inactivation of fhua at the cell surface of escherichia coli k-12 by a
TRANSCRIPT

Vol. 176, No. 15
Inactivation of FhuA at the Cell Surface of Escherichia coliK-12 by a Phage T5 Lipoprotein at the Periplasmic
Face of the Outer MembraneV. BRAUN,* H. KILLMANN, AND C. HERRMANN
Mikrobiologie II, Universitdt Tubingen, D-72076 Tubingen, Germany
Received 18 March 1994/Accepted 16 May 1994
Inactivation of phage T5 by lysed cells after phage multiplication is prevented by a phage-encodedlipoprotein (Llp) that inactivates the FhuA outer membrane receptor protein (K. Decker, V. Krauel, A.Meesmann, and K. Heller, Mol. Microbiol. 12:321-332, 1994). Using FhuA derivatives carrying insertions of 4and 16 amino acid residues and point mutations, we determined whether FhuA inactivation is caused bybinding of Llp to FhuA and which regions of FhuA are important for inactivation by Llp. Cells expressing Llpwere resistant not only to phage T5 but to all FhuA ligands tested, such as phage +80, colicin M, andalbomycin, and they were strongly reduced in the uptake of ferrichrome. Most of the FhuA derivatives whichwere not affected by Llp were, according to a previously published FhuA transmembrane topology model,located in periplasmic turns and in the TonB box close to the periplasm. Since the ligands bind to the cellsurface, interaction ofFhuA with Lip in the periplasm may induce a FhuA conformation which impairs bindingof the ligands. This conclusion was supported by the increase rather than decrease of colicin M sensitivity oftwo mutants in the presence of Llp. The only Llp-resistant FhuA derivatives with mutations at the cell surfacecontained insertions of 16 residues in the loop that determines the permeability of the FhuA channel and serves
as the principal binding site for all FhuA ligands. This region may be inactivated by steric hindrance in thata portion of Llp penetrates into the channel. Outer membranes prepared with 0.25% Triton X-100 from cellsexpressing Llp contained inactivated FhuA, suggesting Llp to be an outer membrane protein whose interactionwith FhuA was not abolished by Triton X-100. Llp solubilized in 1.1% octylglucoside prevented T5 inactivationby FhuA dissolved in octylglucoside.
Decker et al. (4) described a new mechanism of phage-encoded phage resistance which they designated lytic conver-
sion. Phage T5, a lytic phage, expressed a lipoprotein (Llp)which prevented binding of T5 to the outer membrane recep-
tor FhuA of viable cells. By this means, T5 within cells excludessuperinfection by T5. Inactivation of FhuA by Llp also pre-vents inactivation of released progeny T5 by binding to FhuAof lysed cells after T5 multiplication. We took advantage of thissystem to examine whether inactivation of FhuA involvesbinding of Llp and, if so, to determine which regions of FhuAare involved in Llp binding (periplasmic turns, transmembraneregions, or surface loops). Binding of Llp to surface regions ofFhuA would suggest inactivation by steric hindrance, whereasbinding to periplasmic regions would suggest inactivation byinduction of a conformational change in FhuA. In addition, wewanted to see whether the results obtained were consistentwith a previously published model describing the FhuA trans-membrane topology (14). The models proposed for the trans-membrane arrangement of the receptors FhuA (12, 14), FoxA(1), FepA (17, 20), and BtuB (2, 25) are similar in their basicoutline in that they predict 1-barrel structures containing turnsin the periplasm and larger loops at the cell surface, but theydiffer considerably in detail. Therefore, it is important toobtain additional data to test the models.
In order to determine interaction of Llp with FhuA, we usedpreviously isolatedfhuA mutants (12-14, 21) which showed fullor partial activity to the FhuA ligands (phages T5, Ti, and 480and colicin M, albomycin, and ferrichrome). Mutations which
* Corresponding author. Mailing address: Mikrobiologie II, Auf derMorgenstelle 28, D-72076 Tubingen, Germany. Phone: 7071-292096.Fax: 7071-294634.
rendered FhuA unresponsive to Llp defined sites of FhuAwhich are important for interaction with Llp. Among themutants tested were FhuA insertion derivatives which carriedtetrapeptides and hexadecapeptides inserted at various sites ofFhuA. The insertions rendered FhuA susceptible to degrada-tion by proteases which were added either to viable cells or tospheroplasts. Since cleavage of FhuA occurred within or closeto the peptide insertions, accessibility to proteases localizedthe insertion sites at the cell surface and in the periplasm,respectively. These data were used to predict the transmem-brane arrangement of FhuA (14). The model was supported bythe inactivity of an Asp-348 deletion mutant assigned to thelargest loop of FhuA at the cell surface (13). In addition,deletion of 34 amino acids in this loop converted FhuA into an
open channel which rendered cells sensitive to sodium dodecylsulfate (SDS) and bacitracin (12). The rate of ferrichromeuptake into the FhuA deletion mutant was no longer deter-mined by the TonB-dependent transport through the outermembrane but was diffusion controlled. In black lipid mem-branes, the FhuA deletion derivative formed stable channels ofa defined size three times as large as that of the channels of theporins, in contrast to wild-type FhuA, which failed to increasemembrane conductivity (12). It was concluded that wild-typeFhuA forms outer membrane channels which are closed by thelargest loop and opened by interaction with the TonB-ExbB-ExbD complex (6, 7, 12, 21). In this paper, we show thatmutations in periplasmic regions of FhuA confer resistance toLlp. This implies interaction of Llp with periplasmic regions ofFhuA which induces a conformational change in FhuA thatinactivates the ligand binding sites at the cell surface. Inaddition, insertions of 16 amino acid residues in the largestloop at the cell surface rendered FhuA resistant to Llp. Since
4710
JOURNAL OF BACrERIOLOGY, Aug. 1994, p. 4710-47170021-9193/94/$04.00+0Copyright © 1994, American Society for Microbiology
on January 31, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

FhuA INACTIVATION BY PHAGE T5 LIPOPROTEIN 4711
TABLE 1. E. coli K-12 strains and plasmids used
Strain or plasmid Genotype or phenotype Reference or source
StrainsH1443 aroB rpsL Alacl69 araD139 thi 9K0483 fliuA derivative of H1443 W. KosterAB2847 aroB malT tsx 9
PlasmidspWSK29 pSC101 ori; Ampr 24pSUKS1 pl5A ori; Cmr 18pHK763 pT7-6fnuA wild type 12pVK-L3 llp of phage T5 on pT7-3 4pHE101 llp of pVK-L3 on pSUKS1 This studypHE100 pWSK29 FhuA wild type This studypHE213 pWSK29 FhuA(19P) This studypHE214 pWSK29 FhuA(19S) This studypHE215 pWSK29 FhuA(D7G I9N) This studypHE102 pWSK29 FhuA3(L1O6P) This studypHE310 pWSK29 FhuA(L126R) This studypHE311 pWSK29 FhuA(D225Y) This studypHE103 pWSK29 FhuA4(AD348) This studypHE350 pWSK29 FhuA(E350V) This studypHE210 pWSK29 FhuA(A322-333 P-321 PDLA P-334) This studypHE312 pWSK29 FhuA(D395Y) This studypHE313 pWSK29 FhuA(R576G) This studypHE104 pWSK29 FhuA(RPL576-578ARC) This studypHE023-16 pWSK29 FhuA (G-23 LGTWNSRSPGIPGTQ P-24) This studypHE162-16 pWSK29 FhuA (K-162 EGGTAEFPWEFCGLPK E-163) This studypHE223-16 pWSK29 FhuA (P-223 PDPSSSTCRSSSTDLA D-224) This studypHE321-4 pWSK29 FhuA (P-321 PDLA A-322) This studypHE321-16 pWSK29 FhuA (P-321 PDPSSSTCRSSSTDLA A-322) This studypHE333-4 pWSK29 FhuA (A-333 PDLA P-334) This studypHE333-16 pWSK29 FhuA (A-333 PDPSSSTCRSSSTDLA A-334) This studypHE338-4 pWSK29 FhuA (G-338 TCRS H-339) This studypHE369-16 pWSK29 FhuA (D-369 LGTWNSRGSPGIPGTQ 1-370) This studypHE369-4 pWSK29 FhuA (D-369 DLQV 1-370) This studypHE405-16 pWSK29 FhuA (P-405 PDPSSSTCRSSSTDLA V-406) This studypHE531-16 pWSK29 FhuA (D-531 WVPGIPGDPREFQVPS R-532) This studypHE646-16 pWSK29 FhuA (P-646 PDPSSSTCRSSSTDLA A-646) This study
this loop determines the permeability of the FhuA channel, itmay bend into the channel, where it contacts Llp.
MATERIALS AND METHODSBacterial strains, plasmids, and media. The Escherichia coli
K-12 strains and plasmids used are listed in Table 1. Thestrains were grown in TY medium or in NB medium asdescribed previously (12). To reduce the available iron, the NBmedium was supplemented with 0.2 mM 2,2'-dipyridyl toobtain NBD medium. If required, the media contained theantibiotics ampicillin (40 ,ug/ml) and chloramphenicol (40pLg/ml).Plasmid pHE101 (Table 1) was constructed by ligating the
EcoRI-SalI fragment of pVK-L3 harboring the llp gene ofphage T5 (4) into EcoRI-SalI-cleaved pSUKS1. PlasmidspHE100, pHE102, pHE103, pHE104, and pHE350 were con-structed by ligating the EcoRI-HindIlI fhuA fragments ofpHK582, pHK102, pHK103, pHK104, and pHK350 (13), re-spectively, into EcoRI-HindIll-cleaved pWSK29. PlasmidspHE213, pHE214, and pHE215 were constructed by ligatingthe EcoRI-HindIII fragments of the pMc5-8 derivatives carry-ingfhuA TonB box mutations (21) into EcoRI-HindIII-cleavedpWSK29. Plasmids pHE023-16, pHE162-16, pHE223-16,pHE321-4, pHE321-16, pHE333-4, pHE333-16, pHE338-4,pHE369-4, pHE369-16, pHE405-16, pHE531-16, and pHE646-16 were constructed by ligating the XbaI-ClaI fhuA frag-
ments of the corresponding pSKF derivatives (14) into XbaI-ClaI-cleaved pWSK29. Plasmid pHE210 was constructed byligating the XbaI-ClaI flhuA fragment of pHK210 (12) intoXbaI-ClaI-cleaved pWSK29 (Table 1).
Recombinant DNA techniques. Isolation of plasmid DNA(27), use of restriction enzymes, ligation, transformation, aga-rose gel electrophoresis, and recovery of DNA from agarosewere done by standard techniques.
Site-directed mutagenesis. Single-nucleotide replacementswere introduced in fhuA by the gapped duplex DNA methodusing the pMa/c phasmid vectors as described elsewhere(23). The oligonucleotides used were as follows (mismatchesare underlined): p310, 5'-CGTATATGCGGGAACGCGC-3'; p311, 5'-CGTCCGATTATAAAACC-3'; p312, 5'-GGTTACGACIACTCTGTGC-3'; and p313,5'-CGAAGCGAAA£iGGCCGCTGTCG-3'. The resulting plasmids (pHK310,pHK311, pHK312, and pHK313) were digested with EcoRI-HindIII, and the resulting JhuA fragments were reclonedinto EcoRI-HindIII-cleaved pWSK29 to obtain the plasmidspHE310, pHE311, pHE312, and pHE313, respectively.
Phenotype assays. Sensitivity of cells to albomycin, colicinM, and phages T5 and 480 was tested by spotting 10-folddilutions (3 pAl) of the FhuA ligands onto TY plates overlaidwith 3 ml ofTY soft agar which contained 108 cells of the strainto be tested. When strains carrying the plasmid pHE101 weretested, the TY top agar contained 1 mM IPTG.
VOL. 176, 1994
on January 31, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

4712 BRAUN ET AL.
Growth promotion assay. Strain K0483 mutated in the aroBgene could not produce enterochelin and was therefore unableto grow on NBD agar plates. When K0483 transformed withthe various fhuA derivatives was able to use ferrichrome as aniron source, a growth zone developed around filter paper disks(diameter, 6 mm) loaded with the siderophore. NBD agarplates were overlaid with 3 ml of NB top agar containing 108cells of the strain to be tested. Filter paper disks supplementedwith 10 ,ul of 1, 0.3, and 0.1 mM ferrichrome solutions wereplaced on the agar plates and incubated at 37°C overnight. Totest strains carrying plasmid pHE101, the NB top agar con-tained 1 mM IPTG.
Transport assays. Cells grown on TY agar plates overnightat 37°C were washed in M9 salts (16)-0.4% glucose andadjusted to an optical density at 578 nm of 0.5 in M9salts-0.4% glucose at 37°C. [5 Fe3+]ferrichrome was added,and iron transport was determined as described previously (1).Phage T5 adsorption assays. Binding of the T5 lipoprotein
to the FhuA protein was examined by determining the rate ofT5 adsorption to whole cells, the total membrane fraction, andthe outer membrane fraction.
Cells (5 ml) were grown in TY medium to an optical densityat 578 nm of 1.0, collected by centrifugation, and suspended inadsorption buffer (M9 salts, 1 mM MgSO4, 0.1 mM CaCl2).For preparing cell envelopes, cells were grown to an opticaldensity at 578 nm of 1.0 and then lysed with EDTA-lysozyme.Media in which strains carrying plasmid pHE101 were culti-vated were supplemented with 1 mM isopropyl-p-D-thiogalac-topyranoside (IPTG). To isolate the outer membrane from thetotal membrane fraction, the cell envelope was treated with0.25% Triton X-100 and then subjected to differential centrif-ugation, as described previously (5). The isolated total mem-brane fraction (20 ,ul) and the outer membrane fraction (20 ,ul)were each mixed with 480 ,ul of adsorption buffer. To eachreaction mixture, 50 RI1 of phage T5 (5 x 105 phage) was addedand the mixture was incubated at 37°C for 15 min, after which300 ,u1 of the suspensions was mixed with 200 ,ul of ice-coldsaline. After centrifugation, 10-fold dilutions (3 ,ul) of thesupernatant were spotted onto a TY plate overlaid with TY topagar which contained 108 cells of the indicator strain AB2847.After incubation overnight at 37°C, single plaques werecounted.
For inactivation of FhuA by Llp synthesized in differentcells, 1 ml each of overnight cultures of E. coli K0483 fhuA(pHE100), K0483 llp(pHE101), and K0483(pHE100)(pHE101)was diluted in 40 ml of TY medium and grown for 1 h withshaking at 37°C. IPTG was added to cells carrying plasmidpHE101 to a final concentration of 1 mM, and growth wascontinued for 3 h. Cells were harvested and suspended at 0°Cin a solution consisting of 8 ml of 0.2 M Tris-hydrochloride (pH8)-i M sucrose-0.8 ml of 10 mM Na-EDTA (pH 8)-0.08 ml ofhen egg white lysozyme (4 mg/0.1 ml)-24 ml of distilled water,kept at 20°C. Cells were lysed after 10 min of incubation at20°C, after which 4 ml of a solution composed of 50 mMTris-hydrochloride (pH 8), 10 mM MgCl2, 2% Triton X-100,and 0.08 ml of DNase (2 mg/0.1 ml) was added. Cells wereharvested after 5 min, and the sediments were washed twicewith 5 ml of distilled H20. Finally, the sediments weresuspended in 1 ml of H20, which was separated into threeportions (each 0.33 ml) and centrifuged for 25 min at highspeed in a Biofuge. Two of the sediments were suspended in380 p.1 and one was suspended in 38 p.1 of M9 medium (M9 saltsolution [16] supplemented with 1 mM MgSO4 and 0.1 mMCaCl2) and kept overnight at 0°C. One sample remaineduntreated, one was treated with ultrasound twice for 30 s onice, and to the 38-,ul sample was added 342 p.l of 50 mM
Tris-hydrochloride-1 mM Na-EDTA-1.1% octylglucoside, pH8. After 30 min on ice, the last sample was centrifuged for 15min at 38,000 x g and the supernatant was used further (60 p.1added to 380 p.l of M9 medium). The outer membranesuspensions (60 p.l) of K0483(pHE100) and K0483(pHE101)were mixed and added to 380 p.l of M9 medium. Sampleswhich had been treated with ultrasound were mixed by treat-ment once more with ultrasound. As controls, unmixed sam-ples (60 p.l) were diluted in 440 p.1 of M9 medium. The outermembrane suspensions isolated from about 3 x 109 cells wereincubated for 30 min at 37°C, after which 50 p.l of phage T5(2.5 x 106 phage per test) was added. The suspensions wereslowly shaken for 15 min at 37°C, and then 0.3 ml was dilutedwith 0.2 ml of ice-cold 0.9% NaCl and centrifuged and 10-folddilutions (3 p.1) were dropped on the indicator strain AB2847on TY agar plates.
RESULTS
Construction of the test system. Llp of phage T5 encoded onplasmid pVK-L3 inactivated wild-type FhuA encoded on thechromosome (4) or on the low-copy-number plasmid pWSK29(Table 2), whereas too much FhuA was formed by high-copy-number plasmids to be neutralized by Llp (see Fig. 2A). Sinceour mutated fnuA genes described previously were on multi-copy plasmids (12-14, 21), we recloned them on the low-copy-number plasmid pWSK29 (Table 1) to reduce the amount ofFhuA in the outer membrane. High-level expression of Llp tobalance overexpressed FhuA was not possible in strain UL3fhuA (13), which lysed. Presumably, the recA mutation ren-dered UL3 unstable so that overexpression of Llp resulted inkilling. In contrast, strain K0483 recA+, which was derivedfrom a different parent than UL3, tolerated Llp. It was alsoimportant to transform K0483 first with plasmids carryingwild-type and mutated JhuA and then with the llp plasmid.Presumably, expression of llp rendered cells unstable so thatthey did not survive the transformation procedure. The llpgene was recloned on plasmid pSUKS1 under lacI control sothat cells could be transformed with the resulting pHE101plasmid and handled at a low level of llp expression before llptranscription was induced with 1 mM IPTG to determineinactivation of FhuA. The fluA mutation of strain K0483displayed no polar effect on the expression of the downstreamflhuCDB genes, so fhuA transformants could be examined forwhether they took up ferrichrome and albomycin. Expressionof Llp rendered E. coli KO483fhuA+(pHE100) resistant to thephages T5 and +80 and to colicin M and albomycin (Table 2,compare results for cells without Llp with those for cellsexpressing Llp). This shows that Llp inactivated FhuA not onlywith regard to infection by phage T5 but also with regard to allthe other FhuA ligands tested.
Inactivation of FhuA derivatives mutated at periplasmicsites. All tested insertion derivatives of FhuA except one,which according to the FhuA transmembrane topology model(Fig. 1) are located in the periplasm, showed no or only slightinactivation by Llp (Table 2). Cells expressing the FhuAderivatives FhuA369-16, FhuA223-16, and FhuA531-16, con-taining insertions of 16 amino acids at residues 369, 223, and531, respectively, were as sensitive or only slightly less sensitiveto the FhuA ligands in the presence of Llp. Plaque formationand growth inhibition were obtained with the same or 10-fold-higher ligand concentrations in the presence of Llp. FhuA369-4 was somewhat inactivated by Llp, in contrast toFhuA369-16, showing that the 16-residue insertion renderedFhuA more resistant to Llp than the 4-residue insertion.Sensitivity to colicin M of cells expressing FhuA369-4 was
J. BACTERIOL.
on January 31, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
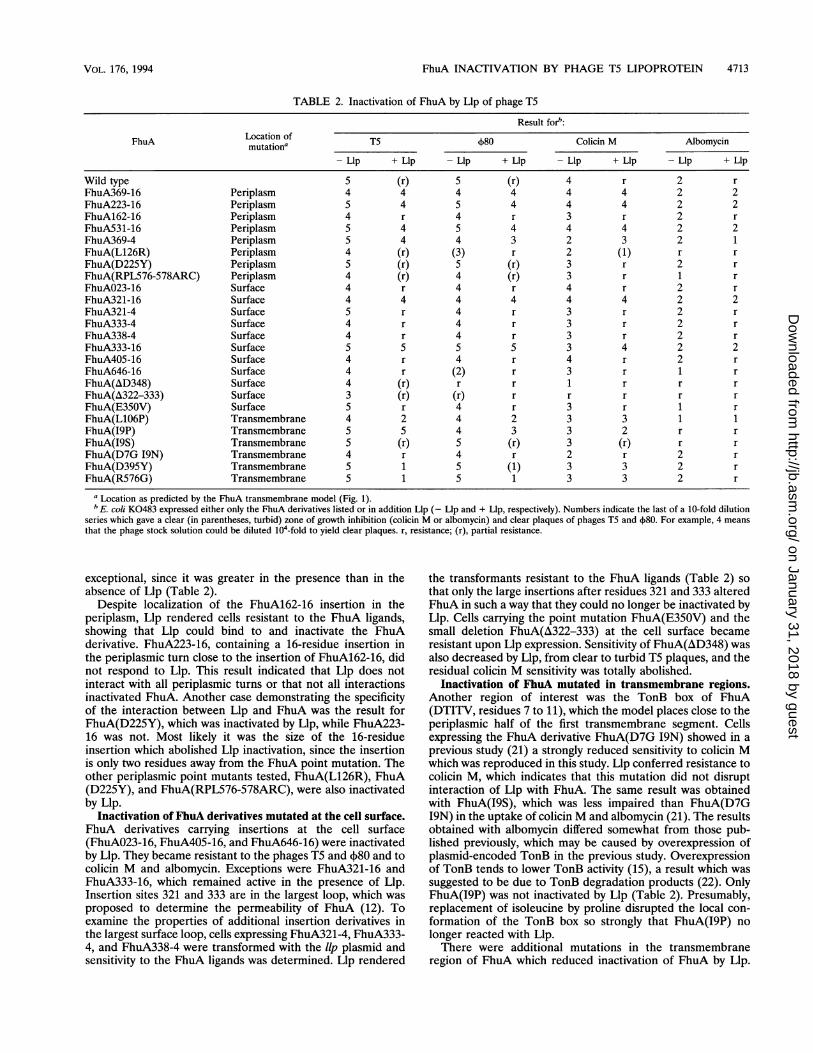
FhuA INACTIVATION BY PHAGE T5 LIPOPROTEIN 4713
TABLE 2. Inactivation of FhuA by Llp of phage T5
Result forb:
FhuA Location of T5 4)80 Colicin M Albomycinmutation
a
-Llp + Llp - Llp + Llp -Llp + Llp -Llp + Llp
Wild type 5 (r) 5 (r) 4 r 2 rFhuA369-16 Periplasm 4 4 4 4 4 4 2 2FhuA223-16 Periplasm 5 4 5 4 4 4 2 2FhuA162-16 Periplasm 4 r 4 r 3 r 2 rFhuA531-16 Periplasm 5 4 5 4 4 4 2 2FhuA369-4 Periplasm 5 4 4 3 2 3 2 1FhuA(L126R) Periplasm 4 (r) (3) r 2 (1) r rFhuA(D225Y) Periplasm 5 (r) 5 (r) 3 r 2 rFhuA(RPL576-578ARC) Periplasm 4 (r) 4 (r) 3 r 1 rFhuAO23-16 Surface 4 r 4 r 4 r 2 rFhuA321-16 Surface 4 4 4 4 4 4 2 2FhuA321-4 Surface 5 r 4 r 3 r 2 rFhuA333-4 Surface 4 r 4 r 3 r 2 rFhuA338-4 Surface 4 r 4 r 3 r 2 rFhuA333-16 Surface 5 5 5 5 3 4 2 2FhuA405-16 Surface 4 r 4 r 4 r 2 rFhuA646-16 Surface 4 r (2) r 3 r 1 rFhuA(AD348) Surface 4 (r) r r 1 r r rFhuA(A322-333) Surface 3 (r) (r) r r r r rFhuA(E350V) Surface 5 r 4 r 3 r 1 rFhuA(L1O6P) Transmembrane 4 2 4 2 3 3 1 1FhuA(I9P) Transmembrane 5 5 4 3 3 2 r rFhuA(I9S) Transmembrane 5 (r) 5 (r) 3 (r) r rFhuA(D7G 19N) Transmembrane 4 r 4 r 2 r 2 rFhuA(D395Y) Transmembrane 5 1 5 (1) 3 3 2 rFhuA(R576G) Transmembrane 5 1 5 1 3 3 2 r
aLocation as predicted by the FhuA transmembrane model (Fig. 1).bE. coli K0483 expressed either only the FhuA derivatives listed or in addition Llp (- Lip and + Llp, respectively). Numbers indicate the last of a 10-fold dilution
series which gave a clear (in parentheses, turbid) zone of growth inhibition (colicin M or albomycin) and clear plaques of phages T5 and 4)80. For example, 4 meansthat the phage stock solution could be diluted 104-fold to yield clear plaques. r, resistance; (r), partial resistance.
exceptional, since it was greater in the presence than in theabsence of Llp (Table 2).
Despite localization of the FhuA162-16 insertion in theperiplasm, Llp rendered cells resistant to the FhuA ligands,showing that Llp could bind to and inactivate the FhuAderivative. FhuA223-16, containing a 16-residue insertion inthe periplasmic turn close to the insertion of FhuA162-16, didnot respond to Llp. This result indicated that Llp does notinteract with all periplasmic turns or that not all interactionsinactivated FhuA. Another case demonstrating the specificityof the interaction between Llp and FhuA was the result forFhuA(D225Y), which was inactivated by Llp, while FhuA223-16 was not. Most likely it was the size of the 16-residueinsertion which abolished Llp inactivation, since the insertionis only two residues away from the FhuA point mutation. Theother periplasmic point mutants tested, FhuA(L126R), FhuA(D225Y), and FhuA(RPL576-578ARC), were also inactivatedby Llp.
Inactivation of FhuA derivatives mutated at the cell surface.FhuA derivatives carrying insertions at the cell surface(FhuAO23-16, FhuA405-16, and FhuA646-16) were inactivatedby Llp. They became resistant to the phages T5 and 480 and tocolicin M and albomycin. Exceptions were FhuA321-16 andFhuA333-16, which remained active in the presence of Llp.Insertion sites 321 and 333 are in the largest loop, which wasproposed to determine the permeability of FhuA (12). Toexamine the properties of additional insertion derivatives inthe largest surface loop, cells expressing FhuA321-4, FhuA333-4, and FhuA338-4 were transformed with the llp plasmid andsensitivity to the FhuA ligands was determined. Llp rendered
the transformants resistant to the FhuA ligands (Table 2) sothat only the large insertions after residues 321 and 333 alteredFhuA in such a way that they could no longer be inactivated byLlp. Cells carrying the point mutation FhuA(E350V) and thesmall deletion FhuA(A322-333) at the cell surface becameresistant upon Llp expression. Sensitivity of FhuA(AD348) wasalso decreased by Llp, from clear to turbid T5 plaques, and theresidual colicin M sensitivity was totally abolished.
Inactivation of FhuA mutated in transmembrane regions.Another region of interest was the TonB box of FhuA(DTITV, residues 7 to 11), which the model places close to theperiplasmic half of the first transmembrane segment. Cellsexpressing the FhuA derivative FhuA(D7G I9N) showed in aprevious study (21) a strongly reduced sensitivity to colicin Mwhich was reproduced in this study. Llp conferred resistance tocolicin M, which indicates that this mutation did not disruptinteraction of Llp with FhuA. The same result was obtainedwith FhuA(19S), which was less impaired than FhuA(D7G19N) in the uptake of colicin M and albomycin (21). The resultsobtained with albomycin differed somewhat from those pub-lished previously, which may be caused by overexpression ofplasmid-encoded TonB in the previous study. Overexpressionof TonB tends to lower TonB activity (15), a result which wassuggested to be due to TonB degradation products (22). OnlyFhuA(19P) was not inactivated by Llp (Table 2). Presumably,replacement of isoleucine by proline disrupted the local con-formation of the TonB box so strongly that FhuA(I9P) nolonger reacted with Llp.There were additional mutations in the transmembrane
region of FhuA which reduced inactivation of FhuA by Llp.
VOL. 176, 1994
on January 31, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

4714 BRAUN ET AL.
A R
A 0
T A
A T
A G
P T
2
W T
A D
S T
E P
Q I
P Q
R L
Y T
S G
Y L
V A
G R
D S
D A
D N
L A
AK
333H
L Y
AL
A A
C
K
SW L
G P
S a Y K A
D Y E N
S K G G A
F G T T P
D S E V 32itF E P E S
G E E P C
K S T N L D V
A S
G N
A N
R P
K D
Y K
V A
V N
F
D D
q50 F
K D
L T
a N
N V
F 40-)
S
Gp
y
R
I
N
K
a
K
a
T
A G
V T
R T
N D
L K
S R
E D
D
D K
A
W F
D G
K N
G I
V F
A
S P
S S
P K
E G
P S K
D F Y G
A F T N
M S T
L V D P
N E T A
N G T C
T G Y V
K E E F
T I A K
R D H
T A
P 6J)I N
A D S
Q G F
V Y K
P S V
K S G
H G S
T
N Y
F G
C C
S F
A U
V G
Y A
E E
R R
A K R T N G ,S Q R Q P N T G S R N G D T E G N A T M T Y D Q
P V T Y G F Y P F Y F N F F Y V F Y V Y W F' K Y R Y A Y T F V
A PSG D Y I G L A Y G E T G D DR YR R S V G TSR V LV
A K V N V GSAISNK V Y T V N N V G G E Y A V Y G V NA
T S S LK D SIL D A L R D R V G D L G G S E G D N T
LT LD IV Y VT I G
A T A V
TI
T E G I G V P N G SN N N P TN S E GL91)V P R N I G MN A S LA V A V Y F V V A V Y L V ILTVT G ~~~D R V K
T N D
G R S K T U SQL IL Y K I K N
g)TY F P 14MS F W T F V F
T ) V F P Y UVE A SA N Y I K R K N A K A D GD D K D T V R ®()S F G D N
K E L A N E R V 22-EXq25 E K E N T I Y NV P%31 (tJA D S S
P E A E D©(DA P E D G E G @369 G E S GIL AG
E M E G S E R T ®62 A N 395 P R A
V A K T K R 576-578 V M
A L V
H S
Q K
126 E P N S
106
G
N Y
T
p
FIG. 1. Tentative transmembrane organization of the FhuA protein (14). Circles indicate the mutation sites, which for the transmembraneregions (boxed regions) are numbered below the circles.
Sensitivity of cells expressing FhuA(L1O6P) to T5 and 4)80 wasreduced 100-fold by Llp, while sensitivity to colicin M andalbomycin remained unaltered. FhuA(D395Y) and FhuA(R576G) showed a 10-fold-reduced sensitivity to colicin Mwhich was not further reduced by Llp. The latter FhuAderivatives were also incompletely inactivated by Llp withregard to T5 and 4)80 infection. The FhuA model predicts allthree point mutations to be located at the inner half of theouter membrane, like the TonB box mutations.
Inhibition of phage T5 adsorption to cells, cell envelopes,and outer membranes by Lip. Phage T5 was incubated withcells, cell envelopes (total membranes), and outer membranesto see whether Llp inactivation of FhuA affected T5 adsorptionand to examine whether treatment with 0.25% Triton X-100used to isolate the outer membrane fraction abolished inacti-vation of FhuA by Llp. After 15 min of incubation, cells wereremoved by centrifugation and the unadsorbed phage wasdetermined by counting the plaques on nutrient agar platesseeded with the indicator strain AB2847. The PFU in thesupernatant of strain K0483 JuA were 20-fold higher than in
the supernatant of the JhuA' transformant (Table 3). Thenumber of phage particles in the supernatant of theflzuA + llp+transformant was as high as that in the supernatant of thefhuAcells, demonstrating inhibition of phage T5 binding to FhuA in
the presence of Llp. Similar results were obtained with cellenvelopes and outer membranes. The latter result shows that0.25% Triton X-100 did not abolish inactivation of FhuA byLlp. To examine the specificity of FhuA inactivation by Llp, T5adsorption to FhuA369-16, which in the infection assay hadretained activity in the presence of Llp, was determined.Adsorption to cells, envelopes, and outer membranes was notreduced by Llp.
Studies of adsorption of phage 480 to cells, cell envelopes,
TABLE 3. Inhibition of phage T5 adsorption to cellsby the T5 lipoprotein
103 phage/ml after incubation witha:Strain
Cells Cell envelopes Outer membranes
K0483 280 312 232K0483 FhuA 14 23 12K0483 FhuA Llp 285 302 335K0483 FhuA369-16 9 18 66K0483 FhuA369-16 Llp 6 10 58
a Values indicate the number of unadsorbed phages or PFU, determined intriplicate.
J. BACrERIOL.
on January 31, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

FhuA INACTIVATION BY PHAGE T5 LIPOPROTEIN 4715
TABLE 4. Inhibition of phage T5 adsorption to isolatedmembranes by T5 lipoprotein
104 phage/ml'Strain In the presence of Ultrasound Untreated
octylglucoside treated
K0483 FhuA 23 14 11K0483 Llp 220 160 180K0483 FhuA/Llpb 230 130 180K0483 FhuA + Llpc 150 13 22
a Values indicate the number of unadsorbed phages or PFU, determined intriplicate.bFhuA/Llp, FhuA and Llp coexpressed in the same cell.FhuA + Llp, FhuA and Lip expressed in different cells and mixed afterwards.
and outer membranes yielded results which were similar tothose obtained with phage T5 (data not shown).We examined whether Llp synthesized in one cell inacti-
vated FhuA synthesized in another cell. Phage T5 was incu-bated with outer membranes of cells producing FhuA, Llp, orFhuA and Llp and with mixtures of outer membranes contain-ing FhuA and outer membranes containing Llp. The superna-tant of outer membranes of cells coexpressing both FhuA andLlp contained 10 times more unadsorbed phages (PFU) thanthe supernatant of outer membranes expressing only FhuA(Table 4). Llp in one membrane inhibited T5 adsorption toFhuA in another membrane only in the presence of octylglu-coside (Table 4), which is known to solubilize FhuA (11, 12).This treatment also solubilized Llp, since the supernatant ofthe membrane after centrifugation was added to solubilizedFhuA. Treatment of the membrane mixture with ultrasoundleft FhuA active (Table 4), suggesting that Llp was not broughtinto proper contact with FhuA.
Inhibition of ferrchrome transport by Llp. Transport offerrichrome was determined in order to measure FhuA activityfor a substrate small in size and to obtain quantitative data onFhuA inactivation by Llp. Plasmid-encoded Llp completelyinhibited [55Fe3+]ferrichrome transport into wild-type E. coliAB2847 fhuA+ (Fig. 2A). Ferrichrome transport into strainAB2847 transformed with wild-typefhuA plasmid pHK763 wasnot inhibited by Llp, demonstrating that too much FhuA(chromosomal and plasmid encoded) was formed for inactiva-tion of all active FhuA molecules by Llp (Fig. 2A). Llp largelyinhibited (80%) transport into the fhuA+ llp+ transformant ofE. coli K0483 (Fig. 2B). In contrast, Llp reduced the rate oftransport into E. coli K0483 FhuA369-16 by only 20% (datanot shown) and the rate of transport into K0483 FhuA369-4by 35% (Fig. 2B). These values agree qualitatively with thephage infection and adsorption data. Transport of ferrichromeinto E. coli K0483 FhuA(R576G) was 75% inhibited by Llp(data not shown), which is comparable with the strong inhibi-tion of phage infection and sensitivity to albomycin (Table 2).
DISCUSSION
Llp inactivated FhuA with regard to all ligands tested. Llpdid not inactivate the FepA receptor, since cells remained fullysensitive to colicin B (data not shown). Llp had to reach acertain concentration relative to FhuA to inactivate FhuA,which was noticed previously (4). Inactivation of FhuA by Llpwas inhibited by mutations in FhuA which, according to theFhuA transmembrane topology model, were located inperiplasmic turns or in transmembrane regions close to theperiplasm. Since the ligands bind to FhuA at the cell surface,it is likely that Llp inactivates FhuA by inducing an inactive
25
§20
15
00
aO 10t
0 2 4 6 8 10 15 30Time in min
FIG. 2. (A) Transport of ["5Fe3+Iferrichrome into E. coli AB2847,AB2847 llp(pHE101), AB2847 phuA(pHK763) llp(pHE101), K0483JznuA(pHE100) expressing FhuA wild-type (WT), K0483 J7huA(pHE100) llp(pHE101), K0483 J7huA(pHE369-4), and K0483 JhuA(pHE369-4) llp(pHE101).
conformation. The conformational change induced by Llpproceeds from the periplasm through the outer membrane tothe surface-exposed regions of FhuA. The finding that Llp notonly inactivated but could also activate FhuA strongly suggestsinduction of a conformational change. Sensitivity of E. coliK0483 FhuA369-4 and E. coli FhuA333-16 to co'licin Mincreased 10-fold in the presence of Llp. It is unlikely that theincrease in colicin M sensitivity comes from an unspecificincrease of outer membrane permeability caused by FhuA369-4 and FhiuA33-16, since no other FhuA derivativesincreased colicin M sensitivity upon Llp expression. A confor-mational change in FhuA was also assumed for the opening ofthe FhuA channel (12) and for the conversion of the reversibleinto irreversible binding sites for the phages Tl and J)80 (8)through the action of TonB, ExbB, and ExbD. Also, the FhuAmutation I9P, impaired in TonB-related activities, did notrespond to Llp. This mutation may block binding of Llp andTonB to FhuA or may prevent a conformational change ofFhuA induced by Llp and TonB as well. However, Llp did notact as an antagonist of TonB, since it inhibited TS binding,which is unaffected by TonB. None of the FhuA mutantsstudied were differentially affected by Llp in their reactivity toTonB-dependent and TonB-independent FhuA activities.The only surface-exposed insertion derivatives, FhuA321-16
and FhuA333-16, which were inactivated by Llp were locatedin the largest loop, which was proposed to control the perme-ability of FhuA (12). Both FhuA derivatives were highly activetoward all ligands, showing a native conformation and proper
I-000-
(I)C
._C0
Time in min
VOL. 176, 1994
on January 31, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

4716 BRAUN ET AL.
insertion of the FhuA derivatives into the outer membrane.They were also not locked in a conformation which did notrespond to regulating proteins, since they reacted with TonB.For regulating the permeability of the FhuA channel, thelargest surface loop could be folded into the channel, similar tothe loop which determines the permeability of the porins (3,26). A portion of Llp may penetrate into the channel from theperiplasm and inactivate FhuA. The 16-residue insertionscould sterically prevent access of FhuA so that these deriva-tives are unaffected by Llp. The 4-residue insertions ofFhuA321-4, FhuA333-4, and FhuA338-4 would be too smallfor steric hindrance and are inactivated by Llp. However, thesesites, although present in the same loop, are not identical withregard to their access to proteases, since FhuA321-16 andFhuA321-4 were cleaved by subtilisin added to living cells,whereas FhuA333-16 was not cleaved (14).
Llp did not reduce the amount of FhuA in the outermembrane by binding to FhuA during FhuA synthesis, result-ing in interference with export of FhuA across the cytoplasmicmembrane or insertion into the outer membrane. SDS-poly-acrylamide gel electrophoresis revealed the same amounts ofFhuA in the outer membrane (data not shown).The outer membrane fraction prepared from cell envelopes
by treatment with Triton X-100 contained Llp, which supportsthe previous localization of Llp in the outer membrane (4). Llpwas not degraded by proteases added to cells, so it may not beexposed at the cell surface (4). Inactivation of FhuA by Llp wasnot abolished by Triton X-100. Apparently, the Llp-FhuAcomplex was not dissociated by the detergent, or binding of Llpto FhuA was restored upon removal of most of the TritonX-100 by washing the membranes. The latter possibility wasexamined by mixing separately synthesized FhuA (11, 12) andLlp, both proteins solubilized in octyglucoside. The mixturedisplayed 10-fold-reduced T5 binding, indicating that the de-tergent did not affect inactivation of FhuA by Llp.
In contrast to the proposed mode of action of the phage T5Llp, TraT, another lipoprotein, which prevents mating betweenF+ cells, is thought to block the OmpA receptor at the cellsurface. This proposal was based on the inhibition of phage K3binding to OmpA and on sequence homologies between TraTand the phage receptor-binding protein 38 (19). The homolo-gous sequence of TraT was localized at the cell surface, whereit inhibited mating-pair formation (10).
Inhibition of FhuA activity by phage T5 lipoprotein was notso specific as to inhibit only T5 adsorption. Llp inactivatedFhuA for all FhuA ligands tested. The inactive FhuA confor-mation induced by Llp must differ from the FhuA conforma-tions of unenergized cells and of TonB, ExbB, or ExbDmutants, which confer T5 infection, and from the energizedconformation induced by TonB, ExbB, and ExbD which allowsinfection by phages T5, Tl, and 480 and uptake of colicin M,ferrichrome, and albomycin. Since binding of ferrichrome alsoalters the FhuA conformation (11), FhuA apparently is able toassume quite a number of different conformations in the outermembrane.
ACKNOWLEDGMENTSWe thank K. Heller for providing plasmid pVK-L3 and results prior
to publication and U. W. Konninger for initial experiments. We areindebted to W. Koster for suggesting the use of his strain K0483.
This work was supported by the Deutsche Forschungsgemeinschaft(SFB 323).
REFERENCES1. Baumler, A. J., and K. Hantke. 1992. Ferrioxamine uptake in
Yersinia enterocolitica: characterization of the receptor FoxA.Mol. Microbiol. 6:1309-1321.
2. Bradbeer, C., and A. Gudmundsdottir. 1990. Interdependence ofcalcium and cobalamin binding by wild-type and mutant BtuBprotein in the outer membrane of Escherichia coli. J. Bacteriol.172:4919-4926.
3. Cowan, S. W., T. Schirmer, G. Rummel, M. Stert, R. Gosh, R. A.Pauptit, J. N. Jansonius, and J. P. Rosenbusch. 1992. Crystalstructures explain functional properties of two E. coli porins.Nature (London) 358:727-733.
4. Decker, K., V. Krauel, A. Meesmann, and K. Heller. 1994. Lyticconversion ofEscherichia coli by bacteriophage T5: blocking of theFhuA receptor protein by a lipoprotein expressed early duringinfection. Mol. Microbiol. 12:321-332.
5. Eick-Helmerich, K., and V. Braun. 1989. Import of biopolymersinto Escherichia coli: nucleotide sequences of the exbB and exbDgenes are homologous to those of the tolQ and tolR genes,respectively. J. Bacteriol. 171:5117-5126.
6. Fischer, E., K. Gunter, and V. Braun. 1989. Involvement of ExbBand TonB in transport across the outer membrane of Escherichiacoli: phenotypic complementation of exb mutants by overexpressedtonB and physical stabilization of TonB by ExbB. J. Bacteriol.171:5127-5134.
7. Gunter, K., and V. Braun. 1990. In vivo evidence for FhuA outermembrane interaction with the TonB inner membrane protein ofEscherichia coli. FEBS Lett. 274:85-88.
8. Hancock, R. E. W., and V. Braun. 1976. Nature of the energyrequirement for the irreversible adsorption of bacteriophages Tiand 4)80 to Escherichia coli. J. Bacteriol. 125:409-415.
9. Hantke, K. 1983. Identification of an iron uptake system specificfor coprogen and rhodotorulic acid in Escherichia coli. Mol. Gen.Genet. 191:301-306.
10. Harrison, J. L., I. M. Taylor, K. Platt, and D. C. O'Connor. 1992.Surface exclusion specificity of the TraT protein is determined bysingle alterations in a 5-amino acid region of the protein. Mol.Microbiol. 6:2825-2832.
11. Hoffmann, H., E. Fischer, H. Kraut, and V. Braun. 1986. Prepa-ration of the FhuA (TonA) receptor protein from cell envelopes ofan overproducing strain of Escherichia coli K-12. J. Bacteriol.166:404-411.
12. Killmann, H., R. Benz, and V. Braun. 1993. Conversion of theFhuA transport protein into a diffusion channel through the outermembrane of Escherichia coli. EMBO J. 12:3007-3016.
13. Killmann, H., and V. Braun. 1992. An aspartate deletion mutationdefines a binding site of the multifunctional FhuA outer mem-brane receptor of Eschenchia coli K-12. J. Bacteriol. 174:3479-3486.
14. Koebnik, R., and V. Braun. 1993. Insertion derivatives containingsegments of up to 16 amino acids identify surface- and periplasm-exposed regions of the FhuA outer membrane receptor of Esch-erichia coli K-12. J. Bacteriol. 175:826-839.
15. Mann, B. J., C. D. Holroyd, C. Bradbeer, and R. J. Kadner. 1986.Reduced activity of TonB-dependent functions in strains of Esch-enchia coli. FEMS Microbiol. Lett. 33:255-260.
16. Miller, J. H. 1972. Experiments in molecular genetics. Cold SpringHarbor Laboratory, Cold Spring Harbor, N.Y.
17. Murphy, C., V. I. Kalve, and P. E. Klebba. 1990. Surface topologyof the Escherichia coli K-12 ferric enterobactin receptor. J. Bac-teriol. 172:2736-2747.
18. Ondraczek, R., S. Hobbie, and V. Braun. 1992. In vitro activationof the Serratia marcescens hemolysin through modification andcomplementation. J. Bacteriol. 174:5086-5094.
19. Riede, I., and M.-L. Eschbach. 1986. Evidence that TraT interactswith OmpA of Escherichia coli. FEBS Lett. 205:241-245.
20. Rutz, J. M., J. Liu, J. A. Lyons, J. Goranson, S. K. Armstrong,M. A. MacIntosh, J. B. Feix, and P. E. Klebba. 1992. Formation ofa gated channel by a ligand-specific transport protein in thebacterial outer membrane. Science 258:471-475.
21. Schodfler, H., and V. Braun. 1989. Transport across the outermembrane of Escherichia coli K12 via the FhuA receptor isregulated by the TonB protein of the cytoplasmic membrane. Mol.Gen. Genet. 217:378-383.
22. Skare, J. T., and K. Postle. 1991. Evidence for a TonB-dependentenergy transduction complex in Escherichia coli. Mol. Microbiol.5:2883-2890.
J. BACTERIOL.
on January 31, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

FhuA INACTIVATION BY PHAGE T5 LIPOPROTEIN 4717
23. Stanssens, P., C. Opsomer, Y. M. McKeown, W. Kramer, M.Zabeau, and H.-J. Fritz. 1989. Efficient oligonucleotide-directedconstruction of mutations in expression vectors by the gappedduplex DNA method using alternating markers. Nucleic AcidsRes. 17:4441-4454.
24. Wang, R., and S. Kushner. 1991. Construction of versatile low-copy-number vectors for cloning, sequencing and gene expressionin Escherichia coli. Gene 100:195-199.
25. Ward, R. J., S. E. Hufton, N. A. C. Bunce, A. J. P. Fletcher, and
R. E. Glass. 1992. A structure-function analysis of BtuB, the E. colivitamin B12 outer membrane transport protein, p. 271-296. In R.James, C. Lazdunski, and F. Pattus (ed.), Bacteriocins, microcins andlantibiotics. Series H: cell biology, vol. 65. Springer-Verlag, Berlin.
26. Weiss, M. S., and G. E. Schulz. 1992. Structure of porin at 1.8 Aresolution. J. Mol. Biol. 227:493-509.
27. Zhou, Y. C., Y. Yang, and A. Y. Jong. 1990. Miniprep in tenminutes. BioTechniques 8:172-173.
VOL. 176, 1994
on January 31, 2018 by guesthttp://jb.asm
.org/D
ownloaded from