influences of hypothyroidism on the taste detection ... of hypothyroidism on the taste detection...
TRANSCRIPT

Behavioral Neuroscience1992, Voll06, No 6,992-998
Copyright 1992 by the American Psychological Association, Inc0735-70441921$3.00
Influences of Hypothyroidism on the Taste Detection Perform~nce of
Rats: A Signal Detection Analysis
Gary M. BrbsvicDepartment of Psychology
and Biopsychology LaboratoryRider College
Richard L. Doty , " ,Smell and Taste Center
and Department of Otorhinolaryngology-Head and Neck Surgery
School of MedicineUniversity of Pennsylvania .
M. Michelle RoweTemple University
Amy Har!on and Nicholas KolodiyDepartment of Psychology and Biopsychology Laboratory, Rider College
The influences of hypothyroidism on behavioral measures of the taste function in male and femaleLong-Evans rats were determined. Experimental rats' preferences for and ability to detect NaCI,HCI, sucrose, and quinine sulfate were examined before, during, and after 9 weeks of maintenanceon 0.1% propylthiouracil (PTU), an ag~nt that produces marked hypothyroidism, with similardeterminations made for control animals. Despite sigrificant decreases in PTy-treated rats' serumtriiodothyronine (T3) and thyroxin (T4), there were no changes in sensitivity or responsivity to thetarget tastants. However, alte;red preferences for NaCI, HCI, and quinine sulfate were observed i~rPTU-treated rats; elevated consumption of HCI and quinine sulfate was present at the end of thej c cstudy when serum T3 and T4 had returned to near-baseline levels. The data confirm observations
,that PTU-induced hypothyroidism alters rats' taste preference behavior.
reported to be completely corrected afteI: 16 days of thyroidhormone replacement therapy, and for most patients in that
study, detection and recognition thresholds were lowered after3 months of replacement therapy.
In a recent study of 750 subjects who had presented withcomplaints of chemosensory dysfunction (Deems et al., 1991 ),patients who were taking thyroid medication (e.g., levothyrox-ine) complained of taste loss more frequently than patientswho were not. H~wever, these p~tients scored higher on a taste
ide,I)tification test and rated low concentrations of caffeine asmore, intense than did comparison patients. Furthermore,their taste thresholds for citric acid, NaCI, and sucrose werewithin normal limits, which suggests that subjective reports oftaste in such patients are not reflected in detection thresholdmeasures (see also Lewitt et al., 1989).
To date, no animal studies have evaluated the influences ofhypothyroidism on taste sensitivity per se, although prefer-ences and ingestive behavior have been examined. For exam.ple, rats made hypothyroid by propylthiouracil (PTU) added todrinking water demonstrated increased consumption of andlowered preference thresholds for NaCI (Fregly, 1962; Fregly,
Cade, Waters, Straw,& Taylor, 1965; Fregly &Waters, 1965,1966). These effects, attributed to hypothyroidism, were re-versed by administrations of graded dosages of desoxycorticos-terone (Fregly & Waters, 1966). ~oltzman rats made hypothy-roid by:administration of radioactive iodide were reported to, ,increase consumption of normally aversive concentrations ofsour aI)d bitter tastants, with a return of ingestjve behayior tonear-baseline levels after 2-3 weeks oL .thyroid hormone
The influence of hypothyroidism, on taste sensitivity iscontroversial. In humans, several studies have reported noeffects of this condition on taste threshold sensitivity. Forexample, Pittman and Beschi (1967) evaluated the thresholdsensitivity of normal subjects, hyperthyroid patients, andhypothyroid patients to sodium chloride (NaCI), potassiumchloride (KCI), sodium bicarbonate, sucrose, urea, and hydro-chlorit; acid (HCI) and found no differences between thesethree study groups. Similarly, Lewitt; Laing, Panhuber, Cor"bett, and Carter (1989) reported finding no differences be-tween hypothyroid and control subjects in the recognition oflow concentrations of NaCI, sucrose, citric acid, and caffeine,or iii the identification of orange juice and pineapple juice.
In contrast, McConnell, Menendez, Smith, and Henkin(1975) reported significant elevations of both detection andrecognition thresholds for NaCI, sucrose, urea, and HCI.Similar threshold elevations for the odorants pyridine andnitrobenzene were also noted. The return of the thresholds tol1ormallevels after thyroid ho!mone replacement therapy andthe absence of dysgeusia and dysosmia before hypothyroidismsuggested to McConnel1 et al. a prominent role for thyroidhormones in the modulation of olfactory and gustatory sensitiv-ity .Indeed, for 1patient, abnormalities of sm~11 and taste were
This research was supported by National Institute on Deafness andOther Communicative Disorders GrantDC 00161.
Correspondence concerning this article should be addressed to GaryM. Brosvic, Department of Psychology, Biopsychology Laboratory,Rider College, 2083 Lawrenceville Road, Lawrenceville, New Jersey08648-3099.
992

993HYPOTHYROIDISM AND TASTE
General Procedure
Animals were weighed daily, placed on a 23V2-hr water deprivationschedule!for 14 days, and trained and tested on the taste detection,taste-mixture discrimination, and preference tests described later(baseline phase). Two groups of 10 rats (5 male and 5 female) werematched on the signal detection measures of sensitivity and responsiv-ity to each tastant, preference behavior, and serum Tz and T4, all ts(18)< 1, all ps > .75. One group was randomly designated as theexperimental group and received PTU dissolved in drinking wateravailable ad libitum for 9 weeks (treatment phase ). PTU was subse-quently removed from the experimental rats' water bottles and testingresumed for 9 weeks (posttreatment phase ). PTU-treated rats' meandaily water consumption was measured and the same amount of waterwas made available to the yoked control animals. Blood samples foranalysis of free serum T 3 and T 4 were collected at baseline and at theconclusion of the treatment and posttreatment phases. Signal detec-tion measures of sensitivity and responsivity to NaCI, sucrose, quininesulfate, and HCI were determined weekly during treatment andposttreatment in 260-trial test sessions. Taste-mixture discriminationperformance and preference for NaCI, sucrose, quinine sulfate, andHCI were deterQ1ined weekly.
replacement (McConnell et al., 1975). Such changes in inges;.tive behavi~r are prima facie evidence that hypothyroidisminfluences the discrimination performance 0! rats but do notdemonstrate that taste sensitivity per se is influenced by thiscondition.
In our study we examined the effects of PTU-inducedhypothyroidism on taste preference and sensitivity meas\ireSwith sucrose, NaC/, HCI, and quinine sulfate as representa.:tives of the four basic taste qualities. In addition, we examinedthe influences of such hypothyroidism on the ability of rats toperform a taste-mixture discrimination task. As revealedbelow, our findings show that although taste preferences arealtered by PTU-induced hypothyroidism, neither taste sensitiv-ity nor taste discrimination performance is similarly altered.This is true in spite of the fact that PTU produced markeddecreases in free-serum triiodothyronine (T 3) and thyroxin
(T4).
Method
Biochemical Verification of HypothyroidismSubjects
Ten male and 10 female Long-Evans rats, approximately 90 days ofage at the beginning of the study, served as subjects. The rats werehoused in pairs in 30 x 30 x 60 cm polystyrene cages in a temperature-and humidity-controlled vivarium maintained on a 12:12-hr light-darkcycle. All testing and training occurred within the light phase of thelight-dark cycle. Chow was available ad libitum. During pre- andposttreatment all animals were maintained on a 23V2-hr water-deprivation schedule. During the treatment period the experimentalanimals were maintained on ad libitum access to 0.1% PTU dissolvedin water; water consumption was liIeasured, and control animals
received an equivalent amount of water.
After anesthesia with ketamine (0.75 ml ip ), approximately 2 ml ofblood was taken by orbital puncture with 1.5-mm glass pipettes dippedin heparin. Blood samples were centrifuged at 4,000 revolutions perminute for 7 min, and the serum was stored in separate 1.7-ml conicalplastic containers at 8 °C until TJ and T4 could be analyzed byradioimmunoassay procedures (see Cullen, Doherty, & Ingbar, 1973;Dozin & De Nayer, 1983; Griessne & Lemarchand-Beraud, 1973;Jacoby, Mueller, & Wurtman, 1975).
Operant Training and T est frocedures
Apparatus
Rats were trained to perform all taste discriminations in a 30 x 30 x
45cm Plexiglas chamber fitted with a 10-barrel stimulus delivery tubeand a single-barrel reinforcement tube mounted on one wall of thechamber (see Brosvic & Slotnick, 1986). The responses of subjectswere recorded, and the stimulus deliveries controlled by computer(IBM PC XT; Med Associates interface; MED-PC).
Stimuli and PTU
Briefly, animals were trained on a modified discrete trials go-no-goprocedure in which a variable (10-30) number of licks at the stimulusdelivery tube defined the attending response and produced a 0.01-mlsample of the S+ or S- tastant. On delivery of S+, licking at thereinforcement tube was required to initiate the next trial. If the animalswitched to the reinforcement tube before making 10 licks at thestimulus delivery tube, the trial was counted as a correct detection(hit), and the rat received a 0.02-ml water reward. If it perseverated onthe stimulus delivery tube (making 10 licks before switching), the trialwas counted as a miss, and the subsequent response on the reinforce-ment tube produced a 0,005-ml water reward. On S- trials the rat wastrained to continue responding on the stimulus delivery tube un~il thenext stimulus was delivered. Trials in which it did so were scored ascorrect rejections. If, on the other hand, it switched to the reinforce-ment tube before making. 10 responses on the stimulus delivery tube,the switching behavior was punished by a 3-s time-out period signaledby operation of a house light, and these trials were counted as false
alarms.Initially, rats were trained in, sessions in which only S + was
presented. After the rat attained 90% correct responding in 100consecutive trials, nonreinforced S- trials were introduced. After a ratachieved 90% correct responding in 100 trials of 50 S+ and 50 S-,descending and ascending series of trials were introduced. A daily testsession consisted of 260 trials, the first 20 of which consisted of 10 S+and 10 S- (triple-deionized water) warm-up trials not used in thesignal detection calculations. After these warm-up trials, blocks of 20trials (10 S+ and 10 S- ) were presented according to a 120-trialdescending concentration series. In each 20-trial block, the positive
Reagent-grade aqueous solutions of NaCI and sucrose at molarconcentrations from 5 x 10-4 to 2 x 10-1 were used. Reagent-grade
aqueous solutions of quinine sulfate and HCI at molar concentrationsfrom 5 x 10-7 to 5 x 10-4 were also used. The molar concentrations ofthe tastants permitted comparison with results of the detectionperformance of normal and control animals tested previously (seeBrosvic & Hecht, 1989; Brosvic et al., 1991; Brosvic & Hoey, 1990a,1990b). Triple-deionized water served as the solvent for all solutions,
the S- stimulus, and reinforcement.PTU was added to drinking water at the concentration of 0.1 %
during the treatment phase. PTU administered at this concentrationproduces marked hypothyroidism in mice when added to drinkingwater (Beard & Mackay-Sim, 1986, 1987; Mackay-Sim & Beard, 1987)and rats when added to laboratory chow (Fregly, 1962; Fregly et al.,
1965; Fregly & Waters, 1965, 1966).

994 BROSVIC, D0Ty;:ROW;E, HARRON, ~OLODIY
and negative trials were presented ip a random order with therestriction that not more than ,3 trials of one type ,occurred insuccession and that there was an equal number of positive andnegative trials in successive blocks. A 120-trial ascending concentra-tion series was then initiated for a total of 40 trials for each S+ teststimulus per test session. ) \
comp.ute~ and the data collapsed into 3-week summary ~cores for
baselIne; treatment,'and posttreatme~. Analyses were then per-formed separately for each group and tastant with Friedlriah's test with
multiple comparisons (Conover, 1980). Similar analyses were used toexamine potential differences in taste-mixture discrimination perfor-
mance, taste preff:rence behavior, and free serum T 3 and T 4.
Taste Perfonnance Measures Control Procedure
The proportion of hits and false alarms were used to calculate
nonparametric indexes of sensitivity (SI) and responsivity (RI) de-scribed by Frey and Colliver (1973). SI was calculated (rorn: theproportion of hits, P(HIT), and false alarms, P(FA), recorded duringeach test session with the formula:
At the conclusion of~ach dflY's testing, all stimulus delivery solenoid
reservoirs and solenoid valves were flushed with 500 ml of triple-deionized water (at 22 "C) to remove potential residuals. Tygon tubingand stimulus reservoirs were replaced every 4 weeks, and the solenoidvalves used to deliver tastant samples(S+ andS- ) were routinelyrotated to minimize potential extraneous cues for discriminativeresponding. In multiple3,000-trial sessions, two randomly selectedchannels of the stimulus delivery tube were filled with triple-deionized
water, sucrose, KCI, NaCI, or sodium-saccharin. In these tests,equimolar concentrations of the same tastant (or water) served as S+and S- to examine potential solenoid valve cues for discriminative
responding.
P(HLT) -P(FA)SI = ,2[P(HIT) + P(FA)] -[P(HIT) +P(FA)l
The SI can theoretically range from O (no detection) to 1 (error-freedetection ).
RI represents the general tendency to respond independent of thetype, of lit.imulus presented. RI was calculated from the proportion of
hits, J'~HIT), and false alarms, P(FA), recorded during each testsession according to the formula:
Results
No signific~nt differences on any depend~nt measure wereobserved as a function of sex of subject (Mann- Whitey testsjallps > .5), so the data were collapsed across sexes for subse-
quent analyse&;.
The RI can range from -1 ( very conservative response criterion) to + 1(very liberal response criterion ). Sensory Perfonnance Measures
For each representative of the four taste qualities, therewere no significant between-groups differences in SI values at
anystimul}Jscon<;entration (Mann-Whitney te&ts, allps >. .5).Similarly, there were no between-groups differences when SIvalues were combined across the stimulus concentrations(Mann-Whitney tests, allps > .5). SI values combined acrossconcentrations are presented in Figure 1, respectively, forNaCI and sucrose.
Taste-Mixture Discrimination Task
All rats were tested oha discrimination task in which the siandard
(5+ ) stimulus consisted of a mixture of 2.9 x 10-2-mol sucrose and 3.4x 10-2-mol NaCI, The comparison 5- stimuli were mixtures of
sucrose and N.itl with 2.9 x 10-2-mol sucrose and NaCI at the molarconcentrations of 4 x 10-3,8 X 10-3, 1.2 x 10-2, 1.6 x 10-2,2 x 10~2,2.~ x 10~2, or 2.8 x 10-2. Initially, rats were tr1!ined to discriminate
between the standard stimulus and the 2.9 x 10-2-mol sucrose with4 x 10-3-mol NaCI comparison stimulus. When the criterion of 75%
correct responding (15 or fewer errors in 60 consecutive trials) wasachieved, the next highest 5- stimulus was introduced, and testingcontinued until the animal failed to achie\ie\criterion in 400 consecu-tive trials.
NonsensoryPerfonnance Measures
For each of)he fo~r taste stim~li, there werc no significal1t
between-gro~ps differences observed for RI values at anystimulus concentration (Mann- Whitnc;y tests, all ps .;> .5).Similarly, there were no between-groups differences when RIvalues were combined across the stimulus concentrations(Mann- Whitney tests, all ps > .5). RI values combined acrossconcentratiol1s for NaCl and sucrose are shown in Figure 2,
respectively.
Taste Preference Tests
Preference for NaCI,(3 x 10-1 mol), sucrose (1.5 x 10-1 mol);quinine sulfate (1.28 x 10-6 mol), and HCI (2.5 x 10-3 mol) wasexamined in 2-hr tests for each animal during baseline, treatment, andposttreatment. Two 150-ml glass bottles, one with tastant dissolved intriple-deionized water and the other with triple-deionized water, werepresented, and bottle positions were reversed halfway through eachtest session. Results of these tests are expressed in terms of thepercentage of preference according to the formula: Test SolutionIntake x 100 + Test Solution + Water Intake. (Intake was measured
in milliliters. )
Taste-Mixture Discrimination Task, ;.
No significant between-groups differences for taste-mixt1,1rethresholds were observed (Mann,.Whitney and WilcoxQn tests,a)1 ps ;> :5). AIl rats could discrimina~e .(75% accuracy orhigher) between the S+ (2.9 x lO-2-mol sucrose and 3.4 xlO-2-mol NaCI) and the highest s- (2.9 x lO-2-mol sucroseand 2.4 x lO-2-mol NaCI) stimuli, but only 7 rats coulddiscriminate between the S+ and the next highest S- stimulus(?,9 x lO-2.mol sucr9se and 2.8 x lO-2-mol NaCI). As
determincd by graphic interpolation, the mean taste mixture
Statistical Analysis
To determine wheth~r sensitivity or responsivity to each ta~tantdiffered between the two study groups, SI and RI scor~ w~re
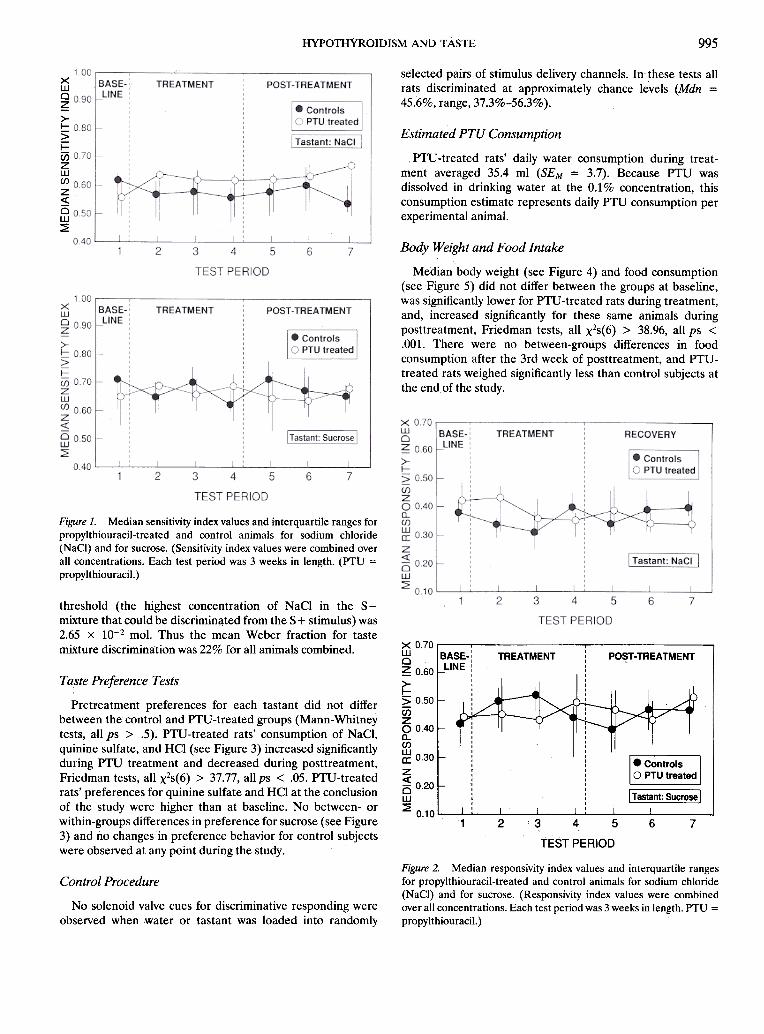
995HYPornyRoIDIsM AND TAsTE
selected pairs of stimulus delivery channels. In these tests allrats discriminated at approximately chance levels (Mdn =
45.6%, range, 37.3%-56.3%).
xw
01Z
>-I-I>i=Ci51ZW
(/)1Z
~
0(W
~
Estimated PTU Consumption
PTU-treated rats' daily water consumption during treat-ment averaged 35.4 ml (SEM = 3.7). Because PTU was
dissolved in drinking water at the 0.1% concentration, thisconsumption estimate represents daily PTU consumption perexperimental animal.
Body Weight and Food Intake
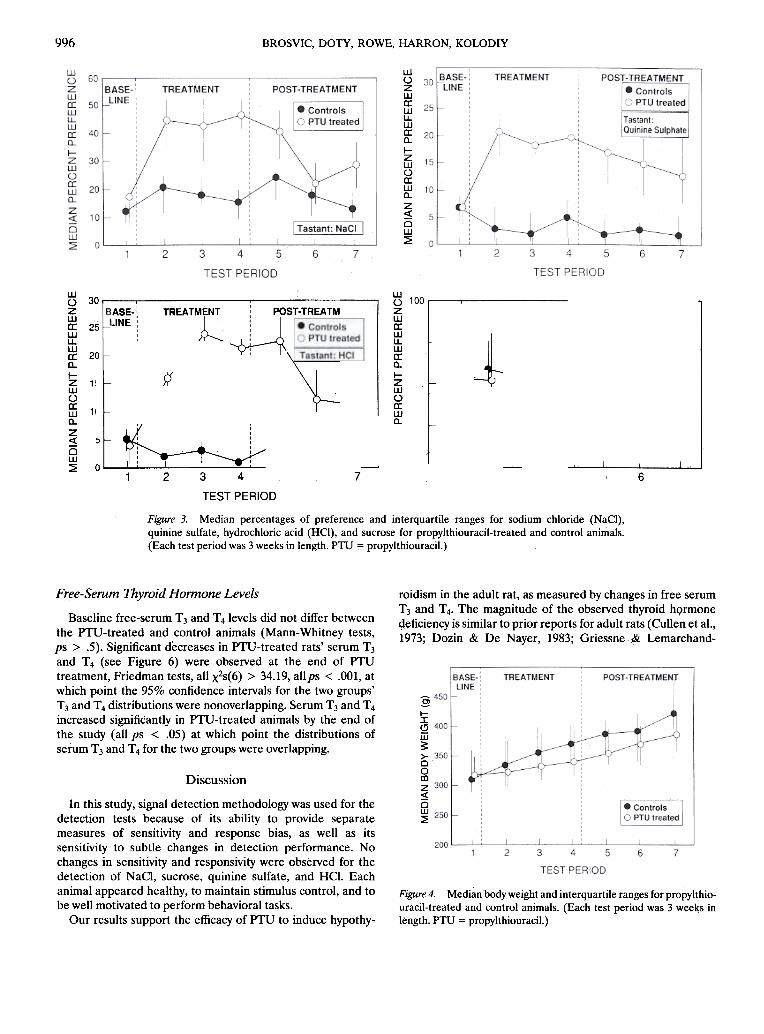
Median body weight (see Figure 4) and food consumption(see Figure 5) did not differ between the groups at baseline,was significantly lower for PTU-treated rats during treatment,and, increased significantly for these same animals duringposttreatment, Friedman tests, all X2s(6) > 38.96, all ps <.001. There were no between-groups differences in foodconsumption after the 3rd week of posttreatment, and PTU-treated rats weighed significantly less than control subjects atthe end of the study.
Figure 1. Median sensitivity index values and interquartile ranges forpropylthiouracil-treated and control animals for sodium chloride(NaCI) and for sucrose. (Sensitivity index values were combined overall concentrations. Each test period was 3 weeks in length. (pTtJ =
propylthiouracil. )
threshold (the highest concentration of NaCI in the S-mixture that could be discrimin&ted from the S+ stimulus) was2.65 x 10-2 mol. Thus the mean Weber fraction for tastemixture discrimination was 22% for all animals combined.
Ta,ste Preference Tests
Pretreatment preferences for each tastant did not differbetween the control and PTU-treated groups (Mann- Whitneytests, all ps > .5). PTU-treated rats' consumption of NaCI,quinine sulfate, and HCI (see Figure 3) increased significantlyduring PTU treatment and decreased during posttreatment,Friedman tests, all X2s(6) > 37.77, allps < .05. PTU-treatedrats' preferences for quinine sulfate and HCI at the conclusionof the study were higher than at baseline. No between- orwithin-groups differences in preference for sucrose (see Figure3) and rio changes in preference behavior for control subjectswere observed at any point during the study.
x 0.70
~ BASE- TREATMENT PO~T-TREATMENT
~ 0.60 LINE
>-1-~ 0.50
cnZO 0.40a.cn
W030 ~a: ..Controls
~ O PTU treatedO 0.20W I Tastant: Sucrose I~ 0.10
1 2 .3 4 5 6 7
TEST PERIOD
Figure 2. Median responsivity index values and interquartile rangesfor propylthiouracil-treated and control animals for sodium chloride(NaCI) and for sucrose. (Responsivity index values were combinedover all concentrations. Each test period was 3 weeks in length. PTU =
propylthiouracil. )
Control Procedure
No solenoid yalve cues for discriminative responding wereobserved when water or tastant was loaded into randomly
996 BROSVIC, DOTY, ROWE, HARRON, KOLODIY
wuzwa:wu.wa:0..f-zwua:w0..z~ow~
POST -TREATMENTBASE-
LlNF
TREATMENT
.Controlsi PTU treated
w 30 ., ,() , ' ,Z BASE-' TREATMENT: POST-TREATMENT
~ 251~INEi ~ ~ iw , ,u. ' ,
w ,c: 20 ,
~ Iz 15W()
ffi 101C-Z<x:C5w~
~ ~:,,,
~
4
~
}1 e '"'f(
I
--,,--+-1I 1 I
5 6
I. Contr~ls I10 PTU treated I
~~
UJ()zUJa:UJLL 85UJa:a.1-Z 70UJ()a:UJa. 55Z~OUJ ,~ ,40' I: I
1 2
.,
3 4 5
TEST PERIOD
6
~ . , ,
, :, ,: ,, ,
u1 2 3 4 7
TEST PERIOD
Figure 3. Median percentages of preference and interquartile ranges for sodium chloride (NaCI),quinine sulfate, hydrochloric acid (HCI), and sucrose for propylthiouracil-treated and control animals.(Each test period was 3 weeks in length. PTU = propylthiouracil.)
Free-Serum Thyroid Hormone Levels roidism in the adult rat, as measured by changes in free serumT3 and T4. The magnitude of the observed thyroid hQrmone~~ficiency is similar to prior reports for adult rats (Cullen et al.,1973; Dozin & De Nayer, 1983; Griessne & Lemarchand-
Baseline free-serum T3 and T4levels did not differ betweenthe PTU-treated and control animals (Mann-Whitney tests,ps > .5). Significant decreases in PTU-treated rats' serum T3and T4 (see Figure 6) were observed at the end of PTUtreatment, Friedman tests, all X2s(6) > 34.19, allps < .001, atwhich point the 95% confidence intervals for the two groups'T3 and T4 distributions were nonoverlapping. Serum T3 and T4increased significantly in PTU-treated animals by the end ofthe study (all ps < .05) at which point the distributions ofserum T3 and T4 for the two groups were overlapping.
§I-I(:Ji:ij$:>-0Omz<I:OUJ~
Discussion
Figure 4. Median body weight and interquartile ranges for propylthio-uracil-treated and control animals. (Each test period was 3 weeks inlength. PTU = propylthiouracil.)
In this study, signal detection methodology was used for thedetection tests because of its ability to provide separatemeasures of sensitivity and response bias, as well as itssensitivity to subtle changes in detection performance. Nochanges in sensitivity and responsivity were observed for thedetection of NaCI, sucrose, quinine sulfate, and HCI. Eachanimal appeared healthy, to maintain stimulus control, and tobe well motivated to perform behavioral tasks.
Our results support the efficacy of PTU to induce hypothy-

997HYPOTHYROIDI~M AND TASTE
confirmed in additional testing in which separate groups of ratsreceiveq thyroidectomy with supplemental thyroxin, thyroidec-tomy without supplemental thyroxin, or sham operations. Nobetween-orwithin-groups differences in SIs or Rls or prefer-ence for sucrose were observed, and there were no differencesin preference behavior between sham-operated rats and thy-roidectomized fats that received supplemental thyroxin. Signif-icant increases in preferences for NaCI, HCI, and quininesulfate were observed for thyroidectomized rats that did not
receive supplemental thyroxin.Also note that PTU, at the concentration used in this and
other studies, likely produces a bitter taste. Therefore, werecently completed a study to establish whether exposing ratsto bitter solutions ( other than PTU) over time would producesimilar alterations in their preferences. Accordingly, separategroups of rats were maintained on ad libitum access to PTU(0.1% ), caffeine (5 x 10-3 mol), urea (2.5 x 10-1 mol), orwater and tested on preference and detection tasks. Nobetweel1- or within-groups differences in SIs or Rls wereobserved. Taste preferences of rats maintained on caffeine orurea didllot differ from those of rats maintained on water,which indicates that exposure to bitter stimuli per se does notproduce pr~ference changes analogous to those observed with
PTU.
Figure 5. Median food consumption and interquartile ranges forpropylthiouracil-treated and control animals. (The baseline periodwas 3 weeks; all other test periods were 1 week each. Pru
propylthiouracil. )
2
TEST PFRIOD
=00E-~1-
~=>[I:w00z~ow~
Beraud, 1913; Ito, Valcanil, & Timiras, 1977; Jacoby et al.,1975). Simifarly, PTU-treated rats' decreased food consump-tion and failure to gain weight, in relation to that {)f controlsubjects, is similar to prior reports for adult inice (Beard &Mackay-Sim, 1986,1987; Mackay~Sim & Beard, 1987).
Significant alterations in the ingestion of NaCI (normallypreferred) and of quinine sulfate and HCI (normally aversive )were observed forea:ch Pru-treated animal, with prt;:ft;:rencesfor HCI and quinine sulfate influenced more than that forNaCI. This pattern is similar to prior reports for rats madehypothyroid by PTU (Fregly, 1962; Fregly, et al., 1965; Fregly& Waters, 1965, 1966) and for humans with either untreatedprimary hypothyroidism'(Mattes, Heller,& Rivlin, 1985; Pitt-man & Beschi, 1967) or pseudohypothytoidism (Henkin,1968). These changes in ingestive behavior also parallel thoseobserved for zinc-deficient (Jakinovich & Osborn, 1981;McConnell & Henkin, 1974) and selectively desalivated rats(Brosvic & Hoey, 1990b), that is, altered preferences for alltastants except for those that are generally considered sweet.However, preferences for sucrose in this and the prior st~dieswere mitigated by a ceiling effect.
The alterations in preference for NaCI reversed shortly afterdiscontinuation of PTU and within 6 weeks returned tobaseline. However, the alterations in preference for HCI andquinine sulfate, although attenuated after discontinuation ofPTU, remained significantly higher than baseline. In follow-up
,tests copducted in our laboratory, we found that considerablylonger recovery periods (range, 2--6 months) are required forPTU-tr~ted rats' preferences for sour and bitter tastants toreturn to baseline, and this range is consistent with the 3months of thyroxin replacement therapy reported to returnhuman hypothyroid patients' th~esholds to more normal levels(McConnell et al., 1975). Alterations in preference behavi!:>rappear to be closely related to thyroid-hormone deficiency, as
Figure 6. Mediarlree-serum triiodothyronine (T3) and thyroxin (T4)and interquartile ranges for propylthiouracil-treated and controlanimals. (Pro = propyltl\iouraci!. )

998 BROSVIC, DOTY, ROWE, HARRON, KOLODIY
The addition of PTU to the rat's drinking water reducedfree-serum T3 and T4 and altered preferences for NaCI, HCI,and quinine sulfate. However, preference tests are not suffi-cient measures of sensory capacity, and when measured with asignal detection p!ocedure, changes in neither sensitivity norresponsivity to representatives of the four taste qualities wereobserved. The reversibilityof altered NaCI intake during theposttreatment phases, the gradual return of preferences forbitter and sour tastants to baseline levels in follow-up studies,and the changes in preference behavior in thyroidectomizedanimals that did not receive supplemental thyroxin indicatethat thyroid hormones playa role in altering ingestive behav-iof, presumably by somehow influencing chemosensory input;however, no specific mechanisms have been identified. Thechanges observed in this study may be similar to alterations insynaptic delay and specific changes in peripheral, sensory, andmotor function reported in humans (Rivlin, 1971,1975) or mayparallel decreased auditory and visual evoked potentials ob-Served in animals and humans with untreated hypothyroidism(Bradley, Eayrs, Glass, & Heath, 1961; Bradley, Eayrs, &Schmalbach, 1960; Harris, Della Rovere, & Prior, 1965).
References
Dozin, B., & De Nayer, 0. (1983). Triiodothyronine receptors in adultrat brain: Topographical distribution and effect of hypothyroidism.
Neuroendocrinology, 39, 261-266.Pregly, M. J. (1962). Effect of hypothyroidism on sodium chloride
aversion of renal hypertensive rats. Endocrinology, 71, 683-692.Pregly, M. J., Cade, J. R., Waters, I. W., Straw, J. A., & Taylor, R. E.
(1965). Secretion of aldosterone by adrenal glands of propylthioura-cil-treated rats. Endocrinology, 77; 777-784.
Pregly, M. J., & Waters, I. W. (1965). Effect of propylthiouracil on
preference threshold of rats for NaCI solutions. Proceedings of the
Society for Experimental Biology and Medicine, 120, 637-640.Pregly, M. J., & Waters, I. W. (1966). Effect of desoxycorticosterone
acetate on NaCI appetite of propylthiouracil-treated rats. Physiologyand Behavior, 1, 133-138.
Prey, P. W., & Colliver, J. A. (1973). Sensitivity and responsivitymeasures for discrimination learning. Learning and Motivation, 4,327-342.
Griessne, M., & Lemarchand-Beraud, T. (1973). Thyrotropin secre-tion and metabolism in rats during PTU treatment. Endocrinology,
92; 166-173.Harris, R., Della Rovere, M., & Prior, P. (1965). Electroencephalo-
graphic studies in infants with hypothyroidism. Archives of Diseases
ofChildren, 40,612-621.Henkin, R. I. (1968). Impairment of olfaction and of the taste of sour
and bitter in pseudohypothyroidism. Journal of Clinical Endocrinol-ogy and Metabolism, 28, 624.
Ito, J. M., Valcana, T., & Timiras, P. S. (1977). Effect of hypo- andhyperthyroidism on regional monoamine metabolism in the adult ratbrain. Neuroendocrinology, 24, 55-64.
Jacoby, J. H., Mueller, G., & Wurtman, R. J. (1975). Thyroid state andbrain monoamine metabolism. Endocrinology, 97, 1332-1335.
Jakinovich, W., & Osborn, D. W. (1981). Zinc nutrition and saltpreference in rats. American Journal of Physiology, 241, 233-239.
Lewitt, M. S., Laing, D. G., Panhuber, H., Corbett, A., & Carter, J. N.
(1989). Sensory perception and hypothyroidism. Chemical Senses,14, 537-546.
Mackay-Sim, A., & Beard, M. D. (1987). Hypothyroidism disruptsneural development in the olfactory epithelium of adult mice.
Developmental Brain Research, 36, 190-198.Mattes, R. D., Heller, A. D., & Rivlin, R. S. (1985). Abnormalities in
suprathreshold taste function in early hypothyroidism in humans. InH. L. Meiselman & R. S. Rivlin (Eds. ), Clinical measurement of tasteand smell (pp. 467-486). New York: Macmillan.
McConnell, S. D., & Henkin, R. I. (1974). Altered NaCI preference forsodium chloride, anorexia, and changes in plasma and urinary zincin rats fed a zinc deficient diet. Journal ofNutritioil, 104, 1108-1112.
McConnell, S. D., Menendez, C. E., Smith, P. R., & Henkin, R. I.(1975). Defects of taste and smell in patients with hypothyroidism.American Journal of Medicine, 59, 354-364.
Pittman, J. A., & Beschi, R. J. (1967). Taste thresholds in hyper- andhypothyroidism. Journal of Clinical Endocrinology and Metabolism,27, 895--896.
Rivlin, R. S. (1971). Hypothyroidism: Vitamins. In S. C. Werner & S.H. Ingbar (Eds.), The thyroid (pp. 750-765). New York: Harper &Row.
Rivlin, R. S. (1975). Defects of taste and smell in patients with
hypothyroidism. American Journal of Medicine, 59, 354-364.
Beard, M. D., & Mackay-Sim, A. (1986). Thyroid control of olfactoryfunction in adult mice. Neuroscience Letters, 23, S32.
Beard, M. D., & Mackay-Sim, A. (1987). Loss of sense of smell in
adult, hypothyroid mice. Developmental Brain Research, 36, 181-189.Bradley, P. B., Eayrs, I. T., Glass, A., & Heath, R. W. (1961). The
maturational and metabolic consequences of neonatal thyroidec-tomy upon the recruiting response in the rat. Electroencephalo-graphic Clinical Neurophysiology, 13,577-592.
Bradley, P. B., Eayrs, I. T., & Schmalbach, K. (1960). The electroen-cephalogram of normal and hypothyroid rats. Electroencephalo-graphic Clinical Neurophysiology, 12,467-473.
Brosvic, G. M., & Hecht, G. S. (1989). Effects of copper and vitaminB-6 deprivation on taste sensitivity in the rat: A signal detection
analysis. Physiology and Behaviol; 45,1139-1145.Brosvic, G. M., Hecht, G. S., LaHaye, S. E., Rowe, M. M., Risser, I.
M., Clementson, E., & Dihoff, R. E. (1991). Quality-specificdifferences in rat taste detection performance as a function ofstimulus volume. Physiology and Behaviol; 50, 711-716.
Brosvic, G. M., & Hoey, N. E. (1990a). Dietary sodium deprivationdoes not influence taste sensitivity in the rat. Physiology andBehaviol; 47, 881-888.
Brosvic, G. M., & Hoey, N. E. (1990b). Taste detection and discrimina-tion of rats following selective desalivation. Physiology and Behavior,
48, 617-623.Brosvic, G. M., & Slotnick, B. M. (1986). Absolute and intensity-
difference thresholds in the rat: Evaluation of an automatedmulti-channel gustometer. Physiology and Behaviol; 38, 711-717.
Conover, W. I. (1980). Practical nonparametrics. New York: Wiley.Cullen, M. I., Doherty, G. F., & Ingbar, S. H. (1973). The effect of
hypothyroidism and thyrotoxicosis on thyroxine metabolism in therat. Endocrinology, 92, 1028-1033.
Deems, D. A., Doty, R. L., Settle, R. G., Moore-Gillon, V., Shaman,P., Mester, A. F., Kimmelman, C. P., Brightman, V. I., & Snow, I. B.(1991). Smell and taste disorders: A study of 750 patients from theUniversity of Pennsylvania Smell and Taste Center (1981-1986).Archives of Otolaryngology, Head and Neck Surgery, 117; 519-528.
Received June 13, 1991Revision received July 13, 1992
Accepted July 13, 1992 .