induction cytolytic t-and b-cell responses against ... fileagainst different influenza aviruses...
TRANSCRIPT

Vol. 28, No. 3INFECTION AND IMMUNITY, June 1980, p. 799-8110019-9567/80/06-0799/13$02.00/0
Induction of Cytolytic T- and B-Cell Responses AgainstInfluenza Virus Infections
D. ARMERDING* AND H. ROSSITER
Immunology Department, Sandoz Forschungs Institut GmbH, Vienna, Austria
Inoculation of mice with live influenza virus results in the induction of cytotoxicthymus-derived (T) lymphocytes and of bone marrow-derived (B) cells producingantiviral antibodies. An assay system was developed to evaluate both types ofimmune responses on a cellular basis within the same lymphocyte pool with noneed to separate out the different effector cell classes. The test system representeda modification of the 51Cr-release assay. T-cell activity was measured exclusivelyin the absence of active complement using targets that were compatibile fordeterminants encoded by the mouse major histocompatibility gene complex, H-2.H-2-different and even xenogeneic target cells were lysed in the presence of eithernon-inactivated fetal calf serum or normal rabbit serum as a complement source.Cytotoxicity was mediated in the latter case by direct interaction of B-cell-produced immunoglobulin directed to viral antigens expressed by the target celland complement. Antibody-dependent cell-mediated cytotoxicity mechanisms didnot contribute to cytotoxicity in the test system described. It was demonstratedthat the cytolytic B-cell responses of one particular strain of mice (BALB/c)against different influenza A viruses were restricted to the immunizing virus onthe effector cell level. In another strain of mice (C3H), B cells revealed a broadcross-reactive response resembling that of killer T cells.
Immunization of mice with influenza viruseswill induce specific T- and B-lymphocyte re-sponses. In recent years both effector functionshave been studied quite extensively, though sep-arately. Activity of cytotoxic T cells was meas-ured predominantly by use of the 51Cr releaseassay (7). B-cell responses were assayed inde-pendently by estimation of antiviral immuno-globulin levels in the serum or other body fluidsusing standard serological tests (2, 35). B-lym-phocyte responses to influenza viruses have alsobeen analyzed, at least in mice, on the cellularlevel using a hemolytic plaque assay, a splenicfocus technique, and cell transfers (8, 24, 36).Cytotoxic T cells are measured by their activityagainst virus-encoded products on target cellsurfaces, whereas anti-influenza antibodies canoften only be detected if reacting with determi-nants on the envelope of free viruses. Hence itwas shown that type A influenza viruses inducehighly cross-reactive T cells which recognizedeterminants of virus antigens on the target cellsurfaces induced by influenza A subtypes asbeing identical or at least closely related (4, 9,12, 42, 44, 45). The group-specific matrix (M)protein is under discussion as one causativeagent for the cross-reactivity (1, 3, 5). Protectiveantibodies in humans and mice to influenza vi-ruses have been described to be directed pre-dominantly against determinants on the hemag-glutinin or the neuraminidase subunits, or both,
of the virion (reviewed in 20, 33, 39). Not muchis known about the induction of cross-reactiveB-cell products during the course of an influenzainfection. Although some evidence exists thatinternal virus proteins do induce B-cell-me-diated immune functions (40), other studies in-dicate instead the absence of an antibody re-sponse against the M protein during a primaryinfection (31, 32, 37).The biological significance of the existence of
cross-reactive T and B lymphocytes becomesobvious when one asks the question whether ornot these lymphocyte populations might be ableto protect against influenza virus infections in adrift or even a shift situation. It is important toknow whether either immunocyte populationalone, T or B cells, or both together, present aprotective mechanism enabling the organism tofight the infection. For vaccination purposes, itwould be necessary to know whether or not theimmunization procedure results in a cross-reac-tive response at all.The aim of this study was to provide the
means for measuring T- and B-cell responses inparallel in an easy way using the same assaysystem. It has been amply demonstrated that inmice antiviral activity of cytotoxic T cells isrestricted by products of the major histocom-patibility gene complex, H-2 (10, 12, 13, 15, 16,22, 23, 30, 34, 43, 44), whereas cytolytic activityof B-cell products is not. Use of H-2-identical or
799
on May 14, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

800 ARMERDING AND ROSSITER
-different effector-target combinations in the as-say should allow discrimination between bothtypes of responses. It will be shown here thatlysis of syngeneic virus-infected targets in theabsence of active complement (C') representsexclusively immune T-cell activity, whereas de-struction of allogeneic or xenogeneic target cellsin the presence of active complement caused bylymphocytes within the same cell pool is due toB-cell activity. The optimal conditions for dem-onstrating T- and B-cell-mediated cytotoxic ac-tivity in parallel will be worked out.
MATERIALS AND METHODSAnimals. BALB/c, CBA/J, C57Br, and C3H/Tif
mice were obtained from GI. BomholtgArd Ltd. (Ry,Denmark). DBA/2, DBA/1, SWR/J, C57BL/6, andC57BL/10 mice were derived from Jackson Labora-tories (Bar Harbor, Maine). C3H/He Han mice werepurchased from Zentralinstitut fur Versuchstierzucht(Hannover, Germany). Only male mice aged 10 weeksor older were used.
Cell lines. Exponentially growing L929 fibroblasts(L), P815 mastocytoma cells (P), 3T3 cells, neuro 2Aneuroblastoma cells (Neuro 2A), and human hepa-toma (HEP) cells, all maintained in tissue culture,were used as target cells in the cytotoxicity assays.Neuro 2A and 3T3 cells have been typed by us for H-2 (not published). Neuro 2A express H-2k' d, and 3T3cells express H-2q determinants recognizable by cyto-toxic T lymphocytes.
Viruses. AOPR8 (A/PR8/34) (HON1), A/NJ (A/New Jersey/76) (HswlNl), A2/Asia (A/Asia/57)(H2N2), and A/HK (A/Hong Kong/68) (H3N2) weregrown in 10-day embryonated eggs and stored as al-lantoic fluid at -700C.
Immunizations. Mice were sensitized with livevirus by the intravenous route. Doses of virus neces-sary for induction of significant cytolytic responseswere estimated for each lot. Mice usually received 0.5ml of a 1:5 to 1:20 diluted virus preparation corre-sponding to about 150 international units (IU) permouse. For primary responses, spleens were removedfrom at least three mice on day 5 or 6 and pooled asspleen cell suspensions.
Cell separation techniques. Preparation of anti-thyl.2 antiserum, estimation of anti-thyl.2 cytotoxic-ity, and anti-thyl.2 plus complement treatment ofspleen cells have been previously described (19). Pu-rification of splenic T lymphocytes was done as de-scribed by Julius et al. (18) using nylon wool columns.
Cytotoxicity assay. A sample of 5 x 106 targetcells was labeled in suspension with 100,Ci of 5'Cr(sodium chromate, Amersham, Buckinghamshire,England; no. CJS 1; specific activity, 100 to 400 mCi/mg of Cr) in 100,l of culture medium (Eagle minimumessential medium plus pyruvate, nonessential aminoacids, and glutamine as described in reference 11),infected with influenza virus simultaneously for 1.5 hat 37°C, washed twice, and dispensed into round-bot-tom microtiter plates (Flow Laboratories GmbH,Bonn, Germany) at a concentration of 1 x 104 or 2 x104 cells per well in 100 td of culture medium. Eachvirus preparation was titrated previously to yield op-
timal results in the T-cell-mediated cytotoxicity assayas described below. Amount of virus varied between100 and 400 1I of undiluted allantoic fluid, correspond-ing to about 300 IU/ml per 5 x 106 cells. Labeled andinfected target cells (or noninfected cells for controls)were cultured in an atmosphere of 10% C02, 8% 02,and 82% N2 for about 4 h at 37°C before addition ofeffector cells. Normal or virus-immune spleen cellswere added in a further 100 ,l of medium. Culturesupernatants were collected using the Titertek super-natant harvester (Flow Laboratories GmbH). Cultureswere terminated either 4 to 6 h (short-term assay) or12 to 16 h (long-term assay) after addition of effectorcells and incubation at 37°C. Hence, long-term incu-bations resulted in a high spontaneous 5'Cr release.The test system was modified, therefore, by loweringthe temperature, after short-term incubation at 37°C,to 30°C. Culture supernatants were collected in thatcase after overnight incubation (total incubation of 14to 16 h). Specific cytotoxicity was equivalent to thatof 10 to 12 h of incubation at 37°C or higher with abackground cytolysis between 30 and 40%. The culturemedium was supplemented with either 10% non-inac-tivated fetal calf serum (FCS; lot. no. L985101S, FlowLaboratories) or heat-inactivated FCS (30 min at56°C) or was used with no serum supplement at all asindicated in the text.
Results are expressed as percent specific 5tCr releasecalculated from the geometric mean of the counts perminute (cpm) of four replicas using the formula: (E-LC)/(HC-LC) x 100, where E is the total 5tCr releasedby the experimental groups (sensitized effector cells),LC is the low control determined by the activity ofnormal spleen cells, and HC represents the amount ofisotope released from the target cells after 10 to 15min of incubation with 10% Triton X-100 (Serva Fein-biochemica, Heidelberg, Germany) at 37°C. Standarderror of the mean in all experiments was usually lessthan 1.05.
Sensitized or normal spleen cells never showed anysignificant cytotoxicity (higher than that of medium)against noninfected targets. The respective data weretherefore omitted from the results. Furthermore,spleen cells from mice immunized with allantoic fluidalone did not express cytolytic activity against influ-enza virus-infected target cells. Target cells incubatedwith allantoic fluid were not lysed by spleen cells frommice sensitized either with virus containing allantoicfluid or with allantoic fluid alone, (data not shown).Specificity controls using influenza B viruses, as wellas other viruses such as herpes simplex 1 and 2 orvaccinia, for immunization or target cell infection didnot reveal any cross-reactivity on induction or effectorcell level with influenza A strain viruses (data notshown). Experiments were usually performed morethan twice (up to 12 times).
Inhibition of cytotoxic assays. Aggregated bo-vine gamma globulin (Serva Feinbiochemica) and ag-gregated human gamma globulin (Serva Feinbiochem-ica) were used at concentrations ranging from 12.5 to200 ig/ml. A 50-ul volume was added to each culturewell. Aggregation was done at a concentration of 1mg/mil in phosphate-buffered saline at 600C for 1 h.Protein A (Staphylococcus aureus, Pharmacia FineChemicals, Uppsala, Sweden) was used at concentra-tions ranging from 12.5 to 200 pg/mI, 50 ,l per culture
INFECT. IMMUN.
on May 14, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

CYTOLYTIC T- AND B-CELL RESPONSES 801
well. Sheep erythrocytes were coated with subagglu-tinating amounts of rabbit anti-sheep erythrocyte im-munoglobulin and served as inhibitors at concentra-tions between 107 and 108 cells per well. Rabbit anti-bovine serum albumin and rabbit anti-mouse immu-noglobulin hyperimmune antisera (a gift from E. Liehl,Sandoz Forschungs Institut) were used at dilutions of1:10 and 1:50, 50 td per well, or as indicated in the text.Antisera were inactivated for 30 min at 560C andextensively absorbed before use with mouse spleencells in the case of the anti-bovine serum albuminantiserum, and with mouse thymus cells in the case ofanti-mouse immunoglobulin antisera. Anti-bovine se-rum albumin immune serum was tested for inhibitoryactivity in cytotoxic assays in medium containing 10%FCS. All inhibitors were added immediately after dis-pensing of the effector cells to the culture wells andwere left there throughout the entire culture period.Complement. Normal rabbit sera were tested for
natural cytotoxicity against 5"Cr-labeled P815 or L929target cells. We chose the sera that had the lowestbackground toxicity but, after absorption with normalmouse spleen cells, the highest complement activity inantibody-mediated cytotoxicity against virus-infectedtarget cells. Optimal concentration for each comple-ment lot was estimated in the appropriate assay sys-tem. For experiments performed here, complementdilutions between 1:5 and 1:20 in medium were used(25 pi per well). Complement was stored at -70'C insmall aliquots and never thawed or frozen more thantwice. In most of the experiments presented here, non-inactivated FCS served as a complement source. TheFCS used had no background cytotoxicity as comparedto serum-free medium and was used without furthertreatment at a 10% concentration in medium.
RESULTS
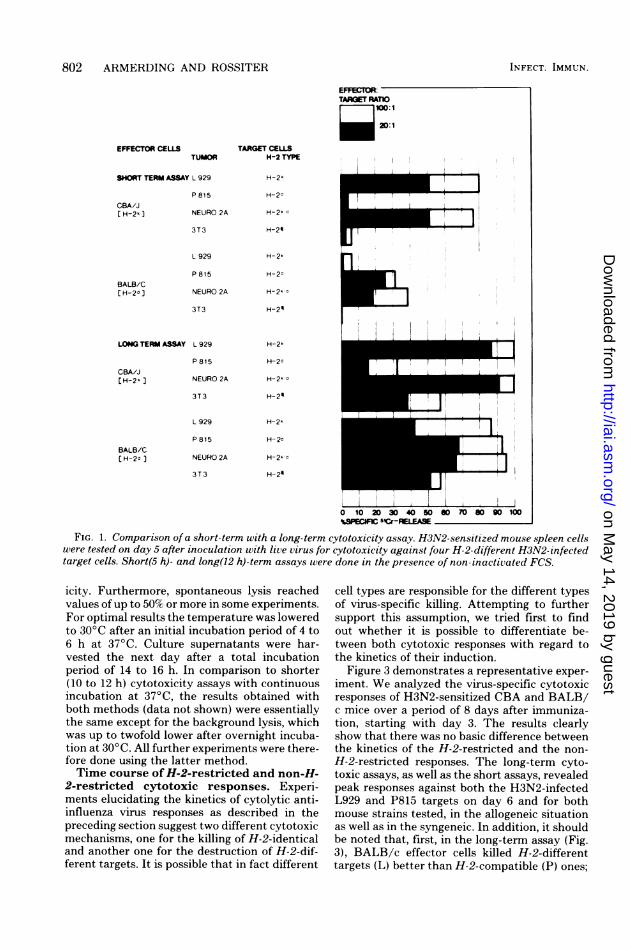
Cytotoxic responses of various mousestrains against virus-infected H-2-compat-ible or H-2-incompatible target cells. Virus-sensitized mouse T lymphocytes will only lysetarget cells infected with related or homologousvirus preparations if both effector and targetcells share the K or D region of the H-2 genecomplex, or both, and express the respectivegene products. This observation has been de-scribed by many authors (10, 12, 13, 15, 16, 22,23, 30, 34, 43, 44). The assay systems usuallyinvolve a short-term incubation period (around4 h) for the effector and target cell mixture andabsence of active complement. Our results con-firmed these findings even if we used non-inac-tivated FCS as medium supplement. Figure 1demonstrates typical results obtained in a short5"Cr release assay using H3N2-sensitized CBA/J (H-2 k) and BALB/c (H-2d) effector cells andfour different target cells infected with the samevirus. It is obvious from the results that signifi-cant cytotoxicity expressed as percent specific5"Cr release only occurred in H-2-compatibleeffector-target combinations, L929 (H-2k) forCBA/J and P815 (H-2d) for BALB/c lympho-
cytes. 3T3 (H-2") target cells, different fromboth effector cell populations with respect to H-2, were not lysed. However, Neuro 2A cells,which are H-2k d, were killed by BALB/c as wellas by CBA immunocytes. Whereas the short-term assay reiterated the H-2 restriction as re-ported by others, responses were no longer de-pendent upon H-2 expression of the target cellsafter a long-term incubation. CBA and BALB/ceffector cells were cytotoxic for H-2-compatibleand for H-2-incompatible targets as well.
Induction of non-H-2-restricted cytotoxic ac-tivity of virus-immune mouse lymphocytes wasalso tested in a broader range of different mousestrains using a human cell line in addition to thetarget cells studied in the preceding experiment(Table 1). Only data from a long-term cytotox-icity assay done in the presence of non-inacti-vated FCS are given. The data clearly demon-strate the absence ofany H-2-restricted cytolyticresponse. All H3N2-infected mouse tumor targetcells were lysed by all effector cell populations(H3N2 sensitized), regardless of whether or notthe H-2 types matched. Moreover, human HEPcells were also destroyed to a highly significantdegree by all immune mouse cells.Kinetics of cytotoxic responses in H-2-
compatible or H-2-incompatible effector-target cell combinations. It has been dem-onstrated in the preceding section that long-term assays in the presence of non-inactivatedFCS in contrast to short-term incubations nolonger yield H-2-restricted cytotoxic antiviralresponses. It was important to find out at whichtime of the assay cytolysis in H-2-different, incomparison to H-2-compatible, effector-targetcell combinations would start, and whether therewas a significant difference in the kinetics ofboth cytotoxic responses. Figure 2 demonstratesthe results obtained with H3N2-immune spleencells from CBA and DBA/2 mice tested over aperiod of 12 h for cytotoxicity against H3N2-infected L929 (H-2k) and P815 (H-2d) targetcells. The following results are of interest: (i)specific 5"Cr release mediated by both effectorcell preparations in both target cell populationsaccumulated continuously, at a similar rate, overthe whole incubation period; (ii) onset of thelysis of H-2-incompatible targets was delayed,about 4 h for the DBA/2-L cell combinationand about 6 h for the CBA-P815 combination,if a 10% 5"Cr release is considered to be thesignificance level of the assay; (iii) highest spe-cific 51Cr release values were reached for allcultures at 12 h. Incubation periods for longerthan 12 h at 370C resulted in an increased rateof the spontaneous isotope release, higher thanthat induced by immune cells (data not shown).This causes a false decrease of specific cytotox-
VOL. 28, 1980
on May 14, 2019 by guest
http://iai.asm.org/
Dow
nloaded from
802 ARMERDING AND ROSSITER
EFFECTOR:TARGET RATIO
20:1
EFFECTOR CELLSTUMO
TARGET CELLSH-2 TYPE
SHORT TERM ASSAY L 929 H-2'
P 815 H-2aCBA/JCH-2k] NEURO 2A H-2k a
3T3 H-24
L 929 H-2k
P 815 H- 2BALB/CCH-2 J NEURO 2A H-2i a
3T3 H-24
LONG TERM ASSAY L 929 H-2k
P 815 H -2CBA/J[H-2] NEURO 2A H-2k a
3T3 H-24
L 929 H-2k
P 815 H- 2BALB/CC H-2d] NEURO 2A H-2 a
3T3 H- 2
1:
_ i _~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~_5:L~~~~~~~
0 1o 20 30 40 50 so to so 90 1oo%SPAC 5'Cr-RELEA.SE
FIG. 1. Comparison of a short-term with a long-term cytotoxicity assay. H3N2-sensitized mouse spleen cellswere tested on day 5 after inoculation with live virus for cytotoxicity against four H-2-different H3N2- infectedtarget cells. Short(5 h)- and long(12 h)-term assays were done in the presence of non-inactivated FCS.
icity. Furthermore, spontaneous lysis reachedvalues of up to 50% or more in some experiments.For optimal results the temperature was loweredto 30'C after an initial incubation period of 4 to6 h at 37"C. Culture supernatants were har-vested the next day after a total incubationperiod of 14 to 16 h. In comparison to shorter(10 to 12 h) cytotoxicity assays with continuousincubation at 370C, the results obtained withboth methods (data not shown) were essentiallythe same except for the background lysis, whichwas up to twofold lower after overnight incuba-tion at 30'C. All further experiments were there-fore done using the latter method.Time course of H-2-restricted and non-H-
2-restricted cytotoxic responses. Experi-ments elucidating the kinetics of cytolytic anti-influenza virus responses as described in thepreceding section suggest two different cytotoxicmechanisms, one for the killing of H-2-identicaland another one for the destruction of H-2-dif-ferent targets. It is possible that in fact different
cell types are responsible for the different typesof virus-specific killing. Attempting to furthersupport this assumption, we tried first to findout whether it is possible to differentiate be-tween both cytotoxic responses with regard tothe kinetics of their induction.
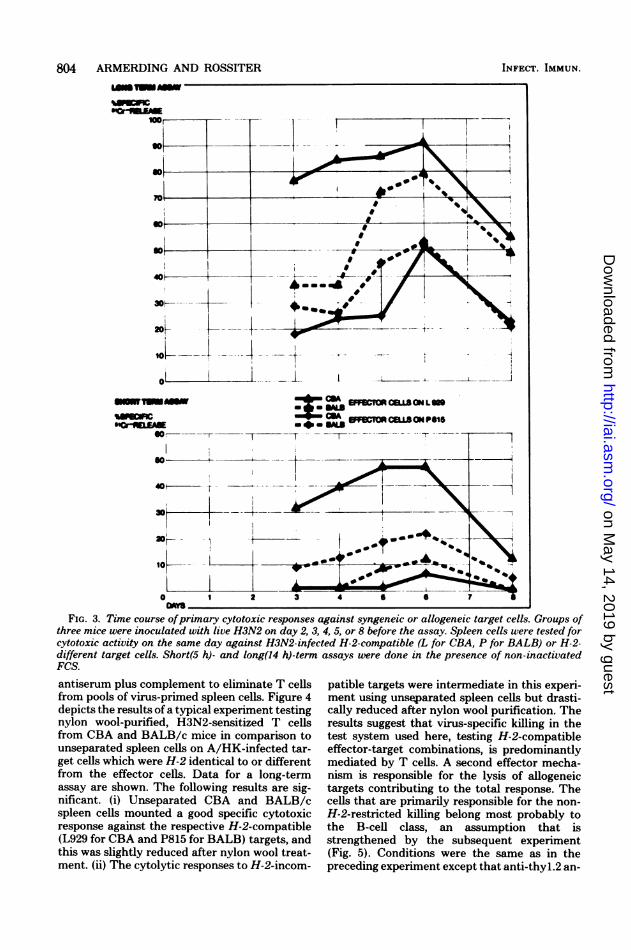
Figure 3 demonstrates a representative exper-iment. We analyzed the virus-specific cytotoxicresponses of H3N2-sensitized CBA and BALB/c mice over a period of 8 days after immuniza-tion, starting with day 3. The results clearlyshow that there was no basic difference betweenthe kinetics of the H-2-restricted and the non-H-2-restricted responses. The long-term cyto-toxic assays, as well as the short assays, revealedpeak responses against both the H3N2-infectedL929 and P815 targets on day 6 and for bothmouse strains tested, in the allogeneic situationas well as in the syngeneic. In addition, it shouldbe noted that, first, in the long-term assay (Fig.3), BALB/c effector cells killed H-2-differenttargets (L) better than H-2-compatible (P) ones;
IN FECT. IMMUN.
on May 14, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

CYTOLYTIC T- AND B-CELL RESPONSES 803
TABLE 1. Cytotoxic activity of various inbred mouse strains sensitized with A/HK against different A/HK-infected target cells'
Effector cells % Specific 5'Cr release
L929 (H-2k) P815 (H-2d) 3T3 (H-2q) HEPMouse strain H-2 type
100:1b 20:1 100:1 20:1 100:1 20:1 100:1 20:1
C57BL/6J H-2b 53.6 34.1 47.8 36.4 86.5 77.4 27.6 17.6C57BL/LOJ H-2b 37.8 31.9 35.0 26.2 82.8 65.6 28.3 16.1BALB/c H-2d 39.6 27.8 60.8 40.5 81.5 57.9 19.3 13.5DBA/2 H-2d 46.5 29.3 53.7 28.0 89.2 60.6 27.5 15.0CBA/J H-2k 66.4 42.7 21.9 15.0 75.1 49.1 17.2 14.1C3H/Tif H-2k 45.2 45.4 29.5 16.6 75.1 49.0 29.4 16.2C57Br H-2k 58.2 48.4 43.2 17.0 92.7 58.3 18.4 0.0DBA/1 H-2q 48.3 44.2 17.0 13.7 70.5 70.7 17.4 10.9SWR/J H-2q 45.4 26.1 30.2 13.5 85.4 69.9 19.3 14.3
a Mice were sensitized with live H3N2 5 days before. Assay was done in the absence of non-inactivated FCS.Long-term incubation (see the text). Responses of H-2-compatible effector-target combinations are shown inboldface type.
b Effector:target cell ratio.
1: ,,00.0-- -!90p .,.'p6 -
p.1140
TI. LHUR3JFIG. 2. Kinetics of cytotoxic responses against syngeneic or allogeneic target cells. H3N2-sensitized mouse
spleen cells were tested on day 6 after inoculation with live virus for cytotoxicity against H3N2-infected H-2-compatible (L for CBA, P for DBA/2) or H-2-different target cells. Assays were done in the presence of non-inactivated FCS. Cultures were terminated after 2, 4, 6, 8, 10, and 12 h.
second, a low allogeneic response could also beobserved on day 6 in the short-term system (Fig.3).Identification of cell types responsible
for virus-specific cytotoxic responses. To
identify the cellular basis of the H-2-restrictedand the non-H-2-restricted cytotoxicity ob-served in our assay system, we designed cellseparation experiments. We used nylon woolcolumns to enrich for T cells and anti-thyl.2
VOL. 28, 1980
O'Cr-FABIAK100I
on May 14, 2019 by guest
http://iai.asm.org/
Dow
nloaded from
804 ARMERDING AND ROSSITERn| -
10L 1
0-i-* 1
smurne
O'0r._EA
CDA EFFOIONCO I O L N
F CI ON r
It =I0
-7
10
0 1 2 3 4 6 6 7
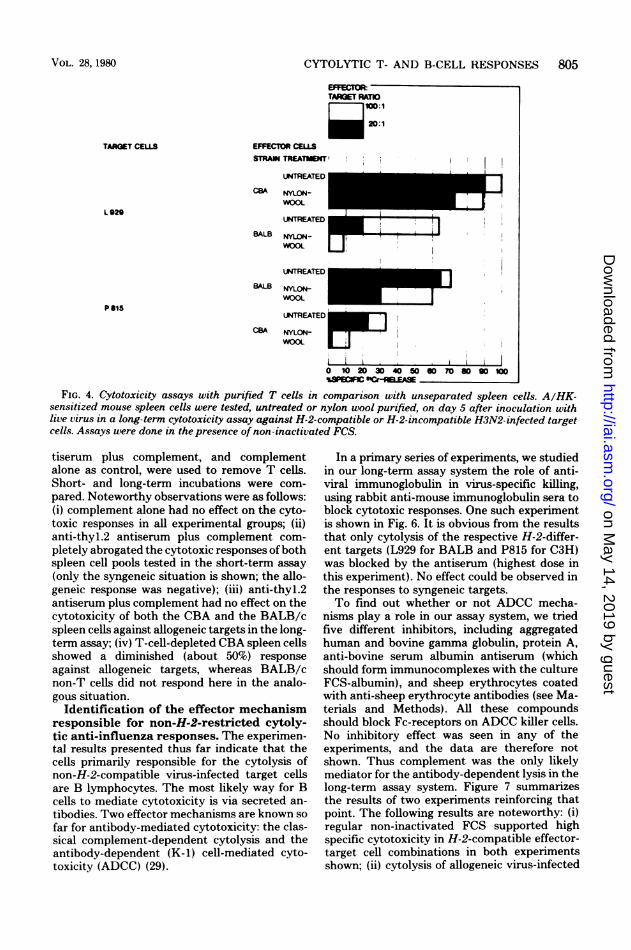
FIG. 3. Time course ofprimary cytotoxic responses against syngeneic or allogeneic target cells. Groups ofthree mice were inoculated with live H3N2 on day 2, 3, 4, 5, or 8 before the assay. Spleen cells were tested forcytotoxic activity on the same day against H3N2-infected H-2-compatible (L for CBA, P for BALB) or H-2-different target cells. Short(5 h)- and long(14 h)-term assays were done in the presence of non-inactivatedFCS.antiserum plus complement to eliminate T cellsfrom pools of virus-primed spleen cells. Figure 4depicts the results of a typical experiment testingnylon wool-purified, H3N2-sensitized T cellsfrom CBA and BALB/c mice in comparison tounseparated spleen cells on A/HK-infected tar-get cells which were H-2 identical to or differentfrom the effector cells. Data for a long-termassay are shown. The following results are sig-nificant. (i) Unseparated CBA and BALB/cspleen cells mounted a good specific cytotoxicresponse against the respective H-2-compatible(L929 for CBA and P815 for BALB) targets, andthis was slightly reduced after nylon wool treat-ment. (ii) The cytolytic responses to H-2-incom-
patible targets were intermediate in this experi-ment using unseparated spleen cells but drasti-cally reduced after nylon wool purification. Theresults suggest that virus-specific killing in thetest system used here, testing H-2-compatibleeffector-target combinations, is predominantlymediated by T cells. A second effector mecha-nism is responsible for the lysis of allogeneictargets contributing to the total response. Thecells that are primarily responsible for the non-H-2-restricted killing belong most probably tothe B-cell class, an assumption that isstrengthened by the subsequent experiment(Fig. 5). Conditions were the same as in thepreceding experiment except that anti-thyl.2 an-
INFECT. IMMUN.
on May 14, 2019 by guest
http://iai.asm.org/
Dow
nloaded from
CYTOLYTIC T- AND B-CELL RESPONSES
EFFECTOR:TARGET RATIO
20:120:1
TAoGT CELLS EFFECTOR CELLSSTRAIN TREATMENT
UNTREATEDCBA NYLON-
UNTREATED I
BALB NYLON-
WOOL Ul
UNTREATEC IBALB NYLON-
WOOL a _
UNTREATED
CBA NYLON- _LWOOL
i0 10 20 30 40 50 60 70 80 90 100RSPeCM HE
FIG. 4. Cytotoxicity assays with purified T cells in comparison with unseparated spleen cells. A/HK-sensitized mouse spleen cells were tested, untreated or nylon wool purified, on day 5 after inoculation withlive virus in a long-term cytotoxicity assay against H-2-compatible or H-2-incompatible H3N2-infected target
cells. Assays were done in the presence of non-inactivated FCS.
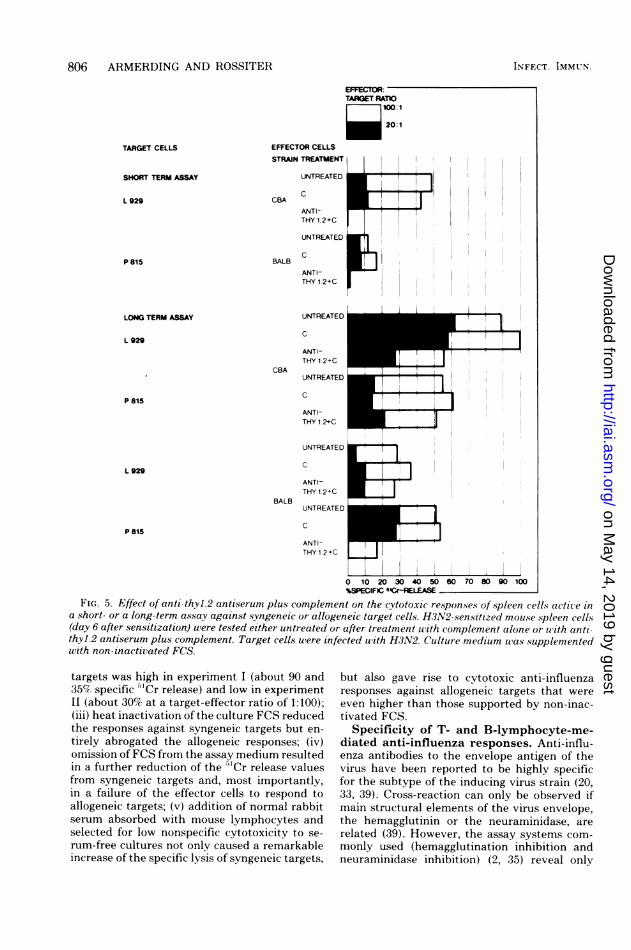
tiserum plus complement, and complementalone as control, were used to remove T cells.Short- and long-term incubations were com-
pared. Noteworthy observations were as follows:(i) complement alone had no effect on the cyto-toxic responses in all experimental groups; (ii)anti-thyl.2 antiserum plus complement com-
pletely abrogated the cytotoxic responses ofbothspleen cell pools tested in the short-term assay
(only the syngeneic situation is shown; the allo-geneic response was negative); (iii) anti-thyl.2antiserum plus complement had no effect on thecytotoxicity of both the CBA and the BALB/cspleen cells against allogeneic targets in the long-term assay; (iv) T-cell-depleted CBA spleen cellsshowed a diminished (about 50%) responseagainst allogeneic targets, whereas BALB/cnon-T cells did not respond here in the analo-gous situation.
Identification of the effector mechanismresponsible for non-H-2-restricted cytoly-tic anti-influenza responses. The experimen-tal results presented thus far indicate that thecells primarily responsible for the cytolysis ofnon-H-2-compatible virus-infected target cellsare B lymphocytes. The most likely way for Bcells to mediate cytotoxicity is via secreted an-
tibodies. Two effector mechanisms are known so
far for antibody-mediated cytotoxicity: the clas-sical complement-dependent cytolysis and theantibody-dependent (K-1) cell-mediated cyto-toxicity (ADCC) (29).
In a primary series of experiments, we studiedin our long-term assay system the role of anti-viral immunoglobulin in virus-specific killing,using rabbit anti-mouse immunoglobulin sera toblock cytotoxic responses. One such experimentis shown in Fig. 6. It is obvious from the resultsthat only cytolysis of the respective H-2-differ-ent targets (L929 for BALB and P815 for C3H)was blocked by the antiserum (highest dose inthis experiment). No effect could be observed inthe responses to syngeneic targets.To find out whether or not ADCC mecha-
nisms play a role in our assay system, we triedfive different inhibitors, including aggregatedhuman and bovine gamma globulin, protein A,anti-bovine serum albumin antiserum (whichshould form immunocomplexes with the cultureFCS-albumin), and sheep erythrocytes coatedwith anti-sheep erythrocyte antibodies (see Ma-terials and Methods). All these compoundsshould block Fc-receptors on ADCC killer cells.No inhibitory effect was seen in any of theexperiments, and the data are therefore notshown. Thus complement was the only likelymediator for the antibody-dependent lysis in thelong-term assay system. Figure 7 summarizesthe results of two experiments reinforcing thatpoint. The following results are noteworthy: (i)regular non-inactivated FCS supported highspecific cytotoxicity in H-2-compatible effector-target cell combinations in both experimentsshown; (ii) cytolysis of allogeneic virus-infected
L 920
P615
VOL. 28, 1980 80v'5
on May 14, 2019 by guest
http://iai.asm.org/
Dow
nloaded from
806 ARMERDING AND ROSSITER
EFFEIR:
TARGET RATIOI l0:1
_020:1
TARGET CELLS
SHORT TERM ASSAY
L929
P 815
EFFECTOR CELLS
CCBA
ANTI-THY 1.2+C
BALBANTI-THY 1 2+C
UNTREATED
C
ANTI-THY 1 2+C0
CBAUNTREATED
CANTI-THY 1. 2+C
UNTREATED
CANTI-THY 1.2+C
BALBUNTREATED
ANTI-
ANTI I 1THY12C |
0 10 20 30 40 50 60 70 80 90 100%SPECIFC 5'rLE
FIG. 5. Effect of anti-thyl.2 antiserum plus complement on the cytotoxic responses of spleen cells active in
a short- or a long-term assay against syngeneic or allogeneic target cells. H3N2-sensitized mouse spleen cells(day 6 after sensitization) were tested either untreated or after treatment with complement alone or with anti-thyl.2 antiserum plus complement. Target cells were infected with H3N2. Culture medium was supplementedwith non-inactivated FCS.
targets was high in experiment I (about 90 and35Cr specific "'Cr release) and low in experimentLI (about 309 at a target-effector ratio of 1:100);(iii) heat inactivation of the culture FCS reducedthe responses against syngeneic targets but en-
tirely abrogated the allogeneic responses; (iv)omission of FCS from the assay medium resultedin a further reduction of the '1Cr release valuesfrom syngeneic targets and, most importantly,in a failure of the effector cells to respond toallogeneic targets; (v) addition of normal rabbitserum absorbed with mouse lymphocytes andselected for low nonspecific cytotoxicity to se-rum-free cultures not only caused a remarkableincrease of the specific lysis of syngeneic targets,
but also gave rise to cytotoxic anti-influenzaresponses against allogeneic targets that wereeven higher than those supported by non-inac-tivated FCS.
Specificity of T- and B-lymphocyte-me-diated anti-influenza responses. Anti-influ-enza antibodies to the envelope antigen of thevirus have been reported to be highly specificfor the subtype of the inducing virus strain (20,33, 39). Cross-reaction can only be observed ifmain structural elements of the virus envelope,the hemagglutinin or the neuraminidase, arerelated (39). However, the assay systems com-
monly used (hemagglutination inhibition andneuraminidase inhibition) (2, 35) reveal only
LONG TERM ASSAY
L 929
P 81S
L 929
P 815 B
INFECT. IMMUN
iI
II
11
on May 14, 2019 by guest
http://iai.asm.org/
Dow
nloaded from
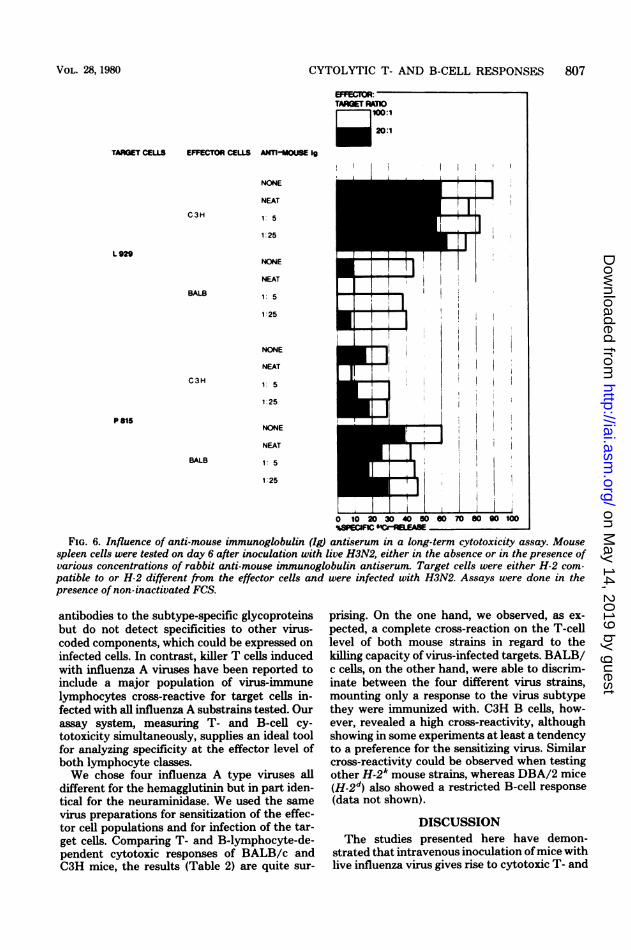
CYTOLYTIC T- AND B-CELL RESPONSES 807
EREC1VOR:TARGET RATIO
mb0:1
20O:1
TARGET CELLS EFFECTOR CELLS AMTI-MOUSE 19
C3H
L 929
BALB
C3H
palS
BALB
NONE
NEAT
1: 5
1:25
NONE
NEAT
1: 5
1:25
NONE
NEAT
1: 5
1:25
NONE
NEAT
1: 5
1:25
A~~~~~~~~i I!_ ,~~~
1o 0 so 40 100oloSUR F __E.-
FIG. 6. Influence of anti-mouse immunoglobulin (Ig) antiserum in a long-term cytotoxicity assay. Mousespleen cells were tested on day 6 after inoculation with live H3N2, either in the absence or in the presence ofvarious concentrations of rabbit anti-mouse immunoglobulin antiserum. Target cells were either H-2 com-patible to or H-2 different from the effector cells and were infected with H3N2. Assays were done in thepresence of non-inactivated FCS.
antibodies to the subtype-specific glycoproteinsbut do not detect specificities to other virus-coded components, which could be expressed oninfected cells. In contrast, killer T cells inducedwith influenza A viruses have been reported toinclude a major population of virus-immunelymphocytes cross-reactive for target cells in-fected with all influenza A substrains tested. Ourassay system, measuring T- and B-cell cy-totoxicity simultaneously, supplies an ideal toolfor analyzing specificity at the effector level ofboth lymphocyte classes.We chose four influenza A type viruses all
different for the hemagglutinin but in part iden-tical for the neuraminidase. We used the samevirus preparations for sensitization of the effec-tor cell populations and for infection of the tar-get cells. Comparing T- and B-lymphocyte-de-pendent cytotoxic responses of BALB/c andC3H mice, the results (Table 2) are quite sur-
prising. On the one hand, we observed, as ex-pected, a complete cross-reaction on the T-celllevel of both mouse strains in regard to thekilling capacity of virus-infected targets. BALB/c cells, on the other hand, were able to discrim-inate between the four different virus strains,mounting only a response to the virus subtypethey were immunized with. C3H B cells, how-ever, revealed a high cross-reactivity, althoughshowing in some experiments at least a tendencyto a preference for the sensitizing virus. Similarcross-reactivity could be observed when testingother H-2k mouse strains, whereas DBA/2 mice(H-2d) also showed a restricted B-cell response(data not shown).
DISCUSSIONThe studies presented here have demon-
strated that intravenous inoculation ofmice withlive influenza virus gives rise to cytotoxic T- and
VOL. 28, 1980
on May 14, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

808 ARMERDING AND ROSSITER
EFFECTOR:TARGET RAnO
~100:1
201
I~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~
EXPERIMENT MEDIUMSUPPLEMENT
BALB- EFFECTOR CELLSL 929
REGULAR FCS P 815
L 929
INACTIVATED FCS P 815
L 929
NO FCS
2I
P815
L 929
NO FCS. -NRS P 815
C 3 H-EFFECTOR CELLSL 929
REGULAR FCS P 815
L 929
INACTIVATED FCS P 815
L 929
P 815
-zszz
[zIzL 929
NO FCS -NRS P815
0 10 20 30 40 50 60 70 80 90 100%SPECIFIC 5rCr-RELEASE
FIG. 7. Role of active complement in the cytotoxic response to vtirus-infected syngeneic or allogeneic targetcells. H3N2-sensitized mouse spleen cells were tested on day 6 after inoculation with live virus for cytotoxicitvagainst H3N2-infected L929 or P815 target cells. Assay uas done in the presence of either non-inactivatedFCS, FCS inactivated for 30 min at 560C, no FCS, or normal rabbit serum. Medium supplements tere presentthroughout the long-term assay.
B-lymphocyte responses specific for virus-in-fected target cells. The kinetics of inductionappear to be identical for both responses, witha peak on day 6. The onset of cytolysis in theassay system was delayed for the B-cell-me-diated cytotoxicity as compared to that of Tlymphocytes. No cell separation was necessaryto evaluate T- and B-cell immunity. T-cell cy-
totoxicity against virus-infected targets was ab-solutely H-2 restricted in both short- and long-term assays, confirming the results of others (4,12, 13). There was no requirement for serum
substituents for killer T-cell activity. In contrast,to measure exclusively B-cell-dependent cyto-toxic mechanisms, two requirements must befulfilled: (i) allogeneic or xenogeneic cells whichexpress determinants of virus-encoded antigens,
and (ii) a complement source. It is possible touse either non-inactivated FCS or normal rabbitserum for complement. FCS works best if pres-ent throughout the whole assay (data notshown). Rabbit complement should be addedlate and for a restricted time period (I to 2 h) toreduce nonspecific cytotoxicity. Both comple-ment sources would have to be chosen for highcomplement activity but low background toxic-ity.
It has been shown here that it is B-cell im-munoglobulin, most likely to be secreted duringthe incubation period, that mediates the cyto-toxic response. The immunoglobulin may haveto reach a critical concentration before revealinga cytolytic response, and this could explain thedelayed onset of cytolysis. This assumption is
TARGET CELLS
]
Il
NO FCS
IN FFCT. IMMUN!9.
on May 14, 2019 by guest
http://iai.asm.org/
Dow
nloaded from
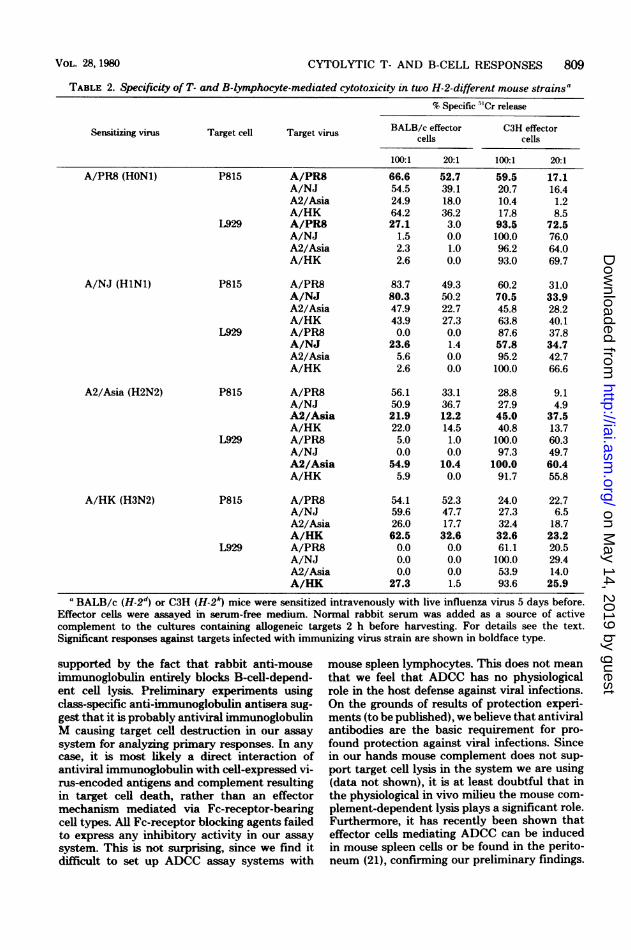
CYTOLYTIC T- AND B-CELL RESPONSES 809
TABLE 2. Specificity of T- and B-lymphocyte-mediated cytotoxicity in two H-2-different mouse strains"
% Specific "Cr release
Sensitizing virus Target cell Target virus cells CM effect
100:1 20:1 100:1 20:1
A/PR8 (HON1) P815 A/PR8 66.6 52.7 59.5 17.1
L929
A/NJ (HlNl)
A2/Asia (H2N2)
A/HK (H3N2)
P815
L929
P815
L929
P815
L929
A/NJA2/AsiaA/HKA/PR8A/NJA2/AsiaA/HK
A/PR8A/NJA2/AsiaA/HKA/PR8A/NJA2/AsiaA/HK
A/PR8A/NJA2/AsiaA/HKA/PR8A/NJA2/AsiaA/HK
A/PR8A/NJA2/AsiaA/HKA/PR8A/NJA2/AsiaA/HK
54.524.964.227.1
1.52.32.6
83.780.347.943.90.0
23.65.62.6
56.150.921.922.05.00.0
54.95.9
54.159.626.062.50.00.00.0
27.3
39.118.036.23.00.01.00.0
49.350.222.727.30.01.40.00.0
33.136.712.214.51.00.0
10.40.0
52.347.717.732.60.00.00.01.5
20.710.417.893.5100.096.293.0
60.270.545.863.887.657.895.2100.0
28.827.945.040.8100.097.3
100.091.7
24.027.332.432.661.1100.053.993.6
16.41.28.5
72.576.064.069.7
31.033.928.240.137.834.742.766.6
9.14.9
37.513.760.349.760.455.8
22.76.5
18.723.220.529.414.025.9
aBALB/c (H-2d) or C3H (H-2k) mice were sensitized intravenously with live influenza virus 5 days before.Effector cells were assayed in serum-free medium. Normal rabbit serum was added as a source of activecomplement to the cultures containing allogeneic targets 2 h before harvesting. For details see the text.Significant responses against targets infected with immunizing virus strain are shown in boldface type.
supported by the fact that rabbit anti-mouseimmunoglobulin entirely blocks B-cell-depend-ent cell lysis. Preliminary experiments usingclass-specific anti-imnmunoglobulin antisera sug-gest that it is probably antiviral immunoglobulinM causing target cell destruction in our assaysystem for analyzing primary responses. In anycase, it is most likely a direct interaction ofantiviral immunoglobulin with cell-expressed vi-rus-encoded antigens and complement resultingin target cell death, rather than an effectormechanism mediated via Fc-receptor-bearingcell types. All Fc-receptor blocking agents failedto express any inhibitory activity in our assaysystem. This is not surprising, since we find itdifficult to set up ADCC assay systems with
mouse spleen lymphocytes. This does not meanthat we feel that ADCC has no physiologicalrole in the host defense against viral infections.On the grounds of results of protection experi-ments (to be published), we believe that antiviralantibodies are the basic requirement for pro-found protection against viral infections. Sincein our hands mouse complement does not sup-port target cell lysis in the system we are using(data not shown), it is at least doubtful that inthe physiological in vivo milieu the mouse com-plement-dependent lysis plays a significant role.Furthermore, it has recently been shown thateffector cells mediating ADCC can be inducedin mouse spleen cells or be found in the perito-neum (21), confirming our preliminary findings.
VOL. 28, 19%0
on May 14, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

810 ARMERDING AND ROSSITER
It is therefore reasonable to assume that duringa viral infection cells effective in ADCC areinduced and play an analogous role to T killercells, especially in situations where a T killermemory might not be available, e.g., after dead-virus vaccination (6, 14). We plan to study theseproblems in further experiments.Another important question we asked was
whether or not T and B effector cells differ intheir specificity for cell-associated viral antigen,as suggested by studies on humoral antiviralantibodies on one hand (20, 33, 39) and by datapublished on killer T-cell activities on the other(4, 12, 13). In regard to the broad cross-reactivityof influenza-induced cytotoxic T cells, the pointhas been stressed by several authors that theinternal vision matrix protein might be the tar-get cell structure against which main T-cell ac-tivity is directed (1, 3, 5, 28). While this assump-tion still awaits final proof, it is at least safe toassume that M protein is immunogenic in mice(38) and that it is expressed on the surface ofsome cells after live virus infection (1, 3, 5, 28).It is also reasonable to assume that other inter-nal virion proteins (such as nucleoprotein) mightbe expressed by influenza-infected cells contrib-uting to cross-reactivity. An alternative expla-nation is that nonviral determinants-at leastnone specific for influenza virus-are also in-duced on the cell surface during infection, caus-ing cross-reactive immunity and serving as tar-get antigen in the assay. It has been shown, e.g.,that after mitogenic and also antigenic stimula-tion in vitro, lymphocytes would express tumorvirus determinants (26, 27, 40). Another possi-bility is that the infecting virus might causemodification of cell surface structures in a waythat they are recognized as foreign, a possibilitywhich is still under discussion for products ofthe H-2 gene complex (41). Whatever the basicmechanism causing T-cell cross-reactivityagainst influenza-infected target cells might be,B cells reveal a similar behavior. As shown herefor C3H mice, there is definite cross-reaction onthe B-cell level between target cells infectedwith influenza A strains that differ in regard tothe hemagglutinin and the neuraminidase sub-type. Although preference for the sensitizingvirus strain is expressed in some experiments,the response to unrelated viruses (in regard toH and/or N determinants) is highly significant.In contrast, BALB/c lymphocytes are quite dis-criminative in their killing activity in the pri-mary response, since they lyse only target cellsinfected with the immunizing virus. It is inter-esting that neuraminidase determinants, closelyrelated in two of the virus pairs used (HON1 andHsusNl, and H2N2 and H3N2), do not seem toplay a role in target cell lysis by sensitized B
lym hocytes here. In further experiments otherH-2 mice also expressed cross-reactivity whileDBA/2 mice did not. Further experiments arenecessary to elucidate the role of H-2-linked orpossibly H-2-unrelated genes in determining an-tibody specificity in the system.
Finally, it should be noted that in an assaysystem comparable to ours, where human pe-ripheral blood lymphocytes were tested for cy-totoxicity against influenza virus-infected targetcells, Greenberg et al. showed that the cell-me-diated cytotoxicity was of neither T nor B origin(17). Greenberg et al. concluded from their re-sults that their assay system measured ADCCactivity, since apparently cytotoxicity was alsocomplement independent. Hence it seems thatwith sensitized human lymphocytes, the 51Crrelease assay allows one to measure three differ-ent effector functions within the same cell pool:the HLA-restricted T-cell killing (25), the null-cell and antibody-dependent cytotoxicity, andthe in vitro B-lymphocyte-secreted antibody-and complement-dependent cytolysis. The latterwould require either early testing after in vivosensitization (day 4 to 6) or, better, cultivationand specific in vitro stimulation before the assayto test for B-cell memory.
ACKNOWLEDGMENTWe acknowledge the excellent technical assistance of An-
drea Hren.LITERATURE CITED
1. Ada, G. L., and K. L. Yap. 1977. Matrix protein ex-pressed at the surface of cells infected with influenzaviruses. Immunochemistry 14:643-651.
2. Aymard-Henry, M., M. T. Coleman, W. R. Dowdle,W. G. Laver, and R. G. Webster. 1973. Influenzavirus neuraminidase and neuraminidase-inhibition testprocedures. Bull. W.H.O. 48:199-202.
3. Biddision, W. E., P. C. Doherty, and R. G. Webster.1977. Antibody to influenza A matrix protein detects acommon antigen on the surface of cells infected withtype A influenza viruses of different subtypes. J. Exp.Med. 146:690-697.
4. Braciale, T. J. 1977. Immunological recognition of virus-infected cells. I. Generation of virus-specific and cross-reactive subpopulation of cytotoxic T cells in the re-sponse to type A influenza viruses of different subtypes.Cell. Immunol. 33:423-436.
5. Braciale, T. J. 1977. Immunologic recognition of influ-enza virus-infected cells. II. Expression of influenza Amatrix protein on the infected cell surface and its rolein recognition by crossreactive cytotoxic T cells. J. Exp.Med. 146:673-689.
6. Braciale, T. J., and K. L. Yap. 1978. Role of viralinfectivity in the induction of influenza virus-specificcytotoxic T cells. J. Exp. Med. 147:1236-1252.
7. Brunner, K. T., J. M. Mauel, J. C. Cerottini, and B.Chapuis. 1968. Quantitative assay of the lytic action ofimmune lymphoid cells on 5"Cr-labelled allogeneic tar-get cells in vitro: inhibition by isoantibody and by drugs.Immunology 14:181-196.
8. Cancro, M. P., W. Gerhard, and N. R. Klinman. 1978.The diversity of the influenza-specific primary B-cellrepertoire in Balb/c mice. J. Exp. Med. 147:776-787.
9. Doherty, P. C., R. B. Effros, and J. Bennink. 1977.
INFECT. IMMUN.
on May 14, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

CYTOLYTIC T- AND B-CELL RESPONSES 811
Heterogenity of the cytotoxic response of thymus-de-rived lymphocytes after immunization with influenzaviruses. Proc. Nati. Acad. Sci. U.S.A. 74:1209-1213.
10. Doherty, P. C., and R. M. Zinkernagel. 1976. Specificimmune lysis of paramyxovirus-infected cells by H-2compatible thymus-derived lymphocytes. Immunology31:27-32.
11. Eagle, H. 1959. Amino acid metabolism in mammaliancell cultures. Science 130:432-441.
12. Effros, R. B., P. C. Doherty, W. Gerhard, and J.Bennink. 1977. Generation of both cross-reactive andvirus-specific T-cell populations after immunizationwith serologically distinct influenza A viruses. J. Exp.Med. 145:557-568.
13. Ennis, F. A., W. J. Martin, M. W. Verboritz, and G.M. Butchko. 1977. Specificity studies on cytotoxic thy-mus-derived lymphocytes reactive with influenza virus-infected cells: evidence for dual recognition of H-2 andviral hemagglutinin antigen. Proc. Natl. Acad. Sci.U.S.A. 74:3006-3010.
14. Ertl, H. C. J., R. H. W. Gerike, and U. Koszinowski.1977. Virus-specific T-cell sensitization. Requirementfor vaccinia specific T-cell sensitization in vivo. Immu-nogenetics 4:515-522.
15. Gardner, L. D., N. A. Bowern, and R. U. Blanden.1975. Cell-mediated cytotoxicity against ectromelia vi-rus-infected target cells. III. Role of the H-2 gene com-plex. Eur. J. Immunol. 5:122-127.
16. Hale, A. H., N. W. Owen, D. Baltimore, and H. N.Eisen. 1978. Vesicular stomatitis virus glycoprotein isnecessary for H-2 restricted lysis of infected cells bycytotoxic T lymphocytes. Proc. Natl. Acad. Sci. U.S.A.75:970-974.
17. Greenberg, S. B., B. S. Criswell, H. R. Six, and R. B.Couch. 1977. Lymphocyte cytotoxicity to influenza vi-rus-infected cells. II. Requirement for antibody andnon-T-lymphocyte. J. Immunol. 119:2100-2106.
18. Julius, M. H., E. Simpson, and L. A. Herzenberg.1973. A rapid method for the isolation of functionalthymus-derived murine lymphocytes. Eur. J. Immunol.3:645-649.
19. Katz, D. H., and D. P. Osborne, Jr. 1972. The allogeneiceffect in inbred mice. II. Establishment of the cellularinteraction required for enhancement of antibody pro-duction by the graft-versus-host reaction. J. Exp. Med.136:455-465.
20. Kilbourne, E. D. 1975. Epidemiology of influenza, p. 483-538. In E. D. Kilbourne (ed.), The influenza viruses andinfluenza. Academic Press, New York.
21. Kohl, S., D. L. Cahall, D. L. Walters, and V. E. Schaff-ner. 1979. Murine antibody-dependent cellular cytotox-icity to herpes simplex virus-infected target cells. J.Immunol. 123:25-30.
22. Koszinowski, U., and H. Ertl. 1977. Cytotoxic interac-tions of virus specific effector cells with virus infectedtargets of different cell type. J. Immunogen. 4:107-114.
23. Koszinowski, U., and R. Thomssen. 1975. Target cell-dependent T cell mediated lysis of vaccinia virus-in-fected cells. Eur. J. Immunol. 5:245-251.
24. McLaren, C., G. E. Grubbs, and F. A. Ennis. 1979.Detection of cells producing surface antigen-specificantibody to influenza virus. J. Clin. Microbiol. 8:438-444.
25. McMichael, A. 1978. HLA restriction of human cytotoxicT lymphocytes specific for influenza virus. J. Exp. Med.148:1458-1467.
26. Moroni, C., and G. S. Schumann. 1975. Lipopolysac-charide induces C-type virus in short-term cultures ofBalb/c spleen cells. Nature (London) 254:60-61.
27. Moroni, C., G. Schumann, M. Robert-Guroff, and D.Martin. 1975. Induction of endogenous murine C-type
virus in spleen cell cultures tested with mitogens and 5-bromo-2'-deoxyuridine. Proc. Natl. Acad. Sci. U.S.A.72:535-538.
28. Oxford, J. S., and G. C. Schild. 1976. Immunologicaland physico-chemical studies of influenza matrix (M)polypeptides. Virology 74:394-402.
29. Pearson, G. P. 1978. In vitro and in vivo investigation onantibody-dependent cellular cytotoxicity. Curr. Top.Microbiol. Immunol. 80:65-96.
30. Pflzermaier, K., H. Jung, A. Starzinski-Powitz, M.Roliinghoff, and H. Wagner. 1977. The role of T cellsin antiherpes simplex virus immunity. I. Induction ofantigen-specific cytotoxic T lymphocytes. J. Immunol.3:939-944.
31. Potter, C. W., and J. S. Oxford. 1979. Determinants ofimmunity to influenza infection in man. Br. Med. Bull.35:69-75.
32. Schild, G. C., and W. R. Dowdle. 1975. Influenza viruscharacterization and diagnostic serology, p. 243-268. InE. D. Kilbourne (ed.), The influenza viruses and influ-enza. Academic Press, New York.
33. Schulman, J. L. 1975. Immunology of influenza, p. 373-393. In E. D. Kilbourne (ed.), The influenza viruses andinfluenza. Academic Press, New York.
34. Sethi, K. K., and H. Brandis. 1977. Specifically immunemouse T cells can destroy H-2 compatible target cellsinfected with herpes simplex virus type 1 or 2. Z. Im-munitaetsforsch. 150:162-173.
35. U.S. Department of Health, Education and Welfare.1975. Advanced laboratory techniques for influenza di-agnosis. Immunological series no. 6. U.S. Departmentof Health, Education and Welfare, Washington, D.C.
36. Virelizier, J. L. 1975. Host defenses against influenzavirus: the role of anti-hemagglutinin antibody. J. Im-munol. 115:434439.
37. Virelizier, J. L., A. C. Allison, and G. C. Schild. 1979.Immune responses to influenza virus in the mouse andtheir role in control of infection. Br. Med. Bull. 35:65-68.
38. Webster, R. G., and V. S. Hinshaw. 1977. Matrix pro-tein from influenza A virus and its role in cross-protec-tion in mice. Infect. Immun. 17:561-566.
39. Webster, R. G., and W. G. Laver. 1975. Antigenicvariation of influenza viruses, p. 269-314. In E. D.Kilbourne (ed.), The influenza viruses and influenza.Academic Press, New York.
40. Wecker, E., A. Schimpl, and T. Hunig. 1977. Expres-sion of Mu LV gp 71-like antigen in normal mousespleen cells induced by antigenic stimulation. Nature(London) 269:598-600.
41. Zinkernagel, R. M., and P. C. Doherty. 1974. Immu-nological surveillance against altered self componentsby sensitized T lymphocytes in lymphocytic choriomen-ingitis. Nature (London) 251:547.
42. Zinkernagel, R. M. 1976. H-2 restriction of virus-specificcytotoxicity across the H-2 barrier. J. Exp. Med. 144:933-945.
43. Zinkernagel, R. M., and P. C. Doherty. 1974. Restric-tion of in vitro T cell-mediated cytotoxicity in lympho-cytic choriomeningitis within a syngeneic or semiallo-geneic system. Nature (London) 248:701-703.
44. Zweerink, H. J., B. A. Askonas, D. Millican, S. A.Courtneidge, and J. J. Skehel. 1977. Cytotoxic Tcells to type A influenza virus; viral hemagglutinininduces A-strain specificity while infected cells confercrossreactive cytotoxicity. Eur. J. Immunol. 7:630-635.
45. Zweerink, H. J., S. A. Courtneidge, J. J. Skehel, M.J. Crumpton, and B. A. Askonas. 1977. Cytotoxic Tcells kill influenza virus infected cells but do not distin-guish between serologically distinct type A viruses. Na-ture (London) 267:354-336.
VOL. 28, 1980
on May 14, 2019 by guest
http://iai.asm.org/
Dow
nloaded from