immunohistological distribution of the tight junction components zo-1 and occludin in regenerating...
TRANSCRIPT

Cutaneous Biology
Immunohistological distribution of the tight junctioncomponents ZO-1 and occludin in regenerating humanepidermis
M . M A L M I N E N , * V . K O I V U K A N G A S ,– J . P E L T O N E N , * , ‡, § S - L . K A R V O N E N , * *
A . O I K A R I N E N ‡ A N D S . P E L T O N E N †, *
Departments of *Medical Biochemistry and †Dermatology, University of Turku, Kiinamyllynkatu 4–8, 20520 Turku, Finland
Departments of ‡Dermatology, §Anatomy and Cell Biology and –Surgery, University of Oulu, Oulu, Finland
**Department of Dermatology, University of Helsinki, Helsinki, Finland
Accepted for publication 28 January 2003
Summary Background Molecular characterization of tight junction proteins during the past few years has
provided novel methods for studying these specialized junctions. Tight junctions have recently been
characterized in the granular cell layer of human epidermis, and the role of these junctions in the
epidermal barrier is now being re-evaluated.
Objectives To investigate the expression of tight junction components during the re-
epithelialization of suction blisters and the regeneration of the corneal layer after tape stripping.
Methods Suction blisters were induced in eight healthy volunteers, and skin biopsies were taken 4
or 6 days afterwards. The restoration of epidermal barrier function was evaluated by measuring
water evaporation (WE) from the wound area. Tape stripping was performed on three volunteers to
remove the corneal layer. The tissues were immunolabelled using indirect immunofluorescence or
the avidin–biotin method.
Results Prior to the biopsies, WE from the blister wounds was markedly elevated in comparison
with normal skin. In the epidermis surrounding the blister, occludin and ZO-1 were expressed in the
granular cell layer only. In the hyperproliferative zone adjacent to the border of the blister, the
expression of ZO-1 was redistributed into several spinous cell layers, while occludin expression was
restricted to the upper epidermis. In the leading edge of migrating keratinocytes, both proteins were
expressed exclusively in the most superficial layer of keratinocytes. Double labelling for ZO-1 and
involucrin showed expression of both proteins in the same layers of hyperproliferative
keratinocytes, while the expression patterns were clearly different in the migrating keratinocytes.
Conclusions Tight junctions of regenerating epidermis may provide a functional barrier prior to
regeneration of the corneal layer.
Key words: epidermal diffusion barrier, intercellular junctions, involucrin, tight junction, wound
healing
Tight junctions form a paracellular permeability barrier
that regulates the movement of water, solutes and
immune cells in simple epithelia. Furthermore, tight
junctions divide the plasma membrane into the apical
and the basolateral membrane domains.1–4 For exam-
ple, in the intestinal epithelium, tight junctions (zonula
occludens) form the most apical component of the
lateral junctional complex. Several structural compo-
nents of tight junctions have been characterized during
recent years.Correspondence: Sirkku Peltonen.
E-mail: [email protected]
British Journal of Dermatology 2003; 149: 255–260.
� 2003 British Association of Dermatologists 255

Occludin is a transmembrane protein that only
occurs in tight junctions. Occludin has four hydropho-
bic transmembrane helices, and both the NH2 and the
COOH terminals point towards the cytoplasmic side of
the plasma membrane. ZO-1 is found in the cytoplas-
mic plaques of tight junctions, and also in adherens
type junctions in cells that lack tight junctions, such as
fibroblasts and cardiac myocytes.1,4,5 ZO-1 is a member
of the MAGUK (membrane-associated guanylate kinase
homologue) protein family. ZO-1 is believed to contrib-
ute to the proper organization of proteins within the
tight junction plaque.1,4,5 In addition, the family of
claudins as well as the cytoplasmic plaque proteins
ZO-2, ZO-3, cingulin, symplekin and 7H6 have been
characterized as components of tight junctions.1–5
Structures resembling tight junctions were demon-
strated in the granular cell layer of human epidermis in
the early 1970s using electron microscopy.6 Elias et al.
used freeze-fracture scanning electron microscopy to
study human and mouse stratified epithelia and
concluded that tight-junctional elements were either
fragmentary or absent.7 Thus, the epidermal diffusion
barrier has been considered not to consist of tight
junctions, but of epidermal lipids, especially the lipid
bilayers present in the cornified layer.8 However, the
characterization of the structural components of tight
junctions has provided a new means to study tight
junctions, and the debate on the presence and function
of tight junctions in epidermis has resumed.9,10 Recent
work on claudin-1 knockout mice strongly suggested
that tight junctions function as components of the
diffusion barrier.11 We have previously shown that
occludin and ZO-1 are located in the granular cell layer
of adult human epidermis, while tight junctions have
been localized to the intercellular contacts of perider-
mal cells in fetal skin.10 Pathological conditions affect-
ing the stratum corneum, such as psoriasis, lichen
planus and ichthyosis vulgaris, showed altered expres-
sion of ZO-1 and occludin.10,12 Thus, tight junctions
may play a more important role in the formation of the
epidermal diffusion barrier if the epidermal lipid barrier
is abnormal or absent.
Wound healing provides a model for studying
epidermal keratinocytes in the proliferating, migrating
and differentiating phases.13,14 The present study was
designed to investigate the expression of the tight
junction components ZO-1 and occludin during the re-
epithelization of skin after the induction of suction
blisters and the regeneration of the corneal layer after
tape stripping.
Materials and methods
Suction blisters
Eight healthy medical students aged 23–26 years
(median 24; six men and two women) participated in
the study. All gave informed consent. Suction blisters
were induced on clinically normal abdominal skin
according to the method of Kiistala.15 Prolonged
suction of the skin leads to a detachment of the
dermoepidermal junction. The roof of the blister
consists of the epidermis and the blister floor the
dermis, while the basal lamina remains principally
intact. In the present study, a 150–250 mmHg
negative pressure was applied for 1–2 h.15–17 Dispo-
sable plastic suction devices (Dermovac; Ventipress,
Lappeenranta, Finland) with five pores were used.
The blister roofs were removed after the blister
induction, and the wounds were left open to heal.
The healing process was observed by measuring
water evaporation (WE) from the blister area. WE
was measured using an Evaporimeter EP1 (Servomed,
Stockholm, Sweden).18 This instrument records the
humidity of air in an open cylinder probe of 12-mm
diameter.
Four biopsies were taken on the fourth day and four
biopsies on the sixth day of healing. The WE measure-
ments were performed both after the blister induction
and before the biopsy. The value of daily evaporation
from healthy skin was subtracted from the wound area
values, because the diameter of the cylindrical probe
was larger than that of the wound. The measurements
were done in a supine position in a room with standard
temperature. The biopsies were taken under local
anaesthesia (1% lidocaine) in an operating room. A
small scalpel was used, and the wound was closed
using interrupted sutures. The sutures were removed
after 1 week. There were no complications in the
healing of the biopsy wounds.
Tape stripping
Tape stripping was performed on three volunteers. To
remove the corneal layer,19 cellophane tape was
applied 90 times repeatedly to an area of normal arm
skin. Three-millimetre punch biopsies were taken
immediately after the tape stripping and 1, 2, 3 and
5 days later. Part of the tape-stripped skin area of two
subjects was covered with cotton swabs soaked in
distilled deionized water and occlusive tape for 24 h.
Biopsies were taken after 24 h of water occlusion.
� 2003 British Association of Dermatologists, British Journal of Dermatology, 149, 255–260
2 5 6 M . M A L M I N E N et al.

Control samples were taken from normal skin adjacent
to the treated area.
Antibodies
The following antibodies were used: mouse monoclonal
antibodies to human ZO-1 (33–9100) and occludin
(33–1500), polyclonal rabbit antibodies to human
ZO-1 (61–7300) and occludin (71–1500) (all were
from Zymed Laboratories Inc., San Francisco, CA,
U.S.A.), and mouse monoclonal antibody to human
involucrin (Ab-1, MS-126-P0; NeoMarkers, Fremont,
CA, U.S.A.). The secondary antibody for indirect
immunofluorescence was tetramethylrhodamine isoth-
iocyanate (TRITC)-conjugated rabbit antimouse IgG
(R0270; Dako A ⁄ S, Glostrup, Denmark). In double
labellings, TRITC-conjugated swine antirabbit IgG
(R0156; Dako A ⁄ S) was mixed with Alexa FluorTM
488-conjugated goat antimouse IgG (Molecular Probes,
Eugene, OR, U.S.A.).
Avidin–biotin immunolabelling
To reveal the tight junction antigens from formalin-
fixed and paraffin-embedded tissue, the sections were
deparaffinized and then exposed to microwaves at
560 W in a water solution containing 0Æ1 mmol L)1
sodium citrate (pH 6) for a total of 10 min. Formalin-
fixed and paraffin-embedded specimens were immuno-
labelled with the avidin–biotin method using a
Histostain-Plus Kit (Zymed Laboratories Inc.) according
to the protocol supplied with the kit by the manufac-
turer. The antibody localization was visualized using
3,3¢-diaminobenzidine tetrahydrochloride (DAB-Plus
Kit; Zymed Laboratories Inc.). The slides were count-
erstained with Mayer’s haematoxylin (Oy Reagena Ltd,
Kuopio, Finland).
Indirect immunofluorescence labelling
Frozen sections of suction blisters were cut on silanated
glass slides. The samples were fixed with 100% methyl
alcohol at )20 �C for 10 min.10 To prevent non-
specific binding, the samples were preincubated in
phosphate-buffered saline (PBS) supplemented with 1%
bovine serum albumin (BSA) for 10 min. The primary
antibodies were diluted in 1% BSA–PBS and incubated
on the samples at 4 �C for 20 h. Following five 10-min
washes in PBS, the slides were incubated with secon-
dary antibodies at 20 �C for 1 h. After the incubation,
the samples were washed five times in PBS and
mounted with Glycergel (Dako A ⁄ S). In the control
immunoreactions, the primary antibody was replaced
with 1% BSA–PBS. In addition, the secondary and
primary antibodies used in double labellings were
tested for cross-reactions. In all controls, only faint
uniform background fluorescence was observed.
Confocal laser scanning microscopy
Confocal laser scanning microscopy was carried out
using a Leica TCS SP spectral confocal laser scanning
microscope equipped with an air-cooled argon–krypton
ion-laser system (Leica Microsystems Heidelberg
GmbH, Heidelberg, Germany) and Leica TCS NT
software (version 1.6.551). The final images were
saved in a tagged image file format.
Results
Water evaporation from suction blisters
Based on previous experiments on healing suction
blisters, the time-points of 4 and 6 days after the
induction of blisters were chosen for tissue sampling.20
The healing process was evaluated by measuring WE
from the wound area. WE in the skin indicates the
epidermal barrier function.18,20–23 In the normal skin
of the eight volunteers, WE ranged from 6 to
25 g m)2 h)1 (median 17Æ5). After the blister induc-
tion, WE ranged from 96 to 130 g m)2 h)1 (median
113Æ5). On the fourth day, the WE values from blister
wounds were 9, 25, 18 and 29 g m)2 h)1 higher than
the normal skin values, and on the sixth day, the WE
values were 7, 12, 12 and 13 g m)2 h)1 above the
baseline. The data show barrier function restoration at
these time points. However, the barrier was not yet
complete.
Avidin–biotin immunolabelling for ZO-1 and occludin
A histological analysis of all samples showed that
keratinocytes were migrating on the wound bed, but
that the wound was not fully covered by epithelium.
Three morphologically different zones could be detec-
ted: (i) the normal epidermis surrounding the blister;
(ii) the hyperproliferative zone at the border of the
blister; and (iii) the leading edge of migrating keratino-
cytes (Fig. 1A,B). Avidin–biotin immunolabelling
showed that, in the normal epidermis, ZO-1 and
occludin were expressed in the granular cell layer, as
reported earlier.10,24 The hyperproliferative zone
� 2003 British Association of Dermatologists, British Journal of Dermatology, 149, 255–260
Z O - 1 A N D O C C L U D I N I N R E G E N E R A T I N G H U M A N E P I D E R M I S 2 5 7
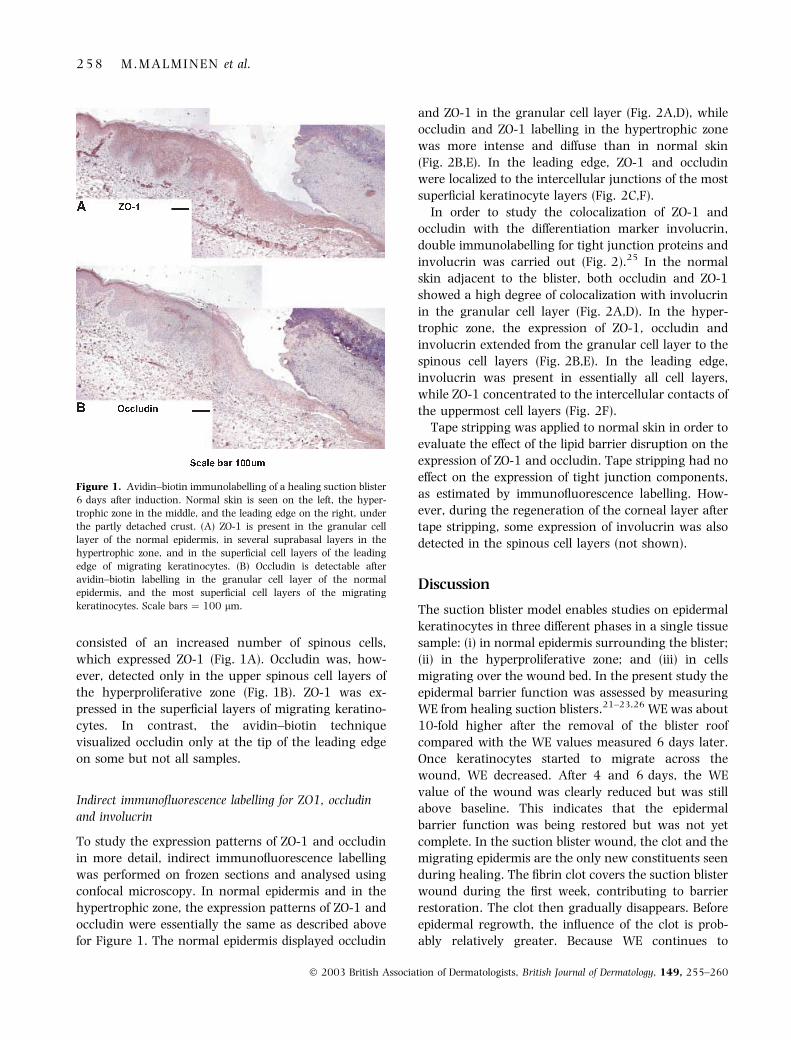
consisted of an increased number of spinous cells,
which expressed ZO-1 (Fig. 1A). Occludin was, how-
ever, detected only in the upper spinous cell layers of
the hyperproliferative zone (Fig. 1B). ZO-1 was ex-
pressed in the superficial layers of migrating keratino-
cytes. In contrast, the avidin–biotin technique
visualized occludin only at the tip of the leading edge
on some but not all samples.
Indirect immunofluorescence labelling for ZO1, occludin
and involucrin
To study the expression patterns of ZO-1 and occludin
in more detail, indirect immunofluorescence labelling
was performed on frozen sections and analysed using
confocal microscopy. In normal epidermis and in the
hypertrophic zone, the expression patterns of ZO-1 and
occludin were essentially the same as described above
for Figure 1. The normal epidermis displayed occludin
and ZO-1 in the granular cell layer (Fig. 2A,D), while
occludin and ZO-1 labelling in the hypertrophic zone
was more intense and diffuse than in normal skin
(Fig. 2B,E). In the leading edge, ZO-1 and occludin
were localized to the intercellular junctions of the most
superficial keratinocyte layers (Fig. 2C,F).
In order to study the colocalization of ZO-1 and
occludin with the differentiation marker involucrin,
double immunolabelling for tight junction proteins and
involucrin was carried out (Fig. 2).25 In the normal
skin adjacent to the blister, both occludin and ZO-1
showed a high degree of colocalization with involucrin
in the granular cell layer (Fig. 2A,D). In the hyper-
trophic zone, the expression of ZO-1, occludin and
involucrin extended from the granular cell layer to the
spinous cell layers (Fig. 2B,E). In the leading edge,
involucrin was present in essentially all cell layers,
while ZO-1 concentrated to the intercellular contacts of
the uppermost cell layers (Fig. 2F).
Tape stripping was applied to normal skin in order to
evaluate the effect of the lipid barrier disruption on the
expression of ZO-1 and occludin. Tape stripping had no
effect on the expression of tight junction components,
as estimated by immunofluorescence labelling. How-
ever, during the regeneration of the corneal layer after
tape stripping, some expression of involucrin was also
detected in the spinous cell layers (not shown).
Discussion
The suction blister model enables studies on epidermal
keratinocytes in three different phases in a single tissue
sample: (i) in normal epidermis surrounding the blister;
(ii) in the hyperproliferative zone; and (iii) in cells
migrating over the wound bed. In the present study the
epidermal barrier function was assessed by measuring
WE from healing suction blisters.21–23,26 WE was about
10-fold higher after the removal of the blister roof
compared with the WE values measured 6 days later.
Once keratinocytes started to migrate across the
wound, WE decreased. After 4 and 6 days, the WE
value of the wound was clearly reduced but was still
above baseline. This indicates that the epidermal
barrier function was being restored but was not yet
complete. In the suction blister wound, the clot and the
migrating epidermis are the only new constituents seen
during healing. The fibrin clot covers the suction blister
wound during the first week, contributing to barrier
restoration. The clot then gradually disappears. Before
epidermal regrowth, the influence of the clot is prob-
ably relatively greater. Because WE continues to
Figure 1. Avidin–biotin immunolabelling of a healing suction blister
6 days after induction. Normal skin is seen on the left, the hyper-
trophic zone in the middle, and the leading edge on the right, under
the partly detached crust. (A) ZO-1 is present in the granular cell
layer of the normal epidermis, in several suprabasal layers in the
hypertrophic zone, and in the superficial cell layers of the leading
edge of migrating keratinocytes. (B) Occludin is detectable after
avidin–biotin labelling in the granular cell layer of the normal
epidermis, and the most superficial cell layers of the migrating
keratinocytes. Scale bars ¼ 100 lm.
� 2003 British Association of Dermatologists, British Journal of Dermatology, 149, 255–260
2 5 8 M . M A L M I N E N et al.

decrease, it is likely that the migrating epidermis and its
evaporation-controlling elements, tight junctions, cre-
ate a new barrier. As the stratum corneum does not
exist in the new epidermis20,23 on the fourth or sixth
days, the lipid barrier has no effect on evaporation at
that time.
The purpose of the present study was to map the
distribution of tight junction proteins in the healing
epidermis. Recent studies have emphasized the role of
tight junctions in the regulation of epidermal permeab-
ility. Genetic ablation of the claudin-1 gene in mice
results in neonatal death, probably because of signifi-
cant dehydration.11 Mice overexpressing claudin-6 die
within 2 days of birth, apparently due to barrier
dysfunction and increased water loss.27 Other recent
studies demonstrate that tight junctions are present in
fetal epidermis10,24,27 which lacks the corneal layer.
Thus, it is feasible to speculate that tight junctions form
a functional barrier in premature skin. Regenerating
epidermis is analogous to fetal skin in its lack of a
normal corneal layer. In this study, the two most
superficial layers of migrating keratinocytes on the
wound bed expressed occludin and ZO-1, suggesting
the presence of tight junction proteins between these
cells. The expression of the tight junction proteins
claudin-1, occludin and ZO-1 has recently been studied
in a human skin organ culture model, showing
occludin and ZO-1 in the first ingrowing epithelial
cells early during wound healing.24 The results of
Brandner et al.24 are thus in agreement with the results
of our study on the in vivo wound healing model. Taken
together, these studies indicate that at least three tight
junction components are present in the regenerating
epithelium before the reconstruction of stratum cor-
neum. These observations further emphasize the role of
tight junctions in epidermis, especially in conditions
where no normal lipid barrier is present.
The hyperproliferative zone, which develops adjacent
to the original border of the blister, displayed a unique
labelling pattern for ZO-1 and occludin. The expression
of ZO-1 was redistributed into several spinous cell
layers. Involucrin was also detected in the granular cell
and spinous cell layers to approximately the same level
as ZO-1. In analogy with healing epidermis, ZO-1 and
involucrin are also coexpressed in the granular and
spinous cell layers in psoriasis.10,12 The expression of
ZO-1 may thus be linked with the differentiation of
keratinocytes. Claudin-1 is expressed in the granular
Figure 2. Indirect immunofluorescence
labelling of a healing suction blister for
occludin (Occl, red), and double immuno-
labelling for tight junction proteins and invo-
lucrin (Inv, green). (A,D) In normal epidermis,
double labelling for occludin or ZO-1 (red) and
involucrin reveals colocalization in the gran-
ular cell layer, while the polyclonal antibody
also detects occludin in the uppermost spinous
cell layers (A, arrow). (B,E) In the hyper-
trophic zone, both tight junction proteins and
involucrin are detected in a wider zone
extending to the spinous cell layers. (C,F) In
the leading edge, both ZO-1 and occludin are
present in the most superficial cell layers. The
white dotted line indicates the location of the
dermal–epidermal junction. (F) Involucrin is
expressed in essentially all cell layers, while
ZO-1 is restricted to the intercellular contacts
of the uppermost cell layers. Panel F repre-
sents a sample taken 4 days after the induc-
tion of the blister, while other figures
represent the sixth day of healing. Scale
bars ¼ 20 lm.
� 2003 British Association of Dermatologists, British Journal of Dermatology, 149, 255–260
Z O - 1 A N D O C C L U D I N I N R E G E N E R A T I N G H U M A N E P I D E R M I S 2 5 9

and spinous cell layers in normal epidermis and at the
border of the wound.24 However, it is currently not
known why the distribution of occludin differs from
that of ZO-1 and claudin-1. To conclude, these
results suggest that tight junctions may have a role
in the development of the diffusion barrier in healing
epidermis.
Acknowledgments
This work was supported by Turku University Foun-
dation, Turku University Central Hospital (grant
13338), Oulu University Hospital, Cancer Societies of
Finland, and Department of Dermatology, University of
Oulu, Oulu, Finland.
References
1 Stevenson BR, Keon BH. The tight junction: morphology to
molecules. Annu Rev Cell Dev Biol 1998; 14: 89–109.
2 Tsukita S, Furuse M. Occludin and claudins in tight-junction
strands: leading or supporting players? Trends Cell Biol 1999; 9:
268–73.
3 Tsukita S, Furuse M. Pores in the wall: claudins constitute tight
junction strands containing aqueous pores. J Cell Biol 2000; 149:
13–16.
4 Tsukita S, Furuse M, Itoh M. Structural and signalling molecules
come together at tight junctions. Curr Opin Cell Biol 1999; 11:
628–33.
5 Anderson JM, Itallie CM. Tight junctions and the molecular basis
for regulation of paracellular permeability. Am J Physiol 1995;
269: G467–75.
6 Hashimoto K. Intercellular spaces of the human epidermis as
demonstrated with lanthanum. J Invest Dermatol 1971; 57: 17–
31.
7 Elias PM, McNutt NS, Friend DS. Membrane alterations during
cornification of mammalian squamous epithelia: a freeze-fracture,
tracer, and thin-section study. Anat Rec 1977; 189: 577–94.
8 Wertz PW. Lipids and barrier function of the skin. Acta Derm
Venereol (Stockh) 2000; 208 (Suppl.): 7–11.
9 Morita K, Itoh M, Saitou M et al. Subcellular distribution of tight
junction-associated proteins (occludin, ZO-1, ZO-2) in rodent
skin. J Invest Dermatol 1998; 110: 862–6.
10 Pummi K, Malminen M, Aho H et al. Epidermal tight junctions:
ZO-1 and occludin are expressed in mature, developing, and
affected skin, and in vitro differentiating keratinocytes. J Invest
Dermatol 2001; 117: 1050–8.
11 Furuse M, Hata M, Furuse K et al. Claudin-based tight junctions
are crucial for the mammalian epidermal barrier: a lesson from
claudin-1-deficient mice. J Cell Biol 2002; 156: 1099–111.
12 Yoshida Y, Morita K, Mizoguchi A et al. Altered expression of
occludin and tight junction formation in psoriasis. Arch Dermatol
Res 2001; 293: 239–44.
13 Singer AJ, Clark RA. Cutaneous wound healing. N Engl J Med
1999; 341: 738–46.
14 Martin P. Wound healing—aiming for perfect skin regeneration.
Science 1997; 276: 75–81.
15 Kiistala U. Suction blister device for separation of viable epidermis
from dermis. J Invest Dermatol 1968; 50: 129–37.
16 Eaglstein WH, Mertz PM. New methods for assessing epidermal
wound healing: the effects of triamcinolone acetonide and poly-
ethylene film occlusion. J Invest Dermatol 1978; 71: 382–4.
17 Oikarinen A, Savolainen E-R, Tryggvason K et al. Basement
membrane components and galactosylhydroxylysyl glycosyl-
transferase in suction blisters of human skin. Br J Dermatol 1982;
106: 257–66.
18 Nilsson GE. Measurement of water exchange through skin. Biol
Eng Comput 1977; 15: 209–18.
19 Ahn SK, Hwang SM, Jiang SJ et al. The changes of epidermal
calcium gradient and transitional cells after prolonged occlusion
following tape stripping in the murine epidermis. J Invest Dermatol
1999; 113: 189–95.
20 Kallioinen M, Koivukangas V, Jarvinen M, Oikarinen A. Expres-
sion of cytokeratins in regenerating human epidermis. Br J Der-
matol 1995; 133: 830–5.
21 Silverman RA, Lender JL, Elmets CA. Effects of occlusive and
semiocclusive dressings on the return of barrier function to
transepidermal water loss in standardized human wounds. J Am
Acad Dermatol 1989; 20: 755–60.
22 Levy JJ, Rosen J, Gabmuller J et al. Validation of an in vivo wound
healing model for the quantification of pharmacological effects on
epidermal wound healing. Dermatology 1995; 190: 136–41.
23 Koivukangas V, Annala A-P, Salmela PI, Oikarinen A. Delayed
restoration of the epidermal barrier function after suction blister
injury in patients with diabetes. Diabet Med 1999; 16: 563–7.
24 Brandner JM, Kief S, Grund C et al. Organization and formation of
the tight junction system in human epidermis and cultured ker-
atinocytes. Eur J Cell Biol 2002; 81: 253–63.
25 Ekanayake-Mudiyanselage S, Aschauer H, Schmook FP et al.
Expression of epidermal keratins and the cornified envelope pro-
tein involucrin is influenced by permeability barrier disruption.
J Invest Dermatol 1998; 111: 517–23.
26 Mathias CG, Wilson DM, Maibach HI. Transepidermal water loss
as a function of skin surface temperature. J Invest Dermatol 1981;
77: 219–20.
27 Turksen K, Troy TC. Permeability barrier dysfunction in trans-
genic mice overexpressing claudin 6. Development 2002; 29:
1775–84.
� 2003 British Association of Dermatologists, British Journal of Dermatology, 149, 255–260
2 6 0 M . M A L M I N E N et al.