ichthyofauna of a near-natural pyrenean river: spatio-temporal
TRANSCRIPT

Environmental Engineering and Management Journal June 2012, Vol.11, No. 6, 1111-1124
http://omicron.ch.tuiasi.ro/EEMJ/
“Gheorghe Asachi” Technical University of Iasi, Romania
ICHTHYOFAUNA OF A NEAR-NATURAL PYRENEAN RIVER:
SPATIO-TEMPORAL VARIABILITY AND REACH-SCALE HABITAT
Pedro M. Leunda1, Manu Sistiaga2, Javier Oscoz1, Rafael Miranda1
1Department of Zoology and Ecology, University of Navarra, PO Box 177, E-31080, Pamplona/Iruña, Navarra, Spain 2SINTEF Fisheries and Aquaculture AS, Postbox 4762 Sluppen, NO-7465 Trondheim, Norway
Abstract The characterization of fish communities and their relationships with abiotic factors is necessary for a number of environmental management issues. This study aims to analyze the spatio-temporal variability in the structure of fish communities in an undisturbed and unpolluted Pyrenean river in relation to aquatic habitat features. Fish and habitat were sampled seasonally in 2002 at fourteen sampling sites along the Erro River, a tributary of the Ebro basin (Navarra, northern Spain). Fish communities comprised eight native species, and their capture efficiency estimates were high and consistent along the river. Differences between species in estimated capture probability values were related to attributes such as body size and/or abundance, underscoring the importance of using species-specific capture probabilities. The observed low variability of species capture probabilities allows their use as reference values for future population size estimations based on single-run electrofishing surveys in similarly sized rivers. The longitudinal fish species distribution was analyzed by means of separate hierarchical cluster analyses performed on total species’ densities and species’ densities partitioned by size-classes, both of which displayed similar patterns of fish community composition along the Erro River. The changes on species presence and relative contributions to local fish communities along the river were also evident from diversity indices, which showed strong and significant positive relationships with distance from the source. Seasonal values in diversity indices were also significantly different within sampling sites, indicating that low-order streams support lower diversity and less-structured fish communities because seasonal flow variations lead to high variability in hydrochemical parameters, habitat size and diversity. Two separate canonical correspondence analyses were performed to jointly summarize the variation in aquatic habitat parameters in relation to fish community size structure in terms of densities or biomasses. In both analyses, the first axis was related to the along-river gradient and the second axis accounted for seasonal variation. The relative positions of fish species along the spatio-temporal gradients are in accordance with their habitat preferences described in the Pyrenean area. These results provide a framework for understanding how the spatio-temporal variations of reach-scale aquatic habitat features structure fish communities in a Pyrenean river with low anthropogenic disturbance. Key words: aquatic habitat, Erro River, fish communities, Iberian Peninsula, longitudinal zonation, seasonality Received: December, 2011; Revised final: May, 2012; Accepted: June, 2012
Author to whom all correspondence should be addressed: e-mail: [email protected]
1. Introduction Characterization of fish communities and their
habitats is a basic requirement for a number of environmental management issues in fluvial ecosystems. Knowledge and understanding of the links between fish species and their habitat enhance the chances of successful fishery management plans (Naiman and Latterell, 2005), ecological restoration
projects (Bond and Lake, 2003; Chessman et al., 2006), conservation plans for threatened species (Naiman and Latterell, 2005) and water quality bioassessments (DeAngelis and Curnutt, 2002; Schmutz et al., 2007). However, fish communities and their relationships with habitat features are highly variable both in space and time, making their study complex and difficult to extrapolate (Lammert and Allan, 1999). This variability is hierarchically nested

Leunda et al./Environmental Engineering and Management Journal 11 (2012), 6, 1111-1124
1112
across spatial (from microhabitat to ecoregions) and temporal (from diel to geological) scales, and the relative importance attributed to the different abiotic (chemical and physical) factors in structuring fish communities is dependent on the spatio-temporal resolution of the study (Jackson et al., 2001; Tejerina-Garro et al., 2005). Such scale-related differences in the estimation of the relative importance of abiotic factors are the result of the broad range of the measured variables that large-scale studies are able to encompass. In contrast, small-scale studies often analyze data that are less spatially variable but have higher resolution (Jackson et al., 2001). In fact, large-scale studies of stream fishes tend to detect and emphasize biogeographic, climatic and historical abiotic factors, whilst locally more variable abiotic factors are more influential at finer spatial resolutions. Even though large- and small-scale approaches are complementary and relevant for ecological science, knowledge achieved from research at finer spatio-temporal resolutions may be more interesting and useful for dealing with certain environmental management issues.
Regarding the spatial scale, drainage basins or river systems share features of large environmental abiotic factors (biogeography, climate, etc.); therefore, the longitudinal source of variation emerges as the most influential factor shaping fish community composition and structure. The strong longitudinal pattern observed in river ecosystems is a consequence of the great length of rivers relative to their width (Jackson et al., 2001).
Studies focused at river scale that examine its entire length usually identify differences in fish communities associated with longitudinal patterns in habitat features, such as water flow and current velocity, substrate composition, water depth, cover and temperature. Therefore, the along-river succession in fish communities is attributed to adaptations to local habitat characteristics associated with the longitudinal variation of abiotic factors (e.g., Vannote et al., 1980). Jackson et al. (2001) recognize that much of the current knowledge about the importance of environmental gradients in shaping riverine fish communities come from studies carried out along rivers at small-to-intermediate spatial scales. However, small-scale abiotic factors and their correlation with fish communities are more variable in time; therefore, their study requires surveys designed to account for temporal variability.
With respect to the temporal dimension, seasonal variation is highly influential because it is directly linked to the life cycles of fish species (e.g., growth and reproductive period) and causes seasonal changes in habitat preferences, availability and use (e.g., Schlosser, 1982). Moreover, the different life-stages of some fish may require different habitats to complete the life cycle, as in the case of potamodromous species that require migratory movements within river systems.
Consequently, the analysis of habitat requirements needs to be conducted while taking into
account fish size, age and life-cycle stage (Santos et al., 2011).
Because of their relevance for ecological science and environmental management, abiotic factors have received much interest as determinants of the composition and structure of fish communities. Even if some of the variability patterns are commonly detected, local habitat and faunal characteristics lead to significant differences in the relative influence of specific abiotic factors, which make difficult the extrapolation between biogeographic areas.
The Iberian Peninsula is considered one of the freshwater fish biodiversity hotspots within Europe (Reyjol et al., 2007; Smith and Darwall, 2006) and its highly endemic freshwater fish fauna is composed mainly of small to medium-sized Cyprinid, Cobitid and Cyprinodontid species (Encina et al., 2006; Filipe et al., 2009; Granado-Lorencio, 1992; Hernando and Soriguer, 1992). Several studies have analyzed the spatio-temporal variation of the native fluvial fish fauna and its relationship with abiotic factors in the Iberian Peninsula (e.g., Benejam et al., 2008; Bravo et al., 2001; Vila-Gispert et al., 2002). Within the Iberian Peninsula, however, freshwater habitats and fish communities are diverse, and biogeographic studies usually distinguish the Ebro River basin from the other major river basins. At the peninsular scale, a biogeographic subgroup that includes the Ebro basin, the Cantabrian rivers and the eastern Pyrenees (Ebro–Cantabrian) is recognized (Ferreira et al., 2007; Hernando and Soriguer, 1992), and at a European scale, the Ebro basin is considered part of the Central Peri–Mediterranean region (Reyjol et al., 2007). At a global scale of analysis, the Ebro River is recognized as the central basin of the Eastern Iberia bioregion (Abell et al., 2008).
Few studies have been carried out on the spatio-temporal variability of native fish communities and their relationships with abiotic factors in the Pyrenean rivers draining to the Ebro River basin in Spain or to the Cantabric Coast–Languedoc bioregion in the French slope (Abell et al., 2008).
The Erro River in northern Spain is a near-natural Pyrenean tributary catchment within the Ebro River Basin. Previous studies have confirmed its good ecological status based on aquatic habitat assessment, hydrochemical parameters and benthic macroinvertebrate communities (Leunda et al., 2004; Leunda et al., 2009b; Oscoz et al., 2005b). These studies have furthermore pointed out that many reaches of the Erro River could be used as reference sites for Pyrenean rivers within the Ebro River Basin for the implementation of the European Community Water Framework Directive (WFD, 2000/60/CE). The objectives of the present study are (1) to describe the seasonal fish community structure and distribution patterns along the Erro River; (2) to evaluate the capture probability and efficiency estimates of the electrofishing surveys; and (3) to identify the most important aquatic habitat characteristics influencing the spatio-temporal variability of the fish community composition and structure.

Ichthyofauna of a near-natural Pyrenean river: spatio-temporal variability and reach-scale habitat
1113
2. Material and methods 2.1. Study area
This study was carried out in the Erro River, a 48.5-km-long tributary of the Irati River (Ebro Basin, Northern Spain). The Erro River flows north to south through a valley characteristic of the western Pyrenean valleys in Navarra Province. The river’s source is at an elevation of 1,125 m a.s.l. and meets the Irati River at 460 m a.s.l., draining an area of approximately 214 km2 (Fig. 1). The mean total annual flow is 167 Hm3, with a maximum of 599 Hm3 (1960/1961) and a minimum of 28 Hm3 (1948/1949) (time series 1932–2002, Confederación Hidrográfica del Ebro, 2011). The flow regime type is pluvio-nival, and the period of greatest discharge lasts from November until April (Bejarano et al., 2010).
Maximum winter (December to February) flows are related to oceanic rains with a daily mean flow of about 10 m3·s-1. The flow decreases of about c. 7 m3·s-1 during the spring, as winter rain discharges are substituted by melting waters. Finally, the summer low-flow period discharge remains below c. 1 m3·s-1 from July to September. The human population density in the Erro valley is low, with scattered settlements administratively attached to the villages of Erro (pop. 759), Lizoain (pop. 238) and Urroz (pop. 369) (north-to-south). The riparian vegetation along the Erro River is dominated by deciduous species (Salix alba, Salix eleagnos, Alnus glutinosa, Corylus avellana, Populus nigra, Fraxinus angustifolia, Acer campestre etc.), whereas reed (Scirpus sp.) and bulrush (Typha sp.) constitute the emergent vegetation.
Fourteen sampling sites (E01 to E14) along the Erro River were selected according to aquatic habitat representativeness, equidistant distribution and accessibility (Fig. 1). The sites E01 to E03 are located in the headwaters where 54% of the land is covered by deciduous forests, 17% is covered by scrubland and pastures cover 27%. The upper-middle Erro catchment area from E03 to E06 is mainly covered by deciduous (25%) and coniferous (39%) forests, with pastures (25%) and scrublands (10%) occupying the remaining land. A similar forest and scrubland cover is found in the lower-middle Erro (E07–E12), but crop fields (24%) replace pastures (4%) in this area. Crop fields (57%) and scrublands (18%) dominate the lower Erro (E13–E14), whereas deciduous (14%) and coniferous (5%) forests and pastures (5%) occupy the rest of the area. E02 is located 70 m upstream from the Sorogain Flow Gauging Weir (FGW). The lack of sampling sites between E03 and E04 is justified by the fact that summer drought affects this reach and the water infiltrates to flow through substrate interstices and the subsurface.
There are low-head dams (2–3 m crest height) between E02–E03 and E12–E13; and only the latter is equipped with a vertical-slot fish-way. E14 is affected by the channelized stretch of the Villaveta FGW (Miranda et al., 2005). A more detailed description of
the study area and sampling sites is available elsewhere (Leunda et al., 2004).
Fig. 1. Map of the western Pyrenean rivers in Navarra (Spain) showing the sampling sites along the Erro River
(Ebro Basin)
2.2. Field sampling
Electrofishing surveys were carried out seasonally (winter 2001–2002, summer 2002 and autumn 2002) at the fourteen sampling sites (E01–E14). During the summer surveys, fish were captured using streambank generator-powered electrofishing gear (ACUITEC, 300–600 V, 0.2–2 A), and a three-run (or two-run) depletion method was applied between two stop nets. In winter and autumn, a generator-powered backpack electrofishing unit (ACUITEC Martín Pescador III, 300–600 V, 0.2–2 A) was used, following a single-run depletion methodology between two stop nets (Lobón-Cerviá, 1991). The same electrofishing equipment and personnel were used in all surveys. To conduct sampling, one person with the anode pole moved upstream in a zigzag pattern, followed by two persons with dip-nets and another two persons carrying buckets. Surveys were performed in accordance with the procedures set out in CEN standard EN 14011 and with the corresponding authorizations from the Department of Rural Development and Environment of the Government of Navarra. Fish collected on each run were kept in different open retention cages so that they could be processed separately.
The walls of the cages were made from a plastic grid to ensure good water flow and they were always placed downstream to prevent the fish inside from suffering new electrical shocks. After being anaesthetized with 2-phenoxyethanol (0.5 ml·l-1), all the specimens were identified and measured (TL, total length to the nearest mm). Fish were released into open cages and then into the river after total recovery from anesthesia. Common and scientific names of fishes hereinafter follow Kottelat and Freyhof (2007) and Leunda et al. (2009a).
Common hydrochemical parameters, such as temperature (T, ± 0.1ºC), dissolved oxygen (Ox, ±

Leunda et al./Environmental Engineering and Management Journal 11 (2012), 6, 1111-1124
1114
0.01 mg·l-1) and its saturation (Os, ± 0.1%) (WTW-OXI 320/SET), conductivity (Cd, ± 1 μS·cm-1) and total dissolved solids (TDS, ± 1 mg·l-1) (WTW-LF 323/SET), and pH (RA-PHM210, ± 0.01) were measured in situ at each sampling sited in every sampling occasion. Physical aquatic habitat features were assessed along a variable number of transects at each sampling site using the transect-point method (Bain and Stevenson, 1999). Specifically, we measured total river reach length (± 0.1 m), river width at each transect (Wd, ± 0.1 m), and, at a variable number of points per transect: depth (Dp, ± 1 cm), water current velocity at 0.6 depth (V, ± 0.1 m·s-
1), presence of shade (SH), presence of aquatic vegetation (VG), and a visual estimation of the relative composition of substrate types. The number of transects per sampling site and the number of measurement points per transect depended on total reach length and width, accessibility and aquatic habitat heterogeneity (Bain and Stevenson, 1999). However, the number and exact location of each transect at a given site remained the same during the study period. 2.3. Statistical analyses
Population size (N) and capture probability (P’) for each fish species were estimated by analyzing summer electrofishing survey data. Depending on whether a three- or two-run depletion method was followed, the estimation formulas of Lelek (1974) or of Seber and Le Cren (1967) were used, respectively, because of their high precision and the simplicity of their calculation (in Lobón-Cerviá, 1991). Capture efficiency was estimated using the following indices: P1, percentage of fish caught in the first run (C1) in relation to the total catch (Ct); P2, percentage of fish caught in the first run (C1) in relation to N; and EM1, percentage of Ct in relation to N (in Lobón-Cerviá, 1991). In the single-run electrofishing surveys carried out in winter and autumn, the results were analyzed according to Seber and Le Cren (1967) (in Lobón-Cerviá, 1991) to estimate fish species population size ( N C1 P ' ). To increase survey sample size and improve estimation accuracy, survey capture efficiency indices and species-specific capture probabilities were obtained from the summer 2002 survey data set and from an additional survey data set from a previous study carried out in the summer 2001. Both data sets were collected in the same river reaches and sampled by the same team using the same methodology. Before pooling the data from both summer surveys to calculate the mean, capture efficiency values were compared by using the Mann–Whitney U-tests with exact p-values. The relationships between capture probabilities, efficiency indices, and river distance from source (DFS) were tested using Pearson correlation tests and least-squares linear regression analyses.
Population size estimates (N) were used to calculate fish densities (number of fish · 100 m-2) for each species. Fish densities were proportionately
distributed along the observed length-frequency distributions, and the density values were transformed into fish biomass estimates (g · 100 m-2) by using species-specific length-weight relationships available for the geographical area (Leunda et al., 2007; Leunda et al., 2006; Miranda et al., 2006; Oscoz et al., 2005a). Density and biomass estimates were partitioned into size-class intervals as follows: for Salmo trutta and Luciobarbus graellsii (1: < 50; 2: 50–149; 3: 150–249; 4: > 250 mm), for Barbus haasi, Parachondrostoma miegii and Gobio lozanoi (1: < 40; 2: 40–79; 3: 80–119; 4: > 120 mm), for Phoxinus bigerri and Barbatula quignardi (1: < 40; 2: 40–59; 3: 60–79; 4: > 80 mm) and for Cobitis calderoni (1: < 40; 2: 40–49; 3: 50–59; 4: > 60 mm). Partitioning fish into size classes makes it possible to analyze conspecific young-of-the-year, juveniles and adults separately, which is useful because spatial and trophic resource utilization varies ontogenetically for many stream fishes (Gillette et al., 2005; Santos et al., 2011).
From fish community compositional data, richness (Margalef’s index, DMg (S 1) ln N ),
diversity (Shannon–Wiener index, H ' pi ln pi )
and dominance (Simpson’s index, D p12 )
measures were calculated; where, S is the number of species, N is the total number of individuals in the sample, and pi is the proportion of individuals found for the ith species (Krebs, 1999; Magurran, 2004; Washington, 1984). Diversity measures for each site were compared between seasons based on a probability (P) value computed using bootstrapping as resampling procedure (1000 runs). The relationships between diversity indices and DFS were tested using Pearson correlation tests and least-squares linear regression analyses.
Hierarchical cluster analyses were performed to detect patterns of similarity in the compositions of fish communities along the Erro River. Two separate cluster analyses were carried out on each fish species’ density matrix and on the estimates partitioned by size-classes that were obtained during the summer sampling occasions. Horn’s overlap index was used as a quantitative similarity measurement for each pair of sampling occasions because it is recommended for abundance data (Legendre and Legendre, 1998). The Unweighted Pair-Group Method using Arithmetic Averages algorithm (UPGMA) was used to cluster the resulting similarity matrices.
Canonical Correspondence Analyses (CCA) were performed to jointly summarize the variation in fish community structure in relation to aquatic habitat parameters. CCA is a direct gradient multivariate analysis in which species abundances are considered to be a direct response to the gradient in environmental variables (teer Braak and Verdonschot, 1995).
The fish community structure matrices used for the CCA consisted of fish estimates partitioned into size-class intervals (8 species x 4 size-classes) for the 14 sampling sites in the three seasonal surveys (42
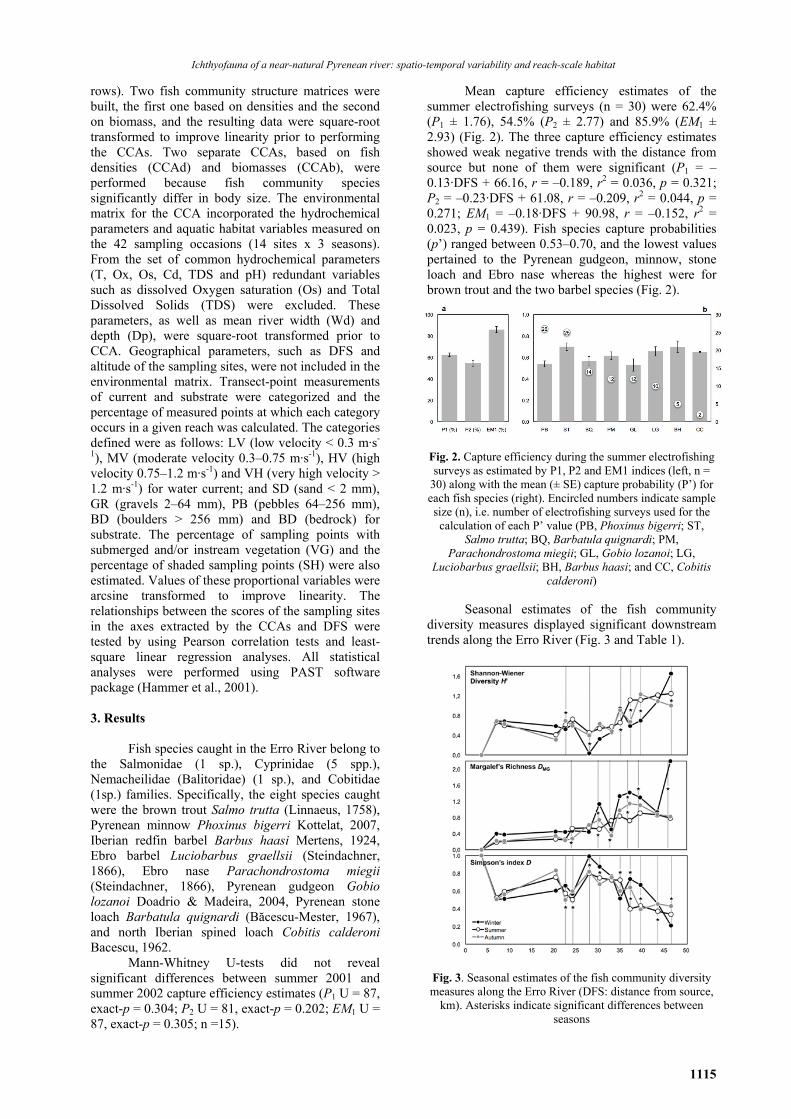
Ichthyofauna of a near-natural Pyrenean river: spatio-temporal variability and reach-scale habitat
1115
rows). Two fish community structure matrices were built, the first one based on densities and the second on biomass, and the resulting data were square-root transformed to improve linearity prior to performing the CCAs. Two separate CCAs, based on fish densities (CCAd) and biomasses (CCAb), were performed because fish community species significantly differ in body size. The environmental matrix for the CCA incorporated the hydrochemical parameters and aquatic habitat variables measured on the 42 sampling occasions (14 sites x 3 seasons). From the set of common hydrochemical parameters (T, Ox, Os, Cd, TDS and pH) redundant variables such as dissolved Oxygen saturation (Os) and Total Dissolved Solids (TDS) were excluded. These parameters, as well as mean river width (Wd) and depth (Dp), were square-root transformed prior to CCA. Geographical parameters, such as DFS and altitude of the sampling sites, were not included in the environmental matrix. Transect-point measurements of current and substrate were categorized and the percentage of measured points at which each category occurs in a given reach was calculated. The categories defined were as follows: LV (low velocity < 0.3 m·s-
1), MV (moderate velocity 0.3–0.75 m·s-1), HV (high velocity 0.75–1.2 m·s-1) and VH (very high velocity > 1.2 m·s-1) for water current; and SD (sand < 2 mm), GR (gravels 2–64 mm), PB (pebbles 64–256 mm), BD (boulders > 256 mm) and BD (bedrock) for substrate. The percentage of sampling points with submerged and/or instream vegetation (VG) and the percentage of shaded sampling points (SH) were also estimated. Values of these proportional variables were arcsine transformed to improve linearity. The relationships between the scores of the sampling sites in the axes extracted by the CCAs and DFS were tested by using Pearson correlation tests and least-square linear regression analyses. All statistical analyses were performed using PAST software package (Hammer et al., 2001). 3. Results
Fish species caught in the Erro River belong to the Salmonidae (1 sp.), Cyprinidae (5 spp.), Nemacheilidae (Balitoridae) (1 sp.), and Cobitidae (1sp.) families. Specifically, the eight species caught were the brown trout Salmo trutta (Linnaeus, 1758), Pyrenean minnow Phoxinus bigerri Kottelat, 2007, Iberian redfin barbel Barbus haasi Mertens, 1924, Ebro barbel Luciobarbus graellsii (Steindachner, 1866), Ebro nase Parachondrostoma miegii (Steindachner, 1866), Pyrenean gudgeon Gobio lozanoi Doadrio & Madeira, 2004, Pyrenean stone loach Barbatula quignardi (Băcescu-Mester, 1967), and north Iberian spined loach Cobitis calderoni Bacescu, 1962.
Mann-Whitney U-tests did not reveal significant differences between summer 2001 and summer 2002 capture efficiency estimates (P1 U = 87, exact-p = 0.304; P2 U = 81, exact-p = 0.202; EM1 U = 87, exact-p = 0.305; n =15).
Mean capture efficiency estimates of the summer electrofishing surveys (n = 30) were 62.4% (P1 ± 1.76), 54.5% (P2 ± 2.77) and 85.9% (EM1 ± 2.93) (Fig. 2). The three capture efficiency estimates showed weak negative trends with the distance from source but none of them were significant (P1 = –0.13·DFS + 66.16, r = –0.189, r2 = 0.036, p = 0.321; P2 = –0.23·DFS + 61.08, r = –0.209, r2 = 0.044, p = 0.271; EM1 = –0.18·DFS + 90.98, r = –0.152, r2 = 0.023, p = 0.439). Fish species capture probabilities (p’) ranged between 0.53–0.70, and the lowest values pertained to the Pyrenean gudgeon, minnow, stone loach and Ebro nase whereas the highest were for brown trout and the two barbel species (Fig. 2).
Fig. 2. Capture efficiency during the summer electrofishing surveys as estimated by P1, P2 and EM1 indices (left, n =
30) along with the mean (± SE) capture probability (P’) for each fish species (right). Encircled numbers indicate sample size (n), i.e. number of electrofishing surveys used for the calculation of each P’ value (PB, Phoxinus bigerri; ST,
Salmo trutta; BQ, Barbatula quignardi; PM, Parachondrostoma miegii; GL, Gobio lozanoi; LG,
Luciobarbus graellsii; BH, Barbus haasi; and CC, Cobitis calderoni)
Seasonal estimates of the fish community
diversity measures displayed significant downstream trends along the Erro River (Fig. 3 and Table 1).
Fig. 3. Seasonal estimates of the fish community diversity measures along the Erro River (DFS: distance from source,
km). Asterisks indicate significant differences between seasons

Leunda et al./Environmental Engineering and Management Journal 11 (2012), 6, 1111-1124
1116
Table 1. Linear regression parameters of seasonal fish community Shannon–Wiener diversity (H’), Margalef’s richness (DMG) and Simpson’s dominance (D) indices against distance from source (DFS) in the Erro River (a, slope; b, intercept; SE, standard
error; r, Pearson’s correlation; r2, coefficient of determination; p, probability; ns, not significant p > 0.05; * p < 0.05; ** p < 0.01; *** p < 0.001)
Season y = a·x + b a SE b SE r r2 p Winter H' = 0.017·DFS + 0.165 0.007 0.221 0.563 0.317 0.033 *
DMG = 0.036·DFS – 0.151 0.007 0.221 0.813 0.660 <0.001 *** D = –0.005·DFS + 0.787 0.004 0.132 –0.322 0.104 0.265 ns
Summer H' = 0.02·DFS + 0.164 0.005 0.143 0.774 0.598 0.002 ** DMG = 0.021·DFS – 0.036 0.002 0.066 0.939 0.882 <0.001 *** D = –0.008·DFS + 0.823 0.003 0.102 –0.563 0.317 0.038 *
Autumn H' = 0.017·DFS + 0.216 0.005 0.149 0.695 0.482 0.007 ** DMG = 0.024·DFS – 0.07 0.004 0.131 0.842 0.709 <0.001 *** D = –0.006·DFS + 0.791 0.003 0.099 –0.471 0.222 0.093 ns
Shannon–Wiener diversity (H’) and
Margalef’s richness (DMG) indices showed significant positive relationships with DFS of the sampling site in all three seasons. In contrast, the relationship between Simpson’s dominance index (D) and DFS was negative and weak, being significant only in summer. Significant differences were detected in the comparisons of seasonal diversity measures within each site. Specifically, significant differences were detected at E05, E07, E10–E12 and E14 for H’; at E06, E08, E09, E11, E12 and E14 for DMG; and at E05–E08 and E10–E14 for D.
The two separate hierarchical cluster analyses carried out on fish community composition displayed equivalent similarity patterns at the fourteen sampling sites (Fig. 4). The dendrogram constructed on fish species’ densities separates the sampling sites into two main groups at a 42% similarity level (Fig. 4, left). The upper six sites form the first group, from which the uppermost E01 departs at the 73% similarity level, whereas the rest split out at around 90% similarity in E02–E03 and E04–E06. Brown trout was the only species occurring at E01, but it co-
occurred with Pyrenean minnow at E02–E03. Below the drought-affected reach, E04 was again dominated by brown trout but Pyrenean minnow densities increased at E05–E06.
The second main group covered the lower eight sites, which were separated into two subgroups at a 77% similarity level in E07–E10 and E11–E14. In the subgroup just downstream of the gorge, E07–E09 group together because of the addition of Pyrenean stone loach and Iberian redfin barbel to the fish community; however, the latter species did not occur at E10 and hence splits from the other three sites of the subgroup. In the lowermost subgroup, localities grouped consecutively in a downstream fashion with the addition and increased density of more cyprinid species such as the Ebro nase, Ebro barbel and Pyrenean gudgeon (Fig. 4, left). E14 splits from the other three sites at about 90% similarity level.
The dendrogram built on fish species’ density estimates partitioned by size-classes displayed similar grouping and patterns along the Erro River, but was less well resolved (Fig. 4, right).
Fig. 4. Dendrograms resulting from the UPGMA cluster analysis of the Horn’s overlap index between the fourteen sampled reaches in the Erro River in summer 2002 according to the fish species’ densities (left) and to these estimates partitioned by size-
classes (right)

Ichthyofauna of a near-natural Pyrenean river: spatio-temporal variability and reach-scale habitat
1117
The first two axes of the CCAd and CCAb
jointly explained 62.46% and 64.96% of the total variance, respectively, for the relationships between fish community structure and aquatic habitat features (Table 2).
The first axis of the CCAd (CCAd1) had an eigenvalue of 0.387 and explained 44.46% of the variance in the species–habitat relationship, whereas CCAb1 had an eigenvalue of 0.531 and explained
51.32% of the variance. Regardless of the specific scores of the abiotic parameters on each CCA, the presence of aquatic vegetation, river width, conductivity, water temperature and bedrock substrate presented strong negative scores in both the CCAd1 and CCAb1 axes, whereas the presence of shade as well as sand and gravel substrates scored positive (Table 2).
Table 2. Factor loadings of each hydrochemical and aquatic habitat variable in the first four axes extracted by the canonical
correspondence analyses performed based on either fish densities (CCAd) or biomasses (CCAb)
Variable CCAd1 CCAd2 CCAd3 CCAd4 CCAb1 CCAb2 CCAb3 CCAb4 River Width Wd –0.594 0.383 –0.016 –0.389 –0.531 –0.224 0.334 0.117 Water Depth Dp –0.148 0.616 –0.432 0.081 –0.043 0.130 0.485 0.397
Dissolved Oxygen Ox –0.152 0.034 –0.025 0.083 –0.127 –0.026 0.048 0.146 pH pH –0.154 0.050 0.002 –0.021 –0.068 –0.074 –0.129 0.150
Conductivity Cd –0.564 0.514 0.013 –0.035 –0.476 0.048 0.425 0.209 Water Temperature T –0.429 0.057 0.073 0.094 –0.399 –0.018 –0.126 0.267 Presence of Shade SH 0.559 –0.087 –0.440 0.068 0.600 0.242 –0.193 0.135 Aquatic Vegetation VG –0.768 0.308 0.328 –0.271 –0.780 –0.337 0.461 –0.075
Sand Substrate SD 0.360 0.161 –0.201 –0.078 0.469 –0.038 –0.021 0.084 Gravel Substrate GR 0.434 –0.193 –0.069 0.264 0.338 0.354 –0.087 –0.272 Pebble Substrate PB –0.001 0.073 0.167 0.557 0.006 –0.013 0.166 0.146 Boulder Substrate BD 0.061 0.276 –0.109 –0.104 0.112 0.009 0.283 0.316 Bedrock Substrate BR –0.284 –0.233 –0.060 –0.442 –0.307 –0.187 –0.322 –0.093
Low Water Velocity LV –0.079 0.131 0.217 0.049 0.059 –0.330 –0.010 0.309 Moderate Water Velocity MV 0.144 –0.275 –0.008 –0.061 0.005 0.285 –0.034 –0.395
High Water Velocity HV –0.043 0.079 –0.276 –0.014 –0.091 0.184 0.041 –0.045 Eigenvalue 0.387 0.156 0.083 0.076 0.531 0.141 0.126 0.078 % Variance 44.46 17.98 9.58 8.75 51.32 13.64 12.14 7.57
% Cumulative Variance 44.46 62.44 72.02 80.77 51.32 64.96 77.10 84.67
Table 3. Linear regression parameters of seasonal fish community Shannon–Wiener diversity (H’), Margalef’s richness (DMG) and Simpson’s dominance (D) indices as well as distance from source (DFS) against sampling sites scores in the first axis
extracted by the canonical correspondence analyses performed based on either fish densities (CCAd1) or biomasses (CCAb1) (a, slope; b, intercept; SE, standard error; r, Pearson’s correlation; r2, coefficient of determination; p, probability; ns, not significant p
> 0.05; * p < 0.05; ** p < 0.01; *** p < 0.001)
Season y = a·x + b a SE b SE r r2 p DFS = –0.05·CCAd1 + 1.781 0.006 0.192 –0.915 0.837 <0.001 ***
DFS = –0.051·CCAb1 + 1.568 0.008 0.254 –0.866 0.750 <0.001 *** H' = –0.342·CCAd1 + 0.78 0.129 0.107 –0.608 0.369 0.019 *
H' = –0.294·CCAb1 + 0.691 0.126 0.099 –0.559 0.313 0.033 * DMG = –0.69·CCAd1 + 1.106 0.117 0.097 –0.862 0.744 <0.001 ***
DMG = –0.671·CCAb1 + 0.942 0.094 0.074 –0.899 0.809 <0.001 *** D = 0.109·CCAd1 + 0.601 0.078 0.065 0.371 0.138 0.187 ns
Winter
D = 0.08·CCAb1 + 0.632 0.075 0.059 0.294 0.086 0.306 ns DFS = –0.058·CCAd1 + 1.887 0.008 0.238 –0.904 0.818 <0.001 *** DFS = –0.052·CCAb1 + 1.546 0.010 0.286 –0.845 0.714 <0.001 *** H' = –0.355·CCAd1 + 0.821 0.057 0.050 –0.876 0.767 <0.001 *** H' = –0.365·CCAb1 + 0.759 0.062 0.050 –0.862 0.743 <0.001 ***
DMG = –0.317·CCAd1 + 0.623 0.037 0.033 –0.928 0.861 <0.001 *** DMG = –0.311·CCAb1 + 0.565 0.050 0.041 –0.873 0.762 <0.001 ***
D = 0.152·CCAd1 + 0.558 0.047 0.042 0.683 0.467 0.008 **
Summer
D = 0.155·CCAb1 + 0.584 0.050 0.040 0.670 0.449 0.010 * DFS = –0.051·CCAd1 + 1.773 0.008 0.233 –0.885 0.783 <0.001 *** DFS = –0.044·CCAb1 + 1.465 0.008 0.226 –0.862 0.743 <0.001 *** H' = –0.346·CCAd1 + 0.803 0.067 0.056 –0.832 0.692 <0.001 *** H' = –0.377·CCAb1 + 0.767 0.079 0.057 –0.808 0.653 <0.001 ***
DMG = –0.432·CCAd1 + 0.736 0.064 0.054 –0.890 0.792 <0.001 *** DMG = –0.473·CCAb1 + 0.692 0.078 0.056 –0.869 0.756 <0.001 ***
D = 0.141·CCAd1 + 0.57 0.051 0.043 0.626 0.392 0.017 *
Autumn
D = 0.147·CCAb1 + 0.586 0.059 0.042 0.584 0.341 0.031 *
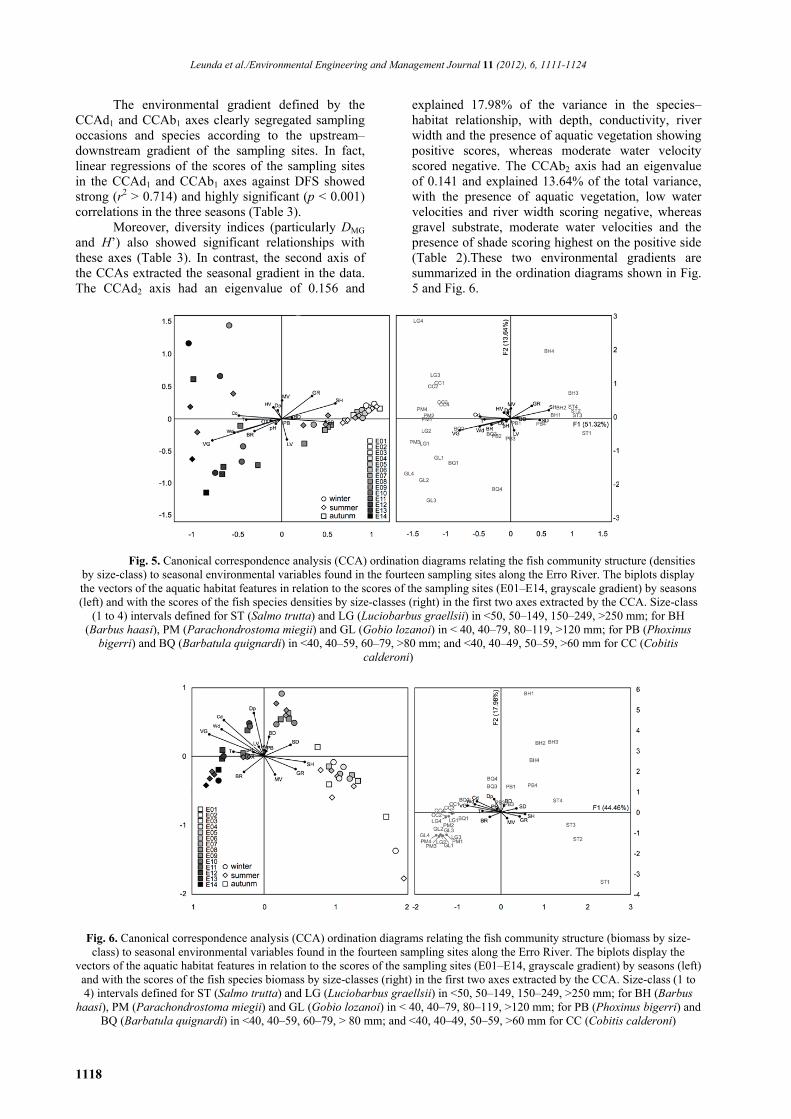
Leunda et al./Environmental Engineering and Management Journal 11 (2012), 6, 1111-1124
1118
The environmental gradient defined by the
CCAd1 and CCAb1 axes clearly segregated sampling occasions and species according to the upstream–downstream gradient of the sampling sites. In fact, linear regressions of the scores of the sampling sites in the CCAd1 and CCAb1 axes against DFS showed strong (r2 > 0.714) and highly significant (p < 0.001) correlations in the three seasons (Table 3).
Moreover, diversity indices (particularly DMG and H’) also showed significant relationships with these axes (Table 3). In contrast, the second axis of the CCAs extracted the seasonal gradient in the data. The CCAd2 axis had an eigenvalue of 0.156 and
explained 17.98% of the variance in the species–habitat relationship, with depth, conductivity, river width and the presence of aquatic vegetation showing positive scores, whereas moderate water velocity scored negative. The CCAb2 axis had an eigenvalue of 0.141 and explained 13.64% of the total variance, with the presence of aquatic vegetation, low water velocities and river width scoring negative, whereas gravel substrate, moderate water velocities and the presence of shade scoring highest on the positive side (Table 2).These two environmental gradients are summarized in the ordination diagrams shown in Fig. 5 and Fig. 6.
Fig. 5. Canonical correspondence analysis (CCA) ordination diagrams relating the fish community structure (densities by size-class) to seasonal environmental variables found in the fourteen sampling sites along the Erro River. The biplots display the vectors of the aquatic habitat features in relation to the scores of the sampling sites (E01–E14, grayscale gradient) by seasons (left) and with the scores of the fish species densities by size-classes (right) in the first two axes extracted by the CCA. Size-class
(1 to 4) intervals defined for ST (Salmo trutta) and LG (Luciobarbus graellsii) in <50, 50–149, 150–249, >250 mm; for BH (Barbus haasi), PM (Parachondrostoma miegii) and GL (Gobio lozanoi) in < 40, 40–79, 80–119, >120 mm; for PB (Phoxinus
bigerri) and BQ (Barbatula quignardi) in <40, 40–59, 60–79, >80 mm; and <40, 40–49, 50–59, >60 mm for CC (Cobitis calderoni)
Fig. 6. Canonical correspondence analysis (CCA) ordination diagrams relating the fish community structure (biomass by size-class) to seasonal environmental variables found in the fourteen sampling sites along the Erro River. The biplots display the
vectors of the aquatic habitat features in relation to the scores of the sampling sites (E01–E14, grayscale gradient) by seasons (left) and with the scores of the fish species biomass by size-classes (right) in the first two axes extracted by the CCA. Size-class (1 to 4) intervals defined for ST (Salmo trutta) and LG (Luciobarbus graellsii) in <50, 50–149, 150–249, >250 mm; for BH (Barbus
haasi), PM (Parachondrostoma miegii) and GL (Gobio lozanoi) in < 40, 40–79, 80–119, >120 mm; for PB (Phoxinus bigerri) and BQ (Barbatula quignardi) in <40, 40–59, 60–79, > 80 mm; and <40, 40–49, 50–59, >60 mm for CC (Cobitis calderoni)

Ichthyofauna of a near-natural Pyrenean river: spatio-temporal variability and reach-scale habitat
1119
The biplot on the left displays the vectors of
the aquatic habitat features against the scores of the sampling occasions, namely the E01–E14 sampling sites coded in a grayscale gradient in combination with different symbols for each survey season. The biplot on the right displays the vectors of the aquatic habitat features, along with the scores of the fish species size-classes.
4. Discussion
This study focused on the spatio-temporal variation of fish community composition and structure and their relationships with reach-scale habitat features along the minimally disturbed Erro River, a Pyrenean tributary of the Ebro Basin (Navarra, northern Spain). The fish communities were found to be composed exclusively of native species, mostly endemic, which are expected to occur in the area: brown trout, Pyrenean minnow, Iberian redfin barbel, Ebro barbel, Ebro nase, Pyrenean gudgeon, Pyrenean stone loach and north Iberian spined loach (García-Fresca, 2003).
The fish community capture efficiency estimates obtained for the summer electrofishing surveys were high and consistent between years as well as along the river. Several along-river longitudinal patterns of fish communities (e.g., species diversity, abundance, size structure) and aquatic habitat features (e.g., conductivity, temperature, turbidity, discharge, width, depth) are known to affect electrofishing capture efficiency (Reynolds, 1996; Zalewski and Cowx, 1990). Even though regression analyses between capture efficiency estimates against distance from source displayed a gentle negative trend, regression parameter values revealed weak or no correlation. Therefore, the effect of such biotic and abiotic factors proved not to be strong enough to have affected the efficiency of the survey methodology used in this study along the Erro River. Regarding capture probabilities, all estimated species-specific values were satisfactorily high, presented narrow standard deviations and overlapped those previously reported for the same or equivalent species (e.g., Lobón-Cerviá, 1991; Penczak and Glowacki, 2008). However, differences in estimated capture probability values between fish species highlight the importance of using species-specific capture probabilities for population size estimation based on single-run electrofishing surveys (Hense et al., 2010; Penczak and Glowacki, 2008).
These differences were related to species attributes such as body size and abundance. The larger and less abundant brown trout, Iberian redfin barbel and Ebro barbel exhibited higher capture probability values compared to smaller and abundant species like Ebro nase, Pyrenean stone loach, Pyrenean minnow and Pyrenean gudgeon. It is known that larger fish are more easily captured by electrofishing because body voltage increases with
size for a given voltage gradient within the electric field applied in the water (Reynolds, 1996). Furthermore, larger fish are more easily detected by dip-netters and consequently are unintentionally preferentially captured (Reynolds, 1996). Hense et al. (2010) pointed out that several research studies have used multiple-run depletion survey data to produce capture probability values for use in fish abundance estimation from single-run electrofishing survey data. However, most of these studies were limited to salmonid (or centrarchid) species, which may make them inapplicable to temperate streams that have fish communities are dominated by cyprinids (or cottids) (Hense et al., 2010). The present study followed this strategy in a near-natura
l Pyrenean river, where fish community changes from salmonid to cyprinid domination, and the observed low variability within species-specific capture probabilities allows their use as reference values for future single-run electrofishing surveys in similar rivers.
The longitudinal fish species addition and succession was analyzed by means of separate hierarchical cluster analyses performed on fish species densities and on theses estimates partitioned by size-classes, and both analyses displayed similar patterns of fish community composition along the sampling sites in the Erro River. The relatively dense distribution of sites surveyed along the Erro River for this study (14 sites along c. 50 km) allowed for great detail in illustrating the fluent transition from Epirithral to Epipotamal zones, which was roughly suggested in a previous study based on only five sampling sites (García-Fresca, 2003). Both dendrograms are able to separate sampling sites into two main groups, E01–E06 and E07–E14, which correspond to the geographic separation provided by a gorge. Within the upper group, the uppermost site E01 can be attributed to Epirhitron because brown trout is the only species present.
Brown trout and Pyrenean minnow co-occur in river reaches from E02 to E06 and thus represent the Metharhitron. There is a drought-affected reach between E03 and E04, where the low-flows in summer infiltrate and any superficial flow disappears, and this disruption was detected by the similarity cluster analyses. The lower group further split in two subgroups, E07–E10 and E11–E14, which correspond to Hyporhithron and Epipotamon, respectively. Even though brown trout was still present in the Hyporhithron, the Pyrenean minnow –small-sized cyprinid– dominated the fish community, and the Ebro nase, Ebro barbel, Iberian redfin barbel and Pyrenean stone loach also occurred in lower numbers. Finally, Ebro barbel, Pyrenean gudgeon, and Ebro nase dominated the Epipotamon, but significant numbers of Pyrenean stone loach and Pyrenean minnow were still present. Sampling site E14 was in the channelized section of a compound flow-gouging weir, and the cluster analyses confirmed that its fish community composition was the most exceptional

Leunda et al./Environmental Engineering and Management Journal 11 (2012), 6, 1111-1124
1120
within the Epipotamon group (Miranda et al., 2005). The scarcity of north Iberian spined loach in the Epipotamon of the Erro River has been previously discussed in detail elsewhere (Leunda et al., 2007). The clear Hyporhithron / Epipotamon separation between E10 and E11 detected by the cluster analyses confirmed the division established in the Erro River by regional managers for recreational fishery management between the ‘trout zone’ and the ‘mixed cyprinid zone’ (BON DF 157/1995). Several studies have adapted the terminology proposed by Illies and Botosaneanu (1963) to describe similar zonal distributions in other rivers in the Iberian Peninsula (García de Jalón and González del Tánago, 1983; Gómez-Caruana and Díaz-Luna, 1991). However, to the authors’ best knowledge, the longitudinal fish distribution of a Pyrenean river has never been described with the level of detail presented here.
The described longitudinal changes in fish species presence and relative contributions to local fish community compositions were also evident from the calculated diversity indices. Shannon–Wiener’s diversity and Margalef’s richness indices showed strong and significant positive relationships with distance from source of the sampling sites along the Erro River.
The continuous addition of species in rivers from headwaters to downstream sites is a universal pattern that has been demonstrated for stream fish communities in temperate and tropical latitudes (Ibañez et al., 2009; Matthews, 1998). In fact, this phenomenon and its effects on the increase of local species richness and diversity along the upstream-downstream gradient have usually been attributed to smooth gradients in environmental abiotic factors in rivers from all continents (e.g., "The River Continuum Concept", Vannote et al., 1980). Specifically, these abiotic factors can be grouped into those related to hydrochemical parameters like temperature and conductivity (e.g., Starmach et al., 1991), those related to habitat size, e.g., stream width, depth and discharge (e.g., Grenouillet et al., 2004; McGarvey and Hughes, 2008; Reyes-Gavilán et al., 1996), and those related to habitat diversity, e.g., depth range, channel morphology, current velocity and substrate composition (e.g., Schlosser, 1982). Because many of these abiotic variables are inherently interrelated and correlated, multivariate statistical approaches are more adequate for the analysis of aquatic habitat variables and their relationships with fish community composition (Ahmadi-Nedushan et al., 2006; Jackson et al., 2001). A previous study on the spatio-temporal variation of aquatic habitat in the Erro River by means of principal component analysis (PCA) identified the longitudinal gradient as the main source of variation (Leunda et al., 2004).
The PCA analysis performed in that study showed a clear separation of upstream and downstream sites on the basis of shade presence, gravel substrate, river width, aquatic vegetation and water conductivity. Here, two separate canonical
correspondence analyses (CCA) were performed to jointly summarize the variation in aquatic habitat parameters in relation to fish community structure; one analysis was based on fish densities (CCAd) and the other was based on biomasses (CCAb). In the two separate CCAs performed, the first two axes largely explained the spatio-temporal variation in the relationship between the size-structured fish community composition and the aquatic habitat features along the Erro River.
The first axis of both CCA analyses clearly segregated sampling sites and fish species according to an upstream-downstream gradient, which was confirmed by the strong and significant correlation between CCA scores and distance from source in the three seasons. This first axis explains nearly 50% of the variance on both CCA analyses, and the aforementioned fluent fish community transition by continuous species addition is clearly displayed along the axis (ST, BH, PB, BQ, CC, GL, LG, PM). In both CCA analyses, the presence of aquatic vegetation and shade, river width, water conductivity, water temperature and substrate types had the highest (absolute) scores in the first axis, which accounted for the longitudinal (spatial) gradient.
The relative position of the fish species along this environmental gradient in the Erro River is in accordance with their habitat preferences described in the Pyrenean area. Brown trout preferred the upper reaches, where cooler water with lower conductivity flows over gravel and pebble substrates in shaded reaches of the Erro River, displaying size-structured differences along these habitat gradients (Ayllón et al., 2010; García de Jalón et al., 1996; Reyjol et al., 2001). Small-to-medium-size reophilic cyprinids occurring in the Erro River can be split into resident and potamodromous species.
Among resident species, the Iberian redfin barbel only occurred at sites E07–E09, probably because of its reported preference for deeper areas with abundant cover and shelter and its preference for coarser substrates in mountain streams (Aparicio and de Sostoa, 1999; Grossman and de Sostoa, 1994; Grossman et al., 1987). The small-bodied resident Pyrenean minnow occurred in all sampling sites except the uppermost site, suggesting that it finds suitable habitats all along the Erro River –i.e., medium depth areas with lower flow velocities, softer substrates and abundant shelter provided by aquatic and riparian vegetation (Mastrorillo et al., 1996; 1997a; 1997b). Potamodromous cyprinids, like the Ebro barbel and the Ebro nase, and the resident Pyrenean gudgeon occurred in the lowermost sites associated with less shaded, wider, and deeper reaches, with increased water temperature and conductivity and lower water velocities (García-Fresca, 2003; Mastrorillo et al., 1996).
In contrast to the latter species group, the distribution of the small-bodied benthonic Pyrenean stone loach was wider and encompassed the entire lower half of the Erro River. This pattern is in accordance with previous studies that reported the

Ichthyofauna of a near-natural Pyrenean river: spatio-temporal variability and reach-scale habitat
1121
presence of Pyrenean stone loach in a wide range of heterogeneous habitats except for those with strong erosive forces (Reyjol et al., 2001; Santoul et al., 2005b). A set of biological traits related to fish life history attributes (e.g., fecundity, potential size, maximum age and reproductive factor) have been found to be significantly related to these species’ longitudinal spatial distributions in the nearby Garonne River system (Santoul et al., 2005a).
Fish life stages usually require habitats with different abiotic features, which may be distributed along the river gradient. Potamodromous fishes perform significant migrations within fluvial ecosystems to reach specific habitats and successfully complete their life cycles (Lucas and Baras, 2001). There have been many studies on the ontogenetic habitat preferences and migratory behavior of fishes with great socioeconomic value like salmonids (e.g., brown trout), but little information is available for most Iberian potamodromous fishes (Santos et al., 2011). Several fish species occurring in the Erro River are potamodromous (brown trout, Ebro barbel and Ebro nase); therefore, differences in the relative position of size-classes were expected along the described spatio-temporal gradient of habitat features (Santos et al., 2011).
However, logistical problems and bad weather conditions prevented sampling in the spring, when the migratory movements of Ebro barbel and Ebro nase are related to the search for suitable substrates during their reproductive period. Only brown trout, which migrate in autumn to reproduce in early winter, displayed a subtle size-structure along the gradients, with young-of-the-year occurring upstream and larger sizes positioning gradually further downstream. In both CCA analyses, the relative positions of fish size-classes were more variable along the second axis than in the first axis, which accounts for the longitudinal gradient. In fact, the second axis in both CCA analyses can be interpreted as representing the seasonal variation in the species–habitat relationship. Depth, river width, water velocity, the presence of aquatic vegetation and shade had the highest (absolute) scores on this axis. Seasonal change in these physical habitat features is closely related to temporal variation in stream flow.
Flow-related variability constitutes an important factor influencing the composition and stability of fish communities in the Iberian Peninsula (Bravo et al., 2001; Godinho et al., 2000). Indeed, low-order streams like the Erro River support less-diverse and less-structured fish communities because seasonal and unpredictable flow variations shape a highly variable environment regarding hydrochemical parameters, habitat size and diversity (Jackson et al., 2001; Tejerina-Garro et al., 2005). Observed differences in the relative positions of fish size-classes along the second axis in both CCA analyses were attributed to community structure changes between seasons. For example, the presence of young-of-the-year cyprinids was related to summer surveys. Accordingly, significant differences were
detected in the comparisons of seasonal values of fish community richness, diversity and dominance indices within the sampling sites of the Erro River. These within-site differences in fish community diversity measures were detected only downstream of E06 because brown trout and Pyrenean minnow were the only species occurring in the headwater sampling sites. Research on freshwater fish communities and their habitats is necessary for dealing with environmental management issues such as fisheries management, the conservation and recovery of threatened species, ecological restoration and water quality bioassessments. These results provide a framework to understand how the spatio-temporal variation of reach-scale aquatic habitat features structure fish communities in a Pyrenean river with low anthropogenic disturbance.
The Erro River is not regulated and the low human population pressure allow the good ecological status of the flowing waters, as demonstrated by previous water quality assessments based on hydrochemical parameters and biotic indices of macroinvertebrates (Leunda et al., 2009b; Oscoz et al., 2005b). However, according to the national dam inventory, there are 274 dams within the Ebro basin and, consequently, most Pyrenean rivers are subject to flow regulation (MARM, 2011). Flow regulation by dams in river ecosystems eliminates their natural dynamic character and has significant ecological effects (see review in Poff and Zimmerman, 2010), including the establishment of non-native fish species (Clavero et al., 2004). This study shows that the fish in the Erro River are all native species (with a high proportion of endemics) that are structured along the described natural spatio-temporal abiotic gradients. The conservation value of the Erro River has also been highlighted by the near-natural spatio-temporal succession of the occurrence of macroinvertebrate families (Leunda et al., 2009b).
As a result, fish fauna of the Erro River could be considered to have a “reference status” for Pyrenean rivers in the implementation of the European Community Water Framework Directive (WFD, 2000/60/CE).
5. Conclusions
This study provide a framework for understanding the spatial (along-river) and temporal (seasonal) variability in the structure of the native fish communities in relation to reach-scale aquatic habitat features in a Pyrenean river with low anthropogenic disturbance. The different statistical analyses performed on species’ densities and biomasses partitioned by size-classes show the prevalence of the along-river gradient over the seasonality in structuring the native fish communities. The relative positions of fish species along the spatio-temporal gradients are in accordance with their habitat preferences described in the Pyrenean area.

Leunda et al./Environmental Engineering and Management Journal 11 (2012), 6, 1111-1124
1122
Acknowledgment We are very grateful to biology students, department colleagues and friends that collaborated in the fieldwork. We also thank Esteban Iriarte, Department of the Environment of the Provincial Council of Gipuzkoa, for lending us the electrofishing equipment. All electrofishing surveys were collected under permits issued to the authors by the Department of Rural Development and the Environment of the Government of Navarra. This research was carried out as part of the project ‘Actuaciones humanas en ríos de Navarra – Su incidencia en la conservación de la biodiversidad’ supported by the Government of Navarra and the Consejo Superior de Investigaciones Científicas of the Government of Spain (2000–2003). Pedro M. Leunda benefitted from a predoctoral grant from the Asociación de Amigos – University of Navarra (2001–2005). References Abell R., Thieme M.L., Revenga C., Bryer M., Kottelat M.,
Bogutskaya N., Coad B., Mandrak N., Balderas S.C., Bussing W., Stiassny M.L.J., Skelton P., Allen G.R., Unmack P., Naseka A., Ng R., Sindorf N., Robertson J., Armijo E., Higgins J.V., Heibel T.J., Wikramanayake E., Olson D., López H.L., Reis R.E., Lundberg J.G., Sabaj Pérez M.H., Petry P., (2008), Freshwater ecoregions of the World: a new map of biogeographic units for freshwater biodiversity conservation, BioScience, 58, 403-414.
Ahmadi-Nedushan B., St-Hilaire A., Bérubé M., Robichaud É., Thiémonge N., Bobée B., (2006), A review of statistical methods for the evaluation of aquatic habitat suitability for instream flow assessment, River Research and Applications, 22, 503-523.
Aparicio E., de Sostoa A., (1999), Pattern of movements of adult Barbus haasi in a small Mediterranean stream, Journal of Fish Biology, 55, 1086-1095.
Ayllón D., Almodóvar A., Nicola G.G., Elvira B., (2010), Ontogenetic and spatial variations in brown trout habitat selection, Ecology of Freshwater Fish, 19, 420-432.
Bain M.B., Stevenson N.J., (1999), Aquatic Habitat Assessment: Common Methods, American Fisheries Society, Bethesda, Maryland, 216.
Bejarano M.D., Marchamalo M., García de Jalón D., González del Tánago M., (2010), Flow regime patterns and their controlling factors in the Ebro basin (Spain), Journal of Hydrology, 385, 323-335.
Benejam L., Aparicio E., Vargas M.J., Vila-Gispert A., García-Berthou E., (2008), Assessing fish metrics and biotic indices in a Mediterranean stream: effects of uncertain native status of fish, Hydrobiologia, 603, 197-210.
BON DF 157/1995, Decreto Foral por el que se aprueba definitivamente el Plan Director de Ordenación Piscícola de Salmónidos de Navarra. Boletín Oficial de Navarra Nº 103, 18/08/1995.,
Bond N.R., Lake P.S., (2003), Characterizing fish-habitat associations in streams as the first step in ecological restoration, Austral Ecology, 28, 611-621.
Bravo R., Soriguer M.C., Villar N., Hernando J.A., (2001), The dynamics of fish populations in the Palancar stream, a small tributary of the river Guadalquivir, Spain, Acta Oecologica, 22, 9-20.
Chessman B.C., Fryirs K.A., Brierley G.J., (2006), Linking geomorphic character, behaviour and condition to fluvial biodiversity: implications for river management,
Aquatic Conservation: Marine and Freshwater Ecosystems, 16, 267-288.
Clavero M., Blanco-Garrido F., Prenda J., (2004), Fish fauna in Iberian Mediterranean river basins: biodiversity, introduced species and damming impacts, Aquatic Conservation: Marine and Freshwater Ecosystems, 14, 575-585.
Confederación Hidrográfica del Ebro, (2011), Automatic Hydrologic Information System (SAIH): Historic data, On line at: http://www.chebro.es.
DeAngelis D.L., Curnutt J.L., (2002), Integration of population, community, and landscape indicators for assessing effects of stressors, In: Biological indicators of aquatic ecosystem stress, Adams S.M. (Ed.), American Fisheries Society, Bethesda, Maryland, 509-531.
Encina L., Rodríguez A., Granado-Lorencio C., (2006), The Iberian ichthyofauna: Ecological contributions, Limnetica, 25, 349-368.
Ferreira T., Caiola N., Casals F., Oliveira J.M., De Sostoa A., (2007), Assessing perturbation of river fish communities in the Iberian Ecoregion, Fisheries Management and Ecology, 14, 519-530.
Filipe A.F., Araújo M.B., Doadrio I., Angermeier P.L., Collares-Pereira M.J., (2009), Biogeography of Iberian freshwater fishes revisited: the roles of historical versus contemporary constraints, Journal of Biogeography, 36, 2096–2110.
García de Jalón D., González del Tánago M., (1983), Estudio biotipológico de las comunidades piscícolas de la cuenca del Duero, Boletín de la Estación Central de Ecología, 12, 57-66.
García de Jalón D., Mayo M., Molles M.C., (1996), Characterization of Spanish Pyrenean stream habitat: Relationships between fish communities and their habitat, Regulated Rivers: Research & Management, 12, 305-316.
García-Fresca C., (2003), Datos sobre la biología de ciprínidos y salmónidos en los Ríos Erro y Urederra (Navarra) – Relación con el medio y análisis metodológico, PhD Thesis, University of Navarra, Pamplona/Iruña, Spain.
Gillette D.P., Tiemann J.S., Edds D. R., Wildhaber M.L., (2005), Spatiotemporal patterns of fish assemblage structure in a river impounded by low-head dams, Copeia, 2005, 539-549.
Godinho F.N., Ferreira M.T., Santos J.M., (2000), Variation in fish community composition along an Iberian river basin from low to high discharge: Relative contributions of environmental and temporal variables, Ecology of Freshwater Fish, 9, 22-29.
Gómez-Caruana F., Díaz-Luna J.L., (1991), Handbook of Freshwater Fishes of the Iberian Peninsula (in Spanish), Editorial Acción Divulgativa S.L., Madrid.
Granado-Lorencio C., (1992), Fish species ecology in Spanish freshwater ecosystems, Limnetica, 8, 255-261.
Grenouillet G., Pont D., Hérissé C., (2004), Within-basin fish assemblage structure: The relative influence of habitat versus stream spatial position on local species richness, Canadian Journal of Fisheries and Aquatic Sciences, 61, 93-102.
Grossman G.D., de Sostoa A., (1994), Microhabitat use by fish in the upper Rio Matarraña, Spain, 1984–1987, Ecology of Freshwater Fish, 3, 141-152.
Grossman G.D., de Sostoa A., Freeman M.C., Lobón-Cerviá J., (1987), Microhabitat use in a Mediterranean riverine fish assemblage: fishes of the upper Matarraña, Oecologia 73, 501-512.

Ichthyofauna of a near-natural Pyrenean river: spatio-temporal variability and reach-scale habitat
1123
Hammer Ø., Harper D.A. T., Ryan P.D., (2001), PAST: Paleontological Statistics software package for educational and data analysis, Paleontologica Electronica, 4, 1-9.
Hense Z., Martin R.W., Petty J.T., (2010), Electrofishing capture efficiencies for common stream fish species to support watershed-scale studies in the central Appalachians, North American Journal of Fisheries Management 30, 1041-1050.
Hernando J.A., Soriguer M.C., (1992), Biogeography of the freshwater fish of the Iberian Peninsula, Limnetica, 8, 243-253.
Ibañez C., Belliard J., Hughes R.M., Irz P., Kamdem-Toham A., Lamouroux N., Tedesco P.A., Oberdorff T., (2009), Convergence of temperate and tropical stream fish assemblages, Ecography, 32, 658-670.
Illies J., Botosaneanu L., (1963), Problems and methods of the classification and of the ecological zonation of running waters, considered primarily in terms of fauna (in French), Mitteilungen der Internationale Vereinigung für Theoretische und Angewandte Limnologie, 12, 1-57.
Jackson D.A., Peres-Neto P.R., Olden J.D., (2001), What controls who is where in freshwater fish communities - the roles of biotic, abiotic, and spatial factors, Canadian Journal of Fisheries and Aquatic Sciences, 58, 157-170.
Kottelat M., Freyhof J., (2007), Handbook of European Freshwater Fishes, Kottelat, Cornol, Switzerland and Freyhof, Berlin, Germany646 pp.
Krebs C.J., (1999), Ecological Methodology, 2nd ed., Benjamin/Cummings, Menlo Park.
Lammert M., Allan J.D., (1999), Assessing biotic integrity of streams: effects of scale in measuring the influence of land use/cover and habitat structure on fish and macroinvertebrates, Environmental Management, 23, 257-270.
Legendre P., Legendre L., (1998), Numerical Ecology, 2nd ed., Elsevier, Amsterdam.
Lelek A., (1974), Toward a method of evaluation of fish populations in streams based on successive fish removals, Proceedings of the Aviemore Symposium FAO EIFAC/74/I/Symp, 38, 2-8.
Leunda P.M., Elvira B., Ribeiro F., Miranda R., Oscoz J., Alves M.J., Collares-Pereira M.J., (2009a), International standardization of common names for Iberian endemic freshwater fishes, Limnetica, 28, 189-202.
Leunda P.M., Miranda R., Escala M.C., (2004), Seasonal aquatic habitat assessment of the Erro River (Navarre) (in Spanish with English abstract), Munibe (Ciencias Naturales - Natur Zientziak), 55, 103-126.
Leunda P.M., Miranda R., Oscoz J., (2007), Occurrence and conservation of the threatened endemic fish Cobitis calderoni in the Erro River (Ebro Basin, Spain), Cybium, 31, 13-18.
Leunda P.M., Oscoz J., Miranda R., (2006), Length-weight relationships of fishes from tributaries of the Ebro River, Spain, Journal of Applied Ichthyology, 22, 299-300.
Leunda P.M., Oscoz J., Miranda R., Ariño A.H., (2009b), Longitudinal and seasonal variation of the benthic macroinvertebrate community and biotic indices in an undisturbed Pyrenean river, Ecological Indicators, 9, 52-63.
Lobón-Cerviá J. (Ed.) (1991), Dinámica de poblaciones de peces en ríos - Pesca eléctrica y métodos de capturas sucesivas en la estima de abundancias, Museo
Nacional de Ciencias Naturales, Consejo Superior de Investigaciones Científicas, Madrid, 156.
Lucas M.C., Baras E., (2001), Migration of freshwater fishes, Blackwell Science Ltd., Oxford, 420.
Magurran A.E., (2004), Measuring Biological Diversity, Blackwell Publishing, Oxford.
MARM, Ministerio de Medio Ambiente y Medio Rural y Marino, (2011), Inventario de Presas y Embalses de España, On line at: http://sig.marm.es/snczi/.
Mastrorillo S., Dauba F., Belaud A., (1996), Utilisation des microhabitats par le vairon, le goujon et la loche franche dans trois rivières du sud-ouest de la France, Annales de Limnologie - International Journal of Limnology, 32, 185-195.
Mastrorillo S., Lek S., Dauba F., (1997a), Predicting the abundance of minnow Phoxinus phoxinus (Cyprinidae) in the River Ariege (France) using artificial neural networks, Aquatic Living Resources 10, 169-176.
Mastrorillo S., Lek S., Dauba F., Belaud A., (1997b), The use of artificial neural networks to predict the presence of small-bodied fish in a river, Freshwater Biology 38, 237-246.
Matthews W.J., (1998), Patterns in Freshwater Fish Ecology, Chapman & Hall, New York.
McGarvey D.J., Hughes R.M., (2008), Longitudinal zonation of Pacific northwest (U.S.A.) fish assemblages and the species-discharge relationship, Copeia, 2008, 311-321.
Miranda R., Oscoz J., Leunda P.M., Escala M.C., (2006), Weight-length relationships of cyprinid fishes of the Iberian Peninsula, Journal of Applied Ichthyology, 22, 297-298.
Miranda R., Oscoz J., Leunda P.M., García-Fresca C., Escala M.C., (2005), Effects of weir construction on fish population structure in the River Erro (North of Spain), Annales de Limnologie - International Journal of Limnology, 41, 7-13.
Naiman R.J., Latterell J.J., (2005), Principles for linking fish habitat to fisheries management and conservation, Journal of Fish Biology, 67, 166-185.
Oscoz J., Campos F., Escala M.C., (2005a), Weight-length relationships of some fish species of the Iberian Peninsula, Journal of Applied Ichthyology, 21, 73-74.
Oscoz J., Leunda P.M., Miranda R., Escala M.C., (2005b), Biological wáter quality of the Erro River (Navarre, N Spain) (2001-2002) (In Spanish), Ecología, 19, 59-74.
Penczak T., Glowacki L., (2008), Evaluation of electrofishing efficiency in a stream under natural and regulated conditions, Aquatic Living Resources 21, 329-337.
Poff N.L., Zimmerman J.K.H., (2010), Ecological responses to altered flow regimes: a literature review to inform the science and management of environmental flows, Freshwater Biology 55, 194-205.
Reyes-Gavilán F.G., Garrido R., Nicieza A.G., Toledo M.M., Braña F., (1996), Fish community variation along physical gradients in short streams of northern Spain and the disruptive effect of dams, Hydrobiologia, 321, 155-163.
Reyjol Y., Hugueny B., Pont D., Bianco P.G., Beier U., Caiola N., Casals F., Cowx I., Economou A., Ferreira T., Haidvogl G., Noble R., de Sostoa A., Vigneron T., Virbickas T., (2007), Patterns in species richness and endemism of European freshwater fish, Global Ecology and Biogeography, 16, 65-75.
Reyjol Y., Lim P., Belaud A., Lek S., (2001), Modelling of microhabitat used by fish in natural and regulated flows in the river Garonne (France), Ecological Modelling, 146, 131-142.

Leunda et al./Environmental Engineering and Management Journal 11 (2012), 6, 1111-1124
1124
Reynolds J.B., (1996), Electrofishing, In: Fisheries Techniques, Murphy B.R., Willis D.W. (Eds.), 2nd edn, American Fisheries Society, Bethesda, Maryland, 221-254.
Santos J., Reino L., Porto M., Oliveira J., Pinheiro P., Almeida P., Cortes R., Ferreira M., (2011), Complex size-dependent habitat associations in potamodromous fish species, Aquatic Sciences - Research Across Boundaries, 73, 233-245.
Santoul F., Cayrou J., Mastrorillo S., Céréghino R., (2005a), Spatial patterns of the biological traits of freshwater fish communities in south-west France, Journal of Fish Biology, 66, 301-314.
Santoul F., Mengin N., Céréghino R., Figuerola J., Mastrorillo S., (2005b), Environmental factors influencing the regional distribution and local density of a small benthic fish: the stoneloach (Barbatula barbatula), Hydrobiologia, 544, 347-355.
Schlosser I.J., (1982), Fish community structure and function along two habitat gradients in a headwater stream, Ecological Monographs 52, 395-414.
Schmutz S., Cowx I.G., Haidvogl G., Pont D., (2007), Fish-based methods for assessing European running waters: a synthesis, Fisheries Management and Ecology, 14, 369-380.
Seber G.A. F., Le Cren E.D., (1967), Estimating population parameters from catches large relative to population, Journal of Animal Ecology, 36, 631-643.
Smith K.G., Darwall W.R.T. (Eds.), (2006), The Status and Distribution of Freshwater Fish Endemic to the Mediterranean Basin, IUCN, Gland, Switzerland and Cambridge, UK.
Starmach J., Fleituch T., Amirowicz A., Mazurkiewicz G., Jelonek M., (1991), Longitudinal patterns in fish communities in a Polish mountain river: their relations to abiotic and biotic factors, Acta Hydrobiologica, 33, 353-366.
teer Braak C.J.F., Verdonschot P.F.M., (1995), Canonical correspondence analysis and related multivariate methods in aquatic ecology, Aquatic Sciences 57, 255-289.
Tejerina-Garro F.L., Maldonado M., Ibañez C., Pont D., Roset N., Oberdorff T., (2005), Effects of natural and anthropogenic environmental changes on riverine fish assemblages: A framework for ecological assessment of rivers, Brazilian Archives of Biology and Technology, 48, 91-108.
Vannote R.L., Minshall G.W., Cummins K.W., Sedell J.R., Cushing C.E., (1980), The River Continuum Concept, Canadian Journal of Fisheries and Aquatic Sciences, 37, 130-137.
Vila-Gispert A., García-Berthou E., Moreno-Amich R., (2002), Fish zonation in a Mediterranean stream: Effects of human disturbances, Aquatic Sciences 64, 163-170.
Washington H.G., (1984), Diversity, biotic and similarity indices - A review with special relevance to aquatic ecosystems, Water Research, 18, 653-694.
Zalewski M., Cowx I.G., (1990), Factors Affecting the Efficiency of Electric Fishing, In: Fishing with electricity - Applications in Freshwater Fisheries Management, Cowx I.G., Lamarque P. (Eds.), Fishing News Books, Oxford, 89-111.