in vitro study of the long-term cortisol treatment effects on the growth rate and...
TRANSCRIPT

In vitro study of the long-term cortisoltreatment effects on the growth rate andproliferation of the neural stem/precursorcells
Alireza Abdanipour1, Mohsen Sagha2, Ali Noori-Zadeh1, Iraj Pakzad3,Taki Tiraihi1
1Shefa Neuroscience Research Center, Khatam Al-Anbia Hospital, Tehran, Iran, 2Department of AnatomicalSciences and Pathology, Research Laboratory for Embryology and Stem Cells, School of Medicine, ArdabilUniversity of Medical Sciences, Ardabil, Iran, 3Department of Microbiology, Faculty of Medicine, Ilam Universityof Medical Sciences, Ilam, Iran
Adult neural stem/precursor cells (NSPCs) residing in the subventricular zone of the lateral ventricles andthe subgranular zone of the dentate gyrus of the hippocampus are involved in the memory formations andpsychological problems. It is believed that basal levels of glucocorticoids are essential for neuronaldevelopment, plasticity, and survival, while stress-mediated levels of glucocorticoids produce neuronalloss. Degeneration of NSPCs by the apoptotic and necrotic stimuli have great devastating outcomes on thebrain and contributes to the pathophysiology of neurological as well as psychological disorders. Using MTTassay, acridine orange, and TUNEL assay, we have demonstrated that cortisol at high and excessive (morethan 5 mM) levels had anti-proliferative effects on the NSPCs derived from subventricular and subgranularzones in a dose- and time-dependent manner through apoptosis as well as necrosis. These outcomes canhighlight the role of stress-mediated decline of adult neurogenesis in the aging brain and interconnectstress-mediated cortisol secretion with brain aging diseases.
Keywords: Cortisol, NSPCs, Adult neurogenesis, Subventricular zone, Dentate gyrus, Neurosphere
IntroductionAdult neurogenesis or the production of new neural
stem/precursor cells (NSPCs) has been shown to
occur in two major regions of the brain including
the subventricular zone of the lateral ventricles and
the subgranular zone of the dentate gyrus of the
hippocampus. In the dentate gyrus, thousands of
granule cells were born daily; however, only a small
portion of the NSPCs survives and develops into
mature granule cells.1 It has been shown that adult
neurogenesis is the key component of the pattern
separation function of the dentate gyrus, and recent
studies also shed light on the significant relationships
between adult neurogenesis and cognition.2–4 The
functional theories of new NSPCs production have
consolidated several aspects of adult neurogenesis
and maturation.5 The immature granule cells demon-
strate an increased intrinsic excitability and plasti-
city,6,7 and interestingly, it is believed that this
immature state of granule cells represents a critical
developmental period in which they encode signifi-
cant features of their environments.8,9 Dentate gyrus
and cornu ammonis 3 interact via the mossy fibers
to process high-resolution spatial information.10–12
Further, the dentate gyrus has been proposed to
mediate spatial pattern separation of similar spaces
for the detection of small changes within an environ-
ment. It is presumed that cornu ammonis 3 mediates
a similar pattern separation process on much larger
scale spaces,11 and they may contribute to neurolo-
gical problems such as Alzheimer’s disease (AD).
Moreover, NSPCs located within the subventricular
zone exert an innate homeostatic regulatory function
protecting striatal neurons from glutamate-mediated
excitotoxicity13, and thus adult neurogenesis may
potentially play a crucial role in Huntington’s disease
(HD) as well, in which loss of the striatal projection
neurons occurs.14 Moreover, striatum is the main
input nucleus of the basal ganglia and a key neural
substrate for procedural learning and memory.15 The
vast majority of the striatal neurons are GABAergic
Correspondence to: Alireza Abdanipour, Shefa Neuroscience ResearchCenter, Khatam Al-Anbia Hospital, Tehran, Iran. Email: [email protected]
� W. S. Maney & Son Ltd 2014DOI 10.1179/1743132814Y.0000000431 Neurological Research 2014 VOL. 000 NO. 000 1

projection neurons, which exhibit slow but, tempo-
rally precise spiking. In Parkinson’s disease (PD), a
broad spectrum of non-motor symptoms and deficits
exist and they may partly rely on hippocampal
function and indeed it is assumed that some of them
are related to defective adult neurogenesis.16 In a
general sense, according to the roles of NSPCs in the
adult brain functions, any disturbances in the adult
brain ability to regenerate NSPCs may contribute to
the pathophysiological conditions, and it is believed,
however, that adult neurogenesis steadily declines
during brain aging causing diseases such as AD, PD,
HD, innate anxiety, major depressive disorder (MDD),
etc. The progressive decline of NSPCs proliferation
during aging raises the questions of whether the
precursor cells become unresponsive to cellular niche
cues or whether the cellular niche does not provide the
stimuli for evoking proliferation or a niche factor itself
is already dampening this process. However, adult
neurogenesis is regulated by a variety of factors such as
learning,17 exercise,18 seizures,19 age,20,21 and stress.22
It has been shown that in response to stress, cortisol
secretion from the zona faciculata of the adrenal gland
at high concentrations suppresses the proliferation of
the neurons.23,24 We hypothesized that cortisol secre-
tion at high concentrations may suppress the prolifera-
tion of NSPCs, and this phenomenon can contribute to
the brain aging during adulthood and senescence. In
this regard, we have evaluated the chronological effects
of cortisol application in the cell culture medium at
different concentrations on the apoptosis and necrosis
rate of NSPCs using MTT and TUNEL assays as well
as acridine orange staining. We found that long-term
treatments of NSPCs by high and excessive levels of
cortisol decreased the viability and proliferation rate of
NSPCs. Also, the number of apoptotic and necrotic
cells increased in a dose- and time-dependent manner.
These outcomes can highlight the role of stress-
mediated decline of adult neurogenesis in the aging
brain and interconnect stress-mediated cortisol secre-
tion with the brain pathophysiology.
MethodsIsolation of NSPCs and neurosphere formationNeural stem/precursor cells were isolated from sub-
granular and subventricular zones of the rats (8–
12 weeks old) under deep ketamine/xylazine anesthesia.
The brains were removed quickly and washed in the ice-
cold phosphate buffered saline (PBS) supplemented
with 4.5 g/L glucose. The dissected hippocampi and
subventricular zones were washed with a 10 mL PBS/
glucose solution and then centrifuged down for
5 minutes at 1600 g and 4uC. After removal of the
supernatant, the tissues were homogenized with scissors
and scalpels. The collected tissues were dissociated by
digestion with a mixture of papain (Sigma), dispase II
(Sigma) and accutase (Invitrogen). The tissues were
incubated for 30 minutes at room temperature (RT)
and the solution was mixed gently every 10 minutes.
The cell mixture was passed through a 70 mm cell
strainer (Falcon) and then, centrifuged for 5 minutes, at
800 g and 4uC. Following the centrifugation, the pellets
were resuspended in a 1 mL Dulbecco modified eagle
medium/F12 medium (DMEM/F12; Invitrogen) sup-
plemented with B27 (Gibco), 100 units/mL penicillin,
and 100 units/mL streptomycin; 20 ng/mL epidermal
growth factor (EGF, Sigma); and 20 ng/mL fibroblast
growth Factor-2 (bFGF, Millipore).25 The isolated cells
were seeded in a six-well non-adherent plate and then
incubated at 37uC in 5% CO2 to form neurospheres for
1–2 weeks. For the preparation of a single-cell layer
from the neurospheres, they were centrifuged for
5 minutes, at 300 g and 4uC. Then, the cells were
washed in the basic media and dissociated with 0.02%
PBS-EDTA and then expanded on the poly-L-lysine
(Sigma) coated six-well chambers and passaged up to
three times.
Immunofluorescence stainingThe NSPCs were seeded on the poly-L-lysine (Sigma)-
coated cover slips and cultured for 4 hours at 37uC.
The cells were fixed (4% paraformaldehyde, 20 min-
utes, RT), washed with PBS, and then permeabilized
(0.2% Triton X-100, 10 minutes, RT). Subsequently,
they were treated with the blocking buffer (PBS with
0.1% goat serum; 30 minutes, RT) before the incuba-
tion with the primary antibodies. Then, the cells were
incubated overnight at 4uC with the primary polyclonal
antibodies against rat nestin (1 : 100; Santa Cruz) and
Sox2 (1 : 200; Santa Cruz) proteins, separately. After
washing with PBS, 60 minutes at RT, incubations with
the species-relevant fluorescent-labeled secondary anti-
bodies were followed. The nuclei were counterstained
with ethidium bromide, and the images were obtained
using a fluorescence microscope. For negative controls,
the immunocytochemistry was performed as for the
samples with the except that the primary antibody was
omitted. For positive controls, neonatal rat dentate
gyrus was used for the Sox2 and nestin expression.
MTT assay and cell viability evaluationSubconfluent cultures of the NSPCs at the third
passage were seeded on the 96-well microtiter plates
and maintained in the DMEM/F12 supplemented with
B27, 20 ng/mL EGF, 20 ng/mL bFGF, and 3% FBS
for 24 hours. To determine the cell viability at various
concentrations of the cortisol, the NSPCs were treated
with cortisol prepared in water (hydrocortisone-water
soluble, synonym: cyclodextrin-encapsulated hydro-
cortisone, H0396, Sigma) at 0, 0.25, 0.5, 1, 2.5, 5, 10,
15, and 20 mM concentrations at different exposure
time courses (24, 48, 72, 96, and 120 hours) and
then, cortisol toxicity were evaluated using MTT
Abdanipour et al. Long-term cortisol treatment effects
2 Neurological Research 2014 VOL. 000 NO. 000

(3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium
bromide) assay.26 Briefly after treatment, the medium
was removed and replaced with 200 mL of 0.5 mg/mL
MTT in 10% FBS-containing medium, and the cells
were incubated in the CO2 incubator at 37uC for
4 hours. The supernatants were removed from the
wells, and the reduced MTT dye was solubilized in the
200 mL/well of the DMSO. Absorbance at 570 nm was
determined by a plate reader, and the percentage of the
relative cell viability was calculated as follows: A570 of
the treated samples/A570 of the untreated samples 6100.
Acridine orange/ethidium bromide stainingCell morphological changes in the necrosis were
assessed by acridine orange/ethidium bromide staining.
The NSPCs were treated with cortisol at 5, 10, 15, and
20 mM concentrations at each aforementioned exposure
time courses. The cells were washed with the PBS buffer,
and fixed with 4% paraformaldehyde for 15 minutes
and then, stained with acridine orange/ethidium bro-
mide (Sigma), 100 mg/mL of each, for 5 minutes at
RT.27 Two hundred of the cells were counted and the
number of the necrotic cells was assessed as their
cytoplasm had been stained orange/yellow, and also
had a nuclear morphology resembling that of the
viable cells without any condensed chromatin and
fragmented nuclei. The cells were observed using a
fluorescence microscope.
TUNEL assayPotential DNA fragmentation and thus apoptosis was
visualized with a fluorescent staining method using In
Situ Cell Death Detection Kit (Roche, Germany)
according to the manufacturer’s instructions. After
each exposure time course treatment at 5, 10, 15, and
20 mM concentrations of cortisol, the cells were fixed
with a 4% paraformaldehyde solution prepared in the
PBS buffer for 15 minutes at RT and then permeabi-
lized with a 0.4% Triton X-100 solution for 10 minutes
at RT. Then, the cells were subjected to a terminal
deoxynucleotidyl transferase-mediated deoxyuridine
triphosphate nick end labeling assay (TUNEL assay).
In order to stain the nuclei, DAPI (4’, 6-diamidino-2-
phenylindole) was subsequently added. Hematoxylin
was also used for counterstaining. The percentage of
TUNEL-positive cells was assessed in the five ran-
domly selected fields for each well.28
Statistical analysisThe quantification was performed based on three
independent experiments for the MTT and TUNEL
and acridine orange assays, separately. The results
were normalized with the control groups, which were
presented as a mean percentage ¡ SEM and analyzed
by one-way ANOVA followed by Tukey’s post hoc
multiple group comparison test. A difference between
groups was considered statistically to be significant if
P , 0.05.
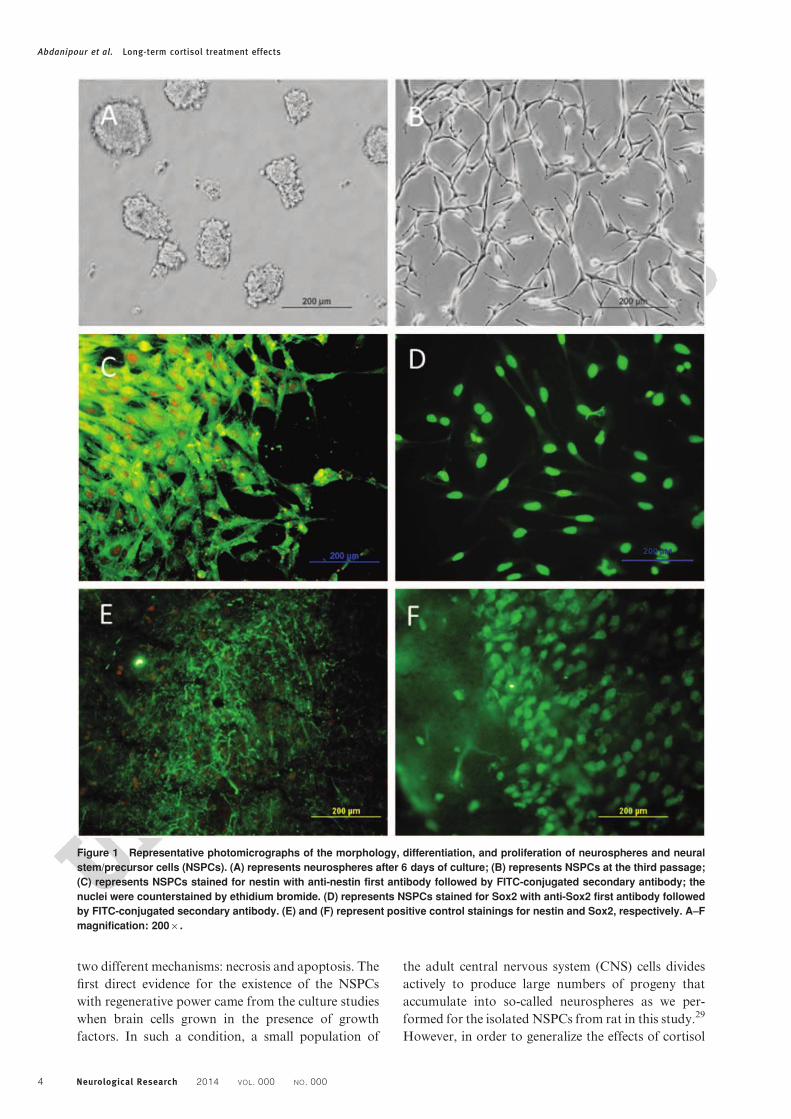
ResultsNeurosphere formationThe isolated NSPCs were grown in the appropriate
medium and clusters of the dividing cells formed into
the neurospheres. After 5–6 days of the incubation,
average diameter of the floating neurospheres grew up
to around 120 mm (Fig. 1A). Then, the individual
neurospheres were transferred into a fresh growth
medium containing bFGF and EGF. They formed
new neurospheres within the next 24 hours, eventually
reaching sizes close to that of the primary neu-
rospheres. After 6–10 days in the culture medium,
primary neurospheres were used for further experi-
ments (Fig. 1B). The results of the immunocytochem-
ical staining of the neurospheres showed that the
NSPCs were positive for nestin (98.34 ¡ 2.45) and
Sox2 (90.66 ¡ 3.12) markers (Fig. 1C and D) as we
performed control stainings confirming the specificity
of the labeling (Fig. 1E and F).
MTT assayMTT assay was used to show the toxicity effects of
cortisol on the NSPCs. As shown in Fig. 2, cortisol
caused a significant decline in the cell viability rate of
the NSPCs and the highest inhibition of cell prolifera-
tion was observed at the highest concentration and
longer duration time course of cortisol, i.e. 20 mM for
120 hours (30.16 ¡ 3.42). However, we did not detect
any decline in the cell viability between 0 and 5 mM
concentrations of cortisol at any aforementioned time
courses.
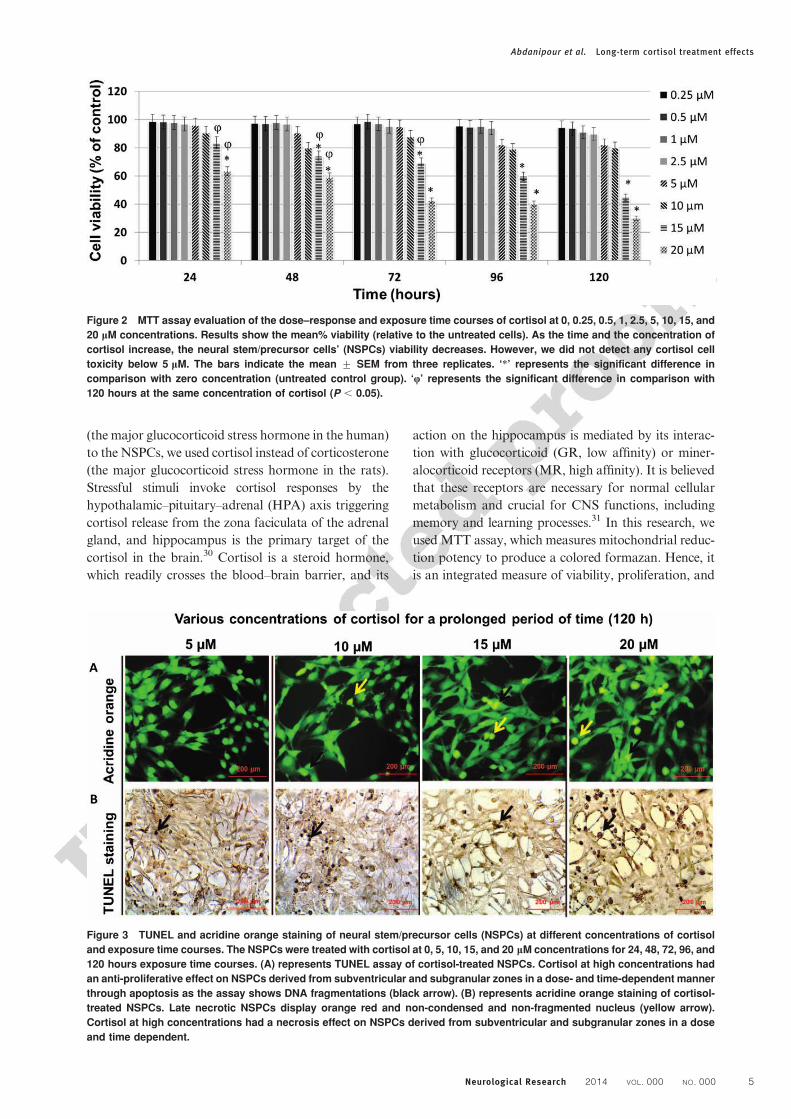
Assessment of necrosisThe results of the necrosis evaluation of the NSPCs in a
dose–response and exposure time courses of corti-
sol were shown in Figs. 3A and 4A. As it has been
demonstrated in these two figures, higher concentration
of cortisol and longer exposure time courses resulted in
a significant increase in the acridine orange/ethidium
bromide-positive cells compared to the control (0 mM).
Assessment of apoptosisAs cortisol concentrations and time exposures were
increased, TUNEL staining results showed that there
were significant increases in the percentage of the
apoptotic cells (shown in Figs. 3B and 4B). TUNEL
staining results showed that there were significant
increases in the percentage of the apoptotic cells as
the cortisol concentration were increased (shown in
Table 1).
DiscussionIn the present study, the stress hormone, cortisol,
significantly decreased the in vitro cell viability and
proliferation of the NSPCs expressing nestin and
Sox2 in a dose- and time-dependent manner through
Abdanipour et al. Long-term cortisol treatment effects
Neurological Research 2014 VOL. 000 NO. 000 3
two different mechanisms: necrosis and apoptosis. The
first direct evidence for the existence of the NSPCs
with regenerative power came from the culture studies
when brain cells grown in the presence of growth
factors. In such a condition, a small population of
the adult central nervous system (CNS) cells divides
actively to produce large numbers of progeny that
accumulate into so-called neurospheres as we per-
formed for the isolated NSPCs from rat in this study.29
However, in order to generalize the effects of cortisol
Figure 1 Representative photomicrographs of the morphology, differentiation, and proliferation of neurospheres and neural
stem/precursor cells (NSPCs). (A) represents neurospheres after 6 days of culture; (B) represents NSPCs at the third passage;
(C) represents NSPCs stained for nestin with anti-nestin first antibody followed by FITC-conjugated secondary antibody; the
nuclei were counterstained by ethidium bromide. (D) represents NSPCs stained for Sox2 with anti-Sox2 first antibody followed
by FITC-conjugated secondary antibody. (E) and (F) represent positive control stainings for nestin and Sox2, respectively. A–F
magnification: 2006.
Abdanipour et al. Long-term cortisol treatment effects
4 Neurological Research 2014 VOL. 000 NO. 000
(the major glucocorticoid stress hormone in the human)
to the NSPCs, we used cortisol instead of corticosterone
(the major glucocorticoid stress hormone in the rats).
Stressful stimuli invoke cortisol responses by the
hypothalamic–pituitary–adrenal (HPA) axis triggering
cortisol release from the zona faciculata of the adrenal
gland, and hippocampus is the primary target of the
cortisol in the brain.30 Cortisol is a steroid hormone,
which readily crosses the blood–brain barrier, and its
action on the hippocampus is mediated by its interac-
tion with glucocorticoid (GR, low affinity) or miner-
alocorticoid receptors (MR, high affinity). It is believed
that these receptors are necessary for normal cellular
metabolism and crucial for CNS functions, including
memory and learning processes.31 In this research, we
used MTT assay, which measures mitochondrial reduc-
tion potency to produce a colored formazan. Hence, it
is an integrated measure of viability, proliferation, and
Figure 2 MTT assay evaluation of the dose–response and exposure time courses of cortisol at 0, 0.25, 0.5, 1, 2.5, 5, 10, 15, and
20 mM concentrations. Results show the mean% viability (relative to the untreated cells). As the time and the concentration of
cortisol increase, the neural stem/precursor cells’ (NSPCs) viability decreases. However, we did not detect any cortisol cell
toxicity below 5 mM. The bars indicate the mean ¡ SEM from three replicates. ‘*’ represents the significant difference in
comparison with zero concentration (untreated control group). ‘Q’ represents the significant difference in comparison with
120 hours at the same concentration of cortisol (P , 0.05).
Figure 3 TUNEL and acridine orange staining of neural stem/precursor cells (NSPCs) at different concentrations of cortisol
and exposure time courses. The NSPCs were treated with cortisol at 0, 5, 10, 15, and 20 mM concentrations for 24, 48, 72, 96, and
120 hours exposure time courses. (A) represents TUNEL assay of cortisol-treated NSPCs. Cortisol at high concentrations had
an anti-proliferative effect on NSPCs derived from subventricular and subgranular zones in a dose- and time-dependent manner
through apoptosis as the assay shows DNA fragmentations (black arrow). (B) represents acridine orange staining of cortisol-
treated NSPCs. Late necrotic NSPCs display orange red and non-condensed and non-fragmented nucleus (yellow arrow).
Cortisol at high concentrations had a necrosis effect on NSPCs derived from subventricular and subgranular zones in a dose
and time dependent.
Abdanipour et al. Long-term cortisol treatment effects
Neurological Research 2014 VOL. 000 NO. 000 5

Figure 4 Apoptosis and necrosis quantification of different concentrations of cortisol and exposure time courses. (A) Higher
concentrations of cortisol and longer exposure time courses resulted in a significant increase in the necrosis process. (B) As
cortisol concentrations and time exposures were increased, TUNEL staining results also showed that there were significant
increases in the percentage of the apoptotic cells as well. The results represent the percentage of the necrosis ¡ SEM and also
the apoptosis ¡ SEM. ‘*’ represents the significant difference in comparison with 24 hours. ‘a,’ ‘b,’ and ‘c’ represent the
significant difference compared to 0, 5, and 10 mM at the same time, respectively (P , 0.05).
Table 1 The mean percentages of the apoptotic neural stem/precursor cells (NSPCs) (stained with TUNEL assay)
Cortisol concentrations
Exposure time courses
24 hours 48 hours 72 hours 96 hours 120 hours
0 mM 1.14 ¡ 0.44 1.62 ¡ 0.48 2.45 ¡ 0.29 3.14 ¡ 0.3 2.85 ¡ 0.355 mM 2.57 ¡ 1 4.51 ¡ 0.64 4.88 ¡ 0.62 8.82 ¡ 0.77 11.44 ¡ 1.2410 mM 6.81 ¡ 1.41 13.27 ¡ 1.15 17.02 ¡ 1.53 22.32 ¡ 2.78 29.77 ¡ 0.8415 mM 16.11 ¡ 1.58 20.66 ¡ 1.34 18.94 ¡ 1.76 25.89 ¡ 1.92 35.33 ¡ 2.3220 mM 18.26 ¡ 0.94 27.48 ¡ 1.55 40.96 ¡ 1.32 48.2 ¡ 0.92 49.48 ¡ 1.97
The NSPCs were treated with cortisol at 0, 5, 10, 15, and 20 mM concentrations for 24, 48, 72, 96, and 120 hours time courses.
Abdanipour et al. Long-term cortisol treatment effects
6 Neurological Research 2014 VOL. 000 NO. 000

metabolic activity, and thus, it is a reasonable assay for
cell health assessment. However, cortisol concentra-
tions lower than 5 mM did not show any cell toxicity
effects on the NSPCs in comparison with untreated
group (zero concentration) and thus, for cortisol
toxicity evaluation, concentrations higher than 5 mM
were further considered. Although basal levels of
glucocorticoids are essential for neuronal development,
plasticity, and survival, stress-mediated and excessive
levels of glucocorticoids produce NSPCs loss (in the
case of cortisol, higher than 5 mM). We showed that
high and excessive doses of cortisol and longer periods
of treatment had profound inhibitory effects on the
NSPCs viability and proliferation through both of the
necrosis and apoptosis processes. Glucocorticoids are
known to be able to inhibit cell differentiation and
synaptic development in the various brain regions at
high concentrations.32–35 The outstanding result of
cortisol neurotoxicity is brain functionality decline.
Because any decline in the adult neurogenesis may
underlie cognitive impairments associated with the
brain aging and the neurodegenerative disorders. If
stress alters the set point for growth of NSPCs in the
adult brain, it could give credence to many studies that
learning disabilities and even mental illnesses might
have association with cortisol effects on neurogenesis in
the human. However, of the hippocampal structures,
dentate gyrus sensitivity to glucocorticoids is of par-
ticular concern that is associated with learning and
memory formation.36 Indeed, in support of this hypo-
thesis, studies showed that the plasma cortisol levels are
increased in AD.37–40 Corticotrophin-releasing hor-
mone (CRH) hyperdrive occurs when the mRNA levels
of CRH increase in the hypothalamic paraventricular
nucleus (PVN), and this phenomenon causes the
hyperactivity of the HPA axis in AD.41 Also, the
evidence for the primary hyperdrive of CRH involv-
ing MDD comes from multiple sources: biochemical
studies, functional HPA-axis tests, neuroimaging, and
postmortem studies, as well as clinical trials with HPA-
axis-related compounds.42 In this research, glucocorti-
coid toxicity was investigated in the neural stem cells;
however, data from other studies showed that gluco-
corticoids have toxic effects on the adult neurons as
well.23,24 As the hippocampus has a pivotal role in the
negative feedback inhibition of cortisol secretion,43 its
neuronal damage may lead to the inefficient inhibitory
control of the CRH producing cells resulting in the
increased circulating cortisol and further hippocampus
damages44 and NSPCs toxicity as well. This feed-
forward cascade are known as the ‘glucocorticoid
cascade hypothesis’ and was proposed to be the major
pathogenetic mechanism in the human neurodegenera-
tive diseases associated with HPA-axis alterations.45
The subject is more attractive as it has been shown that
the NSPCs of the subventricular zone exert an ‘innate’
homeostatic regulatory role by protecting striatal neu-
rons from glutamate-mediated excitotoxicity.13 More-
over, memory and the HPA-axis dysfunction are
synchronously present in HD, AD, and MDD46 and
also adult neurogenesis in the animal models and
postmortem studies is severely affected.16 We found
that the majority of the NSPCs promoted to necrosis as
well as apoptosis after exposure to high concentrations
of cortisol. Thus, cortisol toxicity may underlie such
brain diseases. Necrosis and apoptosis concomitantly
occurs as these phenomena were detected by TUNEL
and acridine orange assays. Acridine orange can detect
necrotic cells and TUNEL assay also is a sensitive
method for in situ detection of DNA fragmentation in
the apoptotic cells based on labeling of the blunt ends of
DNA breaks. So, cortisol may have the lion’s share in
the degeneration of the NSPCs residing in the brain
albeit at higher than 5 mM concentration. At molecular
level, it has been shown that dexamethasone inhibited
the cell cycle through ubiquitin-mediated degradation
of Cyclin D147 and also it has been proposed that
glucocorticoids endanger hippocampal neurons through
exacerbating the excitotoxic glutamate-calcium-reac-
tive oxygen species cascade.48 Further investigations
are needed to determine the exact mechanisms of
cortisol-mediated NSPCs apoptosis and also necrosis
at molecular levels. Understanding the underlined
mechanisms can pave the way to design new ther-
apeutic drugs targeting brain aging processes.
Disclaimer StatementsContributors Alireza Abdanipour: designer, monitoring
projects, projects, corresponding author; Mohsen
Sagha: cooperative research project; Ali Noori-Zadeh:
cooperative research project, molecular biology advi-
sor; Iraj Pakzad: main advisor; Taki Tiraihi: supervisor.
Funding Shefa Neuroscience Research Center,
Khatam Al-Anbia Hospital, Tehran, Iran.
Conflicts of interest The authors have no conflict of
interest to declare with respect to this manuscript.
Ethics approval
AcknowledgementsWe are grateful to all experts and students of the
Khatam Al-Anbia Hospital for their help to carry out
this project.
References1 Kempermann G, Wiskott L, Gage FH. Functional significance
of adult neurogenesis. Curr Opin Neurobiol. 2004;14(2):186–91.2 Aimone JB, Wiles J, Gage FH. Potential role for adult
neurogenesis in the encoding of time in new memories. NatNeurosci. 2006;9(6):723–7.
3 Clelland C, Choi M, Romberg C, Clemenson G, Fragniere A,Tyers P, et al. A functional role for adult hippocampalneurogenesis in spatial pattern separation. Science. 2009;325(5937):210–3.
Abdanipour et al. Long-term cortisol treatment effects
Neurological Research 2014 VOL. 000 NO. 000 7

4 Sahay A, Wilson DA, Hen R. Pattern separation: a commonfunction for new neurons in hippocampus and olfactory bulb.Neuron. 2011;70(4):582–8.
5 Aimone JB, Deng W, Gage FH. Adult neurogenesis: integratingtheories and separating functions. Trends Cogn Sci (Regul Ed).2010;14(7):325–37.
6 Esposito MS, Piatti VC, Laplagne DA, Morgenstern NA,Ferrari CC, Pitossi FJ, et al. Neuronal differentiation in theadult hippocampus recapitulates embryonic development. JNeurosci. 2005;25(44):10074–86.
7 Ge S, Yang CH, Hsu KS, Ming GL, Song H. A critical periodfor enhanced synaptic plasticity in newly generated neurons ofthe adult brain. Neuron. 2007;54(4):559–66.
8 Kee N, Teixeira CM, Wang AH, Frankland PW. Preferentialincorporation of adult-generated granule cells into spatialmemory networks in the dentate gyrus. Nat Neurosci. 2007;10(3):355–62.
9 Tashiro A, Makino H, Gage FH. Experience-specific functionalmodification of the dentate gyrus through adult neurogenesis: acritical period during an immature stage. J Neurosci. 2007;27(12):3252–9.
10 McNaughton BL, Morris RG. Hippocampal synaptic enhance-ment and information storage within a distributed memorysystem. Trends Neurosci. 1987;10(10):408–15.
11 Rolls ET, Kesner RP. A computational theory of hippocampalfunction, and empirical tests of the theory. Progress Neurobiol.2006;79(1):1–48.
12 Rolls ET, Treves A, Robertson RG, Georges-Francois P,Panzeri S. Information about spatial view in an ensemble ofprimate hippocampal cells. J Neurophysiol. 1998;79(4):1797–813.
13 Butti E, Bacigaluppi M, Rossi S, Cambiaghi M, Bari M, SillaAC, et al. Subventricular zone neural progenitors protectstriatal neurons from glutamatergic excitotoxicity. Brain.2012;135(11):3320–35.
14 Reiner A, Albin RL, Anderson KD, D’Amato CJ, Penney JB,Young AB. Differential loss of striatal projection neurons inHuntington disease. Proc Natl Acad Sci U S A. 1988;85(15):5733–7.
15 Kreitzer AC. Physiology and pharmacology of striatal neurons.Annu Rev Neurosci. 2009;32:127–47.
16 Marxreiter F, Regensburger M, Winkler J. Adult neurogenesisin Parkinson’s disease. Cell Mol Life Sci. 2013;70(3):459–73.
17 Gould E, Tanapat P, Hastings NB, Shors TJ. Neurogenesis inadulthood: a possible role in learning. Trends Cogn Sci (RegulEd). 1999;3(5):186–92.
18 Van Praag H, Shubert T, Zhao C, Gage FH. Exercise enhanceslearning and hippocampal neurogenesis in aged mice. JNeurosci. 2005;25(38):8680–5.
19 Parent JM, Yu TW, Leibowitz RT, Geschwind DH, SloviterRS, Lowenstein DH. Dentate granule cell neurogenesis isincreased by seizures and contributes to aberrant networkreorganization in the adult rat hippocampus. J Neurosci.1997;17(10):3727–38.
20 Kuhn HG, Dickinson-Anson H, Gage FH. Neurogenesis in thedentate gyrus of the adult rat: age-related decrease of neuronalprogenitor proliferation. J Neurosci. 1996;16(6):2027–33.
21 Seki T, Arai Y. Age-related production of new granule cells inthe adult dentate gyrus. NeuroReport. 1995;6(18):2479–82.
22 Gould E, Tanapat P, McEwen BS, Flugge G, Fuchs E.Proliferation of granule cell precursors in the dentate gyrus ofadult monkeys is diminished by stress. Proc Natl Acad Sci U SA. 1998;95(6):3168–71.
23 Sapolsky RM, Packan DR, Vale WW. Glucocorticoid toxicityin the hippocampus: in vitro demonstration. Brain Res.1988;453(1–2):367–71.
24 Sapolsky RM. A mechanism for glucocorticoid toxicity in thehippocampus: increased neuronal vulnerability to metabolicinsults. J Neurosci. 1985;5(5):1228–32.
25 Yang Q, Mu J, Li Q, Li A, Zeng Z, Yang J, et al. A simple andefficient method for deriving neurospheres from bone marrowstromal cells. Biochem Biophys Res Commun. 2008;372(4):520–4.
26 Radad K, Rausch WD, Gille G. Rotenone induces cell death inprimary dopaminergic culture by increasing ROS productionand inhibiting mitochondrial respiration. Neurochem Int.2006;49:379–86.
27 Singhal PC, Reddy K, Ding G, Kapasi A, Franki N, Ranjan R,et al. Ethanol-induced macrophage apoptosis: the role of TGF-b. J Immunol. 1999;162:3031–6.
28 Li H, Zhao J, Chen M, Tan Y, Yang X, Caudle Y, et al. Toll-like receptor 9 is required for chronic stress-induced immunesuppression. Neuroimmunomodulation. 2014;21(1):1–7.
29 Reynolds BA, Weiss S. Generation of neurons and astrocytesfrom isolated cells of the adult mammalian central nervoussystem. Science. 1992;255(5052):1707–10.
30 Meijer OC, de Kloet ER. Corticosterone and serotonergicneurotransmission in the hippocampus: functional implicationsof central corticosteroid receptor diversity. Crit Rev Neurobiol.1998;12(1–2).
31 Roozendaal B. Glucocorticoids and the regulation of memoryconsolidation. Psychoneuroendocrinology. 2000;25(3):213–38.
32 Gould E, McEwen BS, Tanapat P, Galea LA, Fuchs E.Neurogenesis in the dentate gyrus of the adult tree shrew isregulated by psychosocial stress and NMDA receptor activa-tion. J Neurosci. 1997;17(7):2492–8.
33 Muneoka K, Mikuni M, Ogawa T, Kitera K, Kamei K,Takigawa M, et al. Prenatal dexamethasone exposure altersbrain monoamine metabolism and adrenocortical response inrat offspring. Am J Physiol. 1997;273(5 Pt2):R1669–75.
34 Slotkin T, Zhang J, McCook E, Seidler F. Glucocorticoidadministration alters nuclear transcription factors in fetal ratbrain: implications for the use of antenatal steroids. Dev BrainRes. 1998;111(1):11–24.
35 Takahashi L, Goh CS. Glucocorticoid facilitation of choliner-gic development in the rat hippocampus. Neuroscience.1998;83(4):1145–53.
36 Kempermann G. Why new neurons? Possible functions foradult hippocampal neurogenesis. J Neurosci. 2002;22(3):635–8.
37 De Bruin V, Vieira M, Rocha M, Viana G. Cortisol anddehydroepiandosterone sulfate plasma levels and their relation-ship to aging, cognitive function, and dementia. Brain Cogn.2002;50(2):316–23.
38 Murialdo G, Nobili FA, Rollero A, Gianelli MV, Copello F,Rodriguez G, et al. Hippocampal perfusion and pituitary-adrenal axis in Alzheimer’s disease. Neuropsychobiology. 2000;42(2):51–7.
39 Rasmuson S, Nasman B, Carlstrom K, Olsson T. Increasedlevels of adrenocortical and gonadal hormones in mild tomoderate Alzheimer’s disease. Dement Geriatr Cogn Disord.2002;13(2):74–9.
40 Umegaki H, Ikari H, Nakahata H, Endo H, Suzuki Y, OgawaO, et al. Plasma cortisol levels in elderly female subjects withAlzheimer’s disease: a cross-sectional and longitudinal study.Brain Res. 2000;881(2):241–3.
41 Raadsheer FC, van Heerikhuize JJ, Lucassen PJ, HoogendijkWJ, Tilders FJ, Swaab DF. Corticotropin-releasing hormonemRNA levels in the para ventricular nucleus of patients withAlzheimer’s disease and depression. Am J Psychiatry. 1995;152(9):1372–6.
42 Claes SJ. CRH, stress, and major depression: a psychobiolo-gical interplay. Vitam Horm. 2004;69:117–50.
43 Smith SM, Vale WW. The role of the hypothalamic-pituitary-adrenal axis in neuroendocrine responses to stress. DialoguesClin Neurosci. 2006;8(4):383–95.
44 McEwen BS. The neurobiology of stress: from serendipity toclinical relevance. Brain Res. 2000;886(1):172–89.
45 Zubenko GS, Zubenko WN, McPherson S, Spoor E, MarinDB, Farlow MR, et al. A collaborative study of the emergenceand clinical features of the major depressive syndrome ofAlzheimer’s disease. Am J Psychiatry. 2003;160(5):857–66.
46 Shirbin CA, Chua P, Churchyard A, Hannan AJ, Lowndes G,Stout JC. The relationship between cortisol and verbal memoryin the early stages of Huntington’s disease. J Neurol. 2013;260(3):891–902.
47 Sundberg M, Savola S, Hienola A, Korhonen L, Lindholm D.Glucocorticoid hormones decrease proliferation of embryonicneural stem cells through ubiquitin-mediated degradation ofcyclin D1. J Neurosci. 2006;26(20):5402–10.
48 Young EA, Haskett RF, Grunhaus L, Pande A, Weinberg VM,Watson SJ, et al. Increased evening activation of thehypothalamic-pituitary-adrenal axis in depressed patients.Arch Gen Psychiatry. 1994;51(9):701–7.
Abdanipour et al. Long-term cortisol treatment effects
8 Neurological Research 2014 VOL. 000 NO. 000