how hydroperiod and species richness affect the balance of resource flows across aquatic-terrestrial...
TRANSCRIPT

RESEARCH ARTICLE
How hydroperiod and species richness affect the balanceof resource flows across aquatic-terrestrial habitats
Tiffany A. Schriever • M. W. Cadotte •
D. Dudley Williams
Received: 17 February 2013 / Accepted: 9 October 2013 / Published online: 18 October 2013
� Springer Basel 2013
Abstract Ecosystem functioning is influenced by the
flow of nutrients, detritus, and organisms. Variation in
these flows, like that found in temporary ecosystems,
affects temporal and spatial patterns of community diver-
sity and secondary production. We evaluated the influence
of hydroperiod and ecosystem size on the bi-directional
flow of subsidies from intermittent ponds and surrounding
forests by quantifying litter deposition and the abundance
and biomass of emerging insects and amphibians. In
addition, we assessed whether amphibian and insect
diversity influenced the magnitude of cross-habitat
resource flux. We found substantial spatial and temporal
variation in the magnitude, composition, and timing of
cross-habitat resource subsidies. Overall, deposition into
ponds far exceeded biomass exported via insect and
amphibian emergence. We found a negative association
between resource flux and the diversity of amphibians and
insects. Different species groups contributed to flux pat-
terns unequally, with insects having higher diversity but
lower flux compared to amphibians. Organismal flux varied
among ponds with amphibians having the highest flux in
the shortest hydroperiod pond and insect flux was highest
from an intermediate hydroperiod pond. This work reveals
how variation in pond size and permanence affects species
diversity and ecosystem flows. Species composition played
a major role in flux differences across ponds. Further, given
the general lack of research and conservation prioritization
of temporary ponds, uncovering how these ponds contrib-
ute to cross-habitat linkages is necessary to develop fully
integrated management strategies.
Keywords Amphibians �Biodiversity–ecosystem function � Biomass �Cross-habitat energy flow � Disturbance � Insects �Subsidy hypothesis
Introduction
Habitats are invariably open systems linked by the flow
of nutrients, detritus, and organisms, all of which have
the potential to influence population and community
dynamics, food webs, and diversity-stability relation-
ships in the recipient habitat (Polis et al. 1997; Nowlin
et al. 2008; McCoy et al. 2009). Spatial flows may also
have a large influence at the meta-ecosystem scale,
thereby strengthening connections between local eco-
systems (Loreau et al. 2003). Spatial subsidies are
resources that are spatially and temporally variable,
originating in one donor habitat and moving into another
(Polis et al. 1997). The flow of resources across habitat
boundaries is influenced by the distance separating
habitats, habitat size, perimeter-to-area ratio of the focal
habitat, adjacent habitat type, and the behavior of fluxes
Electronic supplementary material The online version of thisarticle (doi:10.1007/s00027-013-0320-9) contains supplementarymaterial, which is available to authorized users.
T. A. Schriever � M. W. Cadotte � D. D. Williams
Ecology and Evolutionary Biology, University of Toronto,
25 Willcocks Street, Room 3055, Toronto M5S 3B2, Canada
T. A. Schriever � M. W. Cadotte � D. D. Williams
Department of Biological Sciences, University of Toronto
Scarborough, 1265 Military Trail, Toronto, ON M1C1A4,
Canada
Present Address:
T. A. Schriever (&)
Department of Zoology, Cordley Hall 3029, Oregon State
University, Corvallis, OR 97331-2914, USA
e-mail: [email protected]
Aquat Sci (2014) 76:131–143
DOI 10.1007/s00027-013-0320-9 Aquatic Sciences
123

that move between habitats (Cadenasso et al. 2004).
Spatial subsides can enter the food web at multiple tro-
phic levels (Polis et al. 1997) and vary in quantity and
quality (Massol et al. 2011).
Aquatic food webs are intimately linked with the sur-
rounding terrestrial landscape through a number of energy-
flow pathways. For example, aquatic systems contribute to
the terrestrial food web via aquatic insect emergences
(Benke 1993; Stagliano et al. 1998; Gratton et al. 2008) and
the reciprocal return of terrestrial material to aquatic food
webs through litterfall, such as organic matter deposition in
streams (Fisher and Likens 1973; Hutchens and Wallace
2002; Rubbo and Kiesecker 2004; Rubbo et al. 2006),
insect infall (periodical cicada deposition into streams and
ponds; (Pray et al. 2009), and nitrogen deposition (geese
deposit nitrogen-rich waste into wetlands gained from
foraging in agricultural land; (Post et al. 1998; Kitchell
et al. 1999). Prey subsidies, such as emerging freshwater
insects, have been shown to increase the abundance of
riparian arthropod assemblages (Hoekman et al. 2011;
Dreyer et al. 2012), or in a case where the aquatic insect
subsidy has been restricted, the abundance and biomass of
riparian lizards and arthropods was reduced (Sabo and
Power 2002). In addition, recent work has revealed that not
only the magnitude, but also the functional role of the
spatial prey subsidy is important in determining the impact
on recipient food webs (Leroux and Loreau 2008; Wesner
2010, 2012).
Woodland ponds have diverse communities of insects
and amphibians. Species in both groups undergo meta-
morphosis and leave the aquatic habitat and enter the
terrestrial realm, making them important sources of sub-
sidies between habitats (Schreiber and Rudolf 2008). These
ponds are often relatively small, have high perimeter-to-
area ratios, and are abundant in the landscape (Palik et al.
2001, 2006). In aquatic systems, environmental variation in
the form of duration (i.e., hydroperiod) influences species
richness, diversity, community composition, and survivor-
ship and has direct consequences for secondary production
(Welborn et al. 1996; Sabo and Post 2008). Freshwater
habitats along the hydroperiod gradient range from tem-
porary habitats with relatively few small-bodied predators,
transitioning to permanent fishless habitats with large
invertebrate predators, and finally habitats with fish (Wel-
born et al. 1996). It is unclear how environmental
variability interacts with species richness to influence
cross-habitat energy flow. A considerable accumulation of
published research has found that greater biodiversity
results in higher ecosystem functioning (i.e. productivity
and energy flow; Loreau et al. 2001). The question remains
of how resource subsidies and environmental variability
affects the biodiversity–ecosystem function relationship
(Romanuk et al. 2010).
There are few studies that have quantified the impor-
tance and reciprocal connection of temporary ponds to the
surrounding landscape (but see Palik et al. 2006; Palik and
Kastendick 2010). Given the extensive occurrence of
temporary ponds in temperate regions, ignoring their con-
nections to surrounding habitat hinders our ability to
adequately conserve and manage biota and the resulting
ecosystem services provided by these systems.
Here we present a case study that examined the envi-
ronmental (hydroperiod and ecosystem size) and
community (species richness, diversity) influences on the
magnitude of subsidy flows across habitat boundaries
(strength of habitat coupling). We conducted a natural
experiment using seasonally intermittent freshwater ponds
(classification follows Williams 2006) varying in hydro-
period and size. We quantified the aquatic export of insect
emergence and amphibian metamorphosis leaving ponds
and the reciprocal terrestrial input of organic matter
deposition of litterfall entering the ponds. Using this data,
we tested three hypotheses: First was that the predictable
shift in community composition across a hydroperiod
gradient will have a noticeable and positive effect on the
magnitude and/or composition of the cross-habitat resource
flows. Second, we hypothesized that pond hydroperiod and
size would influence the magnitude of the aquatic-terres-
trial linkage. In particular, we predict that longer
hydroperiods and larger ecosystem size will have higher
amphibian and insect emergence and biomass than shorter
hydroperiod ponds. Finally, we tested for a relationship
between diversity and productivity among ponds. Little is
known about how energy flow influences these biodiversity
effects. Since temporary woodland ponds are often small
and their food webs rely on detrital leaf litter, it is likely
that litter fall will scale with pond size, and we predict the
amount of litter deposition will be directly proportional to
the amount of animal export.
Methods
Study sites and physical habitat
We conducted this investigation at the Queen’s Biological
Station north of Kingston, Ontario, Canada (44.565977 N,
-76.324223 W). The area is predominantly mixed conif-
erous-deciduous forest and supports numerous types of
ponds. Ponds are filled by snowmelt in early spring and
lose water as summer progresses, resulting in some ponds
drying completely. We used four temporary ponds that
varied in hydroperiod and size (hereafter, Short, Interme-
diate 1, 2, 3; Table 1). Our experimental design took
advantage of a natural environmental gradient, capitalizing
on spatial and temporal variation; but this resulted in a
132 T. A. Schriever et al.
123

trade-off in replication. We were most interested in mea-
suring responses to natural environmental variation and
laying a foundation for an unknown system. We used
similar, non-replicated natural studies to check the gener-
ality of our results.
Hydroperiod was measured as the consecutive number
of days with water following ice off to complete drying in
2010. We measured pond area each month by mapping the
perimeter of each using a Trimble TSC1 GPS connected to
a ProXRS satellite and RTCM receiver. All mapping was
completed in one or two consecutive days within each
month. The area (m2) of each pond was then calculated in
ArcMap10 (Esri, Redlands, CA, USA) and averaged to
yield an average pond area for the entire study period.
Every month, we measured dissolved oxygen (mg/L),
water depth (m) and percent canopy cover measured at the
center deepest point in each pond. We used a hand-held
Hydrolab Quanta multiparameter probe (Hach Environ-
mental, Loveland, CO) to measure dissolved oxygen. We
took four canopy readings facing each cardinal direction,
counting the number of open squares on a spherical den-
sitometer (Forest Densiometers Bartlesville, OK, USA).
The four readings were averaged, multiplied by 4.17, and
the product subtracted from 100 % to obtain percent can-
opy density (California Department of Pesticide
Regulation, Environmental Monitoring Branch, SOP
number: FSOT.002.01). Data loggers placed at the center
of each pond *3 cm from pond bottom (StowAway Tid-
bit; Onset computer, Pocasset, MA, USA) recorded water
temperature every 5 h. These were averaged for a 24 h
period to acquire a daily temperature reading during the
period of 7 April to 14 October 2010 or until the pond
dried.
Field and laboratory methodology
To quantify emerging aquatic insects, we placed three
floating emergence traps (total trap area 0.63 m2) in each
pond. Traps were deployed in each pond from May to
August 2010 on the same day in each pond for *48 h
collection periods. We made four collections in May, three
collections each in June and July, and two in August for a
total of 2032.5 trap hours. Insects were aspirated from the
traps and placed into vials containing 80 % ethanol. All
aquatic insects were counted and identified to family.
Insects of non-aquatic origin (e.g. ants) were removed and
not used in further analyses. We stopped trapping for
emergent insects after the pond dried. Since ponds did not
dry on the same day, the number of trapping days per pond
varied. We measured the body length (mm) of a subsample
of random individuals in each family (refer to Online
Resource Table 1 for sample numbers) using a Nikon
SMZ1500 microscope connected to a computer using the
NIS Elements D3.1 software (Shinagaw-ku, Tokyo, Japan).
We used published length-mass conversions (Online
Resource Table 2) to estimate dry mass (DM) for each
individual and calculated biomass for each family by
summing biomass of all constituent individuals (Online
Resource Table 1). Studies have found preservation in
alcohol reduces invertebrate dry mass due to leaching
(Leuven et al. 1985; von Schiller and Solimini 2005;
Edwards et al. 2009), but this loss of dry mass was cor-
rected by using length-mass equations from non-preserved
specimens (von Schiller and Solimini 2005). Aquatic insect
emergence (Ei; g C m-2) was calculated for each pond
using dry mass estimates divided by trap area.
To capture metamorphosing amphibians, on June 4–6
we encircled each pond and sank pitfall traps (five gal
buckets) every 10 m flush with the fence and the ground
along the pond side of the fence (Gibbons 1974). Ponds
were encircled once breeding was over and before meta-
morphs emerged. We monitored tadpole development in
ponds on a bi-weekly schedule starting April 5, 2010 to
ensure we did not miss metamorph emergence. Each trap
contained a wet sponge to provide moisture to sustain
trapped amphibians. This method allowed us to capture
every amphibian moving out of the ponds, although tree-
frogs can climb over fences, and therefore their abundance
Table 1 Physical characteristics of the study ponds measured in 2010
Measurement Pond
Short Intermediate 1 Intermediate 2 Intermediate 3
Mean depth (m) 0.26 (0.20) 0.47 (0.33) 0.25 (0.16) 0.36 (0.17)
Mean water temperature (�C) 14.43 (5.23) 18.32 (4.86) 14.67 (4.29) 16.01 (4.14)
Mean dissolved oxygen (mg/L) 6.68 (1.06) 6.69 (2.15) 5.43 (1.34) 7.18 (1.40)
Mean canopy cover (%) 74.54 (5.57) 31.67 (11.59) 87.31 (12.72) 26.48 (8.93)
Mean pond area (m2) 182.68 (76.47) 987.48 (343.97) 249.13 (157.30) 188.52 (84.56)
Mean perimeter:area (m) 0.43 (0.13) 0.20 (0.03) 0.61 (0.34) 0.37 (0.11)
Hydroperiod (days) 78 145 168 168
Standard deviation is in parentheses
Spatial subsidies between pond and forest habitats 133
123

may have been underestimated. Pitfall traps were opened
and checked daily for all ponds during the periods of 8–10
and 20–29 June, 7–13 and 19–23 July, 9–12 and 16–18
August. Intermediate 3 pond traps were also open 28–29
August because it was the only pond remaining with water.
In total, traps were open for 99 collection days. We stopped
trapping for metamorphs once the pond completely dried
and we collected zero metamorphs for several trapping
days following drying. All amphibians collected were
counted, identified to species, weighed, and the snout-to-
vent length (SVL) measured with digital calipers. We
euthanized a small sample of individuals for biomass
estimation (sample sizes listed in Online Resource
Table 3). Euthanized individuals were dried at 80 �C for
5–7 days, weighed (DM), then ashed at 500 �C for 1–2 h
and reweighed to determine ash-free dry mass (AFDM).
We established species-specific length–DM and length-
AFDM relationships using linear regression to estimate dry
mass and AFDM from the SVL of all individuals collected
(Online Resource Table 3). The DM values were summed
to provide the total biomass of amphibians from each pond.
Amphibian emergence production (Ea; g C m-2) was cal-
culated for each pond using dry mass estimates divided by
pond area.
Litterfall is the main source of terrestrial organic matter
and is a main food resource for many invertebrates and
amphibians within these woodland ponds. Therefore, we
collected overhead inputs of coarse terrestrial particulate
organic carbon (i.e., litterfall) once a month from two litter
traps (0.5 m2 each) placed in each pond from August 29
through November 14, 2010. Trap locations were haphaz-
ardly selected within the wetted area of the ponds and
remained in the initial location throughout the 3 months of
sampling. Litterfall was removed from each trap and
placed in paper bags. In the laboratory, bags of collected
litter were emptied onto a white enamel tray, sorted into
deciduous leaf (mostly Quercus sp., Acer sp., Betula sp.),
coniferous needle (Pinus sp.), woody material (twigs,
branches, bark, cones, etc.), and miscellaneous (bud scales,
pollen, etc.) components, dried at 55 �C for at least 96 h,
and weighed (g DM). We quantified litter dry mass depo-
sition for each trap (Dareal; g m-2). Total litter deposition
(i.e., biomass) for each pond was the sum of litter dry mass
collected over the sampling period.
We used Gratton and Vander Zanden’s (2009) measure
of flux because it incorporates habitat geometry into the
calculation of cross-habitat resource flow. They define ‘‘the
flux (F) of emerging aquatic insects to land as the amount
of insect production annually leaving the body of water per
meter of aquatic shoreline (Fi; g C m-1 year-1)’’. We used
this metric of flux for amphibians (Fa) as well since they
are equivalent in terms of leaving the aquatic habitat. The
calculation was: F ¼ E � ðA=pÞ, where E is the emergence
in g C m-2 year-1of insects (Ei) or amphibians (Ea), A is
the pond area and p is pond perimeter. C represents grams
of dry mass carbon. The study ponds were not perfectly
circular, therefore a shoreline development factor ðDL ¼p=½ pAð Þ1=2�Þ corrected for irregularities in pond shape. We
used the equation F ¼ Eðr=2DLÞ, where r is pond radius.
Litter input per meter of shoreline (Fl), and flux to pond
was calculated as Dareal r=2DLð Þ. We also used a measure
of total flux or productivity to estimate the total aquatic to
terrestrial carbon flux (Etotal) and total terrestrial to aquatic
carbon flux (Dtotal) (Vander Zanden and Gratton 2011). We
calculated total emergence numbers and total production
for each site for insects (Eitotal) and amphibians (Eatotal)
separately because we were interested in the magnitude of
each subsidy type (Ei total ¼ Ei � A ; Ea total ¼ Ea � AAn
Etotal was calculated for each insect family and amphibian
species per pond as well as the Etotal for each pond. Lit-
terfall total was calculated in a similar fashion
(Dtotal = Dareal 9 A; g C y-1).
Statistical analyses
To test if the magnitude of resource flows changed over the
study period, we used general linear models. We calculated
Friedman rank sum test using friedman.test function in R, a
nonparametric version of one-way ANOVA with repeated
measures to evaluate differences in biomass export and
litterfall input among the ponds across repeated sampling
events. We did not use one-way ANOVA because the
assumption of equal variances of the differences between
repeat observations was not met and because of the small
sample size (four ponds). For these data, emergence and
litterfall are the dependent variables, the day of year
(DOY) the sample was collected was the repeated mea-
sures factor (‘‘block’’) and pond was the group factor. An
analysis of covariance (ANCOVA) tested for differences in
total flux between species. We calculated Bray–Curtis
distances on a matrix of insect family flux and amphibian
species flux values by pond. Dissimilarities were multiplied
by 100 and the result subtracted from 100 to get a measure
of percent similarity. We used simple linear correlations to
calculate Pearson product-moment correlation coefficients
in R using the cor and cor.test functions (R Development
Core Team 2009) to compare the relationship between
pond size and hydroperiod on the export fluxes of insects
(i.e., Eitotal, Fi) and amphibians (i.e., Eatotal, Fa), and the
input flux of litterfall (i.e., Dtotal, Fl). Ponds vary monthly
in size and shape, so we used mean pond area (m2) as
calculated from the available wetted months as the pond-
size variable.
We calculated species richness and Shannon diversity
(H0) for amphibians and insects (familial richness and
diversity) using the Biodiversity Calculator provided by J.
134 T. A. Schriever et al.
123

Danoff-Burg and C. Xu (http://www.columbia.edu/itc/cerc/
danoff-burg/Biodiversity%20Calculator.xls) using abun-
dance data (total numbers of each taxon collected per site).
We tested for differences between emergent amphibian and
insect communities in richness, diversity, and evenness
with t tests. We also looked at the relationship between
litterfall and pond canopy cover. Our rule of thumb for
interpreting strengths of correlations was very strong if r =
±0.70 or higher, strong if r = ±0.40–0.69, moderate if
r ± 0.30–0.39, weak if r = ±0.20–0.29, and there was no
relationship if r = ±0.01 to 0.19. Our results are reported
as r (df) = correlation coefficient, P = P value.
Results
Timing of resource flows and environmental influences
Different forms of resource flows occurred at distinctly
different times. Insects emerged prior to amphibian meta-
morphosis from the ponds (Fig. 1) and litterfall input
occurred after emergence (day 263, 20 September).
Emergent insect export began before and lasted almost as
long as the export from amphibians. Insect emergence
abundance peaked first in Short pond (14–16 May) fol-
lowed by Intermediate 1 (19–21 July) and then
Intermediate 2 and 3 ponds (16–18 August). Insect emer-
gence increased over the four-month sampling period
(R2 = 0.1, F48 = 5.34, P = 0.025; Fig. 1) with higher
emergences just prior to pond drying. Amphibians had two
major waves of emergence that were responsible for 43 %
of the total abundance collected (23–25 June and 8–10
July) which corresponded to the drawdown in one pond and
the increase in temperature in another (Online Resource
Fig. 1). The greatest number of amphibians (n = 154)
leaving the ponds at one time occurred on 24 June with
most individuals leaving from Short pond (121 wood frog
metamorphs, Lithobates sylvaticus, and 1 newt, Notoph-
thalmus viridescens) and the remainder from Intermediate
1 (33 wood frog metamorphs). The number of metamor-
phosing amphibians declined over the sampling period
(R2 = 0.09, F98 = 9.331, P = 0.003; Fig. 1).
Litter fall amounts were similar across sampling months
(mean g m-2 ± SD; 430.60 ± 147.68 in Aug.–Sep.,
413.93 ± 169.50 in Sept.–Oct., and 474.05 ± 166.54 in
Oct.–Nov.). However, the timing of maximum litter fall
drop varied among ponds.
Magnitude of fluxes
We collected 1,050 individuals representing a total insect
flux (Eitotal) of 3418.85 g C year-1 from aquatic to terrestrial
habitat. This constituted a flux of 498.24 g C m-1 year-1 or
the emergent insect flux per meter of pond margin. Inter-
mediate 1 accounted for 95 % of the total insect flux with
the highest abundance of emerging insects (791 individuals
m-2), emergence production (Ei = 3.32 g C m-2) and flux
(Fi = 474.07 g C m-1 year-1) (Table 2). High flux was
driven by the presence of odonate taxa. In terms of abun-
dance, Chironomidae midges made up 68 % of collected
individuals, but contributed only 2.4 % to insect flux, Fi
(6.7 % of Eitotal) of all insect families (Fig. 2a). Lestid
damselflies were important contributors to the emergent
insect flux (83 %; Eitotal = 2840.63 g C year-1) even
though they represented only 0.004 % of collected individ-
uals (Fig. 2b). Limnephilidae caddisflies were collected
from all ponds and, although not abundant (n = 26), they
contributed a flux of 25.27 g C m-1 year-1 (Fig. 2a) or 5 %
of Fi and Eitotal (172.16 g C year-1). The number of
emerging insects from each pond was heterogeneous across
sampling events (Friedman rank sum test: v2 (3,
N = 12) = 12.897, P = 0.005). We found that the simi-
larity of insect total flux was the lowest for Short and
Intermediate 1 ponds (0.5 %), whereas the highest similarity
was between Short and Intermediate 2 ponds (31 %). Dis-
similarity between Intermediate 1 and Short ponds resulted
from differences in the flux from Chironomidae, Lestidae,
and Limnephilidae which made up 95 % of total flux
0
50
100
150
200
250
300
Day of year
Em
erge
nce
(no.
/day
)
amphibianinsect
124145
157172
174176
178180
189191
193200
202221
223228
230
Fig. 1 Temporal variation in insect and amphibian emergence
throughout the study period. Insect collection was from day 124 to
230 (May 4th–August 18th), amphibian collection occurred between
day 171 and 241 (June 8th–August 29th). Each point represents the
mean number of captures and the SD across ponds per collection day.
Overall, amphibian peak emergence occurred between days 174 and
176 (23–25 June), while insect peak emergence occurred between
days 200 and 202 (19–21 July)
Spatial subsidies between pond and forest habitats 135
123

leaving Intermediate 1 pond and only 0.5 % from Short. It
also is important to note Intermediate 1 had the highest
insect familial richness (25) and Short the lowest (9).
The total amphibian flux and flux per meter of pond
margin was nearly 6 9 higher than that from insects
(Eatotal = 18,833.9 g C year-1; Fa = 2899.33 g C m-1
year-1, Fig. 2c) and the emergence 18 9 higher (mean
Ea = 18.03 g C m-2) than insect emergence (mean
Ei = 1.00 g C m-2). The maximum number of amphibians
emerging from the four ponds ranged from 123 individuals
at Intermediate 3 pond to 571 individuals from Short pond
between late June and late August 2010 (total number
collected, n = 1197). A Friedman test was conducted to
evaluate differences in the number of individuals emerging
among the ponds across sampling events. The test was
significant, indicating that the number of emerging
amphibians varied among ponds and time steps (v2 (3,
N = 28) = 10.1774, P = 0.02), and signifying emergence
is heterogenous across ponds and time. The pond with the
shortest hydroperiod (78 days) had the densest (3 individ-
uals/m2) and highest emergence (Ea = 41.9 g C m-2
year-1) of amphibians. Emerging wood frog metamorphs
(L. sylvaticus) represented 71 % of all individuals collected
and 63 % of the amphibian flux (Fa) across all ponds to the
terrestrial landscape (Fig. 2c). Intermediate 1 and 2 were
most similar (49 %) in terms of amphibian flux and that
Intermediate 3 and Short ponds were the least similar
(1 %). These differences are because Intermediate 3 was
dominated by P. crucifer, whereas Short pond had very
little biomass from this species. The majority of flux from
Short ponds originated from L. sylvaticus, which was not
present in Intermediate 3, indicating species composition
played a major role in flux differences across ponds. Eatotal
significantly differed among species (ANCOVA: df = 7,
F = 3.66, P = 0.01). A post hoc holm adjustment showed
pairs of L. sylvaticus–A. maculatum (P = 0.035), L. syl-
vaticus–Ambystoma sp. (P = 0.016) and N. viridescens–L.
sylvaticus (P = 0.016) were significantly different from
each other in their contribution to the total flux of energy
from aquatic to terrestrial habitats.
Overall, there was a carbon deposition into ponds that
exceeded that exported via insect and amphibian emer-
gence (Fig. 3). The average net energy gain [Dtotal-
(Eatotal ? Eitotal)] across ponds was 41,279.76 g C year-1
(range 21,019.83–88,312.94 g C year-1). This subsidy
showed spatial variation across ponds (32.5 % difference
in Dtotal from lowest input to highest input), but resource
subsidy input did not differ across sampling events
(Friedman rank sum test: v2 (3, N = 12 = 0.2, P = 0.98).
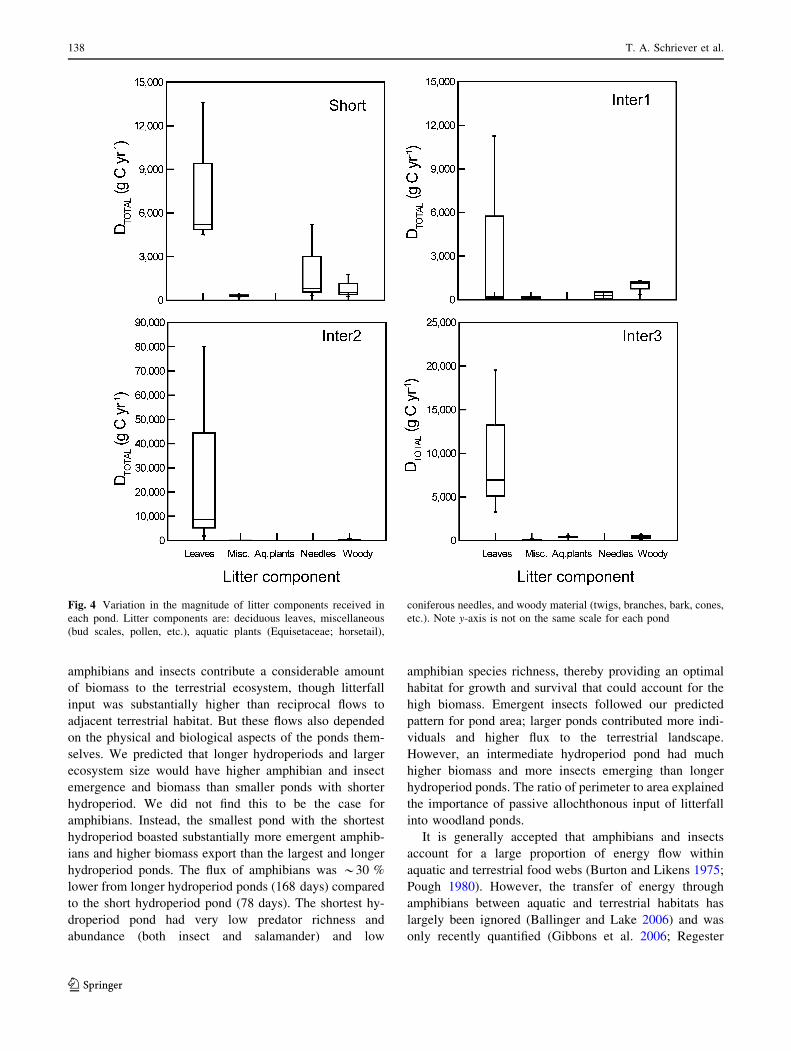
Deciduous leaves were the most important litter resource
for all ponds (Fig. 4). Coniferous needles were of sec-
ondary importance to one pond (Short). As assumed,
litterfall deposition into ponds was directly related to
canopy cover over the pond (GLM, poisson: P \ 0.0001,
df = 3, Dtotal = 9.795 ? 0.0159mean percent cover).
Diversity–ecosystem function response
We found that insects and amphibians had different
diversity–ecosystem function relationships. Total insect
flux (Eitotal) and flux (Fi) per meter of pond margin were
highest from the least diverse pond (Fig. 5). However, total
insect flux increased with increasing taxa richness (Fig. 5).
The amphibian flux (Fa) per meter of pond margin did not
show a consistent pattern with higher flux amounts from
the least and most diverse ponds (Fig. 5). Amphibian total
flux (Eatotal) also failed to produce a pattern with amphibian
richness, though ponds varied little in amphibian richness.
We also evaluated the flux by diversity relationships using
family level resolution for amphibians because that is the
level of resolution we have for insects, and found the
relationships were the same regardless of using amphibian
species richness or family richness.
Ponds with longer hydroperiod tended to have higher
amphibian diversity (r (2) = 0.56, P = 0.44) and evenness
(r (2) = 0.68, P = 0.32). Amphibian species richness was
not linearly related to hydroperiod (r (2) = -0.11,
P = 0.89), but rather peaked at intermediate hydroperiod.
Measures of amphibian diversity showed strong positive
relationships with pond area (richness: r (2) = 0.90,
Table 2 Summary of flux
values for insects and
amphibians
Emergent flux per m pond
margin (F) takes into
consideration the shoreline
developmental factor (DL), total
flux is the total aquatic to
terrestrial flux of carbon (Etotal)a Eshore in Vander Zanden and
Gratton (2011)b Eareal in Vander Zanden and
Gratton (2011)
Short Intermediate 1 Intermediate 2 Intermediate 3
Insects
Emergent flux (Fi, g C m-1 year-1)a 1.23 474.07 3.58 19.35
Emergence (Ei, g C m-2)b 0.04 3.32 0.12 0.53
Total flux (Eitotal, g C year-1) 7.4 3281.2 30.2 100.04
Amphibians
Emergent flux (Fa, g C m-1 year-1)a 1273.26 806.01 420.11 399.94
Emergence (Ea, g C m-2)b 41.91 6.17 13 11.06
Total flux (Eatotal, g C year-1) 7655.35 5569.05 3541.57 2067.93
136 T. A. Schriever et al.
123

P = 0.10; diversity: r (2) = 0.88, P = 0.12; evenness:
r (2) = 0.81, P = 0.20). Insect family richness showed a
strong positive correlation with hydroperiod (r (2) = 0.87,
P = 0.13) and pond area (r (2) = 0.56, P = 0.44). How-
ever, insect diversity (r (2) = -0.62, P = 0.38) and
evenness (r (2) = -0.73, P = 0.27) declined from small to
larger ponds. Insect evenness declined from shorter to
longer hydroperiod ponds (r (2) = -0.49, P = 0.51), but
diversity was only weakly related to hydroperiod
(r (2) = 0.27, P = 0.73).
Influence of pond area and hydroperiod
The subsidy flows of amphibians and insects differed in
their relationships to pond hydroperiod and area. Hydro-
period and the total amphibian flux to terrestrial habitat
were strongly negatively correlated (Eatotal: r = -0.92).
The longer the hydroperiod the lower the magnitude of
amphibian export (Fig. 6a). Pond area had very little
relation to the total amphibian flux to the terrestrial eco-
system (r = 0.21). Conversely, total insect flux (Eitotal)
showed a very strong positive relationship with pond area
(r = 0.99; Fig. 6b), however the positive pattern with hy-
droperiod was less clear (log Eitotal; r = 0.44). Ponds very
small and very large received similar amounts of litter
input. The intermediate sized pond had the highest P/A
ratio, and therefore received more litter biomass per unit
area (Online Resource Fig. 3).
Discussion
Aquatic food webs are not isolated from the surrounding
terrestrial landscape, as they are coupled through multiple
flow paths from multiple trophic levels. This is especially
true for temporary woodland ponds. We have shown that
Coe
nagr
ioni
dae
Lest
idae
Libe
llulid
ae
0
100
200
300
400
Odonata family
B C
A. l
ater
ale
A .m
acul
atum
Am
byst
oma
sp.
B. a
mer
ican
us
H. v
ersi
colo
r
L. s
ylva
ticus
N. v
iride
scen
s
P. c
ruci
fer
0
200
400
600
800
1000
1200
Amphibian speciesInsect family
0
5
10
15
20
25
30
Flu
x (g
C m
−1
yr−
1 )
Bae
tidae
Bra
coni
dae
Cae
nida
eC
erat
opog
onid
aeC
haob
orid
aeC
hiro
nom
idae
Cic
adel
lidae
Cul
icid
aeD
olic
hopo
dida
eE
phyd
ridae
Hyd
ropt
ilida
eIc
hneu
mon
idae
Lim
neph
ilida
eM
etre
topo
dida
eM
usci
dae
Mym
arid
aeP
horid
aeP
sych
odid
aeS
arco
phag
inae
Sca
thop
hagi
dae
Sce
lioni
dae
Sci
arid
aeS
ciom
yzid
aeS
imul
idae
Tab
anid
aeT
ipul
idae
A
Fig. 2 Flux (mean and standard error) attributed by each a insect
family and b Odonata family and c amphibian species collected.
Odonata are separated from other insect families because of
drastically different values compared to other taxa. No variation
exhibited in flux by some families because individuals were collected
from1 pond only
Fig. 3 Comparison of total carbon flux from pond-to-land (Etotal)
(upper portion of figure) and land-to-pond (Dtotal) (lower portion of
figure) attributed from each subsidy type across study ponds. The
inset zooms in on the contribution from insects
Spatial subsidies between pond and forest habitats 137
123
amphibians and insects contribute a considerable amount
of biomass to the terrestrial ecosystem, though litterfall
input was substantially higher than reciprocal flows to
adjacent terrestrial habitat. But these flows also depended
on the physical and biological aspects of the ponds them-
selves. We predicted that longer hydroperiods and larger
ecosystem size would have higher amphibian and insect
emergence and biomass than smaller ponds with shorter
hydroperiod. We did not find this to be the case for
amphibians. Instead, the smallest pond with the shortest
hydroperiod boasted substantially more emergent amphib-
ians and higher biomass export than the largest and longer
hydroperiod ponds. The flux of amphibians was *30 %
lower from longer hydroperiod ponds (168 days) compared
to the short hydroperiod pond (78 days). The shortest hy-
droperiod pond had very low predator richness and
abundance (both insect and salamander) and low
amphibian species richness, thereby providing an optimal
habitat for growth and survival that could account for the
high biomass. Emergent insects followed our predicted
pattern for pond area; larger ponds contributed more indi-
viduals and higher flux to the terrestrial landscape.
However, an intermediate hydroperiod pond had much
higher biomass and more insects emerging than longer
hydroperiod ponds. The ratio of perimeter to area explained
the importance of passive allochthonous input of litterfall
into woodland ponds.
It is generally accepted that amphibians and insects
account for a large proportion of energy flow within
aquatic and terrestrial food webs (Burton and Likens 1975;
Pough 1980). However, the transfer of energy through
amphibians between aquatic and terrestrial habitats has
largely been ignored (Ballinger and Lake 2006) and was
only recently quantified (Gibbons et al. 2006; Regester
Fig. 4 Variation in the magnitude of litter components received in
each pond. Litter components are: deciduous leaves, miscellaneous
(bud scales, pollen, etc.), aquatic plants (Equisetaceae; horsetail),
coniferous needles, and woody material (twigs, branches, bark, cones,
etc.). Note y-axis is not on the same scale for each pond
138 T. A. Schriever et al.
123

et al. 2006). Organisms of aquatic origin represent a spatial
subsidy to the terrestrial habitat in three ways: (1) as a prey
source for terrestrial vertebrate and insect predators; (2) as
a nitrogen and phosphorus source via excretion for soil and
plant uptake; and (3) as a release of nutrients, through
death, for decomposers, primary producers, and scaveng-
ers. Thus, spatial subsidies provided by emerging insects
and amphibians could influence recipient food webs by
directly and indirectly affecting predators, decomposers,
herbivores, and plants.
Few studies have simultaneously quantified multiple
types of resource flow from a single habitat, and here we
provide a unique picture of the energy exchange between
the aquatic and terrestrial ecosystem by quantifying the
magnitude of resource flows from obligate habitat couplers
(amphibians and emergent aquatic insects) and a passive
allochthonous subsidy (litterfall) input across a gradient of
environmental variation. Regester et al. (2006) provided
one of the first estimates of energy flow for amphibians
using ambystomatid salamanders from woodland ponds to
the surrounding forest. From one temporary and two per-
manent natural ponds, they collected 662 emerging
juvenile salamanders contributing 210 g ash-free dry mass
to the adjacent forest ecosystem (no anurans were reported
in their study). In comparison, we collected almost twice
the number of amphibians (n = 1197) emerging from four
ponds, but this represented half as much export (68 g ash-
free dry mass). In our study, cross-habitat energy flow was
dominated by anurans (64.5 %), which generally weigh
less on a per individual basis and thus contribute less
biomass than salamanders. Other studies like that of Gib-
bons et al. (2006) found anurans made up *95 % of
exported amphibian biomass from one large open canopy
pond in South Carolina, USA. Differences in amphibian
community composition can have a profound effect on the
amount of cross-habitat resource flow and striking conse-
quences for, and influences on, the terrestrial ecosystem.
This is another aspect of cross-habitat linkages that is
understudied.
In addition to hydroperiod, canopy cover also strongly
influences the performance and composition of freshwater
communities (Schiesari 2006). In general, closed canopy
ponds have lower water temperature and dissolved oxygen
(DO) than open canopy ponds (Skelly et al. 2002).
Increased canopy cover has been shown to decrease insect
abundance (Palik et al. 2001) and amphibian abundance
(Halverson et al. 2003; Binckley and Resetarits 2007) and
performance (Schiesari 2006). The ponds in our study with
higher canopy cover (Short and Intermediate 2) had lower
average water temperatures, lower salamander abundance
and total flux, and lower insect flux than open canopy
ponds (Intermediate 1 and 3; Table 2). Anurans on the
other hand showed the opposite pattern. We collected over
2.5 times more anuran metamorphs and total flux (Eatotal) in
closed canopy ponds. Anuran abundance and flux was
dominated by wood frog metamorphs. Rubbo et al. (2008)
found increasing leaf litter inputs into mesocosms
increased developmental rate and survival of larval wood
0.0 0 .5 1.0 1 .5 2.0
2000
3000
4000
5000
6000
7000
8000
Amphibian diversity (H)
Eto
tal
0 2 4 6 8 10
Amphibian species richness
0.0 0 .5 1.0 1 .5 2.0
1
10
100
1000
5000
0 5 10 15 20 25 30
Insect diversity (H) Insect richness
log
(Eto
tal)
A B
C D
Fig. 5 The relationship between a amphibian diversity, b amphibian
species richness, c insect diversity, and d insect family richness with
the magnitude of cross habitat energy flow (Etotal; g C year-1)
Fig. 6 The influence of
a hydroperiod and b pond size
on the total flux (g C year-1) of
three types of spatial subsidies.
Insect flux values were log
transformed for better
visualization of relationship
Spatial subsidies between pond and forest habitats 139
123

frogs. Our observed abundance and flux of wood frogs
from natural ponds was highest in the pond with higher
canopy cover and litter input. The few studies that have
investigated salamander performance in response to can-
opy cover have shown mixed results. In a mesocosm study,
Earl et al. (2011) found ambystomatid salamanders had
greater biomass export from high shade ponds compared to
low shade pond treatments. However, other studies have
found reduced growth and developmental rates in newts
from closed canopy ponds (Van Buskirk 2009, 2011) which
could translate to lower biomass. Our results are in contrast
to Earl et al. (2011), but follow Van Buskirk (2009, 2011)
results in that salamander abundance and flux to terrestrial
habitat was highest from more open canopy ponds (Inter-
mediate 1 and 3). Canopy cover, litter input, and pond
water chemistry influenced amphibian and insect biomass
in different ways. These results highlight the need for more
studies examining environmental influences on cross-hab-
itat energy flow.
Magnitude, rate, and functional role of spatial subsidies
can differentially impact the recipient food web (Wesner
2010) and influence the variation and strength of trophic
cascades (Leroux and Loreau 2008; Wesner 2010). Some
insects (odonates) and amphibians act as predators in the
terrestrial food web and can have a high consumptive affect
on arthropods as they disperse from the natal pond (McCoy
et al. 2009). Wesner (2010) found the majority of aquatic
insects emerging from small streams were non-consumers
as adults, thus contributing as a major prey subsidy to the
terrestrial food web. Ponds have emerging insects and
amphibians. Salamanders play vital roles in forest-floor
detrital food webs and nutrient cycling (Davic and Welsh
2004 and citations within) while anurans consume a wide
variety of arthropods on the forest floor (e.g. wood frog)
and in tree canopies (e.g. grey treefrog). The most abun-
dant emerging insect in our study was chironomid midges,
which are non-consumers as adults and therefore act as a
prey subsidy to terrestrial food webs. However, emergent
aquatic insects made up only a small proportion (on aver-
age 39 % compared to 89 % for amphibians) of the total
pond to land flux. This illustrates the trophic structure of
the flux is dominated by consumer biomass. Intermittent
ponds, therefore, have a dual role as providers of prey
subsidies to terrestrial predators and a seasonal contributor
of intermediate predators to terrestrial food webs, making a
case that these aquatic habitats form multiple connections
within discrete food webs.
Studies on the flows of invertebrate subsidies to adjacent
terrestrial habitats have been strongly stream focused
(Jackson and Fisher 1986; Nakano and Murakami 2001;
Sabo and Power 2002; Kato et al. 2003; Baxter et al. 2005;
Ballinger and Lake 2006; Whiting et al. 2011) and some-
times limited to one or two insect families (Fisher and
Likens 1973; Alvarez and Pardo 2005; Runck 2007). There
are surprisingly few studies that have measured the entire
insect assemblage production from wetlands. We collected
individuals representing 36 insect families. Insect flux was
two times higher in our study ponds compared to that found
by Whiles and Goldowitz (2001) from open canopy wet-
lands with and without fish in Nebraska. Gratton and
Vander Zanden (2009) estimated insect emergence rates
across a range of ecosystem size and habitat types. Of the
three ponds used in Gratton and Vander Zanden (2009), the
largest pond (800 m2) had comparable emergence pro-
duction rates to a similar size pond (987.48 m2) in our
study (our empirical value 3.32 vs. theirs 3.29 g C m-2
year-1). Our estimates of emergence numbers and magni-
tude of flux are likely conservative given that insect
trapping started in May and ice off was in March, odonates
are not trapped with the same efficiency as other aquatic
insects, and treefrogs can climb drift fences avoiding
capture (Gibbons et al. 2006); therefore we are most likely
underestimating flux.
Terrestrial primary production in the form of leaf litter is
a significant resource in woodland ponds forming the base
of the aquatic food web (Rubbo et al. 2006; Batzer and
Palik 2007). Litterfall is a donor controlled seasonally
recurring resource subsidy to woodland ponds. We pre-
dicted the amount of litter deposition would be
proportional to the amount of animal export. However, the
highest input of litter was deposited in a pond with high
P:A and resulted in fairly low export via amphibian and
insect emergence. We measured almost 8.5 times more
litter input (Dtotal = 187,371.8 g C year-1) than the com-
bined total export flux of amphibian and insect resources
(Etotal = 22,252.75 g C year-1) from four ponds, resulting
in an overall net carbon input. These results are consistent
with Vander Zanden and Gratton’s (2011) modeled results
of lake-to-land flux (Dtotal) that exceed insect emergence
(Etotal) in small lakes (\2,000 ha). This net carbon input
may be common to most lakes and streams (Jackson and
Fisher 1986; Leroux and Loreau 2008).
A positive diversity and ecosystem function response is
typically found (Balvanera et al. 2006), especially for pri-
mary producers from terrestrial ecosystems (Schlapfer and
Schmid 1999; Bouchard et al. 2007), but more complex
responses are possible when studying multiple trophic
levels of consumer groups (Duffy 2002; Hooper et al.
2005). We found that communities with higher insect or
amphibian diversity result in lower flux to the terrestrial
ecosystem (Etotal and emergent flux; Fig. 5). We studied
four ponds, and therefore are limited in our ability to
generalize our results to pond ecosystems as a whole. That
said, we found an interesting pattern exhibited by two
different taxonomic groups, which indicates further
research on a multiple consumer groups and pond systems
140 T. A. Schriever et al.
123

is warranted. The amphibian export flux was dominated by
one species, wood frog, from one small, intermittent
woodland pond with low insect family and amphibian
species richness. This result is contrary to the sampling
effect model that states there is a higher chance of sam-
pling a productive species in a species rich assemblage
(Tilman et al. 1997; Aarssen 1997), but is consistent with
the dominance effect (Norberg 2004). It is important to
note that we looked across multiple trophic levels, whereby
most experiments look at single trophic levels. Wood frogs
had higher biomass than the cumulative biomass of a more
diverse amphibian community in an intermediate hydro-
period pond; perhaps because its development is best suited
to temporary ponds (Paton and Crouch 2002). We specu-
late that predation, presence of salamander larvae, and
insect predators, as well as competition from other larval
anurans decreased wood frog productivity in other ponds.
Mechanisms controlling community structure change along
the hydroperiod gradient (i.e., abiotic forces in short hy-
droperiod ponds versus strong biotic factors in longer
hydroperiod ponds) and thus can have strong and possibly
different, influences on the biodiversity–ecosystem func-
tion relationship. Further, perhaps the positive relationship
between biodiversity and ecosystem function may not be a
general pattern for aquatic vertebrates (Cardinale et al.
2006).
Conclusions
Intermittent ponds had a tight coupling with the terrestrial
landscape in two reciprocal ways: ponds are heterotrophic
depending on allochthonous carbon inputs to sustain the
aquatic food web and aquatic emergences of amphibians
and insects become part of the terrestrial food web as
consumers (e.g., amphibian juveniles) and as prey subsidy
(e.g. chironomids) to terrestrial organisms. Cross-habitat
resource flows were variable in time, space, and taxonomic
group. Environmental variation in hydroperiod had a strong
influence on amphibian flux to the adjacent terrestrial
habitat, but less influential to insect flux. Diversity was
higher in ponds with lower productivity, thus providing a
‘non-experimental’ view of the BEF relationship. The
magnitude of spatial subsidies in this system is a complex
product of interactions among pond hydroperiod, size,
perimeter-to-area relationships, and species diversity.
Therefore, landscapes comprised of ponds varying in hy-
droperiod are essential to conserving insect and amphibian
diversity and to maximize aquatic-terrestrial linkages.
Acknowledgments Construction of the drift fences and setting
pitfall buckets could not have been completed without the help of:
Katherine Bannar-Martin, Devin Bloom, Kristen Brochu, Kirsten
Comberford, Maria Modanu, Stephen Pynn, David Stitt, and Caroline
Tucker. We also thank Mark Conboy, Klara Jaspers-Fayer, Tristan
Willis and Monica Candelaria for helping with field work. The tre-
mendous dedication of Siao Ryan Yang and Ruby Sambi in
processing samples is greatly appreciated. We thank Karen Pope for
generously donating the emergence traps and Nathan Lovejoy for
microscope and laboratory use. This research was supported by a
Natural Sciences and Engineering Research Council of Canada Dis-
covery Grant awarded to DDW.
References
Aarssen LW (1997) High productivity in grassland ecosystems:
effected by species diversity or productive species? Oikos
80(1):183–184
Alvarez M, Pardo I (2005) Life history and production of Agapetus
quadratus (Trichoptera: Glossosomatidae) in a temporary,
spring-fed stream. Freshw Biol 50:930–943. doi:10.1111/j.
1365-2427.2005.01370.x
Ballinger A, Lake PS (2006) Energy and nutrient fluxes from rivers
and streams into terrestrial food webs. Mar Freshw Res 57:15–28
Balvanera P, Pfisterer AB, Buchmann N, He J-S, Nakashizuka T,
Raffaelli D, Schmid B (2006) Quantifying the evidence for
biodiversity effects on ecosystem functioning and services. Ecol
Lett 9:1146–1156
Batzer DP, Palik BJ (2007) Variable response by aquatic invertebrates
to experimental manipulations of leaf litter input into seasonal
woodland ponds. Fund Appl Limnol 168(2):155–162
Baxter CV, Fausch KD, Saunders WC (2005) Tangled webs:
reciprocal flows of invertebrate prey link streams and riparian
zones. Freshw Biol 50:201–220. doi:10.1111/j.1365-2427.2004.
01328.x
Benke AC (1993) Concepts and patterns of invertebrate production in
running waters. In: Congress of the International Assoc. of
Theoretical and Applied Limnology, Barcelona, Spain, 1993.
International association of theoretical and applied limnology
proceedings. pp 15–38
Binckley CA, Resetarits WJ Jr (2007) Effects of forest canopy on
habitat selection in treefrogs and aquatic insects: implications for
communities and meta-communities. Oecologia 153:951–958
Bouchard V, Frey SD, Gilbert JM, Reed SE (2007) Effects of
macrophyte functional group richness on emergent freshwater
wetland functions. Ecology 88(11):2903–2914
Burton TM, Likens GE (1975) Salamander populations and biomass
in Hubbard Brook experimental forest, New Hampshire. Copeia
3:541–546
Cadenasso ML, Weathers KC, Pickett STA (2004) Integrating food
web and landscape ecology: subsidies at the regional scale. In:
Polis GA, Power ME, Huxel GR (eds) Food webs at the
landscape level. The University of Chicago Press, Chicago,
pp 263–267
Cardinale BJ, Srivastava DS, Duffy JE, Wright JP, Downing AL,
Sankaran M, Jouseau C (2006) Effects of biodiversity on the
functioning of trophic groups and ecosystems. Nature
443(7114):989–992
Davic RD, Welsh HH (2004) On the ecological roles of salamanders.
Ann Rev Ecol Evol Sys 35:405–434
Dreyer J, Hoekman D, Gratton C (2012) Lake-derived midges
increase abundance of shoreline terrestrial arthropods via
multiple trophic pathways. Oikos 121:252–258
Duffy JE (2002) Biodiversity and ecosystem function: the consumer
connection. Oikos 99(2):201–219
Spatial subsidies between pond and forest habitats 141
123

Earl JE, Luhring TM, Williams BK, Semlitsch RD (2011) Biomass
export of salamanders and anurans from ponds is affected
differentially by changes in canopy cover. Freshw Biol
56:2473–2482
Edwards FK, Lauridsen RB, Armand L, Vincent HM, Jones JI (2009)
The relationship between length, mass and preservation time for
three species of freshwater leeches (Hirudinea). Fund Appl
Limnol 173(4):321–327
Fisher SG, Likens GE (1973) Energy flow in Bear Brook, New
Hampshire: integrative approach to stream ecosystem metabo-
lism. Ecol Monogr 43(4):421–439
Gibbons JW, Bennett DH (1974) Determination of anuran terrestrial
activity patterns by a drift fence method. Copeia 1:236–243
Gibbons JW, Winne CT, Scott DE, Willson JD, Glaudas X, Andrews
KM, Todd BD, Fedewa LA, Wilkinson L, Tsaliagos RN, Harper
SJ, Greene JL, Tuberville TD, Metts BS, Dorcast ME, Nestor JP,
Young CA, Akre T, Reed RN, Buhlmann KA, Norman J,
Croshaw DA, Hagen C, Rothermel BB (2006) Remarkable
amphibian biomass and abundance in an isolated wetland:
implications for wetland conservation. Con Biol
20(5):1457–1465
Gratton C, Vander Zanden MJ (2009) Flux of aquatic insect
productivity to land: comparison of lentic and lotic ecosystems.
Ecology 90(10):2689–2699
Gratton C, Donaldson J, Vander Zanden MJ (2008) Ecosystem
linkages between lakes and the surrounding terrestrial landscape
in Northeast Iceland. Ecosystems 11:764–774
Halverson MA, Skelly DK, Kiesecker JM, Freidenburg LK (2003)
Forest mediated light regime linked to amphibian distribution
and performance. Oecologia 134:360–364
Hoekman D, Dreyer J, Jackson RD, Townsend PA, Gratton C (2011)
Lake to land subsidies: experimental addition of aquatic insects
increases terrestrial arthropod densities. Ecology 92:2063–2072
Hooper DU, Chapin FS, Ewel JJ, Hector A, Inchausti P, Lavorel S,
Lawton JH, Lodge DM, Loreau M, Naeem S, Schmid B, Setala
H, Symstad AJ, Vandermeer J, Wardle DA (2005) Effects of
biodiversity on ecosystem functioning: a consensus of current
knowledge. Ecol Monogr 75(1):3–35
Hutchens JJ, Wallace JB (2002) Ecosystem linkages between
southern Appalachian headwater streams and their banks: leaf
litter breakdown and invertebrate assemblages. Ecosystems
5(1):80–91
Jackson JK, Fisher SG (1986) Secondary production, emergence, and
export of aquatic insects of a Sonoran desert stream. Ecology
67(3):629–638
Kato Y, Hori M, Okuda N, Tayasu I, Takemon Y (2003) Spatial
heterogeneity of trophic pathways in the invertebrate community
of a temperate bog. Freshw Biol 55:450–462
Kitchell JF, Schindler DE, Herwig BR, Post DM, Olson MH, Oldham
M (1999) Nutrient cycling at the landscape scale: the role of diel
foraging migrations by geese at the Bosque del Apache National
wildlife refuge, New Mexico. Limnol Oceanogr 44(3):828–836
Leroux SJ, Loreau M (2008) Subsidy hypothesis and strength of
trophic cascades across ecosystems. Ecol Lett 11(11):1147–1156
Leuven RSEW, Brock TCM, van Druten HAM (1985) Effects of
preservation on dry- and ash-free dry weight biomass of some
common aquatic macro-invertebrates. Hydrobiologia 127:
151–159
Loreau M, Naeem S, Inchausti P, Bengtsson J, Grime JP, Hector A,
Hooper DU, Huston MA, Raffaelli D, Schmid B, Tilman D,
Wardle DA (2001) Ecology–Biodiversity and ecosystem func-
tioning: current knowledge and future challenges. Science
294(5543):804–808
Loreau M, Mouquet N, Holt RD (2003) Meta-ecosystems: a
theoretical framework for a spatial ecosystem ecology. Ecol
Lett 6(8):673–679
Massol F, Gravel D, Mouquet N, Cadotte MW, Fukami T, Leibold
MA (2011) Linking community and ecosystem dynamics
through spatial ecology. Ecol Lett 14:313–323
McCoy MW, Barfield M, Holt RD (2009) Predator shadows: complex
life histories as generators of spatially patterned indirect
interactions across ecosystems. Oikos 118(1):87–100. doi:10.
1111/j.1600-0706.2008.16878.x
Nakano S, Murakami M (2001) Reciprocal subsidies: dynamic
interdependence between terrestrial and aquatic food webs.
Proc Natl Acad Sci USA 98(1):166–170. doi:10.1073/pnas.98.
1.166
Norberg J (2004) Biodiversity and ecosystem functioning: a complex
adaptive systems approach. Limnol Oceanogr 49(4):1269–1277
Nowlin WH, Vanni MJ, Yang LH (2008) Comparing resource pulses
in aquatic and terrestrial ecosystems. Ecology 89(3):647–659
Palik B, Kastendick D (2010) Response of seasonal pond plant
communities to upland forest harvest in northern Minnesota
forests, USA. For Ecol Man 260:628–637
Palik B, Batzer D, Buech R, Nichols D, Cease K, Egeland L, Streblow
D (2001) Seasonal pond characteristics across a chronosequence
of adjacent forest ages in northern Minnesota. Wetlands
21:532–542
Palik B, Batzer D, Kern C (2006) Upland forest linkages to seasonal
wetlands: litter flux, processing, and food quality. Ecosystems
9(1):142–151. doi:10.1007/s10021-005-0010-0
Paton PWC, Crouch WB (2002) Using the phenology of pond-
breeding amphibians to develop conservation strategies. Con
Biol 16(1):194–204. doi:10.1046/j.1523-1739.2002.00260.x
Polis GA, Anderson WB, Holt RD (1997) Toward an integration of
landscape and food web ecology: the dynamics of spatially
subsidized food webs. Ann Rev Ecol Sys 28:289–316
Post DM, Taylor JP, Kitchell JF, Olson MH, Schindler DE, Herwig
BR (1998) The role of migratory waterfowl as nutrient vectors in
a managed wetland. Con Biol 12(4):910–920. doi:10.1046/j.
1523-1739.1998.97112.x
Pough FH (1980) Advantages of ectothermy for tetrapods. Am Nat
115(1):92–112
Pray CL, Nowlin WH, Vanni MJ (2009) Deposition and decompo-
sition of periodical cicadas (Homoptera: Cicadidae: Magicicada)
in woodland aquatic ecosystems. J N Am Benthol Soc
28(1):181–195. doi:10.1899/08-038.1
R Development Core Team (2009) R: a language and environment for
statistical computing. R Foundation for Statistical Computing,
Vienna
Regester KJ, Lips KR, Whiles MR (2006) Energy flow and subsidies
associated with the complex life cycle of ambystomatid
salamanders in ponds and adjacent forest in southern Illinois.
Oecologia 147(2):303–314. doi:10.1007/s00442-005-0266-2
Romanuk TN, Vogt RJ, Young A, Tuck C, Carscallen MW (2010)Maintenance of positive diversity–stability relations along a
gradient of environmental stress. PLoS One 5(4):1–9
Rubbo MJ, Kiesecker JM (2004) Leaf litter composition and
community structure: translating regional species changes into
local dynamics. Ecology 85(9):2519–2525
Rubbo MJ, Cole JJ, Kiesecker JM (2006) Terrestrial subsidies of
organic carbon support net ecosystem production in temporary
forest ponds: evidence from an ecosystem experiment. Ecosys-
tems 9(7):1170–1176. doi:10.1007/s10021-005-0009-6
Rubbo MJ, Belden LK, Kiesecker JM (2008) Differential responses of
aquatic consumers to variations in leaf-litter inputs. Hydrobio-
logia 605:37–44
Runck C (2007) Macro-invertebrate production and food web
energetics in an industrially contaminated stream. Ecol Appl
17(3):740–753. doi:10.1890/05-1026
Sabo JL, Post DM (2008) Quantifying periodic, stochastic, and
catastrophic environmental variation. Ecol Monogr 78(1):19–40
142 T. A. Schriever et al.
123

Sabo JL, Power ME (2002) Numerical response of lizards to aquatic
insects and short-term consequences for terrestrial prey. Ecology
83:3023–3036
Schiesari L (2006) Pond canopy cover: a resource gradient for anuran
larvae. Freshw Biol 51:412–423
Schlapfer F, Schmid B (1999) Ecosystem effects of biodiversity: a
classification of hypotheses and exploration of empirical results.
Ecol Appl 9(3):893–912. doi:10.2307/2641337
Schreiber S, Rudolf VHW (2008) Crossing habitat boundaries:
coupling dynamics of ecosystems through complex life cycles.
Ecol Lett 11(6):576–587. doi:10.1111/j.1461-0248.2008.01171.x
Skelly DK, Freidenburg LK, Kiesecker JM (2002) Forest canopy and
the performance of larval amphibians. Ecology 83(4):983–992
Stagliano DM, Benke AC, Anderson DH (1998) Emergence of
aquatic insects from two habitats in a small wetland of the
southeastern USA: temporal patterns of numbers and biomass.
J N Am Benthol Soc 17(1):37–53
Tilman D, Lehman CL, Thomson KT (1997) Plant diversity and
ecosystem productivity: theoretical considerations. Proc Natl
Acad Sci USA 94(5):1857–1861
Van Buskirk J (2009) Natural variation in morphology of larval
amphibians: phenotypic plasticity in nature? Ecol Monogr
79:681–705
Van Buskirk J (2011) Amphibian phenotypic variation along a
gradient in canopy cover: species differences and plasticity.
Oikos 120:906–914
Vander Zanden MJ, Gratton C (2011) Blowin’ in the wind: reciprocal
airborne carbon fluxes between lakes and land. Can J Fish Aquat
Sci 68:170–182. doi:10.1139/F10-157
von Schiller D, Solimini AG (2005) Differential effects of preserva-
tion on the estimation of biomass of two common mayfly
species. Arch Hydrobiol 164(3):325–334
Welborn GA, Skelly DK, Werner EE (1996) Mechanisms creating
community structure across a freshwater habitat gradient. Ann
Rev Ecol Sys 27:337–363
Wesner JS (2010) Seasonal variation in the trophic structure of a
spatial prey subsidy linking aquatic and terrestrial food webs:
adult aquatic insects. Oikos 119(1):170–178
Wesner JS (2012) Emerging aquatic insects as predators in terrestrial
systems across a gradient of stream temperature in North and
South America. Freshw Biol 57:2465–2474
Whiles MR, Goldowitz BS (2001) Hydrologic influences on insect
emergence production from central Platte River wetlands. Ecol
Appl 11(6):1829–1842
Whiting DP, Whiles MR, Stone ML (2011) Patterns of macro-
invertebrate production, trophic structure, and energy flow along
a tallgrass prairie stream continuum. Limnol Oceanogr
56(3):887–898. doi:10.4319/lo.2011.56.3.0887
Williams DD (2006) The biology of temporary waters. Oxford
University Press, Oxford
Spatial subsidies between pond and forest habitats 143
123