in vitro and in vivo haemolytic studies of tentacle-only extract from jellyfish cyanea capillata
TRANSCRIPT

Toxicology in Vitro 24 (2010) 1203–1207
Contents lists available at ScienceDirect
Toxicology in Vitro
journal homepage: www.elsevier .com/locate / toxinvi t
In vitro and in vivo haemolytic studies of tentacle-only extract from jellyfishCyanea capillata
Liang Xiao a,1, Jing Zhang b,1, Qian-qian Wang a,1, Qian He c, Si-hua Liu a, Yue Li a, Li-ming Zhang a,*
a Department of Chemical Defense Medicine, Faculty of Naval Medicine, Second Military Medical University, Shanghai 200433, Chinab Department of Pharmacology, School of Pharmacy, Second Military Medical University, Chinac Department of Gynecology and Obstetrics, Changhai Hospital, Second Military Medical University, China
a r t i c l e i n f o a b s t r a c t
Article history:Received 4 September 2009Accepted 5 February 2010Available online 10 February 2010
Keywords:JellyfishCyanea capillataTentacle-only extractHaemolyticIn vitroIn vivo
0887-2333/$ - see front matter � 2010 Elsevier Ltd. Adoi:10.1016/j.tiv.2010.02.009
* Corresponding author. Tel./fax: +86 21 8187 1128E-mail address: [email protected] (L.-m
1 Co-first authors.
To approach the real haemolytic process of jellyfish toxins, both in vitro and in vivo haemolysis of tenta-cle-only extract (TOE) from jellyfish Cyanea capillata has been studied. Dose–response curves of the hae-molytic activity of TOE in vitro were sigmoid shaped in both erythrocyte suspension and diluted wholeblood, with the former more sensitive to TOE. The in vivo haemolysis increased sharply in the first10 min and was followed by a gradual increase in the following 3 h, with increasing blood potassiumand lactic acid accordingly. SC5b-9 complexes were significantly up-regulated in vitro, but not in vivo.These results showed that the haemolysis of TOE in diluted whole blood and in vivo is not totally consis-tent with that in the erythrocyte suspension, and blood plasma might play a protective role againsthaemolysis. Thus we suggested that erythrocyte suspension can be used to test the damage of toxinon erythrocyte membrane, while the diluted whole blood may be more suitable to test the haemolysisof toxins.
� 2010 Elsevier Ltd. All rights reserved.
1. Introduction
Amongst Scyhozoa, Cyanea capillata is an off-shore jellyfish, alsocalled the ‘‘Sea Blubber”, the ‘‘Hair Stinger” or ‘‘Snottie”. C. capillatahas a worldwide distribution and is a major kind of jellyfish insoutheast Chinese coastal waters. There are no reported humanfatalities caused by C. capillata envenomation. However, contactwith the tentacles may produce a burning feeling, severe pain,swelling, red streak, nausea and abdominal pains. At times, profusesweating, muscle cramp and respiratory distress are seen (Tibballs,2006). Recently, the number of C. capillata has increased dramati-cally in east Chinese coastal waters and the number of patientsstung by this kind of jellyfish has increased correspondingly. Crudeor partially purified C. capillata nematocyst venoms have demon-strated a wide spectrum of biological activities, such as haemolytic,enzymatic, cardiovascular activities and so on in a few in vivo andin vitro studies (Rice and Powell, 1972; Walker, 1977a,b; Walkeret al., 1977; Long and Burnett, 1989; Helmholz et al., 2007).
Haemolytic activity of jellyfish toxin has been widely reported(Edwards et al., 2000; Ishikawa et al., 2004; Bailey et al., 2005;Marino et al., 2008). Almost all the previous studies utilized eryth-rocyte suspension as the exclusive test system for haemolysis of
ll rights reserved.
.. Zhang).
venoms from various specimens of jellyfish in vitro, but this proce-dure may not be consistent with the actual haemolytic processeither in vitro or in vivo because of the lack of blood plasma, whichmay interfere with haemolytic interactions. To approach the realhaemolytic process of jellyfish toxin in vivo or in whole blood,we compared the in vitro haemolysis of tentacle-only extract(TOE) using both erythrocyte suspension and diluted whole bloodwith the same erythrocyte concentration, and then studied thein vivo haemolysis by directly injecting (i.v.) TOE in the anaesthe-tized Sprague Dawley (SD) rats. In addition, as a potential influenc-ing factor, complement activity was also analyzed both in vitro andin vivo.
2. Materials and methods
2.1. TOE preparation from jellyfish C. capillata
Specimens of C. capillata were collected in June 2008 on theSanmenwan coast of the East China Sea in Zhejiang province,China, and identified by Professor Hui-xin Hong from the FisheriesCollege of Jimei University (Xiamen, China). The removed tentacleswere preserved in plastic bags on dry ice and immediately shippedto Shanghai, where the samples were frozen at �70 �C until use.The TOE devoid of nematocysts was prepared following the meth-od described by us before (Xiao et al., 2009). Briefly, frozen tenta-cles were thawed at 4 �C and immersed in seawater (prepared in

1204 L. Xiao et al. / Toxicology in Vitro 24 (2010) 1203–1207
the laboratory by solving 28 g of NaCl, 0.8 g of MgCl2�6H2O, and1.033 g of CaCl2 in 1000 ml water) at the mass and volume ratioof 1:1 to allow autolysis of the tissues for 4 days. The mixturewas stirred for 30 min twice daily. The autolyzed mixture was fil-tered by a 100 mesh cell strainer thrice and the filtrate was centri-fuged at 10,000g for 15 min thrice. The resultant supernatant is theTOE. All the procedures were performed at 4 �C or in ice bath. Be-fore being used for injection, the TOE was centrifuged at 10,000gfor 15 min to remove the sediments, followed by dialysis in phos-phate buffer saline (PBS, 0.01 mol/L, pH 7.4) for over 8 h. The pro-tein concentration in the preparations was determined using themethod of Bradford (1976), with fetal bovine serum as the control.
2.2. Haemolytic assay in vitro
Arterial blood was drawn by a syringe through a catheter whichwas inserted into the left femoral artery of anaesthetized male SDrats (220–240 g, provided by the Laboratory Animal Center of theSecond Military Medical University, Shanghai). Rat whole bloodwas diluted in PBS to the final v/v of 1%, and erythrocytes werecentrifuged from the heparinized blood, washed three times withPBS and resuspended in the same buffer to a final erythrocyte con-centration of 0.45%, approximately the same as 1% rat whole blood.Aliquots of both 1% rat blood and 0.45% erythrocyte suspensionwere incubated with TOE at 37 �C for 30 min and then centrifugedat 3,000g for 10 min to precipitate both intact erythrocytes andghosts. An aliquot of the supernatant was then taken and the opti-cal density (OD) was spectrophotometrically measured at 414 nmto determine the percentage of hemoglobin released from the lysederythrocytes. The haemolytic activity of TOE was expressed as %absorbance compared with that observed after maximal lysis un-der saponin (25 lg/ml). The supernatant of untreated 1% rat bloodor 0.45% erythrocyte suspension was taken as the background andsubtracted. All the animal experiments were approved by the Eth-ics Committee of the Second Military Medical University.
2.3. Haemolytic assay in vivo
Male SD rats (220–240 g, provided by the Laboratory AnimalCenter of the Second Military Medical University, Shanghai) wereanaesthetized with urethane (1.2 g/kg i.p.). A heparinized catheterwas inserted into the external jugular vein for intravenous admin-istration of TOE (5 mg/kg i.v.). Arterial blood was drawn throughanother heparinized catheter inserted into the left femoral arteryat preinjection (0 min), 10, 20, 30, 60 and 180 min after TOEadministration, respectively. Similarly, the arterial blood drawnat the corresponding time point from anaesthetized SD rats aftertreatment with PBS was taken as the control. Aliquots of bloodsamples were centrifuged at 3000g for 10 min to precipitate botherythrocytes and ghosts. The supernatant was then taken and theOD was spectrophotometrically measured at 414 nm as the markerof haemolytic activity in vivo.
2.4. Blood potassium and lactic acid
Surgical preparation of SD rats and TOE administration wereperformed as above in Section 2.3. Arterial blood sample was
Table 1Determination of erythrocyte osmotic fragility by TOE from jellyfish C. capillata (%). After Tfragilities increased (n = 6).
Time (min) 0 5 10 20
Maximal fragility 0.50 0.50 0.55 0.55Minimal fragility 0.35 0.35 0.35 0.35
drawn through a catheter which was inserted into the left femoralartery by a heparinized syringe; afterwards the syringe was sealedquickly with a rubber stopper. Arterial blood potassium and lacticacid were checked within 30 min by an arterial blood gas analyzer(GEM Premier 3000, International Laboratory, USA). Measurementswere divided into six groups at preinjection (0 min), 5, 10, 20, 60and 180 min after TOE administration (5 mg/kg i.v.).
2.5. Erythrocyte osmotic fragility
Surgical preparation of SD rats and TOE administration wereperformed as above in Section 2.3. Femoral arterial blood wasdrawn at preinjection (0 min), 5, 10, 20, 30, 60, 90, 120, 150 and180 min after TOE administration, and then dropped in the aliquotsof different hypotonic NaCl solutions (0.7%, 0.65%, 0.6%, 0.55%, 0.5%,0.45%, 0.4%, 0.35% and 0.3%, respectively). After 10 min reaction,the rat erythrocyte osmotic fragility and haemolysis were esti-mated according to the color and clarity of the solutions. The max-imal erythrocyte osmotic fragility is defined as the concentration ofthe hypotonic NaCl solution with beginning haemolysis and theminimal erythrocyte osmotic fragility is defined as the concentra-tion of the hypotonic NaCl solution with complete haemolysis.
2.6. Transmission electron microscopy
Surgical preparation of SD rats was performed as above in Sec-tion 2.3. Femoral arterial blood was drawn and fixed in 4% parafor-maldehyde after TOE administration (5 mg/kg i.v.) for 30 min,rinsed twice in 0.1 mol/L sodium phosphate, followed by 3% glutar-aldehyde for 6 h. Then the specimens were fixed by 1% osmiumtetroxide for 20 h, rinsed twice and followed by dehydration andembedment. The sections (100–120 nm) were cut and stained withlead citrate and uranyl acetate. Microscopic examination was per-formed with a transmission electron microscope (Hitachi H-800) atthe Electron Microscope Laboratory of the Second Military MedicalUniversity.
2.7. SC5b-9 assay
Enzyme Immunoassay (EIA) of SC5b-9 for the quantitation ofSC5b-9 neoantigens in rat serum was measured utilizing a micro-assay plate coated with the goat monoclonal IgG specific for therat SC5b-9 neoantigens. Briefly, either mixed rat serum/dilutedTOE in vitro or serum obtained from the rat arterial blood afterTOE administration (5 mg/kg i.v.) in vivo was incubated with bio-tinylated SC5b-9 in microtitration wells, which have been coatedwith another SC5b-9 of defined and unique epitope specificity.After incubation and washing, the wells were treated with strepta-vidin labeled with enzyme horseradish peroxidase (HRP). The wellswere again washed; a TMB substrate solution was added to thewells and color developed in proportion to the amount of SC5b-9bound. The stop solution changed the color from blue to yellow,and the intensity of the color was measured at 450 nm. SC5b-9neoantigens were obtained using the standard curve with markersprovided in the USCNLIFE™ SC5b-9 complete kit (USCNLIFE™, Wu-han, China).
OE administration (5 mg/kg i.v.), both the maximal and minimal erythrocyte osmotic
30 60 90 120 150 180
0.55 0.55 0.60 0.65 0.65 0.650.35 0.35 0.35 0.40 0.40 0.40
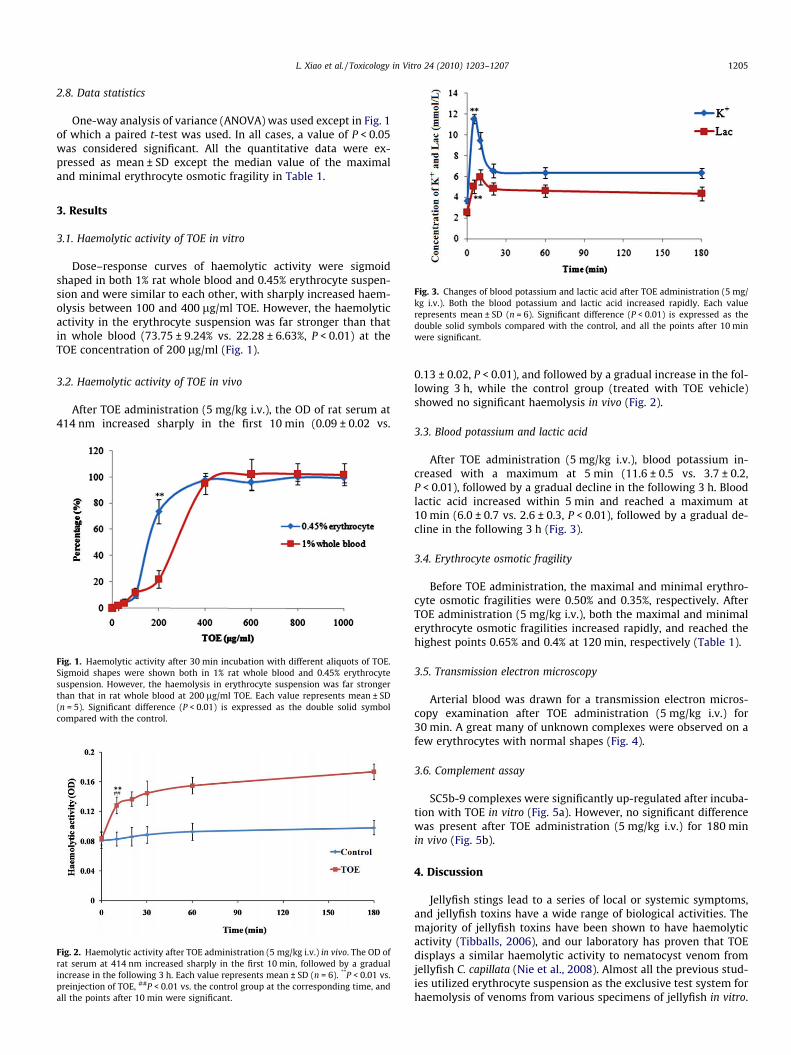
L. Xiao et al. / Toxicology in Vitro 24 (2010) 1203–1207 1205
2.8. Data statistics
One-way analysis of variance (ANOVA) was used except in Fig. 1of which a paired t-test was used. In all cases, a value of P < 0.05was considered significant. All the quantitative data were ex-pressed as mean ± SD except the median value of the maximaland minimal erythrocyte osmotic fragility in Table 1.
Fig. 3. Changes of blood potassium and lactic acid after TOE administration (5 mg/kg i.v.). Both the blood potassium and lactic acid increased rapidly. Each valuerepresents mean ± SD (n = 6). Significant difference (P < 0.01) is expressed as thedouble solid symbols compared with the control, and all the points after 10 minwere significant.
3. Results
3.1. Haemolytic activity of TOE in vitro
Dose–response curves of haemolytic activity were sigmoidshaped in both 1% rat whole blood and 0.45% erythrocyte suspen-sion and were similar to each other, with sharply increased haem-olysis between 100 and 400 lg/ml TOE. However, the haemolyticactivity in the erythrocyte suspension was far stronger than thatin whole blood (73.75 ± 9.24% vs. 22.28 ± 6.63%, P < 0.01) at theTOE concentration of 200 lg/ml (Fig. 1).
3.2. Haemolytic activity of TOE in vivo
After TOE administration (5 mg/kg i.v.), the OD of rat serum at414 nm increased sharply in the first 10 min (0.09 ± 0.02 vs.
Fig. 1. Haemolytic activity after 30 min incubation with different aliquots of TOE.Sigmoid shapes were shown both in 1% rat whole blood and 0.45% erythrocytesuspension. However, the haemolysis in erythrocyte suspension was far strongerthan that in rat whole blood at 200 lg/ml TOE. Each value represents mean ± SD(n = 5). Significant difference (P < 0.01) is expressed as the double solid symbolcompared with the control.
Fig. 2. Haemolytic activity after TOE administration (5 mg/kg i.v.) in vivo. The OD ofrat serum at 414 nm increased sharply in the first 10 min, followed by a gradualincrease in the following 3 h. Each value represents mean ± SD (n = 6). **P < 0.01 vs.preinjection of TOE, ##P < 0.01 vs. the control group at the corresponding time, andall the points after 10 min were significant.
0.13 ± 0.02, P < 0.01), and followed by a gradual increase in the fol-lowing 3 h, while the control group (treated with TOE vehicle)showed no significant haemolysis in vivo (Fig. 2).
3.3. Blood potassium and lactic acid
After TOE administration (5 mg/kg i.v.), blood potassium in-creased with a maximum at 5 min (11.6 ± 0.5 vs. 3.7 ± 0.2,P < 0.01), followed by a gradual decline in the following 3 h. Bloodlactic acid increased within 5 min and reached a maximum at10 min (6.0 ± 0.7 vs. 2.6 ± 0.3, P < 0.01), followed by a gradual de-cline in the following 3 h (Fig. 3).
3.4. Erythrocyte osmotic fragility
Before TOE administration, the maximal and minimal erythro-cyte osmotic fragilities were 0.50% and 0.35%, respectively. AfterTOE administration (5 mg/kg i.v.), both the maximal and minimalerythrocyte osmotic fragilities increased rapidly, and reached thehighest points 0.65% and 0.4% at 120 min, respectively (Table 1).
3.5. Transmission electron microscopy
Arterial blood was drawn for a transmission electron micros-copy examination after TOE administration (5 mg/kg i.v.) for30 min. A great many of unknown complexes were observed on afew erythrocytes with normal shapes (Fig. 4).
3.6. Complement assay
SC5b-9 complexes were significantly up-regulated after incuba-tion with TOE in vitro (Fig. 5a). However, no significant differencewas present after TOE administration (5 mg/kg i.v.) for 180 minin vivo (Fig. 5b).
4. Discussion
Jellyfish stings lead to a series of local or systemic symptoms,and jellyfish toxins have a wide range of biological activities. Themajority of jellyfish toxins have been shown to have haemolyticactivity (Tibballs, 2006), and our laboratory has proven that TOEdisplays a similar haemolytic activity to nematocyst venom fromjellyfish C. capillata (Nie et al., 2008). Almost all the previous stud-ies utilized erythrocyte suspension as the exclusive test system forhaemolysis of venoms from various specimens of jellyfish in vitro.

Fig. 5. SC5b-9 complexes were determined by EIA both in vitro and in vivo. (a)SC5b-9 of rat serum significantly increased after TOE treatment with the v/v ratio of1:10 in vitro. (b) SC5b-9 complexes did not increase significantly after TOEadministration (5 mg/kg i.v.) within 180 min in vivo. Each value representsmean ± SD (n = 4). Significant difference (P < 0.01) is expressed as the double solidsymbol compared with the control.
Fig. 4. Erythrocyte membrane examination by a transmission electron microscopy. (a) Control erythrocytes with smooth membrane and normal shape. (b) A great many ofunknown complexes (indicated by the arrows) were observed on the erythrocyte membrane after TOE administration (5 mg/kg i.v.) for 30 min. Scale bar represents 2 lm.
1206 L. Xiao et al. / Toxicology in Vitro 24 (2010) 1203–1207
But in fact, the test system of erythrocyte suspension shows onlythe damage of toxins on cell membrane, which cannot completelysimulate the real process of haemolysis in vivo, for that erythro-cytes do not exist alone in the body, but as a whole in the blood.It is well known that some proteins in blood plasma (albumin,globulin, etc.) can absorb toxins, which may reduce the availabilityto the erythrocytes. Therefore, we speculated that the haemolysisof TOE tested in diluted whole blood could be different from thatin erythrocyte suspension.
So, the rat erythrocyte suspension and diluted whole bloodapproximately with the same erythrocyte concentration were uti-lized for TOE haemolytic study in vitro. It was found that dose–response curves of haemolytic activity were both sigmoid shapedin the two test systems and haemolytic activity increased be-
tween 100 and 400 lg/ml TOE. However, at 200 lg/ml TOE, thehaemolytic activity in erythrocyte suspension was far strongerthan that in rat whole blood, indicating that the haemolysis ofTOE in whole blood is not totally consistent with that in erythro-cyte suspension, and blood plasma might play a protective roleagainst haemolysis, which was quite consistent with our previousestimate. These results implied that the two different effects ‘‘cellmembrane injury” and ‘‘haemolysis” caused by jellyfish toxinsshould be distinguished. Erythrocyte suspension could be usedto test the damage of toxin on cell membrane, while the dilutedwhole blood would be more suitable to test the haemolysis oftoxins.
The toxic effect of a jellyfish sting, local or systemic, is a sys-temic response made by the body as a whole. Therefore, we spec-ulated that the haemolysis process may also be different betweenin vivo and in vitro. Thus, we directly detected haemolysis of TOEin vivo by determination of the OD of arterial blood serum at414 nm after TOE administration (5 mg/kg i.v.). Meanwhile, wealso examined the blood potassium and lactic acid, which couldreflect the in vivo haemolysis indirectly. Results showed thatthe OD sharply increased in the first 10 min, implying that a ra-pid and severe haemolysis takes place immediately with a greatdeal of erythrocytes lysed by TOE. Then it was followed by agradual increase in the following 3 h, which may be attributedto a partial lesion of some erythrocytes’ membrane. The indirectindices, blood potassium and lactic acid, increased rapidly andreached a maximum within 10 min, which were consistent withthe results of TOE haemolysis in vivo. The following decrease inboth blood potassium and lactic acid may have been due to thecompensatory response in vivo. Besides, we showed that boththe maximal and minimal erythrocyte osmotic fragilities in-creased, further supporting a partial lesion on some erythrocytes’membrane by TOE (5 mg/kg i.v.) in vivo. These results clearly elu-cidate that the haemolysis of TOE in vivo consists of two phases:the first, a rapid and severe haemolysis, and followed by a grad-ual haemolysis. The in vivo haemolysis is not consistent with thein vitro haemolysis and this further illustrated that the test sys-tem erythrocyte suspension cannot completely imitate the haem-olysis in vivo.
Pore-formation is a possible mechanism of haemolysis by jel-lyfish venom. It was reported by Edwards et al. (2000) that theMan-of-War venom markedly increased both Ca2+, Na+ influxand K+ efflux in the primary cultured, embryonic chick heartcells, and the Ca2+ influx was inhibited by certain trivalent(La3+, Gd3+) and divalent (Zn2+, Ni2+, Cu2+, Mn2+) metals but notorganic calcium blockers (diltiazem, verapamil, nifedipine, nimo-dipine and mibefradil), concluding that the venom most likelyproduces those effects by either activating dormant, nonspecificcation channels or forming nonselective channels or pores in

L. Xiao et al. / Toxicology in Vitro 24 (2010) 1203–1207 1207
the sarcolemma. Apparent membrane pore-formation was re-vealed in cultured cells by transmission electron microscopicexamination after exposure to jellyfish venoms (Edwards et al.,2002; Bailey et al., 2005). In our studies, complexes were ob-served on a lot of erythrocytes with normal shapes after TOEadministration (5 mg/kg i.v.) for 30 min by a transmission elec-tron microscopy. As mentioned above, some plasma proteinssuch as albumin and globulin could absorb toxins, thus these‘‘unknown complexes” may be the combination products of tox-in–plasma proteins which have been absorbed onto the erythro-cyte membrane, and the haemolytic components of TOE werecontained in the complexes.
Ishikawa et al. (2004) reported that jellyfish venom in contactwith human serum was able to induce complement activation byalternative pathway activation and SC5b-9 formation was an inte-gral part of the toxin’s action process. It was consistent with our re-sults of SC5b-9 examination in vitro. However, we did not detect asignificant increase of SC5b-9 within 180 min after TOE adminis-tration (5 mg/kg i.v.). Thus, complement activation may not beone of the major factors of haemolysis in vivo. However, nonspe-cific channels and oxidative damage are the other potential mech-anisms proposed in previous studies (Chung et al., 2001; Edwardset al., 2002; Bailey et al., 2005; Heidi et al., 2007; Marino et al.,2009), and may contribute to the TOE haemolysis in vivo.
In conclusion, we, for the first time, proposed that the two dif-ferent effects ‘‘cell membrane injury” and ‘‘haemolysis” caused byjellyfish toxins should be distinguished, taking into account the po-tential protective effect of plasma on TOE haemolysis, and foundthat the haemolysis of TOE in vivo involved two phases, which isdifferent from the single phase of haemolysis in vitro. So, it wassuggested that erythrocyte suspension be used to test the damageof toxin on cell membrane, while the diluted whole blood be moresuitable to test the haemolysis of toxins. In addition, it was shownthe complement activation may not be one of the major factors forpathogenesis by TOE in vivo.
Acknowledgement
This research was funded by the Natural Science Foundationof Shanghai Municipal Government (06ZR14111) and the Founda-tion of the ‘‘Eleventh Five-Year Plan” for Medical Science Devel-opment of PLA (06Z020). The authors thank Fang Wei of theForeign Languages Department of the Second Military MedicalUniversity for his careful revision of the English language for thismanuscript.
References
Bailey, P.M., Bakker, A.J., Seymour, J.E., Wilce, J.A., 2005. A functional comparison ofthe venom of three Australian jellyfish – Chironex fleckeri, Chiropsalmus sp., andCarybdea xaymacana – on cytosolic Ca2+, haemolysis and Artemia sp. lethality.Toxicon 45, 233–242.
Bradford, M.M., 1976. A rapid and sensitive method for the quantitation ofmicrogram quantities of protein utilizing the principle of protein dye binding.Anal. Biochem. 72, 248–254.
Chung, J.J., Ratnapala, L.A., Cooke, I.M., Yanagihara, A.A., 2001. Partial purificationand characterization of a hemolysin (CAH1) from Hawaiian box jellyfish(Carybdea alata) venom. Toxicon 39, 981–990.
Edwards, L., Luo, E., Hall, R., Gonzalez, R.R., Hessinger, D.A., 2000. The effect ofPortuguese Man-of-war (Physalia physalis) venom on calcium, sodium andpotassium fluxes of cultured embryonic chick heart cells. Toxicon 38, 323–335.
Edwards, L.P., Whitter, E., Hessinger, D.A., 2002. Apparent membrane pore-formation by Portuguese Man-of-war (Physalia physalis) venom in intactcultured cells. Toxicon 40, 1299–1305.
Heidi, I.M., Lourdes, S.P., Sonia, G.A., Abel, S., Judith, S.R., 2007. The crude venomfrom the sea anemone Stichodactyla helianthus induces haemolysis and slightperoxidative damage in rat and human erythrocytes. Toxicol. in Vitro 21, 398–402.
Helmholz, H., Ruhnau, C., Schütt, C., Prange, A., 2007. Comparative study on the celltoxicity and enzymatic activity of two northern scyphozoan species Cyaneacapillata (L.) and Cyanea lamarckii (Péron & Léslieur). Toxicon 50, 53–64.
Ishikawa, T., Vucenik, I., Shamsuddin, A., Niculescu, F., Burnett, J.W., 2004. Two newactions of sea nettle (Chrysaora quinquecirrha) nematocyst venom: studies onthe mechanism of actions on complement activation and on the central nervoussystem. Toxicon 44, 895–899.
Long, K.O., Burnett, J.W., 1989. Isolation, characterization, and comparison ofhaemolytic peptides in nematocyst venoms of two species of jellyfish(Chrysaora quinquecirrha and Cyanea capillata). Comp. Biochem. Phys. B 94,641–646.
Marino, A., Morabito, R., Pizzata, T., La Spada, G., 2008. Effect of various factors onPelagia noctiluca (Cnidaria, Scyphozoa) crude venom-induced haemolysis.Comp. Biochem. Physiol. A Mol. Integr. Physiol. 151, 144–149.
Marino, A., Morabito, R., La Spada, G., 2009. Factors altering the haemolytic power ofcrude venom from Aiptasia mutabilis (Anthozoa) nematocysts. Comp. Biochem.Physiol. A 152, 418–422.
Nie, F., Xiao, L., Zhang, J., He, Q., Fan, J.W., Li, Y.L., Zhang, L.M., 2008. Comparison ofhaemolytic activities of venom separations from jellyfish Cyanea capillata andtheir influencing factors. Acad. J. Sec. Mil. Med. Univ. 29, 83–86.
Rice, N.E., Powell, W.A., 1972. Observations on three species of jellyfish fromChesapeake Bay with special reference to their toxins. II. Cyanea capillata. Biol.Bull. 143, 617–622.
Tibballs, J., 2006. Australian venomous jellyfish, envenomation syndromes, toxinsand therapy. Toxicon 48, 830–859.
Walker, M.J.A., 1977a. The cardiac actions of a toxin-containing material from thejellyfish, Cyanea capillata. Toxicon 15, 15–27.
Walker, M.J.A., 1977b. Pharmacological and biochemical properties of a toxincontaining material from the jellyfish, Cyanea capillata. Toxicon 15, 3–14.
Walker, M.J.A., Martinez, T.T., Godin, D.V., 1977. Investigations into thecardiotoxicity of a toxin from the nematocysts of the jellyfish, Cyaneacapillata. Toxicon 15, 339–346.
Xiao, L., He, Q., Guo, Y., Zhang, J., Nie, F., Li, Y., Ye, X., Zhang, L., 2009. Cyanea capillatatentacle-only extract as a potential alternative of nematocyst venom: itscardiovascular toxicity and tolerance to isolation and purification procedures.Toxicon 53, 146–152.