h. floresiensis - kaifu 2011
TRANSCRIPT

at SciVerse ScienceDirect
Journal of Human Evolution 61 (2011) 644e682
Contents lists available
Journal of Human Evolution
journal homepage: www.elsevier .com/locate/ jhevol
Craniofacial morphology of Homo floresiensis: Description, taxonomic affinities,and evolutionary implication
Yousuke Kaifu a,b,*, Hisao Baba a, Thomas Sutikna c, Michael J. Morwood d, Daisuke Kubo b,E. Wahyu Saptomo c, Jatmiko c, Rokhus Due Awe c, Tony Djubiantono c
aDepartment of Anthropology, National Museum of Nature and Science, 4-1-1 Amakubo, Tsukuba-shi, Ibaraki Prefecture JapanbDepartment of Biological Sciences, The University of Tokyo, 3-1-1 Hongo, Bunkyo-ku, Tokyo 113-0033, JapancNational Research and Development Centre for Archaeology, Jl. Raya Condet Pejaten No 4, Jakarta 12001, IndonesiadCentre for Archaeological Science, School of Earth and Environmental Sciences, University of Wollongong, Wollongong, NSW 2522, Australia
a r t i c l e i n f o
Article history:Received 5 October 2010Accepted 21 August 2011
Keywords:LB1/1Homo erectusHomo habilisCraniumFace
* Corresponding author.E-mail address: [email protected] (Y. Kaifu).
0047-2484/$ e see front matter � 2011 Elsevier Ltd.doi:10.1016/j.jhevol.2011.08.008
a b s t r a c t
This paper describes in detail the external morphology of LB1/1, the nearly complete and only knowncranium of Homo floresiensis. Comparisons were made with a large sample of early groups of the genusHomo to assess primitive, derived, and unique craniofacial traits of LB1 and discuss its evolution. Prin-cipal cranial shape differences between H. floresiensis and Homo sapiens are also explored metrically.
The LB1 specimen exhibits a marked reductive trend in its facial skeleton, which is comparable to theH. sapiens condition and is probably associated with reduced masticatory stresses. However, LB1 iscraniometrically different from H. sapiens showing an extremely small overall cranial size, and thecombination of a primitive low and anteriorly narrow vault shape, a relatively prognathic face, a roundedoval foramen that is greatly separated anteriorly from the carotid canal/jugular foramen, and a unique,tall orbital shape. Whereas the neurocranium of LB1 is as small as that of some Homo habilis specimens, itexhibits laterally expanded parietals, a weak suprameatal crest, a moderately flexed occipital, a markedfacial reduction, and many other derived features that characterize post-habilis Homo. Other craniofacialcharacteristics of LB1 include, for example, a relatively narrow frontal squama with flattened right andleft sides, a marked frontal keel, posteriorly divergent temporal lines, a posteriorly flexed anteromedialcorner of the mandibular fossa, a bulbous lateral end of the supraorbital torus, and a forward protrudingmaxillary body with a distinct infraorbital sulcus. LB1 is most similar to early Javanese Homo erectus fromSangiran and Trinil in these and other aspects. We conclude that the craniofacial morphology of LB1 isconsistent with the hypothesis that H. floresiensis evolved from early Javanese H. erectus with dramaticisland dwarfism. However, further field discoveries of early hominin skeletal remains from Flores anddetailed analyses of the finds are needed to understand the evolutionary history of this endemic homininspecies.
� 2011 Elsevier Ltd. All rights reserved.
Introduction
Homo floresiensis is a small-bodied, hominin species that livedon the Indonesian island of Flores in the late Pleistocene. Skeletalremains of this species are currently only known from Liang Bua,a limestone cave, where they are dated to between 74 and 17 kyr. Atleast 14 individuals are represented by these remains, whichinclude LB1, an almost complete skeleton and the species holotype,popularly known as ‘Hobbit’ (Brown et al., 2004; Morwood andJungers, 2009; Morwood et al., 2009; Roberts et al., 2009;
All rights reserved.
Westaway et al., 2009). The unusual combination of extremelysmall brain size, short stature, and other unique physical traits ofH. floresiensis have led some to argue that the skeletal remainsrepresent a population of pathological modern humans. However,such proponents have been unable to indicate a specific syndromethat fully explains these traits, and there is now growing supportfor the hypothesis that H. floresiensiswas a late-surviving species ofpre-modern Homo (reviewed in Aiello, 2010).
The origins of this novel species still remain highly controversialdespite lively debate and further studies following the initialreports (Brown et al., 2004; Morwood et al., 2004, 2005). In fact,nominated candidates for ancestral species of H. floresiensis includeJavanese Homo erectus and pre-erectus grade hominins such as

Y. Kaifu et al. / Journal of Human Evolution 61 (2011) 644e682 645
Homo habilis or even Australopithecus (e.g., Brown et al., 2004;Argue et al., 2009; Brown and Maeda, 2009; Lyras et al., 2009;Morwood and Jungers, 2009). All these possibilities have majorimplications for our understanding of the evolution of genus Homo.If H. floresiensis evolved from a habiline-like ancestor on Flores,then H. erectus sensu lato (H. erectus s. l.) was not the first homininspecies to disperse into Eurasia, as assumed in the current Out ofAfrica 1 hypothesis (Morwood and Jungers, 2009). It would alsoimply that H. erectus and another more primitive form of Homocoexisted in Southeast Asia for a substantial period. Alternatively, ifH. floresiensis originated from Asian H. erectus, then insulardwarfing to an unparalleled degree has been a significant factor inearly hominin evolution on Flores (Brown et al., 2004).
Skeletal evidence of the first hominins to colonize Flores wouldprovide direct and conclusive evidence for the evolutionary historyof H. floresiensis, but further study of the Liang Bua homininremains is also essential. In this paper, we provide a detailed
Figure 1. Facial, posterior, right lateral, left lateral, superior, and basal view
description of the external cranial morphology of LB1, and assess itsmorphological affinities.
Background and the scope of this study
The LB1 skeleton is that of an adult individual whose sex ispresumed to be female on the basis of pelvic morphology (Brownet al., 2004; Jungers et al., 2009b). The cranium is almostcomplete (Reference number LB1/1; Figs. 1 and 2) and is the onlyexample of a H. floresiensis cranium yet recovered (Morwood andJungers, 2009). In this section, we review the published studieson its morphological affinities.
In the original reports of H. floresiensis, Brown et al. (2004)and Morwood et al. (2005) described “a mosaic of primitive,unique and derived features not recorded for any other hominin”in the cranium and other skeletal parts of LB1. For instance, theyfound that the endocranial capacity is small and comparable to
s of LB1/1 oriented based on the Frankfurt Horizontal. Scale ¼ 5 cm.

Y. Kaifu et al. / Journal of Human Evolution 61 (2011) 644e682646
Australopithecus; the face was said to be Homo-like, lacking a seriesof characteristic morphologies of Australopithecus such as a greatfacial height, marked prognathism, large tooth crown size, and ananteriorly oriented infraorbital region; the cranial vault is similar tothose of H. erectus s. l. in height-breadth relationships, bonethicknesses, and some basicranial traits; and the frontal resemblesthose of early African and Dmanisi Homo in exhibiting a strongmidsagittal curvature. Furthermore, principal component analysis(PCA) based on 5 cranial vault measurements (Howells’ GOL, WFB,XCB [SMCB of us], ASB, and VRR: see Table 1 and Howells, 1973) alsoshowed that the vault shape of LB1/1 is, among extant and variousfossil hominin crania, most similar to KNM-ER 3733, KNM-ER 3883,Sangiran 2 and another unspecified Indonesian H. erectus (Brownet al., 2004: SOM Fig. 1).
On the basis of the location and age of the find, as well as somemorphological traits, it was initially suggested that H. floresiensis
Figure 2. Surface rendered CT images of LB1/1. T
most probably evolved from an ancestral H. erectus population onFlores as a result of long-term isolation and insular dwarfing(Brown et al., 2004). With the recovery of additional H. floresiensispostcranial remains, however, the Liang Bua research team wereless certain about the genealogy of H. floresiensis e noting thespecies “is not just an allometrically scaled-down version ofH. erectus” (Morwood et al., 2005: 1016).
Subsequently, two studies employed multivariate analyses oflinear cranial measurements to further investigate LB1’s morpho-logical affinities. Argue et al. (2006) conducted canonical variateanalyses (CVA). Their Analysis 3 is based on 5 cranial vaultmeasurements (Howells’ GOL, XCB, BBH, AUB, ASB: data of LB1cited from Brown et al., 2004) and includes a recent modern humansample (Howells’ data), as well as a small sample of australopith-ecine and early Homo specimens (Sts 5; OH 24; KNM-ER 406, 1813,3733, 3883; D2280; Sangiran 17; five NgandongH. erectus). Another
he orientations and scale same as in Fig. 1.
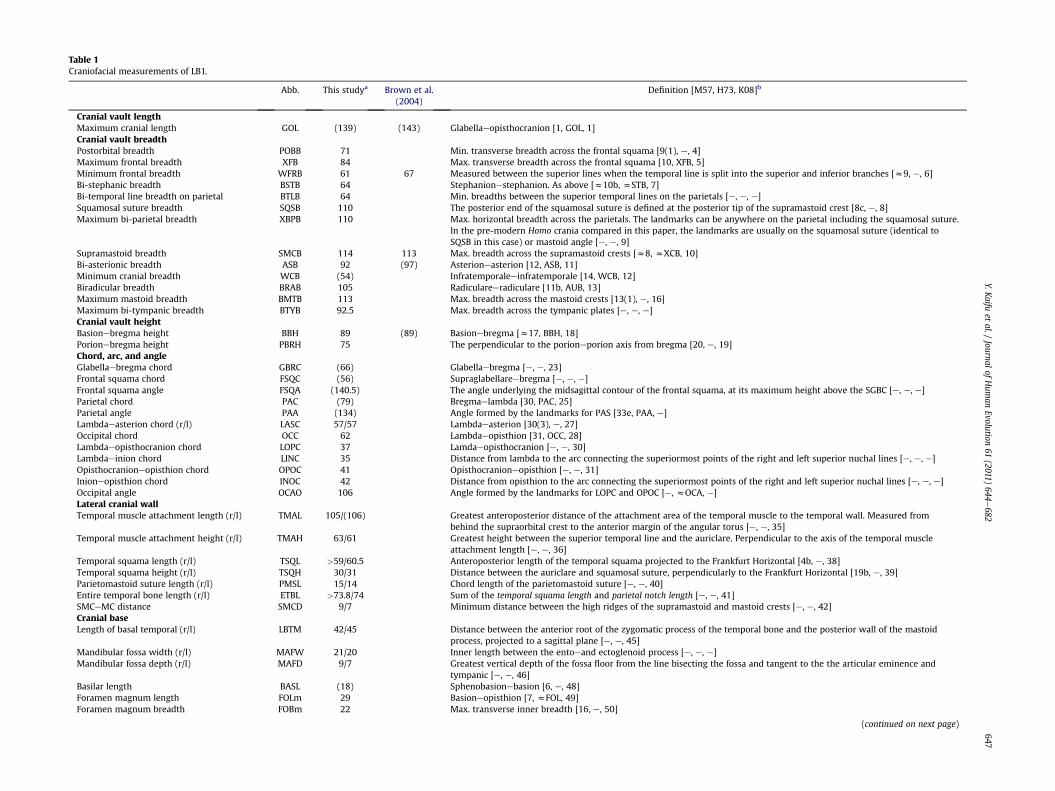
Table 1Craniofacial measurements of LB1.
Abb. This studya Brown et al.(2004)
Definition [M57, H73, K08]b
Cranial vault lengthMaximum cranial length GOL (139) (143) Glabellaeopisthocranion [1, GOL, 1]Cranial vault breadthPostorbital breadth POBB 71 Min. transverse breadth across the frontal squama [9(1), e, 4]Maximum frontal breadth XFB 84 Max. transverse breadth across the frontal squama [10, XFB, 5]Minimum frontal breadth WFRB 61 67 Measured between the superior lines when the temporal line is split into the superior and inferior branches [z9, e, 6]Bi-stephanic breadth BSTB 64 Stephanionestephanion. As above [z10b, zSTB, 7]Bi-temporal line breadth on parietal BTLB 64 Min. breadths between the superior temporal lines on the parietals [e, e, e]Squamosal suture breadth SQSB 110 The posterior end of the squamosal suture is defined at the posterior tip of the supramastoid crest [8c, e, 8]Maximum bi-parietal breadth XBPB 110 Max. horizontal breadth across the parietals. The landmarks can be anywhere on the parietal including the squamosal suture.
In the pre-modern Homo crania compared in this paper, the landmarks are usually on the squamosal suture (identical toSQSB in this case) or mastoid angle [e, e, 9]
Supramastoid breadth SMCB 114 113 Max. breadth across the supramastoid crests [z8, zXCB, 10]Bi-asterionic breadth ASB 92 (97) Asterioneasterion [12, ASB, 11]Minimum cranial breadth WCB (54) Infratemporaleeinfratemporale [14, WCB, 12]Biradicular breadth BRAB 105 Radiculareeradiculare [11b, AUB, 13]Maximum mastoid breadth BMTB 113 Max. breadth across the mastoid crests [13(1), e, 16]Maximum bi-tympanic breadth BTYB 92.5 Max. breadth across the tympanic plates [e, e, e]Cranial vault heightBasionebregma height BBH 89 (89) Basionebregma [z17, BBH, 18]Porionebregma height PBRH 75 The perpendicular to the porioneporion axis from bregma [20, e, 19]Chord, arc, and angleGlabellaebregma chord GBRC (66) Glabellaebregma [e, e, 23]Frontal squama chord FSQC (56) Supraglabellareebregma [e, e, e]Frontal squama angle FSQA (140.5) The angle underlying the midsagittal contour of the frontal squama, at its maximum height above the SGBC [e, e, e]Parietal chord PAC (79) Bregmaelambda [30, PAC, 25]Parietal angle PAA (134) Angle formed by the landmarks for PAS [33e, PAA, e]Lambdaeasterion chord (r/l) LASC 57/57 Lambdaeasterion [30(3), e, 27]Occipital chord OCC 62 Lambdaeopisthion [31, OCC, 28]Lambdaeopisthocranion chord LOPC 37 Lamdaeopisthocranion [e, e, 30]Lambdaeinion chord LINC 35 Distance from lambda to the arc connecting the superiormost points of the right and left superior nuchal lines [e, e, e]Opisthocranioneopisthion chord OPOC 41 Opisthocranioneopisthion [e, e, 31]Inioneopisthion chord INOC 42 Distance from opisthion to the arc connecting the superiormost points of the right and left superior nuchal lines [e, e, e]Occipital angle OCAO 106 Angle formed by the landmarks for LOPC and OPOC [e, zOCA, e]Lateral cranial wallTemporal muscle attachment length (r/l) TMAL 105/(106) Greatest anteroposterior distance of the attachment area of the temporal muscle to the temporal wall. Measured from
behind the supraorbital crest to the anterior margin of the angular torus [e, e, 35]Temporal muscle attachment height (r/l) TMAH 63/61 Greatest height between the superior temporal line and the auriclare. Perpendicular to the axis of the temporal muscle
attachment length [e, e, 36]Temporal squama length (r/l) TSQL >59/60.5 Anteroposterior length of the temporal squama projected to the Frankfurt Horizontal [4b, e, 38]Temporal squama height (r/l) TSQH 30/31 Distance between the auriclare and squamosal suture, perpendicularly to the Frankfurt Horizontal [19b, e, 39]Parietomastoid suture length (r/l) PMSL 15/14 Chord length of the parietomastoid suture [e, e, 40]Entire temporal bone length (r/l) ETBL >73.8/74 Sum of the temporal squama length and parietal notch length [e, e, 41]SMCeMC distance SMCD 9/7 Minimum distance between the high ridges of the supramastoid and mastoid crests [e, e, 42]Cranial baseLength of basal temporal (r/l) LBTM 42/45 Distance between the anterior root of the zygomatic process of the temporal bone and the posterior wall of the mastoid
process, projected to a sagittal plane [e, e, 45]Mandibular fossa width (r/l) MAFW 21/20 Inner length between the entoeand ectoglenoid process [e, e, e]Mandibular fossa depth (r/l) MAFD 9/7 Greatest vertical depth of the fossa floor from the line bisecting the fossa and tangent to the the articular eminence and
tympanic [e, e, 46]Basilar length BASL (18) Sphenobasionebasion [6, e, 48]Foramen magnum length FOLm 29 Basioneopisthion [7, zFOL, 49]Foramen magnum breadth FOBm 22 Max. transverse inner breadth [16, e, 50]
(continued on next page)
Y.Kaifuet
al./Journal
ofHum
anEvolution
61(2011)
644e682
647

Table 1 (continued )
Abb. This studya Brown et al.(2004)
Definition [M57, H73, K08]b
Oval foramen diameter 1 (r/l) OFD1 e/4.2 Inner diameter of the oval foramen measured from its anteromedial to posterolateral corners, parallel to the anterolateralmargin of the petrous bone [e, e, e]
Oval foramen diameter 2 (r/l) OFD2 e/4.6 Max. inner diameter of the oval foramen measured perpendicular to the OFD1 [e, e, e]Oval foramenecarotid canal distance (r/l) OFeCC e/18 Min. inner distance between the oval foramen and carotid canal projected to sagittal plane [e, e, e]Oval foramenejugular foramen distance (r/l) OFeJF e/31 Max. outer distance between the oval foramen and carotid canal projected to sagittal plane [e, e, e]Bi-oval foramen breadth OFeOF 45 Max. outer breadth across the oval foramina [e, e, e]Bi-carotid canal breadth CCeCC 50 Max. outer breadth across the carotid canals [e, e, e]Bi-jugular foramen breadth JFeJF 52 Max. outer breadth across the jugular foramina [e, e, e]Cranial bone thicknessFrontal eminence thickness (r/l) CTFE 7/7c Measured perpendicularly to the external cranial surface [e, e, e]Bregma thickness CTBR e (7.6) As above [e, e, e]Parietal eminence thickness (r/l) CTPE 8/7c 8.5 As above [e, e, e]Lambda thickness CTLA 6c,d 6.3 As above [e, e, e]Asterion thickness (r/l) CTAS (8)/(8)c 11 As above [e, e, e]Opisthocranion thickness CTOP (15)c 16.4 As above [e, e, e]Facial lengthBasionenasion length BNL (78) (81) Basionenasion [5, BNL, 2]Basioneprosthion length BPL (85) (88) Basioneprosthion [z40, BPL, e]Porioneprosthion radius PPRR 91 The perpendicular to the porioneporion axis from prosthion [e, e, e]Facial heightSuperior facial height NPHm (55) (53) Nasionealveolare [48, e, e]Superior facial height NPH (54) Nasioneprosthion [z48, NPH, e]Infraorbital maxillary height (r/l) IOMH 29/29 Min. distance between the inferior orbital margin and the alveolar margin between the M1 and M2 [48(3)’, e, e]Facial breadthSupraorbital torus breadth SOTB 88 Maximum chord distance across the supraorbital torus at or above frontomarale temporale [e, e, 3]Outer bi-orbital breadth OBOB 88 88 Frontomalare temporaleefrontomalare temporale [43, e, e]Inner bi-orbital breadth FMB 76 Frontomalare anteriorefrontomalare anterior [43a, FMB, e]Bi-orbital breadth BOBB 76 Ektoconchioneektoconchion [44, e, e]Bi-zygomatic breadth ZYB (114) Zygionezygion [45, ZYB, e]Bi-jugal breadth JUBm 94 Jugaleejugale [45(1), zJUB, e]Midorbital breadth BZOB 44 Zygoorbitaleezygoorbitale [45(3), e, e]Bi-maxillary breadth ZMBm 77 Zygomaxillareezygomaxillare [46, e, e]Bi-maxillary breadth ZMB 77 Zygomaxillare anteriorezygomaxillare anterior [46b, ZMB, e]Facial subtenseNasospinale subtense (r/l) NASS 15/18 Nasospinale to ZMB [e, e, e]Facial angleFacial profile angle FPFA (105.5) Angle formed below the FH and nasioneprosthion line [z72, e, e]Porionenasioneprosthion angle PNPA (89.5) Angle formed below the porionenasion and nasioneprosthion lines [e, e, e]Infraorbital surface angle IOFA 100.5 Angle formed superoposteriorly to the FH and malar infraorbital surface [76, e, e]Supraorbital torusSOT sagittal length (midorbit) SOTL3 17.1/e Glabellaesupraglabellare [e, e, e]SOT thickness (midorbit) SOTT3 6.8/e Supraorbital torus thickness at the midorbital level [e, e, e]SOT thickness (lateral) SOTT5 8.0/e Supraorbital torus thickness at the lateral quarter point of the superior orbital margin [e, e, e]Orbit and interorbital regionInterorbital breadth DKB 14 Dacryonedacryon [z49a, DKB, e]Anterior interorbital breadth AIOB 13 Maxillofrontaleemaxillofrontale [50, e, e]Interorbital pillar subtense IOPS Nasion to the chord of anterior interorbital breadth [e, e, e]Bi-trochlear fovea breadth BTFB 18 Min. chord distance between trochlear fovea. The landmarks are usually located at the inferior margin of the fovea [e, e, e]Orbital breadth (r/l) OBBm 33/e 32 Maxillofrontaleeectoconchion (Martin) [51, e, e]Superior orbital breadth (r/l) SOBB 30/>29 Min. distance between the trochlrear fovea and frontomalare anterior [e, e, e]Orbital height (r/l) OBHm 32/e 31 Taken at the center of the orbit perpendicularly to the OBBm [52, zOBH, e]Malar regionMaximum malar height (r/l) XMLH 36/e Frontomalare anteriorezygomaxillare [e, e, e]Malar frontal process length (r/l) MFPL 23/e Frontomalare temporaleejugale [e, e, e]Malar frontal process width (superior) (r/l) MFPW1 9/e Frontomalare anteriorefrontomalare temporale [e, e, e]Malar frontal process width (middle) (r/l) MFPW2 12/e Minimum distance from the tip of postmarginal process of the zygomatic frontal process to lateral orbital margin. [e, e, e]
Y.Kaifuet
al./Journal
ofHum
anEvolution
61(2011)
644e682
648

Malar
fron
talprocess
width
(inferior)(r/l)
MFP
W3
14/(15
)Min.d
istance
from
juga
leto
lateralo
rbital
margin[e
,e,e
]Minim
um
malar
heigh
t(r/l)
WMH
17/17
Min.d
istance
betw
eentheinferior
orbitalmarginan
dzy
gomatic
inferior
border
[e,W
MH,e
]Po
sition
ofinfrao
rbital
foramen
(r/l)
PIOF
4.4/6.2
Min.d
istance
from
thesu
periormarginof
theinfrao
rbital
foramen
totheinferior
orbitalmargin[e
,e,e
]Midface
andpalate
Nasal
brea
dth
NLB
m21
21Max
.transverse
inner
brea
dth
[54,
zNLB
,e]
Nasal
heigh
tNLH
m38
Verticald
istance
from
nasionto
nasospinale.
Slightlydifferentfrom
butvirtually
samewithHow
ell’s
NLH
[55,
zNLH
,e]
Cliv
uslength
CLV
L(13)
Nasospinalee
prosthion[48(1),e
,e]
Max
illoa
lveo
larlength
MALL
(52)
Prosthionealve
olon
[z60
,e,e
]Pa
late
length
PALL
51Olareestap
hilion
[62,
e,e
]Ex
tern
alpalatebrea
dth
MAB
5252
Ectomolaree
ectomolare[61,
MAB,e
]Intern
alpalatebrea
dth
IPAB
31En
dom
olaree
endom
olare(M
2leve
l)[63,
e,e
]Pa
latalh
eigh
tPA
LH9
Mea
suredperpen
dicularlyto
thealve
olar
planeat
theleve
lof
theposterior
marginof
M1alve
oli[64,
e,e
]
aEstimates
arein
paren
theses.B
ilateralmea
suremen
tsareindicated
as“(righ
t)/(left)”.
bCorresp
ondingmetricco
des
fortheMartin’smethod
s(Bräuer,1
988),H
owells(197
3),andKaifu
etal.(20
08)a
rein
squareparen
theses.“z”¼differentb
uts
imila
rdefi
nition.“e”¼noeq
uivalen
tmea
suremen
tinthereleva
nt
metricsystem
.cTa
kenfrom
thephysical
replic
aproducedfrom
themicro-CTdata.
dInfluen
cefrom
thetrau
ma-lik
edep
ressiondescribed
inthetext
isproba
blyminim
al.
Y. Kaifu et al. / Journal of Human Evolution 61 (2011) 644e682 649
CVA (Analysis 4) used 9 craniofacial measurements (GOL, XCB, BNL,BBH, AUB, XFB, NPL, BPL, NLB) and included Sts 5, OH 24, KNM-ER1813, KNM-ER 3733, Sangiran 17 and a recent modern humansample.
Gordon et al. (2008) used PCA on 6 size-adjusted, craniofacialmetric variables (GOL, XCB, BBH, ASB, BNL, BPL) to compare thecranial shape of LB1 (data from Brown et al., 2004) with samples ofrecent modern humans (Howells’ data), early modern humans, andearly hominin specimens Sts 5, OH 5, OH 24, KNM-ER 406, KNM-ER3733, Sangiran 17, Kabwe and three Neanderthal crania. They alsoinvestigated possible allometric relationships by examining ex-pected cranial shape in selected comparative subsamples scaled tothe size of LB1 using regression analysis.
Studies using 3D geometric morphometric methods followedconventional 2D morphometric studies. Baab and McNulty (2009)examined both cranial shape and size-related shape variationusing two different data sets: one analysis employed 15 neuro-cranial landmarks while a second analysis used 17 craniofaciallandmarks (a bilateral landmark is counted as one). Their firstanalysis focused on comparisons of LB1/1 (data taken from a ster-eolithographic replica) with modern humans and a number of pre-modernHomo specimens (KNM-ER 1813, 3733, 3883; D2280, 3444;Sangiran 17, two Sambungmacan, four Ngandong H. erectus, twoZhoukoudian H. erectus, and five mid-Pleistocene non-erectusHomo). Their second analysis compared LB1/1 with large, world-wide sample of modern humans, Pan, and Gorilla, as well as a rangeof fossil hominins (Sts 5, 71; OH 5; KNM-ER 406, 1813, 3733; KNM-WT 15000; D2700; Sangiran 17; Zhoukoudian reconstruction;Kabwe; Petralona).
Another 3D morphometric study, by Lyras et al. (2009), wasbased on 13 craniofacial landmarks, and compared the cranialshape of LB1 (data taken from a stereolithographic replica) withthose of 32 non-pathological modern humans, two microcephalicmodern humans, twomid-Holocene skulls from Flores, Sangiran 17,KNM-ER 1813, and Sts 5.
Despite some differences in analytical methods, variables, andcomparative specimens, the results of the previous craniometricstudies consistently showed that LB1 groups with pre-modernHomo specimens in cranial shape. It is well-separated from Aus-tralopithecus and Paranthropus on the one hand, and from post-erectus/ergaster Homo (including modern humans) on the other.The dominant factor in their affiliationwith pre-modern Homowasthe degree of facial prognathism, while the low cranial vault heightin LB1 strongly influenced the latter differences. Studies by Gordonet al. (2008) and Baab and McNulty (2009) further suggested thatthe cranial shape of LB1 can be predicted as a very small specimenof pre-modern Homo but clearly departs from the patterns inmodern human skulls. The primitivemorphology of H. floresiensis isalso documented in its inter-limb proportion, pelvis, wrist and footbones, and various other skeletal elements (Brown et al., 2004;Morwood et al., 2005; Falk et al., 2005, 2007, 2009; Argue et al.,2006; Tocheri et al., 2007; Brown and Maeda, 2009; Larson et al.,2009; Morwood and Jungers, 2009; Jungers et al., 2009a,b).
The morphological affiliation of the LB1 cranium with earliermembers of the genus Homo (H. habilis, Dmanisi Homo, Homoergaster and H. erectus sensu stricto) is more ambivalent. Someprevious studies concluded that LB1 is more similar to TurkanaH. ergaster (KNM-ER 3733 and 3883) and Dmanisi Homo (D2280and 2700) than to Asian H. erectus (Argue et al., 2006; Gordon et al.,2008; Baab and McNulty, 2009). However, the D2700 specimen issubadult and cannot be directly compared with other adult crania(Rightmire et al., 2006). The “scaling relationships” calculated byGordon et al. (2008) for their “non-Asian H. erectus” subsample isbased on KNM-ER 3733 and D2700 only, and is likely affected byvarious other factors such as regional, sexual, and growth variations

Y. Kaifu et al. / Journal of Human Evolution 61 (2011) 644e682650
as well as evolutionary changes. Potential errors in the metric dataused are another issue of concern when one uses casts or replicas,and compile data reported by different researchers. The crushedcranium of H. habilis, OH 24, has been skillfully reconstructed by R.D. Clarke (Tobias, 1991), but its cranial vault length, breadth, andheight measurements should still be used with caution.
In addition, the small sample size of H. erectus from Asia isa major drawback for these previous studies in light of thesubstantial regional and chronological variation observed in thisspecies. Past and recent studies have indicated significant differ-ences between, for example, H. erectus from China and Java as wellas chronologically early and late Javanese H. erectus (Weidenreich,1943; Antón, 2002; Durband et al., 2005; Kaifu et al., 2005b,2008, 2010a; Baab, 2010). Nevertheless, only one of the analysesby Baab and McNulty (2009) included all these major groups ofH. erectus. Brown et al. (2004) noted similarities in cranial vaultshape among LB1, H. ergaster, and some of the Sangiran H. erectus asmentioned above. Thus, comparisons with early Javanese H. erectusare particularly important in investigating morphological affinitiesof H. floresiensis, but the above-cited studies included only one suchspecimens, Sangiran 17. Kaifu et al. (2005b, 2010a) showed thepresence of significant chronological variation in mandibular,dental, and probably cranial morphology even within the strati-graphic sequence of Sangiran that spans at least 400 kyr. Sangiran17 belongs to the chronologically younger and morphologicallyderived subgroup of Sangiran/Trinil H. erectus in this framework(“Bapang-AG” stratigraphic group of Kaifu et al., 2005b). Further-more, as suggested by Jacob (1973) and supported by Kaifu et al.(2008), this specimen exhibits advanced morphological character-istics even within this younger subgroup (e.g., a larger cranial size,a wider frontal squama, a relatively longer occipital plane, anda tendency toward laterally thickened supraorbital torus; see alsoSchwartz and Tattersall, 2005).
Finally, as an approach different from craniometric methods,Argue et al. (2009) recently undertook cladistic analyses of LB1/1.The characters used in their study included 50 cranial plus another10 mandibular, dental and postcranial traits. They hypothesizedthat H. floresiensis represented a very early member of Homo thatwas not related to H. erectus/ergaster. However, apart from somemethodological and interpretational questions discussed byTrueman (2010) and Argue et al. (2010), the following pointsremain as potential problems for the study: The 50 cranial char-acters did not include aspects of overall cranial shape, which areinformative. As many as 12 of these 50 characters variously relate tothe temporomandibular joint, and 4 to the supramastoid crest.Given this biased character selection, the two requirements fortheir analyses, equal importance and genetic independency of eachcharacter, are not supported. In addition, the H. erectus cranialsample of Argue et al. (2009) comprised only three specimens(Sangiran 2, Sangiran 17, Trinil 2).
In our paper, we describe individual, detailed aspects of theexternal cranial morphology of LB1 and report metric data for thespecimen. This study primarily focuses on assessing cranialmorphological affinities and uniqueness of H. floresiensis relative toother early groups of the genus Homo, in order to discuss itsevolution. We also aim tometrically explore principal cranial shapedifferences of H. floresiensis from Homo sapiens. Because thecranium of LB1 is extremely small, cranial shape will be a majorissue of our comparisons. Although estimating effects of cranial sizeon cranial shape variation in individual fossil Homo groups isdifficult with currently available small sample sizes, some attemptsare made to investigate this possibility. As noted by Brown et al.(2004), some osteometric landmarks of LB1 are ambiguousbecause of damage and partially fused cranial sutures. We examineambiguous osteometric landmarks of LB1 based on consulting
internal morphology using micro-CT technology (Kaifu et al., 2008,in press). Morphology of the endocranial surfaces and cranialsinuses of LB1 will be reported elsewhere. Dental and dental rootmorphology is also treated separately from this report.
History of preparation
After its excavation in August 2003, T. Sutikna and P. Browncleaned, dismantled, and reconstructed LB1/1 for the initial publi-cation (Brown et al., 2004). The skeleton of LB1 was “extremelyfragile,” but the cranium was “free of substantial distortion” apartfrom the anterior portion of the right zygomatic arch which wasbroken and depressed medially (Brown et al., 2004). Photographsin Brown et al. (2004) and those taken in October 2004 also showthat, at this time, a thin layer of clay sediments still covered somesurfaces particularly on the cranial base. P. Brown identifieda cranial bone fragment including the bregma and reported thebone thickness at bregma (Brown et al., 2004), but unfortunatelythis piece is currently missing.
In December 2004, the specimen was transferred to GadjahMada University, Yogyakarta (Culotta, 2005; Dalton, 2005). After itsreturn to the original repository, Arkeologi Nasional (ARKENAS),Jakarta, in February 2005, the left zygoma and two teeth hadbroken off, and had been glued back (Morwood and van Oosterzee,2007; Peter Brown, personal communication). The above-mentioned break at the right zygomatic arch had been repaired;the right posterolateral margin of the foramen magnum had beendamaged. The bone surfaces were coated by a good amount of gluemixedwith dirt andmodeling clay, the latter of whichwas probablyused for molding. T. Sutikna and Rokhus Due Awe cleaned much ofthis residue from the facial and superior vault surfaces and appliedParaloid B-72 for reinforcement, but did not clean the cranial baseto avoid risk of damage. In March 2008, I. Kurniawan, together withY. Kaifu, cleaned the entire cranial surfaces using cotton stubsmoistened with acetone, and stabilized the bone by applying a thinor thick acetone solution of Butvar B-76. These procedures wererepeated by Y. Kaifu in March 2009, to further expose anatomicalstructures of interest such as the right lacrimal canal, and tostrengthen the bone. As a result of this latter cleaning, we foundthat fortunately LB1/1 retained most of its surface details ofanatomical importance, although a small area posterior to theopisthion had been damaged by a visitor to ARKENAS sometimebetween our work in 2008 and 2009.
However, it remains possible that the specimen had sufferedfrom other minor damage unrecognized to us or even a slightdegree of overall dimensional alteration after the original study byBrown et al. (2004). In order to assess such potential deformation,in this study, we compare the CT imagery taken by Brown andcolleagues (in April 2004) and us (in April 2009).
Materials and methods
CT scan and physical replicas
High-resolution CT imagery was obtained using the microfocalX-ray CT system TX225-ACTIS (Tesco Co.), at the UniversityMuseum, University of Tokyo, in April 2009. Original scans weretaken at 130 kV and 0.17 mA with a 1-mm-thick copper plateprefilter to lessen beam-hardening effects. Other scanning param-eters included a 512 � 512 matrix, 260 microns pixel size, and 260microns slice thickness and interval. The pixel size and slicethickness/intervals were set at up to 80 microns for close-up scansof some selected portions of the cranium. We created physicalreplicas of the upper and lower cranial vaults separately using an
Y. Kaifu et al. / Journal of Human Evolution 61 (2011) 644e682 651
EDEN 3D printer (Objet Geometries) in order to measure cranialbone thickness.
Comparisons of the 2004 and 2009 CT scans
The 2004 and 2009 CT scans are compared in order to assesspotential dimensional alteration after the former. Both the former(1 mm slice thickness, 0.359375 mm pixel size, a Siemens Emotionmedical CT scanner) and the latter (0.26 mm isometric voxel size,a Tesco industrial micro-CT scanner: see above) were converted tothe isometric voxel size of 0.325 mm with 8 bit gray scale, and thebone surface data were extracted using the HMH (half maximumheight) thresholding between the CT values of air and bone byAnalyze 8.1 (Mayo Clinic, MN). The two surface models were thensuperimposed to minimize their separation, and their dimensionaldifferences were assessed using the software Rapidform 2006(INUS technology, Inc., Seoul).
Cranial sutures
Computed Tomographic imagery (80 microns) confirmed thatthe coronal, sagittal, and lamboid sutures are mostly closed (Brownet al., 2004) and the spongy bone has become continuous amongthe frontal, parietal, and occipital bones. However, some cluesremain for locating these sutures.Coronal suture We previously noted that the coronal suture ofJavanese H. erectus runs along a low, vertical ridge on the lateralcranial surface below stephanion (Kaifu et al., 2008). The samecorrespondence between the suture and a ridge is also found inspecimens from other regions (e.g., KNM-ER 1813, 3733). LB1 hasa similar ridge 20 mm behind the supraorbital torus, and thecoronal suture probably passed on this ridge. The superior cranialvault of LB1 is cracked along an arc that approximately connectsthe right and left ridges, and the position and symmetricalarrangement of this crack suggests that it occurred approximatelyalong the fused coronal suture. As a support for this interpretation,the right and left temporal lines dip slightly downward at thecrack as usually observed at the stephanion of a human skull.
Figure 3. Some clues of the cranial sutures. Coronal (A) and sagittal (B) CT sections at the trwhite arrows may be a remnant of the compact bone between the frontal and parietal. (C)
A compact area is observed in the coronal CT sections close againstthe crack, which may be a remnant of the compact bone betweenthe frontal and parietal (Fig. 3A and B).Sagittal suture We determined the cranial midline for the sake ofosteometrics, but did not identify the exact course of the sagittalsuture.Lambdoid suture The lambdoid suture remains only fragmentallyon the external surface but its course is clearly marked by a distinctfurrow on both sides (Fig. 3C). Such association between thelamboid suture and a furrow is variably observed in othercomparative specimens.Squamosal suture We traced the squamosal suture with referenceto the micro-CT data in order to discriminate it from postmortembreaks (Fig. 4).Sphenooccipital synchondrosis This junction is probably markedby a narrow transverse furrow present on the external surface ofthe basilar part 5 mmposterior to the vomer. However, the junctionis fused and no trace of it was found in our CT imagery (80microns).Nasofronatal suture The interorbital pillar of LB1 has been scrapedaway in front of the right and left lacrimal fossae. Although thenasofrontal suture cannot be identifiedwith confidence, its locationis confined within the compact area formed by the superiormostpart of the nasal bones and outer cortex of the frontal bone (Fig. 5).We estimate the suture at the superior part of this compact area, atthe superior one-third of the orbital height and nearly the samevertical level with the frontomalare anterior.
Osteometric landmarks
Cranial midline The midline of the cranial vault was defined as theline passing through the frontal keel and inion. On the cranial base,the midline was drawn through the vomer, opisthion, and externaloccipital crest.Glabella A significant portion of the left supraorbital, glabellar, andnasal regions are lost, but a small bone preserved medial to thesupraorbital notch indicates the presence of some degree ofglabellar prominence. With reference to the fully adult frontalbones of the early Pleistocene Homo from Africa, Georgia, and Java,
ansverse crack passing around the fused coronal suture. The compact area indicated byThe diagonal furrow (black arrows) that marks the position of the lambdoid suture.
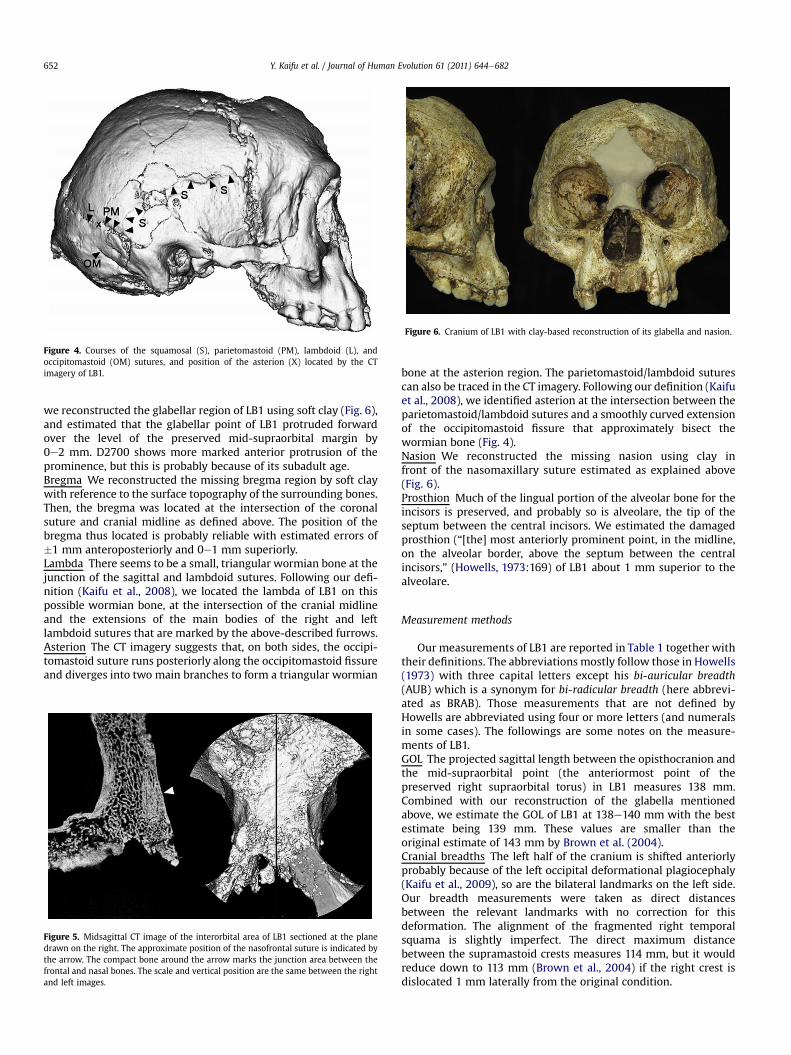
Figure 4. Courses of the squamosal (S), parietomastoid (PM), lambdoid (L), andoccipitomastoid (OM) sutures, and position of the asterion (X) located by the CTimagery of LB1.
Figure 6. Cranium of LB1 with clay-based reconstruction of its glabella and nasion.
Y. Kaifu et al. / Journal of Human Evolution 61 (2011) 644e682652
we reconstructed the glabellar region of LB1 using soft clay (Fig. 6),and estimated that the glabellar point of LB1 protruded forwardover the level of the preserved mid-supraorbital margin by0e2 mm. D2700 shows more marked anterior protrusion of theprominence, but this is probably because of its subadult age.Bregma We reconstructed the missing bregma region by soft claywith reference to the surface topography of the surrounding bones.Then, the bregma was located at the intersection of the coronalsuture and cranial midline as defined above. The position of thebregma thus located is probably reliable with estimated errors of�1 mm anteroposteriorly and 0e1 mm superiorly.Lambda There seems to be a small, triangular wormian bone at thejunction of the sagittal and lambdoid sutures. Following our defi-nition (Kaifu et al., 2008), we located the lambda of LB1 on thispossible wormian bone, at the intersection of the cranial midlineand the extensions of the main bodies of the right and leftlambdoid sutures that are marked by the above-described furrows.Asterion The CT imagery suggests that, on both sides, the occipi-tomastoid suture runs posteriorly along the occipitomastoid fissureand diverges into two main branches to form a triangular wormian
Figure 5. Midsagittal CT image of the interorbital area of LB1 sectioned at the planedrawn on the right. The approximate position of the nasofrontal suture is indicated bythe arrow. The compact bone around the arrow marks the junction area between thefrontal and nasal bones. The scale and vertical position are the same between the rightand left images.
bone at the asterion region. The parietomastoid/lambdoid suturescan also be traced in the CT imagery. Following our definition (Kaifuet al., 2008), we identified asterion at the intersection between theparietomastoid/lambdoid sutures and a smoothly curved extensionof the occipitomastoid fissure that approximately bisect thewormian bone (Fig. 4).Nasion We reconstructed the missing nasion using clay infront of the nasomaxillary suture estimated as explained above(Fig. 6).Prosthion Much of the lingual portion of the alveolar bone for theincisors is preserved, and probably so is alveolare, the tip of theseptum between the central incisors. We estimated the damagedprosthion (“[the] most anteriorly prominent point, in the midline,on the alveolar border, above the septum between the centralincisors,” (Howells, 1973:169) of LB1 about 1 mm superior to thealveolare.
Measurement methods
Our measurements of LB1 are reported in Table 1 together withtheir definitions. The abbreviations mostly follow those in Howells(1973) with three capital letters except his bi-auricular breadth(AUB) which is a synonym for bi-radicular breadth (here abbrevi-ated as BRAB). Those measurements that are not defined byHowells are abbreviated using four or more letters (and numeralsin some cases). The followings are some notes on the measure-ments of LB1.GOL The projected sagittal length between the opisthocranion andthe mid-supraorbital point (the anteriormost point of thepreserved right supraorbital torus) in LB1 measures 138 mm.Combined with our reconstruction of the glabella mentionedabove, we estimate the GOL of LB1 at 138e140 mm with the bestestimate being 139 mm. These values are smaller than theoriginal estimate of 143 mm by Brown et al. (2004).Cranial breadths The left half of the cranium is shifted anteriorlyprobably because of the left occipital deformational plagiocephaly(Kaifu et al., 2009), so are the bilateral landmarks on the left side.Our breadth measurements were taken as direct distancesbetween the relevant landmarks with no correction for thisdeformation. The alignment of the fragmented right temporalsquama is slightly imperfect. The direct maximum distancebetween the supramastoid crests measures 114 mm, but it wouldreduce down to 113 mm (Brown et al., 2004) if the right crest isdislocated 1 mm laterally from the original condition.

Y. Kaifu et al. / Journal of Human Evolution 61 (2011) 644e682 653
Superior facial height Nasioneprosthion height measured basedon the landmarks as defined above was 54 mm. Our estimate ofnasionealveolare height is 55 mm, 2 mm greater than the esti-mation by Brown et al. (2004).Cranial bone thickness Because of the difficulty of thesemeasurements on the original fossil specimen, they were takenfrom physical replicas of the upper and lower cranial vault, afterconfirming that distortions in these replicas were negligible bycomparing external cranial measurements. Because bregma thick-ness could not be obtained as a result of the loss of the bone piece,the value reported by Brown et al. (2004) was used for ourcomparative analyses. Our thickness values at the asterion andopisthocranion were corrected for the presence of the transverse/sigmoid notch and damage, respectively, at their endocraniallandmarks. The lamboid thickness may be affected slightly by thelarge trauma-like feature described below.
Comparative sample
Our comparative fossil Homo specimens are listed in Table 2.They are designated as H. habilis sensu lato, Dmanisi Homo,H. erectus from Java and China, H. ergaster (here denotesw1.8e1.0Ma East African Homo), and some other Asian and Africanpre-modern Homo. In many cases, we took the metric data forcomparative specimens from the original specimens ourselves. Asdescribed elsewhere, efforts were made to minimize various typesof errors when we use casts or published data (Kaifu et al., 2011).
Additionally, three sets of modern human (H. sapiens) samplesare used to investigate principal cranial characteristics of LB1relative to H. sapiens. The first sample is the Howell’s large cra-niometric data set taken from 2524 modern human individuals,which was used for univariate comparisons of various cranial vaultand facial measurements. The second and third samples wereconstructed for multivariate analyses of neurocranial shape (theformer) and some aspects of the cranial base (the latter), respec-tively. The second sample consists of 73 prehistoric and recentmodern humans (H. sapiens). Many of them are Holocene (Jomon,N ¼ 37) or terminal Pleistocene (Minatogawa I) hunteregathererson the Japanese archipelago, but the sample also includes anAfrican (N ¼ 1), Europeans (N ¼ 7), Iranians (N ¼ 7), Andamanese(N ¼ 10), Aboriginal Australians (N ¼ 7), Polynesians (N ¼ 2), and
Table 2Comparative pre-modern Homo specimens (adults).
Regional/chronological group Date
AfricaH. habilis s.l. 2.0e1.8 Maearly African H. erectus 1.8e1.5 Malate African H. erectus 1.4e0.9 Mac. 0.5 Ma African Homo w0.7e0.4 Ma
GeorgiaDmanisi 1.75 Ma
Javaearly Javanese H. erectus w1.5e0.8 Ma
late Javanese H. erectus w0.3e0.05 Ma
ChinaChinese H. erectus w0.8e0.6 Ma
Hexian w0.4e0.2 Mac. 0.2 Ma Chinese Homo w0.3e0.1 Ma
a Data taken from literatures and/or casts with corrections for possible shrinkage etc. ThPublished data were referred for the following specimens: SK 847 (Clarke, 1977;Wood, 192008), Saldanha (Singer, 1954; Rightmire, 2008), Ndutu (Clarke, 1990; Rightmire, 2008)Rightmire et al., 2006), Zhoukoudian (Weidenreich, 1943: exc. Skull 5), Nanjing 1 (WuPoirier, 1995), and Jinniushan (Wu, 1988; Lü, 1990).
Buryato (N ¼ 2). The sample covers a wide range of modern humanvariation in terms of cranial size (small Andamanese and largeBuryato crania) and shape (lengthebreadth index ranges from 65 to88%). The third sample consists of 197 recent humans includingthree different small-bodied groups (7 Andamanese, 17 PhilippineNegritos, and 14 African Pygmies), as well as 25 Papuans, 7Aboriginal Australians, 107 Japanese, and 20 Europeans.
Metric analyses
Somemeasurements of LB1 reported in Table 1 are estimates (asnoted above and indicated in Table 1 and the following scatter-plots), but we included these data in the following metriccomparisons because we believe that the possible errors for themare small enough.
In order to examine cranial shape characteristics of LB1 metri-cally, we perform a number of bivariate scatter plots. As a multi-variate approach, we conduct principal component analyses (PCA)which do not require a priori grouping of the sample.
As variables for the neurocranial PCAs, we chose eightmeasurements (GOL, SOTB, POBB, SQSB, ASB, BRAB, SMCB, PBRH).These are available from a relatively large fossil Homo sample, well-represent the overall cranial vault shape, and were effective todetect spatiotemporal variation in earlier Homo (Kaifu and Baba,2011). The eight measurements were size-adjusted before theanalyses by dividing them by the size variable (SV) for each spec-imen. The size variable used here is the geometric mean of thecranial length (GOL), the arithmetic average of the 6 breadths(SOTB w SMCB), and height (PBRH). The varianceecovariancematrices are used for the PCAs rather than the correlation matricesto retain the original variance structure of the variables.
First, this neurocranial PCA is applied to the pre-modern fossilsample. For this purpose, the PCs are calculated based on the fossilsample without LB1, and the PC scores for LB1 are computedafterward. In order to examine possible effects of cranial size oneach PC, the PC scores are plotted with the SV. Additionally, for thepurpose of evaluating between-group variation in some combina-tions of the PCs, cluster analyses are performed based on Euclideandistance of PC score. Several different joining methods areproposed for cluster analyses (Sneath and Sokal, 1973). Because wecannot be sure what method best represents the actual group
Specimen
KNM-ER 1470, 1590, 1805, 1813, 3732, 3735, 7330; OH 24KNM-ER 730, 1808, 3733, 3883, 3891; SK 847a
OH 9,a 12; Dakaa
Bodoa; Kabwe; Saldanhaa; Ndutua; Saléa
D2280,a 2282,a 3444a
Trinil: T 2Sangiran: S 2, 4, 10, 12, 17, 38, IX; BukuranSambungmacan: Sm 1, 3, 4;Ngandong: Ng 1, 3, 6, 7, 10, 11, 12
Zhoukoudian: ZKD 2,a 5, 10,a 11,a 12a
Nanjing: Nanjing 1a
HexianDalia; Mabaa; Jinniushana
ose without asterisk were measured by the authors based on the original specimens.91), OH 9 (Rightmire, 1990;Wood, 1991), Daka (Asfaw et al., 2008), Bodo (Rightmire,, Salé (Rightmire, 1990), Dmanisi (Gabunia et al., 2000; Lordkipanidze et al., 2006;et al., 2002; Liu et al., 2005; Vialet et al., 2010), Dali (Wu, 2009), Maba (Wu and

Y. Kaifu et al. / Journal of Human Evolution 61 (2011) 644e682654
relationships, two relatively commonly used methods are appliedto each data set. These are WPGMA (weighted pair-group methodusing arithmetic averages) and Ward methods.
For the sake of comparison with H. sapiens, this 8-variable,neurocranial PCA is also applied to a combined sample of pre-modern and modern humans using the varianceecovariancematrix. Then, another PCA is performed using seven size-adjusted,basicranial measurements (OFeCC, OFeJF, OFeOF, CCeCC, JFeJF,OFD1, and OFD2). These variables represent positional relationshipsand shape of some foramina on the cranial base (oval foramen,carotid canal, and jugular foramen). For the purpose of size-adjustment, each of these variables is divided by the square rootof the arithmetic averages of the anteroposterior (OFeCC andOFeJF) and transverse (OFeOF, CCeCC, and JFeJF) distances.
Morphological description
Preservation
The cranium of LB1 is remarkably complete (Figs. 1 and 2). Thebregmatic area, left supraorbital torus, interorbital region, and thesubnasal area were damaged at the time of the discovery. Many ofthe fragile elements on the face, zygomatic arch, and cranial baseare preserved. The parietal, temporal, occipital, and sphenoid bonessuffer from cracking and small portions are fragmented or missing(e.g., along the right coronal and sphenosquamous sutures), butthese did not significantly alter the original cranial form (Brownet al., 2004).
The surface models extracted from the 2004 and 2009 CT scansare superimposed in Fig. 7. Here, the reddish colors indicate thoseareas where the 2004 model projects over the 2009 model, and thebluish colors the opposite. Dark red and dark blue regions corre-spond to the repaired or damaged areas (indicated by the arrows)or the surface cleaning and the application of glue conductedbetween the two scans (see “History of preparation”). Apart fromthese, relatively greater separations between the two models areconfined to the frontal squama, nuchal plane, and palate, where the2004 scan is larger by up to 0.3e0.9 mm. Otherwise, discrepanciesare largely within �0.3 mm. In other words, the superior andinferior surfaces are relatively inflated (by 0.3e0.9 mm) in the 2004compared to the 2009 models, but differences are less clear on theanterior, posterior, and lateral cranial surfaces. No physical damageis evident in themicro-CT sections taken at the locations of greatestdifference, the right frontal and left nuchal plane (sections AeB,CeD, and EeF in Fig. 7), although the fragmented condition alongEeF does not preclude the possibility that the nuchal area had beenslightly altered after the 2004 scan. The pattern and magnitude ofdiscrepancy observed between the two scan sets suggestsa combination of systematic effects along the axis of the 2004 scan(of up to circa 1%) and residual distortion elsewhere in either orboth scan sets (Gen Suwa, personal communications).
In summary, the dimensional differences between the twomodels are small, if any, and most of the observed differences maynot reflect the actual differences of the original cranial specimenbetween 2004 and 2009. We consider that dimensional change inLB1/1 after its initial preparation is negligible and differences in thecraniofacial measurements between Brown et al. (2004) and thepresent study, which are generally minor (Table 1), are mostlybecause of slight differences in methods, landmark identification,and use of estimates where there are missing parts.
Asymmetric distortion and pathology
In superior view, the left parieto-occipital region is flattened andthe entire cranium is skewed slightly in a parallelogram form. The
face also exhibits asymmetric distortion, such as the asymmetricdevelopment of the canine and premolar juga, the horizontalrotation of the maxillary body, the rotated occlusion of the maxil-lary and mandibular dentitions, and various distortions in themandible, as are detailed elsewhere (Jacob et al., 2006; Kaifu et al.,2009, 2010a). These are in all likelihood due to posterior defor-mational (positional) plagiocephaly, that is, the retention of plasticcranial deformation during the infancy (Kaifu et al., 2009). Falk et al.(2010) suggested that the presence of cracks in the LB1 vault duringthe initial laboratory preparation indicates the partial contributionof taphonomic distortion to the observed asymmetry. This may bethe case, although in our view the remarkable preservations of thefragile parts of the cranium (e.g., the vomer and palatines; otherskeletal parts such as the scapula as well) and consistent patterns ofthe distortion between the cranium and mandible suggest thatdeformational plagiocephaly was the dominant factor for all theasymmetries.
The palate of LB1 is rotated rightward relative to its cranial base.The rotation angle measuresw6� when oriented on the basis of theFrankfurt Horizontal, as can be assessed from Figs. 1 and 2. Thelesser value recently claimed by McNulty and Baab (2010), 2.91�,may have resulted from their use of a stereolithographic replica.Furthermore, the unusual observation in LB1 lies not only in itsdegree of maxillary rotation, but also in its resultant asymmetriesin the development of the canine and premolar juga. We infer thatthis situation suggests that the rotation of the maxillary body wasdriven by the rotation of the maxillary dental arch, which occurredin conjunction with the mandibular deformation caused by theanterior shift of the glenoid fossa on the left, affected (flattened)side of the neurocranium (Kaifu et al., 2009).
There is another observation that is consistent with thehypothesis of deformational plagiocephaly. The articular surface ofthe occipital condyle of LB1 (reasonably intact on the right side butposterior portion of the left condyle is also preserved) is notsmoothly convex but is slightly concave with a distinct depressionon its posterior half. The superior articular facet of the LB1’s atlas(LB1/3) also exhibits a similarly roughened surface topology (Fig. 8).This metamorphosis suggests some immobility of the atlanto-occipital joint, and may have resulted from unbalanced right andleft neck muscles inserted on the asymmetrically deformed cranialsurface. Alternatively, LB1 may have suffered from congenitaltorticollis and this condition may have produced posterior defor-mational plagiocephaly during her infancy. Unfortunately, asym-metry in the clavicle cannot be examined because the left claviclehas not been found.
There are a number of small and large irregular depressions onthe external cranial vault surface, which appear to be healed lesions(Fig. 9). The largest one is on the obelion region and measuresw30 mm in diameter. This feature has no taxonomic meaning (cf.Argue et al., 2009) and is probably irrelevant for the deformationalplagiocephaly (cf. Morwood and Jungers, 2009). The second largestdepression is at the center of the frontal squama and measuresw13 mm diameter (Brown et al., 2004). Similar traces of healedlesion are reported in many fossil Homo skulls from Java(Weidenreich, 1951; Indriati, 2006), China (Weidenreich, 1943;Shang and Trinkaus, 2008), Georgia (Rightmire et al., 2006), aswell as Africa (KNM-ER 3732: Wood, 1991).
The maxillary second premolars are bilaterally rotated withtheir buccal cusps situated mesially. Tooth rotation is a relativelycommonly observed dental anomaly in humans and othermammals, with suggested etiologies including some geneticmechanism and a lack of space for the normal tooth eruption(Baccetti, 1998; Jacob et al., 2006; Lukacs et al., 2006; Natsumeet al., 2006). In the hominin fossil record, 90� rotation of premo-lars are reported for an adolescent skull from Dmanisi (the

Figure 7. Color density maps showing differences (distances) between the surface models based on the 2004 and 2009 CT scans of LB1, and three micro-CT sections. Reddish colorsindicate those areas where the 2004 model is larger than the 2009 model, and bluish colors do the opposite. The two dark red areas pointed by the arrows 1 (foramen magnummargin) and 2 (nuchal plane) were damaged after the 2004 scan, and the dark blue areas 3 and 4 (zygomatic bones) had been repaired/modified (see “History of preparation”). The2004 scan was done with the mandible articulated with the cranium, and this affects the colors on the dental occlusal surfaces and the right mandibular fossa. Most other dark redareas (distances > 0.9 mm) indicate the removal of sediments and the dark blue areas (distances > 0.6 mm) reflect the glue used for stabilization after the 2004 scan. Thedistribution shown on the left is calculated only for those areas with separations less than 1.5 mm to eliminate localized, substantial separations caused by the above factors. Thethree micro-CT sections shown below (scale bar ¼ 10 mm horizontally and 1 mm vertically; voxel sizes ¼ 260 and 80 microns for AeB and CeD/EeF, respectively) were taken at theareas where the differences between the 2004 and 2009 models are relatively great (0.3e0.9 mm).
Y. Kaifu et al. / Journal of Human Evolution 61 (2011) 644e682 655
malformed, left P2 of D2700: Rightmire et al., 2006) and an adultmandible from Konso (the left P2 of KGA10-1: Suwa et al., 2007). Asdescribed by Brown and Maeda (2009) and confirmed by Jungersand Kaifu (2011), the maxillary teeth of LB1 are free of dentalcaries but there is heavy dental calculus mainly around the buccalcervical lines of the posterior teeth.
Neurocranial outlines
Viewed superiorly, the lateral ends of the supraorbital torusproject outward only moderately beyond the levels of the temporalfossae. The vault assumes a rounded, teardrop contour. Behind thenarrow forehead, it broadens substantially toward the supra-mastoid region, with a slight degree of outward convexity. In
association with this last morphology, in the basal view, a signifi-cant portion of the sphenoidal greater wing is visible lateral to theinfratemporal crest. The zygomatic arch flares modestly outward sothat the bi-zygomatic breadth (ZYB: 114) is equal to the supra-mastoid breadth (SMCB: 114). In basal view, the area of radiculare ishollowed medially, so that the bi-radicular breadth (BRAB: 105) isdistinctly smaller than the ZYB and SMCB (114). Posterior to thesupramastoid crests, the contour of LB1 flexes posteromedially toform an almost evenly curved, continuous posterior contour of thevault. The posterior extent of the LB1 cranium is comparativelyrestricted partly because of its weak occipital torus development.
In lateral view, despite the missing bregma region, the vertex ofLB1 is apparently located at the anterior third of the parietal. Thevertex coincides with bregma in many of our comparative fossil

Figure 8. Atlas of LB1 (LB1/3) placed in front of its occipital condyle. Note theconcavities and roughened topology in their articular surfaces.
Y. Kaifu et al. / Journal of Human Evolution 61 (2011) 644e682656
Homo crania, but is more posterior in some African and Indonesianspecimens (KNM-ER 3732, 3883; Sm 1; Frankfurt Horizontal wasestimated for those specimens with missing landmarks). Thefrontal and parietal midline curvatures are strong (see “Individualvault bones” below for more details), although the contour of theposterior parietal is affected by the large trauma-like hollowdescribed above. The gently curved occipital plane of LB1 inclinesweakly forward to smoothly continue to its parietal midline viaa weak lamboidal depression. Similar occipital contours are seen inH. habilis, KNM-ER 3733, D2280, D2282, and Sangiran and Zhou-koudian H. erectus, although the presence/absence of lamboidaldepression varies among them. In contrast, the occipital planes ofSambungmacan, Ngandong, Kabwe, KNM-ER 3883, Dali (andpossibly D3444) stand vertically above their occipital tori, and flexforward near the lambda. The opisthocranion of LB1 is on its weak
Figure 9. Two trauma-like depressions mentioned in the text.
external occipital protuberance, which is positioned in a low levelclose to the Frankfurt Horizontal. The nuchal plane is markedlyrounded outward.
The posterior contour of LB1 is relatively wide and low with itsmaximum breadth situated on the weak supramastoid crest orstrong mastoid crest. The gently convex lateral cranial surface ofLB1 stands nearly vertically directly above the lateral edges of itscranial base. Many of our comparative specimens show similarmorphology, although the lateral vault surfaces of Dmanisi, OH 9,Sangiran 4, and Zhoukoudian are inclined medially, and those ofH. habilis and Dmanisi are situated distinctly medially to the lateralmargins of their cranial bases (Santa Luca, 1980; Rightmire et al.,2006). The vault contour of LB1 is evenly rounded with no orminimal development of the parasagittal flattening, parietaleminence, and supramastoid crest. H. erectus from Sangiran andTrinil typically exhibit parasagittal flattening and downward flexionof the contour at the parietal eminence irrespective of overallcranial size (i.e., including smaller crania such as S 10 and 38), butthose from Sambungmacan (Sm 1 and 3) and Ngandong tend toshow rounded profiles similar to LB1. The contour is rounded buttypically more square in H. habilis, H. ergaster, and Dmanisi.
The sharp mastoid crest protrudes laterally to the same level asthe poorly developed supramastoid crest. Such a laterallyprotruding mastoid crest (SMCBz or < XMTB) is more common inAfrica (KNM-ER 1805, 1813, 3733; OH 24, 9; Daka; Kabwe), but isalso observed in H. erectus (S 38; Ng 6, 7; ZKD 11). H. erectus fromSangiran typically exhibit strong supramastoid crests (Villmoare,2005) in contrast to the condition of LB1. Below the mastoidcrest, the lateral side of themastoid process of LB1 (complete on theright side) is straight and strongly slopes inferomedially. LB1 ismore similar to Sangiran H. erectus in this respect.
Ectocranial keeling, sutures, and wormian bones
A short segment (25 mm) of the thick, prominent frontal keel ispreserved in the mid-squama region. The keel appears to continuetoward the supraglabellar fossa, although its anterior segment isobscured by the damage and a trauma-like depression of diameter13mm. The preserved parabregmatic area indicates that coronal andparietal keels were either not present or poorly developed, if any. Afrontal keel is variably present in all the comparative subsampleswith strong expression on the mid-squama similar to LB1 found inKNM-ER 3733, ZKD 10, ZKD 11, ZKD 12, T 2, and to a lesser extent, insuch specimens as Kabwe, S 17, Sm 3, Ng 7, and Dali. Marked coronaland/or sagittal keels are regarded as non-African features (Andrews,1984; Stringer, 1984), although their expression varies withinH. erectus with some of them exhibiting comparatively limiteddevelopment of these structures (e.g., S 12, S IX).
The sutures in the pterion region of LB1 probably assumed an “Hpattern” with the coronal suture, located as explained above,separated from the squamosal suture by w5 mm on the betterpreserved left side. The lambdoid sutures are mostly fused butfurrows, as mentioned above, mark their courses. KNM-ER 1470and 1813 exhibit complicated sutural formations in their lamb-doidal area (Wood, 1991). Such feature is not evident in LB1, but itapparently had a triangular lambdoidal ossicle similar to thoseobserved in Sm 3, Sm 4, and Ng 12. Our CT imagery also indicatesthe presence of a Wormian bone at the asterion of LB1 as notedabove. This is a common observation in Javanese H. erectus (Kaifuet al., 2008).
Temporal line and associated surface structures
Between the right and left superior temporal lines, the externalsurfaces of the frontal, parietal, and occipital bones exhibit a porous

Figure 10. Photograph (left) and surface rendered CT image (right) of the palate and anterior cranial base of LB1/1. Note the posteriorly elongated palatine (PL) and pterygoid plate(PP).
Y. Kaifu et al. / Journal of Human Evolution 61 (2011) 644e682 657
texture, as normally seen in human skulls. The superior and inferiorlines show clear divergence in the middle of the frontal squama.They run closely to each other toward the lambdoid suture, withthe maximum separation being w5e7 mm in the area on theanterior parietal. The temporal lines of LB1 are marked on theanterior two-thirds of the parietal, and weak but traceable on theanterior frontal squama and posterior parietal. In many of theAfrican (e.g., KNM-ER 1813, 3733, 3883, 3891; OH 9; Daka; Kabwe),Georgian (D2280), and Indonesian specimens from Sambungmacanand Ngandong crania, the temporal line is crested or ridge-like onthe anterior frontal squama. Comparatively weak anterior temporallines may be a characteristic of H. erectus from Sangiran andZhoukoudian (cf. Argue et al., 2009).
In superior view of the cranium, the right and left temporal linesare well-separated and maintain similar distances to each other
Figure 11. Diagonal views of LB1/1. Note the short and narrow face, the relatively tall but naperture margin that suggests the presence of a prominent nasal bridge, the forward-facing (deep infraorbital sulcus (IOS), and the canine fossa (CF) that is restricted to the inferior pa
except for a slight degree of posterior divergence. Thus, itsminimum frontal breadth (61) is slightly exceeded by thebi-stephanic breadth (64), and the lines are w70 mm apart in thearea of the parietal eminence. In the mid-parietal area, the linepasses over the indistinct parietal eminence. On the posteriorparietals, the superior lines of LB1 approach the lambdoid suturesleaving only a narrow area (w5 mm) for the development of theangular torus in between them at the mastoid angle. Here, theangular torus is a low, small eminence (better preserved on the leftside). Characterization of this torus is not easy due to variable sizeand prominence, unclear definition, and surface weathering(Kimbel and Rak, 1985; Villmoare, 2005), but the above-describedconfiguration in LB1 is generally similar to those in East AfricanHomo such as KNM-ER 3733 and OH 9. The tori of Asian H. erectusvary from a low, small mound (S 2, 10), to a moderately developed
arrow orbit, the bulbous lateral part of the supraorbital torus, the everted lateral nasalin a horizontal section) infraorbital surface, the forward protruding maxillary body, thert of the infraorbital surface.

110 120 130 140 150 160 170
SMCB
120
160
200
240
GO
L*
h
h
e
o
D
K
NG
G
s
s
ss
m
m
mN
N
NNNN
L
zz z
H
D
e
110 120 130 140 150 160 170
SMCB
70
80
90
100
110
120
PBR
H
h
h
h
ee
oD
K
AG s s
ss
s
s s
s
m
mm
N
N
NN
N
L
z
z
z
H
D
120 160 200 240
GOL*
70
80
90
100
110
120
PBR
H
h
h
ee
oD
K
G s
s
s
s
m
mm
N
N
NN
N
L
z
z
z
H
D
110 120 130 140 150 160
XMTB
100
110
120
130
140
150
160
SQSB
h
h
ee
o
D
K
A
G
s ss
ss
ss
s
m
m
mN
N
N N
N
L
zz
z
HD
110 120 130 140 150 160 170
SMCB
100
110
120
130
140
150
160
SQSB
h
h
h
ee
D
K
A
G
s ss
ss
s s
s
m
m
mN
N
NN
N
L
zz
z
HD
o
100 110 120 130 140 150 160
BRAB
100
110
120
130
140
150
160
SQSB
h
h
ee
D
K
A
G
s ss
sss
s
m
mN
N
NN
N
L
zz
z
HD
o
100 110 120 130 140 150 160
XBPB
60
70
80
90
100
110
120
POBB
h
h
h
he
e
o
D
B
KS
N
AG
G
Ts
s
ss
m
m
m
NNNNN
N
L
zz z
H
D
100 110 120 130 140 150 160
XBPB
80
90
100
110
120
130
140
SOTB
h
h
h
e e
o
D
BK
G
s
s
ss
mm
mNNN
N
N
L
z
zz
H
D
100 110 120 130 140 150 160
XBPB
80
90
100
110
120
130
140AS
B
h
h
ee
o
D
KS
N
GG G
ssss
s
ss
s
m
m
m
NNN
NNN
L
zz z
H
D
h
(1.4)
(1.3)
(1.2)
(0.8)(0.7)
(0.6)
(0.55) (0.5)
(1.1)(1)
(0.9)
(1)
(0.9)
(0.8)
(1)
(0.95)
(0.9)
(1) (0.9)
(0.8)
(0.8)
(0.7)
(0.6)
(1) (0.9)
(0.8)
60 70 80 90 100 110 120
POBB
0
10
20
30
40
SOTB
- PO
BB
h
h
h
h
e
e o
D B
K
G
s
sss
mm
m
NN N
NNL
zzzz
j
H
DM
110 120 130 140 150 160 170
SMCB
-10
0
10
20
30
SMC
B - B
RAB
h
h
ee oDKA
G
s
s
ss
s
sm
m
m
NN
NN
N
L
z
z
z
H
D
A B C
D E F
G H
J K
I
Figure 12. Scatter plots of neurocranial measurements. Lines based on Y ¼ aX are drawn with the coefficients indicated in the parentheses. Metric codes: ASB ¼ bi-asterionicbreadth; BRAB ¼ bi-radicular breadth; GOL ¼ max. cranial length; PBRH ¼ porionebregma height; POBB ¼ postorbital breadth; SMCB ¼ supramastoid breadth; SOTB ¼ supraorbitaltorus breadth; SQSB ¼ squamosal suture breadth; XBPB ¼ max. bi-parietal breadth; XMTB ¼ max. bi-mastoid breadth. “*” indicates that the value for LB1 is an estimate. Symbols:
Y. Kaifu et al. / Journal of Human Evolution 61 (2011) 644e682658

Y. Kaifu et al. / Journal of Human Evolution 61 (2011) 644e682 659
eminence (S 12, 17), to an extensive, triangular raised area centeredbetween their well-separated superior line and lambdoid suture (SIX, Bukuran, Sambungmacan, Ngandong, Zhoukoudian).
The inferior line of LB1 almost sticks to the superior lineposterior to the area of parietal eminence. Then, at the level of thelambda, they diverge from each other to form a crescent-shaped,smooth area between them. Above the angular torus, the superiorand inferior lines approach each other again, and direct anteriorlytoward the supramastoid and mastoid crests, respectively. Thesupramastoid crest is a low, blunt eminence, and runs nearly hor-izontally. LB1 differs in this respect from Sambungmacan andNgandong where the crest stands more vertically to continue intothe anteriorly located posterior segment of the inferior temporalline (see Fig. 12 in Kaifu et al., 2008). The mastoid crest formsa nearly horizontal, sharp ridge below the narrow intertoral sulcuswith a minimum width of w8 mm (supramastoid crest-mastoidcrest distance [SMCD]).
Individual vault bones
Frontal bone (squama) The frontal keel is well-developed andshows a strong curvature in the lateral view, although thesquama sagittal curvature is much less strong beside this midlinekeel. Either side of the LB1 squama is flattened with nodevelopment of the frontal eminences. These surfaces inclineinferolaterally toward the temporal line, so that the squamaassumes a tent-like configuration in its coronal section. Sucha combination of marked frontal keel and flat, sloping right andleft squamae is often found in African post-1.8 Ma Homo (e.g.,KNM-ER 3733, Daka, Kabwe) and early Javanese H. erectus, butnot in H. habilis and H. erectus from Sambungmacan, Ngandong,and Zhoukoudian, where the parasagittal contour is more curveddue to the development of the frontal eminence. In superiorview, the anterior borderline of the frontal squama behind thesupratoral plane of LB1 is convex with anteriorly protrudingmidline area.Parietal bone Viewed laterally, the anteromedial corner of eachparietal is flattened. The cranial midsagittal contour is stronglycurved in the mid-parietal region, and then flattened again in itsposterior one-third. The last morphology is partly affected fromthe posterior deformational plagiocephaly and the large trauma-like depression spanning between the obelion and lambdaregions.
Otherwise, the parietal bones of LB1 are generally convex andnearly evenly curved in both parasagittal and coronal sections.There is no marked parasagittal flattening and the parietaleminence is indistinct as described in the “Neurocranial outlines.”We see no obelionic depression as described byWeidenreich (1943)for ZKD 3 (contra., Argue et al., 2009), but the relevant area of LB1exhibits a large trauma-like depression. Among our comparativespecimens, the parietal surfaces of H. habilis, H. ergaster, Sam-bungmacan and Ngandong H. erectus, and Maba show generalconvexity similar to LB1, whereas the parietals of Sangiran andTrinil H. erectus are characterized by relatively flat or hollowedsuperior surfaces (parasagittal flattening), flexed coronal crosssection, and variably developed sagittal keel.Temporal bone (excluding basal structures) We tracked the supe-rior squamosal margin of LB1 by consulting the micro-CT imagery,as illustrated in Fig. 5. It is relatively straight, despite slight
A ¼ Salé; B ¼ Bodo; D ¼ Dali; D ¼ Daka; e ¼ Turkana H. ergaster; G ¼ Dmanisi; H ¼ Hexian; hN ¼ Ngandong; o ¼ Olduvai H. ergaster; S ¼ Sardanha; s ¼ Sangiran; T ¼ Trinil; z ¼ ZhuokoudHomo; orange ¼ Dmanisi; green ¼ Indonesia; violet ¼ China; red ¼ Flores. African specimereader is referred to the web version of this article.)
damages along the margin (contra Argue et al., 2009). Among thefossil hominins, there is a general chronological trend towardincreasing curvature of the squamosal suture (Terhune and Deane,2008), and a distinctly arched suture is seen in Kabwe and Dali inour sample. Terhune and Deane (2008) suggested that there wasa positive correlation between the curvatures of the cranialmidsagittal contour and the squamosal suture, but themorphology of LB1 apparently does not follow this said trend.
Both a proportionally long temporal squama and short mastoidportion (parietomastoid suture) are present. In addition, thedevelopment of the supramastoid and suprameatal crests is poor,and the zygomatic arch does not show strong lateral flare. Thezygomatic arch of LB1 is thin mediolaterally but thick verticallythroughout its length, and orients horizontally relative to theFrankfurt Horizontal.Occipital bone (excluding basal parts) In the superior/basal viewof the cranium, the LB1 occipital exhibits limited posteriorprotrusion with evenly curved contour, as described above. Inlateral view, the occipital bone of LB1 is flexed at theweak occipitaltorus with the occipital angle being 106�. In sagittal dimensions,the nuchal plane of LB1 slightly dominates over its occipital plane.
Details of the midline contour were described in “Neurocranialoutlines” above. The occipital plane of LB1 is gently convex in bothsagittal and transverse sections. The lambdoid area is curvedforward to continue into the lambdoidal depression. This depres-sion is laterally continuous with the above-described furrows alongthe right and left lambdoid sutures. Similar morphology is variablyobserved among our comparative specimens, but the most similarcase to the LB1 is seen in Sm 1 from Java.
The occipital torus of LB1 is weak but clearly present asa transverse ridge confined to the central one-third of the occipital.Immediately above the torus is a shallow, straight supratoral sulcuswhich is continuous from the right to left sides without interrup-tion. This form of morphology is most typically observed inH. erectus of Asia although the strength of torus development varieswithin this group. An African specimen of Saldanha is also similar toLB1 in this respect but H. habilis and H. ergaster are differentexhibiting pronounced external occipital protuberances and/or lackof supratoral sulcus. The iniac region of LB1 is roughened by thedevelopment of a thin, irregular plate of bone.
Cranial base
Sphenoid bone The posterior margin of the pterygoid plate ispreserved. The medial and lateral laminae of the plate are narrowlyseparated from each other in its inferior half behind the junctionwith the palatine, but the two laminae appear to be fused to forma single plate in their superior portions. This superior portion of theplate is 4e5 mm thick and extends substantially posteriorly andthen posterolaterally toward the blunt sphenoid spine (Fig. 10). Atthe point of the flexion, the root of the plate rims theposteromedial margin of the oval foramen (complete on the leftside). The oval foramen is circular, of w4 mm in diameter, has nobony bridge dividing it, and is well-separated from the posteriormargin of the sphenoid by the above-described root of thepterygoid plate. Lateral to the oval foramen, the preglenoid planeof LB1 is flat andweakly sloping relative to the Frankfurt Horizontal.
The medial and lateral pterygoid plates are well-preserved inOH 24, Kabwe, and Ng 7 among our comparative sample. None of
¼ H. habilis; j ¼ Nanjing (not plotted here); K ¼ Kabwe; L ¼ LB1; m ¼ Sambungmacan;ian. Color codes (online version only): light blue ¼ H. habilis; blue ¼ post-1.8 Ma Africanns are in italic. (For interpretation of the references to color in this figure legend, the

Y. Kaifu et al. / Journal of Human Evolution 61 (2011) 644e682660
these shows extensive fusion between the two components,although their separation is less complete in Ng 7 in terms of thenarrower distance as well as the development of irregular, distinctbony bridges connecting between the two plates. A posteriorlyextended pterygoid plate situating medial to the circular ovalforamen is probably a primitive characteristic of Homo. Thismorphology is found in 2.0e1.5 Ma African and Georgian Homo(KNM-ER 1805, 1813, 3733, 3883; OH 24; D2280, D2700) as well asJavanese H. erectus (Ng 7, and possibly in Sm 4 and Ng 12). Incontrast, post-1.5 Ma African Homo tend to approach the modernhuman condition, in which a constricted oval foramen is situatedbehind the root of the lateral pterygoid lamina, and is situated closeto the posterior border of the sphenoid (i.e., the posteromedialmargin of the oval foramen consists of a thin plate of bone).Unfortunately, the situation in Chinese H. erectus is unknown in theabove respects.Mandibular fossa The mandibular fossa of LB1 is anteroposteriorlywide and shallow partly because of the low, hollowed (transverselyconcave) articular eminence. The shallow fossa of LB1 contrastswith the anteroposteriorly restricted, deep fossae of Ngandong andZhoukoudian.
Medially, the entoglenoid pyramid (process) of LB1 is formed bythe squamous temporal and sphenoid in nearly equal proportion.The pyramid extends posteriorly close to the tympanic plate, and itsconvex posterolateral surface contributes to form a small medialrecess between it and the tympanic plate (Weidenreich, 1943;Tobias, 1991). This configuration is variably observed in ourcomparative Homo crania. The massive pyramids in some H. habilisandH. ergaster specimens show strong inferior projection (KNM-ER1813, 3733, 3891; KNM-WT 15000 [subadult]; OH 13 [subadult]:see Baab, 2008) but this is not the case in Asian fossils includingLB1. As a result of the distinct posterior protrusion of the entogle-noid pyramid mentioned above, the medial portion of the articularsurface of LB1 is flexed and faces laterally. A similarly flexed artic-ular surface is seen in S 2, S 17, ZKD 3, and ZKD 12, but not in Africanand Georgian Homo and H. erectus from Sambungmacan andNgandong.
Bilaterally there is a pinhole-sized perforation at the squa-motympanic fissure. The fossa roof is sagittaly concave and itsposterior half curls downward to form a prominent posteriormargin. Here, the postglenoid process does not take the form ofa distinct, linguiform structure, but is marked by this prominentmargin that fuses with the underlying tympanic plate medially.Such an indistinct postglenoid process is sporadically found inour comparative sample (e.g., KNM-ER 3733, S 2), but this isdifferent from Ngandong where the roof of the mandibular fossatypically deepens posteriorly without forming a downwarddirected margin (except Ng 7: Weidenreich, 1951; Durband, 2002,2008; Baba et al., 2003). The ectoglenoid process of LB1 is a low,blunt, triangular mound situated at the base of the zygomaticprocess.Tympanic The lateral margin of the tympanic plate of LB1 is locatedsubstantially medially to the suprameatal crest. In basal view, theplate is an anteroposteriorly restricted, narrow tube. It is orientedcoronally with the lateral tympanic point e carotid canal lineforming an angle of 93� with the sagittal plane (b of Dean andWood, 1981; average of the right and left sides with damagedlandmark estimated). The anterior portion of the tympanic plate(anterior wall of the external acoustic meatus [EAM]) of LB1 isflattened and stands vertically behind the mandibular fossa.Below the EMA, the inferior surface of the plate is rounded alongits long axis. Medially, the tympanic flexes slightly forward at thecarotid canal to extend for w10 mm onto the petrous bone(complete on the left side). In lateral view, the curved tympanicplate shows almost uniform thickness and supports the lower
half of the EAM. The EAM exhibits an oval contour with its longaxis leaning slightly forward. The lateral margin of the tympanicis incised in a form of “V.” From the base of this V a deeptransverse groove incises the inferior surface of the tympanic forw13 mm. Posterior to this groove is the low tympanic (petrous)crest with a blunt inferior margin. The crest shows only a limiteddegree of inferior projection beside the pit for the styloid process,which is absent. In other words, the structure does not developinto a strong vaginal process as defined by Tobias (1991:94)(Brown et al., 2004).
The indistinct vaginal process of LB1 is most similar to thecondition seen in H. habilis. Although Nevell and Wood (2008)suggested that the first appearance of substantial bony vaginalprocess occurred in H. ergaster, a strong vaginal process appears tobe a general characteristic in post-1.8 Ma Homo including Dmanisi.Otherwise, the above morphology is similar to specimens such asKNM-ER 3733, as well as Sangiran and Zhoukoudian H. erectus. Thetubular form of the tympanic plate close to the LB1 condition isdominant morphology in H. habilis and H. erectus but is also foundin some other specimens (e.g., KNM-ER 3733). A modern human-like condition of a vertical, flat tympanic plate become dominantin post-1.6 Ma African Homo (Nevell and Wood, 2008). The ante-roposteriorly wide and basally thick tympanic plate of Ngandong/Sambungmacan contrasts with the apparently more gracile platein LB1.
The transverse groove on the tympanic, albeit weak, isrecorded in Ngandong (at least one side of Ng 1, 6, 7, 11, andpossibly 12) and Zhoukoudian (ZKD 11, and ZKD 3 has a cleft), andis also present among Sambungmacan, Kabwe, and Dmanisi(Weidenreich, 1943, 1951; Rightmire et al., 2006; Argue et al.,2009; personal observation). Weidenreich (1943) ascribed thisto a poor or incomplete fusion of the anterior and posteriorontogenetic components of the tympanic, and Tobias (1991)further connected this with the notch (¼ the V-shaped incisionin LB1) on the tympanic lateral margin seen in ZhoukoudianH. erectus and OH 24.
Behind the tympanic crest, the tympanomastoid fissure of LB1 isnot extensive, if any (cf. Brown et al., 2004). The extensive fissures(tympanomastoid groove of Tobias, 1991:94) reported for Sam-bungmacan and Ngandong H. erectus are formed between theirwidely separated tympanic crest and mastoid process (the groovein OH 24 is not so extensive compared to these Indonesian indi-viduals, cf. Tobias, 1991), but the tympanic crest of LB1 (see above)is located close to its mastoid process.Petrous portion The inferior surface of the petrous bone is intactonly in its posterior portion on the left side and its morphology isdifficult to detail except for the following (cf. Brown et al., 2004):First, the long axis of the petrous forms an estimated angle ofw40� with the sagittal plane, and the estimated petrous angle (aof Dean and Wood, 1981) is w54�. These figures are similar tothose reported for H. habilis and H. erectus s. l. (Dean and Wood,1982; Tobias, 1991:98). Second, a bony bridge separates theanteromedial and lateral portions of the jugular foramen of LB1.Various forms of jugular foramen bridging are observed in KNM-ER 3883, Kabwe, Sm 4, Ng 7 and 12 (Weidenreich, 1951).Mastoid portion There is no styloid process but its position ismarked by a pit located 4 mm lateral to the carotid canal. Thepresence of a bony styloid process is reported for some Africanand Georgian specimens (KNM-ER 3733, OH 9, OH 12, Kabwe,D3444: Rightmire, 1990; Wood, 1991; Antón, 2004; Lordkipanidzeet al., 2006) but this structure is generally regarded to be absentin H. erectus (Antón, 2003). As in Zhoukoudian H. erectus(Weidenreich, 1943) and Dmanisi Homo (Rightmire et al., 2006),the small stylomastoid foramen of LB1 is located outside of theline connecting the digastric groove and the styloid pit.

Y. Kaifu et al. / Journal of Human Evolution 61 (2011) 644e682 661
The small, pointed mastoid process of LB1 (intact on the rightside) is strongly inclined medially below the prominent mastoidcrest. Its posterolateral aspect that supports neck muscles is flat-tened and faces posteriorly rather than laterally. The abovecombination of morphology is shared with many of the Sangiran(S 2, 12, 17; Bukuran, but not S 4) and Zhoukoudian (ZKD 3 and 11),but not with Ngandong H. erectus. The mastoid process in Ngan-dong (and Sambungmacan, to a lesser degree) is characterized bya large size and a flattened but more laterally-facing posterolateralsurface. The case in Africa is not very clear due to small samplesize. At least some African hominins exhibit inferiorly directingprocesses (KNM-ER 1813, 3883; OH 12) and strong medial incli-nation is not a typical observation except for a juvenile individual(KNM-WT 15000) and a case with a marked mastoid crest (KNM-ER 1805).
The digastric groove of LB1 is narrow and shallow. The surfacemedial to the groove is damaged but is smooth. There is nodevelopment of the occipitomastoid crest and the area of themastoid foramen is deeply grooved along the occipitomastoidsuture.Occipital (basilar and lateral parts) The external surface of thebasioccipital of LB1 is mostly intact except for the damage on itsright lateral margin. The surface is flat transversely and is ata slightly superior level relative to the structures lateral to it(petrous, entoglenoid process, infratemporal surface, tympanicplate, etc.). Posteriorly, the basioccipital broadens and slopes infe-riorly relative to the Frankfurt Horizontal, and flexes downward infront of the slightly damaged basion. The above characteristics aregenerally shared with Javanese H. erectus (S 17, Sm 4, Ng 7, Ng 12)and H. ergaster (KNM-ER 3733, 3883; OH 9). Data for ChineseH. erectus are not available. More fossils are needed to know thecondition in H. habilis, but the basioccipital of OH 24, as well as thatof Sts 19, are not distinctly depressed superiorly. Kabwe and Saléapproach themodern human condition in this respect, showing thebulbous basioccipital which stands out inferiorly from the level ofits lateral structures.
The foramen magnum of LB1 is slightly inclined so that theopisthion is lower than the basion. Its anterior margin is located atthe bi-tympanic line as widely seen in various hominin taxa. Theforamen magnum of chronologically later Javanese H. erectus fromSambungmacan and Ngandong are anteroposteriorly long andexhibit a characteristic narrowing just in front of the opisthion(opisthionic recess : Weidenreich, 1951; Delson et al., 2001; Kaifuet al., 2008). In LB1, the weakly convex posterolateral margin ofthe foramen magnum (nearly intact on the left side) indicatesa slight development of this recess at the opisthion, although theforamen is not as long as Sambungmacan/NgandongH. erectus. Thisrecess with a convex posterolateral foramenmagnummargin is notfound in other comparative specimens.
Posterior to the condyle, the bone surface around the foramenmagnum is damaged. Postcondyloid tuberocities, a characteristicfeature of Sambungmacan and Ngandong H. erectus, are weak ifpresent.Occipital (nuchal plane) Both right and left occipital nuchal planesshow prominent bulges, which largely correspond to deep cere-bellar fossae. Combined with the weak external occipital crest andpoor development of the depressions beside the crest, the entirenuchal plane of LB1 is convex both anteroposteriorly and trans-versely. Extensively bulging nuchal plane morphology similar toLB1 is relatively common in chronologically earlier Homo speci-mens from Africa, Java and China, whereas a substantial area of theplane below the nuchal torus is flattened in Daka, Kabwe, as well asSambungmacan and Ngandong H. erectus. In these latter speci-mens, the surface is even hollowed due to the development of aninferiorly overhanging occipital torus.
Facial skeleton
Supraorbital region The supraorbital torus of pre-modern Homocan usually be divided into three areas: a central glabellarswelling, median arches (superciliary arches), and lateralprominences (Santa Luca, 1980). Only the last component, whichextends from the supraorbital notch to the frontal zygomaticprocess, is intact on the right frontal of LB1. This occupies thelateral three-quarters of the orbital breadth, and arches over thestrongly curved superior orbital margin. The supraorbital tori ofour comparative Homo specimens are mostly weakly curved orstraight in the coronal plane, but a strong curvature similar toLB1 is seen in Daka and Maba.
The preserved torus has a bulbous sagittal cross sectionalcontour throughout. Vertical thickness of the LB1 torus is thinmedially and thickens laterally to the point immediately above thefrontomalare temporale. Another remarkable characteristic of theLB1 torus is that its slightly bulbous lateral end projects laterally tothe level of the frontomalare temporale (Brown et al., 2004). Infacial view, the lateral segment of the torus curves downward andits lateral margin descends vertically before reaching the fronto-malare temporale, so that the supraorbital torus breadth (SOTB) ismeasured 0e1.2 mm above the frontomalare temporale (i.e., SOTBis identical with the outer bi-orbital breadth [OBOB] and can bemeasured anywhere in this area). A similar configuration (i.e., SOTBis measured above the frontomalare temporale, or SOTB> OBOB) isfound only in post-1.5 Ma Homo from Africa and Asia in ourcomparisons (OH 9, Daka, Kabwe, S 17; many of the Sambungma-can, Ngandong, and Zhoukoudian specimens as well as Dali),whereas the gracile lateral margin of the supraorbital torus of2.0e1.5 Ma African Homo slopes inferolaterally and does notprotrude beyond the frontomalare temporale.
The superior orbital margin has a deep supraorbital notch. Insuperior view, the anterior toral surface above the notch is flattenedand coronally oriented (Figs. 1 and 2). A supraorbital notch isa common observation in H. habilis, H. ergaster, and H. erectus, butmany of the post-0.6 Ma African Homo (Bodo, Kabwe, and Ndutu)have supraorbital foramina (cf. Dodo and Sawada, 2010). The flat-tening of the area above supraorbital notch is extensive in LB1 andcomparable to some post-1.8 Ma African (KNM-ER 3733), Georgian(D2280), Javanese (S 17, IX, Bukuran; Ng 11), and Chinese (ZKD 10,12) specimens. The supratoral plane of LB1 gently slopes ante-roinferiorly from the frontal squama and does not form a distinctsulcus or groove as seen in KNM-ER 3733, D2282, and ZhoukoudianH. erectus.Orbit The orbit of LB1 is relatively high and narrow. The roundedsuperior orbital margin of LB1 is uncommon in our comparativesample, but is similar to Maba and Daka as stated previously. Oneach side of the medial orbital walls, the lacrimal fossa extendsupward to the level of the superior one-third of the orbital height,where the fossa is bordered by a diagonally passing sharp crest. Atthe superomedial corner of each orbit is a small, deep trochlearfossa.Nasal region and cavity The interorbital pillar of LB1 is relativelynarrow with the ratio of the anterior interorbital breadth (AIOB) toinnerbiorbital breadth (FMB) being 24%. Surface morphology of theinterorbital region of LB1 is unknown, but the preserved basal partof the left maxillary frontal process faces anterolaterally, suggestingthe presence of a moderately prominent nasal bridge similar to S 17before the break. The same surface tends to face more anteriorly inH. habilis, SK 847, and KNM-ER 3733 (Franciscus and Trinkaus,1988).
The nasal floor of LB1 is transversely concave on the premaxillabut has flat areas posteriorly on the maxillary palatine process.Lateral to the relatively extensive incisive fossa, the nasal floor of

Y. Kaifu et al. / Journal of Human Evolution 61 (2011) 644e682662
LB1 smoothly continues from the premaxilla to the maxillarypalatine process (“smooth” in the scheme of McCollum, 2000). Thisregion is reported to be “stepped” in Au. afarensis and Au. africanusbut appears to be variable in early Afro-Asian Homo (Rightmire,1998; McCollum, 2000). Both the intermaxillary crest and vomeris missing above the nasal floor and its insertion to the premaxilla isunknown.Zygomatic The zygomatic frontal process is slender with minimaldevelopment of the postmarginal process. In the lateral view, thezygomatic frontal process stands nearly vertically and formsa right angle with the zygomatic arch, so that the posteriormargin of the zygomatic is widely open at the jugale. The ante-rior surface of the process of LB1 faces anterolaterally, whereasthe same surfaces of H. habilis, although variably damaged, facemore anteriorly. In H. habilis, this process swings obliquely(anteroinferiorly) in the lateral view (KNM-ER 1470, 1813, 3732;OH 24). The isolated frontal process of KNM-ER 3735 is small insize but otherwise similar to, particularly, KNM-ER 3732. Theanterior surface of the zygomatic frontal process faces morelaterally in post-1.8 Ma East African Homo, Javanese H. erectus(S 10 [isolated zygomatic], S 17), and Maba, but not in Dmanisi,ZKD 12, and Dali. Laterally, the zygomatic bone of LB1 showsa relatively sharp posterior flexion at the point slightly lateralto the zygomaxillare to continue into the zygomatic arch,which exhibits only a modest degree of lateral flare asdescribed above.Infraorbital and paranasal surfaces The infraorbital surface of LB1is extremely short vertically. It faces slightly inferiorly in a sagittalsection, and anteriorly in a horizontal section (Figs. 1, 2, 5 and 11).Immediately below the inferior orbital margin is a thick,transverse bony ridge, and the area below this ridge is hollowedto form a shallow canine fossa. A single, large infraorbital foramenopens relatively high on the surface of LB1 at the superomedialcorner of this hollowed area. As seen in the lateral view (Figs. 1, 2and 5) the maxillary body of LB1 markedly protrudes beyond thelevel of the infraorbital surface. The surface immediately lateral tothe lateral nasal margin is smoothly concave without forminga pillar-like structure seen in Au. africanus (Rak, 1983; Brownet al., 2004). Along the root of the maxillary zygomatic processand below the infraorbital foramen is a deep infraorbital(maxillary) sulcus that creates a “stepped” topography betweenthe maxillary body and infraorbital surface (Fig. 11). Examples ofa markedly protruding maxillary body and associated steppedsurface topography similar to LB1 are found in S IX (Kaifu et al.,2011), ZKD 13, Dali, and to a lesser extent in D2282 and subadultD2700 from Dmanisi, although this morphology is not evident onthe damaged paranasal surface of S 17. In contrast to this possibleAsian morphology, the infraorbital surface continues moresmoothly to the lateral nasal margin without a deep gap in theAfrican specimens of H. habilis (KNM-ER 1470, 1805, 1813; OH 24),SK 847, H. ergaster (variously broken KNM-ER 3733 and 3883;juvenile KNM-WT 15000 as well), as well as Bodo and Kabwe.
The maxillary zygomatic process of LB1 is a thin plate. In thebasal view perpendicular to the Frankfurt Horizontal in Fig. 1 or 2(or to the alveolar plane, which is not shown here), the anteriorsurface of the process is in the plane passing between P2 and M1
(mesial M1), and its lower margin is directly above the mesial M1
(mid-M1). In the facial view, the straight zygomaticoalveolar crestextends diagonally (superolaterally). LB1 is unique among the Asiancomparative specimens in this latter aspect. The crests of S 17, S IX,and Dali are more horizontal while three maxilla of ChineseH. erectus exhibit marked malar incisures (ZKD 11, 13; Nanjing 1).Inclined, straight zygomaticoalveolar crests are seen in some post-1.8 Ma African specimens (SK 847, Bodo, Kabwe, and probablyKNM-ER 3733) but not in KNM-ER 1813 and Dmanisi, suggesting
that the horizontal arrangement of the crest is a plesiomorphy inHomo (Kaifu et al., 2011).Subnasal region and alveolar part The vertically low nasoalveolarclivus of LB1 is damaged except for a small area above the rightlateral incisor, but enough morphology remains to tell that theclivus show neither extensive concavity nor strong anteriorprotrusion present in many H. habilis specimens (KNM-ER 1805,3891, 7330, and OH 24) and KNM-ER 3733. Posteriorly, the lateralsurface of the maxillary alveolar process supporting the almoststraight canine-molar row virtually forms a single plane thatfaces laterally. In facial view, the maxillary body of LB1 narrowssuperiorly, leaving the prominent canine and premolar jugawhose expression is asymmetrical as mentioned above.
A small, circular alveolus is left for the missing left third molar,but no alveolar bone is formed for the congenitally defective rightthird molar. The maxillary alveolar arch of LB1 is small and rela-tively long anteroposteriorly. The dental arch is not parabolic butnearly square with sharp flexion at the canines as seen in H. habilis,H. ergaster, and Sangiran H. erectus (but not in Kabwe and ZKD 13).The damaged anterior tooth row of LB1 may have been slightlyconvex anteriorly, and the nearly straight right and left canine-molar rows show a slight degree of posterior divergence. Medialto the molars, the alveolar bone is thickened to cover the mediallyprotruding lingual root of eachmolar. The large incisive canal opensat a posterior positionwith its posteriormargin situated at a level ofthe mesial second premolars.
Interestingly, the palate of LB1 is markedly elongated posteri-orly, forming a wide space behind the transverse line passing theposterior alveolus wall for the left third molar (w9 mm: Fig. 10).The greater palatine foramen is located far behind the third molar,and the maxillary tuberosity of LB1 is also elongated accordingly tocontinue into the diagonally arranged pterygoid plate of thesphenoid described above. None of the sufficiently-preservedpalatines in our comparative sample show such posterior exten-sion of the palate (OH 24, KNM-ER 1813, D2700, D2822, Kabwe),although further cleaning is necessary to examine the condition inS 17.
Metric comparisons
Bivariate comparisons with pre-modern Homo
In the following bivariate scatter plots that aim to demonstrateshape characteristics of LB1/1, one or more lines based on Y ¼ aXare drawn, where appropriate, to indicate isometric relationships.In some cases,maximum bi-parietal breadth (XBPB) or supramastoidbreadth (SMCB) are used as proxies for the neurocranial size. Theseeffectively increase sample size, and show correlation coefficientsof 0.93 and 0.91, respectively, with the size variable (SV) for the PCAwithin the pre-modern Homo sample including LB1 (N ¼ 26).Overall shape Bivariate plots of various neurocranial measure-ments are shown in Fig. 12. The LB1 vault is anteroposteriorly shortrelative to its breadth (Fig. 12A), and is similar to KNM-ER 1470,D3444, Sm 3, and Hexian in this respect. The maximum craniallength (GOL) of LB1 is an estimate with an inferred error range of�1 mm: this does not significantly influence the aboveinterpretation. The low heightebreadth index of LB1 is sharedwith Turkana H. ergaster, Sangiran H. erectus, and a part of theSambungmacan and Zhoukoudian H. erectus. When theheightelength proportion is also considered (Fig. 12C), LB1 is themost similar to Sm 3 (the smallest ‘m’) in the above three aspects.
Fig. 12DeF compare a midvault breadth (SQSB) relative to threedifferent basal cranial breadths (BRAB, SMCB, XMTB). LB1,H. erectus/ergaster, and other middle Pleistocene fossils generallyshow relatively higher SQSB values in these plots, indicating that

Y. Kaifu et al. / Journal of Human Evolution 61 (2011) 644e682 663
their vaults are expanded transversely compared to the H. habilis orDmanisi conditions (Kaifu and Baba, 2011). As a H. habilis specimenwith a similar basal cranial breadth to that of LB1 (KNM-ER 1813) islocated directly below the position of LB1 in these plots, andbecause the H. habilis crania (and Dmanisi) are generally plottedbelow the isometric lines for the other specimens, the above shapedifferences between H. habilis and H. floresiensis are unlikely to beexplained simply as cranial size-related variation (see below fora multivariate examination of this aspect).
Proportions of the anterior (SOTB, POBB) and posterior (ASB)vault breadths relative to the midvault breadth (XBPB) arecompared in Fig. 12GeI. SOTB/XBPB shows clear separationbetween all the African/Georgian and Asian subsamples, and LB1shows a low ratio which is equal to the Asian condition. AsH. habilis specimens with similar XBPBs to that of LB1 (KNM-ER1813, 3732) are located directly above the position of LB1 inFig. 12G, the above differences between H. habilis andH. floresiensis cannot be simply explained as cranial size-relatedvariation between SOTB and XBPB. In Fig. 12H, low POBB/XBPBratios similar to that of LB1 are observed in the specimens amongH. habilis, Turkana H. ergaster, Sangiran H. erectus, and Hexian, aswell as Ndutu and Salé, but not in 1.5e0.5 Ma African Homo(including Kabwe and Saldanha) and late Javanese H. erectusfrom Sambungmacan and Ngandong. LB1 shows a relativelynarrow ASB that is different from Sangiran H. erectus as well assmall samples of H. habilis and late 1.5e1.0 Ma H. ergaster(Fig. 12I).
Bivariate plots in Fig. 12J and K provide further details aboutLB1’s vault shape. The supramastoid breadth (SMCB) exceeds bi-radicular breadth (BRAB) by only 4 mm or less in KNM-ER 1470 and1813 (Fig. 12J), and similar relationships must have existed in thefragmentary or damaged KNM-ER 3735 and OH 24 which are notplotted here. Some of the Dmanisi and Zhoukoudian specimensalso show strong lateral protrusion of the radiculare (Kaifu andBaba, 2011), whereas H. ergaster and Javanese H. erectus aresimilar to LB1 with the differences between the twomeasurementsranging from 8 to 20 mm. On the other hand, the comparativelysmall difference between supraorbital torus breadth (SOTB) andpostorbital breadth (POBB) in LB1 is a characteristic shared withAsian archaic Homo specimens, but not with African and Georgianfossils compared here (Fig. 12K).Temporal lines The characteristics of temporal lines describedabove are metrically demonstrated in the plots of Fig. 13AeC. Onthe frontal squama, the lines of LB1 are well-separated from eachother and do not show the strong medial incursion seen in someH. habilis specimens (KNM-ER 1805, 3732: Fig. 13A). Most Africanand Georgian crania show strong incursion of the temporal lineson their parietals (bi-stephanic breadth [BSTB] >> bitemporal linebreadth on parietal [BTLB]: Fig. 13B). LB1 does not show this trend(BSTB � BTLB ¼ 0), and is similar to H. erectus from Java andChina in this respect. The temporal muscle attachment length(TMAL) shows chronological reduction in Javanese H. erectus dueto anterior shift of the posterior end of the temporal line inNgandong (Kaifu et al., 2008). LB1 does not show this derivedcondition, showing a higher proportion of TMAL/GOL than theaverage condition in Ngandong (Fig. 13C).Individual vault bones The intact portion of the midline keel andsurrounding bone surfaces suggests that the midsagittal curvatureof the frontal squama (frontal squama angle [FSQA]) of LB1 isstrongest in our comparative sample (Fig. 13D). This curvature isvariable within each of our regional/chronological comparativesubsamples, and several specimens approach the morphology inLB1 (KNM-ER 3733; ZKD 11; Sm 3). The same curvature appearsto be strong in Maba whose FSQA cannot be obtained due todamage. It is worthy to repeat here, however, that the strong
sagittal curvature of the LB1 frontal is restricted to its midlinekeel and not on either side of the squama.
The parietal midsagittal curvature of LB1 is the strongest in ourfossil sample (parietal angle [PAA]¼w134�: Fig. 13E), although thisfigure is slightly affected from the posterior deformational plagio-cephaly and the large trauma-like depression as mentioned above.This angle value is comparable only to Sm 1 from Java, whose anglevalue (134�) may be slightly increased by the surface cracking andinflation (Kaifu et al., 2008).
An upwardly bending supramastoid crest in chronologicallylater Javanese H. erectus produces a relatively short temporalsquama length (TSQL) and a long parietomastoid suture length(PMSL) (Kaifu et al., 2008). Fig. 13F suggests that relative shorteningof the temporal squama and lengthening of the parietomastoidsuture was a general chronological trend in Pleistocene Homo. Inthis respect, LB1, with a long temporal squama and a short parie-tomastoid suture, shares a primitive morphology with 2.0e1.0 MaAfrican and Indonesian Homo.
The occipital squama is relatively low in LB1 and is similar toJavanese H. erectus and Hexian in this respect (OCC/ASB in Fig. 13G).The lamdaeopisthocranion length (LOPC) of LB1 (37 mm) is slightlysmaller than its opisthocranioneopisthion length (OPOC; 41 mm).This proportion does not depart from the conditions exhibited byany of the regional/chronological subsamples examined here(Fig. 13H). Within Java, a shorter occipital plane characterizes thechronologically earlier group from Sangiran, while equality orreversal of proportions are frequently encountered in Sambung-macan and Ngandong. The positions of lambda in KNM-ER 1470and 1813 are complicated by the development of complex lamboidossicles (Wood, 1991). We placed the lambda of these H. habilisspecimens at the posteriormost point of their possible rangesfollowing our own definition (Kaifu et al., 2008).
The occipital angle (OCAO) of LB1 (106�) is in the upper range ofvariation exhibited by H. erectus/ergaster and is more acutecompared to the two H. habilis specimens (KNM-ER 1470 ¼ 111�,1813 ¼ 116�: Fig. 13I). This difference from H. habilis is probablyindependent from the cranial size variation because LB1 and KNM-ER 1813 are similar to each other in cranial size.Cranial base Relative anteroposterior dimensions of the middlepart of the cranial base are extended in late Javanese H. erectuscompared to their predecessors in the region (Kaifu et al., 2008).This midcranial base lengthening is not evident in LB1, as seen inthe plots of the basioccipital length (BASL) and foramen magnumlength (FMGL) with the parietal chord (PAC) (Fig. 14A and B),although LB1 shows incipient development of opisthionic recessas described above. African H. ergaster, Kabwe, and S 17 sharerelatively short BASL and FMGL with LB1. These measurementsare also short in OH 24 (15 and 29 mm, respectively), althoughthe PAC is not available for this H. habilis specimen. Similarly, thepublished photograph (Lordkipanidze et al., 2006) suggests thatmarked lengthening of the midcranial base is not a feature inD3444.
The shallow mandibular fossa of LB1 described above is re-flected in mandibular fossa depth (MAFD) in Fig. 14C, althoughcaution is needed in that this measurement is directly influencedfrom the degree of inferior projection of the tympanic plate (seeTable 1 for the definition). The shallow fossa of LB1 contrasts withthe deep fossae of Ngandong and Zhoukoudian H. erectus. Many ofthe 2.0e1.6 Ma African Homo as well as Sangiran and Sambung-macan H. erectus exhibit shallow fossa morphology more or lesssimilar to LB1, whereas the fossa depth is great or intermediate inothers (OH 9, Daka, S 4).
The relative width of the mandibular fossa (MAFW/SMCB inFig. 14D) of LB1 is close to those of three Sangiran H. erectus (S2, 17,IX) but smaller than the average conditions in H. habilis and

60 70 80 90 100 110 120
POBB
20
30
40
50
60
70
80
90
100
110
WFR
B
h
h
h
ee
DB
KS
A
GG
T
s
s
ss
mm
mNNN N NN
L
z
z
H
D
o
(1)
(0.8)
(0.6)
(0.4)
100 110 120 130 140 150 160
XBPB
-5
0
5
10
15
20
25
30
BSTB
- BT
LB
h
h
ee
o
B
K
GG
T
s
s
s
s
sm
m NNN NNL z
H
70 80 90 100 110 120
PAC*130
140
150
160
PAA*
h
h
he
eD
KS
AG
s
s s s
ss
m
m
m NN
N
N
N
L
z
zz
H
50 60 70 80 90 100 110
FSQC*
130
140
150
160
170
180
FSQ
A*
h
h
h
e
e
D
B
KS
A
G
G
s
s
s
s
mm
mN
N
NN N
N
L
z
z
zH
80 90 100 110 120 130 140
ASB
60
70
80
90
100
OC
C
h
h
e
D
KN
GG
s
ss s
ss
s
mm
mN
N
NN
N
N
L
zzH
(0.8)
(0.7)
(0.6)
40 50 60 70
OPOC
20
30
40
50
60
70
LOPC
h
e
e
DK
s
s
s
s
s
s
s
mm mN
NN
N
N
N
L
H
(1)
(0.8)
(0.6)
A B C
D E F
G H I
60 70 80 90 100
OCC
90
100
110
120O
CAO
h
h
eD
K
NA
G
G
ss s
s
s
s
s m
m
m
N N
N
N
NN
L z
z
H
50 60 70 80 90
TSQL
0
10
20
30
40
PMSL
h
he
e
o
K
Gs
s s
ss
m
m
m
NNN
NN
L
zzz
zH
D (0.3)
(0.2)
120 160 200 240
GOL*
100
110
120
130
140
150
TMAL
h
e
K
G
T
s
s
s
s
m
mm
N
NN
N
N
L
z
z
H
e
(0.8) (0.7)
(0.6)
Figure 13. Scatterplots of neurocranial measurements. Lines based on Y¼ aX are drownwith the coefficients indicated in the parentheses. Metric codes: ASB ¼ biasterionic breadth;BSTB ¼ bistephanic breadth; BTLB ¼ bitemporal line breadth; FSQA ¼ frontal squama angle; FSQC ¼ frontal squama chord; LOPC ¼ lambdaeopisthocranion chord; OCAO ¼ occipitalangle; OCC ¼ occipital chord; OPOC ¼ opisthocranioneopisthion chord; PAA ¼ parietal angle; PAC ¼ parietal chord; PAC ¼ parietal chord; POBB ¼ postorbital breadth;PBRH ¼ porionebregma height; PMSL ¼ parietomastoid suture length; SMCD ¼ supramastoid breadth; TMAL ¼ temporal muscle attachment length; TSQL ¼ temporal squamalength; WFRB ¼ min. frontal breadth; XBPB ¼ max. biparietal breadth. “*” indicates that the value for LB1 is an estimate. Symbols and color codes (online version only) as in Fig. 12.(For interpretation of the references to color in this figure legend, the reader is referred to the web version of this article.)
Y. Kaifu et al. / Journal of Human Evolution 61 (2011) 644e682664
H. ergaster, as well as Kabwe and Dali. The fossa widths of Zhou-koudian H. erectus are not available due to damage, but theyappear to be as narrow as Sangiran H. erectus (ZKD 11, 12, as well as3). The above observation is largely, but not entirely consistentwith that of Terhune et al. (2007) who characterized themandibular fossae of Turkana H. ergaster (KNM-ER 3733, 3883;KNM-WT 15000) as large and shallow and those of H. erectus assmall and deep. It should be noted that, besides the smallernumber of Indonesian H. erectus specimens analyzed by theseauthors (S 4, 17; Sm 1, 3; Ng 6, 7, 12), the study of Terhune et al.(2007) is based on the unrotated principal components extracted
from 3D landmark data for the entire basal temporal, which are notideal for examining variation in individual, local morphologicaltraits.
Cranial breadths across the tympanic plates (BTYB) and supra-mastoid crests (SMCB) are plotted in Fig. 14E. With a few exceptions(ZKD 10, 12; OH 9), H. erectus, H. ergaster, and other middle Pleis-tocene crania show lesser BTYB/SMCB ratios, meaning that theirexternal acoustic meatuses are more medially located in the cranialbase compared to the H. habilis condition. Although the strongerdevelopment of the supramastoid crests in H. erectus contributesto this metric variation, the difference is also evident between
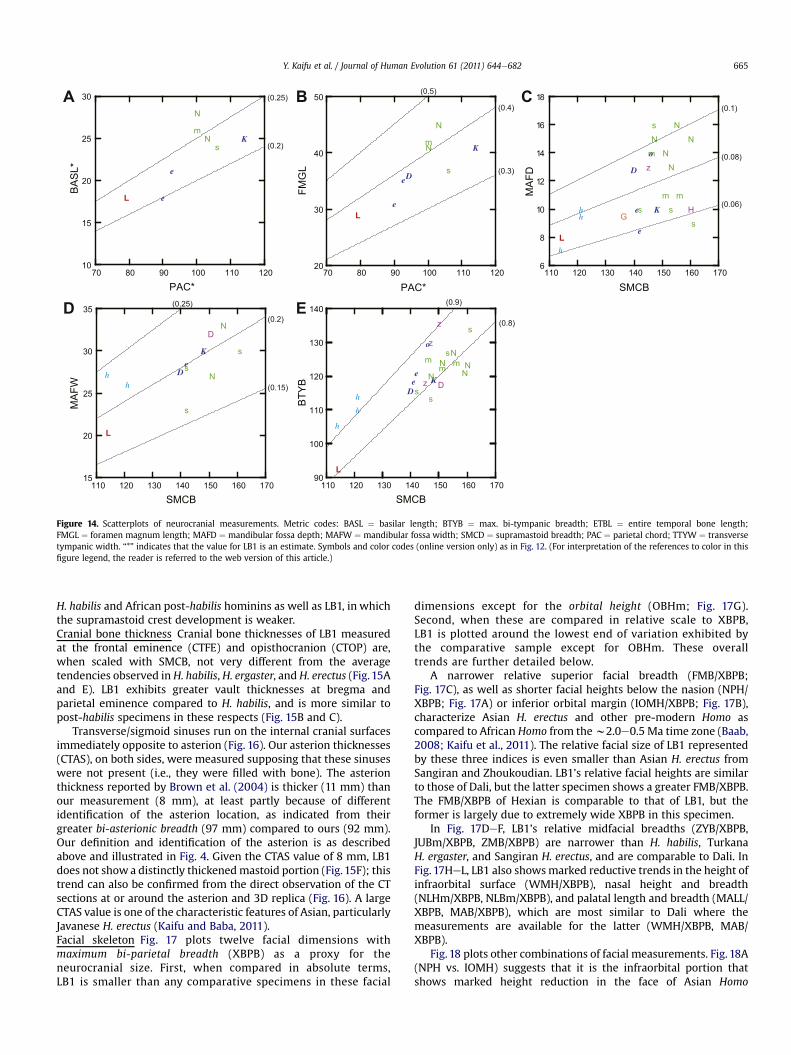
70 80 90 100 110 120
PAC*
20
30
40
50
FMG
L
e
e
D
K
s
mN
N
L
70 80 90 100 110 120
PAC*
10
15
20
25
30
BASL
*
e
e
Ks
m
N
N
L
(0.25)
(0.2)
(0.5)
(0.4)
(0.3)
A B
D E
C(0.1)
(0.08)
(0.06)
110 120 130 140 150 160 170
SMCB
6
8
10
12
14
16
18
MAF
D
h
hh
e
e
D
KG
s
s
ss
m m
N
NN N
N
L
z
H
mo
110 120 130 140 150 160 170
SMCB
15
20
25
30
35
MAF
W
(0.25)
(0.2)
(0.15)h
h
eD
K
s
s
s
N
N
L
D
110 120 130 140 150 160 170
SMCB
90
100
110
120
130
140BT
YB
h
h
h
ee
o
DK
ss
s
s
mm m
N
N NNN
L
z
z
z
D
(0.9)
(0.8)
Figure 14. Scatterplots of neurocranial measurements. Metric codes: BASL ¼ basilar length; BTYB ¼ max. bi-tympanic breadth; ETBL ¼ entire temporal bone length;FMGL ¼ foramen magnum length; MAFD ¼ mandibular fossa depth; MAFW ¼ mandibular fossa width; SMCD ¼ supramastoid breadth; PAC ¼ parietal chord; TTYW ¼ transversetympanic width. “*” indicates that the value for LB1 is an estimate. Symbols and color codes (online version only) as in Fig. 12. (For interpretation of the references to color in thisfigure legend, the reader is referred to the web version of this article.)
Y. Kaifu et al. / Journal of Human Evolution 61 (2011) 644e682 665
H. habilis and African post-habilis hominins as well as LB1, in whichthe supramastoid crest development is weaker.Cranial bone thickness Cranial bone thicknesses of LB1 measuredat the frontal eminence (CTFE) and opisthocranion (CTOP) are,when scaled with SMCB, not very different from the averagetendencies observed in H. habilis, H. ergaster, and H. erectus (Fig.15Aand E). LB1 exhibits greater vault thicknesses at bregma andparietal eminence compared to H. habilis, and is more similar topost-habilis specimens in these respects (Fig. 15B and C).
Transverse/sigmoid sinuses run on the internal cranial surfacesimmediately opposite to asterion (Fig. 16). Our asterion thicknesses(CTAS), on both sides, were measured supposing that these sinuseswere not present (i.e., they were filled with bone). The asterionthickness reported by Brown et al. (2004) is thicker (11 mm) thanour measurement (8 mm), at least partly because of differentidentification of the asterion location, as indicated from theirgreater bi-asterionic breadth (97 mm) compared to ours (92 mm).Our definition and identification of the asterion is as describedabove and illustrated in Fig. 4. Given the CTAS value of 8 mm, LB1does not show a distinctly thickenedmastoid portion (Fig.15F); thistrend can also be confirmed from the direct observation of the CTsections at or around the asterion and 3D replica (Fig. 16). A largeCTAS value is one of the characteristic features of Asian, particularlyJavanese H. erectus (Kaifu and Baba, 2011).Facial skeleton Fig. 17 plots twelve facial dimensions withmaximum bi-parietal breadth (XBPB) as a proxy for theneurocranial size. First, when compared in absolute terms,LB1 is smaller than any comparative specimens in these facial
dimensions except for the orbital height (OBHm; Fig. 17G).Second, when these are compared in relative scale to XBPB,LB1 is plotted around the lowest end of variation exhibited bythe comparative sample except for OBHm. These overalltrends are further detailed below.
A narrower relative superior facial breadth (FMB/XBPB;Fig. 17C), as well as shorter facial heights below the nasion (NPH/XBPB; Fig. 17A) or inferior orbital margin (IOMH/XBPB; Fig. 17B),characterize Asian H. erectus and other pre-modern Homo ascompared to African Homo from thew2.0e0.5 Ma time zone (Baab,2008; Kaifu et al., 2011). The relative facial size of LB1 representedby these three indices is even smaller than Asian H. erectus fromSangiran and Zhoukoudian. LB1’s relative facial heights are similarto those of Dali, but the latter specimen shows a greater FMB/XBPB.The FMB/XBPB of Hexian is comparable to that of LB1, but theformer is largely due to extremely wide XBPB in this specimen.
In Fig. 17DeF, LB1’s relative midfacial breadths (ZYB/XBPB,JUBm/XBPB, ZMB/XBPB) are narrower than H. habilis, TurkanaH. ergaster, and Sangiran H. erectus, and are comparable to Dali. InFig. 17HeL, LB1 also showsmarked reductive trends in the height ofinfraorbital surface (WMH/XBPB), nasal height and breadth(NLHm/XBPB, NLBm/XBPB), and palatal length and breadth (MALL/XBPB, MAB/XBPB), which are most similar to Dali where themeasurements are available for the latter (WMH/XBPB, MAB/XBPB).
Fig. 18 plots other combinations of facial measurements. Fig. 18A(NPH vs. IOMH) suggests that it is the infraorbital portion thatshows marked height reduction in the face of Asian Homo

110 120 130 140 150 160 170SMCB
0
5
10
15
20
CTP
E
h
hh
h
e
eD
KN
G
s
s
s
ss
s
sm
mN
NN
N
NNL
z
z
z
H
Do
110 120 130 140 150 160 170SMCB
0
5
10
15
CTF
E
h
h
h
K
s
s
s
mm
N
N
N
N
N
L
H(0.07)
(0.05)
(0.03)
(0.1)
(0.05)
110 120 130 140 150 160 170SMCB
5
10
15
20
25
CTA
S*
h
hh
e oD
KN
G
ss
ss
s
s
sm
m
NNN
N
NN
L
zzz
H
(0.12)
(0.1)
(0.08)
(0.15)
(0.1)
110 120 130 140 150 160 170SMCB
5
10
15
20
25
30
CTO
P*
hh
h
e
e
o
D
s s
s
s s
sm
m
N
N
N
N
N
N
L z
z
zH
D
A B C
D E F
110 120 130 140 150 160 170SMCB
0
5
10
15
CTB
R*
hh h
h
e
D
G s
ss
ss s
m
mN
NN
N
NNzz
z
H
DKo
s
110 120 130 140 150 160 170SMCB
0
5
10
15
CTL
A**
hh
h
h
e
eD
s
s
s s
s
ss
m mN
NN
NN
L
HK
(0.08)
(0.06)
(0.04)
s
s
s s
(0.07)
(0.05)
(0.03)
L
Figure 15. Scatterplots of cranial bone thicknesses. Metric codes: CTFE ¼ frontal eminence thickness; CTBR ¼ bregma thickness; CTPE ¼ parietal eminence thickness;CTLA ¼ lambda thickness; CTAS ¼ asterion thickness; CTOP ¼ opisthocranion thickness; SMCB ¼ supramastoid breadth. CTBR for LB1 is from Brown et al. (2004). “*” indicates thatthe value for LB1 is an estimate. “**” may be affected by the trauma-like depression slightly. Note that the great CTPE of ZKD 11 (16 mm) reported in Weidenreich (1943) is too greatand is likely a misprint and is thus excluded from the plot here. Symbols and color codes (online version only) as in Fig. 12. (For interpretation of the references to color in this figurelegend, the reader is referred to the web version of this article.)
Y. Kaifu et al. / Journal of Human Evolution 61 (2011) 644e682666
compared to earlier African Homo. LB1 follows this pattern andshows a similar IOMH/NPH to that of Nanjing.
The degree of facial prognathism is here examined by twodifferent measurements, porionenasioneprosthion angle (PNPA)and facial profile angle (FPFA) as plotted in Fig. 18B. In both angles,a higher value means a more prognathic condition. LB1 is moreprognathic than KNM-ER 1470, KNM-ER 3733 and Kabwe, and iscomparable or less prognathic compared to S 17 and KNM-ER 1813.Although caution is needed because of potential errors in recon-struction of the facial orientation in some specimens (KNM-ER1470, S 17; Wood, 1991; Antón, 2003), the above comparisonsbased on a small sample suggest that LB1 is as prognathic as theseearlier Homo taxa. Prognathic faces similar to the LB1 condition arealso found in Bodo, as well as subadult individuals of D2700(Dmanisi) and KNM-WT 15000 (H. ergaster). Potentially deformedor fragmentary specimens of OH 24 and KNM-ER 3732 (H. habilis)and 3883 (H. ergaster) appear to have exhibited strongprognathism.
In the supraorbital torus measurements in Fig. 18C, the LB1torus’ relative thickness value at the midorbital level (SOTT3/SOTB)is comparable to those of H. habilis, SK 847, and KNM-ER 3733 (andD2282 which is not included in this plot due to the lack of SOTB),but these latter specimens do not show the distinct lateral torusthickening (SOTT5 >> SOTT3: Fig. 18D) that characterizes chro-nologically later Javanese H. erectus and some of the Zhoukoudiancrania, and is observed in LB1. Above the midorbital point, theanteroposterior width of the supratoral plane (SOTL3) of LB1 is
moderate when compared to GOL (Fig. 18E). Early JavaneseH. erectus and H. ergaster are distinct in this respect, showingremarkably protruded tori.
The orbital height (OBHm) of LB1 (32 mm) is not the smallest inour comparative sample, but is comparable or slightly higher thanmost of the 2.0e1.7 Ma crania (KNM-ER 1813, 3732; OH 24; D3444)and Nanjing 1 (Fig. 18F). In contrast, LB1’s orbital breadth (OBBm,33 mm) is the narrowest among our comparative specimens. Thus,the orbit of LB1 is relatively high and narrow. An adolescent EastAfrican H. ergaster (KNM-WT 15000) exhibits a similarly narroworbit (not shown in the plot), but this is not a typical observation inthe adult African specimens. Among the adult comparative speci-mens, a narrow orbit closest to the condition of LB1 is seen in one ofthe Asian specimens, Maba.
In Fig. 18GeI, the nasal height (NLHm) of LB1 occupiesa substantial proportion of its NPH, while the nasal breadth (NLBm)is moderate relative to its NLHm and external palatal breadth (MAB).Some post-1.8 Ma African Homo (KNM-ER 3733, Bodo) exhibitextremely wide nasal aperture but this is not the case with LB1.
In the plot of the minimum malar height (WMH) vs. infraorbitalsurface angle (IOFA) (Fig. 18J), the derived, markedly gracile,inferiorly-facing infraorbital surfaces closer to the LB1 condition arefound among the middle Pleistocene specimens from Africa(Kabwe) and Asia (Dali, and probably Zhoukoudian). In contrast tothis, an extensive, vertical or anteroinferiorly sloping infraorbitalsurface is the primitive condition for Homo (Bilsborough andWood,1988).

Figure 16. A diagonal CT section to show the asterion thickness (CTAS) of LB1. Thecorresponding structures between the surface rendered image and the CT section areindicated by the thin arrows. Symbols: ast ¼ asterion; b ¼ a break on the bone surface;S ¼ transverse/sigmoid sinus. Scale ¼ 10 mm.
Y. Kaifu et al. / Journal of Human Evolution 61 (2011) 644e682 667
Fig. 18K shows that the maxilloalveolar length (MALL) of LB1 ismarkedly longer compared to its narrow breadth (MAB). The palateis shallow (palatal height, PATH) and LB1 is more similar to Sangiranand Dmanisi rather than African Homo specimens except for SK 847in this respect (Fig. 18L).
Multivariate analyses with pre-modern Homo
Overall size Comparison of the size variable [SV] in Fig. 19replicates the reported fact that the LB1 neurocranium is smallerthan any known fossil Homo including H. habilis (Brown et al.,2004). LB1 is slightly smaller than the smallest H. habilisspecimen (KNM-ER 1813) in this comparison.Principal component analysis Component loadings of the PCA forthe pre-modernHomo sample are shown in Table 3. The state of LB1in each PC was assessed with reference to these and the originalsize-adjusted variables used in the PCA (Fig. 20). The scores forthe first four PCs, which showed some significant regional ortemporal variation within the comparative fossil sample areplotted with SV in Fig. 19. These plots include least-squareregression lines for the comparatively large, Javanese H. erectussubsample and its 95% prediction ranges.
We first compare the PC scores (Y-axes in Fig. 19) directlywithout considering the possible effect of cranial size (X-axes inFig. 19). PC1 effectively separates H. habilis, D2280, H. ergaster, andKabwe, as well as many of the Chinese fossils from JavaneseH. erectus. The comparatively high PC1 score of LB1, which can beascribed particularly to its great SQSB/SV and smaller SOTB/SV, is inthe middle of the cloud of Javanese H. erectus (Sangiran, Sam-bungmacan, and Ngandong). This observation is supported by theabove bivariate analyses base on larger samples (Fig. 12DeG).
The PC2 tends to distinguish H. habilis, Dmanisi/Turkana/San-giran/Zhoukoudian, and post-1.5 Ma African Homo/Ngandong from
each other. LB1 shows a high value (0.068), mainly because of itssmaller POBB/SV and great SMCB/SV, which is closest to KNM-ER1813 (0.061), S 2 (0.061), Bukuran (0.063), and ZKD 12 (0.055).This observation is consistent with the above bivariate comparisonsbased on larger samples (Fig. 12A, B, and H).
In PC3, the small GOL/SV of LB1 places its score remote fromDmanisi, Sangiran and Zhoukoudian, and within the ranges ofvariation exhibited by H. habilis, H. ergaster, Sambungmacan, andNgandong (as well as Hexian). In PC4, LB1 is closer to H. habilis,Sambungmacan and Ngandong rather than to Sangiran H. erectusand African H. ergaster.
When the possible effect of cranial size is taken into consider-ation in the form of 95% prediction range associated with SV, thescores of LB1 are within the range calculated for Javanese H. erectusin all four PCs (Fig. 19). Whether the cranial shape differencebetween LB1 and other fossil groups (H. habilis, Dmanisi,H. ergaster,etc.) can be explained as size-related variation is difficult toexamine given the small available fossils samples. At least, in regardto PC1, the considerable difference between similarly small-sizedLB1 and KNM-ER 1813 suggests that this PC differs betweenH. habilis andH. floresiensis irrelevant from the cranial size variation.Cluster analysis The cluster trees in Fig. 21 integrate the abovetendencies in PCs1e4 as well as all the PCs (PCs 1e8). The 8 PCanalyses reflect overall neurocranial similarities including allaspects of between-group and within-group variations, while the4 PC analyses, which are based on those PCs showing significantregional/chronological variations, focus more on between-groupvariation. The details of the topology vary among the analyses,but LB1 consistently clusters with Sm 3, Hexian, and the fourSangiran specimens as well as ZKD 11 in all the four trees.
Thus, PCA indicates that the neurocranial shape of LB1 is mostsimilar to those of Sangiran H. erectus, Sm 3, and Hexian from Asia,and it is reasonably predicted as an extremely small-sized spec-imen of Javanese H. erectus.
Comparisons with H. sapiens
Univariate comparisons Z-scores of LB1 relative to the Howell’sworldwide H. sapiens sample (the deviation of LB1 from theH. sapiens general mean [N ¼ 2524] divided by the H. sapiensgeneral standard deviation) are shown in Table 4. LB1 is markedlysmaller than the average H. sapiens condition in all theneurocranial linear measurements (Z-scores � �2.1), particularlyin BBH (�5.9) and OCC (�5.7) which are associated with vaultheight. The small Z-score in PAC (�4.8) may also be a reflection ofthis characteristic. Cranial length (GOL: �4.7) and frontal breadth(XFB: �4.6) are also markedly smaller compared to the basal(AUB), mid- (XCB), and posterior (ASB) cranial breadths. Theoccipital bone of LB1 shows strong flexion compared to H. sapiens(OCA: �2.3).
The Z-scores of LB1 for overall facial height (NPH) and breadths(ZYB, FMB, ZMB) vary from �2.2 to �4.7. These are largelycomparable to the Z-scores for the neurocranial measurements,indicating that the facial skeleton of LB1 is as reduced as those inH. sapiens in terms of its relative size to the neurocranium. Thesmall facial size of LB1 is particularly apparent in some breadthmeasurements (FMB, ZMB, OBBm, and MAB: Z-scores � �2.9), butthe deviations in facial height (NPH: �2.2) and the degree ofprognathism (BPL:�2.0) are more moderate, and the orbital heightis only slightly less than in H. sapiens (OBHm: �0.7).
When aspects of craniofacial shape are examined by shapeindices, LB1’s BPL/BNL and OBHm/OBBm are within the upper2.5% of the H. sapiens variations; LB1’s BBH/XCB and XFB/XCB arewithin the lower 0.02% of the H. sapiens variations; LB1’s OCC/OCA is outside (lower than) the entire range of the H. sapiens
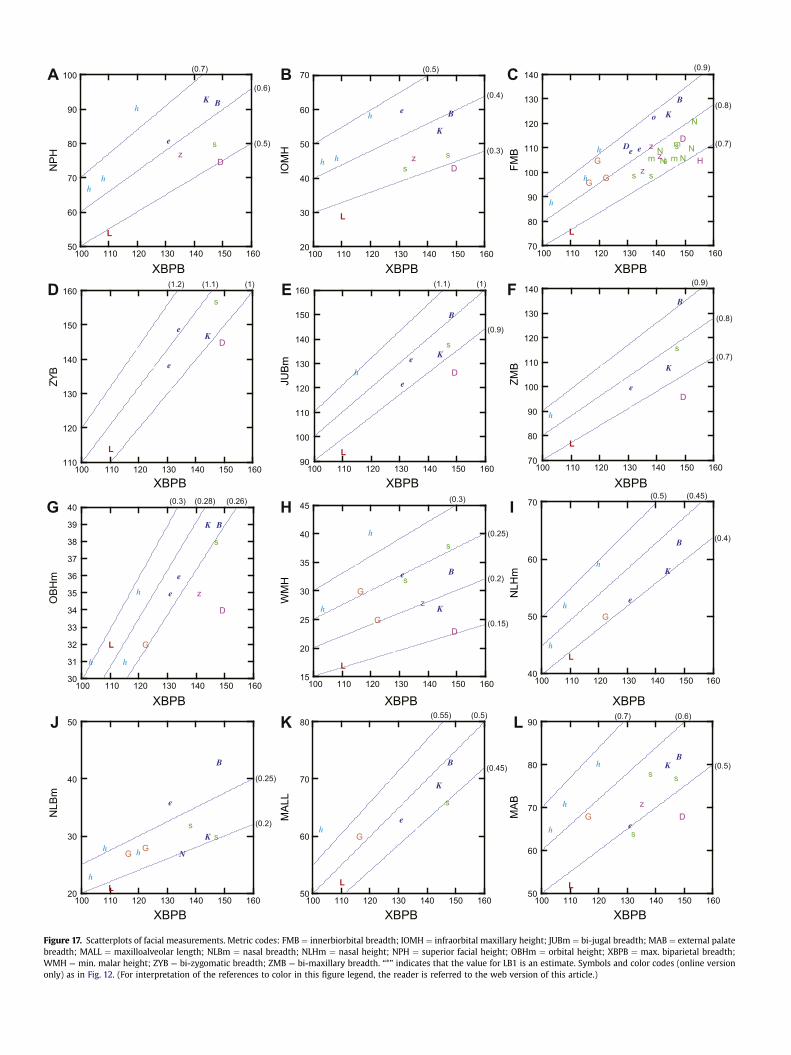
100 110 120 130 140 150 160
XBPB
15
20
25
30
35
40
45
WM
H
h
h
e B
K
G
G
s
s
L
z
D
100 110 120 130 140 150 160
XBPB
70
80
90
100
110
120
130
140
FMB h
h
h
e e
o
D
B
K
G
G G s
s
s
s
mm
m
NNN
N
N
L
z
z
z H
D
100 110 120 130 140 150 160
XBPB
110
120
130
140
150
160
ZYB e
eK
s
L
D
100 110 120 130 140 150 160
XBPB
70
80
90
100
110
120
130
140
ZMB
h
e
B
K
s
L
D
100 110 120 130 140 150 160
XBPB
50
60
70
80
MAL
L
he
B
K
G
s
L
100 110 120 130 140 150 160
XBPB
20
30
40
50
NLB
m
h
h
h
e
B
K
NG G
ss
L
100 110 120 130 140 150 160
XBPB
40
50
60
70N
LHm
h
h
he
B
K
G
L
100 110 120 130 140 150 160
XBPB
90
100
110
120
130
140
150
160JU
Bm he
e
B
Ks
L
D
100 110 120 130 140 150 160
XBPB
50
60
70
80
90
MAB
h
h
h
e
BK
G
s s
s
L
zD
100 110 120 130 140 150 160
XBPB
30
31
32
33
34
35
36
37
38
39
40
OBH
m
h
h h
e
e
BK
G
s
L
z
D
(0.3) (0.28) (0.26)
100 110 120 130 140 150 160
XBPB
20
30
40
50
60
70
IOM
H
h
h h
e B
K
ss
L
zD
(0.5)
(0.4)
(0.3)
100 110 120 130 140 150 160
XBPB
50
60
70
80
90
100
NPH
h
hh
e
BK
s
L
z
(0.7)
(0.6)
(0.5)
D
(0.25)
(0.2)
(0.15)
(0.3)
(0.9)
(0.8)
(0.7)
(1.2) (1.1) (1)
(0.25)
(0.2)
(0.55) (0.5)
(0.45)
(0.7) (0.6)
(0.5)
(1.1) (1)
(0.9)
(0.9)
(0.8)
(0.7)
(0.5) (0.45)
(0.4)
A B C
D E F
G H I
J K L
Figure 17. Scatterplots of facial measurements. Metric codes: FMB ¼ innerbiorbital breadth; IOMH ¼ infraorbital maxillary height; JUBm ¼ bi-jugal breadth; MAB ¼ external palatebreadth; MALL ¼ maxilloalveolar length; NLBm ¼ nasal breadth; NLHm ¼ nasal height; NPH ¼ superior facial height; OBHm ¼ orbital height; XBPB ¼ max. biparietal breadth;WMH ¼ min. malar height; ZYB ¼ bi-zygomatic breadth; ZMB ¼ bi-maxillary breadth. “*” indicates that the value for LB1 is an estimate. Symbols and color codes (online versiononly) as in Fig. 12. (For interpretation of the references to color in this figure legend, the reader is referred to the web version of this article.)

30 40 50 60OBBm
30
31
32
33
34
35
36
37
38
39
40
OBH
m
h hh
e
e
BK
G
s
L
z
N
D
M
jh
(1) (0.9) (0.8) (0.7)
120 160 200 240GOL
15
20
25
30
35SO
TL3
hh
ee
o
D
K
G
s
s
ss
m
m
mN
N
N
N
N
N
L
z
z
zH
D
5 10 15 20 25SOTT3
0
5
10
15
20
SOTT
5
h
hh
h
eew
D
B K S
Gs
s
s
mmm
NN
N
N
L
z
zz z
N
HD
M
o
(1.2) (1) (0.8)
(1.4)
(1.2)
(1)
50 60 70 80 90 100NPH
20
30
40
50
60
70
IOM
H
h
h hh
e
w
B
K
s
L
z
N
(0.6)
(0.5)
(0.7)
15 20 25 30 35 40 45WMH
70
80
90
100
110
IOFA
h
h
h
e
K
s
L
D
20 30 40 50NLBm
40
50
60
70
NLH
m
h
h
he
B
K
G
L
(2.5) (2) (1.5)
50 60 70 80 90MAB
20
30
40
50
NLB
m
h
h
hh
e
B
K
G
ss
L
(0.5)
(0.4)
(0.3)
50 60 70 80 90 100NPH
40
50
60
70
NLH
m
h
h
he
w
B
K
L
(0.8) (0.7)
(0.6)
50 60 70 80 90MAB
50
60
70
80
MAL
L
hh
ew
B
K
G
s
L
(1) (0.9)
(0.8)
50 60 70 80 90MAB
8
10
12
14
16
18
20
22
PATH
h
h
h
h
e
e
w
B
K
Gs
s
L
(0.25)
(0.2)
(0.15)
A B C
D E F
G H I
J K L
D
70 80 90 100PNPA
95
100
105
110
FPFA
h
h
e
K
sL
80 90 100 110 120 130 140SOTB
5
10
15
20
SOTT
3
hh
h
h
e
e
w
oD
B
K
Gs
sss
m
mm
N
N
N
N
L
z
z
z
z
N
H D
M
(0.12)
(0.1)
(0.08)
(0.06)
Figure 18. Scatterplots of facial measurements. Metric codes: GOL ¼ max. cranial length; IOFA ¼ infraorbital surface angle; IOMH ¼ infraorbital maxillary height; MAB ¼ externalpalate breadth; MALL ¼ maxilloalveolar length; NLBm ¼ nasal breadth; NLHm ¼ nasal height; NPH ¼ superior facial height; OBBm ¼ orbital breadth; OBHm ¼ orbital height;PRRD ¼ prosthion radius; SOTB ¼ supraorbital torus breadth; SOTL3 ¼ supraorbital torus length (midorbit); SOTT3 ¼ supraorbital torus thickness (midorbit); SOTT5 ¼ supraorbitaltorus thickness (lateral); WMH ¼ min. malar height. “*” indicates that the value for LB1 is an estimate. Symbols and color codes (online version only) as in Fig. 12 except for w ¼ SK847. (For interpretation of the references to color in this figure legend, the reader is referred to the web version of this article.)
Y. Kaifu et al. / Journal of Human Evolution 61 (2011) 644e682 669

90 100 110 120 130 140 150 160SV
-0.2
-0.1
0.0
0.1
0.2
PC1
hh
eeo
D K
G
s
ss
mm
m
NN NNN
z
z
z
H
D
90 100 110 120 130 140 150 160SV
-0.10
-0.05
0.00
0.05
0.10
PC3
h
h ee
o
D
KG
s
s
ss
m
mm
N
NN
N
N
zzz
H
D
90 100 110 120 130 140 150 160SV
-0.10
-0.05
0.00
0.05
0.10
PC4
h
he
e
o
D
K
G
ss
ss
mm
m
N
N
N
NN
z
z
z
H
D
s
ee
o
D
H
90 100 110 120 130 140 150 160SV
-0.15
-0.10
-0.05
0.00
0.05
0.10
0.15
PC2
h
h
K
G
ss
s
s
m
m
m
N
NN
NN
z
zz
D
L
L
L
L
Figure 19. PC scores plotted with the cranial size variable (SV). Least-square regression lines (solid lines) and 95% prediction ranges (dotted lines) for the Javanese H. erectus sampleare shown. Symbols and color codes (online version only) as in Fig. 13. (For interpretation of the references to color in this figure legend, the reader is referred to the web version ofthis article.)
Y. Kaifu et al. / Journal of Human Evolution 61 (2011) 644e682670
variation. As a result, in BPL/BBH and OBHm/BBHm, which arecombinations of facial and a neurocranial measurements, LB1exhibits larger values (96% and 36%, respectively) than anyH. sapiens individuals recorded by Howells (57e94% and20e33%, respectively). When a PCA is conducted based ona correlation matrix of the above 7 indices, the PC1 whichexplained 29% of the total variance clearly separates LB1 (PC1score ¼ 6.3) from all the H. sapiens specimens (PC1 scores varybetween �3.3 and 3.5). This means that the above combinationof cranial shape indices is unique to LB1 and markedly differentfrom H. sapiens. On the other hand, LB1’s NPH/XCB, ZYB/XCB, andFMB/XCB are well within the variations exhibited by theH. sapiens sample.
Table 3Component loadings of the major PCs of the neurocranial PCA.a
with pre-modern Homo with H. sapiens
PC1 PC2 PC3 PC4 PC1
GOL/SV 0.173 �0.196 0.817 0.215 0.633SOTB/SV �0.721 �0.165 �0.376 0.533 0.796POBB/SV 0.419 �0.611 �0.554 �0.248 0.024SQSB/SV 0.799 0.172 �0.116 0.052 0.153ASB/SV 0.704 0.391 �0.177 0.478 0.847BRAB/SV �0.462 0.775 �0.141 �0.288 0.903SMCB/SV 0.388 0.796 �0.263 0.054 0.882PBRH/SV �0.441 �0.195 �0.304 �0.399 �0.958Proportion 32 27 16 12 62Cum. proportion 32 59 75 87 62
a Statistically significant loadings (p < 0.05, t-test) are in boldface.
Neurocranial PCA In the PCA based on the eight neurocranialmeasurements (Table 3), PC1, which explains 62% of the totalvariation, distinctly separates the pre-modern and modern Homospecimens (Fig. 22A), whereas no clear differences between thetwo groups were observed in the other PCs. Its high, positivecorrelations with GOL, SOTB, ASB, BRAB, and SMCB, and high,negative correlations with PBRH indicate that PC1 mainly reflectsrelative neurocranial height (Table 3, right column). LB1 groupsclearly with the pre-modern Homo in this PC. In Fig. 22A, theentire H. sapiens sample shows positive correlation between PC1and SV (0.308, P < 0.01). LB1 is plotted completely outside the95% prediction range for the H. sapiens sample.Basicranial PCA The results of the PCA based on seven basicranialmeasurements that represent positional (anteroposterior andtransverse) relationships of the oval foramina, carotid canals, andjugular foramina, as well as size and shape of the oval foramen aregiven in Table 5 and Fig. 22B. The PC scores indicated that PC1 andPC3, when combined, distinctly separate late Javanese H. erectus(Sm 4, Ng 7, Ng 12), Turkana H. ergaster (KNM-ER 3733, 3883),and LB1 from the sample of 197 modern human specimens(Fig. 22B). The component loadings in Table 4 indicate that theformer pre-modern Homo group is characterized by relativelygreater anteroposterior separation between the oval foramen andcarotid canal/jugular foramen (OFeCC and OFeJF in PC1),relatively narrow breadths across these foramina (OFeOF, CCeCC,and JFeJF in PC1), and less elongated or rounded shape of theoval foramen (OFD1 and OFD2 in PC1 and PC3). OH 9 is similar tothis fossil group in PC1 but not in PC3, and Kabwe resemblesmodern humans in these respects.

Figure 20. Plots of the size-adjusted variables used in the PCAwith pre-modern Homo. Symbols: D ¼ Dali; D ¼ Daka; e ¼ KNM-ER 3733, 3883; G ¼ D2280; H ¼ Hexian; h ¼ KNM-ER1470, 1813; K ¼ Kabwe; L ¼ LB1; m ¼ Sm 1, 3, 4; N ¼ Ng 6, 7, 10, 11, 12; o ¼ OH 9; s ¼ S 2, 17, IX, Bukuran; z ¼ ZKD 10, 11, 12. Color codes (online version only) as in Fig. 12. (Forinterpretation of the references to color in this figure legend, the reader is referred to the web version of this article.)
Y. Kaifu et al. / Journal of Human Evolution 61 (2011) 644e682 671
Discussion
Cranial morphology of H. floresiensis as compared to H. sapiens
In these metric analyses, the LB1 cranium was comparedto modern humans using three different samples and analyses.Two of these analyses are PCAs focusing on neurocranium and
basicranium, and included Early-Middle Pleistocene Homo speci-mens and a heterogeneous H. sapiens sample including small-bodied populations. The neurocranial PCA effectively confirmedthe previous observation that the pre-modern Homo group isdistinct from H. sapiens primarily in cranial vault height (Day andStringer, 1982; Lieberman et al., 2002; Pearson, 2008). LB1 clearlybelonged to the pre-modern Homo group in this analysis, and its
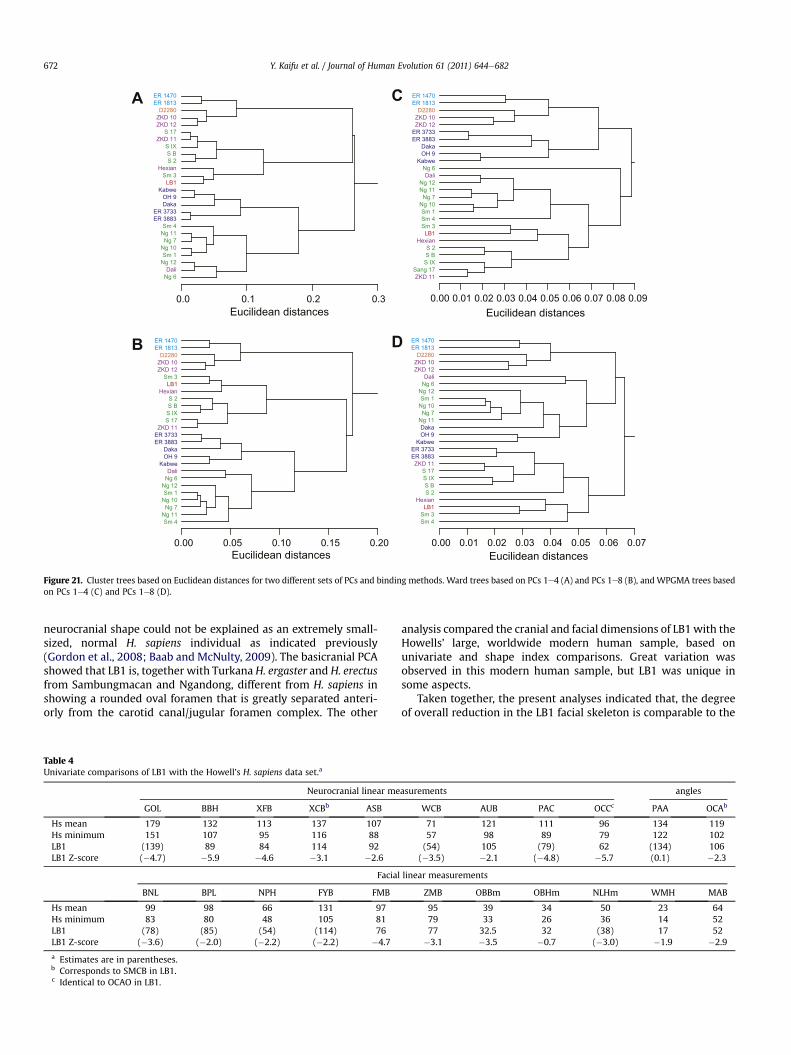
Eucilidean distances
ER 1470ER 1813
D2280ZKD 10ZKD 12
Sm 3LB1
HexianS 2S B
S IXS 17
ZKD 11ER 3733ER 3883
DakaOH 9
KabweDali
Ng 6Ng 12Sm 1
Ng 10Ng 7
Ng 11Sm 4
ER 1470ER 1813
D2280ZKD 10ZKD 12
S 17ZKD 11
S IXS BS 2
HexianSm 3LB1
KabweOH 9Daka
ER 3733ER 3883
Sm 4Ng 11Ng 7
Ng 10Sm 1
Ng 12Dali
Ng 6
Eucilidean distances0.0 0.1 0.2 0.3
0.00 0.05 0.10 0.15 0.20
ER 1470ER 1813
D2280ZKD 10ZKD 12
DaliNg 6
Ng 12Sm 1
Ng 10Ng 7
Ng 11DakaOH 9
KabweER 3733ER 3883ZKD 11
S 17S IXS BS 2
HexianLB1
Sm 3Sm 4
A
B
0.00 0.01 0.02 0.03 0.04 0.05 0.06 0.07 0.08 0.09
ER 1470ER 1813
D2280ZKD 10ZKD 12
ER 3733ER 3883
DakaOH 9
KabweNg 6Dali
Ng 12Ng 11Ng 7
Ng 10Sm 1Sm 4Sm 3LB1
HexianS 2S B
S IXSang 17ZKD 11
0.00 0.01 0.02 0.03 0.04 0.05 0.06 0.07
Eucilidean distances
Eucilidean distances
C
D
Figure 21. Cluster trees based on Euclidean distances for two different sets of PCs and binding methods. Ward trees based on PCs 1e4 (A) and PCs 1e8 (B), and WPGMA trees basedon PCs 1e4 (C) and PCs 1e8 (D).
Y. Kaifu et al. / Journal of Human Evolution 61 (2011) 644e682672
neurocranial shape could not be explained as an extremely small-sized, normal H. sapiens individual as indicated previously(Gordon et al., 2008; Baab and McNulty, 2009). The basicranial PCAshowed that LB1 is, together with Turkana H. ergaster and H. erectusfrom Sambungmacan and Ngandong, different from H. sapiens inshowing a rounded oval foramen that is greatly separated anteri-orly from the carotid canal/jugular foramen complex. The other
Table 4Univariate comparisons of LB1 with the Howell’s H. sapiens data set.a
Neurocranial linear me
GOL BBH XFB XCBb ASB
Hs mean 179 132 113 137 107Hs minimum 151 107 95 116 88LB1 (139) 89 84 114 92LB1 Z-score (�4.7) �5.9 �4.6 �3.1 �2.6
Facial
BNL BPL NPH FYB FMB
Hs mean 99 98 66 131 97Hs minimum 83 80 48 105 81LB1 (78) (85) (54) (114) 76LB1 Z-score (�3.6) (�2.0) (�2.2) (�2.2) �4.7
a Estimates are in parentheses.b Corresponds to SMCB in LB1.c Identical to OCAO in LB1.
analysis compared the cranial and facial dimensions of LB1with theHowells’ large, worldwide modern human sample, based onunivariate and shape index comparisons. Great variation wasobserved in this modern human sample, but LB1 was unique insome aspects.
Taken together, the present analyses indicated that, the degreeof overall reduction in the LB1 facial skeleton is comparable to the
asurements angles
WCB AUB PAC OCCc PAA OCAb
71 121 111 96 134 11957 98 89 79 122 102(54) 105 (79) 62 (134) 106
(�3.5) �2.1 (�4.8) �5.7 (0.1) �2.3
linear measurements
ZMB OBBm OBHm NLHm WMH MAB
95 39 34 50 23 6479 33 26 36 14 5277 32.5 32 (38) 17 52�3.1 �3.5 �0.7 (�3.0) �1.9 �2.9
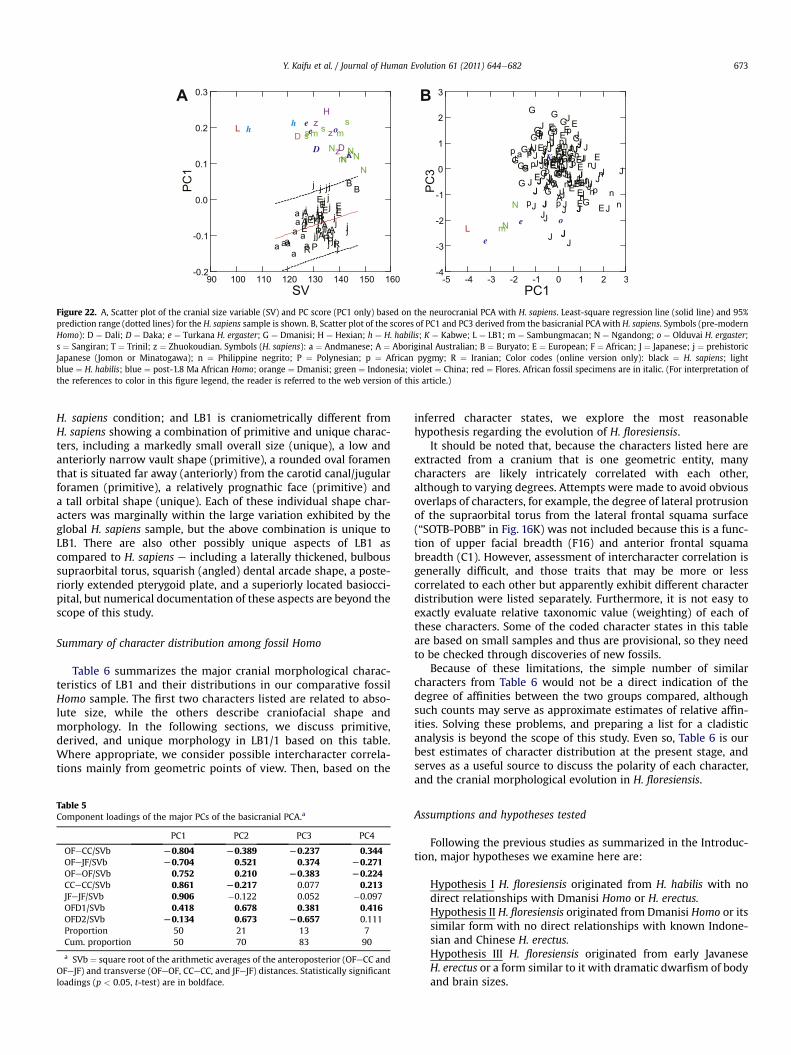
A B
Figure 22. A, Scatter plot of the cranial size variable (SV) and PC score (PC1 only) based on the neurocranial PCA with H. sapiens. Least-square regression line (solid line) and 95%prediction range (dotted lines) for the H. sapiens sample is shown. B, Scatter plot of the scores of PC1 and PC3 derived from the basicranial PCAwith H. sapiens. Symbols (pre-modernHomo): D ¼ Dali; D ¼ Daka; e ¼ Turkana H. ergaster; G ¼ Dmanisi; H ¼ Hexian; h ¼ H. habilis; K ¼ Kabwe; L ¼ LB1; m ¼ Sambungmacan; N ¼ Ngandong; o ¼ Olduvai H. ergaster;s ¼ Sangiran; T ¼ Trinil; z ¼ Zhuokoudian. Symbols (H. sapiens): a ¼ Andmanese; A ¼ Aboriginal Australian; B ¼ Buryato; E ¼ European; F ¼ African; J ¼ Japanese; j ¼ prehistoricJapanese (Jomon or Minatogawa); n ¼ Philippine negrito; P ¼ Polynesian; p ¼ African pygmy; R ¼ Iranian; Color codes (online version only): black ¼ H. sapiens; lightblue ¼ H. habilis; blue ¼ post-1.8 Ma African Homo; orange ¼ Dmanisi; green ¼ Indonesia; violet ¼ China; red ¼ Flores. African fossil specimens are in italic. (For interpretation ofthe references to color in this figure legend, the reader is referred to the web version of this article.)
Y. Kaifu et al. / Journal of Human Evolution 61 (2011) 644e682 673
H. sapiens condition; and LB1 is craniometrically different fromH. sapiens showing a combination of primitive and unique charac-ters, including a markedly small overall size (unique), a low andanteriorly narrow vault shape (primitive), a rounded oval foramenthat is situated far away (anteriorly) from the carotid canal/jugularforamen (primitive), a relatively prognathic face (primitive) anda tall orbital shape (unique). Each of these individual shape char-acters was marginally within the large variation exhibited by theglobal H. sapiens sample, but the above combination is unique toLB1. There are also other possibly unique aspects of LB1 ascompared to H. sapiens e including a laterally thickened, bulboussupraorbital torus, squarish (angled) dental arcade shape, a poste-riorly extended pterygoid plate, and a superiorly located basiocci-pital, but numerical documentation of these aspects are beyond thescope of this study.
Summary of character distribution among fossil Homo
Table 6 summarizes the major cranial morphological charac-teristics of LB1 and their distributions in our comparative fossilHomo sample. The first two characters listed are related to abso-lute size, while the others describe craniofacial shape andmorphology. In the following sections, we discuss primitive,derived, and unique morphology in LB1/1 based on this table.Where appropriate, we consider possible intercharacter correla-tions mainly from geometric points of view. Then, based on the
Table 5Component loadings of the major PCs of the basicranial PCA.a
PC1 PC2 PC3 PC4
OFeCC/SVb L0.804 L0.389 L0.237 0.344OFeJF/SVb L0.704 0.521 0.374 L0.271OFeOF/SVb 0.752 0.210 L0.383 L0.224CCeCC/SVb 0.861 L0.217 0.077 0.213JFeJF/SVb 0.906 �0.122 0.052 �0.097OFD1/SVb 0.418 0.678 0.381 0.416OFD2/SVb L0.134 0.673 L0.657 0.111Proportion 50 21 13 7Cum. proportion 50 70 83 90
a SVb ¼ square root of the arithmetic averages of the anteroposterior (OFeCC andOFeJF) and transverse (OFeOF, CCeCC, and JFeJF) distances. Statistically significantloadings (p < 0.05, t-test) are in boldface.
inferred character states, we explore the most reasonablehypothesis regarding the evolution of H. floresiensis.
It should be noted that, because the characters listed here areextracted from a cranium that is one geometric entity, manycharacters are likely intricately correlated with each other,although to varying degrees. Attempts were made to avoid obviousoverlaps of characters, for example, the degree of lateral protrusionof the supraorbital torus from the lateral frontal squama surface(“SOTB-POBB” in Fig. 16K) was not included because this is a func-tion of upper facial breadth (F16) and anterior frontal squamabreadth (C1). However, assessment of intercharacter correlation isgenerally difficult, and those traits that may be more or lesscorrelated to each other but apparently exhibit different characterdistribution were listed separately. Furthermore, it is not easy toexactly evaluate relative taxonomic value (weighting) of each ofthese characters. Some of the coded character states in this tableare based on small samples and thus are provisional, so they needto be checked through discoveries of new fossils.
Because of these limitations, the simple number of similarcharacters from Table 6 would not be a direct indication of thedegree of affinities between the two groups compared, althoughsuch counts may serve as approximate estimates of relative affin-ities. Solving these problems, and preparing a list for a cladisticanalysis is beyond the scope of this study. Even so, Table 6 is ourbest estimates of character distribution at the present stage, andserves as a useful source to discuss the polarity of each character,and the cranial morphological evolution in H. floresiensis.
Assumptions and hypotheses tested
Following the previous studies as summarized in the Introduc-tion, major hypotheses we examine here are:
Hypothesis I H. floresiensis originated from H. habilis with nodirect relationships with Dmanisi Homo or H. erectus.Hypothesis II H. floresiensis originated from Dmanisi Homo or itssimilar form with no direct relationships with known Indone-sian and Chinese H. erectus.Hypothesis III H. floresiensis originated from early JavaneseH. erectus or a form similar to it with dramatic dwarfism of bodyand brain sizes.

Table 6Major craniofacial characteristics of LB1 and their distribution among comparative.
No. Character state in LB1 (with the opposite state in the parentheses) Character distributiona Hypothesisb
Hh Dm erg KB ST Sm Ng ZN Dal Mb I II III
Overall size (absolute)S1 Cranial vault small (vs. large) : x x x x x x x x x S N NS2 Facial skeleton small (vs. large) x x x x x x x x x x N N NOverall shape of the neurocranium (see also Fig. 15)C1 Anterior frontal squama narrow (vs. wide: Fig. 16H) C C : x C x x C x ? S S SC2 Cranial vault short relative to breadth (vs. long: Fig. 16A) : : x x x : x x x ? S S CC3 Parietals laterally expanded relative to the cranial base (vs. bell-shaped
posterior vault profile: Fig. 16DeF)x x C C C C C C C C? N N S
C4 Suprametal crest depressed medially (vs. protruded laterally: Fig. 16J) x x C C C C C x C ? N N SC5 Cranial vault low relative to breadth (vs. high: Fig. 16B) x x C x C : x : D ? N N SEctocranial keelingsC6 Coronal and sagittal keels poorly developed or absent (vs. well-developed) C x C C : x x x C C? S N SC7 Frontal keel well-developed (vs. poorly developed or absent) x x : D : D D C D ? N N STemporal line and associated surface structuresC8 Temporal line extends posteriorly toward the lamboid suture (vs. limited
posterior extension: Fig. 17C)C C C C : x x :? C ? S S S
C9 Supramastoid crest more horizontally oriented (vs. swings superiorly) C C C C? : x x C C ? S S SC10 Medial incursion of the temporal lines weak on the frontal (vs. marked: Fig. 17A) : C C C C C C C C C S S SC11 Angular torus restricted and weak (vs. large and well-developed) :? C C C : x x x C? ? S S SC12 Temporal line relatively weak on the anterior frontal (vs. marked) : x x x C x x C C x S N SC13 Supramastoid crest poorly developed (vs. well-developed) : x : C x x x x x ? S N NC14 Temporal lines posteriorly divergent on the parietals (vs. medial incursion: Fig. 17B) x x x x : C C C x ? N N SIndividual vault bonesC15 Squamosal suture relatively straight (vs. arched) C C C x C C C C x ? S S SC16 Temporal squama long and parietomastoid suture short (vs. short and long,
respectively: Fig. 17F)C C C x : x x x x ? S S S
C17 Occipital plane gently curved and inclined forward (vs. vertically set) C :? : x C x x C x ? S S? SC18 Parietal rounded (vs. parasagittal flattening present) C x : : x C C x x C S N CC19 Occipital moderately flexed (vs. flexion weaker: Fig. 17I) x : C C C C - - C ? N S SC20 Frontal squama flat on each side with no marked eminence (vs. frontal
eminence distinct)x : : : C x x x x x N S S
C21 Frontal squama strongly curved along the midline (vs. gently curved: Fig. 17D) x x : x x : x : x C? N N CC22 Supratoral sulcus of the occipital straight and continuous (vs. discontinuous) x x? x C C C C C D ? N N? SC23 Low, broad occipital squama (vs. high and narrow: Fig. 17G) x x x x C C C x ? ? N N SC24 Parietal sagittal curvature strong (vs. weak: Fig. 17E) x x x x x : x x x x? N N CCranial baseC25 Nuchal plane strongly convex (vs. flatter) C? C? : x C x x C x? ? S? S? SC26 Root of the pterygoid plate extends posteriorly (vs. posterior extension restricted) C C : x ? C C ? ? ? S S ?C27 Mandibular fossa shallow (vs. deep: Fig. 18D) C C : C C C x x C? ? S S SC28 Postglenoid process present (vs. absent) C C C C C : x C C ? S S SC29 Tympanomastoid fissure not extensive (vs. extensive) C C C C C x x : C ? S S SC30 Postcondyloid tuberocity weak or absent (vs. marked) C C C C C x x ? C ? S S SC31 Midcranial base not extended anteroposteriorly (vs. extended) C? C? C C C x x ? ? ? S? S? SC32 Bony styloid process absent (vs. present) C : x x C C C C ? ? S S SC33 Tympanic tubular along its long axis (vs. flattened) C x : x : C C C x ? S N SC34 Vaginal process absent or ill-defined (vs. well-developed) :? x x x x x x x x ? S? N NC35 Basioccipital flat and located superiorly relative to the surrounding cranial
base structures (vs. located relatively inferiorly)x? ? C x C C C ? C? ? N? ? S
C36 External acoustic meatus medially set (vs. tympanic laterally extensive: Fig. 18F) x C? C C C C C : C ? N S? SC37 Mandibular fossa transversely narrow (vs. wide: Fig. 18E) x ? x x C ? : ? x ? N ? SC38 Anteromedial corner of the mandibular fossa flexed (vs. smoothly concave) x x x x : x x : x ? N N SC39 Mastoid process small, pointed, and medially inclined (vs. large and bulbous
and/or directs directly inferiorly)x x? x x : D x C D ? N N? S
C40 Opisthionic recess incipient (vs. not developed) x x x x? x - - x ? ? N N CC41 Medial and lateral pterypoid plates fused extensively (vs. widely separated) x ? ? x ? ? D ? ? ? N ? ?Cranial bone thicknessC42 Mastoid portion not thickened (vs. thickened: Fig. 19E) C C C D x x x x ? ? S S NC43 Vault bone thick at the bregma and parietal eminence (vs. thin: Fig. 19B and C) x : C C? C C C C C ? N S SFacial size (relative) and morphologyF1 Facial prognathism moderately strong (vs. weak: Fig. 21B) C C? : : C? ? ? ? ? ? S S? S?F2 Supratoral plane with no distinct sulcus (vs. grooved) C : : C C C C x C C S S SF3 Supratoral plane restricted anteroposteriorly (vs. wide: Fig. 21F) C C x x x D C C C C S S CF4 Supraorbital torus (SOT) comparatively thin at the midorbit (vs. thick: Fig. 21D) C : : x x x : x x x S S CF5 Infraorbital surface faces anteriorly (vs. anterolaterally) C C x? x C ? ? ? C ? S S SF6 Maxillary dental arch nearly parallel-sided (vs. parabolic) C C? C x C ? ? x ? ? S S? SF7 Nasal bridge moderately prominent (vs. flattened)c x C? : C C ? ? C C C N S? SF8 Palate shallow (vs. deep: Fig. 21L) x C x x C ? ? ? ? ? N S SF9 Maxillary body protruded forward distinctly beyond the infraorbital surface
with a marked infraorbital sulcus (vs. more smooth junction between thetwo structures)
x D x x : ? ? C C ? N ? S
F10 Lateral end of the SOT bulbous and protrusive laterally (vs. no lateralprojection beyond the frontomalare temporalare)
x x : C : C C C C C N N S
F11 SOT strongly arched (vs. more straight) x x : x x x x x x C N N N
Y. Kaifu et al. / Journal of Human Evolution 61 (2011) 644e682674

Table 6 (continued )
No. Character state in LB1 (with the opposite state in the parentheses) Character distributiona Hypothesisb
Hh Dm erg KB ST Sm Ng ZN Dal Mb I II III
F12 Zygomatic frontal process vertically set in lateral view (vs. inclined anteriorly) x x C C ? ? ? C C C N N ?F13 Infraorbital surface faces slightly inferiorly (vs. vertical or faces superiorly: Fig. 20J) x x C C ? ? ? ? C ? N N ?F14 Zygomaticoalveolar crest straight and diagonally arranged (vs. horizontally set or
with malar incisure)x x C C x ? ? x x ? N N N
F15 Facial height [infraorbital portion] remarkably short (vs. high: Figs. 20A,B and 21A) x x x x D ? ? D? C ? N N ?F16 Upper facial breadth remarkably narrow (vs. wide: Figs. 16G and 20C) x x x x C C D D D C? N N SF17 Maxillary alveolar arch remarkably small (vs. larger: Fig. 20K and L) x x D x : ? ? D? C ? N N SF18 Midfacial breadth remarkably narrow (vs. wide: Fig. 20DeF) x x? x x x ? ? ? C ? N N? NF19 Infraorbital surface remarkably short (vs. high: Fig. 20H) x x x D x ? ? D C ? N N NF20 SOT thickens laterally (vs. thicker medially: Fig. 21E) x x x x x C C : x x N N CF21 Orbit high and narrow (vs. transversely wide: Figs. 20G and 21C) x x x x x x x x x D N N NF22 Palatine posteriorly elongated (vs. no substantial elongation) x x ? x ? ? ? ? ? ? N N ?
a Hh, H. habilis; Dm, Dmanisi; erg, H. ergaster; KB, Kabwe/Bodo; ST, Sangiran/Trinil; Sm, Sambungmacan; Ng, Ngandong; ZN, Zhuokoudian/Nanjing; Dal, Dali; Mb, Maba;C, present; :, present in some specimens; D, not equivalent but close to the LB1 condition; x, not present; ?, not clear; -, present in a more pronounced condition.
b “S” ¼ supported; “N” ¼ not supported; “C” ¼ compatible; “?” ¼ status unknown or unclear.c Based on indirect evidence for LB1.
Y. Kaifu et al. / Journal of Human Evolution 61 (2011) 644e682 675
Hypothesis IV H. floresiensis is not related to any of the abovethree taxa.
Table 6 includes assessments of the first three hypotheses foreach character in four categories (supported, not supported,compatible, or unclear). In interpreting the character distribution inTable 6, we assume that (1) H. habilis is the basal form of the genusHomo (Grine et al., 2009), (2) the Indonesian early and lateH. erectus (Sangiran/Trinil, Sambungmacan, Ngandong) belong toan anagenetically evolving group (Kaifu et al., 2008), (3) DmanisiHomo is close to the basal group that evolved into H. erectus and/orH. ergaster but is derived relative to H. habilis (Martinón-Torreset al., 2008; Rightmire and Lordkipanidze, 2009). Under theseassumptions, for example, if a certain characteristic is found inH. habilis, Dmanisi, and late Javanese H. erectus but not in earlyJavanese H. erectus, the state in late Javanese H. erectus is anexample of evolutionary reversal. If LB1 shares such a characteristic,it may be plesiomorphic retention from H. habilis or Dmanisi(Hypotheses I and II are supported), evidence for its link with lateJavaneseH. erectus (Hypothesis IV is supported), or otherwise it wasacquired secondarily in the lineage of H. floresiensis (compatiblewith Hypothesis III).
LB1 and H. habilis
Of the 67 characters of LB1 listed in Table 6, 31 are shared withH. habilis. All of them are potentially supportive evidence that linkH. habilis and LB1, but 19 of them are also shared with H. habilis,Dmanisi Homo, and early Javanese H. erectus and thus could bederived from any of these groups as plesiomorphic retentions (C1,8e11, 15e17, 25, 27e32; F1, 2, 5, 6). We infer that one additionalcharacter, a posteriorly extended pterygoid plate root (C26), can beadded to the above list. Although the state of this character in earlyJavanese H. erectus is unknown, it is present in chronologically lategroups from the region.
Among the remaining 11 characters that support Hypothesis I, 8are not shared by early Javanese H. erectus. Four of these 8 char-acters are observed in late Javanese H. erectus and thus could occuras a result of evolutionary reversal (compatiblewith Hypothesis III).These are a relatively short cranial length (C2), rounded parietalsurfaces (C18), an anteroposteriorly short supratoral plane (F3), anda thin middle part of the supraorbital torus (F4). Three of theremaining four, a small cranial vault size (S1), and the poor devel-opment of the supramastoid crest (C13) and vaginal process (C34)were found only within H. habilis (S1, C34) or African Homo in
general (C13). These can be used as supportive evidence forHypothesis I. The last one, comparatively thin cranial bone at theasterion (C42), is observed in African and Georgian Homo only, andis a character which possibly links H. habilis or Dmanisi HomowithH. floresiensis.
The remaining three of the 11 characters are shared withH. habilis and early Javanese H. erectus but are not present inDmanisi. These are poor or no development of the coronal andsagittal keels (C6), the weak development of temporal lines on theanterior frontal (C12), and a tubular tympanic plate (C33).
In summary, we found three cranial traits of LB1 (S1, C13, C34)that support Hypothesis I but not Hypotheses II and III. Anothercharacter (C42) supports Hypotheses I and II but not Hypothesis III.
LB1 and Dmanisi
In Table 6, six characteristics of LB1 were present in Dmanisi butnot in H. habilis, including a moderately flexed occipital (C19),a frontal squama flattened on each side with no marked eminence(C20), a medially set external acoustic meatus (C36), thickersuperior vault bones (C43), a moderately prominent nasal bridge(F7), and a shallow palate (F8). However, all of these are sharedwith early Javanese H. erectus and do not exclusively support theHypothesis II. The character C42 (comparatively thin cranial bone atthe asterion) supports Hypotheses I and II with the exclusion ofHypothesis III, as mentioned above.
LB1 and early Javanese H. erectus (cranial vault)
The PC1 of the PCA (without H. sapiens) demonstrated that LB1has a suite of cranial shape characteristics of H. erectus/ergaster andlater taxa, which effectively distinguish them from H. habilis(Fig. 14). Table 6 lists the major contributing factor to this distinc-tion, i.e., a laterally expanded braincase measured on the parietals(C3), as well as many other characteristics which LB1 shares with atleast some of the H. erectus/ergaster groups but not with H. habilisand Dmanisi Homo (C4, 5, 7, 14, 21e24, 38e40; F9e14, 16, 17, 20:a total of 21 characters). All of these can be used to challengeHypotheses I and II, unless the same morphology was acquiredindependently in LB1. The states of two other characteristics (C35,37) are not clear for Dmanisi, but these at least support HypothesisIII and not Hypothesis I.
Among the 21 (or possibly 23) traits that contradict withHypotheses I and II, 12 involve the morphology of the neuro-cranium and the remaining 9 are related to face. Nine of the 12

Y. Kaifu et al. / Journal of Human Evolution 61 (2011) 644e682676
neurocranial characters are present in early Javanese H. erectus, andthus support it. These are related to overall cranial vault shape(C3e5), frontal keel (C7), arrangement of the temporal lines (C14),shape and structure on the occiput (C22, 23), and features of thebasicranium (C38, 39). The character distribution in Table 6suggests that 4 of these 9 traits (C14, 23, 38, 39) are restricted toall or some H. erectus in Asia, and one of these (C23) is unique toearly Javanese H. erectus. Furthermore, our cluster analyses basedon the 8-variable PCA indicated that, in the overall neurocranialshape which reflects C1e5 of Table 6, LB1 is similar to Asian,particularly early Javanese H. erectus (Sm 3, Hexian, four SangiranH. erectus specimens, and ZKD 11). The remaining 3 of the 12characters, which are not observed in early Javanese H. erectus,include a strong midsagittal curvature of the frontal (C21) andparietal (C24), which may be associated with the LB1’s short craniallength (C2), as well as opisthionic recess (C40). All of these arefound among the chronologically later Javanese H. erectus, and thuscan occur independently in the lineage leading to H. floresiensisunder Hypothesis III. In particular, if opisthionic recess is uniquedevelopment in Javanese H. erectus within the fossil Homo groupscompared here, its incipient appearance in LB1 would be a strongsupport for Hypothesis III.
Among the 9 facial characters that contradict with Hypotheses Iand II, four (F9,10,16,17) support, one (F20) is compatiblewith, andanother one (F11) does not support Hypothesis III. The characterstates in the remaining three facial characters (F12e14) are notclear for part or all of the Javanese H. erectus sample.
Thus, we found 12 neurocranial and 5 facial characters of LB1that are supportive or at least compatible with Hypothesis III but donot support Hypotheses I and II (C3e5 [in combinationwith C1 andC2], 7, 14, 21e24, 38e40; F9, 10, 16, 17, 20).
Facial gracilization in LB1
Brown et al. (2004:1057) noted that “[The] face of LB1 lacksmost of themasticatory adaptations evident in Australopithecus andits overall morphology is similar to members of the genus Homo.”Our analyses of metric and morphological comparisons providefurther detailed evidence for this statement.
First, the face of LB1 is small both absolutely and relatively to itsneurocranial size in all dimensions except for the orbital height.This aspect is summarized as the characters F15e19 in Table 6.Combined with its small tooth crown size (Brown et al., 2004;Brown and Maeda, 2009), this substantial facial size reduction inLB1 probably reflects decreasedmasticatory load in H. floresiensis ascompared to the condition in earlierHomo. Presently, the upper andmidfacial morphology of H. floresiensis is known only from LB1/1,but the presence of another mandible (LB6/1), which is slightlysmaller than the LB1 mandible (Brown and Maeda, 2009) suggeststhat a reduced face was a common characteristic of this extinctspecies in Flores.
The existing fossil materials suggest that, during the course ofHomo evolution, marked facial reduction occurred first in the earlyPleistocene H. erectus of Asia (Kaifu et al., 2011). The early Pleisto-cene H. erectus from Java and the earlier middle PleistoceneH. erectus from northern China both exhibit absolutely and rela-tively narrower upper facial breadths [FMB, SOTB] and lower facialheights [NPH, IOMH] compared to their African contemporaries.Although Sangiran H. erectus retains a primitive broad midface(F18) and vertically extensive infraorbital plate (F19), and both theSangiran and Zhoukoudian H. erectus are slightly larger than LB1 inoverall relative facial size (F15e17), a further progressive, reducedfacial skeleton, similar to LB1 is found in Dali from the later middlePleistocene of China. Thus, one parsimonious explanation for themarkedly reduced face of LB1 is that it originated from the small
face of Asian H. erectus. However, as the reduced facial skeletonsfrom the African later Middle Pleistocene (e.g., Eliye Springs, Singa,Florisbad, Jebel Irhoud, Herto) suggests, such facial reduction inHomomay have occurred in parallel at a different place and time. Ifthis was the case, the reduced face of H. floresiensis could haveevolved from any groups of Homo as parallel evolution with AsianH. erectus.
The supraorbital torus of LB1 is not extensive anteriorly (F3)and is less thick compared to the typical conditions in post-1.8 Mapre-modern Homo (F3). This may be a primitive retention suc-ceeded from H. habilis, but a more plausible interpretation is thatthe torus of LB1 reduced from a H. erectus-like form as a part of theoverall facial gracilization. This is because it has a bulbous lateralend, which stands immediately above the frontomalare temporale,a characteristic common to post-1.5 Ma Afro-Asian pre-modernHomo (F10), and because it exhibits a unique Asian (particularlylate Javanese H. erectus) character of lateral thickening (F20).While the strongly arched supraorbital torus is observed in LB1,Daka, and Maba (F11), the narrow orbital shape of LB1 ismarginally shared only with Maba (F21). This LB1 trait is deter-mined by its relatively great (unreduced) orbital height, whichmay reflect that the orbital size was not so reduced in thisH. floresiensis individual compared to the brain and overallcraniofacial sizes. Van Heteren and de Vos (2007) interpreted thehigh orbital index (OBH/OBB) of LB1 as a pedomorphic featureshared with human and chimpanzee juveniles. Although the factthat a juvenile H. ergaster (KNM-WT 15000) has narrow orbits isconsistent with this hypothesis, it does not explain the entirecranial morphology of LB1 including the angled occipital, devel-oped frontal keel, and thick vault bones. Van Heteren (in press),discussed other aspects of H. floresiensis which cannot be viewedas pedomorphic features.
In lateral view, the zygomatic frontal process of LB1 standsvertically (not inclined downward and forward [F12]). Below thisprocess, the short infraorbital surface of LB1 faces slightly inferiorly(F13,19). Suchmorphology is observed in various post-habilis Afro-AsianHomo, and reflects the posterior relocation of their zygomaticbones. However, unlike the case in H. sapiens, LB1 retains theprimitive, prognathic facial morphology (F1) with forwardprotruding maxillary body and a distinct infraorbital sulcus (F9),and a near-parallel arrangement of the canine-molar rows thatmakes its lateral maxillary surface relatively flat and directlylaterally facing (F6). We infer that this forward position of themaxillary body and the shortened tooth rows (reduced toothcrown size) in LB1 create a wide space posterior to their lastmolars, and this spatial relationship results in one of the peculiarcharacteristics seen in the LB1 facial skeleton, the posteriorlyelongated palatine (F22).
In summary, the facial skeleton of LB1 experienced a consider-able degree of reduction that suggests decreased masticatory load,but retains some primitive morphology of Homo including themoderate facial prognathism and squarish maxillary arch shape.The hypothesis of reduced masticatory stress in H. floresiensis isconsistent with the evidence for their great reliance on stone tools(Moore et al., 2009), and meat eating, as inferred from the exca-vated faunal remains and their age profiles (van den Bergh et al.,2009). The overall facial configuration of LB1 is generally similarto Asian groups of pre-modern Homo. This is particularly true in theoverall facial size (F15e17), protruding maxillary body witha marked infraorbital sulcus (F9), supraorbital torus morphology(F20), and orbital shape (F21). These data do not contradict withHypothesis III that connects H. floresiensis with early JavaneseH. erectus, although the former has experienced more substantialfacial gracilization compared to the latter. We find no strongevidence to support Hypotheses I and II in these facial characters.

Y. Kaifu et al. / Journal of Human Evolution 61 (2011) 644e682 677
LB1 shares with H. habilis a few plesiomorphic facial traits (F1, 2, 5,6) but these are also shared with early Javanese H. erectus; therelatively flat nasal bridge characterizes H. habilis, but thismorphology is not evident in LB1 (F7). The straight, diagonalzygomaticoalveolar crest (F14) is found only in LB1 and Africanpost-1.8 Ma Homo in our comparative sample. Because there are noother traits that support direct relationships between these twogroups, the observed crest morphology in LB1 was probably inde-pendent evolution associated with the facial gracilization. Unfor-tunately, morphology of this part is not known in late JavaneseH. erectus from Sambungmacan and Ngandong. A wide nasalaperture is another possible peculiar characteristic in some post-1.8MA African Homo (Fransiscus and Trinkaus, 1988; not listed inTable 6), but this morphology is lacking in LB1.
Unique morphology in LB1
Most of the individual craniofacial characteristics of LB1 listedin Table 6 are shared with one or more of our comparativesubsamples as discussed above, but there are four possible uniquefeatures of LB1 within the fossil Homo compared here. Theseinclude an extremely small facial skeleton (S2), extensively fusedmedial and lateral pterygoid plates of the sphenoid (C41), theproportionally high orbit (F21), and a posteriorly elongated pala-tine (F22), although the character states of C41 and F22 are notknown for many reference groups in Table 6. The character S2reflects the marked facial gracilization in H. floresiensis, and thedevelopments of F21 and F22 may be explained in this context asdiscussed above.
Other phylogenetic hypotheses (Hypothesis IV)
Martin et al. (2006) supposed ancestoredescendant relation-ships between Ngandong H. erectus and H. floresiensis when theysimulated brain size reduction in H. floresiensis. However, LB1lacks many of the important characteristic features of Sam-bungmacan and/or Ngandong H. erectus, including a specializedmandibular fossa morphology represented by the disappearanceof the postglenoid process (C28), a wide tympanomastoid fissure(C29), a strongly developed postcondyloid tuberosity (C30), andan elongated midcranial base (C31). The opisthionic recess (C40)of LB1 is not extensive as those seen in Sambungmacan/Ngan-dong, suggesting the presence of their common ancestor ratherthan direct ancestoredescendant relationships.
Some researchers suggested possible link of H. floresiensiswithaustralopithecines (Dennell and Roebroeks, 2005; Brown andMaeda, 2009). The strongest supportive cranial evidence for thisview would be the LB1’s small neurocranial size (S1), but theabove analyses showed that LB1/1 is clearly derived fromH. habilis showing laterally expanded parietals (C3, PC1 in Fig. 14),and resembles H. erectus in overall neurocranial shape (Fig. 15;C1e5 in Table 6). Most of the other 61 craniofacial characters ofLB1 in Table 6 variably support Hypothesis I, II, and/or III, or canbe regarded as derived characters within Homo, as discussedabove. The possible exception to this is F21 (tall and narrow orbit),in which LB1 apparently resembles Au. africanus (and Au. afar-ensis?) (Kimbel et al., 2004), but not H. habilis, Dmanisi Homo, orJavanese H. erectus. However, given the fact that H. floresiensisexperienced marked facial reduction compared to the Austral-opithecus condition (Brown et al., 2004; Argue et al., 2006;Gordon et al., 2008; Baab and McNulty, 2009; this study), it ispossible that this similarity occurred secondarily. Thus, there isno strong evidence linking H. floresiensis with late JavaneseH. erectus or Australopithecus.
Origins of H. floresiensis: current status of evidence
The conclusive answer as to the ancestry of H. floresiensis willcome from future discoveries of fossils of its ancestors in Flores orneighboring regions. However, comparative morphological anal-yses of the currently available fossil collections still provide valu-able information regarding its origins and mode of evolution.Hypothesis I (H. floresiensis originated from H. habilis) Thishypothesis only requires a moderate degree of brain and body sizereduction in the evolution of Homo floresisensis. Estimated endo-cranial volumes of the known H. habilis individuals range fromw500e750 cm3while that for LB1 was reported to be 380e417 cm3
(Brown et al., 2004; Falk et al., 2005). The relatively short legs ofH. floresiensis and primitive wrist bone morphology are alsoexplained as plesiomorphic retentions derived from this basalHomo species. However, currently there is no direct evidence forsuch a primitive form of Homo in eastern Asia. Instead availablefossil evidence indicates that Java and South China were occupiedby groups of H. erectus for more than 1 Myr. during the later partof the Pleistocene.
If pre-erectus grade hominins were actually present in SoutheastAsia and reached to Flores beforeH. erectus arrived in the region, weexpect that the former was also present in Java where the accesswas easier than to Flores as indicated by its more diverse terrestrialfauna (De Vos et al., 1994). No such evidence has so far beendiscovered from Sangiran and other fossil localities in Java, whichrecord terrestrial mammalian fauna both before and afterH. erectus. At the present stage, the earliest securely recordedhominin fossils of Java are from the stratigraphic levels above theTuff 11 of the Sangiran Formation or in the upper PucanganFormation of Mojokerto (Itihara et al., 1985; Larick et al., 2001;Huffman et al., 2006), and all of them can be regarded asH. erectus (Rightmire, 1990; Anton, 1997; Kaifu et al., 2005b, 2010a;Indriati and Anton, 2008). Below these levels, the fossil evidenceindicates that Java had been inhabited by proboscideans, hippo-potamuses, cervids, bovids and other terrestrial mammals fromhundreds of thousands of years earlier than the first appearance ofH. erectus (De Vos et al., 1994).
Our study indicates that morphological evidence for HypothesisI is weak at best as far as the cranial morphology is concerned. Fourcranial characteristics (S1, C13, C34, C42) may positively supportthis possible link. However, with reference to Hypothesis III,Hypothesis I requires that 21 craniofacial traits (those ways inwhich LB1 resembles early Javanese H. erectus) evolved indepen-dently in the lineage of H. floresiensis (C3e5, 7, 14, 19, 20, 22, 23,35e39, 43; F7e10,16, 17: Fig. 23). Because LB1/1 is close to KNM-ER1813 in overall cranial size, the differences observed between thesetwo specimens are not readily ascribed to size-related cranial shapevariation within H. habilis. It is conceivable that aspects of facialgracilization occurred in parallel in different regional groups ofHomo with a similarly reduced need for powerful mastication.However, similarities between H. floresiensis and H. erectus includecranial vault shape, arrangement of the temporal lines, details ofthe cranial base morphology, and supraorbital torus. If suchextensive parallel evolution between Javanese H. erectus andH. floresiensis actually occurred, it may have been because of similaradaptations to analogous Southeast Asian environments.Hypothesis II (H. floresiensis originated from Dmanisi Homo) Inthis hypothesis the degree of body size reduction in theH. floresiensis lineage was slightly more pronounced than in thecase of Hypothesis I. However, craniofacial morphology does notprovide exclusive support for the hypothesis, while evolutionaryreversal would be required to explain the primitive limb propor-tions of H. floresiensis. In addition, there is currently no evidence forsuch a primitive form of Homo in eastern Asia.
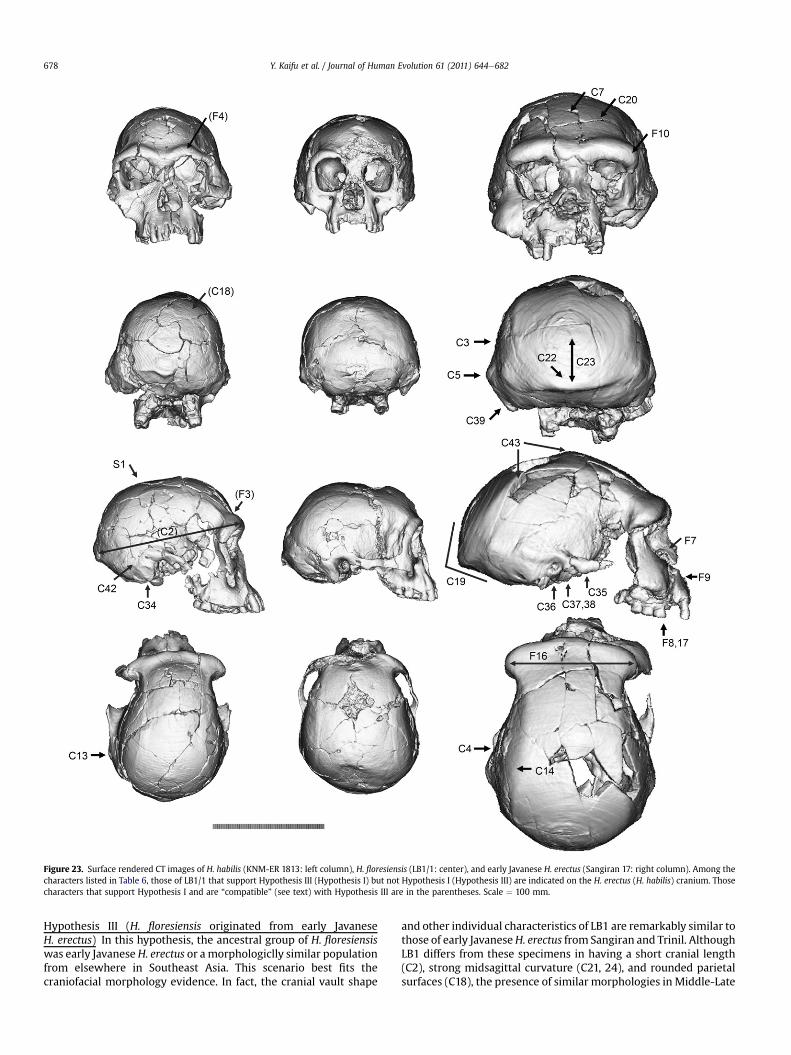
Figure 23. Surface rendered CT images of H. habilis (KNM-ER 1813: left column), H. floresiensis (LB1/1: center), and early Javanese H. erectus (Sangiran 17: right column). Among thecharacters listed in Table 6, those of LB1/1 that support Hypothesis III (Hypothesis I) but not Hypothesis I (Hypothesis III) are indicated on the H. erectus (H. habilis) cranium. Thosecharacters that support Hypothesis I and are “compatible” (see text) with Hypothesis III are in the parentheses. Scale ¼ 100 mm.
Y. Kaifu et al. / Journal of Human Evolution 61 (2011) 644e682678
Hypothesis III (H. floresiensis originated from early JavaneseH. erectus) In this hypothesis, the ancestral group of H. floresiensiswas early JavaneseH. erectus or amorphologiclly similar populationfrom elsewhere in Southeast Asia. This scenario best fits thecraniofacial morphology evidence. In fact, the cranial vault shape
and other individual characteristics of LB1 are remarkably similar tothose of early JavaneseH. erectus from Sangiran and Trinil. AlthoughLB1 differs from these specimens in having a short cranial length(C2), strong midsagittal curvature (C21, 24), and rounded parietalsurfaces (C18), the presence of similar morphologies in Middle-Late

Y. Kaifu et al. / Journal of Human Evolution 61 (2011) 644e682 679
Pleistocene Javanese H. erectus suggests that such characteristicscould evolve independently from the condition in Sangiran/TrinilH. erectus. While the face of LB1 is substantially reduced toa degree close to later Middle Pleistocene pre-modern Homo fromChina, it retains a primitive prognathic face (F1) and squarish(non-parabolic) maxillary arch (F6). Similar facial morphologieswere occasionally found in our sample of Asian pre-modern Homo.
Thus, it is possible that the craniofacial morphology ofH. floresiensis was derived from early Javanese H. erectus, withsubstantial facial gracilization associated with reduced masticatoryactivities. In this scenario, the 4 LB1 cranial characters that supportHypothesis I (a small neurocranium, a weak supramastoid crest,a poorly developed vaginal process, and a thin mastoid portion ofthe cranial bone) evolved uniquely in the lineage of H. floresiensisand were not plesiomorphic retentions from the H. habiliscondition.
This hypothesis is also compatible with the recent findings fromthe Soa Basin on Flores, where the oldest artifacts date back at least1.02 Ma (Brumm et al., 2010). Ages of Sangiran H. erectus fossilcollection are estimated as w1.6e1.0 Ma (Larick et al., 2001) orw1.2e0.8 Ma (Falguères, 2001; Hyodo, 2001).
Recent studies indicate that insular dwarfism can occur in large-bodied primates (Bromham and Cardillo, 2007; Welch, 2009), sothe same adaptive process might be expected in hominins.However, there are other challenges for Hypothesis III; in particular,the degree of brain size reduction required in the evolution ofH. floresiensis (Martin et al., 2006; Conroy and Smith, 2007; Falket al., 2007, 2009; Niven, 2007, 2008; Taylor and van Schaik,2007; Köhler et al., 2008; Aiello, 2010). However, Weston andLister (2009) recently reported that such marked reductionaccompanied by phyletic body size dwarfism actually occurred inan extinct Malagasy pygmy hippopotamus. By applying the modeldrawn from this evidence to the case ofH. floresiensis, these authorspredicted that an ancestral endocranial capacity of w800 cm3 andbody mass of 60 kg could produce the observed brain size ofH. floresiensiswith an estimated bodymass of 23 kg. This estimate isclose to the smallest endocranial capacity estimated for SangiranH. erectus (813 cm3 for S 2, Holloway et al., 2004). However, itremains uncertain if we can directly compare scaling models ofbrain size between Homo and non-primate mammalian species. Inthis regard, Montgomery et al. (2010) suggested that, based on theirestimates of ancestral brain mass for various primate taxa, evolu-tionary brain size reduction occurred in some of the primateevolutionary lineages, accompanied by body size decrease. Whenthe ranges of the absolute and relative brain size decreases thatthey found in such lineages were compared with the severalpossible evolutionary models of the H. floresiensis brain, a hypoth-esis that H. floresiensis originated from “average” H. erectuswith thebrain and body masses of 951 g and 57 kg, respectively, wasunlikely unless H. floresiensis had a considerably small body massapproximating 16 kg.
Another difficulty for Hypothesis III is the need for reverseevolution to explain the Australopithecus-like body proportions inH. floresiensis (Morwood and Jungers, 2009; Brown and Maeda,2009). If reversal occurred, then the relatively long arm and footlengths of LB1 (Morwood et al., 2005; Jungers et al., 2009a,b) musthave been produced by intensive shortening of the lower limbbones. Shortening of the limb bones is a typical observation inendemic quadrupedal large mammals on islands (Sondaar, 1977;van der Geer, 2005; Lyras et al., 2009; Meijer et al., 2010; vander Geer et al., 2010). Such a shift in body proportion mayenhance locomotion stability at the cost of running speed in theabsence of fleet-footed predators. Additionally, there is contro-versy if relative femoral length tends to reduce in small-bodiedpopulations/individuals of modern human and great ape, and if
the short, apelike lower limbs of LB1 can be explained as a simplefunction of body size scaling (Holliday and Franciscus, 2009;Jungers, 2009).
Primitive traits occur throughout the H. floresiensis skeleton.Some are shared with H. erectus/ergaster, while others concernskeletal elements for which there is no morphological informationfor H. erectus. For example, the humerus of LB1 exhibits a primitiveweak torsion angle, as found in H. ergaster and Dmanisi Homo(Lordkipanidze et al., 2007; Larson et al., 2007, 2009), while thewrist and foot bones have Australopithecus-like traits (Tocheri et al.,2007, 2008; Jungers et al., 2009a,b), but few H. erectus foot or handbones are available for comparison.
Other claimed primitive traits for H. floresiensis require moredetailed study and confirmation; for instance, the clavicle of LB1 issaid to be short (Larson et al., 2007), but its broken medial endneeds to be cleaned and reconstructed to allow an accurate esti-mate. The morphologies of the pelvis and limb bones have beenfully described (Larson et al., 2009; Jungers et al., 2009b), butdetailed systematic comparative analyses for them are yet to bepublished.
The LB1 and LB6 mandibles and mandibular teeth were studiedin detail by Brown and Maeda (2009), who reported the followingcharacters that were shared with Australopithecus but which arenot present or very uncommon in Asian H. erectus (their Table 3):(1) a mesiodistally elongated P1 crown; (2) a relatively narrowalveolar arcade; (3) a strongly inclined symphyseal axis; (4) well-developed superior and inferior transverse tori; and (5) anteriorposition of the anterior ramus root (the M1/M2 level). In addition,they emphasized that (6) a tubular, well-developed lingual alveolarprominence comparable to that of LB1 was not present in Sangiranand ZhoukoudianH. erectus. Furthermore, (7) their PCA based on anelliptic Fourier analysis of the midsagittal symphyseal contourseparated the two H. floresiensis mandibles from H. sapiens andH. erectus s. l., and placed the former within the cloud of Au. afar-ensis. Characters (3) and (4) were also supported by Argue et al.(2009). Our own data indicate that (2) is not the case (Table 7).The relative alveolar arcade shapes of LB1 and LB6 are wider (theindices are greater) than Au. afarensis, H. habilis, and DmanisiHomo,and are close to those of Sangiran 9 and 22. Other characters arealso present in H. erectus and other middle Pleistocene Homo,including Sangiran 1b (Sangiran 5 and 6 as well?), Zhoukoudian H1,OH 22, and Arago displaying a strongly inclined symphyseal axis;Sangiran 8, KNM-BK 67 and 8518with well-developed superior andinferior transverse tori; two Sangiran mandibles (Sangiran 9,Bk7905) with an anterior position of the anterior ramus root (theM1/M2 level); and Sangiran 22, OH 22, and Arago 13 with a tubular,well-developed lingual alveolar prominence (Wood and van Noten,1986; Rightmire, 1990; Bräuer and Schultz, 1996; Kaifu et al.,2005a). As for the midsagittal symphyseal contour, the Sangiransample used by Brown and Maeda (2009) did not represent theexisting materials sufficiently: the superior halves of the midlineportions are not preserved in Sangiran 5 and 6 (thus the contoursused are uncertain estimates), and Sangiran 9 and 22 could not beincluded although these two specimens fall short of LB1 and LB6 interms of relative symphyseal thickness. Brown and Maeda (2009)also pointed out that the H. floresiensis mandibles are differentfrom Australopithecus in exhibiting small canine size, molars sizerelationship of M1 � M2 > M3 (their Table 3), and no “distinctiveposterior corpus robusticity” (p. 577). All of these are derivedconditions associated with tooth crown size reduction and lesshorizontal separation between the ramus and lateral corpus (Rosasand Bermúdez de Castro, 1998; Kaifu et al., 2005b). Additionally,LB1 and LB6 lack the lateral surface hollowing between the canine/P2 juga and lateral prominence, another primitive characteristic ofAu. afarensis (White et al., 1981). Taken together, in our view, the

Table 7Comparisons of alveolar arcade index.a
H. floresiensisLB1 105b
LB6 103c
Au. afarensisL.H. 4 125f
A.L. 266-1 121f
A.L. 288-1i 119f
A.L. 400-1a 137g
MAK-VP-1/12 126d
H. habilisKNM-ER 1805 119f
OH13 117f
H. ergasterKNM-ER 730 98f
African middle Pleistocene HomoTighenif 1 99h
Tighenif 3 97h
KNM-BK 8518 113f
Dmanisi HomoD211 123e
D2600 123e
Sangiran H. erectusSangiran 9 108f
Sangiran 22 107f
Zhoukoudian H. erectusZKD H1 88h
a Arcade lengthewidth ratio. Width, distancebetween right and left intersection points of distalcontour of M3 andmidline of themolar row. Length,distance from infradentale to line tangential todistal faces of right and left M3 crowns.
b Measured from Fig. 2 of Kaifu et al. (2009).c Measured from Fig. 1 of Brown and Maeda
(2009).d Measured from Fig. 7 of White et al. (2000).e Measured from Figs. A1.1 and A2.1 of van
Arsdale (2006), respectively.f Measured from occlusal photograph of casts
(Au. afarensis) or the original specimens (theothers), after making a mirror image when only oneside of the arch is complete enough.
g Measured directly from a cast.h Data from Kaifu et al. (2005b).
Y. Kaifu et al. / Journal of Human Evolution 61 (2011) 644e682680
H. floresiensis mandibles are more derived and have morecommonalities with Sangiran H. erectus than Brown and Maeda(2009) suggested. More comprehensive comparisons are neededto assess the former’s morphological affinities.
Kaifu et al. (2005b, 2010a) showed that “early JavaneseH. erectus” from Sangiran and Trinil is not a single, homogeneousentity but exhibits substantial chronological variation. Compared tothe specimens from the upper Bapang-AG stratigraphic levels(above the Grenzbank zone), those from the lower Grenzbank/Sangiran levels of Sangiran (uppermost part of the Sangiran[Pucangan] Formation and the Grenzbank zone) are more primitivein their small cranial vault sizes, large posterior tooth crown sizes,and other morphological details of the cranium, mandible, andteeth. In connection with these findings, Baab and McNulty (2009)suggested that the ancestral group of H. floresiensis could be linkedwith this older, primitive type of early Javanese H. erectus, dated tow1.6 Ma or w1.2 Ma. Although Hypothesis III does not contradictthis possibility, the early Javanese H. erectus cranial sample of thepresent study consists mostly of the Bapang-AG crania (N ¼ 6) andincludes only a small number of the Grenzbank/Sangiran speci-mens (N ¼ 1, or 3 if Trinil and Sangiran 2 are from this level asdiscussed in Kaifu et al., 2010a). Thus, the cranial morphology ofLB1 could also indicate that the ancestors of H. floresiensis camefrom the H. erectus population present in Java around a millionyears ago.
Concluding remarks
After eight years of discovery, intensive studies, discussion anddebate, H. floresiensis still poses difficult questions and challenges.However, we conclude from detailed study of LB1 craniofacialshape and surface morphology that this endemic species couldhave descended from an early Pleistocene H. erectus population inJava or elsewhere in Southeast Asia. If so, then the process wouldhave included drastic body and brain size dwarfism and facialgracilization, as originally proposed (Brown et al., 2004). Whethersuch dramatic dwarfism actually occurred in Homo is still a matterof intense debate, and the cranial evidence alone cannot solve thequestion regarding the phylogenetic origin of H. floresiensis.Ongoing neontological and paleontological studies of mammalianevolution on islands, further morphological studies of H. floresiensisremains, and (most importantly) future discoveries of skeletalevidence for the first hominins to colonize Flores will provide thekey for understanding the evolutionary history and significance ofthis unexpected, but most welcome little hominin.
Acknowledgments
Iwan Kurniawan and Fachroel Aziz kindly helped us withpreparation of the LB1/1. We are grateful to Gen Suwa, and Reiko T.Kono for the micro-CT scanning of LB1, Emma Mbua, Teuku Jacob,Etty Indriati, Friedemann Schrenk, Ottmar Kullmer, John de Vos,Robert Kruszynski, Liu Wu for access to the specimens in their care,and Matthew Tocheri for helpful advice. Comments from the threeanonymous reviewers, associate editor, and Steve Leigh greatlyhelped to improve the text and analyses. The CT data of KNM-ER1813 were provided from the National Museum of Kenya, withassistance from Emma Mbua and Fred Spoor. This study was sup-ported by grants from the National Museum of Nature and Science,Tokyo.
References
Aiello, L.C., 2010. Five years of Homo floresiensis. Am. J. Phys. Anthropol. 142,167e179.
Andrews, P., 1984. An alternative interpretation of characters used to define Homoerectus. Courier Forschungsinstitut Senckenberg 69, 167e175.
Antón, S.C., 1997. Developmental age and taxonomic affinity of the Mojokerto child,Java, Indonesia. Am. J. Phys. Anthropol. 102, 497e514.
Antón, S.C., 2002. Evolutionary significance of cranial variation in Asian Homoerectus. Am. J. Phys. Anthropol. 118, 301e323.
Antón, S.C., 2003. Natural history of Homo erectus. Yearb. Phys. Anthropol. 46,126e170.
Antón, S.C., 2004. The face of Olduvai hominid 12. J. Hum. Evol. 46, 337e347.Argue, D., Donlon, D., Groves, C., Wright, R., 2006. Homo floresiensis: microcephalic,
pygmoid, Australopithecus or Homo? J. Hum. Evol. 51, 360e374.Argue, D., Morwood, M., Sutikna, T., Jatmiko, Wahyu Saptomo, E., 2009. Homo
floresiensis: a cladistic analysis. J. Hum. Evol. 57, 623e639.Argue, D., Morwood, M., Sutikna, T., Jatmiko, Wahyu Saptomo, E., 2010. A reply to
Trueman’s “A new cladistic analysis of Homo floresiensis”. J. Hum. Evol. 59,227e230.
Asfaw, B., Gilbert, W.H., Richards, G.D., 2008. Homo erectus cranial anatomy. In:Gilbert, W.H., Asfaw, B. (Eds.), Homo erectus: Pleistocene Evidence from theMiddle Awash, Ethiopia. University of California Press, Berkeley,pp. 265e327.
Baab, K., 2008. The taxonomic implications of cranial shape variation in Homoerectus. J. Hum. Evol. 54, 827e847.
Baab, K.L., 2010. Cranial shape in Asian Homo erectus: geographic, anagenetic, andsize-related variation. In: Norton, C.J., Braun, D.R. (Eds.), Asian Paleoanthro-pology: From Africa to China and Beyond. Springer, Dordrecht.
Baab, K.L., McNulty, K.P., 2009. Size, shape, and asymmetry in fossil hominins: thestatus of the LB1 cranium based on 3D morphometric analyses. J. Hum. Evol. 57,608e622.
Baba, H., Aziz, F., Kaifu, Y., Suwa, G., Kono, R.T., Jacob, T., 2003. New Homo erectuscalvarium from the Pleistocene of Java. Science 299, 1384e1388.
Baccetti, T., 1998. Tooth rotation associated with aplasia of nonadjacent teeth. AngleOrthod. 68, 471e474.
Bilsborough, A., Wood, B.A., 1988. Cranial morphometry of early hominids: facialregion. Am. J. Phys. Anthropol. 76, 61e86.

Y. Kaifu et al. / Journal of Human Evolution 61 (2011) 644e682 681
Bräuer, G., 1988. Osteometrie. In: Martin, R., Knussman, R. (Eds.), Anthropologie.Gustav Fischer, Stuttgart, pp. 160e232.
Bräuer, G., Schultz, M., 1996. The morphological affinities of the Plio-Pleistocenemandible from Dmanisi, Georgia. J. Hum. Evol. 30, 445e481.
Bromham, L., Cardillo, M., 2007. Primates follow the ‘island rule’: implications forinterpreting Homo floresiensis. Biol. Lett. 3, 398e400.
Brown, P., Maeda, T., 2009. Liang Bua Homo floresiensis mandibles and mandibularteeth: a contribution to the comparative morphology of a new hominin species.J. Hum. Evol. 57, 571e596.
Brown, P., Sutikna, T., Morwood, M.J., Soejono, R.P., Jatmiko, Saptomo, E.W.,Awe, R.D., 2004. A new small-bodied hominin from the Late Pleistocene ofFlores, Indonesia. Nature 431, 1055e1061.
Brumm, A., Jensen, G.M., van den Berg, G.D., Morwood, M.J., Kurniawan, I., Aziz, F.,Storey, M., 2010. Hominins on Flores, Indonesia, by one million years ago.Nature 464, 748e752.
Conroy, G.C., Smith, R.J., 2007. The size of scalable brain components in the humanevolutionary lineage: with a comment on the paradox of Homo floresiensis.Homo 58, 1e12.
Clarke, R.J., 1977. The cranium of the Swartkrans hominid, SK 847 and its relevanceto human origins. Ph.D. dissertation, University of Witwatersrand.
Clarke, R.J., 1990. The Ndutu cranium and the origin of Homo sapiens. Am. J. Phys.Anthropol. 19, 699e736.
Culotta, E., 2005. Discoverers charge damage to ‘Hobbit’ specimens. Science 307,1848.
Dalton, R., 2005. Looking for the ancestors. Nature 434, 432e434.Day, M.H., Stringer, C.B., 1982. A reconsideration of the Omo Kibish remains and the
erectusesapiens transition. In: Première Congrès International de PaleontolgieHumaine, Nice, Prétirage, pp. 814e846.
Dean, M.C., Wood, B.A., 1981. Metrical analysis of the basicranium of extanthominoids and Australopithecus. Am. J. Phys. Anthropol. 54, 63e71.
Dean, M.C., Wood, B.A., 1982. Basicranial anatomy of Plio-Pleistocene hominidsfrom East and South Africa. Am. J. Phys. Anthropol. 59, 157e174.
Delson, E., Harvati, K., Reddy, D., Marcus, L.F., Mowbray, K., Sawyer, G.J., Jacob, T.,Márquez, S., 2001. The Sambungmacan 3 Homo erectus calvaria: a comparativemorphometric and morphological analysis. Anat. Rec. 262, 380e397.
Dennell, R., Roebroeks, W., 2005. An Asian perspective on early human dispersalfrom Africa. Nature 438, 1099e1104.
De Vos, J., Sondaar, P.Y., Van den Bergh, G.D., Aziz, F.,1994. TheHomo bearing depositsof Java and its ecological context. Cour. Forsch.-Inst. Senckenberg 171, 129e140.
Dodo, Y., Sawada, J., 2010. Supraorbital foramen and hypoglossal canal bridgingrevisited: their worldwide frequency distribution. Anthropol. Sci. 118, 65e71.
Durband, A.C., 2002. Squamotympanic fissure in the Ngandong and Sambungmacanhominins: a reply to Delson et al. Anat. Rec. 266, 138e141.
Durband, A.C., 2008. Mandibular fossa morphology in the Ngandong and Sam-bungmacan fossil hominids. Anat. Rec. 291, 1212e1220.
Durband, A.C., Kidder, J.H., Jantz, R.L., 2005. A multivariate examination of theHexian calvaria. Anthropol. Sci. 113, 147e154.
Falk, D., Hildebolt, C., Smith, K., Morwood, M.J., Sutikna, T., Brown, P., Jatmiko,Saptomo, E.W., Brunsden, B., Prior, F., 2005. The brain of LB1, Homo floresiensis.Science 308, 242e245.
Falk, D., Hildebolt, C., Smith, K., Morwood, M.J., Sutikna, T., Jatmiko, Saptomo, E.W.,Imhoh, H., Seidler, H., Prior, F., 2007. Brain shape in human microcephalics andHomo floresiensis. Proc. Natl. Acad. Sci. 104, 2513e2518.
Falk, D., Hildebolt, C., Smith, K., Morwood, M.J., Sutikna, T., Jatmiko, Saptomo, E.W.,Prior, F., 2009. LB1’s virtual endocast, microcephaly, and hominin brain evolu-tion. J. Hum. Evol. 57, 597e607.
Falk, D., Hildebolt, C., Smith, K., Brown, P., Jungers, W., Larson, S., Sutikna, T., Prior, F.,2010. Nonpathological asymmetry in LB1 (Homo floresiensis): a reply to Eck-hardt and Henneberg. Am. J. Phys. Anthropol. 143, 340e342.
Franciscus, R.G., Trinkaus, E., 1988. Nasal morphology and the emergence of Homoerectus. Am. J. Phys. Anthropol. 75, 517e527.
Falguères, C., 2001. Dating layers and fossils in Sangiran Dome: methods and results.In: Simanjuntak, T., Prasetyo, B., Handini, R. (Eds.), Sangiran: Man, Culture, andEnvironment in PleistoceneTimes. YayasanObor Indonesia, Jakarta, pp. 309e319.
Gabunia, L., Vekua, A., Lordkipanidze, D., Swisher III, C.C., Ferring, R., Justus, A.,Nioradze, M., Tvalchrelidze, M., Antón, S.C., Bosinski, G., Jöris, O., de Lumley, M.-A., Majsuradze, G., Mouskhelishvili, A., 2000. Earliest Pleistocene hominidcranial remains from Dmanisi, Republic of Georgia: taxonomy, geologicalsetting, and age. Science 288, 1019e1025.
Gordon, A.D., Nevell, L., Wood, B., 2008. The Homo floresiensis cranium (LB1): size,scaling and early Homo affinities. Proc. Natl. Acad. Sci. USA 105, 4650e4655.
Grine, F.E., Fleagle, J.G., Leakey, R.E. (Eds.), 2009. The First Humans: Origin and EarlyEvolution of the Genus Homo. Springer, Heidelberg.
Holliday, T.W., Franciscus, R.G., 2009. Body size and its consequences: allometry andthe lower limb length of Liang Bua 1 (Homo floresiensis). J. Hum. Evol. 57,223e228.
Holloway, R.L., Broadfield, D.C., Yuan, M.S., 2004. The Human Fossil Record, 3. BrainEndocasts. Wiley, New York.
Howells, W.W., 1973. Cranial variation in man. A study by multivariate analysis ofpattern of difference among recent populations. In: Papers of the PeabodyMuseum, Archaeology and Ethnology, vol. 67. Harvard Univ. Press, Cambridge,MA.
Huffman, O.F., Zaim, Y., Kappelman, J., Ruez, D., de Vos, J., Rizal, Y., Aziz, F., Hertler, C.,2006. Relocation of the 1936 Mojokerto skull discovery site near Perning, EastJava. J. Hum. Evol. 50, 431e451.
Hyodo, M., 2001. The Sangiran geomagnetic excursion and its chronologicalcontribution to the Quaternary geology of Java. In: Simanjuntak, T., Prasetyo, B.,Handini, R. (Eds.), Sangiran: Man, Culture, and Environment in PleistoceneTimes. Yayasan Obor Indonesia, Jakarta, pp. 320e335.
Indriati, E., 2006. Cranial lesions on the late Pleistocene Indonesian Homo erectusNgandong 7. In: Oxenham, M., Tayles, N. (Eds.), Bioarchaeology of SoutheastAsia. Cambridge University Press, Cambridge, pp. 290e308.
Indriati, E., Antón, S.C., 2008. Earliest Indonesian facial and dental remains fromSangiran, Java: a description of Sangiran 27. Anthropol. Sci. 116, 219e229.
Itihara, M., SudijonoKadar, D., Shibasaki, T., Kumai, H., Yoshikawa, S., Aziz, F.,Soeradi, T., WikarnoKadar, A.P., Hashibuan, F., Kagemori, Y., 1985. Geology andstratigraphy of the Sangiran area. In: Watanabe, N., Kadar, D. (Eds.), QuaternaryGeology of the Hominin Fossil Bearing Formations in Java. Geol. Res. Dev.Centre, Spec. Publ., pp. 11e43.
Jacob, T., 1973. New Finds of Lower and Middle Pleistocene Hominines from Indo-nesia and an Examination of Their Antiquity Paper presented at the Conferenceon Early Palaeolithic of East Asia, Montreal.
Jacob, T., Indriati, E., Soejono, R.P., Hsü, K., Frayer, D.W., Eckhardt, R.B.,Kuperavage, A.J., Thorne, A., Henneberg, M., 2006. Pygmoid AustralomelanesianHomo sapiens skeletal remains from Liang Bua, Flores: population affinities andpathological abnormalities. Proc. Nat. Acad. Sci. 103, 13421e13426.
Jungers, W.L., 2009. Interlimb proportions in humans and fossil hominins: vari-ability and scaling. In: Grine, F.E., Fleagle, J.G., Leakey, R.E. (Eds.), The FirstHumans: Origin and Early Evolution of the Genus Homo. Springer, Heidelberg,pp. 93e98.
Jungers, W.L., Harcourt-Smith, W.E.H., Wunderlich, R.E., Tocheri, M.W., Larson, S.G.,Sutikna, T., Awe, R.D., Morwood, M.J., 2009a. The foot of Homo floresiensis.Nature 495, 81e84.
Jungers, W.L., Kaifu, Y., 2011. On dental wear, dental work, and oral health in thetype specimen (LB1) of Homo floresiensis. Am. J. Phys. Anthropol 145, 282e289.
Jungers, W.L., Larson, S.G., Harcourt-Smith, W., Morwood, M.J., Sutikna, T.,Awe, Rokhus Due, Djubiantono, T., 2009b. Descriptions of the lower limbskeleton of Homo floresiensis. J. Hum. Evol. 57, 538e554.
Kaifu, Y., Aziz, F., Baba, H., 2005a. Hominid mandibular remains from Sangiran:1952e1986 collection. Am. J. Phys. Anthropol. 128, 497e519.
Kaifu, Y., Aziz, F., Indriati, E., Kurniawan, I., Jacob, T., Baba, H., 2008. Cranialmorphology of Javanese Homo erectus: new evidence for continuous evolution,specialization, and terminal extinction. J. Hum. Evol. 55, 551e580.
Kaifu, Y., Baba, H., 2011. Craniofacial Evidence for the Evolution of Homo erectus andHomo floresiensis [meeting abstract to be published in PaleoAnthropology 2011].
Kaifu, Y., Baba, H., Aziz, F., Indriati, E., Schrenk, F., Jacob, T., 2005b. Taxonomicaffinities and evolutionary history of the Early Pleistocene hominids of Java:dento-gnathic evidence. Am. J. Phys. Anthropol. 128, 709e726.
Kaifu, Y., Baba, H., Kurniawan, I., Sutikna, T., Saptomo, E.W., Jatmiko, Awe, RokhusDue, Kaneko, T., Aziz, F., Djubiantono, T., 2009. Brief communication: “Patho-logical” deformation in the skull of LB1, the type specimen of Homo floresiensis.Am. J. Phys. Anthropol. 140, 177e185.
Kaifu, Y., Indriati, E., Aziz, F., Kurniawan, I., Jacob, T., Baba, H., 2010a. Cranialmorphology and variation of the earliest Indonesian hominins. In: Norton, C.J.,Braun, D.R. (Eds.), Asian Paleoanthropology: From Africa to China and Beyond.Springer, Heidelberg.
Kaifu, Y., Kaneko, T., Kurniawan, I., Sutikna, T., Wahyu Saptomo, E., Jatmiko,Awe, R.D., Aziz, F., Baba, H., Djubiantono, T., 2010b. Posterior deformationalplagiocephaly properly explains the cranial asymmetries in LB1: a reply toEckhardt and Henneberg. Am. J. Phys. Anthropol. 143, 335e336.
Kaifu, Y., Zaim, Y., Baba, H., Kurniawan, I., Kubo, D., Rizal, Y., Arif, J., Aziz, F., 2011.New reconstruction and morphological description of a Homo erectus craniume Skull IX (Tjg-1993.05) from Sangiran, Central Java. J. Hum. Evol. 61, 270e294.
Kimbel, W.H., Rak, Y., 1985. Functional morphology of the asterionic region in extanthominoids and fossil hominins. Am. J. Phys. Anthropol. 66, 31e54.
Kimbel, W.H., Rak, Y., Johanson, D.C., 2004. The Skull of Australopithecus afarensis.Oxford University Press, New York.
Köhler, M., Moyà-Solà, S., Wrangham, R.W., 2008. Island rules cannot be broken.Trends Ecol. Evol. 23, 6e7.
Larick, R., Ciochon, R.L., Zaim, Y., Sudijono, Suminto, Rizal, Y., Aziz, F., Reagan, M.,2001. Early Pleistocene 40Ar/39Ar ages for Bapang Formation hominins, CentralJawa, Indonesia. Proc. Natl. Acad. Sci. 98, 4866e4871.
Larson, S.G., Jungers, W., Morwood, M.J., Sutikna, T., Jatmiko, Wahyu Saptomo, E.,Due Awe, Rokhus, Djubiantono, T., 2007. Homo floresiensis and the evolution ofhominin shoulder. J. Hum. Evol. 53, 718e731.
Larson, S.G., Jungers, W., Tocheri, M.W., Orr, C.M., Morwood, M.J., Sutikna, T., DueAwe, Rokhus, Djubiantono, T., 2009. Descriptions of the upper limb skeleton ofHomo floresiensis. J. Hum. Evol. 57, 555e570.
Lieberman, D.E., McBratney, B.M., Krovitz, G., 2002. The evolution and developmentof cranial form in Homo sapiens. Proc. Natl. Acad. Sci. USA 99, 1134e1139.
Liu, W., Zhang, Y., Wu, X., 2005. Middle Pleistocene human cranium from Tangshan(Nanjing), southeast China: a new reconstruction and comparisons with Homoerectus from Eurasia and Africa. Am. J. Phys. Anthropol. 127, 253e262.
Lordkipanidze, D., Jashashvili, T., Vekua, A., Ponce de León, M.S., Zollikofer, C.P.E.,Rightmire, G.P., Pontzer, H., Ferring, R., Oms, O., Tappen, M., Bukhsianidze, M.,Agusti, J., Kahlke, R., Kiladze, G., Martinez-Navarro, B., Mouskhelishvili, A.,Nioradzem, M., Rook, L., 2007. Postcranial evidence from early Homo fromDmanisi, Georgia. Nature 449, 305e310.
Lordkipanidze, D., Vekua, A., Ferring, R., Rightmire, G.P., Zollikofer, C.P.E., Ponce deLeón, M.S., Agusti, J., Kiladze, G., Mouskhelishvili, A., Nioradze, M., Tappen, M.,

Y. Kaifu et al. / Journal of Human Evolution 61 (2011) 644e682682
2006. A fourth hominin skull from Dmanisi, Georgia. Anat. Rec. 288A,1146e1157.
Lü, Z., 1990. La découverte de l’homme fossil de Jing-niu-shan Première étude.L’Anthropologie 94, 899e902.
Lukacs, J.R., Nelson, G.C., Walker, C., 2006. Anomalies of dental development inmodern humans and Homo floresiensis. Am. J. Phys. Anthropol. (Suppl. 42),122e123.
Lyras, G.A., Dermitzakis, M.D., van der Geer, A.A.E., de Vos, J., 2009. The Origin ofHomo floresiensis and its Relation to Evolutionary Processes under Isolation, vol.117 33e43.
Martinón-Torres, M., Bermúdez de Castro, J.M., Gómez-Robles, A., Margvelashvili, A.,Prado, L., Lordkipanidze, D., Vekua, A., 2008. Dental remains from Dmanisi(Republic of Georgia): morphological analysis and comparative study. J. Hum.Evol. 55, 249e273.
McCollum, M.A., 2000. Subnasal morphological variation in fossil hominids:a reassessment based on new observations and recent developmental findings.Am. J. Phys. Anthropol. 112, 275e283.
McNulty, K.P., Baab, K.L., 2010. Keeping asymmetry in perspective: a reply to Eck-hardt and Henneberg. Am. J. Phys. Anthropol. 143, 337e339.
Martin, R.D., MacLarnon, A.M., Phillips, J., Dobyns, W.B., 2006. Flores hominid: newspecies or microcephalic dwarf? Anat. Rec. 288A, 1123e1145.
Meijer, H.J., van den Hoek Ostende, L.W., van den Bergh, G.D., de Vos, J., 2010. Thefellowship of the hobbit: the fauna surrounding Homo floresiensis. J. Biogeogr37, 995e1006.
Moore, M.W., Sutikna, T., Jatmiko, Morwood, M.J., Brumm, A., 2009. Continuities instoneflaking technologyat LiangBua, Flores, Indonesia. J. Hum.Evol. 57, 503e526.
Montgomery, S.H., Capellini, I., Barton, R.A., Mundy, N.I., 2010. Reconstructing theups and downs of primate brain evolution: implications for adaptive hypoth-eses and Homo floresiensis. BMC Biol. 8, 9. doi:10.1186/1741-7007-8-9.
Morwood, M.J., Brown, P., Jatmiko, Sutikna, T., Saptomo, E.W., Westaway, K.E.,Awe, Rokhus Due, Roberts, R.G., Maeda, T., Wasisto, S., Djubiantono, T., 2005.Further evidence for small-bodied hominins from the late Pleistocene of Flores,Indonesia. Nature 437, 1012e1017.
Morwood, M.J., Jungers, W.L., 2009. Conclusions: implications of the Liang Buaexcavations for hominin evolution and biogeography. J. Hum. Evol. 57, 640e648.
Morwood, M.J., van Oosterzee, P., 2007. A New Human: The Startling Discovery andStrange Story of the “Hobbits” of Flores, Indonesia Walnut Creek, California.
Morwood,M.J., Soejono, R.P., Roberts, R.G., Sutikna, T., Turney, C.S.M., Westaway, K.E.,Rink, W.J., Zhao, J.-x., van den Bergh, G.D., Awe, Rokhus Due, Hobbs, D.R.,Moore, M.W., Bird, M.I., Fifield, L.K., 2004. Archaeology and age of Homo flor-esiensis, a newhominin from Flores in eastern Indonesia. Nature 431,1087e1091.
Morwood, M.J., Sutikna, T., Saptomoc, E.W., Jatmiko, Hobbs, D.R., Westaway, K.E.,2009. Preface: research at Liang Bua, Flores, Indonesia. J. Hum. Evol. 57, 437e449.
Natsume, A., Koyasu, K., Oda, S., Nakagaki, H., Hanamura, H., 2006. Premolar andmolar rotation in wild Japanese serow populations on Honshu Island. Japan.Arch. Oral Biol. 51, 1040e1047.
Nevell, L., Wood, B.A., 2008. Cranial base evolution within the hominin clade.J. Anat. 212, 455e468.
Niven, J.E., 2007. Brains, islands and evolution: breaking all the rules. Trends Ecol.Evol. 22, 57e59.
Niven, J.E., 2008. Response to Köhler et al.: impossible arguments about possiblespecies? Trends Ecol. Evo. 23, 8e9.
Pearson, O.M., 2008. Statistical and biological definitions of “anatomically modern”humans: suggestions for a unified approach to modern morphology. Evol.Anthropol. 17, 38e48.
Rak, Y., 1983. The Australopithecine Face. Academic Press, New York.Rightmire, G.P., 1990. The Evolution of Homo erectus: Comparative Anatomical
Studies of an Extinct Human Species. Cambridge University Press, Cambridge.Rightmire, G.P., 1998. Evidence from facial morphology for similarity of Asian and
African representatives of Homo erectus. Am. J. Phys. Anthropol. 106, 61e85.Rightmire, G.P., 2008. Homo in the Middle Pleistocene: hypodigms, variation, and
species recognition. Evol. Anthropol. 17, 8e21.Rightmire, G.P., Lordkipanidze, D., 2009. Comparisons of early Pleistocene skulls
from East Africa and the Georgian Caucasus: evidence bearing on the origin andsystematics of genus Homo. In: Grine, F.E., Fleagle, J.G., Leakey, R.E. (Eds.), TheFirst Humans: Origin and Early Evolution of the Genus Homo. Springer, Hei-delberg, pp. 39e48.
Rightmire, G.P., Lordkipanidze, D., Vekua, A., 2006. Anatomical descriptions,comparative studies and evolutionary significance of the hominin skulls fromDmanisi, Republic of Georgia. J. Hum. Evol. 50, 115e141.
Roberts, R.G., Westaway, K.E., Zhao, J., Turney, C.S.M., Bird, M.I., Rink, W.J., Fifield, L.K.,2009. Geochronology of cave deposits at Liang Bua and of adjacent river terracesin theWae Racang valley, western flores, Indonesia: a synthesis of age estimatesfor the type locality of Homo floresiensis. J. Hum. Evol. 57, 484e502.
Rosas, A., Bermúdez de Castro, J.M., 1998. On the taxonomic affinities of the Dmanisimandible (Georgia). Am. J. Phys. Anthropol. 107, 145e162.
Santa Luca, A.P., 1980. The Ngandong Fossil Hominins: A Comparative Study fora Far Eastern Homo erectus Group, vol. 78. Yale University Pub. Anthropol..1e175.
Schwartz, J.H., Tattersall, I., 2005. The human fossil record. In: CraniodentalMorphology of Early Hominins (Genera Australopithecus, Paranthropus,Orrorin), and Overview, vol. 4. Wiley-Liss, New Jersey.
Shang, H., Trinkaus, E., 2008. An ectocranial lesion on the middle Pleistocenehuman cranium from Hulu Cave, Nanjing, China. Am. J. Phys. Anthropol. 135,431e437.
Singer, R., 1954. The Saldanha skull from Hopefield, South Africa. Am. J. Phys.Anthropol. 12, 345e362.
Sneath, P.H.A., Sokal, R.R., 1973. Numerical Taxonomy. W.H. Freeman, San Francisco.Sondaar, P.Y., 1977. Insularity and its effect on mammal evolution. In: Hecht, M.K.,
Goody, P.C., Hecht, B.M. (Eds.), Major Patterns in Vertebrate Evolution. Plenum,New York, pp. 671e707.
Stringer, C.B., 1984. The definition of Homo erectus and the existence of thespecies in Africa and Europe. Courier Forschungsinstitut Senckenberg 69,131e143.
Suwa, G., Asfaw, B., Haile-Selassie, Y., White, T., Katoh, S., Wolde Gabriel, G.,Hart, W.K., Nakaya, H., Beyene, Y., 2007. Early Pleistocene Homo erectus fossilsfrom Konso, southern Ethiopia. Anthropol. Sci. 115, 133e151.
Taylor, A.B., van Schaik, C.P., 2007. Variation in brain size and ecology in Pongo.J. Hum. Evol. 52, 59e71.
Terhune, C.E., Deane, A.S., 2008. Temporal squama shape in fossil hominins: rela-tionships to cranial shape and a determination of character polarity. Am. J. Phys.Anthropol. 137, 397e411.
Terhune, C.E., Kimbel, W.H., Lockwood, C.A., 2007. Variation and diversity in Homoerectus: a 3D geometric morphometric analysis of the temporal bone. J. Hum.Evol. 53, 41e60.
Tocheri, M.W., Orr, C.M., Jacofsky, M.C., Marzke, M.W., 2008. The evolutionaryhistory of the hominin hand since the last common ancestor of Pan and Homo.J. Anat. 212, 544e562.
Tocheri, M.W., Orr, C.M., Larson, S.G., Sutikna, T., Jatmiko, Saptomo, E.W., Due, RokusAwe, Djubiantono, T., Morwood, M.J., Jungers, W.L., 2007. The primitive wrist ofHomo floresiensis and its implications for hominin evolution. Science 317,1743e1745.
Tobias, P.V., 1991. Olduvai Gorge, 4: The Skulls, Endocasts and Teeth of Homo habilis.Cambridge Univ. Press, Cambridge.
Trueman, J.W.H., 2010. A new cladistic analysis of Homo floresiensis. J. Hum. Evol. 59,223e226.
van Arsdale, A.P., 2006. Mandibular variation in early Homo from Dmanisi, Georgia.Ph. D. Dissertation, University of Michigan.
van den Bergh, G.D., Meijer, H.J.M., Awe, R.D., Morwood, M.J., Szabó, K., van denHoek Ostende, L.W., Sutikna, T., Saptomo, E.W., Piper, P.J., Dobney, K.M., 2009.The Liang Bua faunal remains: a 95 kyr. sequence from Flores, East Indonesia.J. Hum. Evol. 57 (5), 527e537.
van der Geer, A.A.E., 2005. Island ruminants and the evolution of parallel functionalstructures. In: Cregut, E. (Ed.), Les ongulés holarctiques du Pliocène et duPléistocène. Actes Colloque International Avignon, 19e22 septembre. Qua-ternair, 2005 hors-série 2, pp. 231e240.
van der Geer, A., Lyras, G., de Vos, J., Dermitzakis, M., 2010. Evolution of IslandMammals. Wiley-Blackwell, Chichester.
van Heteren, A.H., The hominins of Flores: insular adaptations of the lower body. C.R. Palevol., in press.
van Heteren, A.H., de Vos, J., 2007. Heterochrony as a typical island adaptation inHomo floresiensis. In: Indriati, E. (Ed.), Recent Advances on Southeast AsianPaleoanthropology and Archaeology. Laboratory of Bioanthropology andPaleoanthropology. Gadjah Mada University, Yogyakarta, pp. 95e106.
Vialet, A., Guipert, G., He, J., Feng, X., Lu, Z., Wang, Y., Li, T., de Lumley, M.-A., deLumley, H., 2010. Homo erectus from Yunxian and Nankin Chinese sites:anthropological insights using 3D virtual imaging techniques. C. R. Pale 9,331e339.
Villmoare, B., 2005. Metric and nonmetric randomization methods, geographicvariation, and the single-species hypothesis for Asian and African Homo erectus.J. Hum. Evol. 49, 680e701.
Weidenreich, F., 1943. The skull of Sinanthropus pekinensis: a comparative study ona primitive hominin skull. Paleont. Sin. New Ser. D 10, 1e484.
Weidenreich, F., 1951. Morphology of Solo man. Anthropol. Pap. Am. Mus. Nat. Hist.43, 205e290.
Welch, J.J., 2009. Testing the island rule: primates as a case study. Proc. R. Soc. B 276,675e682.
Westaway, K.E., Sutikna, T., Saptomo, W.E., Jatmiko, Morwood, M.J., Roberts, R.G.,Hobbs, D.R., 2009. Reconstructing the geomorphic history of Liang Bua, Flores,Indonesia: a stratigraphic interpretation of the occupational environment.J. Hum. Evol. 57, 465e483.
Weston, E.M., Lister, A.M., 2009. Insular dwarfism is hippos and a model for brainsize reduction in Homo floresiensis. Nature 459, 85e88.
White, T.D., Johanson, D.C., Kimbel, W.H., 1981. Australopithecus africanus: itsphyletic position reconsidered. S. Afr. J. Sci. 77, 445e470.
White, T.D., Suwa, G., Simpson, S., Asfaw, B., 2000. Jaws and teeth of Australopithecusafarensis from Maka, Middle Awash, Ethiopia. Am. J. Phys. Anthropol. 111,45e68.
Wood, B.A., 1991. Koobi Fora Research Project, 4: Hominin Cranial Remains fromKoobi Fora. Clarendon Press, Oxford.
Wood, B.A., van Noten, F.L., 1986. Preliminary observation on the BK 8518 mandiblefrom Baringo, Kenya. Am. J. Phys. Anthropol. 69, 117e127.
Wu, R., 1988. The reconstruction of the fossil human skull from Jinniushan, Yinkou,Liaoning Province and its main features. Acta Anthropol. Sin 7, 97e101.
Wu, R., Zhang, Y., Wu, X., 2002. In: Wu, R., Li, X., Wu, X., Mu, X. (Eds.), Homo erectusfrom Nanjing. Jiangsu Science and Technology, Nanjing, pp. 261e273 (inChinese with English summary).
Wu, X., 2009. A metrical study of the Dali cranium. Acta Anthropol. Sin 28,217e236.
Wu, X., Poirier, F.E., 1995. Human Evolution in China. Oxford Univ. Press, Oxford.