hpml_review
TRANSCRIPT

REVIEW
Behavior at the Choice Point: Decision Making in HiddenPathway Maze Learning
Elizabeth Thomas & Peter J. Snyder &Robert H. Pietrzak &
Paul Maruff
Received: 2 May 2014 /Accepted: 15 September 2014# Springer Science+Business Media New York 2014
Abstract Hidden pathway maze learning tasks (HPMLTs)have been used in neuropsychological research and practicefor more than 80 years. These tasks require the use of visualand auditory task feedback signals to learn the order and direc-tion of a pathway, typically within a grid of stepping-stones, oralleys. Hidden pathway maze learning tasks are purported toassess both visuospatial learning and executive processes. Theoriginal motivation for the HPMLT paradigm for humans was toreduce a complex tactual planning task to one inwhich decisionscould be directly measured by discrete actions at choice pointsguided by visual cues. Hidden maze learning paradigms wereused extensively throughout the 20th century, initially to studyexploratory, anticipatory, and goal-related behavior within thecontext of memory research, and later as an experimental tool inneuropsychology. Computerization of HPMLTs have allowedfor the measurement of different move categories according tothe rule structure and ipso facto, clinically meaningful differ-ences in memory andmonitoring functions during spatial searchand learning. Hidden pathway maze learning tests have been
used to understand the cognitive effects of ageing, neurologicaldisorders, and psychopharmacological challenges.We provide areview of historical antecedents relevant to contemporary appli-cations of HPMLTs in neuropsychology. It is suggested thatcontemporary applications of HPMLTs could be advanced byanalysis of component operations necessary for efficient perfor-mance that can inform theoretical interpretations of this class oftests in clinically meaningful terms.
Keywords Hidden pathwaymaze learning . Executivefunctions . Spatial memory . Goal-related behavior .
Trial-and-error learning
“Much of our thinking goes beyond imagery and in-volves symbolization. Many of our ideas come as insightinto complete relationship of things. Maze learning asan ideation process shows that insight may come grad-ually as the relationships are gradually grasped andcoordinated in some symbolic manner by the mind.”(Chase 1934, pp 438).
Hidden Pathway Maze Learning: An Overview
Hidden pathway maze learning tests (HPMLTs) require a par-ticipant to find the direction of a pathway hidden beneath asmall-scale array of alleys, or stepping-stones across repeatedtrials. These tests differed in structure, requiring the use of astylus to traverse peg-and-grove alleys, or finger/stylus re-sponses to jump from tile-to-tile within a grid of locations.Hidden pathway maze learning tasks were developed byAmerican researchers in the early decades of the 20th centuryto investigate spatial reasoning and learning. The stepping-stonevariant of the HPMLT paradigm has gained prominence incontemporary neuropsychology as a test of visuospatial learning
E. Thomas (*) : P. MaruffPsychological Sciences, University of Melbourne, Parkville,Victoria, Australiae-mail: [email protected]
P. MaruffCentre for Neurosciences, University of Melbourne, Parkville,Victoria, Australia
P. J. SnyderLifespan Hospital System & Department of Neurology, AlpertMedical School of Brown University, Providence, RI, USA
R. H. PietrzakDepartment of Pyschiatry, Yale University School of Medicine, NewHaven, CT, USA
P. MaruffCogState, Ltd., Melbourne, Victoria 3000, Australia
Neuropsychol RevDOI 10.1007/s11065-014-9272-7

and executive function (Bowden and Smith 1994; Walsh 1985).Hidden pathway maze learning has remained a useful model ofcognitive function in neuropsychology because they are rela-tively brief to administer and are understood easily by healthychildren and adults. Furthermore, children with reduced intel-lectual capacity due to brain damage or disorder have been ableto perform HPMLTs because of the simplicity of their rulestructure (Schroder et al. 2004). Despite the simplicity andbrevity of the stepping-stone HPMLT, every move represents adiscrete decision point and consequently a large volume ofinformative data can be obtained within a few minutes of taskperformance (e.g. the Groton Maze Learning Task (GMLT):Snyder et al. 2005a). Computerized versions of HPMLTs haveallowed for greater analytical scope thanmanual predecessors ofthe task because every move can be recorded, classified, andreplayed. When pre-training and alternate pathways of the taskare available, performance by healthy, or brain damaged partic-ipants shows little evidence of practice effects, which is anextremely rare quality in neuropsychological assessment.These factors make HPMLTs ideal for use in repeated measuresresearch designs (e.g. Snyder et al. 2008a). These characteristicshave allowed researchers to build brain-behavior models on thebasis of general performance effectiveness (i.e. from analysis oftotal scores) and also by allowing analysis of the componentcognitive processes as well as their integration (Pietrzak et al.2009a; Thomas et al. 2013). The HPMLT model has been auseful tool in neuropsychology because impairment and im-provement in spatial reasoning and memory functions can bereliably measured over the course of normal development, andacross a range of neuropsychological disorders and treatmentinterventions (Pietrzak et al. 2007; Snyder et al. 2008a; Thomaset al. 2011).
While computerized HPMLTs are relatively new to neuro-psychology, maze learning has been used to study adaptivegoal-related behavior since the formalization of psychologicalscience (Hamilton 1911; Melrose 1922; Ruckmick 1921; Small1901). Current concepts of executive function (Lezak 1982) andspatial memory (Kessels et al. 2001; O’Keefe and Nadel 1978)were derived from maze learning methodologies designed toinvestigate behavior that was structured toward a physically andtemporally remote goal (Melrose 1922; Ruckmick 1921). Theunderlying argument of this review is that historical investiga-tions of motivation and learning through the use of HPMLTs arecontinuous with contemporary investigations of spatial memo-ry, and executive functions such as error monitoring. Analysisof the development of maze learning methodologies is thereforeimportant for understanding the theoretical relevance ofHPMLT paradigms for neuropsychology.
The first section of this review examines the historicalconsiderations through which the stepping-stone variant ofHPMLT was developed and analyzed. It is asserted thatHPMLTs were developed to investigate the processes of cog-nitive integration and co-ordination in goal-directed learning
and these considerations were the precursors to contemporaryaccounts of executive function and spatial navigation. Thesecond half of the review considers the modern uses ofHPMLTs, specifically focusing on use of versions of thestepping-stone maze in neuropsychology. It is argued thatqualitative and quantitative analysis of performance is neces-sary for characterizing change, or difference in cognitiveabilities in clinically meaningful terms.
Hidden Pathway Maze Learning: HistoricalConsiderations
In the introduction to the first edition of the Journal ofExperimental Psychology, Trumball Ladd (1894) outlinedthe historical background to the objectives of the AmericanPsychological Association (APA) by asking the question:“How shall we regard the science of mental life as related tothe methods and conclusions of the most nearly allied physicalsciences, to philosophy, and to human action and character?”(Trumball Ladd, p. 3). One of the aims of the APA at that timewas to establish scientifically credible protocols and methodsfor investigating complexmental and behavioural processes innon-teleological and non-metaphysical terms (Hatfield 2002;Trumball Ladd 1894). Maze learning paradigms were adoptedearly in psychological science because they specifically ad-dressed the problem of how an animal determines the natureof an environment for ecologically salient rewards (such asfood) that are not immediately present, and therefore are notcontiguous determinants of behavior (Small 1901). TheHPMLT paradigm met the standards required of an objectivefactual science from which intelligent, problem-solving be-haviour could be interpreted with regard to a cross-species“natural history of the mind” (Small 1901, p. 222).
From the beginning of comparative psychology (e.g. Darwin1871), the cognitive functions of animals and humans, especial-ly children, inmaze learningwere considered sufficiently similarto regard common behaviours as reflecting a shared intellectualfoundation (Hatfield 2002; Lockard 1971; Tolman 1938; Small1901). Specifically, researchers in comparative psychologybased their models on the assumption that animals and humansrely on common trial-and-error methods for problem-solving(Small 1901, Hamilton 1911). The trial-and-error explorationrequired by maze learning was therefore well suited to theobjectives of early psychological science for investigating selec-tive processes underlying adaptive behavior on the propositionthat discrimination learning was the “basis of each and everyact” (Ruckmick 1921, p. 3–5; Stephens 1935). Tests of discrim-ination learning were also central to advancing theories thatdefined intelligence according to the ability to monitor perfor-mance on a complex task and adapt responses for the efficientpursuit of an end goal (Binet and Simon 1909, cited in Peterson1922, p. 370; Porteus 1918). Decisions made at each choice-
Neuropsychol Rev

point in a maze provided units of behaviour that could beanalysed according to the ability to learn correct turns in thepath from a blind alley (Brown and Buel 1940; Hamilton 1911;Miles 1927; Peterson 1920, 1922; Spence 1932;Warden 1924a,b, 1925). Themaze context was important because it allowed forthe study of behaviour when there is only partial knowledge ofthe environment. It was not simply a method for studying thelearning process, but a method for measuring how an animaldetermines the nature of the environment (Tolman 1948).Researchers investigated how expectancies of a task solutiondeveloped by the extent to which ineffective ‘habits’ of entryinto blind alleyswere eliminated (Brownell 1939; Lashley 1923;Muenzinger 1927; Pepper 1934; Perry 1918; Peterson 1922;Tolman et al. 1932). Thus, in animal studies maze learning wasconsidered an appropriate operational definition for higher cog-nitive processes because it involved “the capacity to take in alarge situation, and by response to remote conditions to delayaction and inhibit motor tendencies aroused by immediate stim-uli until they have been coordinated with the larger circum-stance” (Peterson 1922, p. 387). Maze learning was thereforeregarded as a measure of intelligent learning in which theexperience of previously encountered elements of the mazewas used to guide decisions toward a future state of reward(Peterson 1922), such as food in animals, or task completion inhumans. In general, maze learning was described as a complexprocess of eliminating and integrating behavioral tendencies, orrepresentational elements of a cognitive map, in learning a mazepattern (Hull 1932, 1934a, b, 1938; Tolman 1938, 1948, 1959).
One important characteristic of the rodent maze learningparadigm was that it could be applied largely unchanged tostudy problem solving in humans (Hamilton 1911; Miles 1927;Warden 1924a, b; 1925). Early studies of maze learning inhumans were conducted using small-scale table-topmazes withsubjects blindfolded and using their finger to process tactualcues (Chase 1934; Koch and Ufkess 1926;Melrose 1922; Scott1930; Warden 1924b). Blindfolding in humans was argued toallow for direct comparison of the cognitive processes involvedin maze learning between animals and humans: deprivation ofvision made maze learning dependent on motor cues, asseemed to be the case for rodents (Melrose 1922; Warden1924b). According to Small (1901), tactual-motor experiencewas fundamental to spatial perception. It was generally arguedthat by blindfolding, the primitive basis of experience in kines-thetic and motor cues could be studied directly without theoverlay of latter evolving visual and linguistic processes(Brownell 1939; Peters and McLean 1935; Warden 1924b).
The Development of the Hidden Pathway Maze LearningParadigm
Although the use of blindfolding was intended to make humanand animal maze learning paradigms similar, the deprivation of
vision and forced reliance on tactual-motor information wasnot a normal context for assessing learning and cognition inhumans. For example, the participants in Jensen’s (1934) studyrequired approximately 22 trials and participants made over200 errors, with or without punishment by electric shock, tonegotiate a blindfolded 30 choice point maze (e.g. see Fig. 1).
The blindfolding method used in maze learning studies wasobserved to cause confusion in humans with regard to orientingto the maze for discovery of the pathway direction (Gould andPerrin 1916; Peterson 1920). Errors arising from the disorien-tation in blindfolded maze learning were interpreted as ‘frus-tration’ leading to ‘non-rational’ behaviour such as repeatingmistakes and perseverating at points of disorientation (Peterson1920; Barker 1931). Consequently, apparatus emerged thatwere designed to reduce the artificiality of tactual maze learningtasks for humans and with restricted visual information withoutblindfolding, but which retained the focal construct of mazelearning as a method for studying ‘rational behaviour’. Forexample mazes were developed that allowed indirect visualcontrol such as a map, or ‘two-story duplicate’ maze (Miles1927- see Fig. 2), or the use of hidden stops for hiding the pathin a peg-groove stylus maze (Carr 1921- see Figs. 3 and 4).
In the Miles (1927) maze, the maze layout could be seenfrom the top. However, the top view maze was useful only fornegotiating the bottommaze hidden from view if some form ofvisuo-tactile transformation could be used to bring the twomazes into alignment. In the Carr (1921) maze, the entire mazeproblem is revealed, but slotted grooves hold the stylus in place(Fig. 3), and physical stops beneath the visible apparatus signalentry into blind alleys (Fig. 4). In both examples, the mazelayout could be seen, but required transformations such asimage rotation (Miles 1927), or memory for the location ofblind alleys (Carr 1921). The innovation of partial visual cuesto the maze layout overcame the impedance of blindfolding tomaze learning while preserving the complexity of the task as aspatial learning paradigm. In studies from the early decades of
Fig. 1 Jensen (1934) The effect of punishment by electric shock forerrors on a raised finger maze. The blindfolded participant negotiates amaze with a stylus in the right hand. He receives shocks for errors viaimmersion of left-hand fingers in a salt-bath attached to an electric circuit
Neuropsychol Rev

the 20th century, the role of visual guidance on acts of skill in amotor task was discussed in theoretical and practical terms (seealso, Koch and Ufkess 1926; Warden 1924a). Mainly, re-searchers were concerned with how symbolic knowledge ofspace, or ‘ideation’ is built from the interaction of sensory cueswith intellectual processes (Carr 1921; Chase 1934; Cox 1928;Perrin 1914; Sartian 1940; Tilborg 1936; Warden 1924a, b,1925). Chase (1934) concluded that in ideational learning allsenses are utilized in order to coordinate the central organiza-tion of ideas beyond language and imagery.
The use of indirect visual guidance, as a practical concernin maze learning paradigms, presented methodological prob-lems for designing apparatus for use in humans that had notbeen so important in blindfolded mazes. For example, in ablindfolded maze, the number and length of turns at a choicepoint are not apparent from the outset of the task, as they arewhen the maze layout can be scanned visually. Only tactualand kinaesthetic exploration of the alleys one-at a time canreveal the spatial array, upon which future choice point selec-tion can be made when a location is revisited in future trials(Cox 1928). However, in mazes that are partially revealed, allalleys and turns are immediately present for visual selection atany given choice point. It was recognized that the length andnumber of turns, as well as the direction to the visible end-goalcould influence the probability of selecting an alley. As focusturned from the number of errors or trials to task completion inmaze learning to the probability of selecting particular turns(Hill 1939; Hull 1932; 1934a, b; Tolman 1938), considerationof the maze layout became more important (e.g. Jones andYoshioka 1938; Warden 1924a). Researchers consequentlydesigned standardized maze apparatus in which every alleywas of equal length and with the same number of blind alleys
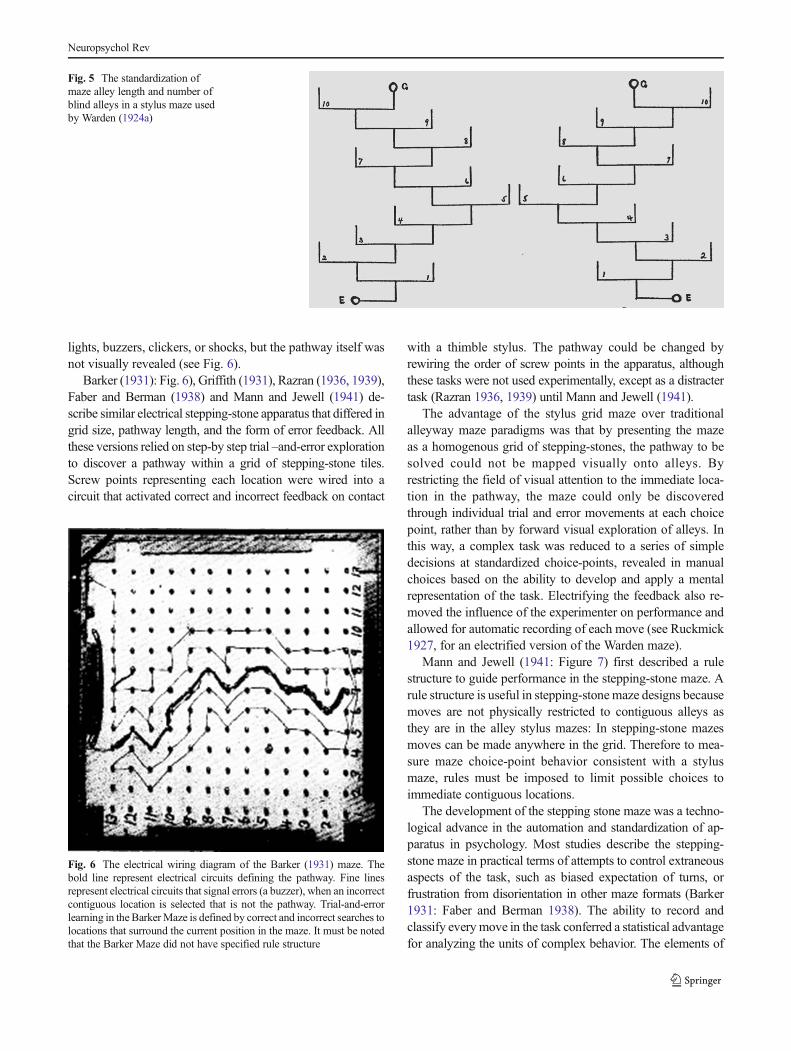
with a single correct alley available for selection at everychoice-point (Griffith 1931; Warden 1924a). By standardizingthe configural aspects of maze pathways, such as the numberand direction of turns, and the length of the alleys, the taskcould be analysed as homogenous units with equal potentialfor selection (Warden 1924a: see Fig. 5).
The introduction of electrically mediated visual and audi-tory feedback signals allowed development of maze learningparadigms in which blind alleys could be signalled symboli-cally, for example by activating a light or a buzzer when ablind alley was entered, rather than relying on physical stopsto indicated dead ends (Barker 1931; Faber and Berman 1938;Gilbert 1934; Gurnee 1938; Razran 1936, 1939).Furthermore, the ability to signal errors at every move madethe need for physical alleys redundant. The maze could berepresented by a field of locations, each of which could beselected one-at-a-time like stepping-stones, with immediatefeedback for each step as correct or incorrect. Hence, move-ment along a series of stepping-stones could be equated withthe forward movement or turns allowed by the alleys in atactual maze. Contiguous points in the surrounding grid de-fined movement options spatially. Movements in the mazewere therefore standardized because every step in the mazepathway gave rise to equal and identical response options forhorizontal and vertical moves in a two-dimensional space. Inelectrical mazes, each location was connected to a contactplate, or screw head contacts wired into an electrical circuit.The circuit was designed so that true points in the pathwaygave different feedback to correct and incorrect locations with
Fig. 2 The two-storey duplicate maze (Miles, 1927). The participantmust learn a pathway through a stylus maze on the lower section that ishidden from view. The maze on the top panel is an inversed version of thebottom panel
Fig. 3 Stylus grooves in the Carr maze (Carr, 1921). Slotted grooves areused to hold the stylus in place within the maze from participant entryuntil completion of the maze
Fig. 4 The pathway of the Carr maze with the position of stops indicatedby dashed lines across the pathway. Location ‘A’ represents the entry, andlocation ‘B’ represents the end position. See Griffith (1931) for anelectrical Carr maze
Neuropsychol Rev
lights, buzzers, clickers, or shocks, but the pathway itself wasnot visually revealed (see Fig. 6).
Barker (1931): Fig. 6), Griffith (1931), Razran (1936, 1939),Faber and Berman (1938) and Mann and Jewell (1941) de-scribe similar electrical stepping-stone apparatus that differed ingrid size, pathway length, and the form of error feedback. Allthese versions relied on step-by step trial –and-error explorationto discover a pathway within a grid of stepping-stone tiles.Screw points representing each location were wired into acircuit that activated correct and incorrect feedback on contact
with a thimble stylus. The pathway could be changed byrewiring the order of screw points in the apparatus, althoughthese tasks were not used experimentally, except as a distractertask (Razran 1936, 1939) until Mann and Jewell (1941).
The advantage of the stylus grid maze over traditionalalleyway maze paradigms was that by presenting the mazeas a homogenous grid of stepping-stones, the pathway to besolved could not be mapped visually onto alleys. Byrestricting the field of visual attention to the immediate loca-tion in the pathway, the maze could only be discoveredthrough individual trial and error movements at each choicepoint, rather than by forward visual exploration of alleys. Inthis way, a complex task was reduced to a series of simpledecisions at standardized choice-points, revealed in manualchoices based on the ability to develop and apply a mentalrepresentation of the task. Electrifying the feedback also re-moved the influence of the experimenter on performance andallowed for automatic recording of each move (see Ruckmick1927, for an electrified version of the Warden maze).
Mann and Jewell (1941: Figure 7) first described a rulestructure to guide performance in the stepping-stone maze. Arule structure is useful in stepping-stonemaze designs becausemoves are not physically restricted to contiguous alleys asthey are in the alley stylus mazes: In stepping-stone mazesmoves can be made anywhere in the grid. Therefore to mea-sure maze choice-point behavior consistent with a stylusmaze, rules must be imposed to limit possible choices toimmediate contiguous locations.
The development of the stepping stone maze was a techno-logical advance in the automation and standardization of ap-paratus in psychology. Most studies describe the stepping-stone maze in practical terms of attempts to control extraneousaspects of the task, such as biased expectation of turns, orfrustration from disorientation in other maze formats (Barker1931: Faber and Berman 1938). The ability to record andclassify every move in the task conferred a statistical advantagefor analyzing the units of complex behavior. The elements of
Fig. 5 The standardization ofmaze alley length and number ofblind alleys in a stylus maze usedby Warden (1924a)
Fig. 6 The electrical wiring diagram of the Barker (1931) maze. Thebold line represent electrical circuits defining the pathway. Fine linesrepresent electrical circuits that signal errors (a buzzer), when an incorrectcontiguous location is selected that is not the pathway. Trial-and-errorlearning in the BarkerMaze is defined by correct and incorrect searches tolocations that surround the current position in the maze. It must be notedthat the Barker Maze did not have specified rule structure
Neuropsychol Rev

cognition could be studied as individual decisions in choice-point behavior, and as a unified whole, such as goals, expec-tations and according to factors that influence immediate be-havioral determinants (Jones and Yoshioka 1938). Theories ofcomplex volitional learning behavior developed throughoutthe 20th century did not focus on the stepping stone maze(e.g. Tolman 1938) because it was rarely used until the latterhalf of the 20th century. However the potential for use ofHPMLT paradigm in theoretical inquiries of spatial reasoningis apparent from quantitative and qualitative analysis of mazeperformance in at least one study (Jones and Yoshioka 1938).
Analysis of Performance in Early Hidden Pathway MazeLearning Studies
Animal and human performance in hidden maze pathwaylearning was usually expressed as a reduction in errors overtrials also termed a learning curve (Gould and Perrin 1916;Lambert and Ewart 1934; Thurstone 1933). However, re-searchers of both animal and human behavior were also inter-ested in qualitative parameters of the learning processes, suchas the rate of elimination of specific choice points in relation tothe end goal (Hull 1934a, b; Jones and Yoshioka 1938;Peterson 1920). It was observed that some turns were harderto learn than others, therefore in accounting for maze behaviorit was important to understand how habits and expectancies ofa maze solution changed with experience (Brown and Buel1940; Hill 1939; Hull 1934a, b, Jones and Yoshioka 1938;Muenzinger 1927; Muenzinger and Vine 1941; Tolman 1938,1932, Tolman et al. 1932, 1938; Voeks, 1948, but see Mann &Jewell 1941). Rather than simply analyzing learning curves, itwas common to analyze qualitatively distinct categories oferrors that related to different types of behaviors, such asanticipation of turns, or avoiding correct turns that lay in theopposite direction to the end goal (Brown & Buel, 1940; Jones
&Yoshioka, 1938). By analyzing how different errors changedwith experience it was possible to describe how ‘schemes ofcontrol’ are built through experience that distinguish learningefficiency (Gould & Perrin 1916, pp 142) and to developtheories of complex behavior (Hull 1934a, b; Tolman, 1938).Theories of maze learning were concerned with anticipatoryand avoidance behavior that could not be simply explained byassociation learning, but required considerations of goals orpurpose through qualitative and quantitative analysis (Hull,1934a; Pepper, 1934; Perrin, 1923; Tolman, 1932).
Although maze learning paradigms were used extensivelyin the first half of the 20th century, only a few studies used thestepping-stone maze before the 1960’s within the theoreticalframework of expectancy and change in motivated behavior(e.g., Jones & Yoshioka 1938; Mann & Jewell 1941: but seeGurnee, 1938; and Razran, 1936, 1939 for quantitative studiesof reinforcement learning in the stepping stone maze).
In one study, a comprehensive analysis of maze learningbehavior in adolescent humans was based on rodent models ofmaze learning (Jones and Yoshioka, 1938). Jones andYoshioka studied 159 adolescents from 11 –to −15 years ona 5×5 stylus maze with 22 and 23 steps (see Figs. 8, 9 and 10)with (1) no feedback, (2) a buzzer, or (3) a light on the fourthand subsequent trials signaling errors.
Results of this study revealed that the nature of task feed-back had no effect on the reduction in errors. In all conditions,the children achieved error free performance after 13 to 15trials. Interestingly, the authors noted that many of the errorsmade by children were incorrect responses made in the gen-eral direction of the end point, where the path continuedlaterally or turned away from the goal. Other frequent errorswere with regard to anticipating turns, and to avoid theperiphery of the maze. Individual differences were shown inthe tendency to retrace and repeat errors and tended to occurmost frequently when the correct pathway turned away fromthe goal. Jones and Yoshioka (1938) described these errors asreflecting (a) goal impetus; (b) drift impetus in the direction ofthe section of the current pathway; and (c) local forward
Fig. 7 The Mann and Jewell bolt head maze (1941). Participants were tofind a pathway traversing the contra-lateral corners of the grid, one step ata time, returning after errors before resuming a search
Fig. 8 The Jones and Yoshioka stylus maze (1938)
Neuropsychol Rev

momentumwhere continuous trajectory is made until an error,where (a) and (b) give rise to anticipatory errors. This study isimportant because it was the first to characterize comprehen-sively the nature, frequency, and change in errors in hiddenpathway electrical stylus maze learning (see Brown & Buel,1940, for a discussion of the findings of Jones & Yoshioka,1938).
Errors such as retracing or repeating the same errorwere often reported in early animal and human maze
learning studies (e.g. Peterson, 1920; Hill, 1939). Theseerrors were described as ‘perseveration’ and were con-sidered to define confused and irrational behavior(Gould & Perin, 1916; Peterson, 1920, 1922).Precursors to modern neuropsychological interpretationsof maladaptive behavior were evident from the time ofBinet and Simon (1909), Hull (1934a, 1943b), Porteus(1958) and Tolman (1959), in so much as they consid-ered variability in adaptation in the maze learning con-text. However, analyses of repeating errors in thestepping-stone maze were not conducted until decadeslater when it was considered an important measure ofperseveration behavior in neuropsychological studies(Karnath et al. 1991; Milner, 1965).
Hidden Pathway Maze Learning and ModernNeuropsychology
Maze learning tasks were considered useful for studies ofcognition in patients with different focal brain lesions becauseof the extensive use of this paradigm in animal learning studies(Orbach, 1955, 1959). Furthermore the processes involved indifferent spatial tasks were well established compared to otherareas of neuropsychological research (Semmes et al. 1955).Non-verbal tasks, such as maze learning, were useful in neuro-psychological experiments because they could be used to studypatients with impairments in aspects of language that were acommon presentation after focal cortical injury (Teuber, 1966).However, the relationship between brain function and aspects ofspatial processing, such as taxon navigation, object-locationlearning, or topographical memory was not well understood(Kessels et al., 2001: Olton, 1985), and consequently the neu-ropsychological interpretations of impairments in hidden path-way maze learning have been speculative.
Fig. 9 The underside wiring diagram of the Jones & Yoshioka (1938)maze. Feedback for each selected location in the maze was given bycircuits A (shaded), which activated a light, and B (unshaded), whichactivated a buzzer. Stops in the pathway were created by the use of brassstrips (C and D) that closed the circuit and activated a signal to indicate anincorrect location
Fig. 10 Pathway patterns used inJones and Yoshioka (1938) areshown in bold with the entryindicated at ‘A’
Neuropsychol Rev

Maze Learning and Lesions of the Temporal Lobes
Hidden pathway maze learning paradigms were most com-monly used in neuropsychological research after the studies ofMilner, (1965), Teuber (1966), and the Oxford group(Newcombe & Russell, 1969) investigating the role of thehippocampus in memory processes. The importance of hippo-campal and medial temporal lobe brain structures to memoryencoding and retrieval was exemplified in patient H.M., whodisplayed severe verbal and spatial anterograde amnesia fol-lowing bilateral mesial temporal lobectomy (Milner et al.1968). Despite this dense amnesia, H.M.’s short term-spatialmemory capacity, as measured by Corsi Block span, waswithin the normal range (Milner, 1971). H.M. and other pa-tients with focal damage to the right medial temporal lobeappeared to have difficulties learning new information thatwas beyond the span of immediate memory (Milner, 1965,1971; Milner et al., 1968). To investigate the extent to whichfocal hippocampal lesions disrupted learning in the spatialdomain, Milner needed a task with a temporal, or sequentialordering component that was beyond the span of immediatespatial memory and therefore required cumulative learning(Milner 1965). Milner used a HPMLT that was similar indesign to the Mann & Jewell (1941) maze. The Milner maze(Fig. 11) and later-developed Oxford HPMLT (Fig. 12) weresimpler than the Porteus or Elithorn perceptual mazes becausethey did not require any development or evaluation of potentialglobal solutions for problem-solving. They were also simplerthan human scale mazes that were navigable by the use ofmaps for orienting in a landscape (Milner, 1965; Semmes et al.,1955). The Semmes et al. (1955) maze required the translationof a 2-dimensional paper map to the location and orientationwithin a physical landscape. Furthermore, the rule structures ofHPMLTs were simple and could be understood easily even bypatients with specific disturbances of language (e.g. arisingfrom focal lesions of Broca’s area). For example, Milner(1965) showed that patients with Broca’s aphasia could per-form the HPMLTwithin normal limits (Milner, 1965).
In Milner’s HPMLT, individuals were required tolocate a 29-step pathway with 11 turns hidden withina 10×10 matrix of stepping stones, or locations tocomplete a single learning trial. The individual had tofind the path traversing contra-lateral corners of the gridusing move rules for the selection of each contiguouslocation using a stylus pen (see Fig. 11). The rules ofthe Milner HPMLT included: go back to the precedingbolthead location when an error is signaled (by anauditory ‘click’); do not retrace sections of the correctpathway (or do not backtrack on the pathway); and donot move diagonally (Milner, 1965).
Milner’s criterion for task completion was the individualobtaining three successive error free trials, or to complete atotal of 25 trials in two blocks given daily, or until criterion
was reached. On the Milner HPMLT, the patient H.M. wasunable to learn the maze within any single session and alsounable to reduce the number of errors made learning the samepathway even after 215 trials across days. The inability tolearn the pathway on the Milner maze was unrelated to anyperceptual or intellectual disability arising from his braininjury (Milner et al., 1968). Subsequent studies interpretedthe deficits in performance on HPMLTs observed in patientswith right retro-Rolandic cerebral damage as topographicalamnesia that was independent of spatial perception or generalintelligence (De Renzi et al. 1977; Newcombe et al. 1987;Newcombe & Russell, 1969; Ratcliff & Newcombe 1973).
The use of HPMLTs to study the effect of lesions orimpairment to the hippocampus in humans was wellestablished by the mid 1970’s, mostly using The OxfordStylus Maze, or the Stylus Maze Test (Newcombe et al.,1987; Newcombe & Russell, 1969; Ratcliff & Newcombe,1973), or HPMLTs that used similar designs (Talland et al.1967; Talland & McGuire, 1967). The Oxford HPMLT wassimilar in design to peg and groove mazes like the Jones &Yoshioka (1938) and the Sartian (1940) HPMLTs. The OxfordStylus HPMLT consisted of an 8×8 grid of raised blockswhere individuals traced a path in an alley between locationsfrom a start point to a finish point with auditory and visualfeedback signals given when the path was traced onto anincorrect location. When errors were signaled the individualreturned to the last choice point before moving again. Task Awas a 6 choice point path in which the examiner demonstratedthe path and the participant was required to relocate thepathway to a criterion of 3 error free trials, or 26 trials. Thiswas used to allow training and practice trial to ensure taskunderstanding (Newcombe & Russell, 1969). Task B had 10choice points and the pathway was not demonstrated,therefore it had to be discovered by trial and error (seeFig. 12).
Fig. 11 The pathway of the Milner HPMLT (Milner, 1965)
Neuropsychol Rev

The Oxford HPMLT was developed as part of a project tostudy the long-term cognitive effects of focal brain injuries,including those affecting the medial temporal lobes and pos-terior parietal cortex (Newcombe & Russell, 1969). The struc-tural similarities between the Milner and Oxford HPMLT areunlikely to have been accidental, given the reported commu-nication between the Montreal and Oxford groups regardingcommon research interests (Newcombe &Russell, 1969). TheOxford HPMLT contained 10 steps with 6 turns. Milner et al.(1968) reported that H.M. showed some evidence of slowmemory acquisition and retention on a modified version ofthe Milner HPMLT that included 8 steps with 3 turns, but wasunable to learn a HPMLT with 10 steps and 4 turns (Milneret al., 1968). H.M.’s ability to learn the smaller HPMLT was
likely due to the capacity to encode the pathway withinworking memory, which was intact. Although it is commonto examine the effect of increasing task difficulty on aspects ofperformance in neuropsychological tests (e.g. De Luca et al.,2003; Luciana &Nelson, 1998, 2002;Welsh et al. 1991), onlytwo studies to date have examined the effect of increasing taskdifficulty, such as grid size and pathway length on a stepping-stone HPMLT (Mathewson et al. 2008; Thomas et al., 2011).Both studies found that manipulation of HPMLT pathwaylength/grid size was associated with qualitative changes inperformance in children (Thomas et al., 2011), as well asneurological differences between older and younger adults(Mathewson et al., 2008). The results of these studies arediscussed later in this review.
Using the Oxford HPMLT, Newcombe and Russell (1969)found evidence of dissociation between visual perception thatwas impaired in patients with occipital lesions, and mazelearning that was specifically impaired in groups withtemporo-parietal lesions, but not in patients with occipitallesions. In accordance with Milner (1965), Newcombe andRussell (1969) argued that performance of the different lesiongroups on the HPMLT could be dissociated such that impair-ment due to right hemisphere lesions could be interpreted as aspatial cartographic problem in posterior parietal groups, and amnestic disturbance in patients with right temporal hippocam-pal excision.
Maze Learning and Lesions of the Frontal Lobes
The importance of the frontal lobes to cognitive models ofproblem solving became evident very early in the history ofneuropsychology (Bigelow, 1850). For example, Teuber(1963, 1966) speculated that lesions of the frontal lobesdisrupted the coordination of cognitive processes thatdepended on other brain structures. Experimental paradigmsused to describe the effects of lesions involving the frontallobes exploited dissociations between the ability to evaluateerrors in performance and to utilize information from errors inplanning future actions (Konow & Pribram, 1970). Anotherway of considering this was as the ability to structure behav-ioral sequences and monitor their outcomes (Luria, 1963,1973). Impairment in this type of complex adaptive behaviorwas observed commonly in patients with frontal lobe lesions.
In order to study disturbances in adaptive behavior inpatients with lesions involving the frontal lobes, maze taskswere utilized often because optimal performance on HPMLTsrequired multi-step forward planning. Paper-and-pencil per-ceptual mazes that depended on visual analysis were used tostudy non-verbal executive functions, as ‘planning and mon-itoring’ tasks (e.g. Elithorn maze: Benton et al. 1963; Elithorn,1955; Porteus maze: Landis & Erlick, 1950; Porteus 1965). Ingeneral, the Porteus and Elithorn mazes measured the ability
Fig. 12 The Oxford Stylus Maze and pathways (from Newcombe andRussell, 1969)
Neuropsychol Rev

to plan the most efficient route through a visual array of alleysor dots that were connected through a series of grids that couldbe increased in size in order to increase the complexity ofdecision-making demands. Route finding efficiency in thePorteus maze was measured by the number of entries intoblind alleys, and by the number of dots traversed in theElithorn maze (Benton et al., 1963; Landis & Erlick, 1950).Failure to follow the task rules was also an important measureof the ability to maintain task set, or rules during mazeperformance (Benton et al., 1963; Landis & Erlick, 1950).Both tasks measured rule break errors, such as deviating fromthe pathway boundary and backtracking (Benton et al., 1963;Landis & Erlick, 1950). Neuropsychological studies indicatedthat performance on paper-and-pencil perceptual maze taskswas impaired in people with focal lesions involving the right/bilateral frontal lobe, as well as the right parietal lobe, sug-gesting that both planning/monitoring and spatial processingfunctions were necessary for optimal performance on thesetasks (Elithorn maze: Benton et al., 1963: Elithorn, 1955;Porteus maze: Landis & Erlick, 1950; Porteus & Kepner,1944). Interestingly, though, the theoretical models of mazelearning, as a measure of adaptive behavior and motivationthat had been developed in earlier decades (e.g. Tolman, 1959;Hull, 1934a) were mostly disregarded in neuropsychologicalstudies. Instead, brain-based models of maze learning focusedon the construct that a task was designed to measure, such aslearning or planning (e.g. Lashley, 1943; Teuber, 1966).
While performance on perceptual mazes was disrupted byfocal lesions of the frontal and parietal lobes, qualitativeanalyses of the errors made by these different patient groupswere not conducted and so it is not possible to determine post-hoc whether the nature of performance impairment differedacross clinical groups (Benton et al., 1963). This is perhapsbecause the errors made on perceptual mazes could not beunderstood easily in terms of prevailing neuropsychologicalmodels of that time (e.g. Landis & Erlick, 1950). For example,solving perceptual mazes required a co-coordinated series ofsteps that take into consideration the entire problem space, aswell as evaluation of possible global solutions. Individualmoves were inextricably tied to a multi-step plan of action.Moves that did not conform to the ideal solution could not beeasily understood according to a theoretical account of thediscrete behavioral processes of maze learning. As HPMLTswere designed to reduce each move to a local series of optionswithin the problem space, each move could be analyzed withregard to behavior at every choice point (Jones & Yoshioka,1938). In this way, maze learning could be regarded as a seriesof conceptually similar choice-point sub-problems. Eachmove constituted a discrete decision unit in a local problemfor finding the next step, and which was guided by the overallgoal of learning the pathway (Griffith, 1931). The HPMLTsretained the focal construct of multi-step forward planning,and spatial learning, but allowed for analysis of each move in
each sub-problem in theoretical terms. In contrast to percep-tual mazes, HPMLTs were amenable to qualitative and quan-titative error analysis, which could be used to guide clinicalinterpretations of focal brain injuries (Milner, 1965). TheHPMLTs were also useful because patient groups with differ-ent cortical lesions were often impaired in both memory andexecutive functions but it was difficult to identify thecontribution of these different impairments to poorperformance on a single neuropsychological task. Milner(1965) was the first to argue that the effects of differentcortical injuries on problem solving and learning could bedissociated using analysis of error types on the stepping-stone version of HPMLT.
Milner recognized that her HPMLT was sensitive to inju-ries of the frontal lobe but that this sensitivity depended onanalysis of the nature of errors made by patients duringpathway learning. For example patients with focal injuriesinvolving the frontal lobe made more errors than controls orpatients with aphasia (arising from lesions of Broca’s area), inthat they revisited locations signaled as incorrect more often,back-tracked along the pathway or jumped to non-contiguouslocations (Milner, 1965). Despite the higher rate of these rulebreak errors, patients with brain injures involving the frontallobes could report that they knew their errors were in violationof the task rules. Subsequent studies have similarly found thatfailing to return, and within search (often termed perseverativeerrors) are more common than other rule break error types inHPMLTs (Karnath et al., 1991; Matson et al. 1997; Milner,1965; Pietrzak et al., 2007; Snyder et al., 2005a; Snyder et al.2008b; Steinberg Chaikelson and Schwartzman 1983;Thomas et al. 2008). This pattern of errors has also beenobserved in children, and occurs most often when the pathwayturns in a direction that is against the end goal, or in anunexpected direction (Thomas, personal observation). Rulebreaking behavior could be discussed from several theoreticalperspectives of error monitoring, behavioral regulation, or‘fixated’ behavior. However theoretical interpretations of rulebreaking behavior and its causes has not been widelydiscussed in relation to HPMLTs.
While the Milner HPMLT had been developed for study ofcognitive impairment in patients with lesions involving thehippocampus, it was applied more broadly to understandcognitive impairment in patients with neuropsychiatric condi-tions associated with deficits in executive functions, such asschizophrenia, obsessive compulsive disorder, as well as inpatients with frontal lobe lesions (Behar et al. 1984; Canavan,1983 [Oxford HPMLT]; Head et al. 1989; Karnath et al. 1991[modified HPMLT]; Matson et al., 1997 [Austin HPMLT];Rettew et al. 1991; Steinberg et al., 1983; Wallesch et al. 1990[modified HPMLT]). In most studies using the MilnerHPMLT and its variants, qualitative and quantitative analysisof errors were used to guide interpretation of impairedperformance in the clinical group under investigation. For
Neuropsychol Rev

example, following Milner (1965) it was common in thestudies cited above to compare the rate of specific errors inHPMLT made by different patient groups, such as repeatingerrors, breaking rules and to differentiate these from errors thatcomply with the rules, but probably reflect difficulty learningthe pathway over trials. Typically errors were classified asqualitatively different forms of behavior that reflected rule-following, or rule-breaking and were interpreted as distinctdisturbances in brain areas associated with error evaluation,and spatial learning and memory. Unfortunately, most studiesusing the Milner HPMLT that have reported separate mea-sures of rule break and spatial memory errors have not includ-ed performance from any of the control groups they alsostudied (for exceptions see: Head et al., 1989; Hymas et al.1991; Rettew et al., 1991). Consequently, no guidelines hadbeen developed to classify normal from impaired performancefor the different types of errors on the HPMLT (Bowden &Smith, 1994). Subsequently, researchers using HPMLT haveshown the sensitivity to detect change or difference in cogni-tion according to analyses of error types reflecting the spatiallearning and rule breaking aspects of performance in terms ofeffect sizes (Pietrzak et al., 2009a; Pietrzak et al., 2009b;Thomas et al., 2008).
The Austin Maze
The Austin HPMLT (see Fig. 13) is a version of the HPMLTMilner maze designed for use in clinical neuropsychologicalsettings as a test of cognitive functions that depended on thefrontal lobes (Darby & Walsh, 2005; Walsh, 1985). Thepopularity of the test in neuropsychology was because it hadproved to be a sensitive test of cognitive dysfunction, at a timewhen there were few tests that were specific to frontal lobeinjuries (Burgess et al., 2006).
Darby and Walsh (2005) describe several case studies thathighlight the sensitivity of the Austin HPMLT to injuries
involving the frontal lobe. In these cases patients with lesionsor injury involving the frontal lobes typically made persistenterrors on the HPMLT showing that like H.M., they wereunable to learn the maze pathway. However H.M. and otherpatients with unilateral or bilateral hippocampal, temporal, orparietal injuries made almost no repeated errors that broke thetask rules. H.M.’s inability to learn the pathway was not due toa failure to follow the task rules, but was attributed to adisturbance of sequential behavior that prevented the acquisi-tion of the pathway in memory (Milner, 1965). Unlike H.M.,the inability to learn the pathway in patients with frontallesions was due to their inability to adjust their behavior onthe basis of the errors made, whether due to disinterest inavoiding errors, or some other dysfunction (Milner, 1965).This inability was considered by Walsh to indicate somedisability to regulate purposive behavior (Darby & Walsh,2005, pp 141, 149). The limited capacity to change behavioron the basis of errors, or repeating the same error continuallydespite negative feedback, was observed in patients who hadsuffered traumatic head injury and in adults with chronicalcoholism (Darby & Walsh, 2005). Walsh was careful toemphasize that these deficits reflected a problem of errorutilization, rather than error evaluation, because all of theirpatients demonstrated that they knew their errors were occur-ring but despite this did not modify their behavior in accordwith this knowledge (Darby & Walsh, 2005, pp 168). Hence,this impairment was framed as a difficulty in implementingbehavioral plans, not impairment in awareness, or in theassessment of the significance of the errors that broke the taskrules. On the HPMLT this failure to learn from errors wasobserved as a rapid reduction in errors across the initiallearning trials with the persistence of a small number of errorsacross the remainder of trials with performance never becom-ing error-free (i.e. defined by Walsh as 3 errorless trials)despite individuals being allowed to continue learning themaze indefinitely. It appears that this criterion of error-freeperformance within 20–30 trials (Walsh, 1985) was selectedbecause Walsh (1960) also reported that patients who hadundergone frontal leucotomy also made rule break errorssimilar to those that had been observed by Milner (1965).However, Walsh found that the rate of rule break errors inpatients with frontal lobe leucotomy was not significantlygreater than controls (Darby & Walsh, 2005 pp, 149, 167).Studies have consistently shown that the rate of rule breakerrors is typically much lower than the rate of pathway mem-ory errors, except in patients with severe frontal lobe injuries(Behar et al., 1984; Canavan, 1983; Milner, 1965; Rettewet al., 1991; Steinberg et al., 1983). Inter-individual differ-ences in HPMLT errors can be shown using metrics thatconsider the relative rate and variability of differenceswithin and between groups (i.e. effect sizes), than theraw rate of different errors (Snyder, et al., 2005a;Snyder et al., 2005b).
Fig. 13 The Austin HPMLT. (Image reproduced with permission ofNCounters Australia). Contralateral red buttons control the timer for thebeginning and end of the maze. Red and green lights are activated at thetop of the maze to indicate correct and incorrect choices. The NCountermaze has a choice of two pathways
Neuropsychol Rev

Bowden and Smith (1994) criticized the use of a criterionrequiring stable error-free performance to define difficulties oferror utilization in HPMLTs, from both a statistical and theo-retical perspective. First, stable error-free performance couldbe achieved in patients with injuries involving the prefrontalcortex, albeit after many more trials than required by healthycontrols (Milner, 1965). Second, in some healthy adults error-free performance required as many as 50 trials (Bowden et al.,1992), and therefore failure to reach criterion in 20 or 30 trials(Walsh, 1985) could not be considered a reliable measure ofcognitive impairment. Finally, Bowden and Smith (1994)showed that the rate of learning (i.e. reduction in errors) overthe initial 10 trials on the Austin HPMLT was highly predic-tive of the number of errors on subsequent trials made by bothhealthy adults and patients with heterogeneous brain lesions(Bowden & Smith, 1994). Although it was traditional toexamine maze performance in terms of trials to a criterion oferror free completion, after the studies of Bowden, it becameincreasingly common to examine errors over a pre-specifiednumber of trials (e.g. Barker et al. 2005; Morrison & Gates,1988; Österberg et al. 2009). The consequence of this was thatit made the time for testing briefer than when trials to error-free performance was used and it also provided a statisticallydefensible basis for standardized task administration.
In the late 1980’s, the Austin HPMLT was used by someresearchers to study impairments in spatial mapping in clinicalconditions proposed to be associated with disruption to thehippocampus, such as alcohol dependence (Bowden, 1988,1989; Bowden & McCarter, 1993; O’Brian et al. 2003). Therapid encoding and recall of object-locations has consistentlybeen shown to depend on the right mesial temporal region(Saling, 2009). However, the neurological impact of long-term chronic alcohol use on brain structures associated withexecutive functions has not, to our knowledge, been assessedusing error analysis in the Austin HPMLT. The AustinHPMLT was used extensively in clinical neuropsychologicalstudies, including normative and validation studies in healthyadults (e.g. Bowden & Smith, 1994; Bowden, et al., 1992;Crowe et al., 1999; Morrison & Gates, 1988; Tucker et al.1987). Studies that administered the Austin HPMLT in neu-rologically impaired populations, in which executive func-tions would likely be affected, typically define impairmentin terms of the total errors, total trials, and total time to reachsome criterion (Darby & Walsh, 2005; Österberg et al., 2009;Österberg et al. 2002; Österberg, Ørbæk, Karlson, Bergendorf,& Seger, 2000; Barker et al., 2005; Kinsella et al., 1995).Despite the extensive use of the Austin HPMLT it is difficultto integrate data across studies because no standard methodfor administration and scoring of the task has been developedor validated. For example, studies using the Austin HPMLTdiffer in the criterion for task completion (10 trials: Barkeret al., 2005; Bowden et al., 1997; O’Brian et al., 2003;Österberg et al., 2009; Walton & Bowden, 1997; two, or three
error free trials: Kilpatrick et al., 1997; Kinsella, et al., 1995;Kinsella et al., 1997; Mathias & Kent 1998; Morrison &Gates, 1988). More importantly, although the task rules havebeen described by Bowden and by Milner (Bowden, 1988,1989; Milner, 1965), studies using the Austin HPMLT havebeen inconsistent in reporting which, or if any rules weregiven to participants prior to assessment (Barker et al., 2005;Mathias & Kent 1998; Matson et al., 1997; Österberg, et al.,2009; Österberg et al., 2002; Österberg et al., 2000).
As described earlier, Bowden & Smith (1994) describedthe wide variance in trials to criterion across control andclinical groups in the Austin HPMLT. Furthermore, trials tocriterion and errors to criterion are also highly correlated inhealthy adults (r=.73-.91: Tucker et al. 1987). Bowden hasalso shown that test-retest reliability on alternative pathwaysof the Austin HPMLT is generally low (errors to criterion:r=.77, trials to criterion: r=.64: Bowden, 1989). The rules ofthe Austin maze are given verbally and task administrationdoes not contain training, or pre-trial practice. This raises thepossibility that improved performance on the second admin-istered pathway (e.g., Bowden, 1989) reflected acquisition ofthe task rules in the first administration that was consolidatedby the second administration.
Searching the same location, or within-search errors wereregarded by Milner (1965) as errors common to patients withfrontal lobe injuries and interpreted as ‘perseveration’, butthey are not included as rules as reported by any studiespublished using the Austin or Milner HPMLT. The definitionsof rule break errors have also been inconsistent, when theyhave been recorded in theMilner HPMLT. Bowden and Smith(1994) criticized the use of rule break errors for makingclinical decisions because of the absence of normative guide-lines for determining impairment in this measure. However,there were at least 5 studies before the mid 1990’s that indi-cated the rates of rule break errors in different clinical groupswas sufficiently high to warrant analyses as a separate mea-sure to other error categories (Behar et al., 1984; approximate-ly 50-to-60 in patients with frontal lobe lesions over 15 trials:Canavan, 1983; 20 rule break errors in patients with frontallobe injuries on trials to criterion:Milner, 1965; approximately2–6 rule break errors in teenagers with Trichotillomania,obsessive compulsive disorder, or Mixed anxiety disordersin 10 trials: Rettew et al., 1991; approximately 42-to32 rulebreak errors in younger and older schizophrenics respectively,in trials to criterion: Steinberg et al., 1983). By comparison,studies that included healthy controls showed that rule breakerrors were infrequent in this group and ranged from approx-imately 1-to 5 errors in trials to criterion (17 trials: Milner,1965; 20 trials: Steinberg et al., 1983: 10 trials: Rettew et al.,1991). Even when rule break errors have been reported, thetheoretical significance of these errors in the stepping-stoneHPMLT has received little attention. Subsequent studies usingthe Austin HPMLT have not considered the quantitative and
Neuropsychol Rev

qualitative nature of errors from which a theoretical analysisof task variability could be advanced.
Although the Austin HPMLT has been used widely inclinical neuropsychological contexts, this review suggests thatit still remains more of an experimental than clinical neuro-psychological test. Researchers have sought to characterizevariability in the rate of pathway learning errors in order tounderstand the metric properties of the task, as much as toinvestigate cognition in different groups. Furthermore theconstruction of the Austin HPMLT, as a commercial manualtask without alternate pathways, limited the administration ofthe task to verbal instructions without practice or task famil-iarization. In order to use HPMLTs for the study of cognitivefunctions, as opposed to being a tool for understanding thenature of maze learning, recent computerized adaptations ofthe HPMLT paradigm have become more standardized, withrules to guide performance and criteria for classification oferrors specified. Two examples of such adaptations arediscussed below.
Computerized Versions of HPMLT
Computerization of the maze format from the late 1980’s hadseveral advantages for administering and analyzing perfor-mance on HPMLTs (Morrison & Gates, 1988). First, comput-erization allowed greater flexibility of HPMLT design andadministration. Second, computerization makes it possible torecord and classify automatically every move, as well as everymove sequence with respect to prior moves or to the mazelayout. Functional neuroimaging studies using HPMLT haveshown changes from frontal to temporoparietal activity overpracticed trials, consistent with a shift from trial-and-errorsearching when the pathway is unknown, to navigation frommemory (Van Horn et al., 1998). This analysis is also consis-tent with the interpretation of HPMLTs as a spatial learningparadigm within the context of executive functions (Milner,1965). At least two versions of this HPMLTwere designed inthe first decade of 2000 for use in clinical research, andimaging environments respectively. This section of the reviewbegins with an examination of the neurological basis ofstepping-stone maze performance in healthy adults from aneuro-imaging study, followed by consideration of computer-ized HPMLTs. The Groton Maze Learning Task (Schroderet al., 2004) preceded the development of the Integneuromaze(Paul et al., 2005). However, in order to critically examinecommon limitations of HPMLT studies in general, theIntegneuro maze will be examined before consideration ofstudies using the Groton Maze Learning Task.
Hidden pathwaymaze learning has been described as a taskinvolving mental mapping (Bowden, 1989), and has beenassociated with topographical memory (De Renzi et al.,1977; Habib& Sirigu, 1987). However topographical amnesia
has been defined as environmental disorientation in pathfinding involving locomotor movement through spaces thatare beyond the immediate perceptual horizon (Habib& Sirigu,1987). It has not been established that the stepping stone mazereflects the same basic processes of way-finding, or navigationin real or virtual landscapes. For example, in hidden pathwaymaze learning the entire problem space is visible (Barker,1931). Hence it is not necessary to create a mental map ofan unseen topology. Milner (1965) suggested that the frame ofthe HPMLT task was fixed in relation to the body and there-fore does not require the transformation of coordinate refer-ence frames that is required by locomotor tasks. It is unlikelythat pathway mapping relies exclusively on visual imagery:One study has shown that attempts to draw the pathway inconjunction with manual maze learning conferred no advan-tage to pathway encoding (Mann & Jewell, 1941). Anotherstudy has shown that there was no transfer of learning betweenan identical stylus and a physical maze (Jones & Batalla,1944). It is therefore probable that maze performance relieson exocentric spatial representations that are dependant on thehippocampus and supporting regions rather than exclusivelyvisual processes.
Although there have been neuro-imaging studies of hiddenpathway maze learning, the tasks used have differed to thestandard Milner versions (i.e. do not allow for most rule breakerrors), have examined non-specific regional measures ofbrain activity or connectivity, or have not examined the per-formance of healthy adults (Chen et al. 2009; Clark et al.,2006; Flitman et al. 1997; Paul et al., 2009; Tate et al., 2010).Only one study has examined stepping-stone maze perfor-mance in healthy adults using positron emission tomography(PET).
Van Horn et al. (1998) used PET imaging to examine theneural correlates of novel, and practiced maze learning using acomputerized 36-step maze. A chase test was used as control.Initial maze performance elicited activity in the right prefron-tal cortex (Brodmann’s areas (BA) 8, 9, 45, and 46). Minoractivity was observed in anterior cingulated and medial pre-frontal area’s (24, and 32), left middle frontal gyrus (10), rightinferior parietal lobule (7, 40), left lingual gyrus (19), andcuneus (18, 19). Practiced trials (2.5 min) elicited strongestparietal activity (bilateral medial parietal and precuneus (7,31), and posterior cingulate, or retrosplenial cortex (29, 30).Little activity was observed in the prefrontal cortex in prac-ticed trials, but activity was observed in left anterior prefrontalcortex in initial trials (10). Together the pattern of activityshowed initial right-sided anterior activity during task acqui-sition, along with BA 34, which has shown to be involved inspatial encoding (Chrastil, 2013; Van Horn et al., 1998), andmedial/posterior activity with recall. Hippocampal andparahippocampal areas were active during both encodingand recall, suggesting the involvement of the place navigationsystem (O’Keefe & Nadel 1978) and the grid system in the
Neuropsychol Rev

entorhinal complex for recall. Brain areas activated in initialand practiced trials are shown in Figs. 14 and 15. The func-tional significance of the regions of interest from this studycan be explored in greater detail in Brodmann’s InteractiveAtlas: http: / /www.fmriconsult ing.com/brodmann/Introduction.html.
It must be noted that responses in the VanHorn et al. (1998)study was via a hand-held device, presumably a joystick or afour-button keypad. If so, then only individual horizontal andvertical steps were possible, thus reducing the rule structure ofthe task and response requirements from the original Milnermaze. Therefore the results from this study should beinterpreted with caution in relation to the 28-step manualstepping-stone maze format.
The Integneuro Maze
The IntegNeuro battery from the Brain Resource Companyhas been used mostly for investigating the brain correlates ofcognitive deficits associated with heterogeneous brain disor-ders (Lipton et al., 2009; Paul et al., 2009; Tate et al., 2010).The Integneuro maze is an 8×8 HPMLT presented on a touchscreen. Moves are made using a 4-directional control panel(with arrows) set on the screen below the maze. Feedback isgiven by auditory and visual responses below the maze (noton each selected tile). Diagonal and jump moves are notpossible.
The Integneuro maze task runs until two error-free trials, ora timeout of anywhere between 7 min (Clark et al., 2006;Mathersul, et al., 2009; Schofield et al., 2009) and 10 min(Gunstad et al., 2007; Paul et al., 2005: Paul et al., 2006; Paulet al., 2009; Silverstein et al., 2007). The computer softwarerecords the number of trials, errors, and time to completion(Clark et al., 2006). Overruns are also recorded, where partic-ipants continue to search locations in a linear trajectory when
Fig. 14 PET activity in Broddmans regions in initial maze learning on amodifiedMilner maze (+−major activity, −minor activity in Brodmann’sarea): +8: Supplementary motor, +9: Middle frontal gyrus (prefrontalcortex), +45: Inferior frontal gyrus Pars triangularis), +46: Anteriormiddle frontal gyrus (prefrontal cortex), −24: Anterior cingulate gyrus,−32: Anterior cingulate gyrus, −10: Middle frontal gyrus (prefrontalcortex), −7: Superior parietal lobule (secondary sensorimotor cortex)−40 Inferior parietal lobule (supramarginal gyrus), −19 Inferior occipitalgyrus (secondary visual cortex), −18: Middle occipital gyrus (secondaryvisual cortex).
Fig. 15 PETactivity in Broddmans regions in practicedmaze learning ona modified Milner maze : medial parietal and precuneus 7, 31, andposterior cingulate 29, 30
Neuropsychol Rev

the pathway turns. The reason for inclusion of this measureand its theoretical significance has not been made explicit inany study that has used the Integneuro maze. Overruns havebeen reported previously and described as pattern factors ofthe maze that generalize to a local forward drift momentum, ororientation direction in choosing pathway locations (Jones &Yoshioka, 1938). In other words, if the pathway is unknown,participants tend to continue searching in the same goal-warddirection until an error is indicated. The pathway consists of24 steps (Integneuro User Manual Version 2007v3: availablefrom www.brainclinics.com), however, the number of turnsincluded in these steps has not been reported and there is noevidence as to whether different pathways are used. TheIntegneuro maze is described as a test of executive functionsbased on the Austin HPMLT, and there is extensive normativedata from 6-years to- 70+ in 1007 participants (Clark et al.,2006), and one validation study (Paul et al., 2005). Howeverthis validation study did not use any other form of maze taskfor comparison (for example the Austin maze on which thetask was purportedly based: Paul et al., 2005).
The Integneuro maze appears to be based on the Morrisonand Gates (1988) computerized version of the Austin maze inthat moves are executed via a 4-directional keypad maderemotely from the maze grid, not within the grid itself. Themove restrictions of this format are similar to the physical‘alleys and stops’ of the Oxford maze and its predecessors inwhich diagonal or jump moves cannot be made (Jones &Yoshioka 1938). However because move decisions are exe-cuted remotely to the maze environment, it is possible thatmove sequences could be coded linguistically according toapplication of a sequence of commands restricted to ‘up,down, left, right’. Because these commands can be appliedand evaluated without visual inspection of the maze layout itis possible that the Integrneuro maze task could be completedefficiently from verbal memory alone, or that any visualdecision making could be supported or enhanced by linguisticprocessing. Therefore differences in the rule structure andadministration of the Integneuro maze limits comparison tomodels of HPMLT maze learning based on performance onthe Austin andMilner HPMLTas measures of either executivefunctions or spatial mapping.
In one study, the correlation between Integneuro mazecompletion time and the Rey Complex Figure Test-delay(RCFT-d) was taken as a validation of the Integneuro maze(Paul et al., 2005). However the selection for the RCFTmeasure in the delayed form as a validating task is unclear.While the authors assert that the Integneuro maze is a measureof executive functions, the delayed trial of the RCFT is con-sidered to be a task of visual memory (Caffarra et al. 2002).The use of the RCFT as a test of executive function dependson the analysis of error patterns, and organization abilities(Meyers & Myers 1995; Meyers & Volbrecht, 1998). Thecommon operations required by the Integneuro maze and the
RTDFT-d were not discussed. Divergent validity wasestablished by the lack of correlation between Inegneuro mazemeasures and the California Verbal Learning Test- Research(short delay). However at least 30 measures were recordedfrom 12 tests administered in the experiment, but an inter-correlation matrix of other measures in the battery was notreported. The veracity of the claim of validation of theIntegneuro maze cannot be assessed without a more completereport of the convergent and divergent relationships with othertests of cognitive functions administered in the experiment.No such data has as yet been published for this version of theHPMLT.
The Integneuro maze has been used extensively in imagingand tractography studies as a task of executive function withinthe context of spatial learning (Lipton et al., 2009; Schofieldet al., 2009; Seckfort et al., 2008; Tate et al., 2010). Howeverimaging studies using the Integneuro maze have used sum-mary measures over trials, not behavioral trial by trial data inrelation to stability or changes in brain functions over trials, aswas shown by Van Horn et al. (1998). The contribution ofstudies using the Integneuro maze for understanding HPMLTperformance is limited, because the maze has not been vali-dated as a HPMLT, and the theoretical significance of themeasures it records, such as overruns are not well understood.
The Groton Maze Learning Task (GMLT)
The Groton Maze Learning Test (GMLT) is a touch screenHPMLT, based on the Austin and Milner HPMLT (seeFig. 16). Originally developed by P.J. Snyder and colleaguesfor use in neuropharmacological studies at the former maincampus for research and development of Pfizer Inc.(Boulanger et al. 2006; Castner et al., 2004; Pietrzak et al.,2007). Maze pathways begin at the top left-hand corner andend at the bottom right-hand corner (see Fig. 16). The pathwayremains hidden and only the most recently selected square is
Fig. 16 The GMLT. The grid shows the current location (blue tile) andthe end goal (red target). The pathway begins in the top left corner andfinishes in the bottom right corner
Neuropsychol Rev

illuminated. When a correct square is touched, it turns whitewith a green tick, accompanied by an auditory tone. When anincorrect tile is touched, it turns white with a red cross, and notone is emitted. Move feedback appears for 250 ms, followingwhich, the square changes color to blue. If two successiveerrors are made, the last correct location (referred to as the“head of path”) flashes 10 times with a tick, as a signal toreturn to the head of path. Correct and error-move feedbackprovides pathway information (Pietrzak et al., 2009a; Pietrzaket al., 2009b). For the GMLT, the grid size, pathway length/number of turns, and rules are the same as for the MilnerHPMLT.
Because the GMLT was specifically designed for clinicaltrials, there was emphasis placed on the ability to generatealternative forms that were resistant to practice effects (Maruffet al., 2006; Pietrzak et al., 2009b; Snyder et al., 2005a;Snyder et al., 2005b; Snyder, et al., 2008a). Hence, there are20 alternate-form maze pathways in the GMLT, and these areall equivalent with respect to the number of correct moves,turns, and individual choice or decision points, within the 10×10 grid. Each time an individual starts the test, a form isselected in pseudo-random order to ensure that no individualsubject will see the same alternate form within three succes-sive test sessions. When a subject attempts to discover thehiddenmaze on the first trial, selection of tiles that are allowedby the task rules for finding the hidden pathway are catego-rized by the GMLTsoftware as exploratory errors. Moves thatviolate the task rules are classified as rule break errors (seeTable 1 for a description of error categories). Researchersusing the GMLT were the first to systematically codify errorcategories in HPMLTs, as well as the first to report analysis of
different errors as a standard output feature (e.g. Maruff et al.,2006; Snyder et al., 2005a; Snyder et al., 2005b). Each step inthe pathway must be located in sequence, according to therules in order to complete a trial.
From any location in the GMLT, it is possible to move toany location within the grid. For the GMLT software torespond with appropriate feedback to every move, it is neces-sary that all moves can be classified according to the task rulesand according to the nature of the move (i.e. see Table 1 forerror classifications). A consequence of this is that the GMLToutput can be rendered to provide raw data for each move andeach response time, as well as trial-by-trial summaries ofdifferent errors and responses. Because the GMLT recordsand classifies every move, there are more categories of ‘erro-neous’ moves than are reported by other HPMLTs (doubletapping tiles, moves between tiles). Correctly returning to thehead of path after errors are signaled can also provide ameasure of error feedback use, although this measure hasnot been previously reported. Most reports using the GMLTinclude incorrect moves that comply with the task rules (legalerrors); errors that do not comply with the task rules (rulebreak errors); and repeating the same error in succession(perseverative errors). These error categories are typicallysummed over the five learning trials and expressed as an effectsize of difference from control, or change from pre-treatmentbaseline score (Pietrzak et al., 2008a; Snyder et al., 2005a;Snyder et al., 2008a; Thomas et al., 2008). Practice trials arean essential component of the GMLT, as it is important toensure that participants fully understand the rules of the taskand can demonstrate stable baseline performance prior toadministering the task in research or clinical applications.Other studies have reported a summary measure of learningefficiency, or a ratio of total moves over trials to task comple-tion (Collie et al. 2006; Maruff, et al., 2006; Pietrzak et al.,2007; Pietrzak et al., 2009a; Pietrzak et al. 2008b; Snyderet al., 2005a). There is ample evidence in the references abovesuggesting the sensitivity of GMLT measures to detect clini-cally significant change or difference using standard outcomemeasures of error and efficiency. However, these along withother GMLT measures can be used to address a variety oftheoretical questions related to the specificity and qualitativenature of cognitive performance on a complex task.
In neuropsychological research and practice, it is importantthat the choice of outcome measures is suitable for specificexperimental and clinical questions. For example, summaryoutcome measures may be preferred to multiple, “process-based” measures when a test is included as part of a large testbattery, or when non-specific brain disease or disorders wouldlikely be associated with diffuse cognitive impairments, suchas in schizophrenia (Pietrzak et al., 2009b). Alternatively,researchers may be interested in the component operationsunderlying task performance in which multiple measures areuseful (e.g. Alderman et al. 2003). As the GMLT is a learning
Table 1 Definition of Errors on the Groton Maze Learning Task. AllMeasures are Recorded as Counts on Each Trial (Five Trials in Total)
Error type Definition
Exploratory Moves in accordance with task rules.
Failure to return Failing to return to the last correct locationafter an incorrect move.
Perseverative The same error repeated after returning to thehead of path.
Back Moving backwards on the pathway.
Jump Moving more than one tile from the currentcorrect location.
Diagonal Moving to locations diagonal to the currentcorrect location.
Double tap Tapping the same incorrect location twice.
Same tile tap Tapping the correct location twice.
Between Moves between tiles.
Out Moves out of grid.
Within Re-searching locations allowed by the task rulesthat have been identified as the incorrect locationfor the next step in the pathway.
Neuropsychol Rev

task, it is also possible to examine the learning processthrough the change in different parameters of performanceover trials, although few studies have examined individualtrial data in the GMLT (Pietrzak, et al., 2009a; Thomas et al.,2013).
One of the most consistent findings in studies using theGMLT is the differential effect of clinical status on measuresof rule break, and legal or exploratory errors. Although there isvast amount of evidence from early studies confirming thatHPMLTs measure some aspects of executive functions inaddition to spatial mapping and learning functions, theseprocesses can be differentiated on the GMLT. For example,exploratory and rule break errors can be independently affect-ed in clinical groups, as well as with pharmacological manip-ulation of neurotransmitter systems. Memory and executivefunction deficits often co-occur in cases of diffuse brain dis-orders and several studies have shown the dissociation of ruleuse and spatial memory processes in hidden pathway mazelearning (Canavan, 1983; Thomas et al., 2008; Snyder et al.,2005a; Snyder et al., 2008b). Rule-break errors in HPMLTsare rare in healthy adults, but common in populations withexecutive function deficits due to anatomical, chemical, orage-related disruption to the prefrontal cortex (Pietrzak et al.,2008a; Snyder et al., 2008a; Snyder et al., 2008b; Thomaset al., 2008). In these studies, spatial memory in the GMLT isgenerally far less affected than executive functions (e.g.Snyder et al., 2008a; b; Thomas et al., 2008). By contrast,patients with focal lesions of the hippocampus, reduced per-formance onHPMLTs are predominantly driven by reductionsin spatial learning and memory, with little or no disruption torule use ability (Milner, 1965; Milner et al., 1968).Importantly, the ability to understand the relative contributionof cognitive systems to performance can inform neuropsycho-logical models of hidden pathway maze learning. For exam-ple, Snyder et al. (2008b) found that children with attentiondeficit hyperactivity disorder (ADHD) made more rule breakerrors than age-matched controls, but not more exploratoryerrors in the GMLT. Administration of stimulant medication inchildren with ADHD led to a greater magnitude reduction inlegal than perseverative errors, suggesting that stimulant med-ication improves both learning/memory and executive pro-cesses in children with ADHD to differing degree’s. Together,these findings are theoretically and clinically important be-cause they suggest that it is possible to isolate and comparedifferent task components of HPMLTs that allows for greaterinterpretive scope in analyzing performance variability.
Because every move in HPMLTs represents a discretedecision, behavioral analysis of move-by move performancecan be coupled with neuroimaging data. Although HPMLTs,other than the Integneuro maze, have not been widely used inimaging studies, they show great potential for coupling simpleresponses with brain activity and cognitive behavior in acomplex task. For example, by examining the brain responses
to individual moves, such as returning after errors as opposedto failing to return after errors, it is possible tomake theoreticalclaims regarding the brain-behavior relationship in error mon-itoring, although such an analysis has yet to be conducted.
Mathewson et al. (2008) investigated error-feedback pro-cessing in aging by examining the event related potentials toerror feedback in older and younger adults performing threelevels of maze difficulty. Fifteen younger females (18–26 years), and 15 older females (65–87 years) competed 84×4 maze (1 learn, 2 test trials), 4 6×6 mazes (1 learn, 3 testtrials), and 2 8×8 mazes (1 learn, 5 test trials). The mainfinding was that in younger adults, electroencephalogram(EEG) feedback related negativity was sensitive to errors atall levels of difficulty in both learn and test trials (it should benoted that Mathewson et al., did not differentiate error types).Consistent with the findings of Van Horn et al. (1998) themaxima of EEG activity was evident in medial brain areas,including the anterior cingulate cortex (ACC), with an anteriorshift evident in more difficult learn trials, and posterior shiftevident in practice trials. For older adults, there was no spe-cific response differentiation in the medial prefrontal cortex;neural responses to correct moves and errors were associatedwith more diffuse brain activity. Behavioral data also showedthat there were no differences in errors on learning trials ateach level of difficulty, but that older adults were less efficientin learning the mazes. Processing speed (Trails A) did notaccount for the variance in error rates, suggesting that perfor-mance was dependent on higher-order learning and executiveprocesses. Older adults also produced smaller feedback-related-negativity (FRN’s) associated with error feedback inall conditions, suggesting that older adults may have recruitedbrain regions less reliant on dopaminergic activity (i.e. thanthe ACC). For both groups, the P3 was sensitive to feedbackvalence. The differentiation between event-related-potentials(ERP) and P3 sensitivity to error feedback (correct and incor-rect moves) with age also support the inference that these twosystems may rely on different neurotransmitter systems. Oneconfounding factor in this study was that neural responseswere not analyzed separately for qualitatively different errortypes (rule break and exploratory). Based on developmentalstudies of maze performance, it would be expected that rulebreaking errors would be more frequent in older adults than inyounger adults (Pietrzak et al., 2007). Analysis of qualitativelydifferent errors could provide information regarding the brain-behavior relationship between internally generated errors (rulebreaking), feedback errors (failing to return, within search),and correct error discrimination (returning after an error). It islikely that qualitatively different errors would be associatedwith distinct patterns of brain activity. If so, then aggregatingerrors (rule break and legal) would appear as more diffuseactivity than consideration of specific categories of error. Thiscould account for the more diffuse error-related neural re-sponses observed in the older adult sample. If this
Neuropsychol Rev

interpretation is correct, it underscores the importance ofassessing different error responses in making inferences re-garding putative brain bases of error monitoring processes.
As noted earlier in this review, Walsh (1985) hypothesizedthat the main difficulty underlying rule break errors in clinicalpopulations with injuries to the prefrontal cortex was aninability to use error information to plan and execute moves,or error utilization. Rule break errors can occur as a failure tofollow rules, such as ‘jumps,’ diagonals, or backtracking, or asa failure to use prior task information to select the next move,such as failing to return after error signals are given, orrepeatedly searching the same location within a trial. Erroranalysis of these different types of errors could be used toidentify the nature of rule use difficulties in normal develop-ment, as well as in clinical samples. Such an analysis wouldthus provide useful information regarding the theoretical basisof variability in maze learning.
An example of componential analysis can be seen in onestudy in which the GMLT was administered to children from5-to-9-years of age (Thomas et al., 2011). Error analysis in thisstudy showed an age-related trend in the profile of differenterror categories (Thomas et al., 2011). Compared to olderchildren, younger children made more errors related to theability to use error information (i.e., failing to return andwithin search errors), than failing to follow the rules (i.e.,making ‘jumps’ and diagonal moves). This finding is consis-tent with the early development of rule use, but the relativelylatter development of error monitoring in childhood (Bunge&Crone, 2009; Marcovitch & Zelazo 2009). It is also consistentwith Walsh’s (1985) hypothesis that error utilization is themain difficulty demonstrated by individuals with executivefunction deficits in performance on the Austin Maze.
Computerization of HPMLTs have allowed for fine-grained analysis of component operations necessary forgoal-directed behavior in trial-and-error learning, such as cor-rect and incorrect error feedback use, spatial learning, and ruleuse. As such, this paradigm shows great potential as a toolfrom which models of complex cognition and its disturbancescan be developed, based on the qualitative and quantitativenature of performance on a single test that is brief to admin-ister, simple to perform, and resistant to practice effects.
Summary and Directions for Future Research
Maze learning was a central methodology for studying funda-mental aspects of cognition and behavior throughout the 20thcentury. HPMLTs were designed to overcome the limitationsof other maze formats in humans, and to improve the psycho-metric properties of such tests in order to augment interpreta-tion of performance. With the advent of brain-based ap-proaches to adult neuropsychology in the 1950’s, maze learn-ing became a useful diagnostic measure of mid-temporal lobe
and frontal lobe function according to the profile of differenterrors made by patient groups. Theoretical accounts of whatHPMLTs measured during the 1980’s and 1990’s were re-stricted by the use of summary outcome measures, such astotal errors or trials to a criterion of task completion. The useof global measures and lack of task standardization haveconsequently prevented comprehensive analysis of brain-based, or cognitive systems that could account for perfor-mance variability both within, and across studies. However,on the weight of evidence, modern HPMLTs are able to assessvarious cognitive abilities sub-served by functionally separateneural systems for spatial learning/memory, and goal-directederror monitoring and rule use. The ability to measure differentaspects of performance, as well as the potential to modify thetask makes the HPMLT paradigm a powerful tool for investi-gating how different cognitive functions change, or remainstable as a function of age, clinical status, or task demands.The flexibility of HPMLTs, in terms of the range of outcomemeasures derived from a move-by move, and trial-by-trialbasis allows for a comprehensive approach to characterizingperformance parameters reflecting component aspects of vi-suospatial learning and executive function.
HPMLTs are learning tasks that are simpler to administerand measure than large-scale or virtual navigation tasks, andappear to elicit neural responses in similar brain regions tolarge-scale tasks. HPMLTwould therefore be useful for inves-tigating factors that facilitate, or disrupt spatial encoding andrecall processes in healthy adults and children, and how theserelate to ‘real-world’ spatial processing, or specific executivefunction disruptions (error feedback use, rule following, goalmaintenance) in patients with focal or diffuse brain disorders.
The review presented here has shown that there is asound neuropsychological basis within which perfor-mance on the HPMLT can be understood. This now pro-vides a firm foundation for exploitation of HPMLTs in thefuture to assist in understanding disorders of the centralnervous system that are characterised by impairment inexecutive function but where the neuropsychological sub-strate of this impairment in now known. For example,GMLT measures are sensitive to cognitive impairment inschizophrenic patients compared to controls, as well astreatment improvement and stability over time (Snyderet al., 2008a). Another aspect of HPMLT that bodes wellfor understanding complex executive functions is that thisparadigm performs well in psychopharmacological stud-ies. Repeated performance is not associated with practiceeffects, therefore HPMLTs can be used to assess theeffects of compounds know to influence executive func-tions and spatial memory, such as dopamine antagonistsor nicotinic agonists. Finally, one notable absence in theliterature of HPMLTs is an understanding of the function-al neuroanatomoy of the different error types. Futurestudies could combine componential analysis of different
Neuropsychol Rev

task parameters with experimental methods to furtherdelineate the functional dynamics of HPMLT perfor-mance, such as imaging, TMS, as well as pharmacologicalmanipulations.
References
Alderman, N., Burgess, P. W., Knight, C., & Henman, C. (2003).Ecological validity of a simplified version of the multiple errandsshopping test. Journal of the International NeuropsychologicalSociety, 9(1), 31–44. doi:10.1017/S1355617703910046.
Barker, R. G. (1931). The stepping-stone maze: A directly visible space-problem apparatus. Journal of General Psychology, 5, 280–285.doi:10.1037/h0072916.
Barker, M. J., Greenwood, K. M., Jackson, M., & Crowe, S. (2005). Anevaluation of persisting cognitive effects after withdrawal fromlogn-term benzodiazépine use. Journal of the InternationalNeuropsychological Society, 11, 281–289. doi:10.1017/S1355617705500332.
Behar, D., Rappoport, J. L., Berg, D. J., Denckla, M. B., Mann, L., Cox,C., & Wolfman, M. G. (1984). Computerized tomography andneuropsychological test measures in adolescents with obsessive-compulsive disorder. American Journal of Psychiatry, 141(3),363––367. Retrieved from: http://ajp.psychiatryonline.org.
Benton, A. L., Elithorn, A., Fogel,M. L., &Kerr, M. (1963). A perceptualmaze test sensitive to brain damage. Journal of Neurology,Neurosurgery and Psychiatry, 26, 540–544. doi:10.1136/jnnp.26.6.540.
Bigelow, H. J. (1850). ART.I- Dr. Harlow’s case of recovery from thepassage of an iron bar through the head. The American Journal ofMedical Sciences, 16(3), 13–22. Retrieved from: http://journals.lww.com/amjmedsci/toc/1850/16390.
Binet, A., & Simon, T. H. (1909). L’Intelligence des Imbeciles. AneePsychol., 1–147. (128ff), cited in Peterson, J. (1922). Intelligenceand learning. Psychological Review, 29(5), 336–389. doi:10.1037/h0069483.
Boulanger, M., Snyder, P.-J., & Cohen, H. (2006). Ralentissementcognitif dans le vieillissement: fonctions exécutives et apprentissageprocédural lors d'une tâche informatisée de labyrinthe (Assessmentof cognitive slowing and error monitoring in healthy older subjectson a computerized maze learning task). Annales Médico-Psychologiques, Revue Psychiatrique, 164(6), 463–469. doi:10.1016/j.amp.2005.01.001.
Bowden, S. C. (1988). Learning in young alcoholics. Journal of Clinicaland Experimental Neuropsychology, 10(2), 157–168. doi:10.1080/01688638808408232.
Bowden, S. C. (1989). Maze learning: Reliability and equivalence ofalternative pathways. The Clinical Neuropsychologist, 3(2), 137–144. doi:10.1080/13854048908403286.
Bowden, S. C., Dodds, B., Whelan, G., Long, C., Dudgeon, P., Ritter, A.,& Clifford, C. (1997). Confirmatory factor analysis of the Wechslermemory scale-revisited in a sample of clients with alcohol depen-dency. Clinical and Experimental Neuropsychology, 19(5), 755–762. doi:10.1080/01688639708403757.
Bowden, S. C., Dumendizic, J., Hopper, J., Kinsella, G., Clifford, C., &Tucker, A. (1992). Healthy adults performance on the Austin Maze.The Clinical Psychologist, 6, 43–52.
Bowden, S. C., & McCarter, R. J. (1993). Spatial memory in alcohol-dependent subjects: Using a push-buttonmaze to test the principle ofequiavailability. Brain and Cognition, 22, 51–62. doi:10.1006/brcg.1993.1024.
Bowden, S. C., & Smith, L. C. (1994). What does the Austin mazemeasure? Australian Psychologist, 29, 34–37.
Brown, W., & Buel, J. (1940). Response tendencies and maze patterns asdeterminers of choice in a maze. Journal of ComparativePsychology, 29(3), 337–399. doi:10.1037/h0059538.
Brownell, W. A. (1939). Theoretical aspects of learning and transfer oftraining. Review of Educational Research, 9(3), 255–273. Retrievedfrom: http://www.jstor.org/stable/1167404.
Bunge, S. A., & Crone, E. A. (2009). Neural correlates of the develop-ment of cognitive control (pp 22–37). In J. M. Rumsey & M. Ernst(Eds.), Neuroimaging in Developmental Clinical Neuroscience.London: Cambridge University Press.
Burgess, P. W., Alderman, N., Forbes, C., Costello, A., Coates, L. M.-A.,Dawson, D. R., & Channon, S. (2006). The case for the develop-ment and use of “ecologically valid”measures of executive functionin clinical and experimental neuropsychology. Journal of theInternational Neuropsychological Society, 12, 194–209. doi:10.1017/S1355617706060310.
Caffarra, P., Vezzadini, G., Dieci, F., Zonato, F., & Venneri, A. (2002).Rey-Osterrieth complex figure: normative values in an Italian pop-ulation sample. Neurological Sciences, 22, 443–447. Retrievedf r om : h t t p : / / l i n k . s p r i n g e r . c om / a r t i c l e / 1 0 . 1 0 0 7 /s100720200003#page-1.
Carr, H. A. (1921). The influence of visual guidance in maze learning.Journal of Experimental Psychology, 4(6), 399–417. doi:10.1037/h0072602.
Canavan, A. G. M. (1983). Stylus-Maze performance in patients withfrontal-lobe lesions: Effects of signal valency and relationship toverbal and spatial abilities. Neuropsychologia, 21(4), 375–382. doi:10.1016/0028-3932(83)90024-6.
Castner, S. A., Andrews, R. D., Ordonez, C. E., Brown, T. F., Strother, S.C., Wernick, M. N., Williams, G. V., & Snyder, P. J. (2004). Contextdependent effects of lorazepam on regional cerebral glucose metab-olism revealed by cognitive testing in FDG-PET. Poster presentationat 17th annual Winter Conference on Brain Research, CopperMountain, CO., 24–30 January
Chase, W. P. (1934). The role of kinesthesis in ideational maze learning.Journal of Experimental Psychology, 17(3), 424–438. doi:10.1037/h72695.
Chen, K. H.M., Chauh, L. Y.M., Sim, S. K. Y., & Chee,M.W. L. (2009).Hippocampal region specific contributions to memory performancein normal elderly. Brain and Cognition, 72, 400–407. doi:10.1016/j.bandc_2009.11.007.
Chrastil, E. E. R. (2013). Neural evidence supports a novel framework forspatial navigation. Psychonomic Bulletin and Review, 20, 208–227.doi:10.3758/s13423-012-0351-6.
Clark, C. R., Paul, R. H., Williams, L. M., Arns, M., Fallahpour, K.,Handmer, C., & Gordon, E. (2006). Standardized assessment ofcognitive functioning during development and aging using an auto-mated touchscreen battery. Archives of Clinical Neuropsychology,21, 449–467. doi:10.1016/j.acn.2006.06.005.
Collie, A., Maruff, P., Snyder, P. J., Darekar, A., & Huggins, J. P. (2006).Cognitive testing in early phase clinical trials: outcome according toadve r s e even t p ro f i l e i n a Phase I s t udy. HumanPsychopharmacology: Clinical and Experimental, 21(7), 481–488.doi:10.1002/hup.799.
Cox, C. (1928). Comparative behavior in solving a series of mazeproblems of varying difficulty. Journal of ExperimentalPsychology, 11(3), 202–218. doi:10.1037/h0074984.
Crowe, S. F., Barclay, L., Brennan, S., Farkas, L., Gould, E.,Katchmarsky, S., & Yayda, S. (1999). The cognitive determinantsof performance on the Austin Maze. Journal of the InternationalNueropsychological Society, 5, 1–9. Retrieved from: http://journals.cambridge.org/action/displayJournal?jid=INS.
Darby, D., & Walsh, K. (2005). Walsh’s Neuropsychology. A ClinicalApproach (5th ed.). Edinburg: Elsevier.
Neuropsychol Rev

Darwin, C. R. (1871). The Descent of Man. London: Murray.De Luca, C. R., Wood, S. J., Anderson, V., Buchanan, J., Proffitt, T. M.,
Mahony, K., & Pantelis, C. (2003). Normative data from CANTAB.I: Development of executive functions over the lifespan. Journal ofClinical and Experimental Neuropsychology, 25(2), 242–254. doi:10.1076/jcen.25.2.242.13639.
De Renzi, E., Faglioni, P., & Villa, P. (1977). Topographical amnesia.Journal of Neurology, Neurosurgery, and Psychiatry, 40, 498–505.doi:10.1136/jnnp.40.5.498.
Elithorn, A. (1955). A preliminary report on a perceptual maze testsensitive to brain damage. Journal of Neurology, Neurosurgeryand Psychiatry, 18, 287–292. doi:10.1136/jnnp.18.4.287.
Faber, P., & Berman, L. (1938). Construction and use of a variable contactmaze. The Psychological Record, 10(2), 302–306. Retrieved from:thepsychologicalrecord.siuc.edu/Vol-2-1938.html.
Flitman, S., O’Grady, J., Cooper, V., & Grafman, J. (1997). PET imagingof maze processing. Neuropsychologia, 35(4), 409–420. doi:10.1016/S0028-3932(96)00086-3.
Gilbert, R. W. (1934). An electrical maze stylus. Journal of GeneticPsychology, 10, 485–486. doi:10.1080/00221309.1934.9917755.
Gould, M. C., & Perrin, F. A. (1916). A comparison of the factorsinvolved in the maze learning of human adults and children.Journal of Experimental Psychology, 1(2), 122–154. doi:10.1037/h0072916.
Griffith, C. R. (1931). A flexible form of the Carr Slot-Maze. TheAmerican Journal of Psychology, 43(2), 283–285. Retrieved from:http://www.jstor.org/stable/i261734.
Gunstad, J., Paul, R. H., Cohen, R. A., Tate, D. F., Spitznagel, M. B., &Gordon, E. (2007). Elevated body mass index is associated withexecutive function dysfunction in otherwise healthy adults.Comprehensive Psychiatry, 48, 57–61. doi:10.1016/j.comppsyc.2006.05.001.
Gurnee, H. (1938). The effect of electrical shock for right responses onmaze learning in human subjects. Journal of ExperimentalPsychology, 22(4), 354–364. doi:10.1037/h0054726.
Habib, M., & Sirigu, A. (1987). Pure topographical disorientation: Adefinition and anatomical basis. Cortex, 23, 73–85. doi:10.1016/S0010-9452(87)80020-5.
Hamilton, M. D. (1911). A study of trial and error reactions in mammals.Journal of Animal Behavior, 1(1), 33–66. doi:10.1037/h0070439.
Hatfield, G. (2002). Psychology, philosophy, and cognitive science:Reflections on the history and philosophy of experimental psychol-ogy. Mind and Language, 17(3), 207–232. Retrieved from: http://scholar.google.com.au/scholar?q=hatfield+2002+mind+and+language&btnG=&hl=en&as_sdt=2005&sciodt=0%2C5&cites=3249434448813365107&scipsc=.
Head, D., Bolton, D., & Hymas, N. (1989). Deficits in cognitive shiftingability in patients with obsessive-compulsive disorder. BiologicalPsychiatry, 25(7), 929–937. Retrieved from: http://www.elsevier.com/wps/ f ind / journa ldescr ip t ion .cws_home/505750/description#description.
Hill, C. J. (1939). Goal gradient, anticipation, and perseveration in com-pound trial-and-error learning. Journal of Experimental Psychology,25(6), 566–585. doi:10.1037/h0059929.
Hull, C. L. (1932). The goal gradient hypothesis and maze learning.Psychological Review, 39(1), 25–43. doi:10.1037/h0072640.
Hull, C. L. (1934a). The concept of the habit-family hierarchy: Part I.Psychological Review, 41(1), 33–54. doi:10.1037/h0070758.
Hull, C. L. (1934b). The concept of the habit-family hierarchy: Part II.Psychological Review, 41(2), 33–54. doi:10.1037/h0072855.
Hull, C. L. (1938). The goal-gradient hypothesis applied to some ‘field-force’ problems in the behavior of young children. The PsychologialReview, 45(4), 271–299. doi:10.1037/h0053885.
Hymas, N., Lees, A., Bolton, D., Epps, K., & Head, D. (1991). Theneurology of obsessional slowness. Brain, 114, 2203–2233. doi:10.1093/brain/114.5.2203.
Jensen, M. B. (1934). Punishment by electric shock as affecting perfor-mance on a raised fingermaze. Journal of Experimental Psychology,17(1), 65–72. doi:10.1037/h0075639.
Jones, H. E., & Batalla, M. (1944). Transfer in children’s maze learning.The Journal of Educational Psychology, 35(8), 474–483. Retrievedfrom: http://csaweb112v.csa.com.ezp.lib.unimelb.edu.au/ids70/browse_archive.php?SID=5h5k0hteisoj9ieljg6sk16j91&db=psycarticles-set-c&issn=0022-0663.
Jones, H. E., & Yoshioka, J. (1938). Differential errors in children’slearning on a stylus maze. Journal of Comparative Psychology,25, 463–480. doi:10.1037/h0054154.
Karnath, H. O., Wallesch, C. W., & Zimmermann, P. (1991). Mentalplanning and anticipatory processes with acute and chronic frontallobe lesions: Comparisons of maze performance in routine and non-routine situations. Neuropsychologia, 29(4), 271–290. doi:10.1016/0028-3932(91)90042-7.
Kessels, R. P. C., de Haan, E. H. F., Jaap Kappelle, L., & Postma, A.(2001). Varieties of human spatial memory: a meta-analysis on theeffects of hippocampal lesions. Brain Research Reviews, 35(3),295–303. doi:10.1016/S0165-0173(01)00058-3.
Kilpatrick, C., Murrie, V., Cook, M., Andrews, D., Desmond, P., &Hopper, J. (1997). Degree of left hippocampal atrophy correlateswith severity of neuropsychological deficits. Seizure, 6, 213–218.doi:10.1016/S1059-1311(97)80008-8.
Kinsella, G., Prior, M., Sawyer, M., Murtagh, D., Eisenmajer, R.,Anderson, V., & Klug, G. (1995). Neuropsychological deficit andacademic performance in children and adolescents following trau-matic brain injury. Journal of Pediatric Psychology, 20(6), 753–767.doi:10.1093/jpepsy/20.6.753.
Kinsella, G., Prior,M., Sawyer,M., Ong, B.,Murtagh, D., Eisenmajer, R.,& Klug, G. (1997). Predictors and indicators of academic outcomein children 2 years following traumatic brain injury. Journal of theInternational Neuropsychological Society, 3(6), 608–616. Retrievedfrom: http://journals.cambridge.org.ezp.lib.unimelb.edu.au/action/displayJournal?jid=INS.
Koch, H. L., & Ufkess, J. (1926). A comparative study of stylus mazelearning by blind and seeing subjects. Journal of ExperimentalPsychology, 9(2), 118–131. doi:10.1037/h0075473.
Konow, A., & Pribram, K. H. (1970). Error recognition and utilizationproduced by injury to the frontal cortex in man. Neuropsychologia,8(4), 489–491. doi:10.1016/0028-3932(70)9004-8.
Lambert, J. F., & Ewart, P. H. (1934). Part I: The effect of verbalinstruction upon stylus maze learning. The Journal of GeneralPsychology, 6(2), 377–399. doi:10.1080/00221309.1932.9711879.
Landis, C., & Erlick, D. (1950). An analysis of the Porteus Maze-Test asaffected by psychosurgery. The American Journal of Psychology,64(4), 557–566. Retrieved from: http://www.jstor.org/stable/1418870.
Lashley, K. S. (1923). The behavioristic interpretation of consciousness I.The Psychological Review, 30(4), 234–272. doi:10.1037/h007383.
Lashley, K. S. (1943). Studies of cerebral functioning in learning. XII.Loss of maze habit after occipital lesion in blind rats. The Journal ofComparative Neurology, 79(3), 431–462. doi:10.1002/cne.900790309.
Lezak, M. D. (1982). The problem of assessing executive functions.International Journal of Psychology, 17, 281–297. doi:10.1080/00207598208247445.
Lipton, M. L., Gulko, E., Zimmerman, M. E., Friedman, B. W., Kim, M.,Gellella, E., & Branch, C. A. (2009). Diffusion-tensor imagingimplicates prefrontal axonal injury in executive function impairmentfollowing very mild traumatic brain injury. Neuroradiology, 252(3),816–824. doi:10.1148/radiol.2523081584.
Lockard, R. B. (1971). The fall of comparative psychology: Is there amessage for us all? American Psychologist, 26(2), 168–179. doi:10.1037/h0030816.
Luciana, M., & Nelson, C. A. (1998). The functional emergence ofprefrontally-guided working memory systems in 4-to 8-year-old
Neuropsychol Rev

children. Neuropsychologia, 36(3), 273–293. doi:10.1016/S0028-3932(97)00109-7.
Luciana, M., & Nelson, C. A. (2002). Assessment of Neuropsychologicalfunction through use of the Cambridge Neuropsychological TestingAutomated Battery: Performance in 4- to 12-year-old children.Developmental Neuropsychology, 22(3), 595–624. doi:10.1207/S15326942DN2203_3.
Luria, A. R. (1963). Restoration of Function after Brain Injury. Oxford,England: Macmillan.
Luria, A. R. (1973). The Working Brain: An Introduction toNeuropsychology. New York: Basic Books.
Mann, C. W., & Jewell, W. O. (1941). Configural aspects of humanlearning on the electrical maze. American Journal of Psychology,54(4), 536–545. Retrieved from: http://www.jstor.org/stable/1417203
Marcovitch, S., & Zelazo, P. D. (2009). A hierarchical competing systemsmodel of the emergence of early development of executive function.Developmental Science, 12(1), 1–18. doi:10.1111/j.1467-7687.2008.00754.x.
Maruff, P., Werth, J., Giordani, B., Caveneny, A. F., Feltner, D., &Snyder, P. J. (2006). A statistical approach for classifying changein cognitive function in individuals following pharmacologic chal-lenge: an example with alprazolam. Psychopharmacology, 186, 7–17. doi:10.1007/s00213-006-0331-5.
Mathias, J. L., & Kent, P. S. (1998). Neuropsychological consequences ofextreme weight loss and dietary restriction in patients with AnorexiaNervosa. Journal of Clinical and Experimental Neuropsychology,20(4), 548–564. doi:10.1076/jcen.20.4.548.1476.
Mathersul, D., Palmer, D. M., Gur, R. C., Gur, R. E., Cooper, N., Gordon,E., &Williams, L. M. (2009). Explicit identification and recognitionof facial emotions: II. Core domains and relationship with generalcognition. Journal of Clinical and Experimental Neuropsychology,31(3), 278–291. doi:10.1080/13803390802043619.
Mathewson, K., Dywan, J., Snyder, P. J., Tays, W., & Segalowitz, S.(2008). Aging and electrocortical response to error feedback duringa spatial learning task. Psychophysiology, 45(6), 936–948. doi:10.1111/j.1469-8986.2008.00699.x.
Matson, D. T., Berk, M., & Lucas, M. D. (1997). A neuropsychologicalstudy of prefrontal lobe function in the positive and negative sub-types of schizophrenia. The Journal of Genetic Psychology, 158(4),487–494. doi:10.1080/00221329709596685.
Melrose, J. A. (1922). The organismal point of view in the study of motorand mental learning. Psychological Review, 29(5), 390–405. doi:10.1037/h0071206.
Meyers, J. E., & Myers, K. E. (1995). Rey complex figure test under fourd i f f e r en t admin i s t r a t i on p rocedu re s . The C l in i ca lNeuropsychologist, 9(1), 63–67. doi:10.1080/13854049508402059.
Meyers, J. E., & Volbrecht, M. (1998). Validation of memory errorpatterns on the Rey Complex Figure and Recognition trial.Applied Neuropsychology, 5(3), 120–131. doi:10.1207/s15324826an0503_2.
Miles, W. (1927). The two-story duplicate maze. Tracing the stylus mazewith a maximum of indirect visual guidance. Journal ofExperimental Psychology, 10(5), 365–377. doi:10.1037/h0073950.
Milner, B. (1965). Visually-guided maze learning in man: Effects ofbilateral hippocampal, bilateral frontal, and unilateral cerebral le-sions. Neuropsychologia, 3(4), 317–338. doi:10.1016/0028-3932(65)90005-9.
Milner, B. (1971). Interhemisheric differences in the localization ofpsychological processes in man. British Medical Bulletin, 27(3),272–277. Retrieved from: http://bmb.oxfordjournals.org.
Milner, B., Corkin, S., & Teuber, H. L. (1968). Further analysis of thehippocampal amnesiac syndrome: 14-year follow-up study of H M.Neuropsychologia, 6, 215–234. doi:10.1016/0028-3932(68)90021-3.
Morrison, P. R., & Gates, G. R. (1988). Assessment of a microcomputer-based version of the Austin maze. The Journal of General
Psychology, 115(3), 307–314. doi:10.1080/00221309.1988.9710567.
Muenzinger, K. F. (1927). Physical and psychological reality.Psychological Review, 34(3), 220–233. doi:10.1037/h0071975.
Muenzinger, K. F., & Vine, D. O. (1941). Motivation in learning: IX. Theeffect of obstacles in human learning. Journal of ExperimentalPsychology, 29(1), 67–71. doi:10.1037/h0059118.
Newcombe, F., Ratcliff, G., & Damasio, H. (1987). Dissociable visualand spatial impairments following right posterior cerebral lesions:clinical, neuropsychological and anatomical evidence.Neuropsychologia, 25(1B), 149–161. doi:10.1016/0028-3932(87)90127-8.
Newcombe, F., & Russell, R. (1969). Dissociated visual perceptual andspatial deficits in focal lesions of the right hemisphere. Journal ofNeurology, Neurosurgery, and Psychiatry, 32, 73–81. Retrievedfrom: jnnp.bmj.com.
O’Brian, C. E., Bowden, S. C., Bardenhagen, F. J., & Cook, M. J. (2003).Neuropsychological correlates of hippocampal and rhinal cortexvolumes in patients with mesial temporal sclerosis. Hippocampus,13, 892–904. doi:10.1002/hipo.10128.
O’Keefe, J., & Nadel, L. (1978). The Hippocampus as a Cognitive Map.Oxford: Clarendon.
Olton, D. S. (1985). The temporal context of spatial memory. PhilosophicalTransactions of the Royal Society of London. Series B. BiologicalSciences, 308(1135), 79–86. doi:10.1098/rstb.1985.0011.
Orbach, J. O. (1955). Nonvisual functioning of occipital cortex in themonkey. Proceedings of the National Academy of Sciences of theUnited States of America, 41(5), 264–267. Retrieved from: http://www.jstor.org/stable/89113
Orbach, J. O. (1959). Disturbances of the maze habit following occipitalcortex removals in blind monkeys. A.M.A. Archives of Neurologyand Psychiatry, 81(1), 49–54. Retrieved from: http://archneurpsyc.jamanetwork.com.
Österberg, K., Karlson, B., & Hansen, A. M. (2009). Cognitive perfor-mance in patients with burnout, in relation to salivary cortisol.Stress, 12(1), 70–81. doi:10.1080/10253890802049699.
Österberg, K., Ørbæk, P., & Karlson, B. (2002). Neuropsychological TestPerformance of Swedish Multiple Chemical Sensitivity Patients—An Exploratory Study. Applied Neuropsychology, 9(3), 139–147.doi:10.1207/S15324826AN0903_2.
Österberg, K., Ørbæk, P., Karlson, B., Bergendorf, U., & Seger, L.(2000). A comparison of neuropsychological tests for the assess-ment of chronic toxic encephalopathy. American Journal ofIndustrial Medicine, 38(6), 666–680. doi:10.1002/1097-0274(200012)38:6<666::AID-AJIM7>3.0.CO;2-8.
Paul, R. H., Brickman, A. M., Cohen, R. A., Williams, L. M., Niaura, R.,Pogun, S., & Gordon, E. (2006). Cognitive status of younger andolder cigarette smokers: Data from the international brain database.Journal of Clinical Neuroscience, 13, 457–465. doi:10.1016/j.jocn.2005.04.012.
Paul, R., Grieve, S. M., Chaudary, B., Gordon, N., Lawrence, J., Cooper,N., & Gordon, E. (2009). Relative contributions of the cerebellarvermis and prefrontal lobe volumes on cognitive function across theadult lifespan.Neurobiology of Aging, 30(3), 457–465. doi:10.1016/j.neurobiolaging.2007.07.017.
Paul, R. H., Lawrence, J., Williams, L. M., Richard, C. C., Cooper, N., &Gordon, E. (2005). Preliminary validity of “IntegneuroTM”: A newcomputerized battery of neurocognitive tests. International Journalo f N e u ro s c i e n c e , 11 5 , 1 5 49–1567 . d o i : 1 0 . 1 0 8 0 /00207450590957890.
Pepper, S. C. (1934). The conceptual framework of Tolman’s purposivebehaviorism. Psychological Review, 41, 108–133. doi:10.1037/h0075220.
Perrin, F. A. C. (1914). An experimental and introspective study of thehuman learning process in the maze. Psychological Monographs,16(70), 1–97.
Neuropsychol Rev

Perrin, F. A. C. (1923). The Psychology of motivation. PsychologicalReview, 30(3), 176–191. doi:10.1037/h0072475.
Perry, R. B. (1918). Docility and purposiveness. The PsychologicalReview, 25(2), 1–20. doi:10.1037/h0072690.
Peters, H. N., & McLean, L. (1935). An experiment on orientation instylus maze learning. Journal of Experimental Psychology, 18(5),633–637. doi:10.1037/h0060054.
Peterson, J. (1920). The backward elimination of errors in mental mazelearning. Journal of Experimental Psychology, 3, 257–280. doi:10.1037/h0072487.
Peterson, J. (1922). Intelligence and learning. Psychological Review,29(5), 336–389. doi:10.1037/h0069483.
Pietrzak, R. H., Cohen, H., & Snyder, P. J. (2007). Spatial learningefficiency and error monitoring in normal aging: An investigationusing a novel hidden maze learning test. Archives of ClinicalNeuropsychology, 22, 235–245. doi:10.1016/j.acn.2007.01.018.
Pietrzak, R. H., Maruff, P., Mayes, L. C., Roman, S. A., Sosa, J. A., &Snyder, P. J. (2008a). An examination of the construct validity andfactor structure of the GrotonMaze Learning Test, a newmeasure ofspatial working memory, learning efficiency, and error monitoring.Archives of Clinical Neuropsychology, 23(4), 433–445. doi:10.1016/j.acn.2008.03.002.
Pietrzak, R. H., Maruff, P., & Snyder, P. J. (2009a). Convergent validityand effect of instruction modification on the Groton Maze LearningTest: A new measure of spatial working memory and error moni-toring. International Journal of Neuroscience, 119, 1137–1149. doi:10.1080/00207450902841269.
Pietrzak, R. H., Snyder, P. J., Jackson, C. E., Olver, J., Norman, T.,Piskulic, D., &Maruff, P. (2009b). Stability of cognitive impairmentin chronic schizophrenia over brief and intermediate re-test intervals.Human Psychopharmacology, 24, 113–121. doi:10.1002/hup.998.
Pietrzak, R. H., Sprague, A., & Snyder, P. J. (2008b). Trait impulsivenessand executive function in healthy young adults. Journal of Researchin Personality, 42, 1347–1351. Retrieved from: http://library.cogstate.com/private/publications/Pietrzak.
Porteus, S. D. (1918). The measurement of intelligence: six hundred andfifty-three children examined by the Binet and Porteus tests. Journalof Educational Psychology, 9(1), 13–31. doi:10.1037/h0075363.
Porteus, S. D. (1958). What do the maze tests measure? AustralianJournal of Psychology, 10(3), 245–256.
Porteus, S. D. (1965). Porteus Maze Test. Fifty Years Application.California: Pacific Books.
Porteus, S. D., & Kepner, R. M. (1944). Mental changes after bilaterallobotomy. Genetic Psychology Monographs, 29, 3–115.
Ratcliff, G., & Newcombe, F. (1973). Spatial orientation in man: effectsof left, right, and bilateral posterior cerebral lesions. Journal ofNeurology, Neurosurgery and Psychiatry, 36(3), 448–454. doi:10.1136/jnnp.36.3.448.
Razran, G. H. S. (1936). Attitudinal control of human conditioning.Journal of Psychology, 2, 27–37. doi:10.1080/00223980.1936.9917464.
Razran, G. H. S. (1939). Studies in configural conditioning. II. The effectof subjects’ attitudes and of task-sets upon configural conditioning.Journal of Experimental Psychology, 24(1), 95–105. doi:10.1037/h0062932.
Rettew, D. C., Cheslow, D. L., Rappoport, J. L., Leonard, H. L., &Lenane, M. C. (1991). Neuropsychological test performance intrichotillomania: A further link with obsessive-compulsive disorder.Journal of Anxiety Disorders, 5, 225–235. Retrieved from: http://www.sciencedirect.com.ezp.lib.unimelb.edu.au/science/journal/08876185
Ruckmick, C. A. (1921). II. William James. In Benteley, M. and themembers of the Psychological Seminary: Dynamic Principles inRecent Psychology. Psychological Monographs: Critical andExperimental Studies in Psychology From the University ofIllinois. Princeton N. J: Psychological Review Company, (pp 3–5)
Ruckmick, C. A. (1927). Some suggestions in Laboratory Apparatus. TheAmerican Journal of Psychology, 38(4), 647–648. Retrieved from:http://www.jstor.org/stable/1414400
Saling, M. M. (2009). Verbal memory in mesial temporal lobe epilepsy:beyond material specificity. Brain, 132(3), 570–582. doi:10.1093/brain/awp012.
Sartian, A. Q. (1940). The conception of the true path and efficiency inmaze learning. Journal of Experimental Psychology, 26(1), 74–93.doi:10.1037/h0063196.
Schofield, P. R., Williams, L. M., Paul, R. H., Gatt, J. M., Brown, K.,Luty, A., & Gordon, E. (2009). Disturbances in selective informa-tion processing associated with the BDNF Val66Met polymor-phism: Evidence from cognition, the P300 and fronto-hippocampalsystems. Biological Psychology, 80, 176–188. doi:10.1061/j.biopsych.2008.09.001.
Schroder, M. D., Snyder, P. J., Sielski, I., & Mayes, L. (2004). Impairedperformance of children exposed in utero to cocaine on a novel testof visuospatial working memory. Brain and Cognition, 55, 409–412. doi:10.1016/j.bandc.2004.02.062.
Scott, T. C. (1930). The retention and recognition of patterns in mazelearning. Journal of Experimental Psychology, 13(2), 164–207. doi:10.1037/h0071831.
Seckfort, D. L., Paul, R., Grieve, S. M., Vanderberg, B. M., Bryant, R. A.,Williams, L. M., & Gordon, E. (2008). Early life stress on brainstructure and function across the lifespan: A preliminary study. BrainImaging and Behavior, 2, 49–58. doi:10.1007/s11682-007-9015-y.
Semmes, J., Weinstein, S., Gent, L., & Teuber, H.-L. (1955). Spatialorientation in man after cerebral injury: I. Analysis by locus oflesion. The Journal of Psychology: Interdisciplinary and Applied,39(1), 227–244. doi:10.1080/00223980.1955.9916172.
Silverstein, S.M., Berten, S., Olson, P., Paul, R.,Williams, L.M., Cooper,N., & Gordon, E. (2007). Development and validation of a World-Wide-Web-based neurocognitive assessment battery: WebNeuro.Behavioral Research Methods, 39(4), 940–949. doi:10.3758/BF03192989.
Small, W. S. (1901). Experimental study of the mental processes of therat. II. The American Journal of Psychology, 12(2), 206–239.Retrieved from: http://www.jstor.org/stable/1412534
Snyder, P. F., Bednar, M. M., Cromer, J. R., & Maruff, P. (2005a).Reversal of scopolamine induced deficits with a single dose ofdonepezil, an acetylcholinesterase inhibitor. Alzheimer’s andDementia, 1, 126–135. doi:10.1016/j.jalz.2005.09.004.
Snyder, P. J., Jackson, C. E., Piskulic, D., Olver, J., Norman, T., &Maruff,P. (2008a). Spatial working memory and problem solving in schizo-phrenia: The effect of symptom stabilization with atypical antipsy-chotic medication. Psychiatry Research, 160(3), 316–326. doi:10.1016/j.psychres.2007.07.011.
Snyder, A. M., Maruff, P., Pietrzak, R. H., Cromer, J. R., & Snyder, P. J.(2008b). Effect of treatment with stimulant medication on nonverbalexecutive function and visuomotor speed in children with attentiondeficit/hyperactivity disorder. Child Neuropsychology, 14(3), 211–226. doi:10.1080/09297040701220005.
Snyder, P. J., Werth, J., Giordani, B., Caveney, A. F., Feltner, D., &Maruff, P. (2005b). A method for determining the magnitude ofchange across different cognitive functions in clinical trials: Theeffect of acute administration of two different doses alprazolam.Human Psychopharmacology: Clinical and Experimental, 20,263–273. doi:10.1002/hup.692.
Spence, K. W. (1932). The order of eliminating blinds in maze learningby the rat. Journal of Comparative Psychology, 14(1), 9–27. doi:10.1037/h0075997.
Stephens, S. S. (1935). The operational definition of psychological concepts.Psychological Review, 42(6), 517–527. doi:10.1037/h005697.
Steinberg Chaikelson, J., & Schwartzman, A. E. (1983). Cognitivechanges with aging in schizophrenia. Journal of ClinicalPsychology, 39(1), 25–30. doi:10.1002/1097-4679(198301)39.
Neuropsychol Rev

Talland, G. A., Hagen, D. Q., & James, M. (1967). Performance tests ofamnesiac patients with Cylert. The Journal of Nervous and MentalDisease, 144(5), 421–429. Retrieved from: http://journals.lww.com/jonmd/pages/currenttoc.aspx.
Talland, G. A., & McGuire, M. T. (1967). Tests of learning and memorywith Cylert. Psychopharmacologia, 10, 445–451. doi:10.1007/BF00403986.
Tate, D. F., Conley, J., Paul, R. H., Coop, K., Zhang, S., Zhou, W., &Tashima, K. (2010). Quantitative diffusion tensor imagingtractography metrics are associated with cognitive performanceamong HIV-Infected patients. Brain Imaging and Behavior, 4, 68–79. doi:10.1007/s11682-009-9086-z.
Teuber, H.-L. (1963). Space perception and its disturbances after braininjury in man. Neuropsychologia, 1, 47–57. doi:10.1016/0028-3932(63)90012-5.
Teuber, H.-L. (1966). Alteration of perception after brain injury. In J. C.Eccles (Ed.), Brain and Conscious Experience (pp. 182–216). NewYork: Springer.
Thomas, E., Reeve, R., Fredrickson,A., &Maruff, P. (2011). Spatialmemoryand executive functions in children. Child Neuropsychology, 17(6),599–615. doi:10.1080/09297049.2011.567980.
Thomas, E., Reeve, R., Pietrzak, R., & Maruff, P. (2013). Disentanglingcomponent learning and executive processes in hidden pathwaymaze learning in children: A process-based approach. ChildNeuropsychology, 19(6), 588–600. doi:10.1080/09297049.2012.704010.
Thomas, E., Snyder, P. J., Pietrzak, R. H., Jackson, C. E., Bednar, M., &Maruff, P. (2008). Specific impairments in visuospatial working andshort-term memory following low dose scopolamine challenge inhealthy older adults.Neuropsychologia, 46(10), 2476–2484. doi:10.1016/j.neuropsychologia.2008.04.010.
Thurstone, L. L. (1933). The error function in maze learning. The Journalof General Psychology, 9(2), 288–301. doi:10.1080/00221309.1933.9920938.
Tilborg, P. W. V. (1936). The retention of mental and finger maze habits.Journal of Experimental Psychology, 19(3), 334–341. doi:10.1037/h0059048.
Tolman, E. C. (1938). The determiners of behavior at a choice point.Psychological Review, 45(1), 1–41. Retrieved from: http://psycnet.apa.org/journals/rev/45/1/.
Tolman, E. C. (1948). Cognitive maps in rats andmen. The PsychologicalReview, 55(4), 189–208. Retrieved from: http://psychclassics.yorku.ca/Tolman/Maps/maps.htm.
Tolman, E. C. (1959). Performance vectors: A theoretical and experimen-tal attack upon emphasis, effect, and repression. The AmericanPsychologist, 14(1), 1–7. doi:10.1037/h0044061.
Tolman, E. C., Hall, C. S., & Bretnall, E. P. (1932). A disproof of the lawof effect and a substitution of the laws of emphasis, motivation anddisruption. Journal of Experimental Psychology, 15(6), 601–614.doi:10.1037/h0073609.
Trumball Ladd, G. (1894). President’s address before the NewYork meeting of the American Psychological Association.The Psychological Reviews, 1(1), 1–21. doi:10.1037/h0064711.
Tucker, A., Kinsella, G., Gawith, M., &Harrison, G. (1987). Performanceon the Austin Maze: Steps towards normative data. AustralianPsychologist, 22(3), 353–359. doi:10.1080/00050068708259536.
Van Horn, J. D., Gold, J. M., Esposito, J. L., Ostrem, V. M., Mattay, V.,Weinberger, D. R., & Berger, K. F. (1998). Changing patterns ofbrain activation during maze learning. Brain Research, 793(1–2),29–38. doi:10.1016/S0006-8993(98)00051-1.
Voeks, V. W. (1948). Postremity, Recency, and Frequency as bases forprediction in the maze situation. Journal of ExperimentalPsychology, 38(5), 495–510. Retrieved from: http://psycnet.apa.org/journals/xge/38/5/.
Wallesch, C.-W., Karnath, H. O., Papagno, C., Zimmerman, P., Deuschl,G., & Lucking, C. H. (1990). Parkinson’s disease patient’s behaviorin a covered maze learning task.Neuropsychologia, 28(8), 839–849.doi:10.1016/0028-3932(90)90008-C.
Walsh, K. W. (1960). Cited in Darby, D., & Walsh, K. (2005). Walsh’sNeuropsychology. A Clinical Approach. (5th Edition). Edinburg:Elsevier. (pp. 149–150, 167)
Walsh, K. W. (1985). Understanding Brain Damage: A Primer onNeuropsychological Evaluation. Edinburough: ChurchillLivingstone
Walton, N. H., & Bowden, S. C. (1997). Does liver dysfunction explainneuropsychological status in recently detoxified alcohol-dependentclients? Alcohol and Alcoholism, 32(3), 287–295.
Warden, C. J. (1924a). Primacy and recency as factors in cul-de-sacelimination in a stylus maze. Journal of Experimental Psychology,7(2), 98–116. doi:10.1037/h0074915.
Warden, C. J. (1924b). The relative economy of various modes of attackin the mastery of the stylus maze. Journal of ExperimentalPsychology, 7(4), 243–275. doi:10.1037/h0073906.
Warden, C. J. (1925). Judgments of certain space relations based upon thelearning of a stylus maze. Journal of Experimental Psychology, 8(6),399–407. doi:10.1037/h0064260.
Welsh, M. C., Pennington, B. F., & Grossier, D. B. (1991). A normative-developmental study of executive function: Awindow on prefrontalfunction in children. Developmental Neuropsychology, 7(2), 131–149. Retrieved from: http://www.erlbaum.com/Journals/journals/DN/dn.htm.
Neuropsychol Rev