identification and intracellular localization of the unc...
TRANSCRIPT

Identification and intracellular localization of the unc-22 gene product of Caenorhabditis elegans D o n a l d G. M o e r m a n , 1 Guy M. Benian, 2 Robert J. Barstead, Lawrence A. Schriefer, and Robert H. Waterston
Department of Genetics, Washington University School of Medicine, St. Louis, Missouri 63110 USA
The unc-22 gene is one of a set of genes identified using classical genetics that affect muscle structure and function in the free-living nematode Caenorhabditis elegans. Since cloning the unc-22 gene by transposon tagging, we have used conventional techniques combined with a set of Tcl transposon insertion alleles to characterize the gene and its products. The gene extends over more than 20 kb of genomic sequence and produces a transcript of approximately 14 kb. A polyclonal antibody raised against an Escherichia coli ~- galactosidase--unc-22 fusion protein recognizes a polypeptide in nematode extracts that is between 500,000 and 600,000 daltons and labels the muscle A-band in indirect immunofluorescent microscopy. The Tcl-induced alleles have been used at every stage to verify these conclusions. The Tcl insertions are spread over much of the region that contributes to the mature transcript; in most alleles, Tc l sequences are incorporated into a composite unc-22-Tcl transcript. The large protein is either absent or severely reduced in amounts in the mutants. In one case, a truncated polypeptide was also identified. The location of the protein in the A-band, along with earlier genetic data, suggests that the unc-22 product may interact with myosin to regulate its function.
[Key Words: Muscle structure; unc-22; myosin; transposon; nematode]
Received July 6, 1987; revised version accepted November 4, 1987.
The discovery of mutator strains of Caenorhabditis elegans in which the element Tcl transposes at high fre- quencies has made possible the recovery of DNA se- quences for genes without prior knowledge of the pro- tein product (Moerman and Waterston 1984; Eide and Anderson 1985; Greenwald 1985; Moerman et al. 1986). Among the first genes isolated via Tcl tagging was the unc-22 locus (Moerman et al. 1986), which is a particu- larly frequent target of the element and has been used extensively in characterizing the mutator activities. The unc-22 locus is also of interest as one of more than 25 genes implicated in normal muscle formation in C. elegans (Brenner 1974; Waterston et al. 1980; Zengel and Epstein 1980). This set of genes has been studied inten- sively, and the genes for many of the major components of the myofilament lattice have been identified, in- cluding myosin heavy-chain isoforms (Epstein et al. 1974; MacLeod et al. 1977b; Karn et al. 1983), actins (Landel et al. 1984), and paramyosin (Waterston et al.
~Present address: Department of Zoology, University of British Co- lumbia V6T 2A9 Canada. 2present address: Department of Pathology, Emory University School of Medicine, Atlanta, Georgia 30322 USA.
1977; H. Kagawa and J. Karn, pers. comm.). The product of the unc-22 gene, however, remained unknown.
Among the muscle-affecting genes, unc-22 is particu- larly intriguing because of its unusual phenotype. Mu- tant alleles of the locus lead to varying degrees of im- paired movement and muscle disorganization (Moerman 1980; Waterston et al. 1980). Weak alleles have little ef- fect on movement or muscle structure. Strong alleles lead to almost complete paralysis of adult animals and to abnormal muscle structure in which thick and thin filaments, though present in normal quantities, are not in their normal register. All alleles, however, produce a nearly constant, subcellular twitch in all the body wall muscle cells. This phenotypic feature otherwise results only from rare alleles of the unc-54 myosin heavy-chain gene (MacLeod et al. 1977a; Dibb et al. 1985) and from one allele of lev-ll (Lewis et al. 1980). Whether the twitch represents an unregulated contraction or relax- ation is unclear, but the twitch is exacerbated by choline agonists (Moerman and Baillie 1979).
Genetic analysis has yielded several insights into the nature of the gene and its function. The gene is an un- usually large mutational target (Zengel and Epstein 1980), and intragenic recombination frequencies for al- leles at opposite ends of the gene are similar to or higher
GENES & DEVELOPMENT 2:93-105 © 1988 by Cold Spring Harbor Laboratory ISSN 0890-9369/88 $1.00 93
Cold Spring Harbor Laboratory Press on August 31, 2018 - Published by genesdev.cshlp.orgDownloaded from

Moerman et al.
than frequencies seen for the most distant allelic pairs of the unc-54 gene (Moerman and Baillie 1979; Waterston et al. 1982), which encodes the 228,000-dalton myosin heavy chain (Karn et al. 1983). Thus, the gene is likely to be much larger than average. Most alleles are condition- ally dominant, that is, heterozygous unc-22/+ animals move normally and have normal muscle structure but can be induced to twitch violently in solutions of cho- line agonists (Moerman and Baillie 1979). As this domi- nance is shown even by likely null alleles, including an amber allele s32 and deletion alleles, the unc-22 gene probably encodes a protein required stoichiometrically. Most intriguing of all, reversion analysis of unc-22 al- leles revealed that certain missense alleles of the myosin heavy-chain gene unc-54 can suppress the unc-22 pheno- type (Moerman et al. 1982). Not only is the twitch sup- pressed and motility partially restored, but muscle structure is also improved. Sequence analysis of the unc-54 suppressors showed that all of these mutations lie in the region coding for the myosin head; several are located near the nucleotide binding domain, and others are near the conserved thiol domain (Dibb et al. 1985; J. Kiff, D.G. Moerman, and R.H. Waterston, unpubl.).
In this study, we have used the recently obtained DNA clones from the unc-22 region (Moerman et al. 1986) to investigate the molecular organization of the gene and the nature of its product. In addition, because unc-22 is a frequent target of the C. elegans var. Bergerac mutator system, and because the mutator-induced al- leles are generally intermediate in strength, we exam- ined several mutator-induced alleles to determine the nature of the mutations and their effects on unc-22 gene expression.
Our results show that the unc-22 gene extends over more than 20 kb, that Tcl insertions probably account for all the mutator-induced alleles of this set, and that the insertions occur throughout much of the unc-22 transcribed region. In addition, a 14-kb transcript has been identified, which is increased in size in the Tcl mutants by the inclusion of the Tcl sequence. The poly- peptide identified with an antibody produced to a fusion protein is larger than rabbit nebulin, which has been re- ported to be approximately 500,000 daltons in size (Wang and Williamson 1980). This protein is absent in amber and deletion alleles and is altered in size in a Tcl- induced allele. The unc-22 polypeptide in wild type is located in muscle cells and is apparently cell auton- omous. Its colocalization with myosin at the light mi- croscopic level in several strains, together with the pre- vious genetic data, suggests that the unc-22 protein in- teracts with components of the thick filament.
R e s u l t s
Sequences important for unc-22 expression extend over more than 20 kb
The region of genomic DNA cloned in previous work extends over 70 kb (Coulson et al. 1986; A. Coulson and J. Sulston, pers. comm.), of which more than 30 kb has
been recovered in phage clones (Fig. 1; Moerman et al. 1986). Various unc-22 alleles were screened for rear- rangements within this region to help define the extent of the unc-22 gene. The positions of the deficiency sDfl 9 (Rogalski and Baillie 1985), which by genetic criteria breaks within the genes, and two Tcl insertions, st136::Tcl and st137::Tcl, were determined previous- ly (Moerman et al. 1986). Several diepoxybutane-induc- ed alleles were found to have small deficiencies to the right of the st136::Tcl position (C. Trent, pers. comm.). In addition, we examined 12 more mutator-induced unc-22 alleles for changes in restriction fragments in the unc-22 region. All 12 alleles showed alterations in their hybridization patterns; in each mutant, a change consis- tent with a single insertion of 1.6 kb, the size of Tcl (Rosenzweig et al. 1983), was found.
The 12 different Tcl insertions were placed into at least 10 different sites by analyzing the patterns pro- duced on Southern transfers of DNA from each allele digested with four different restriction enzymes (Fig. 1). For example, in DNA from the stl40::Tcl allele, the 3.5-kb SacI fragment of the wild type has been replaced by a 5.1-kb fragment (Fig. 2). A weak signal is present in this DNA at 3.5 kb, but this is presumed to be the result of somatic excision of Tcl (Emmons and Yesner 1984). The stl81 ::Tcl allele affects the same SacI fragment, but stl40::Tcl and stl81::Tcl are clearly at different sites as in stl40 DNA the 0.4-kb BglII fragment is replaced with a 2.0-kb fragment whereas in the stl81 DNA the adja- cent 3.4-kb BglII fragment is altered (data not shown). Data for two additional alleles, st185::Tcl and st139::Tcl, are also shown (Fig. 2). Eleven of the inser- tions are distributed over 17 kb. The twelfth insertion, the st139::Tcl allele, lies about 12 kb outside this group. To verify that this insertion indeed represents the rele- vant sequence change producing the Unc-22 phenotype, we recovered a mutator-induced revertant of st139::Tcl; its restriction pattern is indistinguishable from that of wild type (Fig. 2). The st139::Tcl allele is unusual in that it results in such a weak phenotype that it is neces- sary to use the choline agonist nicotine to score the ho- mozygote reliably. The leftmost-located Tcl insertion st185::Tcl, which lies under the sDfl9 deficiency, re- sults in a weak phenotype, intermediate between st139::Tcl and the other Tcl alleles.
unc-22 produces a 14-kb mRNA
To determine what transcripts might arise from the re- gion defined by the rearrangements, we probed Northern transfers of size-fractionated total RNA with various ra- diolabeled fragments from the genomic clones. A 1.6-kb RNA species was recognized only by the 4.6-kb SacI fragment that extends to the left of the sDfl 9 deficiency break point (data not shown). No probes from other re- gions recognized this RNA, so we considered that it was likely to be the product of another gene. A much larger species, estimated to be 14 kb using single-stranded DNA markers (see Fig. 3), also hybridized with the 4.6- kb SacI fragment, as well as with probes throughout the
94 GENES & DEVELOPMENT
Cold Spring Harbor Laboratory Press on August 31, 2018 - Published by genesdev.cshlp.orgDownloaded from

Molecular characterization of uric-22 gene
t s D f l 9
• st136 s t l 4 0 stl41 "--st137
stlS5 • --~ - - - -'- st187 st139
t f TtTTT,T IT ,ft I l * * ~ f 7
c t38 c t 3 6 , . . . . . , - 4 : 3 - - - ~ , - - - o . . . . .
t r a n s c r i p t ~ T c l 3 ' I . . . . 5 '
DM17 ? SacI D M 2 0 ~, B g l ~
D M 2 2 - - 5 k b
I I
unc-22 region Figure 1. The unc-22 region and sites of Tcl insertion. The genomic segment containing the unc-22 gene is shown, along with several DNA rearrangements that define its extent. Three X clones, which together span most of the region, are also shown. The minimum region contributing to the transcript is indicated, and the direction of transcription, as determined using single-stranded probes, is from right to left. The positions of the large deficiency sDfI9 (Rogalski and Baillie 1985; Moerman et al. 1986) and three smaller deficiencies, ct36, ct37, and ct38 (C. Trent, pers. comm.), are shown as open boxes. The break point of sDF19 was used to show that the orientation of the physical map to the genetic map is as depicted (Moerman et al. 1986). The approximate position of 14 Tcl insertions that result in the Unc-22 phenotype are indicated by triangles. The asterisks above two of the triangles indicate that two independent insertions have been localized within the interval. The order and approximate sizes of the SacI fragments, as well as the order of the unc-22::TcI alleles, are from right to left: 7.2 kb (st139), 1.0 kb, 0.9 kb, 4.3 kb, 1.1 kb, 3.7 kb, 0.9 kb (st187), 3.9 kb (st188), 1.7 kb (st137, st136), 0.3 kb, 1.0 kb (st184), 0.2 kb, 1.5 kb ([stl41, stI91]), 3.5 kb [stl8I, stl40, {st186, st182)], 4.6 kb (st183 and st185). The orientation of the Tcl insertion, where known, is shown by an arrow. The striped box denotes the 1.9-kb SacI fragment that was subcloned into the expression vector XRB1 (R.J. Barstead and R.H. Waterston, unpubl.).
region containing the majority of the insertion sites. The r ightmost fragment tested that gave a positive signal was the 1.1-kb SacI fragment. The adjacent 4.3-kb SacI fragment yielded no hybridization signal. Fragments far- ther to the right were not tested. The region covered by these fragments and thus contributing to the 14-kb tran- script extends over about 20 kb. Single-stranded DNA probes showed that the orientation of the transcription uni t is from right to left on the conventional genetic map.
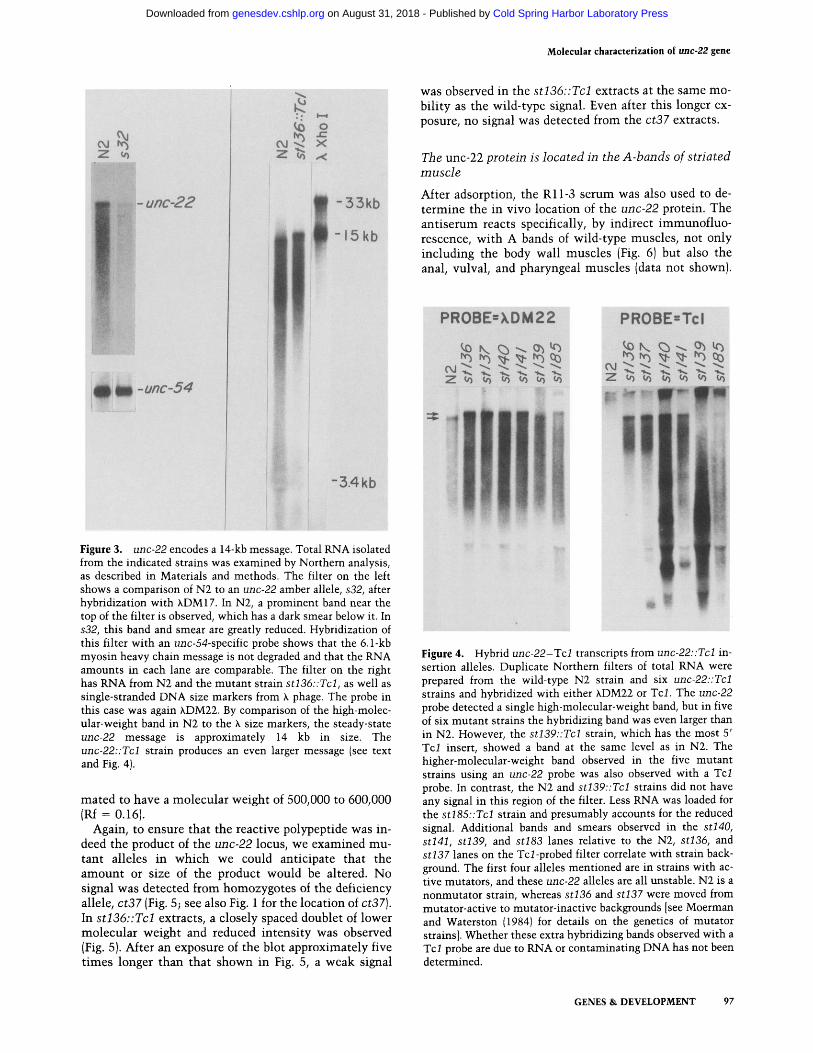
To confirm that the 14-kb transcript is the product of the unc-22 locus, we also examined RNA from several unc-22 mutants that might be expected to have altered m R N A levels or sizes. Amber alleles are known in other genes to reduce the steady-state m R N A levels (Losson and LaCroute 1979; Brown et al. 1982), and one unc-22 amber allele, s32, has been identified (Moerman 1980). In RNA from s32 homozygotes, the signal from the 14-kb band was reduced markedly compared to wild type when hybridized wi th an unc-22 region probe; in control experiments using an unc-54-specific probe against the same filter, the signal from the myosin mes- sage was s imilar between the wild-type and s32 lanes (Fig. 3).
As insert ion of transposable elements into the tran- scribed portion of a gene has been shown to affect the transcript in any of a variety of ways (Doting et al. 1984; Levis et al. 1984; Pirrotta and Brockl 1984; Rowland and Strommer 1985; Zachar et al. 1985), we also examined RNA from several of the Tc~/-induced alleles. These in- clude st139::Tcl, st137::Tcl, st136::Tcl, s t l41::Tcl , s t l40::Tcl , and st185::Tcl, which span the region of in- terest (Fig. 1). When hybridized wi th probes from unc-22
(Fig. 4a), in five of the six cases, the hybridizing tran- script appeared slightly larger than that in wild type, but of s imilar abundance. Because an increased size might result from incorporation of Tc l sequences into the transcript, s imilar Northern transfers were hybrdized wi th a Tcl sequence. In the wild type, no RNA in this region of the transfer hybridized wi th Tcl .
Both orientations of Tc l insertion are represented in these five alleles. The st136::Tcl and st137::Tcl alleles have Tcl inserted in opposite orientations (Moerman et al. 1986). Hybridizat ion of Northern transfers with single-stranded Tc l probes showed that st l40::Tcl has the same orientation as st137::Tcl and that st l41::Tcl has the same orientation as st136::Tcl. The orientation of the st185::Tcl allele was not determined.
The only allele of the six that tested positive wi th unc-22 probes but negative wi th the Tcl probe was st139::Tcl. This is the very weak allele whose site of Tc l insert ion is well to the right of the position of the other D N A rearrangements associated wi th unc-22 phe- notypes.
The unc-22 protein is larger than 500,000 daltons
To determine the size of the unc-22 protein product and its location in muscle, we raised antibodies against a polypeptide produced by the fusion of unc-22 sequences to the ~-galactosidase gene of Escherichia coli. Limited sequencing of a region near the st136::Tcl insertion site revealed an open reading frame extending through a SacI site that would remain in frame when ligated to the SacI site wi th in the ~-galactosidase gene (data not shown). We inserted the corresponding 1.9-kb unc-22 SacI frag-
GENES & DEVELOPMENT 95
Cold Spring Harbor Laboratory Press on August 31, 2018 - Published by genesdev.cshlp.orgDownloaded from

M o e r m a n et al.
Bg111" Sac I I I I I
00 00 C~I - - Od - - - ! - -
Z~Z~ ) : kb
9.5 -8.0
~, . . ,~ - 6.2
-4.6
- 3.0
Bgl rl Sac I l I I I
O O O d - - Z o~ o0
8
-5.1
-3.,.5 -3.4 - 3.0
-2.1
Sacl I I
00
0.,)~ i,<) C'xJ
Z ~ r r
kb
- 8 . 8 O ~ Q D -7.2
o o o - 4 . 3
~ 1 ~ -I.0 "0.9
Figure 2. Spontaneous unc-22 alleles isolated in mutator background have Tcl insertions in the unc-22 region. DNA from each of the 12 mutator-induced unc-22 alleles was isolated and compared to wild-type DNA by Southern blot analysis. By using the k clones in Fig. ! as probes, each mutant strain was shown to contain an insert of 1.6 kb in single SacI and BglII fragment. Once a mutation was localized to a small region, the appropriate SacI or BglII fragment spanning the insertion site was used as a probe to confirm this assignment. The simpler pattern obtained allowed unambiguous interpretation of the results. Shown are two examples, st185 and stl40, where we have probed digested DNA from strains containing these mutations with a SacI fragment spanning the Tcl insert, and one example, st139, where we have used a k clone. The st185 mutation was probed with the left-most 4.6-kb SacI fragment, stl40 was probed with the 3.5-kb SacI fragment adjacent to the 4.6-kb fragment, and st139 was probed with KDM22 containing the right- most region of the gene. In all three cases, a band 1.6 kb larger than the corresponding wild-type band was detected. Weaker hybridiza- tion at the position of the wild-type band was also seen, which was probably a result of somatic excision, a characteristic of all Tcl insertions so far identified. Similar results were obtained for the other 11 mutants described in Fig. 1. A revertant of st139::Tcl was recovered and included in the analysis. The revertant, RW2418, has lost the 8.8-kb SacI hybridizing band and regained the wild-type 7.2-kb hybridizing band.
ment in the desired orientation into a modified kgtl 1 vector that has a unique SacI site within the f3-galactosi- dase gene (Young and Davis 1983; R.J. Barstead and R.H. Waterston, unpubl.). When [3-galactosidase expression was induced, a fusion peptide of about 155 kD appeared in the k lysogens, the size expected if the open reading frame was mainta ined across the entire 1.9-kb SacI frag- ment, wi th a subsequent out-of-frame fusion to the re- mainder of the f~-galactosidase sequence (data not shown).
To obtain antiserum, the polypeptide was partially pu- rified and injected into rabbits. The resultant serum,
called Rl l -3 , was adsorbed extensively against full- length f~-galactosidase and used against Western transfers. The ant iserum reacted strongly against the 155-kD protein in k lysogens (data not shown) and with a very large protein or proteins in either 2-aminoethanol or SDS-soluble proteins from C. elegans wild-type an- imals (Fig. 5). The signal, depending on the preparation, was either a single band or a closely spaced doublet. Ex- trapolating from the mobilities of myosin heavy-chain {225,000 daltons; Rf = 0.66) and rabbit nebulin (500,000 daltons; Wang and Will iamson 1980; Rf = 0.22) markers, the wild-type protein, or proteins, was esti-
96 GENES & DEVELOPMENT
Cold Spring Harbor Laboratory Press on August 31, 2018 - Published by genesdev.cshlp.orgDownloaded from
Molecular characterization of uric-22 gene
Z
l ~ l
unc-22
:-unc-54
,,.<
1 - 15kb
J
-3.4kb
Figure 3. unc-22 encodes a 14-kb message. Total RNA isolated from the indicated strains was examined by Northern analysis, as described in Materials and methods. The filter on the left shows a comparison of N2 to an unc-22 amber allele, s32, after hybridization with XDM17. In N2, a prominent band near the top of the filter is observed, which has a dark smear below it. In s32, this band and smear are greatly reduced. Hybridization of this filter with an unc-54-specific probe shows that the 6.1-kb myosin heavy chain message is not degraded and that the RNA amounts in each lane are comparable. The filter on the right has RNA from N2 and the mutant strain st136::Tcl, as well as single-stranded DNA size markers from k phage. The probe in this case was again XDM22. By comparison of the high-molec- ular-weight band in N2 to the X size markers, the steady-state unc-22 message is approximately 14 kb in size. The unc-22::Tcl strain produces an even larger message (see text and Fig. 4).
ma ted to have a molecu la r we igh t of 500,000 to 600,000 (Rf -- 0.16).
Again, to ensure tha t the react ive polypept ide was in- deed the product of the unc-22 locus, we examined mu- t an t al leles in w h i c h we could an t ic ipa te tha t the a m o u n t or size of the product would be altered. No signal was detected f rom homozygo tes of the def ic iency allele, ct37 (Fig. 5; see also Fig. 1 for the locat ion of ct37). In st136::Tcl extracts, a c losely spaced doublet of lower molecu la r we igh t and reduced i n t e n s i t y was observed (Fig. 5). After an exposure of the blot approx imate ly five t imes longer t han tha t shown in Fig. 5, a weak signal
was observed in the st136::Tcl extracts at the same mo- b i l i ty as the wi ld- type signal. Even after th is longer ex- posure, no signal was detected from the ct37 extracts.
The unc-22 protein is located in the A-bands of striated muscle
After adsorption, the R11-3 se rum was also used to de- t e r m i n e the in vivo loca t ion of the unc-22 protein. The a n t i s e r u m reacts specifically, by indi rec t immunof luo - rescence, w i t h A bands of wi ld- type muscles , not only inc lud ing the body wal l musc les (Fig. 6) but also the anal, vulval, and pharyngea l musc les (data not shown).
PROBE=XDM22
"i
PROBE=Tcl
i
Figure 4. Hybrid unc-22-Tcl transcripts from unc-22::Tcl in- sertion alleles. Duplicate Northern filters of total RNA were prepared from the wild-type N2 strain and six unc-22::Tcl strains and hybridized with either XDM22 or Tcl. The unc-22 probe detected a single high-molecular-weight band, but in five of six mutant strains the hybridizing band was even larger than in N2. However, the st139::Tcl strain, which has the most 5' Tcl insert, showed a band at the same level as in N2. The higher-molecular-weight band observed in the five mutant strains using an unc-22 probe was also observed with a Tcl probe. In contrast, the N2 and st139::Tcl strains did not have any signal in this region of the filter. Less RNA was loaded for the s t185::Tcl strain and presumably accounts for the reduced signal. Additional bands and smears observed in the stl40, stl41, st139, and st183 lanes relative to the N2, st136, and st137 lanes on the Tcl-probed filter correlate with strain back- ground. The first four alleles mentioned are in strains with ac- tive mutators, and these unc-22 alleles are all unstable. N2 is a nonmutator strain, whereas st136 and st137 were moved from mutator-active to mutator-inactive backgrounds [see Moerman and Waterston (1984) for details on the genetics of mutator strains]. Whether these extra hybridizing bands observed with a Tcl probe are due to RNA or contaminating DNA has not been determined.
GENES & DEVELOPMENT 97
Cold Spring Harbor Laboratory Press on August 31, 2018 - Published by genesdev.cshlp.orgDownloaded from

Moerman et al.
#
r--n1 i
oJ c~i z z
q
mhc - ~ ~
~,~
p m
#
c? !
LO rO _
C~J OJ z z
m h ¢
ZeOkE/
i
!
p m
105 kE
I
t-- ro ¢ o - -
Figure 5. Antiserum to an unc-22-f~-galactosidase fusion polypeptide identifies a polypeptide in C. elegans that is larger than 500,000 daltons. Rabbit antibodies were generated to an unc-22-~-galactosidase fusion polypeptide (see text for details), and the resulting serum, R11-3, was absorbed against f~-galactosidase before use. Total C. elegans protein extracts, prepared as described in Materials and methods, were separated on a polyacrylamide gel and either stained with Coomassie blue or transferred to nitrocellulose, reacted with the R11-3 antiserum and ~2SI-labeled protein A, and visualized after autoradiography. The molecular weight markers shown here are paramyosin (pro) at 105 kD and myosin heavy chain (mhc) at 220 kD. The left pair of panels shows that in wild type animals the antisera react with a large protein or proteins of -500 kD. Although not visible here, in some extracts a faint band, corresponding to the band detected with the antiserum, can also be obsrved after Coomassie blue staining. The panels on the right demonstrate that the antibody reactive band is either absent or altered in unc-22 mutants. Mutants bearing the unc-22 internal deficiency ct37 lack this large protein; no signal can be detected even after prolonged exposure. From a st136::Tcl-bearing strain, a shorter polypeptide that runs as a doublet is detected with the R11-3 antiserum. In some preparations, the reactive species in N2 can sometimes be a doublet.
No signal was observed in any of these muscle cells in animals treated wi th the pre immune serum. In the body wall muscles, the R l l - 3 reactive region appears to be nearly coextensive wi th the region stained by an unc-54- specific monoclonal antibody. This colocalization is ob- served even in a mutan t where the unc-54 myosin is present but is not part of a normal A-band (Fig. 7).
Mutan t animals were again used to determine the contr ibut ion of the unc-22 gene to the observed signal (Fig. 6). In ct37 animals, the antibody staining is drasti- cally reduced in the body wall, vulval, and anal muscles. In contrast, the pharyngeal signal is identical to wild type. The slight residual staining of the body wall muscles in wild type is confined to narrow strips be- tween each row of dense bodies. The pattern obtained
wi th the ant i -unc-54 antibody is abnormal, but the signal remains strong. Similar results were obtained wi th the antibody when used on most st136::Tcl an- imals.
unc-22 protein expression is ceil autonomous
Because Tcl is known to excise somatically (Emmons and Yesner 1984; Eide and Anderson 1985), we elected to see if we could identify animals in which one or a few muscle cells contained normal or nearly normal levels of the antibody reactive protein. In previous work we had obtained a strain containing st136::Tcl in which germ line excision is exceedingly rare (< 10-7), yet somatic ex- cision, as judged by Southern analysis of exponential ly
98 GENES & DEVELOPMENT
Cold Spring Harbor Laboratory Press on August 31, 2018 - Published by genesdev.cshlp.orgDownloaded from

Molecular characterization of unc-22 gene
Figure 6. Subcellular localization of the unc-22 protein relative to the unc-54 myosin heavy chain protein. Wild-type (a,b) or unc-22(ct37) mutant animals (c,d) were prepared for immunofluorescence, as described in Francis and Waterston (1985) and Materials and methods. They were then double stained with the monoclonal antibodies DM28.2 (Miller et al. 1983) and DM5-8, which are specific for the unc-54 myosin heavy chain protein (a,c) and with the Rll-3 serum (b,d). The Rll-3 staining in wild type (b) is localized to the same region as the unc-54 myosin (a). Overall, the A-bands are severely disorganized in the mutant, as shown by the myosin distribution (c), and the R11-3 staining is greatly reduced compared to wild type (d). The residual signal in the mutant is midway between the rows of dense bodies (not stained). The large arrows in a and b point to the center of the same A-band. Smaller arrows indicate the cell boundaries. The longitudinal axis of the nematode is horizontal. Bar, 20 ~m.
growing popula t ions , is as h igh as tha t in m u t a t o r s t rains (Moerman and Waters ton 1984; M o e r m a n et al. 1986). In f ragments prepared f rom random popula t ions of this st136::Tcl strain, we observed rare specimens w i th musc le cells tha t had strong R11-3 reac t iv i ty dis- t r ibuted w i t h i n the cells in a s tr iated pat tern (Fig. 8). W i t h i n the same fragments, cells were also present that did not s ta in wi th the ant ibody, and the l imi t s of the s ta in ing clearly corresponded to musc le cell boundaries. As a control , the same an imals were also s tained wi th the unc-22-specif ic ant ibody, and all cells stained. In ad- di t ion, no ce l l - l imi ted s ta in ing was ever observed in wi ld- type an imals prepared the same way. In cells that reacted w i th bo th antibodies, the unc-54 and unc-22
prote ins appear to be colocalized even in ins tances where the A-bands are s o m e w h a t disorganized (Fig. 8).
D i s c u s s i o n
Genet ic s tudies indica ted tha t unc-22 migh t be an un- usua l ly large gene, bo th from the f requency of m u t a t i o n and the r ecombina t iona l dis tances be tween unc-22 al- leles. Our molecu la r studies conf i rm these earlier infer- ences. The genomic region compris ing the unc-22 gene m u s t be at least 20 kb, based on the rear rangements ana- lyzed. The major cluster of Tc l inser t ion sites is distrib- u ted over 17 kb, and the d iepoxybutane- induced dele- t ions extend the region a m i n i m u m of another 3 kb. D N A th roughou t the 20-kb region cont r ibutes to the es-
Figure 7. Colocalization of unc-22 and unc-54 proteins in a thick filament mutant. Shown is an animal homozygous for the domi- nant missense allele unc-54(st273) stained with DMS-8 (a), a monoclonal antibody specific to the unc-54 protein and R11-3 (b), a polyclonal antibody that reacts with the unc-22-encoded protein (see Dibb et al. 1985 for a fuller description of this class of uric-54 mutation). Homozygotes of st273 have severely disorganized thick filaments, as shown in a using the unc-54-specific monoclonal antibody. The unc-22 R11-3 serum reveals a similar subcelluar staining pattern (b) to that observed in a, which indicates that even in cells with disorganized A-bands the unc-22 protein and myosin are colocalized. Arrows in a and b point to similar position. Bar, 20 ~zm.
GENES & DEVELOPMENT 99
Cold Spring Harbor Laboratory Press on August 31, 2018 - Published by genesdev.cshlp.orgDownloaded from

M o e r m a n et al.
DM28.2
R I I -3
d 7
Figure 8. The unc-22 gene product is cell autonomous. Mosaic animals were detected when R11-3, the polyclonal antibody to the unc-22 protein (b,d) and DM28.2, a monoclonal antibody to the major myosin heavy chain (a,c), were used against unc-22(stI36::Tcl) animals. (a) Three body wall muscle cells stained with the myosin antibody; (b) the same three cells as in a treated with the unc-22 antibody; (c) the ends of two body wall muscle cells reacted with the myosin antibody; (d) the same muscle cells as in c treated with the unc-22 antibody. (b,d) Arrows indicate the edge of a nonrevertant muscle cell. Note that the unc-22-expressing cell in c has better organized A-bands than the nonrevertant cell (see also Fig. 6c). Bar, 10 ~m.
timated 14-kb transcript. The gene may be larger still, as the st139::Tcl insertion site lies several kilobase pairs to the right of the small deficiencies. However, this allele is unusual in that the resultant phenotype is very weak and Tcl sequences are not incorporated into the tran- script. Until the 5' end of the gene and its normal regula- tory sequences are defined, we cannot rule out the possi- bility that several large introns or an unusually large reg- ulatory region could place even the st139::Tcl site within the boundaries of the gene. The large stable tran- script detected indicates that most of the 20-kb region is comprised of exons, which is not atypical of C. elegans genes (Kramer et al. 1982; Karn et al. 1983; Blumenthal et al. 1984; Spieth and Blumenthal 1985). Most of this mRNA must be coding sequence to yield the protein product or products of the gene greater than 500,000 daltons, as defined by the antibody and the analysis of the mutants.
The genetic analysis had also suggested that the unc-22 protein might be associated with the myosin heavy-chain product of the unc-54 locus. Only rare al- leles of the unc-54 locus are able to suppress unc-22 mu- tants; no other loci were identified despite exhaustive screening (Moerman et al. 1982; D.G. Moerman, un- publ.). In addition, other rare alleles of the unc-54 locus partially mimic the unc-22 twitching phenotype. The two analyzed alleles of this class have small in-frame deletions that remove a section of the myosin rod (Dibb et al. 1985). The location of the unc-22 protein in the A-band, as determined by the antibody R11-3, supports the notion that the unc-54 and unc-22 proteins are asso- ciated, and studies on unc-54 missense mutants reveal that in mutants where the unc-54 protein is present but displaced from its normal A-band location, the unc-22 protein colocalizes with the unc-54 myosin (Fig. 7). This colocalization of the protein with unc-54 myosin in wild-type and mutant animals suggests that the unc-22 protein might bind directly to myosin or another compo- nent associated with it. Additional biochemical and morphological studies will be necessary to determine whether the unc-22 protein is indeed a component of the thick filament.
The R11-3 antibody not only detects reactive material
in the body wall muscles but also in the anal, vulval, and pharyngeal muscles. All of these muscles, including the pharyngeal, also react with the R11-3 antibody after it has been affinity purified against the fusion polypeptide (D.G. Moerman et al., unpubl.). In unc-22 mutants the antibody staining is significantly reduced in all the muscles except the pharyngeal muscles, where the signal is identical to wild type. These observations on mutants, including genetic mosaics (Fig. 8), clearly indi- cate that staining of the body wall, anal, and vulval muscles in wild-type animals is due to the presence of the unc-22 protein in these cells. That unc-22 mutants have no effect on pharyngeal staining is in agreement with earlier studies showing that pharyngeal pumping and pharyngeal muscle structure were normal in mul- tiple mutant isolates of unc-22 (Waterston et al. 1980). Apparently, an immunologically related protein is ex- pressed in the pharynx, which is not the product of the unc-22 locus. Our inability to detect this second protein on Western blots is perhaps not surprising considering that pharyngeal muscles account for less than 5% of the total muscle mass. The weak residual staining of the body wall muscles observed in unc-22 mutants may be due to this second protein, or possibly even a third. Probing low-stringency genomic Southern blots with unc-22 fragments has not, as yet, revealed any strongly cross-hybridizing bands (D.G. Moerman et al., unpubl.). However, there is precedent for differential expression among gene family members in nematode muscle: Myosin heavy chains are encoded by four genes, two of which are expressed in the pharyngeal muscle and two of which are expressed in the body wall muscle (Epstein et al. 1974; Waterston et al. 1982; Karn et al. 1984; Miller et al. 1984).
The twitching aspect of the phenotype of unc-22 mu- tants has been used to infer that the unc-22 gene product is involved in the regulation of muscle contraction. The location of the protein in the A bands seems to rule out the possibility that the unc-22 protein is part of the muscle membrane or the sarcoplasmic reticulum. Its size also eliminates the possibility that it is one of the known thin-filament regulatory proteins or one of the myosin light chains. One intriguing possibility consis-
100 GENES & DEVELOPMENT
Cold Spring Harbor Laboratory Press on August 31, 2018 - Published by genesdev.cshlp.orgDownloaded from

Molecular characterization of unc-22 gene
tent with our findings is that the unc-22 protein may modulate myosin activity directly.
The intracellular location of the unc-22 protein is dif- ferent from any of the few very large proteins identified in muscles of other animals. Z-nin (Suzuki et al. 1981) and zeugmatin (Mahler et al. 1985) are found in the Z- line region of vertebrate muscle. Another large protein, nebulin, is only slightly smaller than the unc-22 protein, but its location in the N-2 line of the I band (Wang and Williamson 1980) makes it an unlikely analog. Titin is much larger in size, with a molecular weight of more than 1,000,000 (Wang et al. 1979; Trinick et al. 1984). Its apparent position within the sarcomere seems to depend on sarcomere length, but at resting length the immuno- reactive groups are restricted to the A- to I-band junc- tion. Another potentially large protein discovered re- cently is the product of the Duchenne's muscular dys- trophy locus, which encodes an mRNA of about 16 kb (Monaco et al. 1986). However, no hybridization of the Duchenne's cDNA probes to the unc-22 genomic clones was detected even under low stringency (L. Kunkel, pets. comm.). In addition, partial sequences for both are known and they are not similar (unpubl.; L. Kunkel, pers. comm.). Whether any of these proteins or others are analogs of the unc-22 protein must await either the development of functional tests for these large proteins or more complete sequence comparisons.
The Tcl-induced alleles of unc-22 have been crucial at each step in our investigations and, in turn, our investi- gations of these alleles has revealed new aspects of Tcl biology. The wide distribution of insertion sites helped to demonstrate the size of the gene. Although the dis- perse distribution of these insertions might suggest that Tcl has little or no insertion site specificity, a much more limited set of insertion sites has been found for unc-54 (Eide and Anderson 1985), and our own sequence analysis of Tcl insertion sites demonstrates that Tcl has a preferred target sequence (Mori et al. 1987). Appar- ently the unc-22 gene is unusually rich in such sites.
The incorporation of Tcl sequences in whole or in part into a full-length stable message aided in defining the region of the gene contributing to mature mRNA and is an unusual finding for transposable elements. The incorporation is independent of Tcl orientation and largely independent of Tcl position within the gene. In five of the six cases we studied, the insertions are in ei- ther orientation and are spread over a region of about 12 kb. The only exception is the st139::Tcl allele, where a normal-sized message is made.
The incorporation of a transposable element into a full-length RNA has been reported for a Ds insertion ele- ment into the alcohol dehydrogenase (ADH) gene of maize (Doring et al. 1984), but, in general, insertion of a transposon into a gene results in a truncated message (Levis et al. 1984; Pirrotta and Brockl 1984). The inten- sity of signal from the full-length composite unc-22- Tcl transcript in the examples here suggests that very little, it any, termination is occurring within the Tcl sequence, but we have not ruled out that termination occurs either at a variety of sites or results in an un-
stable RNA that is not readily detected on Northern analysis. In fact, a few faint bands can be detected on long exposures of Northern transfers that are of a size consistent with their production via truncation within Tcl (G.M. Benian and D.G. Moerman, unpubl.). Never- theless, our results suggest that Tcl contains either no, or only very weak, transcription-termination signals.
The incorporation of Tcl sequences into the messages of genes with Tcl insertions has several practical impli- cations. First, it may serve to verify that a particular mRNA is the transcript of the gene under study, as was done here. Second, hybridization with a Tcl probe of a Northern transfer of total RNA from a mutant strain might be useful in verifying that a mutation represents a Tcl insertion. Such a composite RNA might even be used as an aid in cloning the gene in unusual circum- stances. At present, we do not know how often Tcl in- sertions will result in a composite mRNA. Results from an analysis of three unc-15::Tcl alleles reveal that none of these mutations generates a hybrid message (R.P. An- derson, S. Rioux, and R.H. Waterston, unpubl.). Recent sequence analysis of these mutations (S. Rioux and R.H. Waterston, unpubl.) and of the paramyosin gene (H. Ka- gawa, pers. comm.) suggests that these insertions lie near the 5' end of the gene. These alleles are non-null alleles and may be analogous to the st139 allele. On the other hand, Tcl insertions into tra-2 result in a tran- script larger than wild type (P. Okkema and J. Kimble, pers. comm.).
A puzzling aspect of the unc-22::Tcl alleles is that they are all phenotypically milder in their effects than are amber alleles or other likely null alleles. This is par- ticularly noticeable at the level of muscle structure (see Figs. 6 and 7; Moerman and Waterston 1984). Pre- viously, we postulated that the insertions might be in introns where they could be processed out (Moerman and Waterston 1984). Instead, the Tcl sequences are in- corporated into the stable transcript, and in the case of st136::Tcl, a likely truncated peptide has been found. Furthermore, the integration sites of the st136::Tcl and st137::Tcl alleles lie within long open reading frames, not introns (I. Mori, G.M. Benian, and D.G. Moerman, unpubl.). Another possible explanation is that somatic excision of Tcl, which results in the appearance of sev- eral percent of the signal at the size of the wild-type band, could result in the production of sufficient protein to yield a weak phenotype. However, the unc-22 product appears to be cell autonomous, and only very rare cells show restoration of near normal levels of antibody-reac- tive material. Apparently, many of the somatic excision events, as judged by Southern analysis, must produce out-of-frame deletions or unstable, nonfunctional poly- peptides. Another explanation is suggested by our present data: perhaps, the truncated polypeptide de- tected in st136::Tcl animals (Fig. 5) is sufficient to pro- duce discernible lattice organization but is unable to in- hibit twitching. In severe alleles, lattice structure is disrupted more severely (Moerman and Waterston 1984). This possibility of different functional domains within the unc-22 locus is supported by partial sequence anal-
GENES & DEVELOPMENT 101
Cold Spring Harbor Laboratory Press on August 31, 2018 - Published by genesdev.cshlp.orgDownloaded from

Moerman et al.
ysis cu r ren t ly under way (J. Kiff, I. Mori, R.H. Waters ton, and G.M. Benian, unpubl.) .
The t en ta t ive a s s ignmen t of unc-22 to the set of genes affect ing the th i ck f i l amen t is s ignif icant . In addi t ion to unc-22, the genes for bo th m y o s i n heavy-cha in isoforms and pa ramyos in have been discovered and subjected to a combined molecu la r and genet ic analysis . The genes for the m y o s i n l ight chains have been cloned (C. C u m m i n s and P. Anderson, pers. comm) and, once local ized to the genet ic map, m a y also be subjected to genet ic analysis . In addi t ion, several genes have t en t a t ive ly been assigned to the th i ck f i l amen t by genet ic criteria, inc lud ing unc-87, unc-82, and unc-89 (Waterston et al. 1980). Mu- ta tor - induced alleles of bo th unc-87 and unc-89 are available, and efforts are under way to recover D N A for these genes (D.G. M o e r m a n et al., unpubl.). A thorough s tudy of these and other genes ye t to be discovered m a y provide us w i t h a comple te picture of t h i ck f i l amen t as- s embly and funct ion .
M a t e r i a l s a n d m e t h o d s
Growth and description of nematode strains
Stock maintenance and strain growth for protein and nucleic acid preparations were as described in Brenner (1974) and Sul- ston and Brenner (1974). All unc-22::Tcl mutants were isolated in either the BO strain of C. elegans var. Bergerac (Nigon 1949; Liao et al. 1983; Moerman and Waterston 1984) or in strains derived from crosses between BO and the N2 strain of C. elegans vat. Bristol (Brenner 1974).
precipitated, washed with absolute ethanol, and dissolved in 0.1-0.3 ml of a solution containing 10 mM Tris and 0.1 mM EDTA at pH 8.
Fifteen micrograms of total RNA was electrophoresed through 0.7% agarose-formaldehyde gels {Maniatis et al. 1982) and transferred to nitrocellulose filters (Thomas 1980; Schleicher and Schuell, Inc., Keene, NH). Filters were pre-hy- bridized for several hours at 42°C in 50% formamide, 5 x SSC, 0.1% SDS, and 100-200 mg/ml denatured calf thymus DNA. This solution was discarded, and the filters were hybridized for 16 hr at 42°C to s2P-labeled DNA or RNA probes in the same buffer, with the addition of dextran sulfate to 10%. Extensive washings were done, initially in 2 x SSC and 0.1% SDS at room temperature, and then in 0.1 x SSC and 0.1% SDS at 55-60°C. Nitrocellulose filters were blotted dry and then exposed to Kodak XAR-2 film with an intensifying screen (DuPont Cronex Lightning-Plus, Dupont Co., Wilmington, DE) for 1-5 days at
- 70°C. To look for unc-22-encoded RNA, nick-translated ~ and
plasmid clones (described above) were used as hybridization probes. For determining the direction of unc-22 transcription, we used strand-specific M13 probes (Brown et al. 1982), which were derived from a 1.1-kb EcoRV fragment near the stI36::Tcl insert (Moerman et al. 1986). The vector used to clone these fragments was mWB3295 (Barnes and Bevan 1983). A 1.6-kb EcoRV Tcl fragment from pDM4 (Moerman et al. 1986) was nick-translated and used to probe the Northern blot shown in Figure 4. To determine the orientation of Tcl within the com- posite unc-22-Tcl transcripts, labeled RNA was prepared from two SP6 recombinants that contain Tcl in opposite orientation (Zinn et al. 1983). The myosin heavy-chain gene used for probing Northern blots was the 2.8-kb BamHI fragment of unc-54 (Karn et al. 1980).
Nucleic acid procedures
High-molecular-weight nematode genomic DNA was isolated following the protocol of Wills et al. (1983). Standard methods were used for restriction digestion, gel electrophoresis, and Southern blotting (Southern 1975; Maniatis et al. 1982) DNA probes were labeled by nick translation (Rigby et al. 1977), and hybridization conditions were as described by Goodman et al. (1977). DNA clones used for localizing the unc-22::Tcl inserts included ~DM17, ?~DM20, and kDM22 (Moerman et al. 1986), which span the 30-kb unc-22 region. In addition to these clones, several single SacI and BglII fragments from unc-22, cloned into pucl8 or pucl9 (Yanish-Perron et al. 1985), were used as hybrid- ization probes. Methods for growth and for DNA isolation from these phage and plasmid clones have been described (Karn et al. 1980; Maniatis et al. 1982).
Total RNA was prepared by a modification of the guanidine hydrochloride method of Chirgwin et al. (1979). A mixed popu- lation of nematodes was harvested from five 100-mm petri dishes, washed to remove E. coli, and frozen at - 70°C. Worms were ground in liquid nitrogen for 10 rain, using a mortar and pestle, and lyophilized for 5 hr. This powder was added to 4 ml of a solution containing 6 M guanidine hydrochloride, 0.5% Sarkosyl, 0.1 M 2-mercaptoethanol, and 0.1% antifoam A emul- sion (Sigma, St. Louis, MO), pH 7.0, and vortexed for 80 sec. Debris was removed by centrifugation at 8000 rpm in a Sorvall SS-34 rotor for 10 rain. The supernatant was then layered onto a 1.25-ml CsC1 pad [5.7 M CsC1, 0.1 M EDTA, pH 7.0] and centri- fuged in a Sorvall AH-650 rotor at 36,000 rpm for 13 hr. The supernatant was discarded, and the pellet was resuspended in 0.3 ml of guanidine hydrochloride solution (described above) by pipetting and heating at 65°C for 5 min. The RNA was ethanol-
Generation of the unc-22 fusion protein and antibody production
The ~, vector ~gt l l (Young and Davis 1983) was modified by oligonucleotide mutagenesis in order to eliminate the SacI re- striction site outside of the [3-galactosidase gene, thereby making the SacI site in the gene unique (R.J. Barstead and R.H. Waterston, unpubl.). Partial sequencing of a portion of the unc-22 gene, as depicted in Figure 1, allowed us to identify a SacI fragment that would produce an in-frame fusion between the [3-galactosidase of the modified vector, ~RB1, and this por- tion of unc-22.
The clone containing the unc-22 insert, KLS1, was used to establish a lysogen in the E. coli strain Y1089 (Young and Davis 1983). The phage can be induced by a heat pulse at 42°C to initiate a lyric cycle in this strain; however, the amber muta- tion S100 in the phage S gene prevents lysis of the cells. The lysogenic strain was grown at 30°C to an OD600 of 0.25 in L broth. The culture was then transferred to a 42-44°C water bath and grown for 15 min after its temperature reached the temperature of the bath. Isopropal-[3-D-thiogalactopyranoside (IPTG) was added to lmM, and growth was continued at 38°C for 1 hr. The cells were collected by centrifugation, resuspended in breaking buffer [50 mM NaH2POa, pH 7.4, 50 mM NaC1, 1 mM EDTA, 1 mM phenylmethylsulfonyl flouride (PMSF), 0.5 M [3-mercaptoethanol, and a mixture of protease inhibitors], and sonicated. The solution was spun at 18K for 15 min, at 4°C in a Sorvall SS-34 rotor. NHaSOa was added to 35%.
The NHaSOa precipitate containing the unv-22 fusion protein was collected by centrifugation. The 155-kD fusion protein, first identified by its reaction with an anti-f3-galactosidase serum (not shown; Cappel Laboratories), was further purified
102 GENES & DEVELOPMENT
Cold Spring Harbor Laboratory Press on August 31, 2018 - Published by genesdev.cshlp.orgDownloaded from

Molecular characterization of unc-22 gene
by preparative SDS-PAGE (Laemmli 1970). Samples were run on 5% polyacrylamide gels, which were fixed and stained using standard procedures. The band containing the fusion protein was excised, neutralized, and passed through a French Press in 0.15 M NaH2PO4 (pH 7.4). The disrupted gel was then emulsi- fied with two volumes of adjuvant by sonication. A New Zea- land white rabbit was immunized with three subcutaneous in- jections over the course of a month with -50 ~g of fusion pro- tein/injection. Immune serum was collected 7 days after the last injection. Anti-~-galactosidase activity was eliminated by adsorption to E. coli [5-galactosidase (Sigma), coupled to cy- anogen bromide (CNBr)-activated Sepharose (Sigma).
Western transfer procedures
To prepare protein samples, 0.1 g of freshly grown nematodes was suspended in 1 ml of a solution containing 50 mM 2-amin- oethanol (pH 11 ), 5 mM dithiothreitol (DTT), 2 mlvi EDTA, and I mM PMSF. The samples were then heated in a microwave oven for 25 sec and transferred to a boiling water bath for 3 min. Laemmli sample buffer (1 ml) was added to each, and the samples were immediately loaded onto a 5% acrylamide-SDS gel (Laemmli 1970). The gels were either stained with 0.05% Coomassie Brilliant Blue R-250 using standard conditions or transferred electrophoretically to a nitrocellulose filter (Towbin et al. 1979). Transfer conditions were as described by Francis and Waterston 11985), except the electrode buffer contained 0.1% SDS. Mter electroblotting, the nitrocellulose filters were preincubated for 15 rain in Tris-buffered saline (TBS: 50 mM Tris-HC1, pH 7, 150 mM NaC1) with 0.1% Tween-20 (Batteiger et al. 1982) and 5% nonfat dried milk (Johnson et al. 1984) as blocking agents. Filters were then incubated with gentle shaking for 1 hr in a sealed plastic bag containing this buffer and a 170-fold dilution of the R11-3 serum. After washing away unbound antibody with the same buffer, the filters were incu- bated with 1 jxCi of affinity-purified ~2SI-labeled protein A (Amersham, Arlington Heights, IL) for 1 hr. Once again, the filters were washed, as described above, and exposed at -70°C to Kodak XAR-2 film with an intensifying screen.
Immunofluorescence techniques
Nematodes were partially sheared in a French Press, according to the procedure of Francis and Waterston (1985), in the fol- lowing rigor buffer: 100 mM KC1, 10 mM KHePO 4 (pH 7.0), 2 m ~ MgCle, 1 mM EDTA, 1 mM DTT, and 1 mM PMSF. They were washed twice in rigor buffer containing 1% NP-40 and fixed at room temperature for 3 hr to overnight in 3% formalde- hyde made up by dilution of formalin in phosphate-buffered sa- line (PBS) (pH 7), with the addition of 5 mM EDTA. The mono- clonal antibody DM28.2 is specific for the unc-54 myosin heavy-chain peptide and is described in Miller et al. (1983). The monoclonal antibody DM5-8 is similar to DM28.2 (D.M. Miller, pers. comm.). Tetramethylrhodamine isothiocyanate- conjugated goat-anti-mouse and fluorescein isothyocyanate- conjugated goat-anti-rabbit secondary antibodies were ob- tained from Jackson Immunoresearch Labs, Inc. {Avondale, Pennsylvania). Antibody incubations were done as in Francis and Waterston (1985). Stained specimens were viewed with a Zeiss Universal microscope (Carl Zeiss, Inc., Thornwood, New York), equipped with epifluorescent illumination. Photomicro- graphs were prepared as in Francis and Waterston (1985).
A c k n o w l e d g m e n t s
The authors gratefully acknowledge Ross Francis for his advice
regarding protein and immunochemical methods. We thank Carol Trent for providing three unc-22 internal deficiency strains, Kuan Wang for his gift of purified titin and nebulin, David Miller for providing a monoclonal antibody to the myosin heavy chain of unc-54, and Wayne Barnes for providing the plasmid mWB3295. We thank Eric Ward for introducing us to dideoxy sequencing. We would also like to thank the fol- lowing people for allowing us to cite their unpublished results: Hiro Kagawa and Jon Karn, Stephen Rioux, Louis Kunkel, Claudia Cummins and Phil Anderson, Ikue Mori, Jane Kiff, and Peter Okkema and Judith Kimble. Secretarial assistance was provided by Kay Webb. R.J.B. was supported by a postdoctoral fellowship from the Muscular Dystrophy Association. This re- search was supported by U.S. Public Health Service grant GM23883 and a Muscular Dystrophy Association Jerry Lewis Neuromuscular Research Center grant to R.H.W.
R e f e r e n c e s
Barnes, W.M. and M. Bevan. 1983. Kilo-sequencing: An ordered strategy for rapid DNA sequence data acquisition. Nucleic Acids Res. 11: 349-368.
Batteiger, B., W.J. Newhall, and R.R. Jones. 1982. Use of Tween 20 as a blocking agent in the immunological detection of proteins transferred to nitrocellulose membranes. J. Im- munol. Methods 55: 297-307.
Blumenthal, T., M. Squire, S. Kirtland, J. Cane, M. Donegan, J. Spieth, and W. Sharrock. 1984. Cloning of a yolk protein gene family from Caenorhabditis elegans. J. Mol. Biol. 174: 1-18.
Brenner, S. 1974. The genetics of Caenorhabditis elegans. Ge- netics 77: 71-94.
Brown, D.M., J. Frampton, P. Goelet, and J. Kam. 1982. Sensi- tive detection of RNA using strand-specific M13 probes. Gene 20: 139-144.
Chirgwin, J.M., A.E. Przybyla, R.J. MacDonald, and W.J. Rutter. 1979. Isolation of biologically active ribonucleic acid from sources enriched in ribonuclease. Biochemistry 18" 5294- 5299.
Coulson, A., J. Sulston, S. Brenner, and J. Karn. 1986. Toward physical map of the genome of the nematode Caenorhab- ditis elegans. Proc. Natl. Acad. Sci. 83: 7821-7825.
Dibb, N.J., D.M. Brown, J. Karn, D.G. Moerman, S.L. Bolten, and R.H. Waterston. 1985. Sequence analysis of mutations that affect the synthesis, assembly, and enzymatic activity of the unc-54 myosin heavy chain of Caenorhabditis elegans. J. Mol. Biol. 183: 543-551.
Doring, H.P., M. Freeling, S. Hake, M.A. Johns, R. Kunze, A. Merchelbach, F. Salamini, and P. Starlinger. 1984. A Ds-mu- tation of the Adhl gene in Zea mays L. Mol. Gen. Genet. 193: 199-204.
Eide, D. and P. Anderson. 1985. The gene structure of sponta- neous mutations affecting a Caenorhabditis elegans myosin heavy chain gene. Genetics 109: 67-69.
Emmons, S.W. and L. Yesner. 1984. High frequency excision of transposable element Tcl in the nematode Caenorhabditis elegans is limited to somatic cells. Cell 36: 599-605.
Epstein, H.F., R.H. Waterston, and S. Brenner. 1974. A mutant affecting the heavy chain of myosin in C. elegans. J. Mol. Biol. 90: 291-300.
Francis, G. and R. Waterston. 1985. Muscle organization in Caenorhabditis elegans: Localization of proteins implicated in thin filament attachment and I-band organization. J. Cell Biol. 101: 1532-1549.
Goodman, H.M., M.V. Olson, and B.D. Hall. 1977. Nucleotide sequence of a mutant eukaryotic gene: The yeast tryosine-
GENES & DEVELOPMENT 103
Cold Spring Harbor Laboratory Press on August 31, 2018 - Published by genesdev.cshlp.orgDownloaded from

Moerman et al.
inserting ochre suppressor sup4-O. Proc. Natl. Acad. Sci. 74: 5453-5457.
Greenwald, I. 1985. lin-12, a nematode homeotic gene, is ho- mologous to a set of mammalian proteins that includes growth factor. Cell 43: 583-590.
Johnson, D.A., J.W. Gautsch, J.R. Sportsman, and J.H. Elder. 1984. Improved techniques utilizing nonfat dry milk for analysis of proteins and nucleic acids transferred to nitrocel- lulose. Gene Anal. Technol. 1: 3-8.
Karn, J., S. Brenner, and L. Barnett. 1983. Protein structural do- mains in the Caenorhabditis elegans unc-54 myosin heavy chain gene are not separated by introns. Proc. Natl. Acad. Sci. 80: 4253-4257.
Karn, J., N.J. Dibb, and D.M. Miller. 1984. Cloning nematode myosin genes. In Cell and muscle motili ty V/(ed. J. Shay) pp. 185-237. Plenum, New York.
Karn, J., S. Brenner, L. Barnett, and G. Cesareni. 1980. Novel bacteriophage K cloning vector. Proc. Natl. Acad. Sci. 77: 5172-5176.
Kramer, J.M., G.N. Cox, and D. Hirsh. 1982. Comparisons of the complete sequences of two collagen genes from Caenor- habditis elegans. Cell 30: 599-606.
Laemmli, U.K. 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227: 680- 685.
Landel, C.P., M. Krause, R.H. Waterston, and D. Hirsh. 1984. DNA rearrangements of the actin gene cluster in Caenor- habditis elegans accompany reversion of three muscle mu- tants. J. Mol. Biol. 180: 497-513.
Levis, R., K. O'Hare, and G.M. Rubin. 1984. Effects of transpos- able element insertions on RNA encoded by the white gene of Drosophila. Cell 38: 471-481.
Lewis, J.A., C.-H. Wu, J.H. Levine, and H. Berg. 1980. Leva- misole resistant mutants of the nematode Caenorhabditis elegans appear to lack pharmacological acetylcholine re- ceptors. Neuroscience 5: 967-989.
Liao, L.W., B. Rosenzweig, and D. Hirsh. 1983. Analysis of a transposable element in Caenorhabditis elegans. Proc. Natl. Acad. Sci. 80: 3583-3589.
Losson, R. and F. LaCroute. 1979. Interference of nonsense mu- tations with eukaryotic messenger RNA stability. Proc. Natl. Acad. Sci. 76: 5134-5137.
MacLeod, A.R., R.H. Waterston, and S. Brenner. 1977a. An in- ternal deletion mutant of a myosin heavy chain in C. elegans. Proc. Natl. Acad. Sci. 74: 5336-5340.
MacLeod, A.R., R.H. Waterston, R.M. Fishpool, and S. Brenner. 1977b. Identification of the structural gene for a myosin heavy chain in C. elegans. J. Mol. Biol. 114: 133-140.
Mahler, P.A., G.F. Cox, and S.J. Singer. 1985. Zeugmatin: A new high molecular weight protein associated with Z lines in adult and early embryonic striated muscle. J. Cell Biol. 101: 1871-1883.
Maniatis, T., E.F. Fritsch, and J. Sambrook. 1982. Molecular cloning: A laboratory manual. Cold Spring Harbor Labora- tory, Cold Spring Harbor, New York.
Miller, D.M., I. Ortiz, G.C. Berliner, and H.F. Epstein. 1983. Differential localization of two myosins within nematode thick filaments. Cell 34: 477-490.
Miller, D.M., F.R. Stockdale, and J. Karn. 1986. Immunological identification of the genes encoding four myosin heavy chain isoforms of Caenorhabditis elegans. Proc. Natl. Acad. Sci. 83: 2305-2309.
Moerman, D.G. 1980. "Genetic analysis of the unc-22 region in Caenorhabditis elegans." Dissertation. Simon Fraser Uni- versity, Burnaby, B.C.
Moerman, D.G. and D.L. Baillie. 1979. Genetic organization in
Caenorhabditis elegans: Fine-structure analysis of the unc-22 gene. Genetics 91" 95-103.
Moerman, D.G. and R.H. Waterston. 1984. Spontaneous un- stable unc-22 IV mutations in Caenorhabditis elegans var. Bergerac. Genetics 108: 859-877.
Moerman, D.G., S. Plurad, R.H. Waterston, and D.L. Baillie. 1982. Mutations in the unc-54 myosin heavy chain gene of Caenorhabditis elegans that alter contractility but not muscle structure. Cell 29: 773-781.
Moerman, D.G., G.M. Benian, and R.H. Waterston. 1986. Mo- lecular cloning of the muscle gene unc-22 in Caenorhab- ditis elegans by Tcl transposon tagging. Proc. Natl. Acad. Sci. 83: 2579-2583.
Monaco, A.P., R.L. Neve, C. Colletti-Feener, C.J. Bertelson, D.M. Kurnit, and L.M. Kunkel. 1986. Isolation of candidate cDNAs for portions of the Duchenne muscular dystrophy gene. Nature 323: 646-650.
Mori, I., G.M. Benian, D.G. Moerman, and R.H. Waterston. 1987. Transposable element Tcl of Caenorhabditis elegans recognizes specific target sequences for integration. Proc. Natl. Acad. Sci. (in press).
Nigon, V. 1949. Les modalites de la reproduction et le deter- minisme du sexe chez quelques nematodes libres. Ann. Sci. Natl. Zool. 11: 1-132.
Pirrotta, V. and C. Brockl. 1984. Transcription of the Droso- phila white locus and some of its mutants. EMBO J. 3(3): 563-568.
Rigby, P.W.J., M. Dieckmann, C. Rhodes, and P. Berg. 1977. La- beling deoxyribonucleic acid to high specific activity in vitro by nick translation with DNA polymerase I. J. Mol. Biol. 113: 237-251.
Rogalski, T.M. and D.L. Baillie. 1985. Genetic organization of the unc-22 IV gene and the adjacent region in C. elegans. Mol. Gene. Genet. 201: 409-414.
Rosenzweig, B., L.W. Liao, and D. Hirsh. 1983. Sequence of the C. elegans transposable element Tcl. Nucleic Acids Res. 11(12): 4201-4209.
Rowland, L.J. and J.N. Strommer. 1985. Insertion of an unstable element in an intervening sequence of maize Adhl affects transcription but not processing. Proc. Natl. Acad. Sci. 82: 2875-2879.
Southern, E.M. 1975. Detection of specific sequences among DNA fragments separated by gel electrophoresis. J. Mol. Biol. 98: 503-517.
Spieth, J. and T. Blumenthal. 1985. The Caenorhabditis elegans vitellogenin gene family includes a gene encoding a dis- tantly related protein. Mol. Cell. Biol. 5(10): 2495-2501.
Sulston, J.E. and S. Brenner. 1974. The DNA of Caenorhabditis elegans. Genetics 77: 95-104.
Suzuki, A., M. Saito, A. Okitani, and Y. Nonami. 1981. Z-nin, a new high molecular weight protein required for reconstitu- tion of the Z-disk. Agric. Biol. Chem. 45: 2535-2542.
Thomas, P.S. 1980. Hybridization of denatured RNA and small DNA fragments transferred to nitrocellulose. Proc. Natl. Acad. Sci. 77: 5201-5205.
Towbin, H., T. Staehelin, and J. Gordon. 1979. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellu- lose sheets: Procedure and some application. Proc. Natl. Acad. Sci. 76: 4350-4354.
Trinick, J., P. Knight, and A. Whiting. 1984. Purification and properties of native titin. J. Mol. Biol. 180: 331-356.
Wang, K. and C.L. Williamson. 1980. Identification of an N2 line protein of striated muscle. Proc. Natl. Acacl. Sci. 77: 3254-3258.
Wang, K., J. McClure, and A. Tu. 1979. Titin: Major myofi- brillar components of striated muscle. Proc. Natl. Acad. Sci.
104 GENES & DEVELOPMENT
Cold Spring Harbor Laboratory Press on August 31, 2018 - Published by genesdev.cshlp.orgDownloaded from

Molecular characterization of uric-22 gene
76: 3698-3702. Waterston, R.H., R.M. Fishpool, and S. Brenner. 1977. Mutants
affecting paramyosin in Caenorhabditis elegans. ]. Mol. Biol. 111: 679-697.
Waterston, R.H., J.N. Thomson, and S. Brenner. 1980. Mutants with altered muscle structure in Caenorhabditis elegans. Dev. Biol. 77: 271-302.
Waterston, R.H., K.C. Smith, and D.G. Moerman. 1982. Ge- netic fine structure analysis of the myosin heavy chain gene unc-54 of Caenorhabditis elegans. J. Mol. Biol. 158: 1-15.
Wills, N., R.F. Gesteland, J. Karn, L. Barnett, S. Bolten, and R.H. Waterston. 1983. The genes sup-7 X and sup-5 III of C. elegans suppress amber nonsense mutations via altered transfer RNA. Cell 33: 575-583.
Yanisch-Perron, C., J. Vieira, and J. Messing. 1985. Improved M13 phage cloning vectors and host strains: Nucleotide se- quences of the M13mpl8 and pUC19 vectors. Gene 33: 103-119.
Young, R.A. and R.W. Davis. 1983. Efficient isolation of genes by using antibody probes. Proc. Natl. Acad. Sci. 80: 1194- 1198.
Zachar, Z., D. Davison, D. Garza, and P.M. Bingham. 1985. A detailed developmental and structural study of the tran- scriptional effects of insertion of the copia transposon into the white locus of Drosophila melanogaster. Genetics 111: 495-515.
Zengel, J.M. and H.F. Epstein. 1980. Identification of genetic elements associated with muscle structure of the nematode Caenorhabditis elegans. Cell Motil. 1: 73-97.
Zinn, K., D. DiMaio, and T. Maniatis. 1983. Identification of two distinct regulatory regions adjacent to the human B-in- terferon gene. Cell 34: 865-879.
GENES & DEVELOPMENT 105
Cold Spring Harbor Laboratory Press on August 31, 2018 - Published by genesdev.cshlp.orgDownloaded from

10.1101/gad.2.1.93Access the most recent version at doi: 2:1988, Genes Dev.
D G Moerman, G M Benian, R J Barstead, et al. of Caenorhabditis elegans.Identification and intracellular localization of the unc-22 gene product
References
http://genesdev.cshlp.org/content/2/1/93.full.html#ref-list-1
This article cites 59 articles, 23 of which can be accessed free at:
License
ServiceEmail Alerting
click here.right corner of the article or
Receive free email alerts when new articles cite this article - sign up in the box at the top
Copyright © Cold Spring Harbor Laboratory Press
Cold Spring Harbor Laboratory Press on August 31, 2018 - Published by genesdev.cshlp.orgDownloaded from