in vitro assessment of osteoblast behavior in craniosynostosis · pdf filein vitro assessment...
TRANSCRIPT

In Vitro Assessment of Osteoblast Behavior in
Craniosynostosis
by
Tatiana Karine Simon Cypel
A thesis submitted in conformity with the requirements for the degree of Master
of Science
The Institute of Medical Science
University of Toronto
© Copyright by Tatiana Karine Simon Cypel – 2011

ii
IN VITRO ASSESSMENT OF OSTEOBLAST BEHAVIOR IN
CRANIOSYNOSTOSIS
Tatiana Karine Simon Cypel
Masters of Science
The Institute of Medical Science
University of Toronto
2011
ASTRACT
Introduction: The objective of this study is to investigate the role of osteoblasts in
the pathophysiology of premature suture fusion in infants.
Hypothesis: Regional variations in osteoblast function and cell signalling exist in
calvaria of infants with craniosynostosis.
Methods: Bone and periosteal tissue from fused and patent cranial sutures and
adjacent bone were harvested from infants undergoing surgery for craniosynostosis and used
to develop primary osteoblast cell cultures. Dural tissue was obtained from neurosurgical
procedures in order to generate an osteoblast-dural co-culture. Osteoblast proliferation,
differentiation, mineralization, protein expression (Noggin, BMP3 and Runx2) and response
to exogenous FGF2 stimulation were assessed.
Results: Cell cultures demonstrated significant (p<0.05) regional variations in
osteoblast proliferation, alkaline phosphatase and in vitro bone nodule formation. The

iii
expression of anti-osteogenic molecules (Noggin and BMP3) was decreased in osteoblasts
from fused suture regions. Expression of Runx2 was increased in fused suture osteoblasts in
dural co-culture.
Conclusion: The creation of a pro-osteogenic environment through the decreased
expression of anti-osteogenic signalling molecules and increased expression of osteogenic
factors may be responsible for premature suture fusion in infants.

iv
AKNOWLEDGEMENTS
I would like to thank Dr. Christopher Forrest for giving me the opportunity to work in the
exciting and very important field of craniofacial care and research. Dr. Forrest has always
taken time to ensure that I had all the support required to complete my work and he was
always keen to provide me opportunities in order to increase my level of knowledge.
Furthermore, his qualified guidance and enthusiastic supervision in my research project,
along with his commitment to patient care, have provided me with a role model of a
contemporary surgeon.
I would like to also thank Dr. Cho Pang for these two years. Dr. Pang taught me how to think
and work as a scientist.
Drs Iona Leong, Cho Pang and Peter Dirks, as members of my Program Advisory
Committee, have been invaluable in providing their expert guidance and ideas to further
enrich my research and make it productive.
The research outlined here would not have been possible without the technical and
intellectual support of my colleagues in the Craniofacial Surgery Department ( Dr. John
Phillips and the craniofacial clinical fellows) and Neurosurgery Department (Dr. Rutka and

v
Dr. Drake). Homa Ashrafpour and Ning Huang, as our lab manager and technician
respectively, have done excellent work in keeping the lab efficient and supporting my
experiments. Thanks to Balram Sukhu, his expertise in bone cell culture and osteoblast
behavior made this work possible.
I also would like to thank Dr. Rinaldo De Angeli Pinto, chair of the Division of Plastic
Surgery (Federal University of Rio Grande do Sul) where I performed my plastic surgery
training in Brazil. Dr. De Angeli was for me, an example of a superb, ethical, and highly
competent surgeon. He created the surgical “personality” I currently have and hopefully I
will carry that for my entire career.
I would like to acknowledge the contribution of Nicole Gojska in assisting with technical
work in the conditioned culture medium project.
No research would be possible without sufficient resources. I would like to thank the
Craniofacial Care and Research Funding, The SickKids Start up Funding, The Komedyplast
and Amercian Society of Craniofacial Surgery, The Physician’s Service Incorporate
Foundation and the American Society of Maxillofacial Surgeons for supporting and making
this work possible.

vi
DEDICATION
This thesis is dedicated to my family: my parents and two sisters who have always supported
me and stood behind me and my husband Marcelo, who has shared my challenges and
successes with the greatest of understanding, patience and support.

vii
TABLE OF CONTENTS
Page
I. List of Tables x
II. List of Figures x
III. List of Appendices xiii
IV. List of Abbreviations xiv
V. Introduction 1
(a) Overview of Craniosynostosis 2
- Classification 3
- Functional Problems Associated with Craniosynostosis 5
- Surgical Treatment 7
(b) Pathogenesis of Craniosynostosis 9
- Embryology of Cranial Suture 9
- Normal Skull and Suture Growth 10
- Normal Suture Fusion 10
- Historical Theories of Craniosynostosis 11
- Current Theories 12

viii
- The Role of Anti-osteogenic Signalling 13
- The Role of Dura Mater 17
- The Role of Runx2 18
- The Role of FGFs 20
- Experimental Models for Craniosynostosis Research 22
- Effect of Culture Medium Composition on Osteoblast Function 23
VI. Hypothesis 27
VII. Material and Methods 30
- In Vitro Human Osteoblast Cell Culture Model 31
- Statistical Analyses 38
VII. Results
- Demographics 40
- Histology 41
- Collagen I Expression 42
- Validation of Cell Culture Model 43
- Medium Composition 45
- Osteoblast Proliferation 47
- Runx2 Expression 52
- Alkaline Phosphatase Activity 54
- Mineralization 58
- Transmission Electron Microscopy 63
- Expression of Anti-osteogenic Signalling Molecules 64
- Dura Mater Expression of FGF2 and TGF-β1 70

ix
- Recombinant Human FGF2 Stimulation 71
VIII. Discussion 77
IX. Conclusion 93
X. Future Directions 96
XI. References 99
XII. Appendices 110
- Detailed Protocols 111

x
LIST OF TABLES
Page
Table 1: Classification of Craniosynostosis 3
Table 2: Demographics of Patients with Craniosynostosis Enrolled in the Study 40
LIST OF FIGURES
Page
Figure 1: Single-suture Synostosis Phenotypic Expression 4
Figure 2: Mechanism that BMP3 and Noggin Cause Inhibition of Bone Formation 16
Figure 3: The Role of Runx2 in Osteogenic Differentiation 19
Figure 4: Potential Mechanisms of Craniosynostosis 26
Figure 5: Hematoxylin-and eosin Staining for Bone Tissue Sampling 41
Figure 6: Immunohistochemistry Staining for Collagen I 42
Figure 7: Evidences of Osteoblasts in our Cell Culture Model 43
Figure 8: MTT Assays of Human Cranial Suture-derived Osteoblasts 46
Figure 9: Cellular Growth Prior and Post-subculture 47
Figure 10: Osteoblast Proliferation Rates (MTT) 48
Figure 11: Proliferation Rates for Syndromic Patients 49

xi
Figure 12: Osteoblast Proliferation Rates in Co-culture with Dura Mater 50
Figure 13: Expression of Runx2 Demonstrated by Immunohistochemistry 52
Figure 14: Expression of Runx2 by Western Blot 53
Figure 15: Alkaline Phosphatase Assay Assessing Osteoblast Differentiation Rates 54
Figure 16: Alkaline Phosphatase Activity 55
Figure 17: Analysis by qRT-PCR of Alkaline Phosphatase and Osteocalcin 56
Figure 18: AP for Osteoblast Co-cultured with Dura Mater Cells 57
Figure 19: Bone Nodule Formation – Alizarin Red Assay 59
Figure 20: Bone Nodule Formation at Days 14 and 18 60
Figure 21: Mineralization at Days 21 and 28 61
Figure 22: Mineralization for Osteoblasts Co-cultured with Dura Mater 62
Figure 23: Transmission Electron Microscopy of Bone Nodules 63
Figure 24: Immunohistochemistry Analysis of Noggin in Tissue Samples 64
Figure 25: Graphic Analysis for Noggin in the Tissue Samples 65
Figure 26: Immunohistochemistry Analysis of BMP3 in Tissue Samples 66
Figure 27: Graphic Analysis for BMP3 in the Tissue Samples 67
Figure 28: Western Blot Analysis of BMP3 and Noggin 68

xii
Figure 29: Expression of Pro-osteogenic Molecules in the Conditioned Medium from
Dura Mater Cells 70
Figure 30: Proliferation Rates of Osteoblasts Stimulated with FGF2 72
Figure 31: Proliferation Rates after Stimulation with FGF2 (MTT) 73
Figure 32: AP Staining after Stimulation with FGF2 75
Figure 33: APA after Stimulation with FGF2 76

xiii
LIST OF APPENDICES
Page
I. Calvarial Osteogenic Cell Culture 111
II. Medium Composition Study 114
III. MTT Assay 116
IV. Alkaline Phosphatase Activity 117
V. Alkaline Phosphatase Staining 118
VI. Alizarin Red Staining 119
VII. Transmission Electron Microscopy 119
VIII. Histology 120
IX. Immunohistochemistry and Immunofluoresence 120
X. Western Blot 121
XI. Real Time PCR 124
XII. Materials 126

xiv
LIST OF ABREVIATIONS
AMEM (αAMEM) Alpha Modified Eagle Medium
ANOVA Analysis of Variance
AP Alkaline Phosphatase
APA Alkaline Phosphatase Activity
BMP2 Bone Morphogenic Protein 2
BMP3 Bone Morphogenic Protein 3
BMP4 Bone Morphogenic Protein 4
BMP7 Bone Morphogenic Protein 7
CT Computed Tomography
DMEM Dulbeco Modified Eagle Medium
Cbfa1 Core binding factor alpha 1
DNA Deoxyribonucleic acid
FBS Fetal Bovine Serum
FGFs Fibroblast Growth Factors
FGFR Fibroblast Growth Factor Receptor
FGF2 Fibroblast Growth Factor 2
FS Fused Suture
IHC Immunohistochemistry
I-SMADS Inhibitory Smads
MTT Methylthiazoletetrazolium
PS Patent Suture

xv
qRT-PCR Real Time Quantitative Reverse Transcripition PCR
REB Research Ethical Board
RNA Ribonucleic Acid
SMAD Sma and Mad Related Proteins
TEM Transmission Electron Microscopy
TGF-β Transforming Growth Factor-β
WB Western Blot

1
Introduction

2
OVERVIEW OF CRANIOSYNOSTOSIS
Craniosynostosis, the premature fusion of one or more cranial sutures, is a relatively
common congenital disorder, affecting as many as 1 in 2000 to 2500 live births worldwide1.
Hippocrates provided the first description of this anomaly in 100 B.C. He noted the variable
appearance of the calvarial deformity and correlated it with the pattern of cranial suture
involvement. In 1851, Virchow was the first one to recognize that cranial sutures were
responsible for growth of the calvarium at right angles to the suture and that premature fusion
resulted in growth arrest at right angles to the suture with compensatory growth at patent
sutures2.
Premature ossification of cranial sutures may lead to a number of serious
morphologic and functional consequences, such as increased intracranial pressure,
developmental delay, visual and hearing impairment and can require major staged
reconstructive procedures to correct the condition. Despite its prevalence, the cause of
craniosynostosis still remains largely unknown.
Craniosynostosis may be classified by the number of involved sutures, etiology or
whether the condition is associated with a syndrome or not (Table 1).

3
Table 1: Classification of Craniosynostosis
Anatomical
Single Suture Multiple Sutures
Sagittal Any combination possible – usually bi-coronal
Metopic
Unicoronal
Lambdoid
Minor suture (fronto-sphenoid, zygomatico-temporal)
Etiology
Primary
Non-Syndromic Syndromic
Scaphycephaly (sagittal) Apert’s syndrome
Trigonocephaly (metopic) Crouzon’s syndrome
Anterior plagiocephaly (unicoronal) Pfeiffer syndrome
Posterior plagiocephaly (lambdoid) Jackson-Weiss syndrome
Turribrachycephaly (bi-coronal) Carpenter syndrome
Oxycephaly (delayed bicoronal) Muenke syndrome
Saethre-Chotzen syndrome
Secondary
Biomechanical (shunt)
Bone metabolic disorders
Nutritional
The major cranial sutures involved (in order of decreasing frequency) are the sagittal,
metopic, coronal and lambdoid sutures and each give a characteristic shape to the cranial
vault (Figure 1).

4
Figure 1: Single-suture Synostosis Phenotypic Expression: Single suture non-
syndromic craniosynostosis phenotypic and radiologic (CT) features.
Single suture craniosynostosis is usually an isolated phenomenon and has a low
incidence of familial occurrence, generally considered to be less than 5%. The highest
incidence of familial occurrence is with sagittal synostosis, which has been reported to be as
high as 8%3.
While the majority of craniosynostosis occur due to unknown causes,
craniosynostosis can occur as a result of metabolic disorders (e.g., hyperthyroidism),
malformations (e.g., holoprosencephaly, microcephaly, shunted hydrocephalus,

5
encephalocele), drug exposure (e.g., valproic acid, phenytoin) or mucopolysaccharidosis
(e.g., Hurler’s syndrome, Morquio’s syndrome).
FUNCTIONAL PROBLEMS ASSOCIATED WITH CRANIOSYNSTOSIS
Premature fusion of the cranial sutures may be associated with a variety of clinical
problems ranging from cranial facial dysmorphism to increased intracranial pressure due to
cranio-cerebral disproportion. The severity and extent of involvement is dependent upon the
number of affected sutures and the presence or absence of an associated syndrome. This
section details the potential functional problems associated with craniosynostosis.
Increased Intracranial Pressure: Elevated intracranial pressure may be associated
with craniosynostosis4. Renier et al. published the first major study to measure the
preoperative and postoperative differences in intracranial pressure in infants with
craniosynostosis. Nonsyndromic patients overall had a 14% incidence of elevated intracranial
pressure before surgery. The number of involved sutures affects the probability that
intracranial pressure will be elevated. Preoperatively, 8.9% of the patients with one suture
affected had elevated intracranial pressure but those with multiple sutures involved had a
45% incidence of intracranial hypertension. The effect of craniosynostosis, elevated
intracranial pressure, and syndromic disease on intelligence is less clear. In general, for most
nonsyndromic patients, intelligence is normal with single-suture involvement (>90% IQ ≥
90) but decreases with multiple suture involvement (78% IQ ≥ 90)5. For syndromic patients,
the degree of developmental delay is highly variable, depending on the syndrome and
severity of disease.

6
Orbits: Exorbitism is a significant component for syndromic patients with
craniosynostosis. Hypoplastic orbits and a retruded midface can cause globe and corneal
exposure, which can result in corneal injury and exposure keratitis. The majority of orbital
problems occur relatively early, in the first 5 years of life which represents the main period of
orbital growth.
Airway: Syndromic craniosynostosis may be associated with airway compromise that
requires early recognition and treatment to prevent cardiopulmonary and neurologic
sequelae6. This may be due to midface retrusion and choanal atresia resulting in airway
obstruction. The reported incidence of airway obstruction in syndromic craniosynostosis
ranges from 40% to 100%6. Many of these children will additionally have lower airway
anomalies that include tracheomalacia, complete cartilaginous tracheas, and granulations.
Airway obstruction and hypoxia during sleep may present as snoring, noisy respirations,
apneic episodes, paradoxical chest movements, persistent restlessness, feeding difficulties,
failure to thrive, hypertension, daytime fatigue, and cardiopulmonary or neurologic
impairment7. Complications of persistent airway obstruction include respiratory infections,
cor pulmonale, neurologic dysfunction and brain damage.7
Neurodevelopment Particularly problematic is the issue of intelligence and
neurocognitive development. Virtanen et al8 contended that in a series of patients undergoing
early operative correction for sagittal craniosynostosis, certain indices of neurocognitive
performance were below those of age-matched control subjects and remained delayed
throughout the period of examination after surgery. Other authors contended that although
young children with craniosynostosis are often normal from a mental standpoint, there is an
increase in frequency of psychomotor problems as they develop5.

7
Aesthetic/Psychosocial Aesthetic considerations are more difficult to quantify than
the objective values of protecting vision, reducing intracranial pressure, or improving
occlusion. Infants with craniosynostosis have visibly altered craniofacial appearance, which
varies in relation to the location and extent of suture involvement. The possibility of
permanent abnormality in facial or cranial appearance seems to greatly
affect parents’
decisions to have their infant undergo craniofacial surgery. Many surgeons believe that the
primary indication for cranioplasty in isolated synostosis is cosmetic
rather than functional
9.
There is clear evidence in other groups of children that even mild deviations from typical
facial appearance can have significant impact on psychological adjustment. Congenital
defects involving an infant’s face and skull seem to evoke particularly strong emotional
responses from parents, who must contend with a host of potentially stressful
events and
circumstances, including the infant’s unusual
appearance, potentially life-threatening
surgeries and other medical procedures, and the possibility of future neuropsychological
and
educational problems. All of these factors can potentially affect parents’ responsiveness
and
adaptation to the infant with craniofacial abnormality10
.
SURGICAL TREATMENT
The field of craniofacial surgery has existed for only a short time in the overall
history of medicine. Of course, the patients have always been there, with a birth prevalence
of craniosynostosis of approximately 1 in 2000 live births1; but it is only since Paul Tessier
began his pioneering work in the late 1960s that the field of craniomaxillofacial
reconstruction developed. Since the 1970s the specialty of craniofacial surgery has grown to
include multidisciplinary teams treating a wide variety of patients.

8
Indications for Treatment
In nonsyndromic patients, individual cranial sutures may be fused, resulting in an
abnormality of shape requiring cranio-orbital reshaping, but the midface is generally
unaffected. In syndromic craniosynostosis, in addition to cranial suture abnormalities, other
facial skeletal anomalies exist including a shortened cranial base, orbital hypoplasia, midface
and zygomatic retrusion to name a few. The reasons for surgical intervention in the
nonsyndromic group may be primarily aesthetic; in the syndromic group, they are
multifactorial. Aesthetic considerations are more difficult to quantify. Nevertheless, the
benefits of improving appearance on psychosocial adjustment have been evaluated across
many different populations11
.
Timing of Surgery
Advances in pediatric anesthesia and intensive care have allowed extensive cranial
reconstructions in infancy that previously were available only to adolescents and adults.
Concerns for the negative effect of early intervention on skeletal growth were reduced by
studies demonstrating that in syndromic patients, the midface was deficient whether patients
had early surgery or not12, 13
.
McCarthy and Cutting14
proposed that the first procedure, cranial vault remodeling
and fronto-orbital advancement, be performed between 6 and 9 months of age. At an earlier
age, the bone is more fragile; at a later age (>18 months), residual calvarial defects will fail
to reossify. Early surgery is also important because of rapid growth of the brain, which more
than doubles in volume during the first year of life15
.

9
Pathogenesis of Craniosynostosis
During cranial development, adjustment to the expanding brain takes place by bone
deposition at sutural margins and on the ectocranial surface of the calvarium, and by bone
resorption on the endocranial surface16
. The cranial sutures, as primary areas of growth
during the expansion of the cranium16, 17
play a pivotal role. Although a descriptive
knowledge of suture morphogenesis and function is well reported16, 18
, the spatial and
temporal regulation of bone deposition, resorption, and remodeling are not well understood19
.
Embryology of Cranial Suture
Sutural morphogenesis occurs in midgestation when enlarging bone plates in the
primordial cranium come into apposition16, 17
. Mesenchymal cell populations along the
expanding osteogenic fronts differentiate into osteoblasts and contribute to the formation of
osteoid and the mineralization of new bone17, 18
, bringing opposing bones into close
approximation. An intervening zone of immature mesenchymal tissue is thereby created
between them, forming the blastema of the suture18, 20
Suture mesenchymal cells continue to
proliferate and organize within the fibrous suture matrix, which acquires the characteristic
appearance of the mature, multilamellar suture17, 18
. Simultaneously, populations of
mesenchymal cells bordering the suture along the osteogenic fronts continue to differentiate
into osteoblasts, contributing to the formation of new bone17, 18
. From the time of their
formation, sutures are extremely active centers of cellular proliferation, cellular
differentiation, tissue synthesis, and remodeling. The intricate regulation of these processes
allows for the completion of cranial morphogenesis while preventing the formation of bone

10
across the sutural space, an event which occurs only in the pathologic condition of
craniosynostosis or part of normal suture fusion later in adult life19, 21, 22
.
Normal Skull and Suture Growth
Intramembranous ossification of the skull begins at the end of the second month of
gestation. A center of osteogenesis develops directly in vascularized mesenchyme. Expansion
of the ossification center proceeds rapidly via appositional growth. Initially cancellous bone
forms, but as trabeculae thicken and the bone becomes less porous, it become compact bone.
Eventually each intramembranous cranial bone has enlarged to the point at which it
articulates with an adjacent bone via a syndesmosis or sutures. Growth then proceeds at the
sutures19
.
Growth at the suture area is a secondary, compensatory, and mechanically obligatory
event following the primary growth of the enclosed brain and ocular globes. The bones of the
calvaria are displaced outward by the enlarging brain. Each bone of the domed skull roof
responds to the expansion of the brain by depositing new bone at the contact edges of the
sutures.
Normal Suture Fusion
Functioning sutures are the sites of continuous bone deposition and resorption.
Initially, sutures are straight edges of bone separated by connective tissue. Gradually,
interdigitations develop and become more prominent with time19
. For interdigitations to
form, develop, and interlock, the distribution of osteoblasts along sutural bone must be
uneven with clumps of osteoblasts at the tip of each interdigitation23
. Sutural interdigitations

11
may permit adjustive movements and/or stress reduction. Their architecture may depend on
the types and distribution of forces.
Suture closure has been attributed to vascular, hormonal, genetic, mechanical, and
local factors. Biomechanical factors have been a perennial favorite mechanism24
. The cause
of suture closure is still unclear. There may be one or possibly more than one mechanism.
The relationship between suture closure, cessation of growth, and functional demands across
sutures poses questions about various biological relationships. Does cessation of growth lead
to suture fusion? The growth of the human brain ceases prior to the onset of osseous fusion
of the cranial sutures. With this is a delay from completion of brain growth to sutural fusion
in the 20s and 30s.
Historical Theories of Craniosynostosis
Although certain cranial deformities arise from mechanical or functional causes (e.g.,
plagiocephaly and hydrocephaly), the molecular basis of the majority of craniofacial
abnormalities is becoming increasingly evident through advancements in molecular biology.
Early explanations of cranial suture fusion included anectodal associations with intrauterine
constraint, uterine malformations, decreased amniotic fluid, or breech presentation. Ozaki et
al25
performed ultrastructural analysis of sagittal sutures in the process of fusion. Their
analysis revealed several new facts:
1. Premature fusion of sutures was found to begin centrally in the suture.
2. It began endocranially as opposed to both endocranially and ectocranially in
normal sutures.

12
3. It exhibited a disorganized ultrastructure of lower density than in normal sutures.
Molecular biology has now taken us beyond the speculative explanations of
mechanical causes to the roots of abnormal suture development. This progression is
particularly evident in autosomal dominant syndromic craniosynostosis subtypes.
Current Theories
Recent work has demonstrated that fusion of the calvarial sutures is mediated by
locally elaborated soluble growth factors, leading some to speculate that external
biomechanical forces play little role in suture development. Historically, the theory that fetal
head constraint may play a critical role in the pathogenesis of many cases of nonsyndromic
craniosynostosis has been supplanted by humoral theories although it is possible that cranial
biomechanical stresses experienced in fetal and early life might be the trigger that leads to
dural cytokine signalling involved with suture fusion and/or patency26
.
On the other hand, research focused on the molecular mechanisms underlying normal
cranial suture fusion has demonstrated the importance of dura mater mediated cell signalling
in the complex process of fusion of normal cranial sutures. It is hypothesized that the dura
mater acts as a regionally specific endogenous tissue engineer, releasing growth factors in a
specific orchestrated fashion that cause the overlying cranial suture to close in a predictable
fashion. Possible candidates for these growth factors include fibroblast growth factor (FGF),
and transforming growth factor-β (TGF-β) isoforms.
In addition to the effects of these locally released growth factors, cranial suture
development has been shown to be influenced by anti-osteogenic signalling molecules such

13
as Noggin and Bone Morphogenic Protein 3 (BMP3), which are upregulated in patent sutures
during the normal process of suture fusion thereby maintaining the cranial suture patency27,
28. Furthermore, the role of Runx2, a transcription factor that is a marker of osteoblast
differentiation, has been implicated in the process of normal cranial suture fusion. Runx2 is
found in osteogenic fronts and sutural mesenchyme and has been demonstrated to be
upregulated in fusing sutures during the process of normal suture fusion. Runx2 up regulation
enhances differentiation and bone production leading to earlier suture fusion. This factor has
been shown to regulate the expression of a number of proteins, including osteocalcin,
produced by the mature osteoblast and responsible for its bone formation29
.
The Role of Anti-osteogenic Signalling
Noggin
Noggin, a secreted BMP2/4 antagonist produced by osteoblasts and released in the
extracellular matrix, is important in the process of normal suture fusion27
. Noggin is a
polypeptide that inhibits Transforming Growth Factor – β (TGF-β) signal transduction by
binding to TGF-β family ligands and preventing them from binding to their correspondent
receptors (Figure 2). Down-regulation of pro-osteogenic BMP signalling (part of the TGF-β
superfamily) is then observed which maintains suture patency27
. On the other hand, down-
regulation of Noggin expression results in disinhibition of pro-osteogenic BMP signalling
(BMP2,4,7) increasing bone formation which leads to suture fusion and may be one of the
molecular mechanisms involved in the pathophysiology of craniosynostosis.
Research findings in a murine model of normal suture fusion demonstrate that down-
regulation of Noggin expression in the normally fusing posterior frontal suture increased

14
bone formation with resulting suture fusion. In a normal non-fusing suture (sagittal),
increased Noggin expression results in suture patency27
. It has also been shown that the
suture-specific dura mater is an independent source of Noggin. Cultured dura mater cells
from patent sutures expressed high levels of Noggin protein, whereas the dura mater from the
fusing posterior frontal suture expressed almost undetectable levels of Noggin30
.
Experiments in a rabbit model of congenital coronal craniosynostosis also
demonstrated the interaction between Noggin and premature suture fusion. Fusing sutures
showed low Noggin expression29
. Underexpression of Noggin was also found in the dura and
coronal mesenchyme prior to suture fusion. In contrast, in the same model, the patent coronal
and sagittal sutures expressed normal levels of Noggin leading to suture patency29
.
Despite the current findings strongly suggesting an important role for Noggin in
maintaining suture patency in animal models of normal suture fusion, there is a lack of
understanding about Noggin expression and its interactions in infants with craniosynostosis.
Bone Morphogenic Proteins
The BMPs are growth factors secreted by osteoblasts and released in the extracellular
matrix. They are part of the TGF-β superfamily, which are well known for their ability to
induce the formation of bone and cartilage. The actions of these growth factors are highly
concentration dependent and influence a number of cellular processes. For instance, BMP2, 4
and 7 have been shown to promote cellular chemotaxis and proliferation at low extracellular
concentrations and to induce cellular differentiation and bone formation at high extracellular
concentrations31
.

15
BMP3 is an antagonist of BMP2 and BMP4 32
. Rather than impeding BMP signalling
of bone formation by binding to a ligand and preventing specific ligand-receptor interactions
(as does Noggin), BMP3 activates a TGF-β/activin–specific response pathway33
(Figure 2).
Activin is a member of the TGF-β superfamily, and antagonizes the BMP pathway by
competing for SMAD proteins; SMAD proteins are transcription factors that regulate the
expression of genes involved in the modulation of the activity of TGF-β ligands involved in
osteoblast differentiation and bone formation34
.
BMP3 has been implicated in the process of normal suture fusion in mice28
. Altough
the source of BMP3 during normal suture fusion is not clear, its expression pattern is
consistent with that of an antagonist playing a role in suture fusion and patency. BMP3 levels
decreased in the posterior frontal suture during suture fusion and were maintained or
increased in the patent sagittal suture35
. It is speculated that BMP3 may be negatively
regulated by osteogenic factors such as FGF2 and TGF-β1 which are differentially expressed
in the fusing posterior frontal and sagittal suture complexes36, 37
. These factors are noted to
increase in the dura mater underlying the fusing posterior frontal suture during fusion when
compared with the patent sagittal suture30
.

16
Activin ResponsePathway
BMP2
Noggin
SMAD Activation andNuclear TranslocationResulting in Bone Formation
Role of Noggin and BMP3 Signalling
Promotes bone formation
Binds to BMP2 blocking the
stimulus for bone formation
Antagonize bone formation by
competing for SMADS protein
BMP-3 activates a TGF-β/activin-specific response antagonizing osteogenic signaling
BMP3
No signalling
Figure 2: Mechanism by which BMP3 and Noggin cause inhibition of bone formation.
Schematic of Noggin and BMP3-mediated antagonism of bone morphogenetic protein (BMP)
signalling. Antagonists such as Noggin bind to BMP ligand and prevent ligand-receptor interactions.
BMP3 binds to TGF-β/activin receptors and blocks BMP signalling downstream of activated BMP
receptor complexes.
Despite the insights in Noggin and BMP3 expression during the normal suture fusion
process, it is not clear if down-regulation of these anti-osteogenic molecules during
premature suture fusion is a cause or the effect of pro-osteogenic activation leading to
premature ossification in craniosynostosis. Additional investigation of the expression of
osteogenic antagonists and their regulation will further advance our knowledge of the

17
complex cascades regulating suture fusion and patency in infants with syndromic and non-
syndromic craniosynostosis.
The Role of Dura Mater
Central to many studies of cranial sutures has been the role of the dura mater.
Historically, dura mater was thought to be a conduit for tensile forces transmitted from the
expanding neurocranium38, 39
. The formation of sutures was seen as a byproduct of this
mechanical phenomenon, forming along dural reflections in both normal and disease states39
.
While several animal models have established the importance of dura mater in the
regeneration of normal cranial bone40
and sutures in developing animals, evidence suggests
that cell signalling and humoral mechanisms are more important than biomechanical forces
with respect to bone regeneration41
.
Recent findings suggest that dura mater modulates calvarial ossification in many
ways, including providing a source of osteoblastic precursors and/or supplying osteogenic
cytokines42
. Evidences suggest that the underlying dura mater also influences the behavior of
the overlying suture complex by means of paracrine signalling43
. The dura mater underlying
the cranial suture complex is one of several sources of FGF2 and TGF-β1cytokines in vivo;
however it remains unclear whether the levels of these growth factors produced by dura
mater are capable of down-regulating anti-osteogenic molecules expression in osteoblasts to
favor a pro-osteogenic enviroment and promote premature suture fusion44
.
Li et al44
demonstrated direct evidence for a paracrine effect of juvenile dura mater
cells on osteoblasts by showing that dura mater derived FGF2 mediates mitogenic activity in
calvarial osteoblasts which is inhibited by neutralizing FGF245
. Osteoblasts demonstrated
significantly increased proliferation when combined with juvenile dura mater cells in co-

18
culture or when dura mater cell-conditioned medium was applied to them. Moreover high
levels of FGF2 protein were detected in juvenile dura mater cells and their conditioned
medium. In contrast low levels of FGF2 protein were detected in adult dura mater cells and
not detectable levels in their conditioned medium. This study reinforced the idea that FGF2
might be an important paracrine signalling factor in vivo supplied by the underlying dura
mater to stimulate the overlying calvarial osteoblasts44
.
The idea that dura mater derived from immature animals is osteoinductive and/or
osteogenic in nature was further supported by studies in which heterotopic transplantation of
the dura mater into epithelial mesenchymal pockets in adult rats caused ectopic bone
formation42
. Furthermore, when dura mater from adult guinea pigs (18 months of age) was
grafted into the base of calvarial defects created in syngeneic infant guinea pigs (3 to 4 weeks
old), incomplete reossification was observed40
. In contrast, dura mater taken from an infant
rat and placed into the calvarial defect of an adult rat markedly enhanced reossification46
.
Therefore immature dura mater seems to have a strong influence on the development
of bone formation in vivo and as such we may expect the dura mater to have similar
importance in suture regulation. Despite the advancements in the understanding of the pivotal
role of dura mater in premature suture fusion there still is a lack of information regarding the
interactions of dura mater pro-osteogenic signalling and anti-osteogenic molecules leading to
premature calvarial ossification in infants with craniosynostosis.
The Role of Runx2
Polyomavirus enhancer binding protein 2/core binding factor Alpha 1 (Cbfa1) or
currently denominated Runx2 is a master transcription factor that has been shown to regulate

19
osteoblast differentiation stimulating osteogenic gene transcription through a cascade,
starting with BMP-2 binding to its receptor (BMPR-II). This binding activates a SMAD
(signal transducers for the members of the transforming growth factor-beta superfamily)
signalling cascade, ultimately activating Runx2 and stimulating osteogenesis (Figure 3). It
also regulates the expression of a wide variety of genes responsible for the osteoblast
phenotype and function including osteocalcin and TGF-β47
. The latter provides a direct link
between transcriptional regulation and growth factor activity.
Figure 3: Role of Runx2 in osteogenic differentiation. BMP2 binds to its receptor
(BMPR-II). This binding activates a SMAD signalling cascade, ultimately activating Runx2
and stimulating osteogenic gene transcription.
Reflecting its major role in bone formation, Runx2 levels have been shown to be
elevated in areas of normal suture formation in mice48
. Mutations in which Runx2 is absent
demonstrate defects in osteogenesis49
. These studies provide a sound basis for an effort to

20
examine the activity of specific transcription factors such as Cbfa1/Runx2 in osteoblasts
derived from fused sutures and to determine whether they play a causative role in sutural
closure associated with craniosynostosis.
The Role of Fibroblast Growth Factors
Fibroblast Growth Factor 2 is a member of the fibroblast growth factor family
involved in angiogenesis, wound healing, and embryonic development50
. The FGFs are
heparin-binding proteins and interactions with cell-surface associated heparin sulfate
proteoglycans have been shown to be essential for FGF signal transduction. FGFs are key
players in the processes of proliferation and differentiation of wide variety of cells and
tissues. Several observations indicate that FGFs may play an important role in the control of
osteogenesis during skeletal development. FGF2 is a potent mesodermal inducer during
embryogenesis and FGF receptors (FGFRs) are strongly expressed in developing bones50
.
Studies in bovine calvaria cells showed that FGF2 is produced by osteoblasts and
accumulates in the bone matrix51
. In bovine and rodent calvaria-derived cells, the effects of
FGF on bone cell proliferation and differentiation appear to be opposite. FGF1 and FGF2
stimulate cell proliferation but inhibit alkaline phosphatase (AP) activity and reduce collagen
type I (ColI) and osteocalcin (OC) expression, indicating that FGF2 has independent effects
on calvarial cell proliferation. The effects of FGFs on osteoblastic cell differentiation and
bone matrix formation in long-term culture are however conflicting, since positive52, 53
and
negative effects54
have been reported, depending on the cell culture system.
Considering that dura mater cells may be a source of FGF2 and anti-osteogenic
molecules such as Noggin may be regulated by FGF2, it is important to know if this

21
molecule is the key element in the pathophysiology of craniosynostosis, which may
corroborate with the hypotheses that dura mater influences osteoblast behavior at the fused
suture site through release of FGF2 leading to decreased Noggin and BMP3 expression. It is
also unclear if osteoblasts from different sites are defective and not able to respond equally to
FGF2 stimulation, or if there is an up-regulation of this molecule at the fused suture site.
While blocking FGF signalling30
or FGF2 activity55
prevents cranial suture fusion or
osteogenesis, respectively, the findings suggest that exogenous FGF signalling is capable of
suppressing Noggin expression during cranial suture fusion.
Many researchers have concentrated efforts on investigating the genetic basis of
syndromic craniosynostosis and the functional consequences of mutations involving the
fibroblast growth factor receptor (FGFR) gene. Recent findings shown that point mutations
in FGFR-1 and FGFR-2 induce premature cranial ossification suggesting that FGF is an
important regulator of bone-forming cells during human calvaria (HC) osteogenesis56
.
Mutations of three FGFRs account for most causes of syndromic craniosynostosis, including
Crouzon’s, Crouzon’s with acanthosis nigricans, Pfeiffer’s, Apert’s, Muenke’s, Beare-
Stevenson, and Jackson-Weiss syndromes57
. On activation, the FGFR immunoglobulin-like
bonding regions form dimers, activating the intracellular tyrosine kinase. Subsequent
downstream effects on the nucleus influence cellular proliferation, differentiation, and
migration. Characterization of specific mutations in genes that cause craniosynostosis is a
step forward understanding the mechanism of normal and abnormal development of calvarial
bone. However, various approaches of research in this area are still needed to help unravel
the complex interaction of gene products that participate in signalling pathways58
.

22
Taken all findings together, FGF2 may guide suture fate (patency versus fusion). It is
also important to clarify the role of FGF2 in proliferation and differentiation, and to
investigate if different concentrations at the suture site are responsible for the regional
differences in osteoblast behavior. Furthermore, it is important to elucidate whether dura
mater is the source of FGF2 and evaluate the ability of osteoblast from different sites to
respond to FGF2 stimulation, clarifying one step of cascade that leads to craniosynostosis in
infants.
Experimental Models for Craniosynostosis Research
Over the past several years, investigation of the biology underlying programmed
posterior frontal suture fusion in rats and mice has been taken as a means of understanding
the pathology seen clinically in human craniosynostosis44
. The murine model has been
thought to be an excellent system with which to study suture development and molecular
specification between mice and humans45
. In the mouse, the posterofrontal suture lies
between two frontal bones, and the sagittal suture lies between the two parietal bones. The
posterofrontal suture undergoes fusion in a predictable manner on postnatal days 8 to 10,
whereas other sutures remain patent28
. This is analogous to humans, in which the metopic
suture fuses in infancy and the other sutures remain patent well into adulthood. Thus, the
murine posterofrontal and the sagittal sutures taken in juxtaposition, as exemplars of normal
suture fusion and patency, respectively, allow for insight into both the normal coordination of
suture fusion and possible mechanisms of craniosynostosis19
.
Currently, the most representative animal model of craniosynsostosis is the rabbit
craniosynostosis strain from the University of Pittsburgh58
. In this model, pathologic suture

23
fusion begins in utero, causing cranial vault deformities such as plagiocephaly in unilateral
coronal suture synostosis and brachycephaly in bilateral synostosis59
. This model has made it
possible to investigate the biomolecular mechanisms involved in craniosynostosis, including
the role of anti-osteogenic molecules such as Noggin and pro-osteogenic factors such as
Runx2.
Although there are few studies utilizing a limited number of discarded samples from
patients with craniosynostosis60
, to date there are no reliable models of cell cultures derived
from human calvarial bone using a large number of patients and well representing the wide
spectrum of craniosynostosis in infants.
Effect of Culture Medium Composition on Osteoblast Function
The use of normal and fused suture osteoblasts derived from cranial bones of patients
with syndromic and non-syndromic craniosynostosis provides a valuable model for
investigating molecular and cellular defects associated with this significant disorder in
infants60
. However, there is little uniformity in the conditions used in human osteoblast cell
cultures, particularly the concentration of reagents present in the media. Initial studies by
Coelho et al61
analyzed the effects of two widely used culture media, Dulbecco’s modified
Eagle’s medium (DMEM) and minimum essential medium Eagle Alpha modification (α-
MEM) on human osteoblastic characteristics, including cell viability and alkaline
phosphatase (AP) activity61
. These studies demonstrated that DMEM, a less nutrient-rich
medium when compared to α-MEM, appears to demonstrate higher values of cell
proliferation and growth61, 62
. Subsequently, many studies have found that the optimal
concentration of fetal bovine serum (FBS) to supplement the culture medium is FBS 10%,

24
for it produces the highest proliferation rates62, 63
. The success of cranial suture biology in
explaining the pathophysiological mechanism of craniosynostosis is predicated in replicable
and efficient cell culture systems that are representative of in vivo cellular dynamics.
Therefore, the standardization of culture conditions, especially the medium and the presence
of essential compounds, is critical for developing future applications of cranial suture
research in reconstructive medicine.
Summary of Research
Current research has proposed the molecular basis of craniosynostosis based on
normal suture fusion animal models. It has been shown that the patency of the suture line
depends on the balance between pro-osteogenic and anti-osteogenic signalling molecules,
and the imbalance between those may be responsible for premature suture fusion. However,
is not clear which changes in the cranial suture enviroment are responsible for this
phenomenon. To elucidate the mechanisms locally involved in premature suture fusion we
propose to create a human model of osteoblast cell culture by obtaining calvarial bone
samples from patent sutures, fused sutures and adjacent bone from infants affected by this
condition. First, we propose to evaluate the samples histologically to confirm regional
variations between suture sites and adjacent bone. We then aim to search in the surrounding
enviroment for factors that may contribute to the imbalance between pro- and anti-osteogenic
signalling. It has been shown that dura mater may be the source of growth factors, such as
FGF2 and TGF-β that orchestrate this complex process of suture fusion and patency.
However, the interaction between osteoblast and the underlying dura mater has not been well
characterized, especially in humans. By combining dura mater cells and osteoblasts from
humans (using a co-culture model) we hope to be able to better evaluate osteoblast behavior.

25
We plan to evaluate important mediators, some pro-osteogenic (FGF2) and other anti-
osteogenic (Noggin and BMP3) that may be regulated by dura mater paracrine signalling or
may be imbalanced at the suture site affecting bone formation. Noggin and BMP3 have been
shown to downregulate ossification at the suture site in order to maintain suture patency
during the normal suture fusion process. Alternatively, it has been hypothesized that the
excessive bone formation at the fused suture site may be due a defect in differentiation,
function or both in fused suture osteoblasts, independent of humoral signalling from the
surrounding microenvironment. A potential candidate to explain these defects is the
transcriptional factor Runx2. Runx2 controls osteoblast differentiation and expression of
proteins such as osteocalcin responsible for bone forming function. As such we plan to
evaluate protein levels of Runx2 in fused and patent sutures.
The strength of this research is based on the establishment of a human model of
osteoblast cell culture harvested from patients affected by craniosynostosis. We also plan to
develop a model that combines human osteoblasts with human dura mater cells, establishing
a more physiological environment to evaluate osteoblast behavior. Using this model, we have
the possibility to surpass the normal suture fusion animal model and search in more detail the
mechanisms of craniosynostosis counting on a large number of human samples representing
the wide spectrum of this condition.
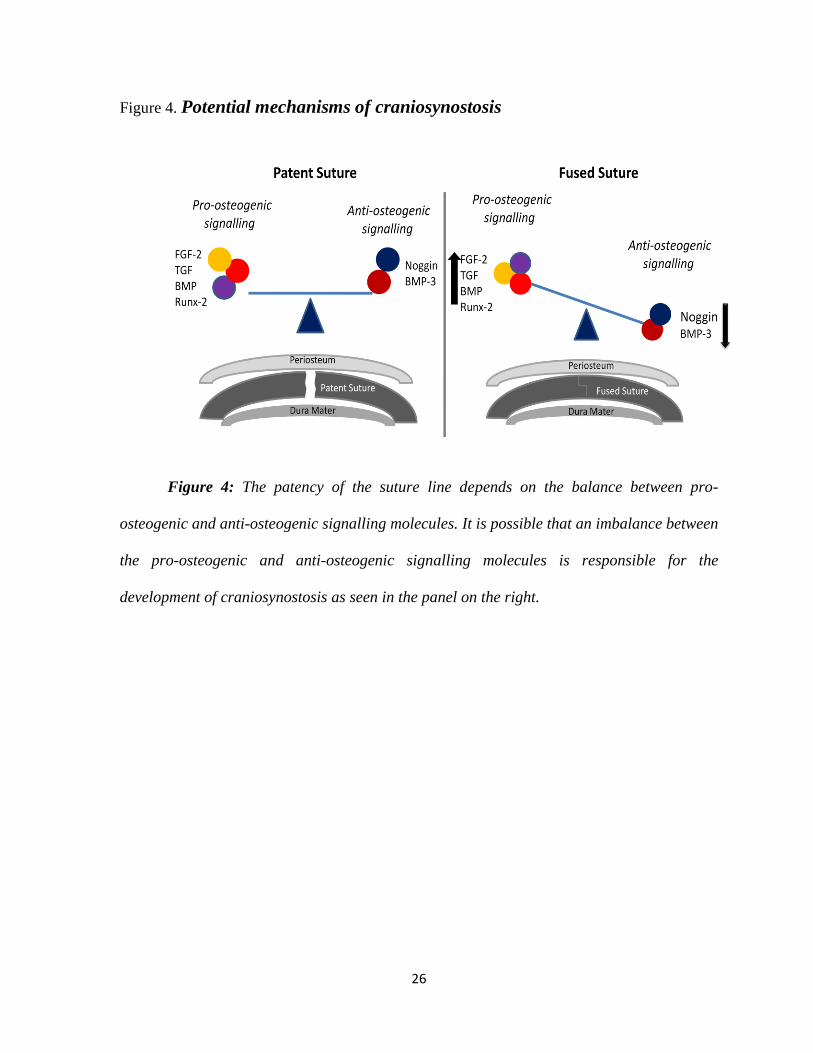
26
Figure 4. Potential mechanisms of craniosynostosis
Figure 4: The patency of the suture line depends on the balance between pro-
osteogenic and anti-osteogenic signalling molecules. It is possible that an imbalance between
the pro-osteogenic and anti-osteogenic signalling molecules is responsible for the
development of craniosynostosis as seen in the panel on the right.

27
Hypothesis
Regional variations in osteoblast function and cell signalling exist in calvaria of
infants with craniosynostosis.
SPECIFIC AIMS:
I) To develop a reliable osteoblast cell culture from calvaria of infants
undergoing surgery for craniosynostosis repair.
Rationale: The use of osteoblast cells derived from cranial bones of patients with
craniosynostosis may provide a valuable model for investigating the molecular and cellular
abnormalities associated with this disorder. In order to develop a valid technique for
osteoblast cell culture, the effects of differing media will be used to determine the optimal
growth conditions. To validate our bone cell culture model and demonstrate the presence of
osteoblasts, we will assess cellular proliferation, Runx2 expression, alkaline phosphatase,
osteocalcin, collagen I expression and mineralization. To further confirm our findings of in
vitro bone formation, ultrastructural analysis of the samples will be performed by
Transmission Electron Microscopy.
II) To assess regional variations in osteoblast behavior with and without dura
mater cells in co-culture.
Rationale: Although variations in osteoblast activity have been shown in murine models of
normal suture fusion, no studies have been performed using human osteoblasts and dura
mater cells in vitro from infants with craniosynostosis. Osteoblast behavior will be studied in

28
osteogenic monocultures and co-cultures with dura mater cells by MTT (proliferation rates),
immunocytochemistry and Western Blot for Runx2 expression, alkaline phosphatase assay
(differentiation), mineralization assay, and Transmission Electron Microscopy (TEM).
Quantitative Real Time – Polymerase Chain Reaction (qRT-PCR) will be performed to
evaluate the DNA expression of alkaline phosphatase and osteocalcin to confirm the cellular
level of differentiation. It is anticipated that these experiments will demonstrate regional
variations in osteoblast proliferation and differentiation which will provide the basis for aim
III.
III) To investigate the role of anti-osteogenic signalling on human osteoblasts in
vitro with and without dura mater cells in co-culture.
Rationale: It has been demonstrated that Noggin and BMP3 are important signalling
molecules in models of normal suture fusion but their role in craniosynostosis is unknown.
Noggin and BMP3 both down-regulate bone formation around the suture promoting suture
patency. These experiments (Western blot and immunohistochemistry) will investigate the
expression of these molecules in patent and fused sutures in infants with craniosynostosis in
order to determine if regional variations in expression exist. This findings may have
significance with respect to understanding the pathophysiology of craniosynostosis in
humans.
IV) To investigate the role of dura mater paracrine signalling in the
pathophysiology of craniosynostosis in humans.
Rationale: Dura mater underlying the cranial suture complex is one of several sources of
FGF2 and TGF-β1cytokines in vivo. However, it remains unclear whether growth factors

29
produced by dura mater are capable of down-regulating BMP3 and Noggin expression in
osteoblasts and up-regulate Runx2 expression thereby influencing osteoblast behavior. The
aim of these experiments is to determine if dura mater cells in co-culture influence
osteoblasts through paracrine signalling through secretion of FGF2 and TGF-β cytokines.
V) To assess the effects of exogenous administration of FGF2 on osteoblast
function in vitro.
Rationale: It has been demonstrated that FGF2 signalling is of central importance for
premature cranial suture fusion and might be an important paracrine signalling factor from
underlying dura mater to overlying calvarial osteoblasts. These experiments will investigate
the effects of exogenous administration of FGF2 to our cell culture and compare with the
effects produced by dura mater cells in a co-culture model.

30
Material and Methods

31
Tissue Sampling:
Patient Population: Patients sequentially selected with syndromic and non-syndromic
craniosynostosis (3 months - 3 years old) scheduled to undergo elective cranial vault
reshaping for craniosynostosis at The Hospital for Sick Children between July 2008 and
September 2010 were enrolled in this study. Informed consent was obtained. This research
received REB approval. Patients with previous cranial vault surgery were excluded from the
study.
Bone samples: During surgery, bone samples (5x5mm) and periosteum (1x1cm) were
obtained from fused and patent cranial sutures and non-suture bone, during the normal course
of the procedure. The sex, age, side, suture site and type of craniosynostosis was to be
registered. Tissue samples from fused suture, patent suture and adjacent non sutural bone
were processed for routine histology to confirm the origin of each sample, confirming the
patency or fusion of the suture line, and collagen I staining (See Appendix pg.120)60
.
Dura Mater: Samples of dura mater (patients between 3 to 17 years old) were obtained
from patients undergoing surgery for epilepsy and used to develop cell co-culture models.
Aim I) To develop a reliable osteoblast cell culture from infants undergoing
surgery for craniosynostosis repair.
Bone and periosteal tissue samples were taken from the operating room,
transported in ice in 50ml tubes containing αMEM and 5x penicillin-streptomycin and
processed immediately for cell culture. Samples were sequentially digested in a collagenase
(Sigma/Aldrich C0130) mixture at 37º C for 20 min and centrifuged at 700 rpm for 8
minutes. The pellet was resuspended in αMEM (Wisent Bio-Cat# 310010) containing 10-7
M

32
dexamethasone (Sigma/Aldrich Cat# D8893) and 15% FBS and plated in 75cm2 tissue
culture flasks (SARSTEDT – Cat#83.1813.002) (See Appendix pg.111). At subconfluence,
cultures were trypsinized (0.05% Trypsin-EDTA-Wisent Bio Cat# 325042) and seeded into
the tissue culture plates – 24 wells/5000 cells per well and 96 wells/1000 cells per well
(SARSTEDT – cat# 83.1835) for analysis (See Appendix pg.112). Medium changes were
done every 2-3 days. At 1 week after subculture, the medium was additionally supplemented
with 1mM ß-glycophosphate (Sigma/Aldrich cat# G6251) and 50μg/ml Ascorbic acid
(Sigma/Aldrich Cat# A2218)64
. Identification of cultured cells was performed by phase
contrast microscopy.
In order to validate our bone cell culture model and demonstrate the presence of
osteoblasts, we assessed cellular proliferation, Runx2 expression, alkaline phosphatase
staining, osteocalcin and collagen I expression and mineralization. To further confirm our
findings of in vitro bone formation, ultrastructural analysis of the samples was performed by
TEM.
i) Proliferation Rates: To assess proliferation in cells derived from infants with
craniosynotosis, the standard MTT assay (Sigma – Ref. 5655) - (See Appendix
pg.116) was employed at days 1,3,5 and 7 following subculture. This assay
assesses mitochondrial dehydrogenase activity and can serve as an indirect
measure of cellular proliferation65
.
ii) Differentiation Rates: At the same time points, alkaline phosphatase activity
was analyzed using the standard ρ-nitrophenil phosphate assay (pNP, Sigma –
Ref. 104-0) - (See Appendix pg.117). The final alkaline phosphatase activity
was adjusted per protein content (µg) and time of assay incubation (h)64
. After

33
confluence in cell culture, imunnocytochemistry for Runx2 (abcan 54868) was
performed (See Appendix pg.120)58
.
iii) qRT-PCR: Osteoblast differentiation was also evaluated by gene expression
of Osteocalcin, which represents the latest marker of osteoblast differentiation
(Primers - Forward: GGCAGCGAGGTAGTGAAGAG and Reverse:
CTGGAGAGGAGCAGAACTGG) and Alkaline Phosphatase (Primers –
Forward: CGTGGCTAAGAATGTCATCATTGTT and Reverse:
TGGTGGAGCTGACCCTTGA) examined by real time PCR. HPRT was
used as the housekeeping gene (See Appendix pg.124)66
.
iv) IHC: Collagen I (Mouse monoclonal to Colagen I – abcan 6308) expression
was evaluated by Immunocytochemistry (See Appendix pg.120).
v) Mineralization assay: Subsequently, cells were grown for 21 and 28 days
following subculture and were analyzed for mineralized bone nodule
formation via Alizarin Red S assay (Sigma Ref. A5533) - (See Appendix
pg.119)64
.
vi) Transmission Electron Microscopy: Mineralized nodules grown on
coverslips were fixed in 2% glutaraldehyde and processed for Transmission
Electron Microscopy to confirm bone formation and structure (See Appendix
pg 119)64
.
The effects of differing media composition on cell culture was assessed in order to
optimize the culture settings for osteoblast growth and differentiation. Human osteogenic
cells from patients (n=7) with craniosynostosis were cultured in αMEM (Wisent Bio-Cat#
310010) containing 10-7
M dexamethasone (Sigma/Aldrich Cat# D8893), supplemented with

34
i) 1% FBS, ii) 10% FBS, iii) 15% FBS or iv) ascorbic acid and β-glycophosphate.
Experimental culture conditions were compared on the basis of active cell growth (MTT
reduction assay) and differentiation (AP assay).
Aim II) To assess regional variations in osteoblast behavior with and without
dura mater cells in co-culture.
Osteoblasts obtained from regions of fused suture, patent suture and adjacent non
sutural bone and periosteum were cultured in αMEM containing 10-7
M dexamethasone,
supplemented with 15% FBS for 7 days and then supplemented with ascorbic acid 50µg/1ml
of medium and 1% β-glycophosphate. Proliferation rates, differentiation, including alkaline
phosphatase, collagen I and osteocalcin and mineralization were assessed in all 3 regions. In
order to determine if dura mater cells exerted any influence on osteoblast behavior, co-
cultures were established with dura mater samples from neurosurgical patients and
proliferation rates, differentiation and mineralization were assessed. Results of proliferation
were also stratified in syndromic and non-syndromic patients.
Dura mater sample was taken from the operating room, transported in ice in 14ml
tube containing αMEM and 5x penicillin-streptomycin and processed immediately for cell
culture. Samples were sequentially digested in a collagenase (Sigma/Aldrich C0130) mixture
at 37º C for 20 min and centrifuged at 700 rpm for 8 minutes. The pellet was resuspended in
αMEM (Wisent Bio-Cat# 310010) containing 10-7
M dexamethasone (Sigma/Aldrich Cat#
D8893) and 1% FBS and plated in 25cm2 tissue culture flasks (SARSTEDT –
Cat#83.1813.002) (See Appendix pg.111)44
. At subconfluence, cultures were trypsinized

35
(0.05% Trypsin-EDTA-Wisent Bio Cat# 325042) and seeded into the tissue culture plates – 6
wells/50000 for analysis (See Appendix pg.112). Medium changes were done every 2-3 days.
i) Co-Culture Model: We plated 5.0x104
first-passage osteoblasts per well in
six-well tissue culture plates (SARSTEDT Cat# 83.1839) and 2.0x104
dura
mater cells onto correspondenting co-culture filter inserts (VWR Cat# 62406-
163). The inserts have a pore size of 0.4µm. Cells are cultured separately in
standard medium until both cell populations were confluent. Dura mater cell-
seeded filter inserts are then combined with the six-well plates of osteoblasts
and cultured in αMEM containing 10-7
M dexamethasone, supplemented with
1% FBS for 10 days. The medium is changed every other day up to 10 days
when cells and medium were collected for Western blot analysis. Osteoblasts
cultured with empty co-culture inserts serve as a control67
.
Aim III) To investigate the role of anti-osteogenic signalling on human
osteoblasts in vitro with and without dura mater cells in co-culture.
Tissue samples from fused suture, patent suture and adjacent non sutural bone were
sent for immunohistochemical analysis of Noggin and BMP3 expression. These molecules
were investigated in tissue samples to obtain true representation of tissue expression of
Noggin/BMP3 in vivo without the influence of cell culture. Osteoblasts obtained from
regions of fused suture, patent suture and adjacent non sutural bone, and periosteum were
cultured as described in Aim II. In order to determine the influence of dura mater cells on
Noggin and BMP3 expression, a co-culture was established and medium from osteoblasts

36
alone or in co-culture was collected at day 10. Protein expression was measured by Western
Blot.
i) Investigation of Noggin and BMP3 expression by immunohistochemistry
Samples were demineralized with EDTA, fixed in 10% formalin, embedded in
paraffin, microtomized (5µm) and stained with rabbit polyclonal antibody to
Noggin (abcam) used in dilution 1:20 or rabbit polyclonal antibody to BMP3
(R&D System) used in dilution 1:5 to analyze their spatial expression patterns
in fused and patent sutures and non-suture bones. Detection was performed
with Goat Polymer (Biocaremedical) for BMP3 and ABC Ellite System
(Vector) for Noggin and Collagen I. 3,3’Diaminobenzidine (DAB) was used
as chromogen. Semi-quantitative analysis of the staining was carried out. The
staining intensity in the extracellular matrix (Noggin and BMP3) was
evaluated using semi-quantitative scoring system: no staining (0), low staining
(1), intermediate staining (2), and strong staining (3). The results were
evaluated by 2 independent investigators and averaged.
ii) Detection of Noggin and BMP3 by Western Blot : Western blot analysis for
Noggin (abcam 16054) and BMP3 (abcam71500) was carried out in
osteoblasts alone and in co-culture with dura mater cells (See Appendix
pg.121). β-actin was used as a positive control. Results were analyzed by
densitometry and expressed as protein density29
.

37
Aim IV) To investigate the role of dura mater paracrine signalling in the
pathophysiology of craniosynostosis in humans.
In order to search for potential growth factors responsible for the dura mater paracrine
signalling, dura mater cells were grown for 10 days and conditioned medium was collected at
different time points to determine the expression of FGF2 and TGF-β, both described in the
literature as the primary growth factors secreted by dura mater that may influence osteoblast
behavior.
i) Detection of FGF2 and TGF-β by Western blot : Conditioned Medium from
dura mater cells culture was collected at days 3,5 7 and 10 after subculture.
Western blot analysis was carried out for FGF2 (ab57059) and TGF-β (ab27969)
expression (See Appendix pg.121). β-actin was used as a positive control. Results
were analyzed by densitometry and expressed as protein density29
.
Aim V) To assess the effects of exogenous administration of FGF2 on osteoblast
function in vitro.
In Aim IV we examined the expression of endogenous FGF2 in dura-mater cells. The
aim in these experiments was to evaluate osteoblast behavior under stimulation by exogenous
human recombinant FGF2. Osteoblasts obtained from regions of fused suture, patent suture
and adjacent non-sutural bone were cultured alone or with increasing doses of human
recombinant FGF2. In order to evaluate osteoblast capacity to respond to stimulation,
proliferation and differentation rates were assessed.
i) Human Recombinant FGF2 Stimulation: Cells were plated onto 6 and 24-well
plates at a density of 50000 and 5000 cells/well respectively. After overnight
attachment, cells were treated with osteogenic differentiation media (αMEM

38
containing 10-7
M dexamethasone, supplemented with 15% FBS, ascorbic acid
50µg/1ml of medium and 1% β-glycophosphate) supplemented with human
recombinant FGF2 (5, 10, 50 and 100ng/ml)58
or only osteogenic media as a
control. Medium was changed every 2-3 days. FGF2 was added at each medium
change. MTT and Alkaline Phosphatase were performed at days 1, 3, 5 and 7.
Alkaline Phosphatase staining was performed at 1 week to assess early osteogenic
differentiation.
Statistical Analyses
Statistical analyses were performed using Graph Pad Prism and data are expressed as
mean ± standard deviation (SD). One-way analysis of variance (ANOVA) was applied for
comparison of sample groups in the MTT, Alkaline Phosphatase and Mineralization Assays,
Western Blot density, Immunohistochemistry density and qRT-PCR. Differences in values
between groups were evaluated using Tukey’s test. Two-way analysis of variance (ANOVA)
with repeated measurements was applied when more variables were evaluated for all three
groups. Significance was established at p < 0.05.
Research Ethics Board: This research has been approved by the Research Ethical Board at
The Hospital for Sick Children (1000013036). Consent was obtained from parents in all
cases.

39
Results

40
Demographics
Forty-five patients were enrolled in the study from July of 2008 to September of
2010. Twenty-eight were male and seventeen female. Mean age of the non-syndromic
patients was 7.5 ± 2.5 months (range from 3 months old to 13 months old). Forty patients had
single suture non-syndromic craniosynostosis. Samples were obtained from five syndromic
patients (3 Apert’s Syndrome, 1 Crouzon Syndrome and 1 patient with chromosome 7p
deletion). The sagittal suture was the most frequently involved followed by the metopic
suture (Table 2). All surgeries were performed uneventfully and there were no mortalities or
significant morbidities.
Cases n=45 Age
(months)
Male Female Sagittal Coronal Metopic Lambdoid
Syndromic
n=5
12.6±5.2 2 3 1 3 1 -
Non-
syndromic
n=40
7.5±2.5 26 14 19 9 11 1
Table 2: Demographics of patients with craniosynostosis enrolled in the study.
Histology
Histology was performed in tissue samples using hematoxylin-and-eosin staining to
confirm the tissue architecture of the cranial site from where the samples were harvested.
Histological evaluation of the patent, fused and adjacent bone confirmed the clinical
observations of presence or absence of a suture. Hematoxylin- and eosin-stained sections of
the control bone showed mature lamellar bone and hematopoietic marrow. Sections of patent
sutures showed fibrous connective tissue flanked on both sides by calvarial bony plates.

41
Sections of fused sutures showed an absence of the fibrous connective tissue zone, which
was replaced by lamellar bone. Some sites showed remnants of fibrous tissue in areas of
partial osseous obliteration and areas of bone remodeling (Figure 5).
Control Patent Suture Fused Suture
DAPI
H&E x100 magnification
H&E x50 magnification
LB
HM FT
FT
B
B
LB
HM
Figure 5: Histological sections of control bone, patent and fused suture bone with H&E
stain. The control bone showed lamellar bone (LB) and hematopoietic marrow (HM). The
patent suture showed fibrous connective tissue (FT) flanked on both sides by calvarial bony
plates (not seen) representing the normal patent suture. The fused suture showed the absence
of the fibrous connective tissue zone, replaced by bone (B).

42
Collagen I Expression
Collagen I is secreted by mature bone in the extracellular matrix. Collagen I
expression in tissue samples from non-sutural adjacent bone, patent and fused sutures was
assessed by IHC. Staining for Collagen I was positive for all three sites. The control bone
showed areas of mature bone. The patent suture showed an intense staining at the fibrous
connective tissue representing the normal patent suture. Stained cross-section of fused suture
showed intense collagen I staining, with areas of mature bone and remodeling (Figure 6).
Negative Control
100x
Fused Suture
Control Bone
Patent Suture
Figure 6: Immunohistochemistry staining for Collagen I. Stained sections show
positive expression of collagen I in fused bone, patent bone and adjacent non-sutural bone,
demonstrating formation of extracellular bone matrix (Magnification 100x).

43
Aim I) To develop a reliable osteoblast cell culture from calvarial of infants
undergoing surgery for craniosynostosis repair.
After culture of bone samples and periosteum, osteoblast growth was evaluated daily
by phase contrast microscopy up to confluence (usually 5 to 7 days after culture). Bone
samples and periosteum demonstrated the same potential for osteoblast retrival (4.5 x 106
versus 4.2 x 106) after 5 days. As such, osteoblasts from bone samples were preferred for all
experiments due to the in vivo relation between dura mater and calvarial bones.
Developing a valid bone cell culture model from calvaria of infants with craniosynostosis
In order to demonstrate the presence of osteoblasts in our cell culture model, we
assessed cellular proliferation, Runx2, alkaline phosphatase, collagen I and osteocalcin
expression and mineralization. To further confirm our findings of in vitro bone formation,
ultrastructural analysis of the samples was performed, demonstrating features of in vitro
osteogenesis, including mineral depositions and collagen fibrils in the extracellular matrix. A
ring pattern for electron diffraction studies was characteristic of normal hydroxyapatite
(Figure 7).
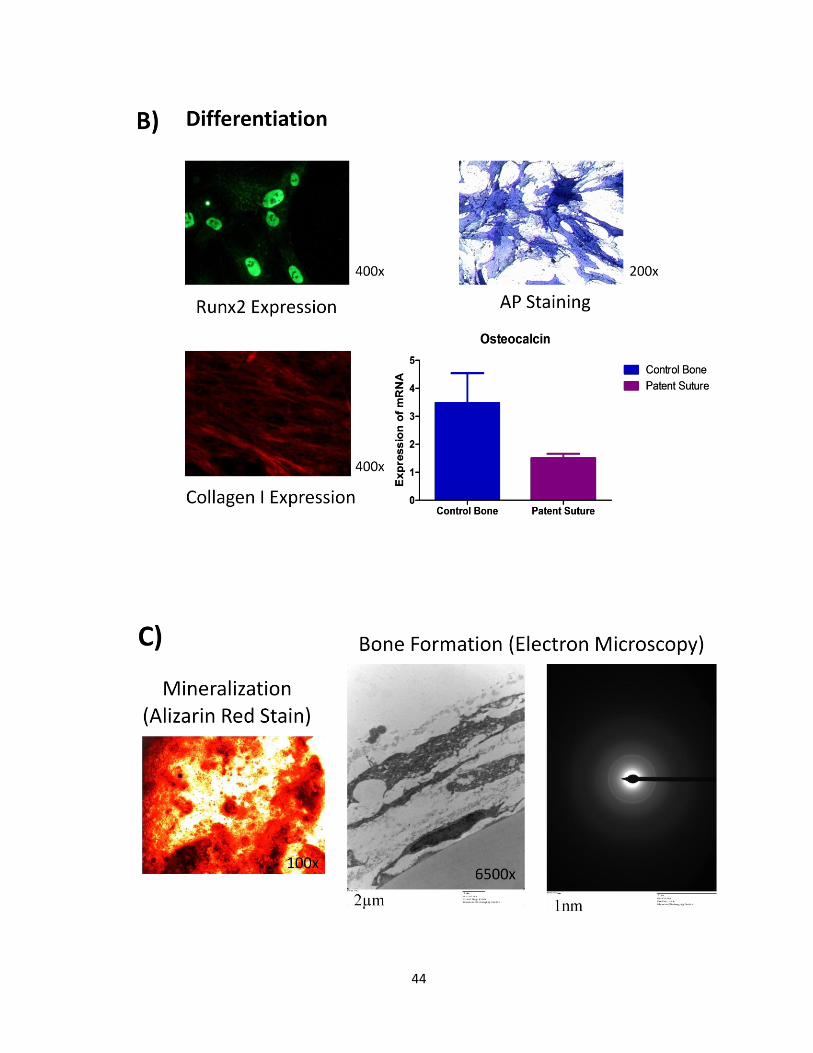
44

45
Figure 7: Evidences of osteoblasts in our cell culture model and in vitro bone
formation: Patent suture cells were able to proliferate in vitro (A) and differentiate in
osteoblasts as shown by the expression of differentiation markers such as alkaline
phosphatase, Runx2, collagen I and osteocalcin (B). Mineralization and Bone Formation was
achieved after 28 days in osteogenic conditions. Ultrastructural analysis of the samples was
performed, demonstrating features of in vitro osteogenesis, including mineral depositions
and collagen fibrils in the extracellular matrix. A ring pattern for electron diffraction studies
was characteristic of normal hydroxyapatite (C).
Effect of medium composition on cellular proliferation
Fused suture osteoblasts, independent of the experimental conditions or time-point,
demonstrated higher growth rates than the patent and control sutures. The addition of
ascorbic acid and β-glycerophosphate to the osteogenic medium resulted in significantly
higher growth rates for the fused suture osteoblasts when compared αMEM (1% FBS) and
αMEM (15% FBS) on day 3 (p<0.05) (Figure 8A). There was no significant differences in
proliferation rates at days 5,7 and 10 independent of medium composition.

46
Figure 8: MTT assays of human cranial suture-derived osteoblasts. All cultures in
1%, 10%, 15% FBS or 15% FBS + ascorbic acid (50µg)/1ml of medium. Values are
mean±SD; n=7, *p<0.05.
Differences in FBS concentration did not significantly affect the growth of cranial
suture-derived osteoblasts from fused suture, patent suture and adjacent bone. Moreover,
there was no significant difference in alkaline phosphatase activity independent of the
medium composition and therefore we chose for the model system the medium consisting of
αMEM containing 10-7
M dexamethasone, supplemented with 15% FBS for 7 days and then
supplemented with ascorbic acid (50µg)/1ml of medium and 1% β-glycerophosphate for the
experiments.
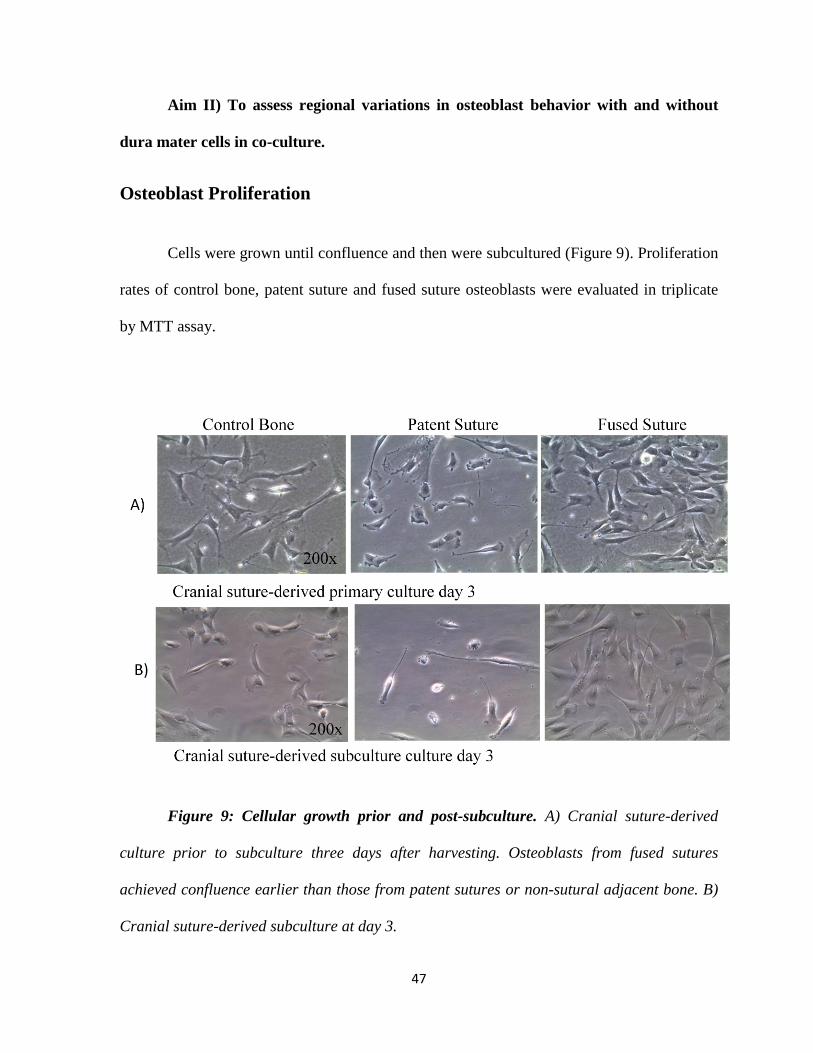
47
Aim II) To assess regional variations in osteoblast behavior with and without
dura mater cells in co-culture.
Osteoblast Proliferation
Cells were grown until confluence and then were subcultured (Figure 9). Proliferation
rates of control bone, patent suture and fused suture osteoblasts were evaluated in triplicate
by MTT assay.
Figure 9: Cellular growth prior and post-subculture. A) Cranial suture-derived
culture prior to subculture three days after harvesting. Osteoblasts from fused sutures
achieved confluence earlier than those from patent sutures or non-sutural adjacent bone. B)
Cranial suture-derived subculture at day 3.

48
The osteoblasts from the fused sutures exhibited a significant (p < 0.01) increased rate
of growth, compared with those derived from the control and patent suture at days 5 and 7
(Figure 10).
All Cases
Day 1 Day 3 Day 5 Day 70
5
10
15Control Bone
Patent Suture
Fused Suture
* *
* p<0.02n=33 per group
*
OD
/1000 c
ells
Figure 10: Osteoblast proliferation rates (MTT): Fused suture cells showed a
significantly higher rate of proliferation at time-point 3 (p<0.05) when compared to control
bone and time-points 5 (p<0.001) and 7 (p<0.001) when compared with control and patent
suture osteoblasts at the same time-points.
Proliferation rates were significantly lower for syndromic cases in all three groups
when compared with non-syndromic patients (Figure 11). For this reason we decided to
exclude samples from syndromic patients for the subsequent experiments in order to not
confound the findings.
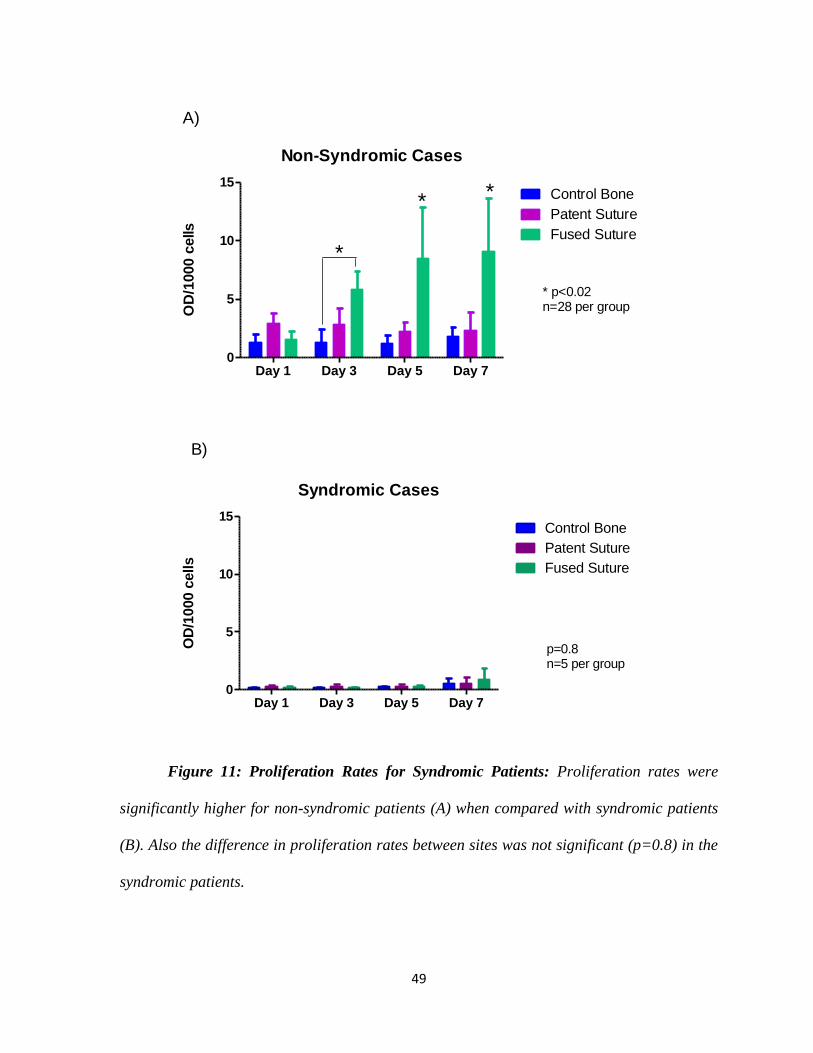
49
Non-Syndromic Cases
Day 1 Day 3 Day 5 Day 70
5
10
15Control Bone
Patent Suture
Fused Suture
* *
* p<0.02n=28 per group
A)
*
OD
/1000 c
ells
Syndromic Cases
Day 1 Day 3 Day 5 Day 70
5
10
15Control Bone
Patent Suture
Fused Suture
p=0.8n=5 per group
B)
OD
/1000 c
ells
Figure 11: Proliferation Rates for Syndromic Patients: Proliferation rates were
significantly higher for non-syndromic patients (A) when compared with syndromic patients
(B). Also the difference in proliferation rates between sites was not significant (p=0.8) in the
syndromic patients.

50
Co-Culture of Osteoblasts and Dura Mater Cells: Proliferation Rates
Osteoblasts from control bone, fused suture and patent suture co-cultured with dura
mater were compared with their counterparts without dura mater cells (Figure 12). Adding
dural cells to the osteoblast culture does not change significantly the proliferation rates for
fused suture and control bone. However, patent suture osteoblasts in co-culture with dura
demonstrated a significantly greater (p=0.001) proliferation rate at day 7 when compared
with patent suture osteoblasts without dura.
Patent Suture
Day 1 Day 3 Day 5 Day 70
5
10
15
Subculture with Dura
Subculture without Dura
p=0.001 (two-way ANOVA)n= 28 each group without duran= 6 each group with dura
B)
OD
/1000 c
ells
*
Control Bone
Day 1 Day 3 Day 5 Day 70
5
10
15Subculture without Dura
Subculture with Dura
p=0.27 (two-way ANOVA)n= 28 each group without duran= 6 each group with dura
A)
OD
/1000 c
ells
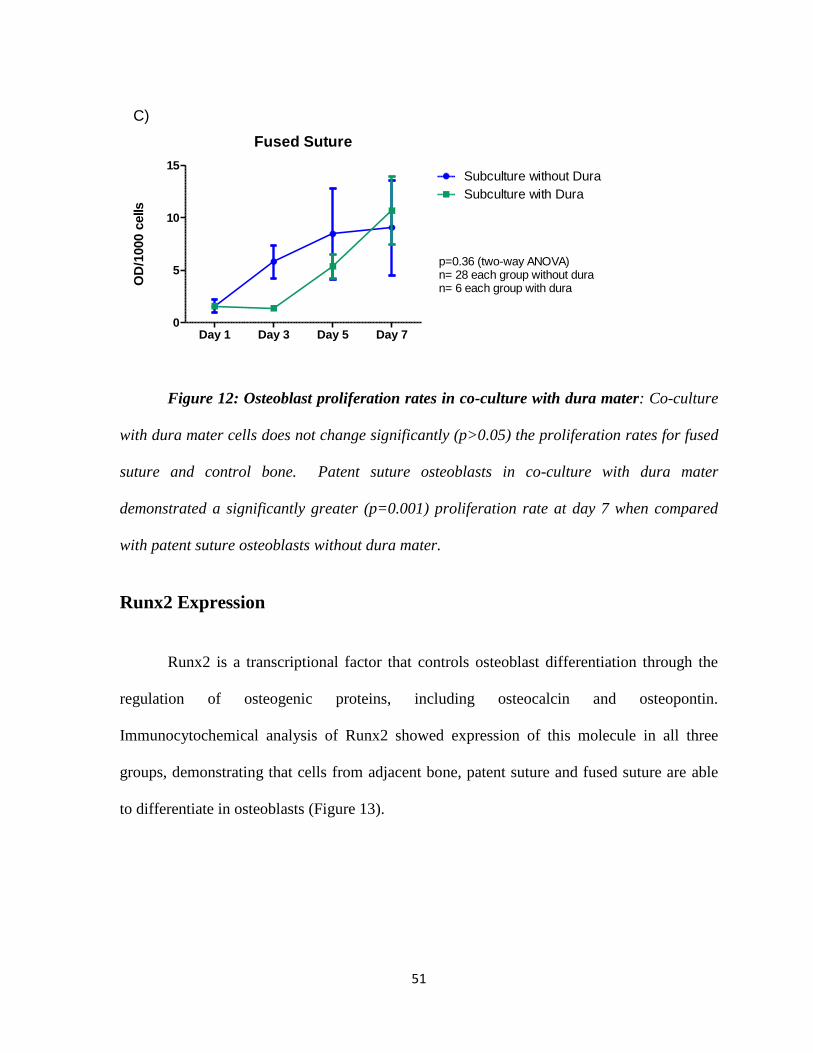
51
Fused Suture
Day 1 Day 3 Day 5 Day 70
5
10
15Subculture without Dura
Subculture with Dura
C)
p=0.36 (two-way ANOVA)n= 28 each group without duran= 6 each group with dura
OD
/1000 c
ells
Figure 12: Osteoblast proliferation rates in co-culture with dura mater: Co-culture
with dura mater cells does not change significantly (p>0.05) the proliferation rates for fused
suture and control bone. Patent suture osteoblasts in co-culture with dura mater
demonstrated a significantly greater (p=0.001) proliferation rate at day 7 when compared
with patent suture osteoblasts without dura mater.
Runx2 Expression
Runx2 is a transcriptional factor that controls osteoblast differentiation through the
regulation of osteogenic proteins, including osteocalcin and osteopontin.
Immunocytochemical analysis of Runx2 showed expression of this molecule in all three
groups, demonstrating that cells from adjacent bone, patent suture and fused suture are able
to differentiate in osteoblasts (Figure 13).

52
Negative Control Control Bone Patent Suture Fused Suture
400x
400x
Dapi
Runx-2
Figure 13: Expression of Runx2 demonstrated by Immunocytochemistry (nuclear
staining). Control bone, patent suture and fused suture cells were similarly positive for
Runx2, indicating that cells from all three groups were able to differentiate.
However, Western blot analysis of Runx2 showed an up regulation of this molecule
in the control group and fused suture group with and without dura mater when compared
with patent suture. This difference was significantly (p<0.05) greater when osteoblasts
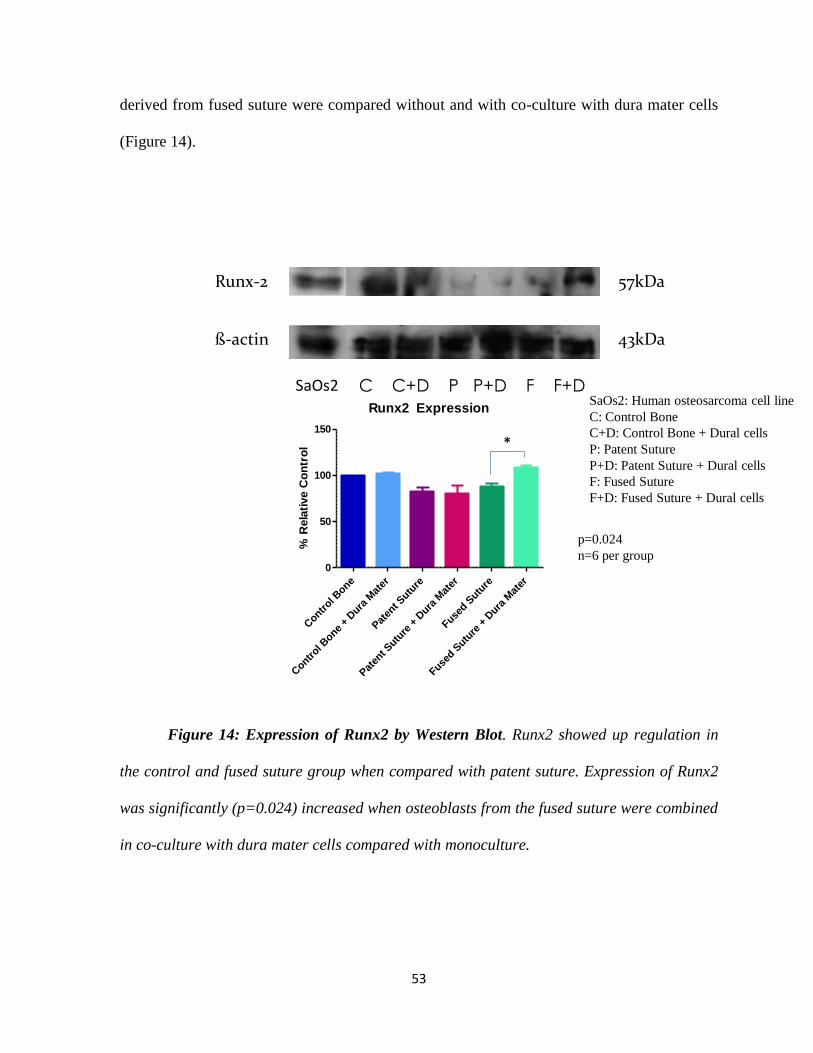
53
derived from fused suture were compared without and with co-culture with dura mater cells
(Figure 14).
57kDaRunx-2
SaOs2 C C+D P P+D F F+D
ß-actin 43kDa
Runx2 Expression
Contr
ol Bone
Contr
ol Bone
+ Dura
Mat
er
Pat
ent S
uture
Pat
ent S
uture
+ D
ura M
ater
Fused S
uture
Fused S
uture
+ D
ura M
ater
0
50
100
150
% R
ela
tive C
on
tro
l
p=0.024
n=6 per group
*
SaOs2: Human osteosarcoma cell line
C: Control Bone
C+D: Control Bone + Dural cells
P: Patent Suture
P+D: Patent Suture + Dural cells
F: Fused Suture
F+D: Fused Suture + Dural cells
Figure 14: Expression of Runx2 by Western Blot. Runx2 showed up regulation in
the control and fused suture group when compared with patent suture. Expression of Runx2
was significantly (p=0.024) increased when osteoblasts from the fused suture were combined
in co-culture with dura mater cells compared with monoculture.
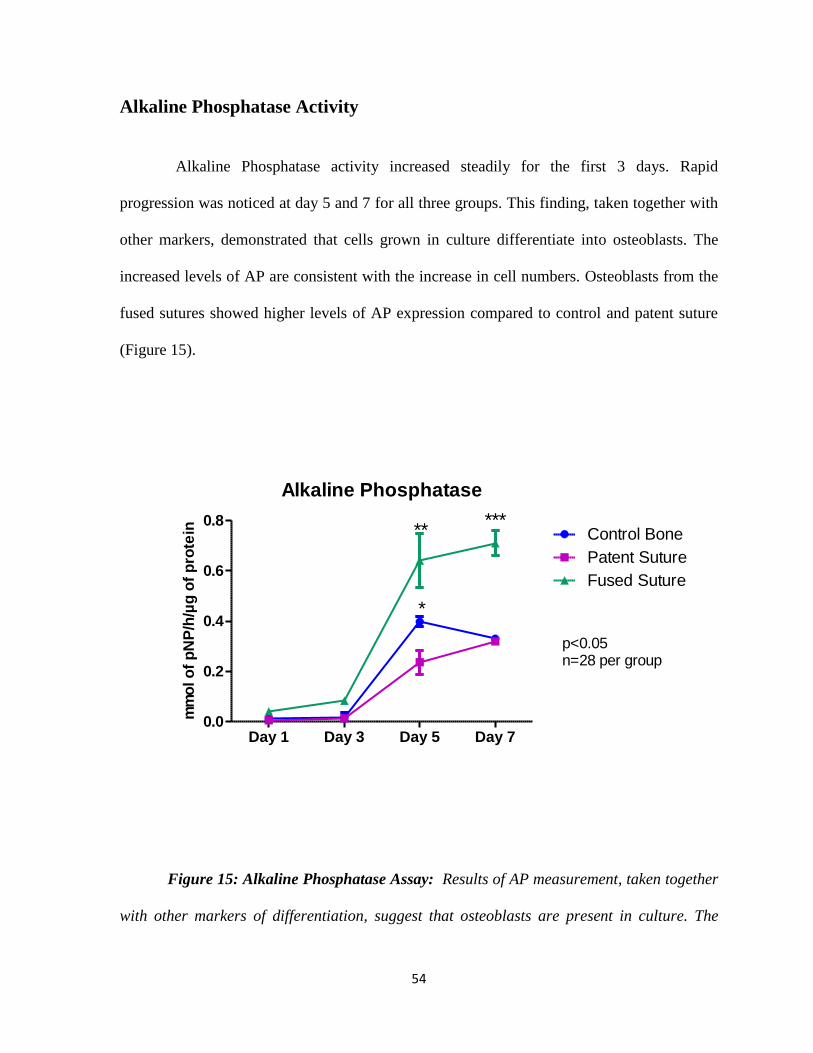
54
Alkaline Phosphatase Activity
Alkaline Phosphatase activity increased steadily for the first 3 days. Rapid
progression was noticed at day 5 and 7 for all three groups. This finding, taken together with
other markers, demonstrated that cells grown in culture differentiate into osteoblasts. The
increased levels of AP are consistent with the increase in cell numbers. Osteoblasts from the
fused sutures showed higher levels of AP expression compared to control and patent suture
(Figure 15).
Alkaline Phosphatase
Day 1 Day 3 Day 5 Day 70.0
0.2
0.4
0.6
0.8Control Bone
Patent Suture
Fused Suture
p<0.05n=28 per group
*
*****
mm
ol o
f p
NP
/h/µ
g o
f p
rote
in
Figure 15: Alkaline Phosphatase Assay: Results of AP measurement, taken together
with other markers of differentiation, suggest that osteoblasts are present in culture. The

55
increased levels of AP are consistent with the increased number of cells at days 5 and 7.
(*p<0.05 control versus patent suture;
** p<0.05 fused suture versus patent suture and
***p<0.05 fused suture versus control and patent bone). Final AP concentration is expressed
as mmol of pNP per hour per µg of protein.
Figure 16: Alkaline Phosphatase Activity (Fast Blue BB salt - dark blue staining)
in cell cultures: Osteoblasts derived from the fused suture demonstrated multiple layers and
an increased number of cells at day 7.

56
qRT-PCR
AP Osteocalcin0
2
4
6Control Bone
Patent Suture
Fused Suture
Exp
ressio
n o
f m
RN
A
p=0.66 (AP)
p=0.11 (Osteocalcin)
n=6 per group
Figure 17: Analysis by qRT-PCR of alkaline phosphatase and osteocalcin gene
expression: Expression of alkaline phosphatase and osteocalcin mRNA are not significantly
different between osteoblasts from various regions of the infant calvarium. mRNA levels
were measured by qRT-PCR and normalized to HPRT mRNA.
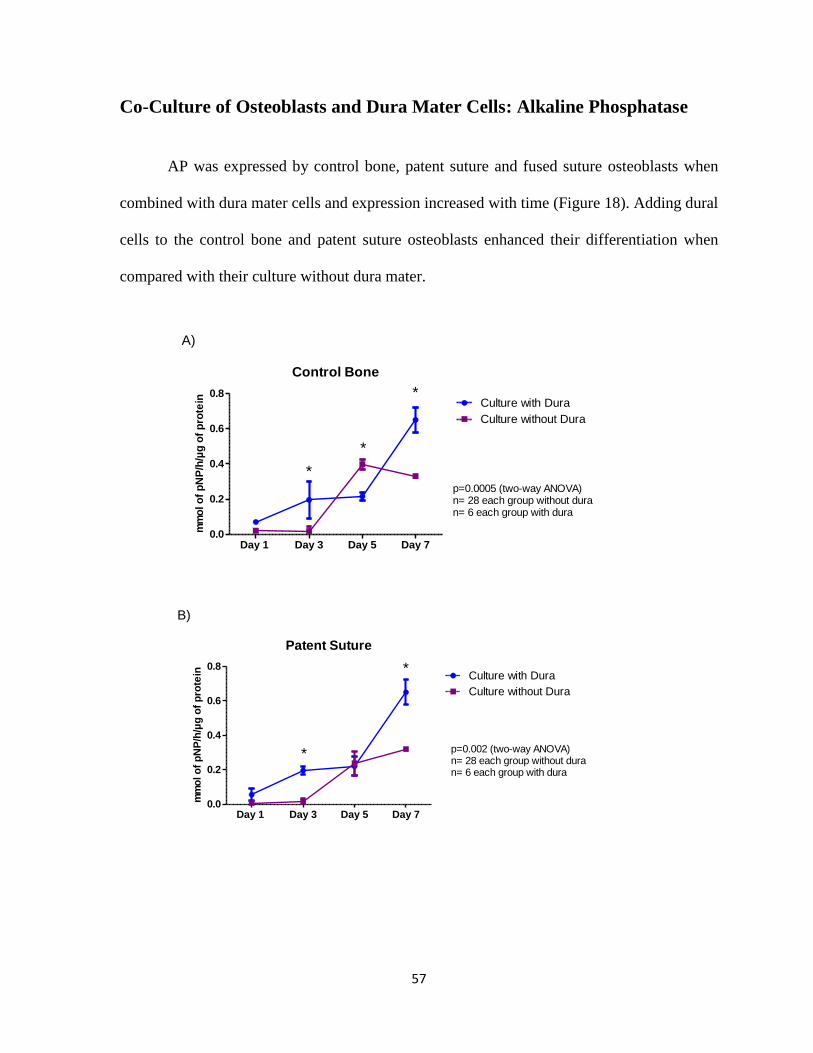
57
Co-Culture of Osteoblasts and Dura Mater Cells: Alkaline Phosphatase
AP was expressed by control bone, patent suture and fused suture osteoblasts when
combined with dura mater cells and expression increased with time (Figure 18). Adding dural
cells to the control bone and patent suture osteoblasts enhanced their differentiation when
compared with their culture without dura mater.
Control Bone
Day 1 Day 3 Day 5 Day 70.0
0.2
0.4
0.6
0.8Culture with Dura
Culture without Dura
p=0.0005 (two-way ANOVA)n= 28 each group without duran= 6 each group with dura
A)
*
*
mm
ol o
f p
NP
/h/µ
g o
f p
rote
in
*
Patent Suture
Day 1 Day 3 Day 5 Day 70.0
0.2
0.4
0.6
0.8Culture with Dura
Culture without Dura
p=0.002 (two-way ANOVA)n= 28 each group without duran= 6 each group with dura
B)
*
*
mm
ol o
f p
NP
/h/µ
g o
f p
rote
in

58
Fused Suture
Day 1 Day 3 Day 5 Day 70.0
0.2
0.4
0.6
0.8Culture with Dura
Culture without Dura
p<0.04 (two-way ANOVA)n= 28 each group without duran= 6 each group with dura
C)
mm
ol o
f p
NP
/h/µ
g o
f p
rote
in
Figure 18: AP in osteoblasts co-culture with dura mater cells: Osteoblasts co-
cultured with dura mater cells demonstrate increased AP expression over time. Co-culture
with dura mater cells significantly enhanced control bone (A) differentiation at days 3,5 and
7 and patent suture osteoblasts (B) differentiation at days 3 and 7 compared with osteoblasts
without dura mater and reduced differentiation of fused suture osteoblasts (C). Final AP
concentration is expressed as mmol of pNP per hour per µg of protein.
Mineralization
In cultures containing β-glycerophosphate (10mM) and ascorbic acid (50µg/ml), the
secretion of extracellular matrix leads to progressive mineralization and eventual bone
nodule formation by day 14 (Figure 20). Osteoblasts from all three sites were capable of
forming bone nodules in vitro, as evidenced by extensive Alizarin Red Staining. However, at
28 days osteoblasts from fused sutures demonstrated significantly (p<0.05) greater
mineralization compared to osteoblasts from patent sutures (Figure 19). Bone nodules
were detected earlier in osteoblasts from fused sutures (11±3 days) compared with control
(13±4 days) and patent sutures (16±4 days) osteoblasts (Figure 20). Formation of bone
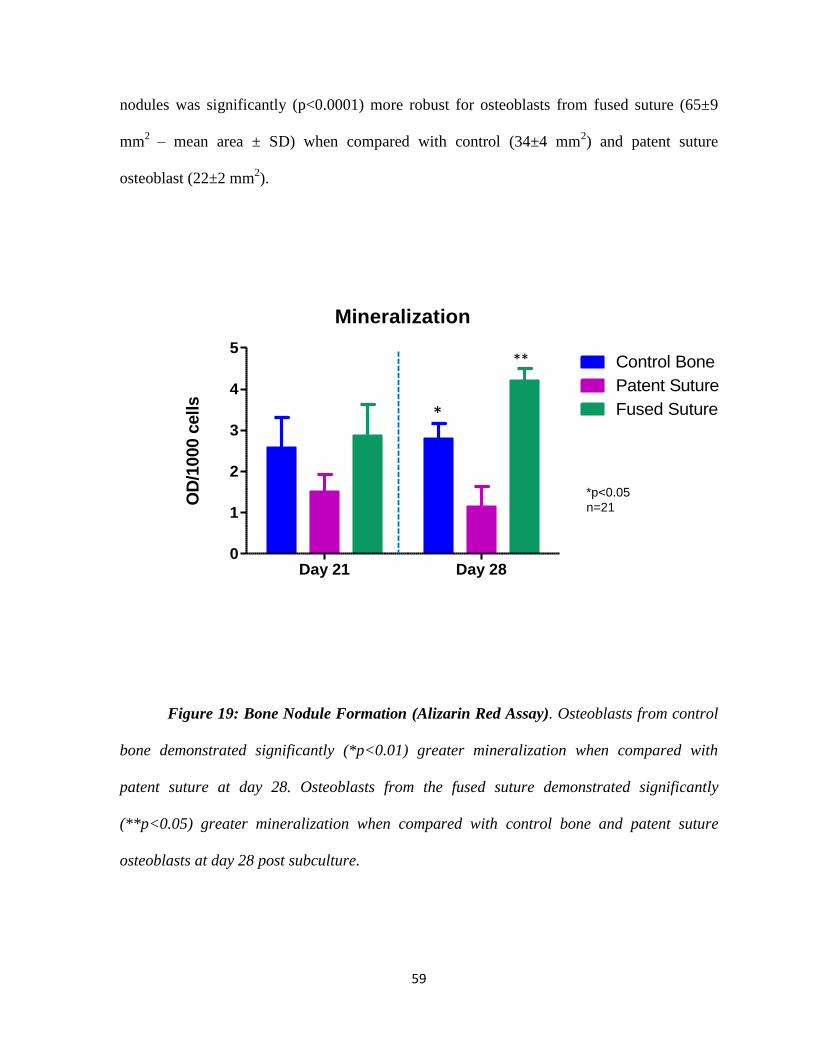
59
nodules was significantly (p<0.0001) more robust for osteoblasts from fused suture (65±9
mm2 – mean area ± SD) when compared with control (34±4 mm
2) and patent suture
osteoblast (22±2 mm2).
Mineralization
Day 21 Day 280
1
2
3
4
5Control Bone
Patent Suture
Fused Suture
OD
/1000 c
ells
*
*p<0.05
n=21
**
Figure 19: Bone Nodule Formation (Alizarin Red Assay). Osteoblasts from control
bone demonstrated significantly (*p<0.01) greater mineralization when compared with
patent suture at day 28. Osteoblasts from the fused suture demonstrated significantly
(**p<0.05) greater mineralization when compared with control bone and patent suture
osteoblasts at day 28 post subculture.

60
Figure 20: Bone Nodule Formation at days 14 and 18 (Alizarin Red S Staining):
Formation of bone nodules in cultures derived from fused sutures was detected earlier (11±3
days) compared with that of control (13±4 days) and patent sutures (16±4 days). Arrow
shows bone nodule formation.

61
Control – 28 days Patent Suture – 28 days Fused Suture– 28 days
Control – 21 days Patent Suture – 21days Fused Suture– 21days
100x 100x 100x
100x 100x 100x
Figure 21: Mineralization at days 21 and 28 (Alizarin Red Staining). Formation of
bone nodules in cultures derived from fused sutures was significantly (p<0.001) more robust,
compared with that of normal sutures and control areas at day 28. Arrow shows bone nodule
formation.

62
Co-Culture of Osteoblasts and Dura Mater Cells: Mineralization
Mineralization rates in osteoblasts co-cultured with dura mater cells were not
different between groups. The addition of dural cells to culture resulted in qualitative
differences in all three groups with reduced nodule formation and mineralization (Figure 22).
Figure 22: Bone nodule formation in osteogenic cultures with and without dural co-
culture at day 28 (Alizarin Red S Staining). Mineralization in osteoblasts co-cultured with
dura mater cells was not different between groups.

63
Transmission Electron Microscopy
Electron microscopy of cell cultures of control bone, patent suture and fused sutures
revealed ultrastructural features of in vitro osteogenesis, including mineral depositions
and collagen fibrils in the extracellular matrix (Figure 23).
Figure 23: Transmission Electron Microscopy of Bone Nodule Formation: Sections
showing ultrastructural features of in vitro osteogenesis, including collagen fibrils,
mineral and osteoblasts. Cell cultures from control bone, patent suture and fused suture
were able to form bone.
Aim III) To investigate the role of anti-osteogenic signalling on human
osteoblasts in vitro with and without dura mater cells in co-culture.

64
Expression of anti-osteogenic signalling molecules
The anti-osteogenic molecules Noggin and BMP3 were analyzed in tissue samples
from control bone, patent suture and fused suture by IHC. The expression of Noggin and
BMP3 in the osteoblasts from patent suture was greater than expression levels in control
bone and fusing suture (Figure 24).
Figure 24: Immunohistochemistry analysis of Noggin in tissue samples. Noggin
expression (expressed as a brown pigment in the extracellular matrix) was found to be
greater in the osteoblasts from patent suture when compared to control bone or fused suture.
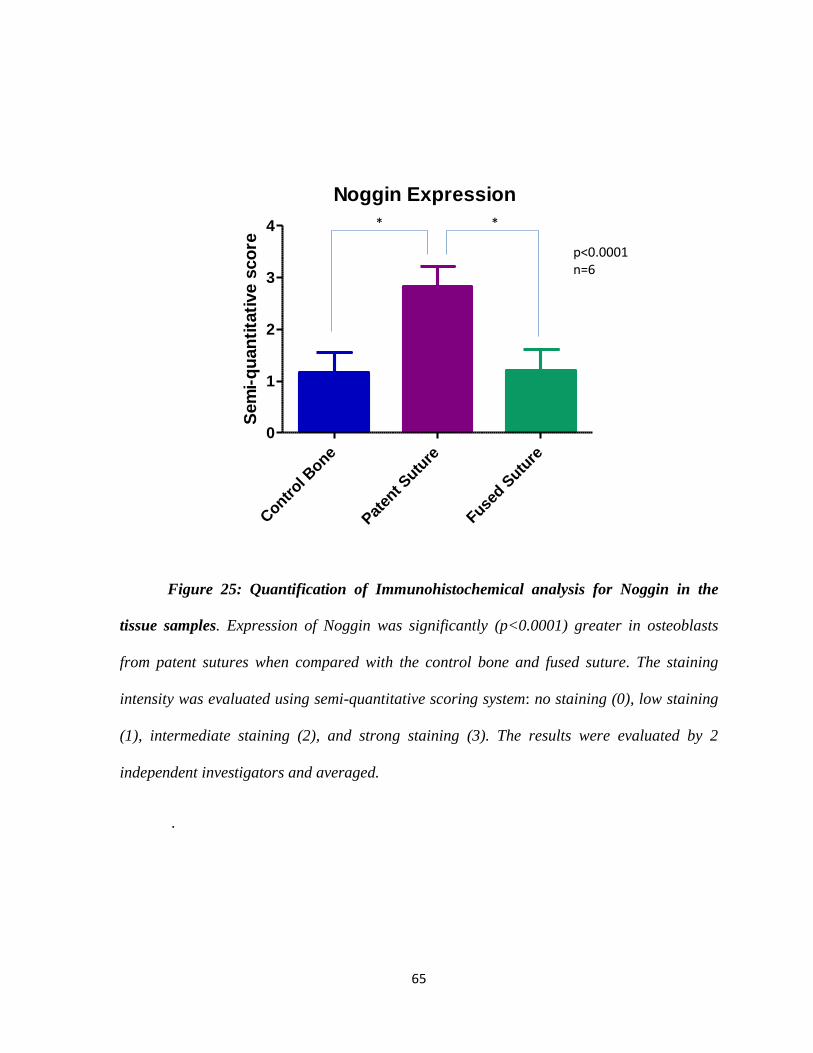
65
Noggin Expression
Contr
ol Bone
Pat
ent S
uture
Fused S
uture
0
1
2
3
4
Sem
i-q
uan
tita
tive s
co
re p<0.0001n=6
* *
Figure 25: Quantification of Immunohistochemical analysis for Noggin in the
tissue samples. Expression of Noggin was significantly (p<0.0001) greater in osteoblasts
from patent sutures when compared with the control bone and fused suture. The staining
intensity was evaluated using semi-quantitative scoring system: no staining (0), low staining
(1), intermediate staining (2), and strong staining (3). The results were evaluated by 2
independent investigators and averaged.
.
[Type a quote from the document or the summary of an interesting point. You can position the
text box anywhere in the document. Use the Text Box Tools tab to change the formatting of the
pull quote text box.]
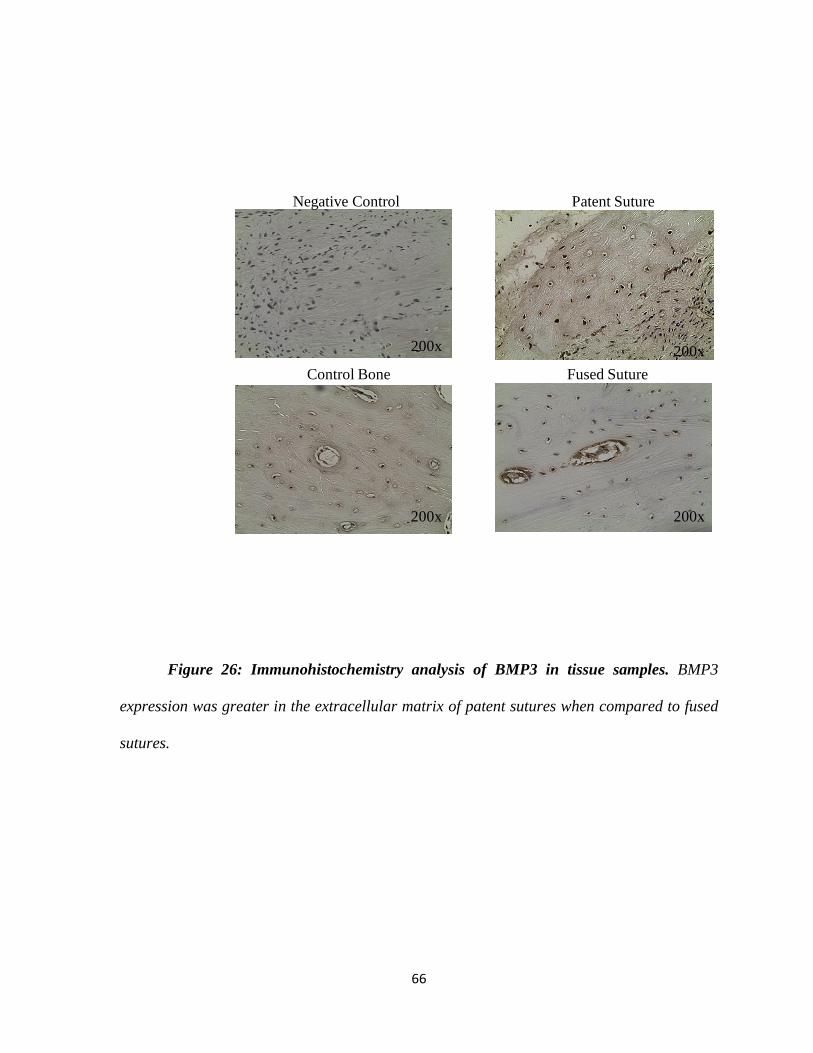
66
Negative Control Patent Suture
Control Bone Fused Suture
200x 200x
200x 200x
Figure 26: Immunohistochemistry analysis of BMP3 in tissue samples. BMP3
expression was greater in the extracellular matrix of patent sutures when compared to fused
sutures.
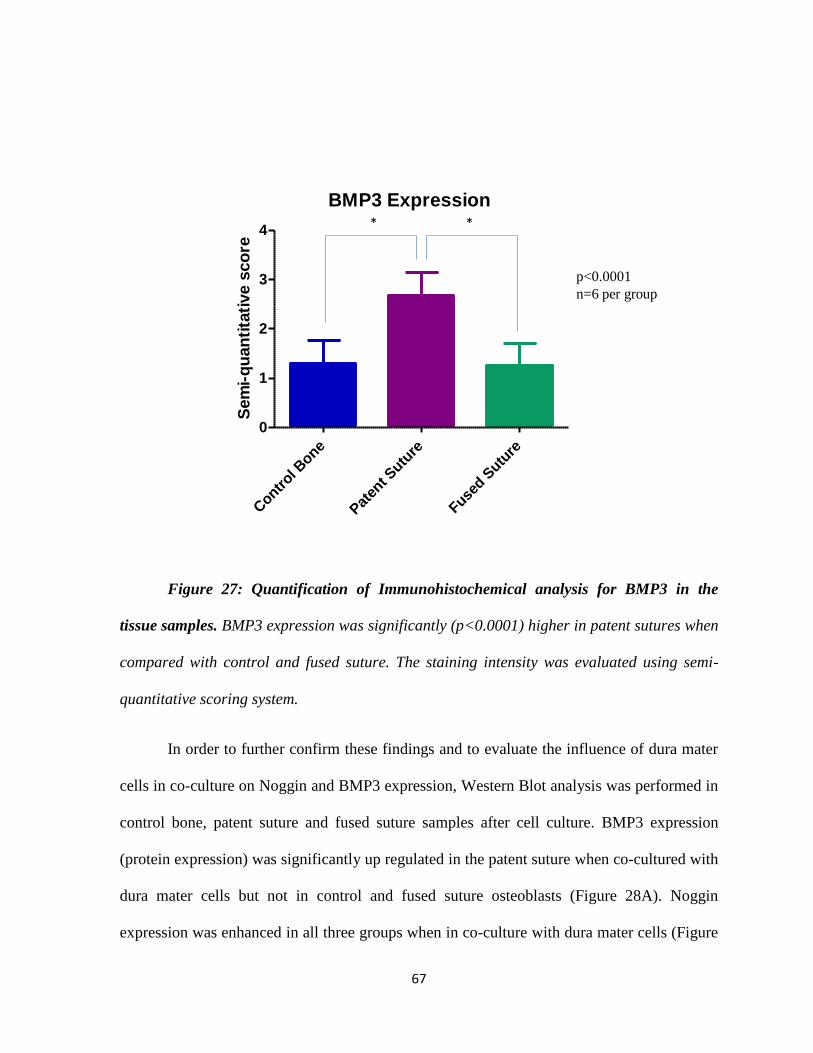
67
BMP3 Expression
Contr
ol Bone
Pat
ent S
uture
Fused S
uture
0
1
2
3
4S
em
i-q
uan
tita
tive s
co
re
p<0.0001
n=6 per group
**
Figure 27: Quantification of Immunohistochemical analysis for BMP3 in the
tissue samples. BMP3 expression was significantly (p<0.0001) higher in patent sutures when
compared with control and fused suture. The staining intensity was evaluated using semi-
quantitative scoring system.
In order to further confirm these findings and to evaluate the influence of dura mater
cells in co-culture on Noggin and BMP3 expression, Western Blot analysis was performed in
control bone, patent suture and fused suture samples after cell culture. BMP3 expression
(protein expression) was significantly up regulated in the patent suture when co-cultured with
dura mater cells but not in control and fused suture osteoblasts (Figure 28A). Noggin
expression was enhanced in all three groups when in co-culture with dura mater cells (Figure
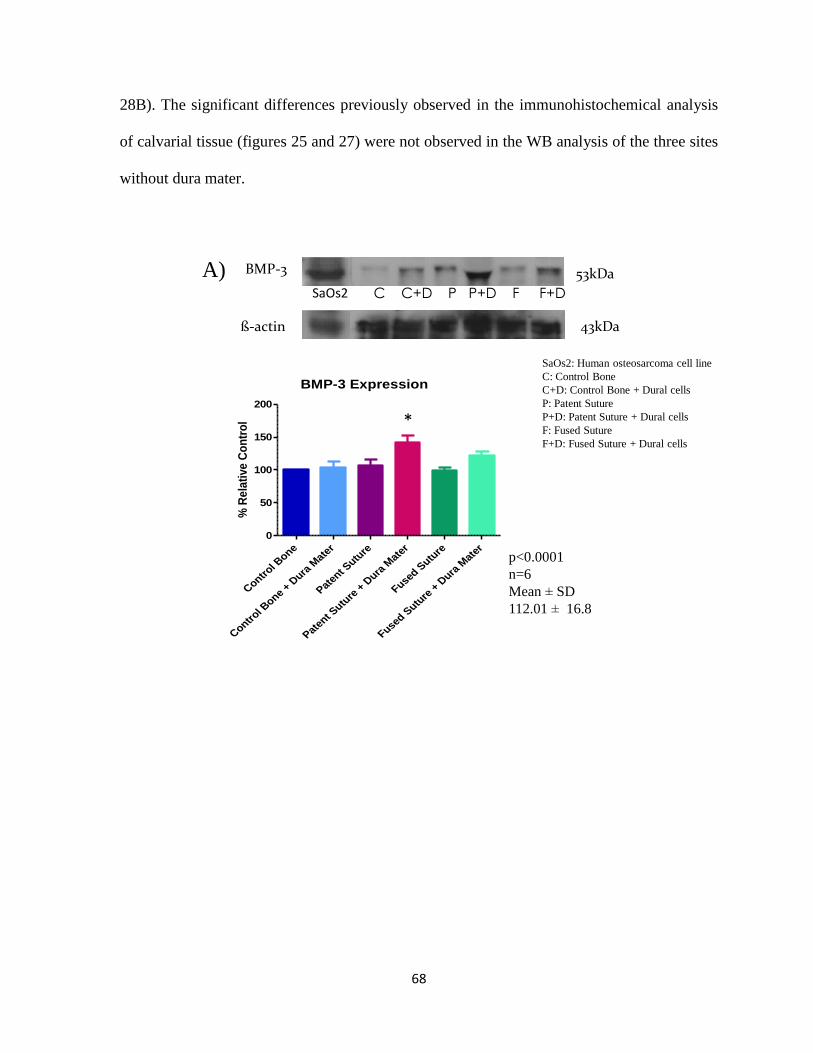
68
28B). The significant differences previously observed in the immunohistochemical analysis
of calvarial tissue (figures 25 and 27) were not observed in the WB analysis of the three sites
without dura mater.
BMP-3 Expression
Contr
ol Bone
Contr
ol Bone
+ Dura
Mate
r
Paten
t Sutu
re
Paten
t Sutu
re +
Dura
Mat
er
Fused S
uture
Fused S
uture
+ D
ura M
ater
0
50
100
150
200
% R
ela
tive
Co
ntr
ol
p<0.0001
n=6
Mean SD
112.01 16.8
*
A) BMP-3 53kDa
C C+D P P+D F F+D
ß-actin 43kDa
SaOs2
SaOs2: Human osteosarcoma cell line
C: Control Bone
C+D: Control Bone + Dural cells
P: Patent Suture
P+D: Patent Suture + Dural cells
F: Fused Suture
F+D: Fused Suture + Dural cells
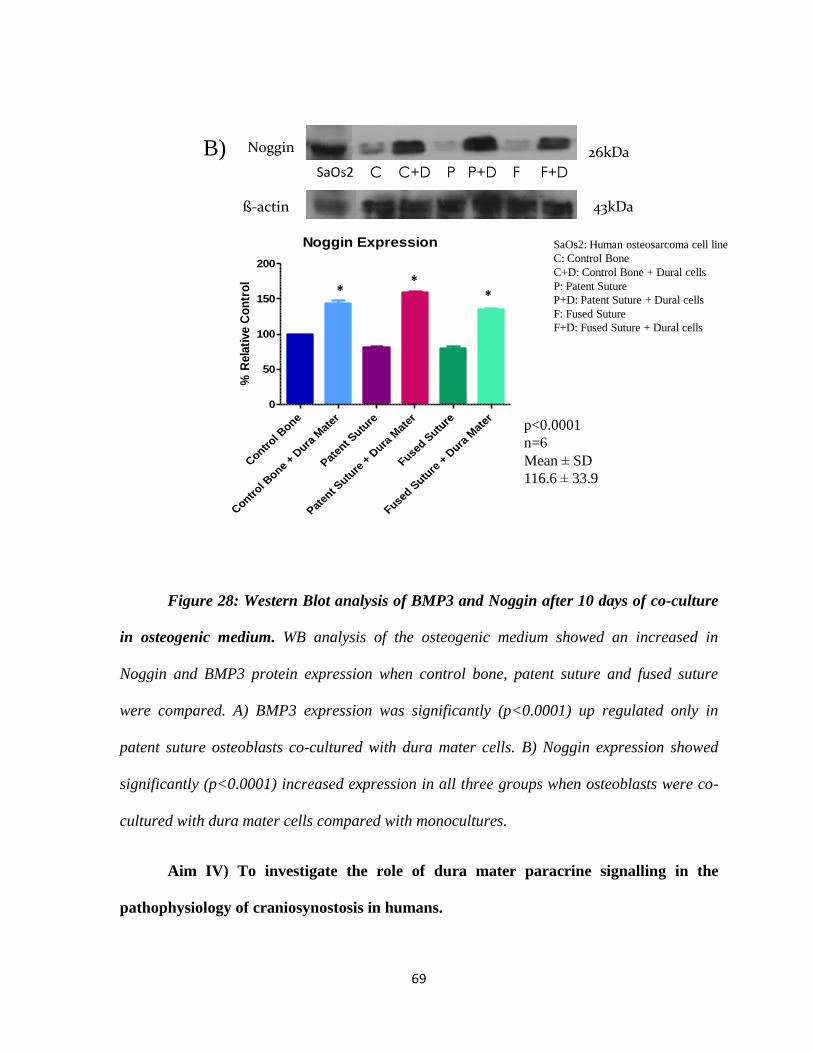
69
B)C C+D P P+D F F+D
ß-actin 43kDa
SaOs2
SaOs2: Human osteosarcoma cell line
C: Control Bone
C+D: Control Bone + Dural cells
P: Patent Suture
P+D: Patent Suture + Dural cells
F: Fused Suture
F+D: Fused Suture + Dural cells
Noggin 26kDa
Noggin Expression
Contr
ol Bone
Contr
ol Bone
+ Dura
Mat
er
Paten
t Sutu
re
Paten
t Sutu
re +
Dura
Mat
er
Fused S
uture
Fused S
uture
+ D
ura M
ater
0
50
100
150
200
% R
ela
tive C
on
tro
l
**
*
p<0.0001
n=6
Mean SD
116.6 33.9
Figure 28: Western Blot analysis of BMP3 and Noggin after 10 days of co-culture
in osteogenic medium. WB analysis of the osteogenic medium showed an increased in
Noggin and BMP3 protein expression when control bone, patent suture and fused suture
were compared. A) BMP3 expression was significantly (p<0.0001) up regulated only in
patent suture osteoblasts co-cultured with dura mater cells. B) Noggin expression showed
significantly (p<0.0001) increased expression in all three groups when osteoblasts were co-
cultured with dura mater cells compared with monocultures.
Aim IV) To investigate the role of dura mater paracrine signalling in the
pathophysiology of craniosynostosis in humans.

70
Dura Mater Expression of FGF2 and TGF-β1
In order to understand the influence of dura mater cells on osteoblast behavior, we
then searched for candidate molecules that may be responsible for dura mater paracrine
signalling modifying osteoblast behavior. We chose FGF2 and TGF-β1 to examine. Western
Blot of the conditioned medium obtained at days 3,5,7 and 10 from dural cultures
demonstrate the presence of FGF2 and TGF-β1. TGF-β1 levels remained consistent
throughout the study period whereas FGF2 levels declined after day 3 (Figure 29).
FGF2
TGF-ß1 44kDa
47.5kDa
D3 D5 D7 D10 D3 D5 D7 D10
Dura from 3 year old patient Dura from 11 years old patient
FGF2
Day 3 Day 5 Day 7 Day 100
10
20
30
40
FG
F-2
Den
sit
y
TGF- B1
Day 3 Day 5 Day 7 Day 100
10
20
30
40
TG
F-
B1 D
en
sit
y
Mean SD
Day 3: 14.8 2.82
Day 5: 12.8 0.28
Day 7:7.35 5.3
Day 10: 10.1 0.424
Mean SD
Day 3: 22.05 0.05
Day 5: 11.6 3.8
Day 7:10.85 4.150
Day 10: 5.5 0.5
Actin 43KDa
Figure 29: Expression of pro-osteogenic molecules in the conditioned medium
from dura mater cells. TGF-ß1 did not show significant variation between days. FGF2 had a
two-fold higher expression at day 3 when compared with the subsequent days. Western blot
analyses by densitometry.

71
Aim V) To assess the effects of exogenous administration of FGF2 on osteoblast
function in vitro.
Recombinant Human FGF2 Stimulation
FGF2 was expressed in higher concentration in the medium from dura mater cells
when compared with TGF-β1. We then decided to add exogenous FGF2 to control bone,
patent suture and fused suture osteoblast cell cultures to evaluate the ability of osteoblasts to
respond to this stimulation. The addition of FGF2 to culture medium enhanced cellular
proliferation at all 4 concentrations for the three groups in a dose dependent fashion.
Osteoblasts from fused suture were more sensitive to lower doses of stimulation (5ng/ml –
p<0.05) at day 7 than the other two groups.

72
Proliferation
Control Bone
5ng/ml 10ng/ml 50ng/ml 100ng/mlNo stimulation
200x
Patent Bone
Fused Bone
200x
200x
Figure 30: Proliferation of osteoblasts stimulated with FGF2. At 3 days of
incubation, an increase in proliferation was observed for all three groups independent of the
FGF2 concentration compared with non-stimulated osteoblasts shown in the photos of cell
culture plates.

73
MTT-Control Bone
Day 1 Day 3 Day 5 Day 70
5
10
15Control Bone
5 ng/ml
10ng/ml
50ng/ml
100ng/ml
A)
** *
p<0.0001(two-way ANOVA)n=4
Co-culture with Dura Mater
* *
OD
/1000 c
ells
MTT-Patent Suture
Day 1 Day 3 Day 5 Day 70
5
10
15Patent Suture
5ng/ml
10ng/ml
50ng/ml
100ng/ml
B)
*
*
p<0.0001(two-way ANOVA)n=4
Co-culture with Dura Mater
OD
/1000 c
ells
MTT-Fused Suture
Day 1 Day 3 Day 5 Day 70
5
10
15Fused Suture
5ng/ml
10ng/ml
50ng/ml
100ng/ml
C)
**
**
***
*
p<0.0001(two-way ANOVA)n=4
Co-culture with Dura Mater
OD
/1000 c
ells
Figure 31: Proliferation Rates after Stimulation with FGF2 (MTT): Stimulation
with recombinant human FGF2 significantly (p<0.0001) increases the rate of proliferation

74
for all three groups (in a dose dependent manner) compared with osteoblasts without
stimulation. A) In the control bone group proliferation is significantly greater with 50ng/ml
(p<0.05) at days 5 and 7; 100ng/ml (p<0.01) at days 5 and in co-culture with dura mater
cells at days 5 (p<0.001) and 7 (p<0.01). B) In the patent suture group proliferation is
significantly greater in co-culture with dura mater cells at day 5 (p<0.05) and 7 (p<0.001)
C) In the fused suture group proliferation is significantly greater with 5ng/ml at day
7(p<0.05); 50 ng/ml at days 3(p<0.05),5 (p<0.01)and 7(p<0.05) and with 100ng/ml at days
5 and 7(p<0.0) and in co-culture with dura mater cells at days 5 and 7 (p<0.001).

75
Differentiation
Osteoblasts cultured with exogenous FGF2 demonstrated an increase in alkaline
phosphatase expression in all three groups at days 3,5 and 7. However the differences
between groups were not significant.
Control Bone
Patent Bone
No stimulation 5ng/ml 10ng/ml 50ng/ml 100ng/ml
Fused Bone
200x
200x
200x
Figure 32: AP expression after stimulation with FGF2. At 3 days of incubation,
increased alkaline phosphatase staining is seen in all three groups treated with FGF2
independent of the FGF2 concentration.

76
Alkaline PhosphataseControl Bone
Day 1 Day 3 Day 5 Day 70.0
0.2
0.4
0.6
0.8Control Bone
5 ng/ml
10ng/ml
50ng/ml
100ng/ml
A)
Co-culture with Dura Mater
p<0.01(two-way ANOVA)n=4
mm
ol o
f p
NP
/h/µ
g o
f p
rote
in
Alkaline PhosphatasePatent Suture
Day 1 Day 3 Day 5 Day 70.0
0.2
0.4
0.6
0.8Patent Suture
5ng/ml
10ng/ml
50ng/ml
100ng/ml
Co-culture with Dura Mater
B)
p=0.1(two-way ANOVA)n=4
mm
ol o
f p
NP
/h/µ
g o
f p
rote
in
Alkaline PhosphataseFused Suture
Day 1 Day 3 Day 5 Day 70.0
0.2
0.4
0.6
0.8Fused Suture
5ng/ml
10ng/ml
50ng/ml
100ng/ml
Co-culture with Dura Mater
C)
p=0.27(two-way ANOVA)n=4
mm
ol o
f p
NP
/h/µ
g o
f p
rote
in
Figure 33: APA after stimulation with FGF2: Osteoblasts cultured with exogenous
FGF2 demonstrated an increase in alkaline phosphatase in all three groups at days 3,5 and
7 compared with day 1. Final AP concentration is expressed as mmol of pNP per hour per
µg of protein.

77
Discussion

78
The work developed herein validates our hypotheses that regional variations in
osteoblast behavior in infants with craniosynostosis do exist.
We first developed a human model of osteoblast cell culture by obtaining calvarial
bone samples from patent sutures, fused sutures and adjacent bone from infants with
craniosynostosis. Histologically, we confirmed regional variations between suture sites and
adjacent bone. The ability to obtain osteoblasts capable of forming bone was demonstrated
by cell proliferation, presence of Runx2 and alkaline phosphatase, collagen I expression,
osteocalcin production, mineralization and transmission electron microscopy. Cells derived
from fused suture sites clearly demonstrated greater osteogenic potential with higher rates of
proliferation and differentiation when compared with cells from control bone and patent
suture. We also searched for factors in the surrounding enviroment that may contribute to the
imbalance between pro- and anti-osteogenic signalling. Noggin and BMP3, both anti-
osteogenic molecules were significantly greater expressed by patent sutures than by fused
sutures and control bone. Runx2 expression was upregulated in the fused suture osteoblasts
combined with dura mater demonstrating that osteoblasts from fused sutures may be more
differentiated that their counterparts. Dura mater cells were shown to mainly influence
osteoblast differentiation and mineralization, downregulating those parameters in fused
suture cells and enhancing proliferation and differentiation in patent suture cells. On the other
hand, exogenous FGF2 appeared to regulate proliferation in a dose dependent fashion, with
osteogenic cells from fused sutures being more sensitive to lower doses of FGF2. Taken
together, our findings demonstrated interactions between calvarial osteogenic cells and dura
mater and also validate our hypothesis that regional variations exist in the calvarial bones of
infants with craniosynostosis.

79
Development of a valid model of osteogenic culture derived from cranial suture sites
Over the past several years, many studies have used animal models to investigate the
biology underlying premature suture fusion28, 29,48
. Specifically, the sagittal suture in a
murine model has been utilized as a model of normal patent human suture biology and the
fusing posterior frontal suture as a model of premature fusion. A rabbit cranial suture model
from a colony of congenitally fused rabbits29
has also been studied in an attempt to
understand the mechanisms underlying syndromic craniosynostosis.
Despite extensive research based on animal models, there is little literature on the use
of human cranial tissue samples to investigate the biomolecular mechanisms involved in
craniosynostosis. Recent studies have been limited by the number of samples obtained from
non-syndromic patients60
. We were able to develop a valid human model of osteogenic cell
culture derived from cranial suture sites, and obtain a large number of samples representing
the wide spectrum of this condition. Our study is the largest one compared to those in the
current literature. We obtained samples from 45 patients with syndromic and non-syndromic
craniosynostosis representative of the epidemiology of craniosynostosis with regards to with
age, suture affected and sex. We also obtained samples from different regions of the cranial
vault in all patients, representing the fused suture, the patent suture and adjacent non-sutural
bone.
In order to validate our bone cell culture model and demonstrate the presence of
osteoblasts in cell culture, we assessed cellular proliferation, Runx2 expression, alkaline
phosphatase activity, collagen I and osteocalcin expression and mineralization. Cells isolated
from patent suture, fused suture and adjacent bone demonstrated characteristics of the
osteoblast phenotype with rapid proliferation, presence of alkaline phosphatase activity,

80
expression of Runx2, and in vitro bone formation. The earliest stages of osteoblast
differentiation can be detected through the measurement of alkaline phosphatase and Runx2.
Although alkaline phosphatase can be expressed by other cell types such as fibroblasts,
Runx2 is a specific marker of osteoblast differentiation and was expressed by all cells of our
culture model, confirming that cells were able to differentiate in osteoblasts. In cultures
containing ascorbic acid and β-glycerophosphate, the secretion of extracellular matrix by
differentiated osteoblasts was followed by progressive mineralization and bone nodule
formation. To further confirm our findings of in vitro bone formation, ultrastructural analysis
of the samples was performed, demonstrating features of in vitro osteogenesis, including
mineral depositions and collagen fibrils in the extracellular matrix. A ring pattern for electron
diffraction studies was characteristic of normal hydroxyapatite.
Taken all together, the above findings allow us to conclude that the human bone cell
culture model described herein is valid and reliable.
We determined the most favorable conditions for osteogenic culture by varying
medium composition based on animal models previously described. The development of
osteoblast function and morphology in vivo strongly depends on the composition of the
culture medium, particularly essential factors that influence various stages of proliferation
and differentiation during the process of osteogenesis68
. Early studies of osteoblastic cell
lines have demonstrated that osteoblasts undergo a temporal sequence of differentiation,
featuring active cell proliferation, expression of osteoblastic markers, synthesis and
deposition of a collagenous extracellular matrix, as well as mineralization, in order to
develop normal osteoblast phenotype and function68
.

81
Osteoblast proliferation rates were higher in the fused sutures, independent of
experimental culture conditions. This observation is consistent with a study that showed bone
formation activity at the fused suture site is increased in non-syndromic craniosynostosis,
leading to premature ossification69
. The same study in humans showed that at all ages (3 – 18
months), the proliferation of osteoblasts obtained from fused suture locations was similar to
that from patent sutures69
and both AP activity and osteocalcin production by osteoblast cells
originating from fused sutures were significantly higher than that shown in cells derived from
normal sutures, upon stimulation with vitamin D69
. The difference in findings between the
previous work69
and our study may be attributed to different growth conditions used in the
two investigations. Studies by Coelho et al. have shown that the presence of dexamethasone
in combination with ascorbic acid and β-glycerophosphate resulted in significantly increased
cell growth and AP activity68
.
The effects of ascorbic acid have been well defined by previous studies, which have
emphasized its critical role in collagen biosynthesis, particularly in the production of the
collagenous bone extracellular matrix68
due to its role as a cofactor for proline hydroxylase
and lysine hydroxylase, which are involved in the hydroxylation of collagen. A Na+-
dependent transporter specific for ascorbic acid is present in the plasma membrane of
osteoblasts and is essential for maintenance of intracellular ascorbate concentrations70
. One
pathway that mediates ascorbate’s effect on osteoblast growth and differentiation is the
collagen receptors or integrins70
. In addition, osteoblast differentiation as mediated by 1,25
(OH)2 Vitamin D3, retinoic acid, and bone morphogenic proteins is also affected by the
concentration of ascorbic acid. Thus, ascorbic acid is essential for normal bone formation70
.
Our study showed that during a 10 day time interval, regional variations in osteoblast

82
proliferation rates are evident in infants with craniosynostosis. Fused suture cell cultures,
supplemented with 15% FBS + ascorbic acid + α-glycerophosphate demonstrated
significantly higher growth rates, in comparison to both αMEM (1% FBS) and αMEM
(15%FBS) on day 3. Studies by Prader et al., which used similar ascorbic acid
supplementation, demonstrated that an optimal amount of ascorbic acid (50µg/mL)
stimulates osteoblastic differentiation, as well as dose-dependent collagen synthesis, leading
to higher AP values62
. This relationship between cell differentiation rates and ascorbic acid
was investigated by Franceschi in 1992, who proposed that the collagen matrix synthesized
by cells treated with ascorbic acid, provides a more favourable environment for tissue-
specific gene expression in mesenchymal-derived tissues like bone, resulting in higher
differentiation rates.
Cell culture models are a critical tool in cranial suture biology research. This study
has described the effects of culture medium composition on proliferation and differentiation
of human calvarial osteogenic cells. Furthermore, for human calvarial osteogenic cells,
αMEM with 10-7
M dexamethasone, supplemented with 15% FBS and ascorbic acid proved to
be the best culture medium, resulting in high growth and differentiation rates. These results
reveal the importance in defining the experimental conditions in prospective osteoblast cell
studies68
.
Assessment of regional variations in osteoblast behavior with and without dura mater
cells in co-culture.
In the current study, MTT and alkaline phosphatase activities, as indices of bone cell
proliferation and differentiation, respectively, were greater in osteogenic cultures from fused
sutures compared with patent sutures and adjacent non-sutural bone. Fused suture cells

83
proliferated faster and differentiated earlier then control and patent suture cells. Proliferation
rates were also higher in younger patients when compared with patients over 12 months of
age (seen in the syndromic group) which is in accordance with the literature71
. Hassler et al71
demonstrated that after surgical treatment of sagittal synostosis in children aged 6 months or
younger, reossification usually started 2 weeks postoperatively and was complete within 6
months. In children aged 7 to 12 months, reossification was prolonged and lasted for 12
months or longer and in children older than 12 months reossification was incomplete with
persistent pseudosutures, compromising surgical results. Patients with syndromic
craniosynostosis in our study were older (mean 12.6 months old) than patients with non-
syndromic craniosynostosis (7.5 months old). The delayed surgical procedure is usually
associated with other comorbidities found in syndromic patients. Cells retrieved from older
patients may not retain the same potential for proliferation and differentiation once they
reach maturity. Animal studies corroborate the influence of age on osteoblast proliferation44
.
It also may suggest that osteoblasts from younger patients are more susceptible to the dura
mater paracrine effect and the paracrine effects may last longer in vitro. The influence of age
in osteoblast behavior will be investigated in depth in further studies using our cell culture
model.
Osteoblasts from non-sutural bone, patent and fused sutures also showed a
significant difference in their alkaline phosphatase activity during the early period of culture;
alkaline phosphatase activity was significantly greater (p<0.05) during this period in the
fused suture cultures.
Formation of bone nodules in cultures derived from fused sutures was not only
detected earlier (11±3 days) compared with control (13±4 days) and patent sutures (16±4

84
days) osteoblasts, but also individual nodules in these cultures were two to three times larger
in diameter than those derived from control and patent sutures (p<0.0001). These results
suggest that osteoblasts derived from fused sutures exhibit an aggressive pattern of bone
formation, compared with their normal counterparts. TEM revealed ultrastructural features of
in vitro osteogenesis, including mineral depositions and dissected collagen fibrils in the
extracellular matrix.
In view of the ability of fused suture osteoblasts to aggressively form bone in vitro,
we examined the levels of Runx2 (also known as Cbfa-1). Runx2, a transcription factor that
is a marker of osteoblast differentiation, has been localized to the critical area of cranial
suture fusion. Runx2 is found in the osteogenic fronts and sutural mesenchyme and has
variable expressivity in patent versus fusing sutures in normal development29
. By
immunofluorescence we were able to show that Runx2 was expressed by adjacent bone,
patent suture and fusing suture, demonstrating that cells from all three sites have the ability to
differentiate. WB analyses demonstrated up-regulation of Runx2 expression at the fused
suture site when osteoblasts were combined with dura mater cells. Based on our findings,
Runx2 did not show significant differences when osteoblast from different sites were
evaluated, suggesting that it may not regulate suture patency and fusion itself, but it may
interact with other molecules or respond to dura mater paracrine signalling determining the
osteoblast differentiation in premature suture fusion.
The findings of accelerated in vitro bone formation parallel the clinical and
histological observations of partial or complete suture fusion suggesting important regional
variations in osteoblast behavior in infants with craniosynostosis.

85
The Role of Dura Mater
Over the last decade, numerous animal studies have suggested that the immature dura
mater is the critical factor governing successful calvarial ossification72, 73
. More recently,
studies have begun to identify the biomolecular and cellular differences between immature
and mature dura mater74
. Collectively, these studies suggest that the immature dura mater
supplies growth factors and/or osteoblast-like cells that participate in successful calvarial
reossification. Longaker67
et al., using a novel co-culture system, demonstrated, for the first
time, that osteoblast proliferation, gene expression, and bone nodule formation can be
directly modulated by paracrine signals from immature, non–suture-associated dural cells.
Furthermore, that study demonstrated that immature, non–suture-associated dura mater,
isolated or in co-culture, produced high levels of osteoinductive cytokines like TGF-β1 and
FGF2.
Although clinical observations and animal studies have demonstrated that young dura
mater (3-days-old mice) is both osteogenic and osteoconductive67
, to date no study has
clarified the interaction between human cranial sutures and human dura mater in
craniosynostosis. To better understand the contributions of dura mater and osteoblasts to
calvarial ossification, we co-cultured mature human dura mater with osteoblasts to examine
the effects of dural cells on the biologic activity of co-cultured osteoblasts. Our co-culture
model allowed us to investigate the in vitro interactions between human dura mater and the
calvarial bones. The dura mater cells were obtained from neurosurgical procedures. A
potential limitation of our study includes the age of neurosurgical patients undergoing
surgery for epilepsy. Although we attempted to age-match our patients, craniofacial patients
were younger at the time of the surgery (3-13 months old) compared with neurosurgical

86
patients (3 to 17 years old). We were also not able to select specific sites where dural samples
were harvested. Specifically we could not match the dural sites to the cranial suture sites and
samples from fused sutures could not be obtained. However, despite these shortcomings,
dural co-culture demonstrated that osteoblasts were influenced by the presence of dura in the
following ways: enhancing proliferation and differentiation for osteoblasts from patent
sutures; enhancing differentiation for osteoblasts from control bone and promoting the same
pattern of mineralization for all three groups.
Herein we demonstrated that osteogenic cultures from the fused suture and the control
bone co-cultured with dura mater showed no difference in proliferation rates compared with
osteogenic monocultures from the same areas cultured without dura. We speculate it may be
due to the age of patients from whom dura mater was harvested. It may influence
proliferation rates, with less mature dura mater from age-matched patients perhaps enhancing
proliferation rates for all three groups. However, co-cultured control bone and patent suture
osteogenic cultures expressed significantly (p<0.05) greater levels of alkaline phosphatase
when compared with osteogenic cultures without dura. There were no differences in pattern
of mineralization seen at day 28 in osteoblasts co-cultured with dura mater cells when
comparing control, patent suture and fused suture, suggesting that interaction with dura mater
cells resulted in down-regulation of mineralization in vitro. These findings suggest that
paracrine signalling from dura mater controls mineralization at the suture site by up or
downregulating osteoblast differentiation.

87
The role of anti-osteogenic signalling on human osteoblasts in vitro with and without
dura mater cells in co-culture
Our data suggest an important role for Noggin in pathologic suture fusion. Low
expression of Noggin was found in the fused suture when osteoblasts were cultured alone. In
the same system, patent suture osteoblasts expressed higher levels of Noggin. Interestingly,
Noggin was expressed in the conditioned medium from all three sites when combined with
dura mater cells, suggesting that dura mater cells may be the source of Noggin, maintaining
the patency of the suture line. These findings are in concordance with the literature. Warren
et al27, 75
have examined postnatal suture mesenchyme in an attempt to determine Noggin’s
role in normal suture fusion in a murine model. They found that Noggin was expressed by
the patent sagittal suture but not by the fused posterior frontal suture. They also found that
expression of Noggin was decreased by FGF2 and that overexpression of Noggin, induced by
gene transfection, resulted in suture patency of the normally fused suture.
BMP3 has also been implicated in the process of normal suture fusion in mice. Our
results confirm the findings from the normal suture fusion model. BMP3 was strongly
expressed by the patent suture as demonstrated by IHC, but BMP3 was not expressed by the
fused suture. Expression of BMP3 was increased in co-cultures of dura mater cells with
patent suture cells, which did not occur in fused suture or control sites osteogenic co-cultures.
This may suggest that Noggin and BMP3 do not have the same responses to dura mater
paracrine signalling. Based on our findings, co-culture with dura mater cells was able to
change only the pattern of Noggin expression leading us to speculate that dura mater could
be a source of Noggin regulating suture patency and fusion. This will be further investigated
in our laboratory.

88
Further studies evaluating the influence of dura mater and/or Noggin in Runx2
expression, are necessary to elucidate osteoblast behavior and regional variations in
craniosynostosis.
The role of dura mater paracrine signalling in the pathophysiology of craniosynostosis
To better understand the influences of dura mater and the biomolecular mechanisms
mediating the observed changes in proliferation, gene expression, and bone nodule formation
of co-cultured osteoblasts, we examined the expression of TGF-β1 and FGF2 in the
conditioned medium from dural cell cultures at days 3, 5, 7 and 10.
We focused on these candidate molecules because evidence suggests dura mater is a
source of FGF2 and TGF-β1 and they promote osteogenesis and osseous repair in vivo72, 76
.
The actions of these growth factors are highly concentration dependent and influence a
number of cellular processes77
. Our results demonstrated dural cells expressed both TGF-β1
and FGF2. It is possible that FGF2 released by dura mater cells may be a key growth factor
responsible for the paracrine effects of dura mater in controlling suture patency and fusion.
Considering we were able to identify the expression of these molecules in the
medium from dura mater cells, we questioned whether the levels of dural cell TGF-β1 and
FGF2 were responsible for the observed changes in proliferation and differentiation of co-
cultured osteoblasts. The literature concerning the effect of TGF-β1 on osteoblasts in vitro
contains many conflicting reports. TGF-β1 may cause either an increase or decrease in
osteoblast proliferation depending on the type of cell and cellular density78, 79
. Irrespective of
its effects on osteoblast proliferation, TGF-β1 is a potent chemoattractant for osteoblast-like

89
cells. Thus, the osteoinductive effects of TGF-β1 in vivo may instead be related to its ability
to recruit osteoprogenitor cells80
.
Interestingly, several studies have demonstrated that TGF-β1 has conflicting effects
on osteoblast type I collagen and alkaline phosphatase expression81-83
. Most notably, in
contrast to its in vivo activity, in vitro TGF-β1 seems to retard osteoblast differentiation and
decrease osteoblast expression of osteocalcin and the formation of mineralized bone
nodules81
. Thus, it is unlikely that TGF-β1 alone could account for the dramatic phenotypic
differences observed in co-cultured osteoblasts. Based on these reports, we did not
investigate the influence of TGF-β1 in osteoblast behavior. We decided to focus on FGF2
effects on osteoblast proliferation and function.
Fibroblast growth factors are a highly conserved family of at least 19 closely related
monomeric peptides. FGF2 is the most abundant ligand, and it has been shown to stimulate
osteoblast proliferation and enhance bone formation in vivo and in vitro84
. FGF2 expression
is elevated during fracture healing, and exogenously applied FGF2 accelerates osteogenesis
in critically sized bone defects and at fracture sites85
. Despite its in vivo osteoinductive
properties, FGF2 had been thought, until recently, to decrease osteoblast proliferation and
expression of markers of differentiation, such as type I collagen, alkaline phosphatase, and
osteocalcin86
. However, a recent study by Debiais et al58
demonstrated that an osteoblast’s
response to FGF2 is stage specific. Immature osteoblasts (3 days old mice) show significant
increase in proliferation under FGF2 stimulation in comparison to mature osteoblasts (30
days old mice). Continuous (7-day) FGF2 treatment of differentiating human calvarial–
derived osteoblasts (immature cells) resulted in increased osteocalcin production and matrix
mineralization. The above-mentioned studies on the isolated effects of TGF-β1 and FGF2 are

90
interesting, but osteoblasts in a calvarial defect are exposed simultaneously to a variety of
cytokines. Could TGF-β1 and FGF2 interact to affect osteoblast activity? In one of the only
studies to investigate the effect of simultaneous TGF-β1 and FGF2 stimulation on osteoblast
behavior, Globus et al87
demonstrated that these cytokines acted in a dose-dependent,
synergistic manner to increase the rate of bovine osteoblast proliferation.
Effect of exogenous human recombinant FGF2 administration on osteoblast function in
vitro
The increased rate of proliferation of our co-cultured patent suture osteoblasts may
have resulted from the synergistic effects of TGF-β1 and FGF2 elaborated by the dural cells.
To date no studies have adequately examined the effect of simultaneous (long-term) TGF-β1
and FGF2 treatment on osteoblast gene expression and differentiation.
To elucidate the effects of FGF2 on osteoblast behavior in short-term cultures we
supplemented our standard osteogenic media with human recombinant FGF2 (5, 10, 50 and
100ng/ml)45
. Cells derived from control bone, patent suture and fused suture demonstrated a
significant (p<0.05) increase of proliferation rates over time and compared with unstimulated
osteoblasts, showing that FGF2 is a potent mitogenic factor in human calvarial cells. In the
FGF2 stimulation experiments, unstimulated osteogenic cells from all three groups
demonstrated similar rates of proliferation at days 5 and 7 when compared with our previous
experiments on proliferation where fused suture osteoblasts demonstrated significantly
higher rates of proliferation. Cell cultures used for FGF2 experiments were derived from
older patients (mean age 11.7 months old) and mainly from coronal sutures, whereas cell
cultures used for the proliferation experiments were mainly from sagittal sutures and younger

91
patients (5.1 months old). These findings support the influence of age on osteoblast behavior
and may also suggest that osteoblasts from different sutures behave differently.
In accordance with previous observations, quantification of alkaline phosphatase was
not significantly increased when comparing the osteoblasts unstimulated with osteoblasts
stimulated by different concentrations of FGF2. When comparing results obtained with FGF2
stimulation and co-culture with dura mater cells, the latest was responsible for a more
consistent enhancing in osteoblast proliferation. These findings suggest that the effects of
FGF2 are dose dependent and may be also depend of the timing of exposure to growth
factors. In vivo, during osteogenesis, osteoblasts are exposed to dura mater influence since
the first trimester, which may explain the behavior of cells at different suture sites.
Unfortunately, we have not been able to develop such an ideal condition to confirm those
suppositions.
These observations highlight the multifactorial nature of calvarial bone development,
suggesting that premature cranial suture fusion most likely results from activation of
osteoprogenitors and osteoblasts in combination with the proliferation and differentiation of a
subpopulation of osteoblast-like cells residing within the dura mater itself.
In summary, the study reported here demonstrated that dura mater can affect
proliferation, rate and degree of differentiation, and bone nodule formation. Furthermore, we
demonstrated that non–suture-associated dura mater in isolation produced high levels of
osteoinductive cytokines (i.e., TGF-β1 and FGF2). Although these candidate cytokines cause
conflicting effects on osteoblasts in vitro and in vivo, they are interesting candidates for the
changes in behavior observed in co-cultured osteoblasts. As a future direction of this work
we aim to decipher and understand the exact nature of paracrine signalling responsible for the

92
observed changes in phenotype and function of co-cultured osteoblasts and include Noggin
as potential candidate for the paracrine signalling by dura mater cells.

93
Conclusion

94
The work herein demonstrates that regional variations in osteoblast behavior exist in
the calvaria of infants affected by craniosynostosis. A successful cell culture technique for
generating a reliable osteoblast culture was developed. Co-culture with dura mater
demonstrated that there was no significant effect on osteoblast proliferation rates from
regions of fused suture and control bone. However, dural influence in vitro was noted in AP
expression and mineralization rates suggestive of a paracrine effect. This was demonstrated
by assays detecting important cytokines (FGF2 and TGF-β1) in the media of the dura mater
cultures.
The increased expression of anti-osteogenic molecules, Noggin and BMP3 in patent
sutures osteoblasts suggests that they play a role in the maintenance of suture patency and
down-regulation at levels similar with control bone was noted in the fused suture osteoblasts
acting as further evidence of their role.
The creation of a pro-osteogenic environment that would support premature fusion of
the cranial sutures was further corroborated with the Runx2 studies. These studies
demonstrated that Runx2, a transcription factor that is important in osteoblast differentiation
was expressed by fused suture cells. This would suggest that these cells are more responsive
and amenable to an environment that would promote osteogenesis and hence, fusion of the
sutures.
Exogenous FGF2 was demonstrated to selectively enhance growth of fused suture
cell cultures. This pro-osteogenic molecule did not have the same impact on cells from patent
sutures, thereby confirming the presence of regional variations in osteoblast behavior.

95
In summary, we would support the theory that activation of the fibroblast growth
factor signalling pathway secreted by dura mater potentiates suture osteogenesis by inhibiting
the expression of osteogenic antagonists such as Noggin and BMP3, allowing agonists such
as bone morphogenic proteins (BMP2 and 4) to increase osteoblastic differentiation from
cells in the suture mesenchyme leading to premature suture fusion.
A better understanding of the mechanism underlying of this condition could lead to
the development of new therapeutic modalities. For example, if nonsyndromic
craniosynostosis derives from aberrant growth factor production or action, correction of this
problem might be possible through selective drug therapy. Gene therapy to correct the
molecular defect(s), however, will clearly require a thorough understanding of the complex
cascades regulating suture fusion and patency.

96
Future Directions

97
As a future direction of this work we aim to decipher and understand the exact nature
of paracrine signalling responsible for the observed changes in phenotype and function of co-
cultured osteoblasts.
Although we demonstrated important differences in osteoblast behavior depending on
the suture site and the interaction with dura mater cells, we were limited by the age of
patients who underwent neurosurgical procedures. In the future, studies obtaining dura mater
samples from age-matched patients and from the fused suture site would shed light on the
mechanisms involved in the local paracrine signalling by dura mater cells. The current co-
culture model described in our work also does not account for the effects of cell-cell
interaction or the effects of conditioned medium collected from dura mater. Those
interactions would be further investigated based on our human bone culture model of
craniosynostosis. By enhancing our understanding of the cellular events which comprise
suture formation and growth, it may be possible to recapitulate, in an experimental setting,
the normal development of a suture at a specific calvarial site.
Based on the findings presented in this work, a further step would be to investigate in
greater depth the influence of age in osteoblast behavior. Osteoblasts from non-syndromic
patients demonstrated regional variations not seen in the osteoblasts from syndromic patients.
Furthermore, proliferation rates were equal for all three groups in the FGF2 experiments
when osteoblasts were not stimulated. Reviewing patients’ age, osteoblasts for those
experiments were retrieved from older patients (mean age 11.7 months) which could explain
the lack of differences. Taken all together, investigation of the influence of age in osteoblast
behavior could shed light some of the contradictory findings of this work.

98
Another important step forward would be to understand in greater depth the role of
Noggin in the pathophysiology and therapeutics of craniosynostosis. Further experiments
will investigate the expression of Noggin by dura mater cells. Noggin also will be added to
cultured cells, as well as, anti-Noggin antibody to neutralize its effects and parameters of
proliferation and function will be reassessed. Patients will be also stratified by age to
determine the ideal time point where cells would be able to respond to a therapy.
Though at present we are just beginning to understand the phenomenon of premature
suture fusion, ultimately, this may lead to a new therapeutic modality as an alternative to
invasive procedures which would restore both the structure and function of the developing
cranium.

99
References

100
1. Shen K, Krakora SM, Cunningham M, et al. Medical treatment of craniosynostosis:
recombinant Noggin inhibits coronal suture closure in the rat craniosynostosis model. Orthod
Craniofac Res 2009;12:254-62.
2. Delashaw JB, Persing JA, Broaddus WC, Jane JA. Cranial vault growth in
craniosynostosis. J Neurosurg 1989;70:159-65.
3. Cohen MM, Jr., Kreiborg S. Birth prevalence studies of the Crouzon syndrome:
comparison of direct and indirect methods. Clin Genet 1992;41:12-5.
4. Renier D, Sainte-Rose C, Marchac D, Hirsch JF. Intracranial pressure in
craniostenosis. J Neurosurg 1982;57:370-7.
5. Kapp-Simon KA, Figueroa A, Jocher CA, Schafer M. Longitudinal assessment of
mental development in infants with nonsyndromic craniosynostosis with and without cranial
release and reconstruction. Plast Reconstr Surg 1993;92:831-9; discussion 40-1.
6. Moore MH. Upper airway obstruction in the syndromal craniosynostoses. Br J Plast
Surg 1993;46:355-62.
7. Lo LJ, Chen YR. Airway obstruction in severe syndromic craniosynostosis. Ann Plast
Surg 1999;43:258-64.
8. Virtanen R, Korhonen T, Fagerholm J, Viljanto J. Neurocognitive sequelae of
scaphocephaly. Pediatrics 1999;103:791-5.
9. Hayward R, Jones B, Evans R. Functional outcome after surgery for trigonocephaly.
Plast Reconstr Surg 1999;104:582-3.
10. Speltz ML, Greenberg MT, Endriga MC, Galbreath H. Developmental approach to
the psychology of craniofacial anomalies. Cleft Palate Craniofac J 1994;31:61-7.

101
11. Pertschuk MJ, Whitaker LA. Psychosocial considerations in craniofacial deformity.
Clin Plast Surg 1987;14:163-8.
12. Coccaro PJ, McCarthy JG, Epstein FJ, Wood-Smith D, Converse JM. Early and late
surgery in craniofacial dysostosis: a longitudinal cephalometric study. Am J Orthod
1980;77:421-36.
13. Kreiborg S, Aduss H. Pre- and postsurgical facial growth in patients with Crouzon's
and Apert's syndromes. Cleft Palate J 1986;23 Suppl 1:78-90.
14. McCarthy JG, Cutting CB. The timing of surgical intervention in craniofacial
anomalies. Clin Plast Surg 1990;17:161-82.
15. DiRocco C, Caldarelli M, Ceddia A, Iannelli A, Velardi F. [Craniostenosis. Analysis
of 161 cases surgically treated during the first year of life]. Minerva Pediatr 1989;41:393-
404.
16. Moss ML. Growth of the calvaria in the rat; the determination of osseous
morphology. Am J Anat 1954;94:333-61.
17. Pritchard JJ, Scott JH, Girgis FG. The structure and development of cranial and facial
sutures. J Anat 1956;90:73-86.
18. Markens IS. Embryonic development of the coronal suture in man and rat. Acta Anat
(Basel) 1975;93:257-73.
19. Cohen MM, Jr. Sutural biology and the correlates of craniosynostosis. Am J Med
Genet 1993;47:581-616.
20. Johansen VA, Hall SH. Morphogenesis of the mouse coronal suture. Acta Anat
(Basel) 1982;114:58-67.

102
21. Kokich VG. Age changes in the human frontozygomatic suture from 20 to 95 years.
Am J Orthod 1976;69:411-30.
22. Winograd JM, Im MJ, Vander Kolk CA. Osteoblastic and osteoclastic activation in
coronal sutures undergoing fusion ex vivo. Plast Reconstr Surg 1997;100:1103-12.
23. Koskinen L, Isotupa K, Koski K. A note on craniofacial sutural growth. Am J Phys
Anthropol 1976;45:511-6.
24. Moss ML. Fusion of the frontal suture in the rat. Am J Anat 1958;102:141-65.
25. Ozaki W, Buchman SR, Muraszko KM, Coleman D. Investigation of the influences
of biomechanical force on the ultrastructure of human sagittal craniosynostosis. Plast
Reconstr Surg 1998;102:1385-94.
26. Kirschner RE, Gannon FH, Xu J, et al. Craniosynostosis and altered patterns of fetal
TGF-beta expression induced by intrauterine constraint. Plast Reconstr Surg 2002;109:2338-
46; discussion 47-54.
27. Warren SM, Brunet LJ, Harland RM, Economides AN, Longaker MT. The BMP
antagonist noggin regulates cranial suture fusion. Nature 2003;422:625-9.
28. Nacamuli RP, Fong KD, Lenton KA, et al. Expression and possible mechanisms of
regulation of BMP3 in rat cranial sutures. Plast Reconstr Surg 2005;116:1353-62.
29. Gabbay JS, Heller J, Spoon DB, et al. Noggin underexpression and runx-2
overexpression in a craniosynostosis rabbit model. Ann Plast Surg 2006;56:306-11.
30. Greenwald JA, Mehrara BJ, Spector JA, et al. In vivo modulation of FGF biological
activity alters cranial suture fate. Am J Pathol 2001;158:441-52.
31. Urist MR. Bone morphogenetic protein: the molecularization of skeletal system
development. J Bone Miner Res 1997;12:343-6.

103
32. Hino J, Matsuo H, Kangawa K. Bone morphogenetic protein-3b (BMP-3b) gene
expression is correlated with differentiation in rat calvarial osteoblasts. Biochem Biophys
Res Commun 1999;256:419-24.
33. Daluiski A, Engstrand T, Bahamonde ME, et al. Bone morphogenetic protein-3 is a
negative regulator of bone density. Nat Genet 2001;27:84-8.
34. Candia AF, Watabe T, Hawley SH, et al. Cellular interpretation of multiple TGF-beta
signals: intracellular antagonism between activin/BVg1 and BMP-2/4 signaling mediated by
Smads. Development 1997;124:4467-80.
35. Fong KD, Song HM, Nacamuli RP, et al. Apoptosis in a rodent model of cranial
suture fusion: in situ imaging and gene expression analysis. Plast Reconstr Surg
2004;113:2037-47.
36. Mehrara BJ, Mackool RJ, McCarthy JG, Gittes GK, Longaker MT.
Immunolocalization of basic fibroblast growth factor and fibroblast growth factor receptor-1
and receptor-2 in rat cranial sutures. Plast Reconstr Surg 1998;102:1805-17; discussion 18-
20.
37. Greenwald JA, Mehrara BJ, Spector JA, et al. Regional differentiation of cranial
suture-associated dura mater in vivo and in vitro: implications for suture fusion and patency.
J Bone Miner Res 2000;15:2413-30.
38. Smith DW, Tondury G. Origin of the calvaria and its sutures. Am J Dis Child
1978;132:662-6.
39. Moss ML. Inhibition and stimulation of sutural fusion in the rat calvaria. Anat Rec
1960;136:457-67.

104
40. Hobar PC, Schreiber JS, McCarthy JG, Thomas PA. The role of the dura in cranial
bone regeneration in the immature animal. Plast Reconstr Surg 1993;92:405-10.
41. Mossaz CF, Kokich VG. Redevelopment of the calvaria after partial craniectomy in
growing rabbits: the effect of altering dural continuity. Acta Anat (Basel) 1981;109:321-31.
42. Yu JC, McClintock JS, Gannon F, Gao XX, Mobasser JP, Sharawy M. Regional
differences of dura osteoinduction: squamous dura induces osteogenesis, sutural dura induces
chondrogenesis and osteogenesis. Plast Reconstr Surg 1997;100:23-31.
43. Slater BJ, Kwan MD, Gupta DM, Amasha RR, Wan DC, Longaker MT. Dissecting
the influence of regional dura mater on cranial suture biology. Plast Reconstr Surg
2008;122:77-84.
44. Li S, Quarto N, Longaker MT. Dura mater-derived FGF-2 mediates mitogenic
signaling in calvarial osteoblasts. Am J Physiol Cell Physiol 2007;293:C1834-42.
45. James AW, Xu Y, Wang R, Longaker MT. Proliferation, osteogenic differentiation,
and fgf-2 modulation of posterofrontal/sagittal suture-derived mesenchymal cells in vitro.
Plast Reconstr Surg 2008;122:53-63.
46. Guzel MZ, Yildirim AM, Yucel A, Seradjmir M, Dervisoglu S. Osteogenic potential
of infant dural grafts in different recipient beds. J Craniofac Surg 1995;6:489-93.
47. Geoffroy V, Corral DA, Zhou L, Lee B, Karsenty G. Genomic organization,
expression of the human CBFA1 gene, and evidence for an alternative splicing event
affecting protein function. Mamm Genome 1998;9:54-7.
48. Nacamuli RP, Fong KD, Warren SM, et al. Markers of osteoblast differentiation in
fusing and nonfusing cranial sutures. Plast Reconstr Surg 2003;112:1328-35.

105
49. Otto F, Thornell AP, Crompton T, et al. Cbfa1, a candidate gene for cleidocranial
dysplasia syndrome, is essential for osteoblast differentiation and bone development. Cell
1997;89:765-71.
50. Kimelman D, Kirschner M. Synergistic induction of mesoderm by FGF and TGF-beta
and the identification of an mRNA coding for FGF in the early Xenopus embryo. Cell
1987;51:869-77.
51. Globus RK, Plouet J, Gospodarowicz D. Cultured bovine bone cells synthesize basic
fibroblast growth factor and store it in their extracellular matrix. Endocrinology
1989;124:1539-47.
52. Noff D, Pitaru S, Savion N. Basic fibroblast growth factor enhances the capacity of
bone marrow cells to form bone-like nodules in vitro. FEBS Lett 1989;250:619-21.
53. Pitaru S, Kotev-Emeth S, Noff D, Kaffuler S, Savion N. Effect of basic fibroblast
growth factor on the growth and differentiation of adult stromal bone marrow cells: enhanced
development of mineralized bone-like tissue in culture. J Bone Miner Res 1993;8:919-29.
54. Tang KT, Capparelli C, Stein JL, et al. Acidic fibroblast growth factor inhibits
osteoblast differentiation in vitro: altered expression of collagenase, cell growth-related, and
mineralization-associated genes. J Cell Biochem 1996;61:152-66.
55. Moore R, Ferretti P, Copp A, Thorogood P. Blocking endogenous FGF-2 activity
prevents cranial osteogenesis. Dev Biol 2002;243:99-114.
56. Wilkie AO, Morriss-Kay GM, Jones EY, Heath JK. Functions of fibroblast growth
factors and their receptors. Curr Biol 1995;5:500-7.
57. Robin NH, Falk MJ, Haldeman-Englert CR. FGFR-Related Craniosynostosis
Syndromes. 1993.

106
58. Debiais F, Hott M, Graulet AM, Marie PJ. The effects of fibroblast growth factor-2
on human neonatal calvaria osteoblastic cells are differentiation stage specific. J Bone Miner
Res 1998;13:645-54.
59. Mooney MP, Losken HW, Siegel MI, et al. Development of a strain of rabbits with
congenital simple nonsyndromic coronal suture synostosis. Part II: Somatic and craniofacial
growth patterns. Cleft Palate Craniofac J 1994;31:8-16.
60. Shevde NK, Bendixen AC, Maruyama M, Li BL, Billmire DA. Enhanced activity of
osteoblast differentiation factor (PEBP2alphaA2/CBFa1) in affected sutural osteoblasts from
patients with nonsyndromic craniosynostosis. Cleft Palate Craniofac J 2001;38:606-14.
61. Coelho MJ, Cabral AT, Fernande MH. Human bone cell cultures in biocompatibility
testing. Part I: osteoblastic differentiation of serially passaged human bone marrow cells
cultured in alpha-MEM and in DMEM. Biomaterials 2000;21:1087-94.
62. Pradel W, Mai R, Gedrange T, Lauer G. Cell passage and composition of culture
medium effects proliferation and differentiation of human osteoblast-like cells from facial
bone. J Physiol Pharmacol 2008;59 Suppl 5:47-58.
63. Reuther T, Kettmann C, Scheer M, Kochel M, Iida S, Kubler AC. Cryopreservation
of osteoblast-like cells: viability and differentiation with replacement of fetal bovine serum in
vitro. Cells Tissues Organs 2006;183:32-40.
64. Gevorgyan A, La Scala GC, Sukhu B, et al. An in vitro model of radiation-induced
craniofacial bone growth inhibition. J Craniofac Surg 2007;18:1044-50.
65. Carmichael J, DeGraff WG, Gazdar AF, Minna JD, Mitchell JB. Evaluation of a
tetrazolium-based semiautomated colorimetric assay: assessment of chemosensitivity testing.
Cancer Res 1987;47:936-42.

107
66. VanGuilder HD, Vrana KE, Freeman WM. Twenty-five years of quantitative PCR for
gene expression analysis. Biotechniques 2008;44:619-26.
67. Spector JA, Greenwald JA, Warren SM, et al. Co-culture of osteoblasts with
immature dural cells causes an increased rate and degree of osteoblast differentiation. Plast
Reconstr Surg 2002;109:631-42; discussion 43-4.
68. Coelho MJ, Fernandes MH. Human bone cell cultures in biocompatibility testing.
Part II: effect of ascorbic acid, beta-glycerophosphate and dexamethasone on osteoblastic
differentiation. Biomaterials 2000;21:1095-102.
69. De Pollack C, Renier D, Hott M, Marie PJ. Increased bone formation and osteoblastic
cell phenotype in premature cranial suture ossification (craniosynostosis). J Bone Miner Res
1996;11:401-7.
70. Ganta DR, McCarthy MB, Gronowicz GA. Ascorbic acid alters collagen integrins in
bone culture. Endocrinology 1997;138:3606-12.
71. Hassler W, Zentner J. Radical osteoclastic craniectomy in sagittal synostosis.
Neurosurgery 1990;27:539-43.
72. Bradley JP, Han VK, Roth DA, Levine JP, McCarthy JG, Longaker MT. Increased
IGF-I and IGF-II mRNA and IGF-I peptide in fusing rat cranial sutures suggest evidence for
a paracrine role of insulin-like growth factors in suture fusion. Plast Reconstr Surg
1999;104:129-38.
73. Bradley JP, Levine JP, McCarthy JG, Longaker MT. Studies in cranial suture
biology: regional dura mater determines in vitro cranial suture fusion. Plast Reconstr Surg
1997;100:1091-9; discussion; 100-2.

108
74. Greenwald JA, Mehrara BJ, Spector JA, et al. Biomolecular mechanisms of calvarial
bone induction: immature versus mature dura mater. Plast Reconstr Surg 2000;105:1382-92.
75. Zimmerman LB, De Jesus-Escobar JM, Harland RM. The Spemann organizer signal
noggin binds and inactivates bone morphogenetic protein 4. Cell 1996;86:599-606.
76. Song HM, Fong KD, Nacamuli RP, et al. Mechanisms of murine cranial suture
patency mediated by a dominant negative transforming growth factor-beta receptor
adenovirus. Plast Reconstr Surg 2004;113:1685-97.
77. Bostrom MP, Saleh KJ, Einhorn TA. Osteoinductive growth factors in preclinical
fracture and long bone defects models. Orthop Clin North Am 1999;30:647-58.
78. Uneno S, Yamamoto I, Yamamuro T, et al. Transforming growth factor beta
modulates proliferation of osteoblastic cells: relation to its effect on receptor levels for
epidermal growth factor. J Bone Miner Res 1989;4:165-71.
79. Robey PG, Young MF, Flanders KC, et al. Osteoblasts synthesize and respond to
transforming growth factor-type beta (TGF-beta) in vitro. J Cell Biol 1987;105:457-63.
80. Lucas PA. Chemotactic response of osteoblast-like cells to transforming growth
factor beta. Bone 1989;10:459-63.
81. Harris SE, Bonewald LF, Harris MA, et al. Effects of transforming growth factor beta
on bone nodule formation and expression of bone morphogenetic protein 2, osteocalcin,
osteopontin, alkaline phosphatase, and type I collagen mRNA in long-term cultures of fetal
rat calvarial osteoblasts. J Bone Miner Res 1994;9:855-63.
82. Noda M, Rodan GA. Type-beta transforming growth factor inhibits proliferation and
expression of alkaline phosphatase in murine osteoblast-like cells. Biochem Biophys Res
Commun 1986;140:56-65.

109
83. Centrella M, McCarthy TL, Canalis E. Transforming growth factor beta is a
bifunctional regulator of replication and collagen synthesis in osteoblast-enriched cell
cultures from fetal rat bone. J Biol Chem 1987;262:2869-74.
84. Nakamura K, Kurokawa T, Kawaguchi H, et al. Stimulation of endosteal bone
formation by local intraosseous application of basic fibroblast growth factor in rats. Rev
Rhum Engl Ed 1997;64:101-5.
85. Kawaguchi H, Kurokawa T, Hanada K, et al. Stimulation of fracture repair by
recombinant human basic fibroblast growth factor in normal and streptozotocin-diabetic rats.
Endocrinology 1994;135:774-81.
86. Canalis E, Centrella M, McCarthy T. Effects of basic fibroblast growth factor on bone
formation in vitro. J Clin Invest 1988;81:1572-7.
87. Globus RK, Patterson-Buckendahl P, Gospodarowicz D. Regulation of bovine bone
cell proliferation by fibroblast growth factor and transforming growth factor beta.
Endocrinology 1988;123:98-105.

110
Appendices

111
Calvarial osteogenic cell culture
Calvarial osteogenic cell culture preparation
The bone sample is kept on ice in a 50 ml Falcon tube with αMEM and 5xpenicillin-
streptomycin until digestion. Samples are processed within 30 min after taken from the
operating room. It is then transferred to a 65-mm Petri dish (Falcon™, Becton, Dickinson
and Co., Franklin Lakes, NJ –Ref. 35002) with 6 ml collagenase mixture (See appendix pg.
126). The bone is diced with scissors into as small as possible pieces (< 0.5 mm3). The
mixture with small bone fragments is transferred to a 14 ml tube (Falcon, Ref. 35-2059) and
incubated at 37° C for 20 minutes for digestion in collagenase mixture (See appendix pg.
126). During digestion, the tube is removed from the incubator and mixed on Vortex for 30
seconds every 5 minutes. After 20 minutes of incubation, the tube is removed from the
incubator and left for the larger pieces to settle to the bottom of the tube. The upper layers of
the mixture are transferred to a new 50 ml Falcon tube, already containing 15 ml of FSGM
(See appendix pg.127). Then, 6 ml of collagenase mixture is added to tube 1, containing
small bone pieces for further digestion at 37° C for 20 minutes.
The above procedure is repeated to a total of 3 times to digest out as many cells as
possible. Cells collected from each digestion are added to the same 50 ml Falcon tube. After
the final digestion, the 50 ml Falcon tube is centrifuged at 4° C, 700 rpm (80.5× g) in IEC
Centra MP4R (International Equipment Company, Needham Heights, MA – Ref. 2438; 28-
tubes, 4 × 250 ml rotor head, Ref. 224) for 15 minutes. The supernatant is removed and
discarded. The cell pellet is diluted in 15 ml FSGM, resuspended and plated into a T75 tissue
culture flask (SARSTEDT Ref. 83.1813.002). The medium is changed every 2-3 days until
confluence.

112
Calvarial osteogenic cell subculture
At confluence, the culture medium is aspirated with a sterile Pasteur’s pipette and
discarded. The flasks are rinsed with 5 ml PBS without Ca and Mg (Wisent Bioproducts Ref.
80450) to remove any residual medium and the liquid is aspirated. 5 ml Trypsin-EDTA 1x
(Wisent Bioproducts Ref. 325042) reconstituted in PBS without Ca and Mg and the flask is
incubated at 37° for 5 minutes. Then, the flask is removed from the incubator and observed
under the phase contrast microscope to insure that all cells are detached. If any cells are
remaining attached, tap the side of the flask to detach the cells. If necessary, a cell scraper
(SARSTEDT Ref. 83.1830) is used to remove any residual cell colony attached to the plate.
Neutralize Trypsin with 5 ml of FSGM, pipetting up and down and washing the
bottom of the flask. The content of the flask is moved to a 14 ml tube (Falcon – Ref. 35-
2098) and the later is centrifuged at 800 rpm (105.2x g), 4°C for 7 minutes [relative
centrifugal force (RCF) value (g) = 1118 × 10E-8 × R × N2, where R is the radius in cm from
the centre of the rotor to the point at which the RCF value is required (for 4, 28 and 48 tubes
=14.7 cm), and N = speed of centrifuge in rpm]). The supernatant is carefully aspirated with
a sterile Pasteur’s pipette and discarded, leaving very little liquid on top of the cell pellet.
Then, precisely 5 ml of FSGM is added to the cell pellet and pipetted up and down 10-20
times to obtain single-cell suspension. If multiple primary cell cultures exist, the suspensions
are combined in a larger tube. If only a small number of cells are expected to be retrieved
from the pellet, little amount (1-2 ml) of FSGM is used.
After resuspending the pellet, 0.1-0.2 ml of the suspension is transferred to an
Eppendorf tube. Cells are counted in a haemocytometer and plated at densities of 1000 cells
per well in flat bottomed 96-well plates (SARSTEDT – Ref. 83.1835) for alkaline

113
phosphatase (AP) (including protein) and mineralization assays. Three columns, comprising
of 4 wells is used for each AP and protein assays on the same plate and 4 wells for
mineralization assay on a separate plate. Concurrently, cells are seeded into 24-well plates, 4
wells per plate for each of AP/Protein and mineralization assays. For collagen type I assay,
50,000 cells per well are seeded in a flat bottomed 6-well tissue culture plate (SARSTEDT
Cat# 83.1839).
Medium change in 96-well plates
Medium (α-MEM with 15% FCS, 10-7 Dexamethasone) was changed on Monday,
Wednesday and Friday. The old medium was removed with a 20G (at later days, especially
with folding, 25-26G) sterile needle connected to a wall suction system, suctioning the fluid
form the edge of the left lower quadrant of the culture well. A new needle was used for each
plate, in order to reduce the risk of contamination. Using a multichannel (8) pipette, 100 μl
(150 μl for the weekend) of medium was delivered to each of the wells in the 96-well plates,
changing the tips between plates, depending on the medium change interval and cell
concentration. For the 6-well plates, 2 ml (3 ml for the weekend) of medium was added. At
around day 7 (1 week in subculture), prior to medium change, vitamin C (Ascorbic acid –
Sigma/Aldrich Cat# A2218), 0.33 μl stock solution (50 μg) / 1 ml of medium, necessary for
collagen synthesis, and β-glycerophosphate 1% (Sigma – Ref. G6251) were added to
promote mineralization.

114
Medium Composition Study
Cells were subcultured by trypsinization (5 mL Trypsin-EDTA) and neutralized in 5 mL
of FSGM to obtain single-cell suspension that was transferred to an Eppendorf tube17
.
Subsequently, cells were counted in a haemocytometer and plated at densities of 1000 cells
per 96-well culture plates. On day 3, 5 and 7 of culture, old medium (αMEM with 15% FBS)
was removed and 200 μl of either, (αMEM containing 10-7
M dexamethasone, supplemented
with 15%, 10%, 1% of FBS, or an osteogenic medium supplemented with ascorbic acid and
β-glycophosphate was added to each well to generate four culture medium conditions. Cells
were incubated at 37°C and the medium was changed every 2-3 days.
Preparation of the periosteum-derived osteoblast cell culture
The periosteum was kept in α-MEM + RNA + DNA + 1× antibiotic on ice until
digestion and then transfered to 3 ml collagenase mixture (for composition, see Materials pg.
120) in a 60-mm Petri dish (Falcon – Ref. 4-1007-0). The periosteum was diced with scissors
into small pieces (< 1 mm3) and transfered to a 14 ml tube (Falcon – Ref. 35- 2059) and
incubated at 37° C for 15'. 5 ml of α-MEM + RNA + DNA + 1× antibiotic (penicillin-
streptomycin) + 15 % FBS + 10-7 M Dexamethasone (FS-GM) were added and mixed to
allow the larger pieces to settle to the bottom of the tube (Tube 1) The upper layers of the
mixture were transferred to a new tube, centrifugated at 4° C and 700 rpm (80.5× g) in IEC
Centra MP4R (International Equipment Company, Needham Heights, MA – Ref. 2438; 28-
tubes, 4 × 250 ml rotor head, Ref. 224) for 8' and then removed and discarded the supernatant
containing collagenase. The cell pellet was diluted with 5 ml FS-GM and seeded into T25
flask with 0.2 μm vented plug cap and incubated at 37° C. 3 ml of collagenase was added to

115
tube 1, containing small periosteal pieces for further digestion at 37° C for 15'. After the
second “redo” 5 ml FS-GM and was added to the tube and centrifuged at 4° C and 700 rpm
(80.5× g) for 8'. After removing and discarding the supernatant containing collagenase, the
pellet was diluted in 5 ml FS-GM, and plated. Medium was changed every 2-3 days until
confluence.
Subculture of the periosteum derived cell culture
The culture medium was aspirated with sterile Pasteur’s pipette and discarded from
flasks. Cells were washed with 5 ml PBS without Ca, Mg (Wisent Bioproducts – Ref
311011) to remove any residual medium.PBS is then removed with sterile Pasteur’s pipette.
3-5 ml Trypsin-EDTA 1x was added (Wisent Bioproducts – Ref. 425042), reconstituted in
PBS without Calcium and Magnesium), and incubate at 37° for 5 minutes. After removal
from the incubator flasks were tapped at the side to detach the cells and a cell scraper
(SARSTEDT Ref. 83.1830) was used to remove any residual cell colony attached to the
plate. Trypsin was neutralized with 5 ml of fully supplemented growth medium (FSGM),
pipetting up and down, and washing the bottom of the flask. All the content was aspirated
and placed into a 50-ml tube (Falcon – Ref. 35-2098) and then centrifugated at 800 rpm
(105.2x g), 4°C for 7 minutes [relative centrifugal force (RCF) value (g) = 1118 × 10E-8 × R
× N2, where R is the radius in cm from the centre of the rotor to the point at which the RCF
value is required (for 4, 28 and 48 tubes =14.7 cm), and N = speed of centrifuge in rpm]).
With a sterile Pasteur’s pipette the supernatant was carefully aspirated and discarded, leaving
some liquid on top of the cell pellet.

116
Exactly 5 ml of FSGM were added to the pellet and pipette up and down 10-20 times to
obtain single cell suspension. If multiple plates exist from the same specimen, the suspension
was transferred to the next pellet-containing tube. If only a small number of cells was
expected to be retrieved from the pellet, less than 5 ml of FSGM (1 or 2 ml) was used. After
the last pellet has been suspended, cells were counting using 0.1-0.2 ml of the liquid.
• Cells were counted as previously described and plated at densities of 500 (mineralization),
and 1,000 per well in a flat bottomed 96-well tissue culture plate (SARSDETD – Ref.
83.1835), 8 wells per dilution) and 50,000 per well in a flat bottomed 6-well tissue culture
plate (Falcon – Ref. 35-3046) for collagen assay.
MTT Assay
3-[4,5-Dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide; Thiazolyl blue – kept in the
freege (4C) The MTT test is a simple, accurate, reproducible means for measuring the
activity of living cells via mitochondrial dehydrogenase activity (Mosmann, T. Rapid
colorimetric assay for cellular growth and survival: application to proliferation and
cytotoxicity assays). The reactant will form formazan crystals in living cells mitochondria
when incubated at 37° for 3-5 hours, and this will be released in solution by the DMSO.
Formazan solution absorbs light at 550-570 nm but not at 620-650 (690) nm, which
absorbance results from cell debris & well imperfections. Final optical density (OD) obtained
from formazan formation can be calculated OD=L1-L2 or automatically by the μQUANT
spectrophotometer.
Reagent
• 50mg of MTT (Sigma – Ref. M2128) dissolved in 10 ml PBS and filtered.
Technique

117
30 μl off the mixture were added to each well (analysis in triplicate), without removing the
medium and then incubated at 37°C for 5 hours. Wells contents were aspirated and 100 μl of
DMSO (Sigma – Ref. D-8779) were added to wells + one “blank” well, left for 5 minutes
and shaked (cells-containing wells produce a purple color).
Reading
In a spectrophotometer, results were read at @ 570nm (primary), with reference
wavelength of 690nm.
Alkaline phosphatase activity, current technique: p-Nitrophenyl
Phosphate (expressed per amount of protein per time of incubation of AP assay)
Reactant
Paranitrophenyl Phosphate (pNP, Sigma – Ref. 104-0) (kept in the -200C freezer) 9
mg/ml was dissolved in 10 ml of Tris buffer at pH=9.0, mix well.
Technique
Medium was removed from wells (analysis in triplicate). Wells were flushed twice
with 100 μl PBS and 200 μl of the reactant solution were added to each well + 1 for “blank”
measurement. Cells were incubated at 37°C, checking for color changes (when many cells,
significant color change in 1 hour, otherwise a few hours may be necessary). Results were
read on Titertek Multiskan MCC/340 MK II (Titertek, Huntsville, AB) at Filter 2 (@
405nm).

118
Protein absorbance
Medium was removed from wells (analysis in duplicate). Wells were flushed twice
with 100 μl NaHCO3 (Sigma – Ref. S-6014) and 100 μl of Bio-Rad protein assay reagent
(Bio-Rad Laboratories, Hercules, CA – Ref. 500-0006) diluted 1:5 in NaHCO3 at pH 7.4
were added to sample wells + to 1 “blank” well. After 10 minutes absorption was measured
on Titertek: Same procedure as above, use Filter 7 (@ 620nm)
Calculation of real values
APA: slope: 324.4 intercept: - 1.383 real value = Abs × 324.4 – 1.383
Proteins: slope: 246 intercept: 0.671 real value = Abs × 246 + 0.671
Final AP concentration is expressed as mmol of pNP per hour per μg of protein.
Alkaline phosphatase staining: Fast Blue BB Salt
Reactant
Naphtol was dissolved as phosphate (Sigma – Ref. N-5625), 3 mg in 50 μl
dimethylformammide (Sigma – Ref. D-4254) and then again in 10 ml of Tris buffer 0.2 M at
pH = 9.0 (Sigma-Fluka – Ref. 98306). 20 mg of Fast Blue BB salt (Sigma – Ref. F-3378)
was added. Wells were shake and left at room temperature for 10 minutes. As soon as the
colour changes to yellow-brownish, it was filtered using non-sterile syringe filter to get rid of
particulate matter.
Technique
100 μl of the reactant solution was added to each well and placed in the incubator at
37°C. Assessment for possible color change was done q 5 minutes (cells stained in dark blue
are considered AP positive cells [osteoblast-like cells]). For 6-well plates, 1 ml was added

119
(just to cover the cell surface) of the reactant solution to each well, placed in the incubator at
37°C, and analyzed similarly.
Alizarin Red Staining for mineralization in 96 well plates
Medium was removed and 100 µl of formalin (Sigma – Ref. HT-50-1-320) added and
left for 5 minutes. Formalin was then removed and washed twice with 150 μl of
distilled water. 50 μl of Alizarin Red S (Cat. No. A5533, Sigma Chemical Co., St.
Louis, MO) 2% staining solution were added and left for 5 minutes (added also to
“blank” well).The staining solution was removed from samples and blank and washed
twice with 150 μl of distilled water. Results were read @ 525nm on Titertek
Multiskan MCC/340 MK II.
Transmission Electron Microscopy
Electron microscopy was used to analyze bone-specific ultrastructure in vitro. Cells were
grown in 24-well plates up to day 28 in subculture, then fixed using 2% glutaraldehyde in 0.1
mol/L sodium cacodylate buffer, and postfixed in 1% osmium tetroxide in 0.1 mol/L sodium
cacodylate buffer. The samples were dehydrated through a graded ethanol series, followed by
propylene oxide, and embedded in Quetol-Spurr resin. Sections 100nm thick were cut on an
RMC MT6000 ultramicrotome, stained with uranyl acetate and lead citrate and viewed in an
FEI Tecnai 20 TEM.

120
HISTOLOGY
The bone samples were fixed in 10% phosphate buffered formalin for 48 hours at 4°C and
then transferred to Bouin’s fixative for 48 hours at 4°C, washed in distilled water, decalcified
in acetic acid/4% formaldehyde/0.85% saline, dehydrated in graded ethanol, and embedded
in paraffin. Serial sections were cut for routine hematoxylin and eosin staining to define areas
of complete ossification, fibrous tissue and remodeling bone.
IMMUNOHISTOCHEMISTRY AND IMMUNOFLUORESENCE
The samples were fixed in 3-4% paraformaldehyde in PBS pH 7.4 for 15 min at room
temperature and then washed twice with ice cold PBS. Samples were incubated for 10 min
with PBS containing 0.25% Triton X-100. Triton X-100 is the most popular detergent for
improving the penetration of the antibody. However, it is not appropriate for the use of
membrane-associated antigens since it destroys membranes. Cells were washed in PBS three
times for 5 min. Cells were then incubated with 1% BSA in PBST for 30 min to block
unspecific binding of the antibodies Following, the same cells were incubated in the diluted
antibody in 1% BSA in PBST in a humidified chamber for 1 hr at room temperature or
overnight at 4°C. The solution was decanted and cells washed three times in PBS, 5 min each
wash. Cells were then incubated with the secondary antibody in 1% BSA for 1 hr at room
temperature in dark. The secondary antibody solution was also decanted and cells washed
three times with PBS for 5 min each in dark. Cells were then incubated on 0.1-1 μg/ml
Hoechst or DAPI (DNA stain) for 1 min and rinsed with PBS. The coverslip was mounted
with a drop of mounting medium and sealed with nail polish to prevent drying and movement
under microscope. Slides were stored in dark at -20 or 4°C.

121
Formalin-fixed, paraffin-embedded tissue sections (5µm) were mounted on positively
charged microscope slides. Tissue samples were then deparaffinized in xylene, 3% H2O2
was used to clear endogenous pigment. Antigen retrieval was then performed with pepsin at
37°C and tissue sections were blocked with 0.5% BSA with casein. Rabbit polyclonal to
Noggin (Abcam) was used in dilution 1:20, mouse monoclonal to Collagen I (Abcam) was
used in dilution 1:400 or rabbit polyclonal to BMP3 (R&D System) was used in dilution 1:5.
Detection was performed with Goat Polymer (Biocaremedical) for BMP3 and ABC Ellite
System (Vector) for Noggin and Collagen I. DAB was used as chromogen.
WESTERN BLOT
Cells were subcultured into 6 well plates to confluence, 500 µl of medium was taken
from each well and centrifuged at 2000 RPM for 10 min. 300 µl was taken and placed
in a new 1.5 ml tube and added 100 µl of 4x sample buffer, boiled at 100°C for 5 min.
The 200 µl were left for protein assay. Residual medium was discarded. 500 µl of
lyses buffer was added to each well, cells were collected with a 3ml Syringe and 25G
needle to small tubes, centrifuged at 14,000 RPM for 6 minutes and transferred to
new tubes. 120 µl of 4x sample buffer was added and samples were boiled at 100°C
for 5 min.
SDS
Resolving (8%) and stacking gels (4%) were prepared just before pouring the gel
between the glasses. Resolving gel (8 ml for mini gel) was poured and the upper surface was
quickly covered with water, and let polymerize for about 30min. Water on top was
eliminated, and then stacking gel (1.5 ml) was poured on top of the resolving gel. Combs
were inserted avoiding bubbles and let polymerize for about 15min. The comb was taken

122
away and inserted the glasses in the Western blotting apparatus. Running buffer was added.
25 or 50 l samples supernatants and cell pellets (cell lysates: loading buffer +1:1)
previously boiled for 2 min were loaded. Proteins were separated by electrophoresis 2 h at
100V. At the end of the electrophoresis gel was directly transferred onto PVDF membrane
which was treated with MeOH for 1 min. The cassette for transferring was prepared in the
following order: black side, sponge, filter paper, gel, PVDF membrane, filter paper, sponge
and clear side. Blotting tank was placed in the cold room overnight at 36V (100mA). After
transfer, the membrane was washing once with 1X TBST and kept in 4oC.
Western blotting
The membrane was treated with MeOH for 1 min then washed with 1X TBST. The
membrane was then blocked with 4% BSA or 5% milk in 1X TBST for 1 hour at room
temperature and then washed 2X with 1X TBST. The membrane was incubated with the first
antibody in 1X TBST for 1h and washed 3 X 5min with 1X TBST. Next, the membrane was
incubated with the second antibody in 1X TBST for 1 hour at room temperature and washed
6X 5min with 1X TBST. Detection was done with Western Lightening Chemiluminescence
Reagent.
The membrane was stripped with stripping buffer and blocked with 4% BSA in
1XTBST overnight at 4oC and then washed the membrane with 1XTBST buffer. The
membrane was incubated with anti-β-actin* (1:3000) dilution in 4% BSA-TBST for 1 hour at
room temperature (*anti-β-actin, Sigma, cat# A5441, arise in Mouse, MW 47KDa) and
washed 3 X 5 min with 1X TBST. Then it was incubated with horseradish peroxidase

123
conjugate anti-rabbit IgG (1:10,000) in 1X TBST for 1h and washed 5 X 5min with 1X
TBST. Detection was done with Western Lightening Chemiluminescence Reagent.
SDS PAGE Gels
Resolving Gel (35ml)
5% 6% 7% 8% 10% 12% 15% 20%
40%
Acrylamide
4.38 5.25 4.38 5.25 4.38 5.25 4.38 5.25
1.5M Tris,
pH 8.8
8.75 8.75 8.75 8.75 8.75 8.75 8.75 8.75
1% APS 2.5 2.5 2.5 2.5 2.5 2.5 2.5 2.5
dH2O 19.4 18.5 19.4 18.5 19.4 18.5 19.4 18.5
*Add 25l of TEMED
Stacking Gel (8ml)
5% 4% 3%
40% Acrylamide 1 0.8 0.6
0.5M Tris, pH6.8 2 2 2
1% APS 1 1 1
dH2O 4 4.2 4.4
*Add 8l of TEMED

124
10X TBST (1L)
100ml 1M Tris pH 7.4 (121.14g Tris= 1L dH2O)
87.7g NaCl
5ml Tween 20
Adjust volume to 1L with dH2O
1X Transfer buffer
Tris 6.06g
Glycine 28.82g
Methanol 200ml
Adjust volume to 2L with dH2O
REAL TIME PCR
Cells were growth for 10 days. After remove the medium from wells incubate cells
with TRIZOL at 15-30° C for 5 minutes. 200 µl of chloroform were added per 1 ml of
TRIZOL used. Tubes were vortex for 15 seconds and then incubate at 15-30° C for
10-15 minutes with vortex half way through. Samples were centrifuge at 12,000 PRM
for 15 minutes at 2-8°C (cold room). For RNA precipitation the aqueous phase was
transferred to an eppendorf tube carefully, using a P-200 pipettor. Lower, DNA phase
was frozen -80°C. 500µl of Isopropyl Alcohol was added, mixed gently, and
incubated at room temperature for 10 minutes. Tubes were centrifuged at 12,000 X g
for 10 minutes at 4°C. The RNA precipitate formed a gel-like pellet at the bottom of
the tube. The supernatant was carefully pulled off with a P-200. The cell pellet was

125
washed once with 1ml of 75% Ethanol (ICE Cold) and centrifuge at 12,000 x g for 15
minutes at 4°C. Again, the supernatant was carefully pulled off with a P-200. The
pellet was air dried completely so there was no residual ethanol. RNA was dissolved
in Rnase- free water (DEPC WATER), volume dependent on number of cells, or size
of pellet (50µl). RNA concentration was checked by spec.
Reverse transcription was carried out with the SuperScript First-Strand
Synthesis System for RT-PC. The following RNA/primer mixture was prepared in
each tube: Total RNA 5µg, Random hexamers (50ng/µ) 3µl, 10 mM dNTO mix 1µl
and DEPC water to 10µl. Samples were incubated at 65°C for 5 min and then on iced
for at least 1 min. The reaction master mixture (10x TR buffer 2µl, 25 mM MgCl2
4µl, 0.1 M DTT 2µl and RNAase OUT 1µl) was added to the RNA/primer mixture,
mixed briefly, and then placed at room temperature for 2 min. Then 1µl (50 units) of
SuperScript II RT was added to each tube, mixed and incubated at 25°C for 10 min.
The tubes were then incubated at 42°C for 50 min, heat inactivated at 70°C for 15
min, and then chilled on ice. 1 µl RNase H was added and incubatet at 37°C for 20
min. The 1st strand cDNA was stored at -20°C until use for reat-time PCR.
The primer concentrations were normalized and mixed-specific forward and
reverse primers pair. Each primer concentration in the mixture was 5ρmol/µl. The
following mixture was prepared for each optical tube: 25 or 12.5 µl SYBR Green Mix
(2x), 0.5 or 0.2 cDNA, 2 or 1 µl primer mix pair (5ρmol/µl each primer), 22.5 or 11.3
µl water. The experiment and the following PCR program were set on ABI Prism
SDS 7000 according to the primers used: 94°C for 90 sec, 60°C for 30 sec and 72°C
for 30 sec – 40 cycles. After PCR was finished, the PCR specificity was examined by

126
3% agarose gel using 25 µl from each reation. The real-time PCR results were
analyzed with a SDS 7000 software.
MATERIALS
Collagenase Incubation Mixture
Substance Supplier 200 ml 300 ml 400 ml Storage
Collagenase Sigma C 0130 or Sigma C9891 0.6 g 0.9 g 1.2 g -20°C
D-sorbitol Fisher S 459 or Sigma S-3889 (500 g) 3.644 g 5.466 g 7.288 g Shelf
(room t°)
Chondroitin
Sulphate
Fluka 27043 or Sigma C-4384 1.2 g 1.8 g 2.4 g +4°C
DNAse Sigma D 4513 0.8 ml
1.2 ml
1.6 ml
-20°C
The above chemicals were combined and added the corresponding amount of Krebs’ II
buffer with Zn2+. Stored at – 20° C.
Krebs’ II A buffer with Zn2+
Substance Concentration 1 litre 2 litres
NaCl 111.2 mM 6.4965 g 12.993 g
Tris Buffer (Base) 21.3 mM 2.5802 g 5.1604 g
Glucose 13.0 mM 2.3421 g 4.6842 g
KCl 5.4 mM 0.4026 g 0.8052 g

127
MgCl2 1.3 mM 0.2643 g 0.5286 g
ZnCl2 0.5 mM 0.0682 g 0.1364 g
The first five chemicals were combined in 900 or 1800 ml of distilled water, pH adjusted to
7.4. ZnCl2 was added in and made up the volume in a 1 or 2 litre volumetric flask. The
solution was filtered sterile and stored at 4° C.
Mediums
Complete medium (15% FCS, Dexamethasone 10E-7
) from α-MEM:
α-MEM volume was divided by 0.85 to obtain the volume of the final solution, the initial α-
MEM volume was subtracted to obtain FCS volume.
(i.e.: α-MEM 500 ml: 500/0.85 = 588 ml; subtract 500 = 88 ml (vol. of FCS to add)).
Dexamethasone is provided in a 10E-3
solution: to obtain 10E-7
: (10E-7
* Volume in ml) /
10E-3
(i.e. (10E-7
* 588) / 10E-3
= 0.0588 ml [58.8 μl] of Dexamethasone 10E-3
solution)
FSGM (1% β-Glycerophosphate 15% FCS, Dexamethasone 10E-7
) from α-MEM:
Alpha-MEM volume was divided by 0.84 to obtain the volume of the final solution, divided
by 100 to obtain the volume of β-GP, subtracted this + the initial α-MEM volume to obtain
FCS volume. (i.e.: α-MEM 500 ml: 500/0.84 = 595 ml; subtract 500 + 6 (vol β-GP) = 89 ml
(vol. of FCS to add).Vitamin C (Ascorbic Acid) stock solution, 0.33 μl (50 μg) / 1 ml
medium only at the time of use.

128