inhibition of woodchuck hepatitis virus gene expression in
TRANSCRIPT

Virology 384 (2009) 88–96
Contents lists available at ScienceDirect
Virology
j ourna l homepage: www.e lsev ie r.com/ locate /yv i ro
Inhibition of woodchuck hepatitis virus gene expression in primary hepatocytes bysiRNA enhances the cellular gene expression
Zhongji Meng a,e, Song Qiu a,b,c, Xiaoyong Zhang a,c, Jun Wu a,b,d, Thomas Schreiter b, Yang Xu c,Dongliang Yang d, Michael Roggendorf a, Jörg Schlaak b, Mengji Lu a,c,⁎a Institute of Virology, University Hospital of Essen, University Duisburg-Essen, Essen, Germanyb Department of Gastroenterology and Hepatology, University Hospital of Essen, University Duisburg-Essen, Essen, Germanyc Department of Microbiology, Tongji Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan, Chinad Division of Clinical Immunology, Tongji Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan, Chinae Department of Infectious Diseases, Taihe Hospital, Yunyang Medical College, Shiyan, China
⁎ Corresponding author. Institute of Virology, UVirchowstr. 179, 45122 Essen, Germany. Fax: +49 201 72
E-mail address: [email protected] (M. Lu).
0042-6822/$ – see front matter © 2008 Elsevier Inc. Aldoi:10.1016/j.virol.2008.11.012
a b s t r a c t
a r t i c l e i n f oArticle history:
Small interfering RNA (siRN Received 3 August 2008Returned to author for revision1 October 2008Accepted 10 November 2008Available online 6 December 2008Keywords:HepadnavirusWoodchuckRNA interferenceRNAiIFN-stimulated genesISGs
A) has been shown to be active to inhibit the hepatitis B virus gene expressionand replication in transient and stable transfection systems. Here in primary hepatocytes prepared fromnaturally woodchuck hepatitis virus (WHV)-infected woodchucks, four siRNAs targeting the WHV preS1, S, C,and X region led to a depletion of WHV transcripts and replicative intermediates with different kinetics and adecreased production of viral particles. Two siRNAs targeting WHV S and X region had the highest efficacy todeplete 70% of WHV transcripts and replicative intermediates. In addition, siRNA-mediated suppression ofWHV enhanced the expression of cellular genes like MxA and MHC I. Specific siRNAs are able to inhibit thehepadnaviral replication and enhance the expression of cellular genes relevant for antiviral actions. Thus,siRNAs might be useful as novel antiviral agents for the treatment of chronic HBV infection.
© 2008 Elsevier Inc. All rights reserved.
Introduction
About 350 million people are chronically infected with hepatitis Bvirus (HBV) world wide. These patients have a relatively high risk ofdeveloping end-stage liver diseases, such as liver cirrhosis andhepatocellular carcinoma (Seeger and Mason, 2000). Despite sig-nificant progress in recent years, the currently available treatmentregiments for hepatitis B such as interferon-α (IFN-α) or nucleoside/nucleotide analogues are costly and have limited long-term efficacy.Only about one-third of patients treated with IFN-α show a sustainedresponse. Nucleoside or nucleotide analogues are not able to eliminatethe virus completely in the vast majority of patients and may selectresistant viral variants during the prolonged treatment (Locarnini andMason, 2006; Marcellin, 2002). Therefore, it is necessary to developnew antiviral strategies against HBV.
RNA interference (RNAi) may provide a promising approach for thespecific treatment of HBV infection. It is mediated by small interferingRNAs (siRNAs) of a length of 21 to 23 nucleotides (Hannon, 2002) thatlead to the sequence-specific degradation of homologous messenger
niversity Hospital of Essen,35929.
l rights reserved.
RNAs (mRNAs). Using chemically synthesized or vector-expressedsiRNAs, many clinically relevant viruses including the humanimmunodeficiency virus, HBV, and hepatitis C virus could be inhibitedin vitro (Randall and Rice, 2004; Stevenson, 2003;Wu and Nandamuri,2004). A number of recent studies have demonstrated the effective-ness of specific siRNAs for inhibiting the HBV gene expression andviral replication (Giladi et al., 2003; Guo et al., 2005; Hamasaki et al.,2003; Klein et al., 2003; Konishi et al., 2003; McCaffrey et al., 2003;Morrissey et al., 2005; Shlomai and Shaul, 2003; Uprichard et al.,2005; Wu et al., 2005). Yet, RNAi has not been tested in a naturalinfection system with a complete viral life cycle so far.
The woodchuck hepatitis virus (WHV) is a member of the familyHepadnaviridae discovered in 1978 (Summers et al., 1978). WHVcauses acute and chronic infections in woodchucks (Marmota monax)like HBV in humans. The woodchuck model has been proven to be aninformative model for studies on hepadnaviral infection and patho-genesis and for evaluation of antiviral drugs (Lu et al., 2007;Roggendorf and Lu, 2005). In this study, we examined the inhibitionof theWHV gene expression and replication inWHV-infected primarywoodchuck hepatocytes (PWHs) by specific siRNAs. In addition, weinvestigated whether the suppression of viral gene expressionmodulates the expression of host genes with antiviral and immuno-logical functions.

Fig.1. Inhibition of theWHV gene expression by specific siRNAs. (a) A schematic presentation of the open reading frames on theWHV genome and the sites targeted by siRNAs used inthis study. (b) The ability of the selected siRNAs to knock down the specificWHV transcripts. HepG2 cells were co-transfected with 20 nM of siRNA and 1 μg of the expression plasmidsfor selected targets. The WHV gene transcripts were detected 72 h later by Northern blot. The quantification of the signals was performed by using Phosphor-Imager. (c) BHK cellswere co-transfected with 20 nM of siRNA and 1 μg of the expression plasmids for selected targets. WHsAg and WHcAg were detected by immunofluorescence staining with specificantibodies. The expression of EGFP was directly examined by fluorescence microscopy. RDLU, relative digital light units.
89Z. Meng et al. / Virology 384 (2009) 88–96

90 Z. Meng et al. / Virology 384 (2009) 88–96
Results
siRNAs targeting the WHV preS1, S, C, and X gene inhibit WHVgene expression
Four siRNAs targeting to theWHV genomewere designed by usingthe HiPerformance siRNA Design Algorithm (Fig. 1a). The specificinhibitory effect of siRNAs was demonstrated by co-transfection ofsiRNAs with the respective expression plasmids forWHpreS1, WHsAg,WHcAg, andWHxAg-EGFP into HepG2 cells or BHK cells. In the HepG2cells, expression of the specific WHV mRNAs was decreased by co-transfection with the respective siRNAs (Fig. 1b). Two siRNAs siWHsand siWHx were highly efficient to reduce the specific mRNAs to 24%and 27% of the mock control while siWHpreS1 and siWHc had onlymoderate effects (Fig. 1b).
Additionally, the inhibitory effects of WHV-specific siRNAs wereinvestigated on the protein level by the direct detection of WHVproteins WHsAg and WHcAg by immunofluorescence staining in BHKcells. The expression of EGFP by the WHxAg-EGFP fusion constructwas examined by fluorescence microscopy. Consistent with the
Fig. 2. Inhibition of the WHV gene expression and replication in PWHs by specific siRNAs. PsiRNA siGFP. The WHV transcripts (a) and replicative intermediates (b) were analyzed onrepresentative result is shown. The data presented asmeans±standard deviation from three iS, siWHs; C, siWHc; X, siWHx; N, siGFP as a negative control; RDLU, relative digital light un
detection of specific mRNAs, the WHV proteins expression wasdecreased by co-transfection with specific siRNAs (Fig. 1c).
Specific siRNAs lead to the depletion of WHV transcripts and replicativeintermediates in PWHs
PWHs from chronically infected woodchucks are uniformlyinfected by WHV and contain usually a high steady-state level ofWHV specific transcripts and replicative intermediates. It has beenshown that there are two major transcripts, 2.1 and 3.7 kb in lengthfound in the liver samples from chronic WHV-infected woodchucks(Moroy et al., 1985). The 2.1 kb RNAs correspond to the transcripts formajor WHsAg while the 3.7-kb RNA is involved in the expression ofother viral proteins. It is of interest to examine whether specificsiRNAs are able to deplete the WHV RNAs and replicative inter-mediates. PWHs with WHV infection were transfected with specificsiRNAs or control siRNA, respectively. The levels of WHV transcriptsand WHV DNA were assayed on days 2, 4, and 7 after transfection byNorthern blot and Southern blot, respectively. The reduction of WHVtranscripts by specific siRNAs was readily detectable on day 2 (Fig. 2a).
WHs were transfected with 100 nM of siWHpreS1, siWHs, siWHc, siWHx, or a controldays 2, 4, and 7 post transfection by Northern blot and Southern blot, respectively. Andependent Southern blotting analysis were also shown (b lower panel). S1, siWHpreS1;its.

Fig. 3. Inhibition of the production of WHV viral particles released into the cell culturesupernatants by specific siRNAs. PWHs were transfected with 100 nM of siWHpreS1,siWHs, siWHc, siWHx, or a control siRNA siGFP. The copy numbers of WHV progenyDNA secreted into culture supernatants were determined on days 2, 4, and 7 posttransfection by real-time PCR. ⁎ Compared with mock control, pb0.05; ⁎⁎ comparedwith mock control, pb0.01.
91Z. Meng et al. / Virology 384 (2009) 88–96
Two siRNAs, siWHpreS1 and siWHs, were directed to the transcripts ofWHV surface proteins. The treatment with siWHpreS1 led to thereduction of WHV 3.7 kb transcripts while the amount of the 2.1 kbtranscripts remained unchanged, since siWHpreS1 has no target siteon the 2.1 kb transcripts. In addition, both 3.7 and 2.1 kb transcriptswere decreased after the transfection with siWHs and siWHx as well.The siRNA siWHc was less effective to knock down the WHVtranscripts. The inhibitory effect on WHV-specific mRNAs reached amaximum on day 4. The transfection with siWHs and siWHx led to areduction of WHV mRNA to 35% and 31%, respectively. The effect ofsiRNAs lasted at least until day 7 (also see below).
After transfection with WHV-specific siRNAs, WHV replicativeintermediates in PWHs decreased with a delay compared with thekinetics of reduction of WHV mRNAs. This can be explained by thefact that siRNAs do not act directly on the level of the WHV DNA
Fig. 4. Suppression and rebound of the WHV replication after a single siRNA transfection. PWwere examined on days 2, 4, 7, 10, and 14 by Southern blot analysis (a). The signals detectedsetting the mock transfection control as 100% (b). X, siWHx; C, siWHc.
intermediates. The decrease of WHV replicative intermediates wasmainly caused by the reduced formation of new WHV ss DNA frompregenomic RNAs. Consistently, transfection with two potent siRNAssiWHs and siWHx reduced the amounts of WHV replicativeintermediates in PWHs to a level of 37% on day 7 as comparedwith the mock control (Fig. 2b). The other two siRNAs were lesseffective.
Furthermore, the release of viral particles into the culture super-natants was assessed. WHV DNA in culture supernatants wasextracted and quantified by real-time PCR. A single transfection ofsiWHpreS1, siWHs, and siWHx reduced the WHV DNA levels in theculture supernatants. A reduction of more than 50% of the WHV DNAlevel was measured on day 2 after treatment with siWHs and siWHxand reached a level of about 90% suppression on the day 4 (Fig. 3). Arepeated transfection of siRNAs 24 h after the first transfection couldfurther enhance the silencing effect and led to a 90% reduction of theWHV DNA concentration in the cell culture supernatants on day 7(data not shown).
Single transfection of siRNA has only a transient inhibitory effect onWHV replication
As siRNAs have only limited stability in cells, the RNAi effect ceasesover time and new transcripts can be produced from WHV cccDNAs(Bartlett and Davis, 2006). Thus, we examined whether new WHVreplicative intermediates are formed in PWHs treated with specificsiRNAs. PWHs were transfected with siRNAs and the amounts of WHVreplicative intermediates in PWHs were determined up to day 14 aftertransfection. As already shown previously, a single transfection withsiWHx led to a reduction ofWHV replicative intermediates up to day 7(Fig. 4). However, the level of WHV replicative intermediates insiWHx-treated PWHs returned gradually to a high level on day 14,suggesting that the therapeutic use of siRNAs against HBV will facesimilar problems with the persistence of cccDNAs as observed duringthe treatment with nucleoside analogues.
Hs were transfected with 100 nM of siWHx or siWHc. WHV replicative intermediatesin Southern blot were analyzed, and the relative strength of signals was calculated by
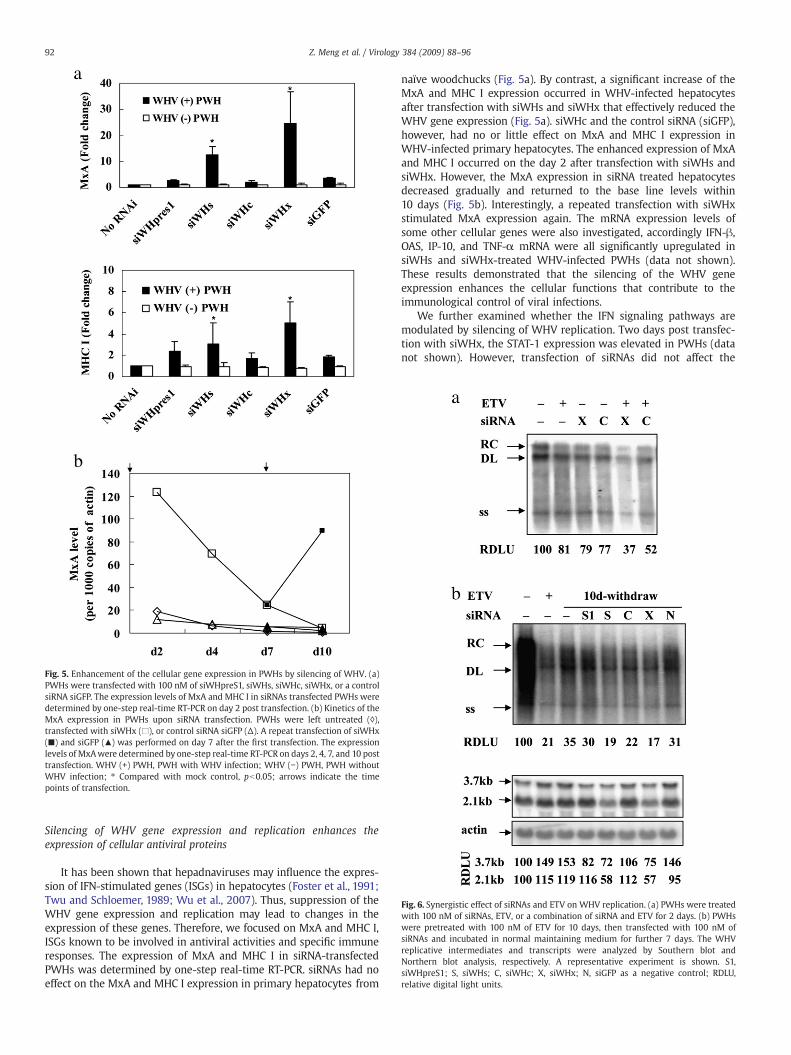
Fig. 5. Enhancement of the cellular gene expression in PWHs by silencing of WHV. (a)PWHs were transfected with 100 nM of siWHpreS1, siWHs, siWHc, siWHx, or a controlsiRNA siGFP. The expression levels of MxA and MHC I in siRNAs transfected PWHs weredetermined by one-step real-time RT-PCR on day 2 post transfection. (b) Kinetics of theMxA expression in PWHs upon siRNA transfection. PWHs were left untreated (◊),transfected with siWHx (□), or control siRNA siGFP (Δ). A repeat transfection of siWHx(■) and siGFP (▴) was performed on day 7 after the first transfection. The expressionlevels of MxAwere determined by one-step real-time RT-PCR on days 2, 4, 7, and 10 posttransfection. WHV (+) PWH, PWH with WHV infection; WHV (−) PWH, PWH withoutWHV infection; ⁎ Compared with mock control, pb0.05; arrows indicate the timepoints of transfection.
Fig. 6. Synergistic effect of siRNAs and ETV on WHV replication. (a) PWHs were treatedwith 100 nM of siRNAs, ETV, or a combination of siRNA and ETV for 2 days. (b) PWHswere pretreated with 100 nM of ETV for 10 days, then transfected with 100 nM ofsiRNAs and incubated in normal maintaining medium for further 7 days. The WHVreplicative intermediates and transcripts were analyzed by Southern blot andNorthern blot analysis, respectively. A representative experiment is shown. S1,siWHpreS1; S, siWHs; C, siWHc; X, siWHx; N, siGFP as a negative control; RDLU,relative digital light units.
92 Z. Meng et al. / Virology 384 (2009) 88–96
Silencing of WHV gene expression and replication enhances theexpression of cellular antiviral proteins
It has been shown that hepadnaviruses may influence the expres-sion of IFN-stimulated genes (ISGs) in hepatocytes (Foster et al., 1991;Twu and Schloemer, 1989; Wu et al., 2007). Thus, suppression of theWHV gene expression and replication may lead to changes in theexpression of these genes. Therefore, we focused on MxA and MHC I,ISGs known to be involved in antiviral activities and specific immuneresponses. The expression of MxA and MHC I in siRNA-transfectedPWHs was determined by one-step real-time RT-PCR. siRNAs had noeffect on the MxA and MHC I expression in primary hepatocytes from
naïve woodchucks (Fig. 5a). By contrast, a significant increase of theMxA and MHC I expression occurred in WHV-infected hepatocytesafter transfection with siWHs and siWHx that effectively reduced theWHV gene expression (Fig. 5a). siWHc and the control siRNA (siGFP),however, had no or little effect on MxA and MHC I expression inWHV-infected primary hepatocytes. The enhanced expression of MxAand MHC I occurred on the day 2 after transfection with siWHs andsiWHx. However, the MxA expression in siRNA treated hepatocytesdecreased gradually and returned to the base line levels within10 days (Fig. 5b). Interestingly, a repeated transfection with siWHxstimulated MxA expression again. The mRNA expression levels ofsome other cellular genes were also investigated, accordingly IFN-β,OAS, IP-10, and TNF-α mRNA were all significantly upregulated insiWHs and siWHx-treated WHV-infected PWHs (data not shown).These results demonstrated that the silencing of the WHV geneexpression enhances the cellular functions that contribute to theimmunological control of viral infections.
We further examined whether the IFN signaling pathways aremodulated by silencing of WHV replication. Two days post transfec-tion with siWHx, the STAT-1 expression was elevated in PWHs (datanot shown). However, transfection of siRNAs did not affect the

93Z. Meng et al. / Virology 384 (2009) 88–96
expression level of TRIF, MyD88, and IRAK1 (data not shown).Furthermore, Bay11-7082, an NF-kappa B inhibitor did not block thestimulation of MxA in PWHs (data not shown).
A synergistic action of siRNAs and ETV on the inhibition ofWHV replication
Nucleoside analogues like ETV are effective antivirals to inhibit theactivity of hepadnaviral polymerases and prevent the formation ofviral replicative intermediates (Hadziyannis, 2006). Treatment ofWHV-infected PWHs with ETV at a concentration of 100 nMsignificantly inhibitedWHV replicationwithin 2 days, while combina-tion of siWHx and ETV exhibited more potent inhibition on WHVreplication than ETV or siRNA treatment alone (Fig. 6a). Treatment ofPWHs with ETV for 10 days led to a strong reduction of WHVreplicative intermediates to a level below 20% of untreated controls(Fig. 6b upper panel). The levels of WHV transcripts remainedunchanged by ETV treatment, while the transfection with siRNAsresulted in a reduction of WHV RNAs (Fig. 6b lower panel). Theinhibitory effect of siRNAs on the WHV replication could be seen afterthe discontinuation of ETV treatment. The WHV replicationrebounded significantly 7 days after ETV discontinuation butremained at the low levels if the hepatocytes were treated withsiWHs and siWHx (Fig. 6b upper panel).
Further, we tested whether inhibition of WHV replication by ETVmay have an influence on the expression of MxA as observed for theuse of WHV-specific siRNAs. ETV treatment did not lead to inductionof MxA in PWHs (Fig. 7), while their expression was enhanced aftertransfection with siWHx but not siWHc. These results indicated thatsiRNAs could contribute to the antiviral therapies through differentways and act synergistically with ETV.
Discussion
In this study, we demonstrated that WHV-specific siRNAs wereable to reduce the levels of WHV mRNAs in primary hepatocytesnaturally infected with WHV. Four siRNAs targeting the differentregions of the WHV genome were used in this study. The siRNAstargeting theWHV S and X region had the highest efficacywhile siRNAagainst the WHV core region was not effective, probably due to theinappropriate target sequence. The siRNA to theWHV preS regionwasable to induce the degradation of the specific 3.7 kb RNAs, while it wasless potent in suppressing WHV replicative intermediates, comparedto siWHs and siWHx. In addition, our results also indicated thatsiRNAs and nucleoside analogues may act synergistically in suppres-sing WHV replication. The control siRNA had slight unspecific effectson WHV replication. However, this effect was not significant whencompared with mock control.
Fig. 7. TheMxA expression in PWHs after siRNA, ETV, or a combination of siRNA and ETVtreatment. PWHs were left untreated or treated with siRNA, ETV, or a combination ofETV and siRNA. The expression levels of MxA were determined by one-step real-timeRT-PCR 2 days post transfection.
There are some interesting features in the action of effectivesiRNAs on WHV-infected PWHs. First, the amounts of WHV RNAs,replicative intermediates, and WHV DNAs in culture supernatantschanged with different kinetics. The decrease of the levels of WHV-specific RNAs was readily measurable on day 2. However, thedepletion of WHV replicative intermediates became visible on day 4,thus with a delayed kinetics compared to that of WHV transcripts.Obviously, a pool of WHV replicative intermediates was present inpersistently infected PWHs and needed about more than 2 days to bedepleted. The production of viral particles was decreased on day 2 asthe amounts of WHV RNAs were reduced in PWHs, probably due tothe reduced expression of WHV proteins. Second, the amounts of viralreplicative intermediates returned to base line levels within twoweeks. Two factors determine the kinetics of viral rebound. One factoris the half-life of siRNAs in PWHs. However, it is difficult to determinethe half-life of siRNA in transfected cells and to correlatewith the RNAieffects (Chiu and Rana, 2003). The formation of new pregenomic RNAsfrom the WHV cccDNA pool in PWHs will determine the final steady-state level of WHV replicative intermediates. Thus, repeated applica-tions of siRNAs or use of modified siRNAs with a longer half-life wouldbe necessary for clinical use.
Furthermore, the knock down of WHV gene transcripts by specificsiRNAs led to an enhanced expression of cellular genes like MxA andMHC I, which are related to specific immune responses and antiviralfunctions. An increased expression of MxA and MHC I was onlydetected in WHV-infected PWHs but not in naïve PWHs. Thus, wecould exclude that the biochemical properties of siRNAs itself wereresponsible for IFN induction. In addition, upregulation of the MxAand MHC I expression occurred only if the siRNAs were effective todegradeWHV RNAs. For example, siWHc did not induce an increase ofthe MxA and MHC I expression in WHV-infected PWHs, as this siRNAwas inactive. Therefore, the mechanisms leading to the enhancedcellular gene expression byWHV-specific siRNAs need to be examinedfurther. One hypothesis is that the knock down of WHV transcriptswould reduce the amount ofWHV proteins and therefore facilitate thecellular gene expression. Early reports indicated that HBV polymerasemay inhibit the IFN signaling pathway (Wu et al., 2007). Further, HBVmay counteract the host antiviral effector mechanisms by down-regulating the IFN-inducible MxA promoter through direct interactionof precore/core proteins or by inhibiting proteasome activities in anHBX-dependentmanner (Fernandez et al., 2003; Zhang et al., 2004). Inthis context, knock down of the viral gene expression would reliefthese inhibitory effects on the cellular gene expression. Anotherhypothesis to be investigated is that the induction of the cellular genesmay occur due to the triggering of IFN responses by degradationproducts of WHV RNAs during RNAi. It has been reported that acleavage of cellular RNAs by RNase L produces small RNAs that are ableto activate IFN-β (Malathi et al., 2007). It should be investigated if asiRNA-directed RNA degradation would also produce small RNAfragments and trigger the activation of IFN-β gene. This processcould not yet be examined in woodchuck cells due to lack ofwoodchuck specific reagents. It is noteworthy, however, that anenhanced expression of MxA and MHC I by siRNA treatment does notoccur in established human hepatoma cell lines with HBV replication(Meng and Lu, unpublished results). This may be due to the defectiveIFN signaling pathways in such hepatoma cells (Stojdl et al., 2000).However, early publications indicated that some other cell lines maybe suitable for such studies (Bridge et al., 2003; Sledz et al., 2003).
Though the mechanisms leading to the enhanced cellular geneexpression need further studies, this effect itself may facilitate theimmunotherapy and lead to an enhanced antiviral activity in vivo. Inchronically HBV-infected patients, there is a deficiency of specificcellular responses to HBV proteins (Chisari and Ferrari, 1995;Rehermann, 2007). The early attempts to stimulate the specificcellular responses in such patients by conventional HBV vaccines orDNA vaccines had only limited effects (Mancini-Bourgine et al., 2004).

Table 1siRNAs target sequences
Designation Target sequences Locationa WHV gene
siWHpreS1 5′-AGCTCAAACTTTCGAAATCAA-3′ 3279–3299s PreS1siWHs 5′-ACGACAGTCAATTGCAGACAA-3′ 632–652 SsiWHc 5′-AAGAACAATTATAGTAAATCA-3′ 2263–2283 CsiWHx 5′-AAAGATCAATTATTAACTAAA-3′ 1827–1847 XsiGFP 5′-cggcaagctgaccctgaagttcat-3′
a The positions of the target sequences according to the WHV sequence M19183.
94 Z. Meng et al. / Virology 384 (2009) 88–96
It is proposed that a pretreatment of chronically HBV-infected patientswith antiviral drugs would enhance the effect of therapeutic vaccines,as an effective antiviral treatment with lamivudine would restore thecellular immune responses in patients to a limited extent (Boni et al.,1998, 2001, 2003). A combined treatment with antiviral drugs andtherapeutic vaccines has not yet been successfully tested in clinicaltrials (Dahmen et al., 2002; Horiike et al., 2005; Vandepapeliere et al.,2007). In the woodchuck model, such combined approaches haveshown an improved antiviral effect (Lu et al., 2008; Menne et al.,2007). An enhancement of cellular IFN responses by siRNA may bebeneficial and is worth future investigation.
Taken together, we demonstrated the potential usefulness andlimitation of siRNAs for inhibition of hepadnaviral gene expressionand replication in naturally WHV infected hepatocytes. The ability ofsiRNAs to modulate the cellular gene expression specifically bysuppression of hepadnaviral gene expression and replication repre-sents a unique aspect and provides new options for treatment ofchronic HBV infection.
Materials and methods
siRNAs and entecavir (ETV)
Four siRNAs were designed by using the HiPerformance siRNADesign Algorithm (Qiagen, Düsseldorf, Germany) in combination withthe algorithm described by Reynolds et al. (Reynolds et al., 2004). Thesequences of the siRNAs are given according to the WHV59 genomesequence (GenBank accession no. M19183). The selected siRNAs,designated as siWHpreS1, siWHs, siWHc, and siWHx, have the specifictarget sequences within the WHV preS1, S, C, and X gene, respectively(Fig.1 and Table 1). siGFP, a siRNA against green fluorescent protein, wasused as the control (Table 1). All siRNAs were purchased from Qiagen.ETV was purchased from Bristol-Myers Squibb (Munich, Germany).
Woodchucks
All animals were purchased from North Eastern Wildlife (Ithaca,NY, USA) and kept in the center of animal laboratory of the Universityof Essen. Experiments were conducted in accordance with the Guide
Table 2Primers used for amplification of WHV preS1, X, and C gene and real-time RT-PCR assay
Amplicon Primer Polarity Nuc
WHV preS1 whpres1 Sense 5′-Cwhs2 Antisense 5′-C
WHV X gene whx-1 Sense 5′-Awhx-2 Antisense 5′-T
WHV C gene wc1 Sense 5′-Twc-149s Antisense 5′-A
Woodchuck β-actin wβ-actin-A1 Sense 5′-Twβ-actin-A2 Antisense 5′-T
Woodchuck MHC I wMHCI-s1 Sense 5′-CwMHCI-a1 Antisense 5′-G
Woodchuck MxA wMxA-s1 Sense 5′-GwMxA-a1 Antisense 5′-C
The underlined part of primer wc-149s indicates the specific cleavage sites of restriction en
for the Care and Use of Laboratory Animals and were reviewed andapproved by the local Animal Care and Use Committee of the districtgovernment (Düsseldorf, Germany).
Primary hepatocytes culture and siRNA transfection
Primary hepatocytes from WHV-infected and uninfected wood-chucks were prepared by liver perfusion with collagenase IV (Sigma,Munich, Germany) as described previously (Baccarani et al., 2003).The viability of prepared hepatocytes was assessed by trypane bluedye exclusion and exceeded 80% in all experiments. The cells wereplated at a density of 1.5×106 cells per well on collagen I-coated six-well plates in Williams medium (Gibco, Karlsruhe, Germany)supplemented with 10% fetal calf serum (Gibco), 1 mg/ml ofhydrocotisone (Sigma), 10 μg/ml of insulin (Serva, Amsterdam, TheNetherlands), 50 μg/ml of gentamycin (Sigma), 50 IU/ml of penicillin–streptomycin (Gibco), 2 mM glutamine (Gibco), 1.25 mg/ml of inosine(Serva), and 1% DMSO (Merck, Darmstadt Germany).
Transfection of primary hepatocytes with siRNAs was carried outon day 2 of in vitro culture except specification. Lipofectamine 2000(Invitrogen, Karlsruhe, Germany) was used according to the manufac-turer's instructions. 150 pmol siRNA and 5 μl of Lipofectamine 2000per well were applied in a final volume of 1.5 ml Opti-MEM. After 5 h,the medium was replaced by fresh culture medium.
To ensurehighlyefficient siRNAdelivery intohard-to-transfect PWHs,a high amount of siRNA is needed (Gilot et al., 2005). A dose-findingexperiment suggested that siWHx inhibited the WHV gene expressionand replication in a dose-dependent manner. 100 nM of siRNAs wereused in the present experiments. Although higher concentration than100 nM led to more potent effect on the mRNA level, no more reductionon WHV replication was observed (data not shown).
Construction of expression vectors for WHV preS1, surface antigen(WHsAg), core protein (WHcAg), and X protein (WHxAg)-enhancedgreen fluorescent protein (EGFP) fusion
The plasmids expressing WHsAg and WHcAg were describedpreviously (Lu et al., 1999). To construct the vector expressing theWHV preS1 protein, the WHV genome region (nt 2992–964) wasamplified by PCR with the primers whpres1 and whs2 (Table 2) andcloned into the pCR2.1 vector (Invitrogen). The fragment containingthe large WHsAg gene was isolated by digestion with EcoRI andinserted into the EcoRI site of the pcDNA3 vector (Invitrogen). Thegenerated plasmid contained the large WHsAg gene under the controlof the cytomegalovirus promoter. The region comprising the WHV Xregion was amplified with the primers whx-1 and whx-2 (Table 2),and cloned into the pCR2.1 vector. The clone sequence was verified byDNA sequencing. The fragment containing the WHV X region was cutby EcoRI and cloned into the EcoRI site of pIRES2-EGFP vector(Clontech, Heidelberg, Germany).
leotide sequence Position Reference
AGCTAGTGCAACATAATGG-3′ 2976 M19183CACCATTTTGTTTTATTGA-3′ 987 M19183TGGCTGCTCGCCTGTGTTGC-3′ 1503 M19183TACAGAAGTCGCATGCATTT-3′ 1928 M19183GGGGCATGGACATAGATCCCTA-3′ 2015 M19183AGATCTCTAAATGACTGTATGTTCCG-3′ 2467 M19183GGAATCCTGTGGCATCCATGAAAC-3′ 1 AY170121.1AAAACGCAGCTCAGTAACAGTCCG-3′ 349 AY170121.1CAGACCCAGGAGATGGAAC-3′ 729 AF146091GACAATCACAGCAGCAACA-3′ 940 AF146091GAGGGAGGAGAAGAGGAAA-3′ 66 EU503128TGGAGATGCGGTTGTGAG-3′ 189 EU503128
zyme BglII.

95Z. Meng et al. / Virology 384 (2009) 88–96
Cell cultures, transfection, and immunofluorescence staining
Culturing and transfection of BHK and HepG2 cells were carriedout as described previously (Zheng et al., 2002). To detect WHsAgand WHcAg, immunofluorescence staining was performed withmonoclonal mouse anti-WHpreS (Waters et al., 2001; Zheng et al.,2002) or anti-WHcAg antibodies (Zhang et al., 2006). The anti-WHpreS antibody recognizes the middle WHsAg that contains themajor WHsAg as a domain. The expression of EGFP by the WHx-EGFP fusion was determined by direct observation and photographyunder a fluorescence microscope with an excitation wavelength of490 nm.
Isolation and analysis of viral RNA and WHV replicative intermediates
Total RNA was extracted from hepatocytes with TRIzol reagent(Invitrogen) according to the manufacturer's instructions. Northernblot analysis was carried out by agarose-glyoxal method according topublished protocols (Meng et al., 2008). WHV replicative intermedi-ates were purified from intracellular core particles according to theprotocol described for HBV previously (Meng et al., 2008). The isolatedWHV DNA was subjected to agarose gel electrophoresis, followed bydenaturation and Southern blotting. WHV RNA and DNA on Northernand Southern blots were detected by hybridization with a 32P-labeledfull length HBV probe. The quantitative analysis of the signals on theSouthern blots and Northern blotswas performed by using a Phospho-Imager (Cyclon, Parkard, Meriden, CT, USA). The total WHV replicativeintermediates shown as a smear in the Southern blots including thethree major forms: RC (relaxed circular), DL (duplex linear), and SS(single-stranded) DNAwere quantified. The relative strength of signals(relative digital light units, RDLU) was calculated by setting the mocktransfection control as 100%. The WHV specific gene transcripts incotransfection experiments and the 3.7 and 2.1 kb RNAs in PWHswerequantified and normalized against β-actin. The RDLU was calculatedby setting the mock transfection control as 100%.
Quantitation of WHV progeny DNA in culture supernatants
WHV progeny DNA in the culture supernatants was extractedusing Qiaamp DNA blood mini kit (Qiagen) and quantified as genomeequivalents (GE) per ml by real-time PCRwith LightCycler DNAmasterSYBR green kit (Roche Diagnostics, Mannheim, Germany). PCR wasperformed with the primers wc1 and wc-149s (Table 2) (Lu et al.,2005, 2008) on Light Cycler™ instrument (Roche) with 95 °C for 0 s,53 °C for 10 s and 72 °C for 12 s. A plasmid containing a full lengthWHV genome was diluted and served as standard. The detection limitof this assay was 103 WHV GE/μl.
Real-time reverse-transcriptase (RT) PCR
Total RNA was purified using the RNeasy Mini Kit (Qiagen)according to the manufacturer's instructions. One-step real-time RT-PCR was performed with 100 ng of total RNA using QuantiFast SYBRGreen RT-PCR Kit (Qiagen) on a Light Cycler™ instrument (Roche)with 50 °C for 10min for reverse transcription followed by 45 cycles ofPCR: 95 °C 10 s, 60 °C 30 s. The primers used are listed in Table 2. Therelative mRNA copies were determined with a standard curveconstructed using corresponding cDNA fragments. The copy numbersof woodchuck MxA and MHC I transcripts were normalized againstwoodchuck β-actin transcripts.
Acknowledgments
We thank Thekla Kemper and Barbara Bleekmann for theirexcellent technical assistance. We are thankful to Delia Cosgrove forthe critical reading of the manuscript.
This work is supported by grants of the Deutsche Forschungsge-meinschaft (Lu 669/2-1, GRK1045/1, and Lu 669/5-1), the FederalMinistry of Education and Research (GU0207), the National NaturalScience Foundation of China (30271170, 30571646) and the NationalHigh Technology Research and Development Program of China(2006AA02Z128).
References
Baccarani, U., Sanna, A., Cariani, A., Sainz-Barriga, M., Adani, G.L., Zambito, A.M., Piccolo,G., Risaliti, A., Nanni-Costa, A., Ridolfi, L., Scalamogna, M., Bresadola, F., Donini, A.,2003. Isolation of human hepatocytes from livers rejected for liver transplantationon a national basis: results of a 2-year experience. Liver Transpl. 9, 506–512.
Bartlett, D.W., Davis, M.E., 2006. Insights into the kinetics of siRNA-mediated genesilencing from live-cell and live-animal bioluminescent imaging. Nucleic Acids Res.34, 322–333.
Boni, C., Bertoletti, A., Penna, A., Cavalli, A., Pilli, M., Urbani, S., Scognamiglio, P., Boehme,R., Panebianco, R., Fiaccadori, F., Ferrari, C., 1998. Lamivudine treatment can restoreT cell responsiveness in chronic hepatitis B. J. Clin. Invest. 102, 968–975.
Boni, C., Penna, A., Ogg, G.S., Bertoletti, A., Pilli, M., Cavallo, C., Cavalli, A., Urbani, S.,Boehme, R., Panebianco, R., Fiaccadori, F., Ferrari, C., 2001. Lamivudine treatmentcan overcome cytotoxic T-cell hyporesponsiveness in chronic hepatitis B: newperspectives for immune therapy. Hepatology 33, 963–971.
Boni, C., Penna, A., Bertoletti, A., Lamonaca, V., Rapti, I., Missale, G., Pilli, M., Urbani, S.,Cavalli, A., Cerioni, S., Panebianco, R., Jenkins, J., Ferrari, C., 2003. Transientrestoration of anti-viral T cell responses induced by lamivudine therapy in chronichepatitis B. J. Hepatol. 39, 595–605.
Bridge, A.J., Pebernard, S., Ducraux, A., Nicoulaz, A.L., Iggo, R., 2003. Induction of aninterferon response by RNAi vectors in mammalian cells. Nat. Genet. 34,263–264.
Chisari, F.V., Ferrari, C., 1995. Hepatitis B virus immunopathology. Springer Semin.Immunopathol. 17, 261–281.
Chiu, Y.L., Rana, T.M., 2003. siRNA function in RNAi: a chemical modification analysis.RNA. 9, 1034–1048.
Dahmen, A., Herzog-Hauff, S., Bocher, W.O., Galle, P.R., Lohr, H.F., 2002. Clinical andimmunological efficacy of intradermal vaccine plus lamivudine with or withoutinterleukin-2 in patients with chronic hepatitis B. J. Med. Virol. 66, 452–460.
Fernandez, M., Quiroga, J.A., Carreno, V., 2003. Hepatitis B virus downregulates thehuman interferon-inducible MxA promoter through direct interaction of precore/core proteins. J. Gen. Virol. 84, 2073–2082.
Foster, G.R., Ackrill, A.M., Goldin, R.D., Kerr, I.M., Thomas, H.C., Stark, G.R., 1991.Expression of the terminal protein region of hepatitis B virus inhibits cellularresponses to interferons alpha and gamma and double-stranded RNA. Proc. Natl.Acad. Sci. U. S. A. 88, 2888–2892.
Giladi, H., Ketzinel-Gilad, M., Rivkin, L., Felig, Y., Nussbaum, O., Galun, E., 2003. Smallinterfering RNA inhibits hepatitis B virus replication in mice. Mol. Ther. 8, 769–776.
Gilot, D., Serandour, A.L., Ilyin, G.P., Lagadic-Gossmann, D., Loyer, P., Corlu, A., Coutant,A., Baffet, G., Peter, M.E., Fardel, O., Guguen-Guillouzo, C., 2005. A role for caspase-8and c-FLIPL in proliferation and cell-cycle progression of primary hepatocytes.Carcinogenesis 26, 2086–2094.
Guo, Y., Guo, H., Zhang, L., Xie, H., Zhao, X., Wang, F., Li, Z., Wang, Y., Ma, S., Tao, J., Wang,W., Zhou, Y., Yang, W., Cheng, J., 2005. Genomic analysis of anti-hepatitis B virus(HBV) activity by small interfering RNA and lamivudine in stable HBV-producingcells. J. Virol. 79, 14392–14403.
Hadziyannis, S.J., 2006. New developments in the treatment of chronic hepatitis B.Expert Opin. Biol. Ther. 6, 913–921.
Hamasaki, K., Nakao, K., Matsumoto, K., Ichikawa, T., Ishikawa, H., Eguchi, K., 2003. Shortinterfering RNA-directed inhibition of hepatitis B virus replication. FEBS Lett. 543,51–54.
Hannon, G.J., 2002. RNA interference. Nature 418, 244–251.Horiike, N., Fazle Akbar, S.M., Michitaka, K., Joukou, K., Yamamoto, K., Kojima, N., Hiasa,
Y., Abe, M., Onji, M., 2005. In vivo immunization by vaccine therapy following virussuppression by lamivudine: a novel approach for treating patients with chronichepatitis B. J. Clin. Virol. 32, 156–161.
Klein, C., Bock, C.T., Wedemeyer, H., Wustefeld, T., Locarnini, S., Dienes, H.P., Kubicka, S.,Manns, M.P., Trautwein, C., 2003. Inhibition of hepatitis B virus replication in vivoby nucleoside analogues and siRNA. Gastroenterology 125, 9–18.
Konishi, M., Wu, C.H., Wu, G.Y., 2003. Inhibition of HBV replication by siRNA in a stableHBV-producing cell line. Hepatology 38, 842–850.
Locarnini, S., Mason, W.S., 2006. Cellular and virological mechanisms of HBV drugresistance. J. Hepatol. 44, 422–431.
Lu, M., Hilken, G., Kruppenbacher, J., Kemper, T., Schirmbeck, R., Reimann, J., Roggendorf,M., 1999. Immunization of woodchucks with plasmids expressing woodchuckhepatitis virus (WHV) core antigen and surface antigen suppresses WHV infection.J. Virol. 73, 281–289.
Lu, M., Isogawa, M., Xu, Y., Hilken, G., 2005. Immunization with the gene expressingwoodchuck hepatitis virus nucleocapsid protein fused to cytotoxic-T-lymphocyte-associated antigen 4 leads to enhanced specific immune responses in mice andwoodchucks. J. Virol. 79, 6368–6376.
Lu, M., Menne, S., Yang, D., Xu, Y., Roggendorf, M., 2007. Immunomodulation as anoption for the treatment of chronic hepatitis B virus infection: preclinical studies inthe woodchuck model. Expert Opin. Investig. Drugs 16, 787–801.
Lu, M., Yao, X., Xu, Y., Lorenz, H., Dahmen, U., Chi, H., Dirsch, O., Kemper, T., He, L.,Glebe, D., Gerlich, W.H., Wen, Y., Roggendorf, M., 2008. Combination of an antiviral

96 Z. Meng et al. / Virology 384 (2009) 88–96
drug and immunomodulation against hepadnaviral infection in the woodchuckmodel. J.Virol. 82, 2598–2603.
Malathi, K., Dong, B., Gale Jr., M., Silverman, R.H., 2007. Small self-RNA generated byRNase L amplifies antiviral innate immunity. Nature 448, 816–819.
Mancini-Bourgine, M., Fontaine, H., Scott-Algara, D., Pol, S., Brechot, C., Michel, M.L.,2004. Induction or expansion of T-cell responses by a hepatitis B DNA vaccineadministered to chronic HBV carriers. Hepatology 40, 874–882.
Marcellin, P., 2002. Advances in therapy for chronic hepatitis B. Semin. Liver Dis. 22(Suppl. 1), 33–36.
McCaffrey, A.P., Nakai, H., Pandey, K., Huang, Z., Salazar, F.H., Xu, H., Wieland, S.F.,Marion, P.L., Kay, M.A., 2003. Inhibition of hepatitis B virus in mice by RNAinterference. Nat. Biotechnol. 21, 639–644.
Meng, Z., Xu, Y., Wu, J., Tian, Y., Kemper, T., Bleekmann, B., Roggendorf, M., Yang, D., Lu,M., 2008. Inhibition of hepatitis B virus gene expression and replication byendoribonuclease-prepared siRNA. J. Virol. Methods 150, 27–33.
Menne, S., Tennant, B.C., Gerin, J.L., Cote, P.J., 2007. Chemoimmunotherapy of chronichepatitis B virus infection in the woodchuck model overcomes immunologictolerance and restores T-cell responses to pre-S and S regions of the viral envelopeprotein. J. Virol. 81, 10614–10624.
Moroy, T., Etiemble, J., Trepo, C., Tiollais, P., Buendia, M.A., 1985. Transcription ofwoodchuck hepatitis virus in the chronically infected liver. EMBO J. 4, 1507–1514.
Morrissey, D.V., Blanchard, K., Shaw, L., Jensen, K., Lockridge, J.A., Dickinson, B.,McSwiggen, J.A., Vargeese, C., Bowman, K., Shaffer, C.S., Polisky, B.A., Zinnen, S.,2005. Activity of stabilized short interfering RNA in a mouse model of hepatitis Bvirus replication. Hepatology 41, 1349–1356.
Randall, G., Rice, C.M., 2004. Interfering with hepatitis C virus RNA replication. VirusRes. 102, 19–25.
Rehermann, B., 2007. Chronic infections with hepatotropic viruses: mechanisms ofimpairment of cellular immune responses. Semin. Liver Dis. 27, 152–160.
Reynolds, A., Leake, D., Boese, Q., Scaringe, S., Marshall, W.S., Khvorova, A., 2004.Rational siRNA design for RNA interference. Nat. Biotechnol. 22, 326–330.
Roggendorf, M., Lu, M., 2005. Woodchuck hepatitis virus, In: Thomas, T.H.,Zuckermann, A., Lemon, S. (Eds.), Viral Hepatitis, 3rd ed. Blackwell PublishingLtd., Oxford, pp. 210–224.
Seeger, C., Mason,W.S., 2000. Hepatitis B virus biology.Microbiol. Mol. Biol. Rev. 64, 51–68.Shlomai, A., Shaul, Y., 2003. Inhibition of hepatitis B virus expression and replication by
RNA interference. Hepatology 37, 764–770.Sledz, C.A., Holko, M., de Veer, M.J., Silverman, R.H., Williams, B.R., 2003. Activation of
the interferon system by short-interfering RNAs. Nat. Cell Biol. 5, 834–839.
Stevenson, M., 2003. Dissecting HIV-1 through RNA interference. Nat. Rev. Immunol. 3,851–858.
Stojdl, D.F., Lichty, B., Knowles, S., Marius, R., Atkins, H., Sonenberg, N., Bell, J.C., 2000.Exploiting tumor-specific defects in the interferon pathway with a previouslyunknown oncolytic virus. Nat. Med. 6, 821–825.
Summers, J., Smolec, J.M., Snyder, R., 1978. A virus similar to human hepatitis B virusassociated with hepatitis and hepatoma inwoodchucks. Proc. Natl. Acad. Sci. U. S. A.75, 4533–4537.
Twu, J.S., Schloemer, R.H., 1989. Transcription of the human beta interferon gene isinhibited by hepatitis B virus. J. Virol. 63, 3065–3071.
Uprichard, S.L., Boyd, B., Althage, A., Chisari, F.V., 2005. Clearance of hepatitis B virusfrom the liver of transgenic mice by short hairpin RNAs. Proc. Natl. Acad. Sci. U. S. A.102, 773–778.
Vandepapeliere, P., Lau, G.K., Leroux-Roels, G., Horsmans, Y., Gane, E., Tawandee, T.,Merican, M.I., Win, K.M., Trepo, C., Cooksley, G., Wettendorff, M., Ferrari, C., 2007.Therapeutic vaccination of chronic hepatitis B patients with virus suppression byantiviral therapy: a randomized, controlled study of co-administration of HBsAg/AS02 candidate vaccine and lamivudine. Vaccine 25, 8585–8597.
Waters, J.A., O'Neill, C., bdul-Gader, A., Goldin, R., Karayiannis, P., Monjardino, J., 2001.Purification of woodchuck hepatitis surface antigen using a monoclonal antibodyraised against the antigen. J. Virol. Methods 93, 97–103.
Wu, J., Nandamuri, K.M., 2004. Inhibition of hepatitis viral replication by siRNA. Expert.Opin. Biol. Ther. 4, 1649–1659.
Wu, H.L., Huang, L.R., Huang, C.C., Lai, H.L., Liu, C.J., Huang, Y.T., Hsu, Y.W., Lu, C.Y., Chen,D.S., Chen, P.J., 2005. RNA interference-mediated control of hepatitis B virus andemergence of resistant mutant. Gastroenterology 128, 708–716.
Wu, M., Xu, Y., Lin, S., Zhang, X., Xiang, L., Yuan, Z., 2007. Hepatitis B virus polymeraseinhibits the interferon-inducible MyD88 promoter by blocking nuclear transloca-tion of Stat1. J. Gen. Virol. 88, 3260–3269.
Zhang, Z., Protzer, U., Hu, Z., Jacob, J., Liang, T.J., 2004. Inhibition of cellular proteasomeactivities enhances hepadnavirus replication in an HBX-dependent manner. J. Virol.78, 4566–4572.
Zhang, Z., Tian, Y., Li, L., Fiedler, M., Schmid, E., Roggendorf, M., Xu, Y., Lu, M., Yang, D.,2006. A conserved linear B-cell epitope at the N-terminal region of woodchuckhepatitis virus core protein (WHcAg). J. Virol. Methods 135, 17–25.
Zheng, X., Schirmbeck, R., Hilken, G., Waters, J.A., Yang, D., Reimann, J., Roggendorf, M.,Lu, M., 2002. Characterization of complex B cell epitopes on woodchuck hepatitisvirus surface antigens by using plasmids encoding chimeric proteins and DNAimmunization. Virology 294, 342–353.