatal lethality, but compound heterozygotes carrying the Tw allele and the transgene are viable. The function of the closely linked ataxia locus is not disrupted by the transgene insertion. This insertional mutant will provide molecular access to genes located in the Twirler region of mouse chromosome 18.
M OLECULAR analysis of mutant mice has be- come an important method for identifying
genes involved in mammalian development (REITH and BERNSTEIN 1991). The roles of growth factors, receptors and transcriptional regulators in normal development have been revealed by analysis of spon- taneous mutants such as undulated (BALLING et al. 1 988), small eye (HILL et al. 199 l), Steel (reviewed in WITTE 1990), Patch (STEPHENSON et al. 199 l), short ear (KINGSLEY et al. 1992) and Snell dwarf (CAMPER et al. 1990; LI et al. 1990). The molecular defects in these spontaneous mutants were identified by posi- tional cloning and by their genetic proximity to can- didate genes. Developmental mutations caused by in- sertion of foreign DNA provide another mode of access: by use of the inserted sequence as a molecular probe for direct isolation of the mutated gene (CON-
This approach has been successfully applied to the isolation of novel genes required for development from transgenic insertional mutants (WOYCHIK et al. 1990; WEIHER et al. 1990; HODCKINSON et al. 1993; HUGHES et al. 1993).
The Twirler mutation, which produces craniofacial abnormalities, was described by LYON (1958). Mice
STANTINI et al. 1989; GRIDLEY 199 1 ; MEISLER 1992).
MC 5041, Chicago, Illinois 60637. ’ Present address: Division of Cardiology, University of Chicago, 0509
‘ The first two authors contributed equally to this work. ’ Present address: Department of Physiology, University of Michigan, Ann Arbor, Michigan 48109-0622.
Genetics 136: 247-254 (January, 1994)
heterozygous for the spontaneous allele, Tw, have a complex phenotype that includes circling behavior, hyperactivity and early onset obesity. The circling behavior is the consequence of abnormal structural development of the crista ampullaris in the inner ear. Tw/Tw homozygotes are born with cleft palate f cleft lip and do not survive. The Twirler locus has been mapped to proximal chromosome 18 (LYON 1958; LANE et al. 198 1). In this paper we describe a trans- gene induced insertional mutation that exhibits many of the phenotypic features of Twirler mice. The trans- gene insertion may have generated a new allele of Twirler, and thus provide molecular access to this developmentally important gene. Alternatively, Twirler and TZz5’ may represent mutations in two closely linked genes that are both required for cranio- facial development.
MATERIALS AND METHODS
Animals: The origin of transgenic line 9257 was previ- ously described (TING et al. 1992). A 25-kb ApaI/MluI fragment containing a human salivary amylase gene was microinjected into fertilized mouse eggs from matings be- tween (C57BL/6J X CSH)FI individuals. Transgenic animals were identified by PCR of tail DNA using the human amylase primers HTA-C and HTA-N (TING et al. 1992). The line was maintained by crossing with strain C57BL/6J.
C57BL/6J and SPRET/Ei mice were obtained from the Jackson Laboratory, Bar Harbor, Maine. Tw/+ mice carry- ing the original Twirler mutation were kindly provided by MARY LYON, Medical Research Council, Harwell. Hetero-

248 C.-N. Ting et al.
zygous axJ/+ mice carrying the ataxia allele were obtained from GLENN RADICE, Massachusetts Institute of Technol- ogy, Cambridge, Mass.
Histology: Seven heterozygous transgenic mice and five C57BL/6J mice were heavily anesthetized with chloral hy- drate and perfused through the heart with phosphate buffer followed by 4% paraformaldehyde in phosphate buffer. Intrascalar perfusion of the same fixative through the round window was followed by immersion in fixative for 16-20 hr, one rinse with phosphate buffer, intrascalar osmication for 1 hr with 1% osmium in phosphate buffer through the round window and another buffer rinse. Inner ears from four transgenic and three normal mice were dissected and drilled to expose the semicircular canals and assessed under a Wild dissection microscope. All inner ears (dissected and nondissected) were decalcified for 1 week in 2% EDTA, dehydrated through a graded series of alcohols and em- bedded in plastic (EmGed 81 2). Six-micron sections were cut through the vestibular and auditory portions of the inner ear and treated with Richardson’s stain. Sections were assessed under normal and differential interference contrast optics on a Leitz photomicroscope.
Pulsed field gel electrophoresis: Spleen DNA was pre- pared in agarose blocks and digested with restriction en- zymes as described (SAMBROOK, FRITSCH and MANIATIS 1989). Digested blocks were electrophoresed on 1 % gels at 200 V for 24 hr, using a BioRad CHEF DR I1 apparatus with 40-100 sec pulse ramp. Gels were stained in 0.5 rg/ ml ethidium bromide and destained in water. Fragments were transferred to a nylon filter and hybridized with radi- olabeled cosmid clone N2 (GUMUCIO et al. 1988). In situ hybridization: Metaphase chromosomes were pre-
pared from spleen cells from a heterozygous transgenic animal after culture for 2 days in the presence of concana- valin A, as previously described (BOYLE, B A L L A R D ~ ~ ~ WARD 1990). Cosmid N2 (GUMUCIO et al. 1988) was labeled by nick translation with biotin-1 1-dUTP (BRIGATI et al. 1983). The L1 banding probe KS13A (FANNING 1983) was labeled with digoxigenin-1 1-dUTP. Fluorescence in si tu hybridiza- tion was carried out exactly as described (BOYLE et al. 1992). All image acquisition and processing was done on a Macin- tosh IIci computer. Images were merged and 24-bit pseu- docolored using custom software developed in DAVID WARD’S laboratory (T. RAND, unpublished data). For addi- tional details on image generation see BOYLE et al. (1992). Images were photographed directly from the computer screen.
Genetic mapping: To generate the interspecific back- cross, heterozygous transgenic mice were crossed with SPRET/Ei and female transgenic offspring were back- crossed to C57BL/6J. This backcross has been used for mapping loci on chromosomes 3 and 6 (BAIN et al. 1993; BARROW et al. 1993). DNA was prepared from newborn offspring by the salting out method of MILLER, DYKES and POLESKY (1988). Southern blotting was carried out as pre- viously described (BARROW et al. 1993). For resolution of the 12-kb and 14-kb SphI fragments detected with the flanking probe, electrophoresis was carried out on 0.4% agarose gels (PFGE Agarose, Boehringer Mannheim).
Restriction fragment length polymorphisms were identi- fied by Southern blot analysis of genomic DNA from strains C57BL/6J, C3H/HeJ and SPRET/Ei after digestion with a variety of restriction endonucleases. Blots were hybridized with the following isolated plasmid inserts: a 1.3-kb EcoRI fragment from mouse Apc cDNA clone 5 1 B (SU et al. 1992); an 0.7-kb EcoRI-Hind111 fragment from mouse Egr-1 cDNA clone pRSV3.1 (SUKHATME et al. 1988); an 0.475-kb EcoRI- XbaI fragment from mouse Fgf-1 cDNA clone fgfa(a1) (HE- BERT et al. 1990); a 1-kb Hind111 fragment from rat Grl-1
TABLE 1
Dominant inheritance of circling behavior in line 9257
Transgenic offspring
Nontransgenic offspring
Cross No. % circling No. % circling
1. Tgl+ X +/+ 53 77 86 0 2. Tgl+ X Tg/+ 23 78 5 0
Transgenic mice were crossed with C57BL16J. Inheritance of the transgene was assayed by PCR of genomic DNA.
cDNA clone pRM16 (MIESFELD et al. 1984); and a 1 .2-kb HincII fragment from mouse Pdgfrb cDNA clone pGR102 (YARDEN et al. 1986).
Cloning of flanking DNA: Genomic DNA from a trans- genic heterozygote was digested with MboI, size selected by sucrose gradient centrifugation, and cloned into a cosmid vector. Nitrocellulose filter replicas of the library were screened by hybridization (SAMBROOK, Fritsch and MANIA- TIS 1989) using two transgene specific probes. Sixty-eight transgene-positive clones were probed with labeled mouse DNA to identify clones that also contain mouse repeated sequences; Cosmid AT 1 was found to contain approximately 15 kb of mouse DNA and 20 kb of transgene DNA.
RESULTS
Behavioral abnormalities in transgenic line 9257: Transgenic line 9257 was generated by microinjection of fertilized mouse eggs with a human genomic frag- ment (TING et al. 1992). The founder animal was crossed with strain C57BL/6J to generate the N1 generation. Three N 1 individuals demonstrated be- havioral abnormalities including bidirectional circling and hyperactivity. This phenotype has persisted through nine generations of breeding. In a swimming test, affected mice are unable to remain at the surface of the water for more than 1-2 min, while normal mice can d o so for at least 5 min.
More than 20 lines expressing similar constructs have been studied [TING et al. ( 1 992) and unpublished data]. None of the other lines exhibit behavioral ab- normalities, indicating that the circling phenotype is a characteristic of the specific insertion site in line 9257.
Autosomal dominant inheritance of circling be- havior: The data are consistent with autosomal dom- inant inheritance of the circling phenotype with ap- proximately 80% penetrance (Table 1). In the cross between heterozygous transgenic mice and C57BL/ 6J, the proportion of transgenic offspring was signifi- cantly lower than 50% (53/139, P C 0.01). However, in three other crosses described below, with SPRET/ Ei, ax/+ and Tw/+ mice, the proportion of transgenic offspring was close to 50%, suggesting that the viabil- ity of Tg/+ mice is influenced by genetic background.
Recessive lethality of the transgene mutation: Cos- mid clone A T 1, containing the junction between the transgene and flanking mouse DNA, was isolated from a library of heterozygous DNA by hybridization with

Mouse Craniofacial Mutation 249
+ @){
23 -
f o m
9.4 -
6.6 -
FIGURE 1.-Detection of a transgene junction fragment by Southern blotting. Genomic DNA was digested with SphI and fragments were separated by electrophoresis on an 0.4% agarose gel. Filters were probed with a 1-kb subclone (2B2) of cosmid A T l . Left, Hind111 fragments from bacteriophage X (kb): arrow, 16 kb transgene junction fragment.
both transgene and mouse genomic DNA probes. Subclones of AT1 containing nonrepetitive mouse DNA were used for RFLP analysis of the [(Tg/+ X SPRET/Ei)F, X C57BL/6J] backcross described be- low (not shown). The lack of recombination between AT1 and the transgene (0/46) is consistent with der- ivation of the mouse DNA in AT1 from the transgene insertion site. A 1-kb subclone of AT1 hybridized with a 12-kb SphI fragment in genomic DNA from strain C57BL/6J and an additional 14-kb fragment in heterozygous transgenic mice (Figure 1, arrow). The transgene specific 14-kb SphI fragment is contained within clone ATl .
T o determine whether transgenic homozygotes are viable, 5 litters containing 33 fetuses were collected from the cross Tg/+ X Tg/+ at embryonic day 17.5. N o evidence of resorbed embryos was observed. Gen- otypes were determined by analysis of the Sphl poly- morphism. We detected 8 individuals homozygous for the 12-kb C57BL/6J allele, 25 heterozygotes with 12- kb and 14-kb hybridizing fragments, and 0 homozy- gotes for the 14-kb transgene allele. The data are inconsistent with the 25% frequency of homozygotes that is expected for a nonlethal mutation (x’ = 16.5, P < 0.00 1). The observed proportion of heterozygotes and homozygotes does not differ significantly from the 2:l ratio expected if the transgenic mutation is a recessive lethal (x2 = 1.23, P > 0.25).
Abnormalities of the semicircular canals in trans- genic mice: The bony labyrinth of the inner ear was dissected from four transgenic animals and three nor- mal C57BL/6J animals. A dramatic reduction of the horizontal canal was evident in all the transgenic ani- mals (Figure 2, A and B). These four samples and additional nondissected inner ears from three trans- genic and two normal mice were assessed with 6-pm plastic sections through the vestibular and auditory sensory epithelium. Sections through the vestibular apparatus showed normal utricle and sacculi with nor-
mal maculae and otoliths in all of the transgenic animals. Abnormalities were evident in the crista am- pullaris, particularly of the horizontal canal. The sen- sory epithelium of the crista ampullaris normally has a convex shape, overlying an evagination (Figure 2C). The cristae of the horizontal canal of the seven trans- genic animals lacked the usual evagination. Instead, the epithelium was either flattened or invaginated (Figure 2, D-F). In the posterior and anterior canals, flattening was only occasionally observed and invagi- nation was never seen. Despite the abnormal appear- ance of the crista, the sensory epithelium including the vestibular hair cells was normal (Figure 2D). The overlying cupula was sometimes underdeveloped and in other cases appeared normal. The flattening of the crista and underdevelopment of the horizontal canal are sufficient to account for the circling behavior of the transgenic mice.
Auditory sensory epithelium and general anatomy of the cochlea appeared normal. The presence of a normal Preyer’s startle reflex suggested that the cir- cling mice are not deaf. This was confirmed by meas- urement of auditory brain stem responses, which were normal (data not shown). Brain histology did not reveal any obvious neuroanatomical abnormalities (R. Albin, unpublished data).
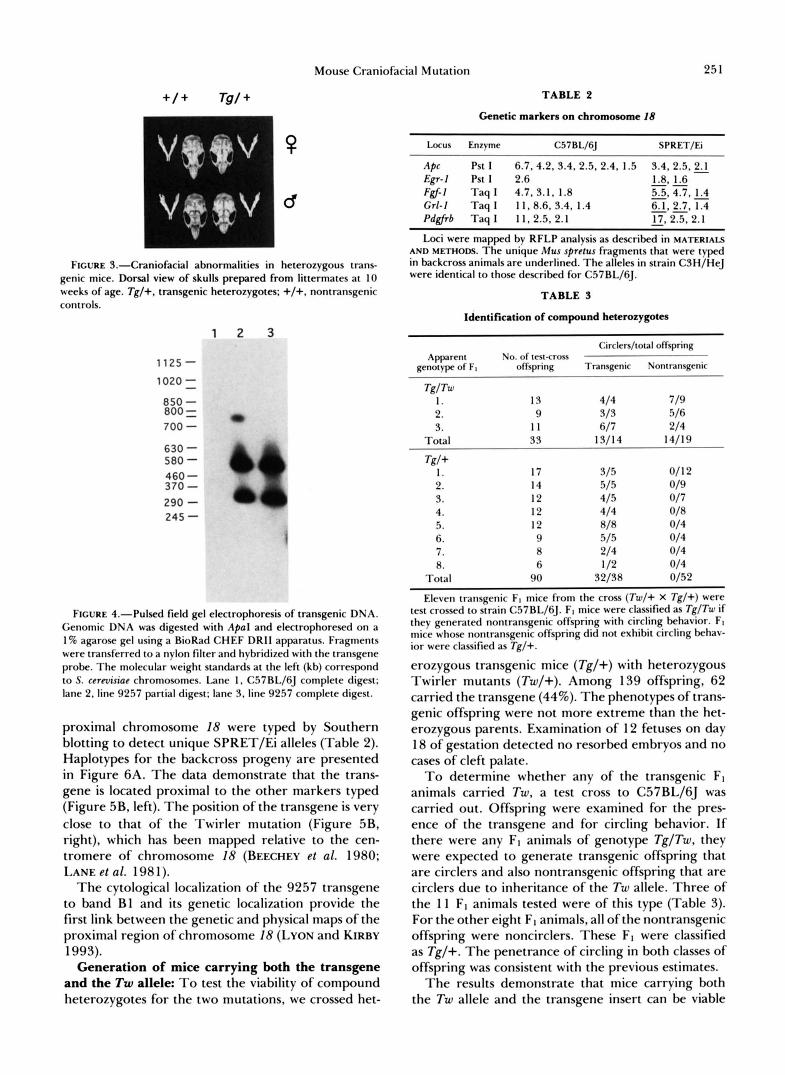
Craniofacial abnormalities: Transgenic mice can be recognized by their abnormal facial appearance resulting from marked hypoplasia of the nasal bones (Figure 3). The length of the nasal bones in transgenic animals is approximately 75% that of nontransgenic littermates, and overall skull length is reduced by 10%. Abnormalities of the maxilla, premaxilla, nasal septum, turbinates and frontal bones are also present.
Structure of the transgene insert: The microin- jected transgene in line 9257 was a linear 25-kb hu- man ApaI/MZuI genomic fragment (TING et aZ. 1992). T o analyze the structure of the insert, which segre- gates as a single locus, genomic DNA was prepared from transgenic mice and analyzed on standard South- ern blots. Comparison of the hybridization intensity of transgenic DNA with copy number standards indi- cated that the transgene was present in more than 20 copies (data not shown). High molecular weight DNA was prepared and digested with ApaI, which does not cut within the transgene. The resulting fragments were separated by pulsed field gel electrophoresis and hybridized with a transgene-specific probe (Figure 4). No hybridization was observed with nontransgenic C57BL/6J DNA (lane 1). Intensely hybridizing frag- ments of 280 kb and 500 kb were detected in trans- genic DNA (lanes 2 and 3). A weakly hybridizing fragment of 800 kb was also generated under partial digestion conditions (lane 2). The data are consistent with a tandem array containing between 20 and 32 copies of the full length transgene. The internal ApaI site may have been generated by a single “head-to-

250 C.-N. Ting et al.
E
Cochlea
C
FIGURE 2.-Anatomical assessment of semicircular canals. (A) Exposed semicircular canals from transgenic mice (left and right) and normal littermate (center). The horizontal canal (H) is missing from the transgenic mice, while anterior (A) and posterior (P) canals have a normal appearance. (B) Diagrammatic representation of semicircular canals. (C) Cross section through the crista ampullaris of the horizontal canal of a normal animal showing the characteristic evagination. (D) Cross section through the crista ampullaris of the horizontal canal from a transgenic animal. (E, F) Sections through the crista ampullaris of the semicircular canal from a second transgenic animal, showing invagination of the crista (arrow). Magnification lOOX to 300X.
head” alignment of transgenes within a larger “head- to-tail” array, or by incorporation of exogenous DNA within the array. In situ hybridization of the transgene to the B1
region of chromosome 28: To determine the chro- mosomal location of the insert, metaphase chromo- somes were prepared from cultured spleen cells and hybridized with a biotin-labeled transgene probe. The hybridization solution also contained a digoxygenin- labeled L1 repetitive sequence probe that generates a unique Giemsa-like banding pattern for each mouse chromosome (BOYLE, BALLARD and WARD 1990). Flu- orescent visualization of the probes demonstrated the presence of the transgene insert in the B1 region of
mouse chromosome 18 (Figure 5). A total of 57 met- aphase spreads were examined and 54 provided a strong signal on chromosome 18. (The other three metaphases were partial spreads.) Fourteen metaphase spreads were imaged and used to make the band assignment.
Linkage to genetic markers in the proximal region of chromosome 18: To locate the transgene insert on the meiotic map of chromosome 18, an interspecific backcross was generated by crossing mice from line 9257 with strain SPRET/Ei. Transgenic female F1 animals were then mated to strain C57BL/6J. Ge- nomic DNA from backcross offspring was analyzed by PCR to identify transgenic mice. Five genes on
Mouse Craniofacial Mutation 25 1
TABLE 4
Genetic markers on chromosome 18
Locus Enzyme C57BLl6J SPRET/Ei
+/+ Tg/+
?
d
FIGURE 3.-Craniofacial abnormalities in heterozygous trans- genic mice. D o r s a l view of skulls prepared from littermates at 10 weeks of age. Tg/+, transgenic heterozygotes: +/+, nontransgenic controls.
1 2 3
1125-
1020 - 850 - 700 - 630 - 580 - 460 - 290 - 245 -
- 800 =
370 -
FIGURE 4.-Pulsed field gel electrophoresis of transgenic DNA. Genomic DNA was digested with Apal and electrophoresed on a 1% agarose gel using a BioRad CHEF DRll apparatus. Fragments were transferred to a nylon filter and hybridized with the transgene probe. The molecular weight standards at the left (kb) correspond to S. cerrvizicre chromosomes. Lane 1, C57BL/6J complete digest; lane 2. line 9257 partial digest: lane 3. line 9257 complete digest.
proximal chromosome 18 were typed by Southern blotting to detect unique SPRET/Ei alleles (Table 2). Haplotypes for the backcross progeny are presented in Figure 6A. The data demonstrate that the trans- gene is located proximal to the other markers typed (Figure 5B, left). The position of the transgene is very close to that of the Twirler mutation (Figure 5B, right), which has been mapped relative to the cen- tromere of chromosome I8 (BEECHEY et al. 1980; LANE et al. 198 1).
The cytological locali7ation of the 9257 transgene to band B1 and its genetic locali7ation provide the first link between the genetic and physical maps of the proximal region of chromosome 18 (LYON and KIRBY
Generation of mice carrying both the transgene and the Tw allele: To test the viability of compound heterozygotes for the two mutations, we crossed het-
1993).
Apc Pst I 6.7.4.2. 3.4, 2.5. 2.4. 1.5 3.4. 2.5.2.1 1.8, 1.6
Fgf-I Taq I 4.7, 3.1. 1.8 5.54.7.1.4 Grl-1 Taq I 11.8.6. 3.4. 1.4 6.1, 2.7. 1.4 Pdgfrb Taq I 11.2.5, 2.1
Loci were mapped by RFLP analysis as described in MATERIALS AND METHODS. The unique Mw sprrrus fragments that were typed in backcross animals are underlined. The alleles in strain CSH/HeJ were identical to those described for C57Bl./fiJ.
&-I Pst I 2.6 -
"
- "
- 17.2.5.2.1
TABLE 3
Identification of compound heterozygotes
Circlcn/total ofTspring
genotype of F, offspring Transgenic Nonrransgenic Apparent No. of test-cross
TglTw I . 13 414 719 2. 9 313 516 3. I 1 617 214
Total 33 1311 4 14/19
Tg/+ 1. 17 315 011 2 2. 14 515 019 3. 12 415 017 4. 12 414 018 n. 12 818 014
7. R 214 014 8. 6 112 014
6. 9 515 014
Total 90 32/38 0152
Eleven transgenic F, mice from the cross (Tw/+ X Tg/+) were test crossed to strain C57BL/6J. F, mice were classified as TglTw if they generated nontransgenic offspring with circling behavior. FI mice whose nontransgenic offspring did not exhibit circling behav- ior were classified as Tg/+.
erozygous transgenic mice (Tg/+) with heterozygous Twirler mutants (Tw/+). Among 139 offspring, 62 carried the transgene (44%). The phenotypes of trans- genic offspring were not more extreme than the het- erozygous parents. Examination of 12 fetuses on day 18 of gestation detected no resorbed embryos and no cases of cleft palate.
To determine whether any of the transgenic F1 animals carried Tw, a test cross to C57BL/6J was carried out. Offspring were examined for the pres- ence of the transgene and for circling behavior. If there were any FI animals of genotype Tg/Tw, they were expected to generate transgenic offspring that are circlers and also nontransgenic offspring that are circlers due to inheritance of the Tw allele. Three of the 11 F1 animals tested were of this type (Table 3). For the other eight F1 animals, all of the nontransgenic offspring were noncirclers. These FI were classified as Tg/+. The penetrance of circling in both classes of offspring was consistent with the previous estimates.
The results demonstrate that mice carrying both the Tw allele and the transgene insert can be viable

252 C.-N. Ting et al.
FIGURE 5.-ln situ hybridization to metaphase chromosomes lo- calizes the transgenic insert to band B1 of nlouse chromosome 18. Metaphase chromosomes from a transgenic heterozygote were pre- pared from cultured spleen cells. Chromosonles were hvbridi7ed with a biotin-labeled 11uma11 amylase transgene probe and a digos- ygenin-labeled L I repeat probe, as described in MATERIALS AND
METHODS. The biotinvlated probe was detected wi th rhodamine- avidin. The digoxygenin probe $vas detected with FIT(:-laheletl antidigoxigenin Fab fi-agment (Boehringel- Mannheim). Chromo- sonles were counterstained wi th DAI'I. The DAPl and L1 images are displayed i n green and the transgene hybridiration signal is red. As indicated by the partial karyotype at the top of the figure. the chromosomes c o n t a i ~ ~ a single insert of~hurnan D N A that is l o c a l i d to band B1 of nwuse chron1osome 18.
and fertile. The number of offspring analyzed is not sufficient to determine whether viability of the com- pound heterozygotes is reduced.
Complementation of the ataxia mutation: Ataxia is a recessive mutation causing progressive paralysis that maps approximately 0.5 cM from the Twirler locus, with a reported recombination frequency of 6/ 1206 (LYON 1955, 1958, 1975). A complementation test was carried out by mating ax/+ females with Tg/+males and scoring progeny for the ataxia phe- notype. Paralysis of the hind limbs in axlax mice is first noticeable at 2-3 weeks of age and progresses to near immobility at 1-2 months (LYON 1955). We examined 47 progeny, including 24 transgenic ani- mals. None of the animals exhibited any sign of pa- ralysis when observed for a minimum of 2 months after birth. This complementation of the ataxia phe- notype by the transgenic chromosome demonstrates that the transgene does not affect expression of the nearby locus ( P < 0.001).
DISCUSSION
The interesting characteristics of this transgenic mutant include failure of normal morphological de-
B
FIGURE 6,"Linkage analysis. (A) Haplotypes of backcross prog- eny from the cross (C57BL/6J-9257 X SPRET/Ei)FI X C57BL/6J. Each column represents the haplotype of the chromosome inherited from the F I parent. The number of mice with each haplotype is given at the bottom of each column. Fgf-1, 01-1 and Pdgfrb were typed on the entire backcross DNA panel: Apc and Egr-1 were typed on all backcross progeny with recombination in the interval between the transgene and Pdgfrb. Haplotypes were inferred by assuming the absence of double crossovers. Solid symbols, SPRET/Ei; open symbols, C57BL/6J. (B) Genetic map of chromosome 18 derived from our data (left) and the consensus map (DAVISSON andJoHNsoN 1992) (right).
velopment of the cristae ampullaris and shortening of the membranous and bony labyrinth. The observed asymmetry of left and right canals is sufficient to account for the circling behavior of transgenic mice, and perhaps for the hyperactivity as well. The appear- ance of the ampullae in transgenic and Twirler mice suggests that the production of extracellular material may be defective in the mutant. The affected struc- tures of the inner ear and cranium develop during the period between embryonic day 11-1 6 . A defective bone morphogen, growth factor or related receptor that is normally present during this period of devel- opment is an attractive model to account for the multiple cranial defects.
The phenotypes of mice heterozygous for the 9257 transgene and for the Twirler mutation are compared in Table 4. The similarities include the virtually iden- tical appearance of the cristae ampullaris (compare this paper and LYON 1958). Three features specific to Twirler are the loss of otoliths, early onset obesity and the birth of homozygotes with cleft palate. The exten- sive similarities in phenotype together with the genetic localization to the same chromosome region make it likely that the two mutations have altered at least one gene in common. In view of the complex phenotypes, both mutations could represent contiguous gene syn- dromes affecting two or more adjacent genes. An-

Mouse Craniofacial Mutation 253
TABLE 4
Phenotypes of mice heterozygous for TC” and Twirler mutations
W 2 ” Twirler
Hearing Normal Normal Horizontal canal Shortened Shortened Cristae ampullaris Flat or inverted Flat or inverted Otoliths Normal Missing Cochlea Normal Normal Facial bones Hypoplastic Normal Behavior Circling, non- Circling, non-
swimmer, swimmer, hyperactive hyperactive
Obesity Normal Early onset
Penetrance of circling 80% 90% Chromosomal map Proximal Chr Proximal Chr
position 18 18 Homozygosity Prenatal lethal Day 1 lethal;
cleft palate
obesity
other possibility is that Twirler and 7’$,,’ contain mutations in two distinct but closely linked genes, both of which are required for craniofacial develop- ment. This could account for the complementation (survival) observed in compound heterozygotes. De- termination of the molecular relationship between these two mutations is a goal of our future work.
According to the classification scheme proposed by Steel and Bock (1985) for inner ear mutants of the mouse, the insertional mutation produces a mild mor- phogenetic defect with limited structural abnormali- ties and no apparent effect on hearing. More than 40 circling mutations have been mapped in the mouse genome (GREEN 1989). Many of these cause extensive defects in the cochlea and bony labyrinth. None of these mutants has yet been characterized at the mo- lecular level.
The mutation described here is one of a growing number of insertional mutations in transgenic mice. The reported frequency of mutations in transgenic mice is 3-5% (MEISLER 1992). Two other insertional mutants with circling behavior have been described, the dominant mutation Wocko with extreme disor- ganization of the vestibular apparatus (CRENSHAW et al. 199 1) and the recessive mutation chakragati, which appears to have a central nervous system defect (RATTY et al. 1990, 1992). In view of the large number of transgenic lines that are currently being generated, transgenic insertions are likely to be an increasingly useful resource for investigation of morphogenesis of the inner ear as well as other developmental processes.
We thank MARY LYON for providing Twirler mice and GLENN RADICE for providing mice carrying the ataxia mutation. We are grateful to ROGER ALBIN for histological analysis of the brain of transgenic mice and DAVID F. DOLAN for measurement of auditory brain stem response. We thank J. ESCOBEDO, K. YAMAMOTO, V. SUKHATME and J. HEBERT for providing probes. Supported by
USPHS grant GM24872 (M.H.M.) and HG00272 to DAVID C. WARD, Yale University. C.N.T. received a predoctoral fellowship from the Michigan Center for Cancer Research. D.C.K. is the recipient of National Research Service Award DC00109 and sup- port from the Developmental Biology Training Grant (HD07247). D.L.B. is a predoctoral fellow of the University of Michigan Ge- netics Training Program (GM07544).
LITERATURE CITED
BAIN, P. A, , M. H. MEISLER, B. A. TAYLOR and A. H. PAYNE, 1993 The genes encoding gonadal and non-gonadal forms of 3P-hydroxysteroid dehydrogenase/A5-A4 isomerase are closely linked on mouse chromosome 3. Genomics 16: 219-223.
BALLING, R., U. DEUTSCH and P. GRUSS, 1988 Undulated, a mutation affecting the development of the mouse skeleton, has a point mutation in the paired box of Pax 1. Cell 55: 53 1-535
BARROW, L. L., K. SIMIN, K. MOHLKE, W. C. NICHOLS, D. GINSBURG and M. H. MEISLER, 1993 Conserved linkage of neurotrophin 3 and von Willebrand factor on mouse Chr 6. Mamm. Genome 4: 343-345.
BEECHEY C . V., M. D. BURTENSHAW, B. B. BROWN, E. P. EVANS and A. G. SEARLE, 1980 Chr 18: distance of Tw from cen- tromere. Mouse News Lett. 63: 17.
BOYLE, A. L., S. G. BALLARD and D. C. WARD, 1990 Differential distribution of long and short interspersed element sequences in the mouse genome: chromosome karyotyping by fluores- cence in situ hybridization. Proc. Natl. Acad. Sci. USA 87: 7757-7761.
BOYLE, A. L., D. M. FELTQUITE, N. C. DRACOPOLI, D. E. HOUSMAN and D. C. WARD, 1992 Rapid physical mapping of cloned DNA on banded mouse chromosomes by fluorescence in situ hybridization. Genomics 12: 106-1 15.
BRIGATI, D. J., D. MYERSON, J. J. LEARY, B. SPALHOLZ, S. Z. TRAVIS, et a l . , 1983 Detection of viral genomes in cultured cells and paraffin-embedded tissue sections using biotin-labeled hybrid- ization probes. Virology 126: 32-50.
CAMPER, S. A., T. L. SAUNDERS, R. W. KATZ and R. H. REEVES, 1990 The pit-1 transcription factor gene is a candidate for the murine Snell dwarf mutation. Genomics 8: 586-590.
CONSTANTINI, F., G. RADICE, J. L. LEE, K. K. CHADA, W. PERRY, et a l . , 1989 Insertional mutations in transgenic mice. Proc. Nucleic Acid Res. Mol. Biol. 36: 159-169.
CRENSHAW 111, E. B., A. RYAN, S. R. DILLON, K. KALLA and M. G. ROSENFELD, 1991 Wocko, a neurological mutant generated in a transgenic mouse pedigree. J. Neurosci. 11: 1524-1 530.
DAVISSON, M. T., and K. R. JOHNSON, 1992 Mouse chromosome 18. Mamm. Genome 3: S2614265.
FANNING, T., 1983 Size and structure of the highly repetitive BAM HI element in mice. Nucleic Acids Res. 11: 5073-5091.
GREEN, M. C., 1989 Catalog of Mutant Genes and Polymorphic Loci. Genetic Variants and Strains of the Laboratory Mouse, Ed. 2, edited by M. F. LYON and A. G. SEARLE. Oxford University Press, Oxford.
GRIDLEY, T., 1991 Insertional versus targeted mutagenesis in mice. New Biol. 3: 1025-1034.
GUMUCIO, D. L., K. WIEBAUER, R. M. CALDWELL, L. C. SAMUELSON and M. H. MEISLER, 1988 Concerted evolution of human amylase genes. Mol. Cell. Biol. 8: 1197-1205.
HEBERT, J. M., C. BASILICO, M. GOLDFARB, 0. HAUB and G. R. MARTIN, 1990 Isolation of cDNAs encoding four mouse FGF family members and characterization of their expression pat- terns during embryogenesis. Dev. Biol. 138: 454-463.
HILL, R. E., J. FAVOR, B. L. M. HOGAN, C. C. T. TON, G. F. SAUNDERS, et al . , 199 1 Mouse Small eye results from mutations in a paired-like homeobox-containing gene. Nature 3 5 4 522- 525.
HODGKINSON, C. A., K. J. MOORE, A. NAKAYAMA, E. STEINCRIMS-

254 C.-N. Ting et al.
SON, N. G., COPELAND, N. A. JENKINS and H. ARNHEITER, 1993 Mutations at the mouse microphthalmia locus are asso- ciated with defects in a gene encoding a novel basic-helix-loop- helix-zipper protein. Cell 74: 395-404.
HUGHES, M. J., J. B. LINGREL, J. M. KRAKOWSKY and K. P. ANDERSON, 1993 A helix-loophelix transcription factor-like gene is located at the mi locus. J. Biol. Chem. 268: 20687- 20690.
KINGSLEY, D. M., A. E. BLAND, J. M. GRUBBER, P. C. MARKER, L. B. RUSSELL, et al., 1992 The mouse short ear skeletal mor- phogenesis locus is associated with defects in a bone morpho- genetic member of the TGF beta superfamily. Cell 71: 399- 410.
LANE, P. W., A. G. SEARLE, C. V. BEECHEY and E. M. EICHER, 1981 Chromosome 18 of the house mouse. J. Hered. 72:
Lr, S., E. B. CRENSHAW 111, E. J. RAWSON, D. M. SIMMONS, L. W. SWANSON, et al., 1990 Dwarf locus mutants lacking three pituitary cell types from mutations in the POU-domain gene pit-1. Nature 347: 528-533.
LYON, M. F., 1955 Ataxia-a new recessive mutant of the house mouse. J. Hered. 46: 77-80.
LYON, M. F., 1958 Twirler: a mutant affecting the inner ear of the house mouse. J. Embryol. Exp. Morphol. 6: 105-1 16.
LYON, M. F., 1975 Recombination between Tw and ax. Mouse Newsl. 52: 36.
LYON, M. F., and M. C. KIRBY, 1993 Mouse chromosome atlas. Mouse Genome 91: 40-80.
MEISLER, M. H., 1992 Insertional mutation of “classica1”and novel genes in transgenic mice. Trends Genet. 8: 341-344.
MILLER, S. A., D. D. DYKES and H. F. POLESKY, 1988 A simple salting out procedure for extracting DNA from human nu- cleated cells. Nucleic Acids Res. 16: 1215.
MIESFELD, R., S. OKRET, A. C. WIKSTROM, 0. WRONGE, J. A. GUSTAFSSON, et al . , 1984 Characterization of a steroid hor- mone receptor gene and mRNA in wild-type and mutant cells. Nature 312: 779-78 1.
RATTY, A. K., L. W. FITZGERALD, M. TITELER, S. D. GLICK, J. J. MULLINS, et al., 1990 Circling behavior exhibited by a trans- genic insertional mutant. Mol. Brain Res. 8: 355-358.
RATTY, A. K., Y. MATSUDA, R. W. ELLIOTT, V. M. CHAPMAN and K. W. GROSS, 1992 Genetic mapping of two DNA markers,
409-4 12.
D16Rosl and DZ6Ros2: flanking the mutation site in the chak- ragati mouse, a transgenic insertional mutant. Mamm. Genome
REITH, A. D., and A. BERSTEIN, 1991 Molecular basis of mouse developmental mutants. Genes Dev. 5: 11 15-1 123.
SAMBROOK, J., E. F. FRITSCH and T. MANIATIS, 1989 Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory Press. Cold Spring Harbor, N. Y.
STEEL, K. P., and G. R. BOCK, 1985 Genetic factors affecting hearing development. Acta Otolaryn. (Stockh) Suppl. 421: 48- 56.
STEPHENSON, D. A., M. MERCOLA, E. ANDERSON, C. Y. WANG, C. D. STILES, et a l . , 199 1 Platelet-derived growth factor receptor a-subunit gene (pdgfra) is deleted in the mouse patch (Ph) mutation. Proc. Natl. Acad. Sci. USA 88: 6-10, 1991.
Su, L.-K., W. KINZLER, B. VOGELSTEIN, A. C. PREISINGER, A. R. MOSER, et al., 1992 Multiple intestinal neoplasia caused by a mutation in the murine homolog of the APC gene. Science
SUKHATME, V. P., X. CAO, L. C. CHANG, C.-W. TSAI-MORRIS, D. STAMENKOVICH, et al., 1988 A zinc finger-encoding gene coregulated with c-jos during growth and differentiation, and after cellular depolarization. Cell 53: 37-43.
TING, C.-N., M. P. ROSENBERG, C. M. SNOW, L. C. SAMUELSON and M. H. MEISLER, 1992 Evolutionary role of an endogenous retrovirus in the tissue specific expression of a human salivary amylase gene. Genes Dev. 6: 1457-1465.
WEIHER, H., T . NOD, D. A. GRAY, A. H. SHARPE and R. JAENISCH, 1990 Transgenic mouse model of kidney disease: insertional inactivation of ubiquitously expressed gene leads to nephrotic syndrome. Cell 62: 425-434.
WITTE, 0. N., 1990 Steel locus defines new multipotent growth factor. Cell 63: 5-6.
WOYCHIK, R. P., R. L. MAAS, R. ZELLER, T. F. VOGT and P. LEDER, 1990 “Formins”: protein’s deduced from the alternative tran- scripts of the limb deformity gene. Nature 346: 850-853.
YARDEN, Y., J. A. ESCOBEDO, W. J. KUANG, T . L. YANG-FENG, T . 0. DANIEL, et al., 1986 Structure of the receptor for platelet- derived growth factor helps define a family of closely related growth factor receptors. Nature 323: 226-232.
3: 5- 10.
2 5 6 668-670.
Communicating editor: R. E. GANSCHOW