growth-inhibitory action of an estrogen-chlorambucil...
TRANSCRIPT

[CANCER RESEARCH 52, 1187-1191, March l, 1992]
Growth-inhibitory Action of an Estrogen-Chlorambucil Conjugate (KM2210) inHuman Breast Cancer Cell Line MCF-7: Its Relation to Reduction of EstrogenReceptor and Transforming Growth Factor-«SecretionHiroshi Kosano,1 Tetsuro Kubota, Nakaaki Ohsawa, Shunji Yamamori, Osahiko Abe, Hirohumi Inagaki,
and Naokazu NagataThird Department of Internal Medicine, National Defense Medical College, 3-2, Namiki, Tokorozawa-ski, Saitama 359 [H. K., N. N.]; Department of Surgery, Schoolof Medicine, Keio University, Tokyo 160 [T. K., O. A.]; First Department of Internal Medicine, Osaka Medical College, Osaka 569 fN. OJ; Mitsubishi Yuka Bio-Clinical Laboratories, Inc., Tokyo 174 fS. YJ; and Department of Hygiene and Public Health, Nippon Medical School, Tokyo 113,[H. I.J Japan
ABSTRACT
We investigated the effects of a benzoate of an estradiol-chlorambucilconjugate (KM2210) and chlorambucil on growth, estrogen receptor, andsecretion of transforming growth factor (TGF)-a in the hormone-dependent human breast cancer cell line MCF-7. In the presence of 10"'°-10~'
M KM2210, the estrogen-induced growth of MCF-7 was completelyinhibited. Inhibited growth of MCF-7 treated with 10"" or 10"* M
KM2210 for 4 days was not rescued by removal of the drug and theaddition of estradiol. By treatment of MCF-7 with KM2210 for 4 days,estrogen receptor-binding sites were decreased at 1(1* M and were notdetected at 10~*Mbut were unaltered by IO"8Mchlorambucil. Moreover,
estrogen receptor immunoreactivity and the level of estrogen receptorniKN'A were decreased through treatment with II)"" M KM2210 for 4
days. These suppressions occurred prior to the onset of inhibitory actionon MCF-7 growth. Secretion of TGF-a from MCF-7 was decreased by 4days of treatment with 10"* and 10"* M KM2210 but not with chloram
bucil. The addition of exogenous TGF-a generally restored the growth ofMCF-7 treated with IO"8 M KM2210.
We concluded that KM2210 has irreversible or at least long-standinginhibitory effect on estrogen-dependent growth of MCF-7. It is conceivable that the decrease of estrogen receptor renders the cell unable torespond to estrogen with increased TGF-a secretion and succeeding cellgrowth.
INTRODUCTION
The goal of site-directed chemotherapy for malignant tumorsis to utilize drugs with a toxicity that is preferential for thetumor cells. KM22102 was initially developed as a target-oriented anticancer agent for the therapy of ER-positive breastcancer (1). The concentrations of KM2210 and its derivativesin the tumor tissue of the sensitive xenografts were 5-10 timeshigher than those in blood and muscle tissue (2).
Recently, it has been reported that the proliferation of hormone-dependent breast cancer cells is controlled by autocrinegrowth factors such as TGF-o (3), TGF-0 (4), and insulin-likegrowth factor type I (5). Induction by estradiol of a TGF-a-likeactivity was reported in a number of hormone-dependent breastcancer cells (3). Accordingly, studies on the mechanism ofanticancer drugs have focused much attention on the role ofsuch autocrine growth factors. In the previous report, we demonstrated that one of the anticancer drugs, alkyl-lysophospho-lipid, which possesses preferential cytotoxicity, inhibited estradiol uptake and TGF-a secretion in a hormone-dependent
Received 5/28/91 ; accepted 12/17/91.The costs of publication of this article were defrayed in part by the payment
of page charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
' To whom requests for reprints should be addressed.2The abbreviations used are: KM2210, estradiol-chlorambucil conjugate; ER,
estrogen receptor; TGF, transforming growth factor; CBL, chlorambucil; DCC,dextran-coated charcoal; MEM, minimal essential medium; EIA, enzyme immunoassay; SDS, sodium dodecyl sulfate; SSC, standard saline-citrate (O.IS MNaCl-0.15 M sodium citrate); EOF, epidermal growth factor; NBS, newbornbovine serum.
breast cancer cell line, MCF-7 (6). These observations maybecome useful for the development of a drug against breastcancer.
In the present study, we investigated the effect of KM2210and CBL on growth, ER, and TGF-a secretion in MCF-7 cells.
MATERIALS AND METHODS
Chemicals. 170-[2,4,6,7-3H]Estradiol(3.85 TBq/mmol) was pur
chased from New England Nuclear (Boston, MA). KM2210 (Fig. 1)and CBL were donated by Kureha Chemical Industry (Tokyo, Japan).17/3-Estradiol was purchased from Sigma Chemical Co. (St. Louis,MO) and all other chemicals used were obtained from Wako PureChemical Industries (Osaka, Japan). KM2210 and CBL as stock solutions (10~2 M in dimethyl sulfoxide) were added to a culture medium
to the desired concentrations with a Final dimethyl sulfoxide concentration of 0.33%.
Cell Lines. The human breast cancer cell line MCF-7 was subculturedin RPMI 1640 (Flow Laboratories, McLean, VA) supplemented with10% fetal bovine serum (Filtron, Altona, Australia) as described previously (7). For all experiments, the cells were passaged for 2-3 days inphenol red-free MEM (Sigma Chemical Co.) supplemented with 1% or5% NBS (Flow Laboratories) treated with DCC (8) and containing IO"8
M estradiol with a final concentration of absolute ethanol of 0.1%.Growth Experiment. MCF-7 cells were plated at a density of 2.5 x
104/well into 6-well multidishes (Falcon No. 3046; Becton-Dickinson,
Lincoln Park, NJ) with phenol red-free MEM supplemented with 1%DCC-NBS (experimental medium) and the incubation was initiated bythe addition of the drugs with or without 10~8 M estradiol. Medium
changes were made every 2 days. Cell number was counted at variousstages.
To test if the effect of KM2210 on the cell growth was reversible,the cells were plated as usual at a density of 2.5 x 104/well in theexperimental medium. After cell attachment (12-24 h), the cells wereexposed to the drugs by changing to the experimental medium containing each drug at the concentrations of IO"8 and 10~6M in the presenceof IO"8 M estradiol and cultured for 4 days. The medium was changed
every 2 days. On day 4, the medium was removed and the cells werewashed twice with the experimental medium. Then, IO"8 M estradiol
was added to one-half of the wells containing drug-treated cells. Thecell number was counted on days 11 and 15.
In order to determine the effect of exogenous TGF-a on the growthof MCF-7 treated with KM2210, the cells were exposed to KM2210 inthe presence of 10~8M estradiol for 4 days and then 10-1000 pg/ml of
human recombinant TGF-a (Bachern, Inc., Terranee, CA) were addedto the wells and cultured for 7 more days. The effect of TGF-a in theabsence of estradiol was also tested. Medium changes were made every2 days. The cell number was counted on day 11.
In all of the experiments, the viability of the cell examined by thetrypan blue exclusion test was more than 95%.
ER Binding Assay. Whole cell uptake of estradiol and Scatchardanalysis were carried out as described previously (9). MCF-7 cells wereplated at 1 x IO7 cells/bottle with the experimental medium andcultured for 1 day. All drugs were then added in the presence of 10~8
Mestradiol and cultured for 4 more days, and the medium was removed.The cells were harvested by trypsinization and washed twice with the
1187
on June 15, 2018. © 1992 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
REDUCTION OF ER AND TGF-n IN MCF-7 BY KM2210
^v
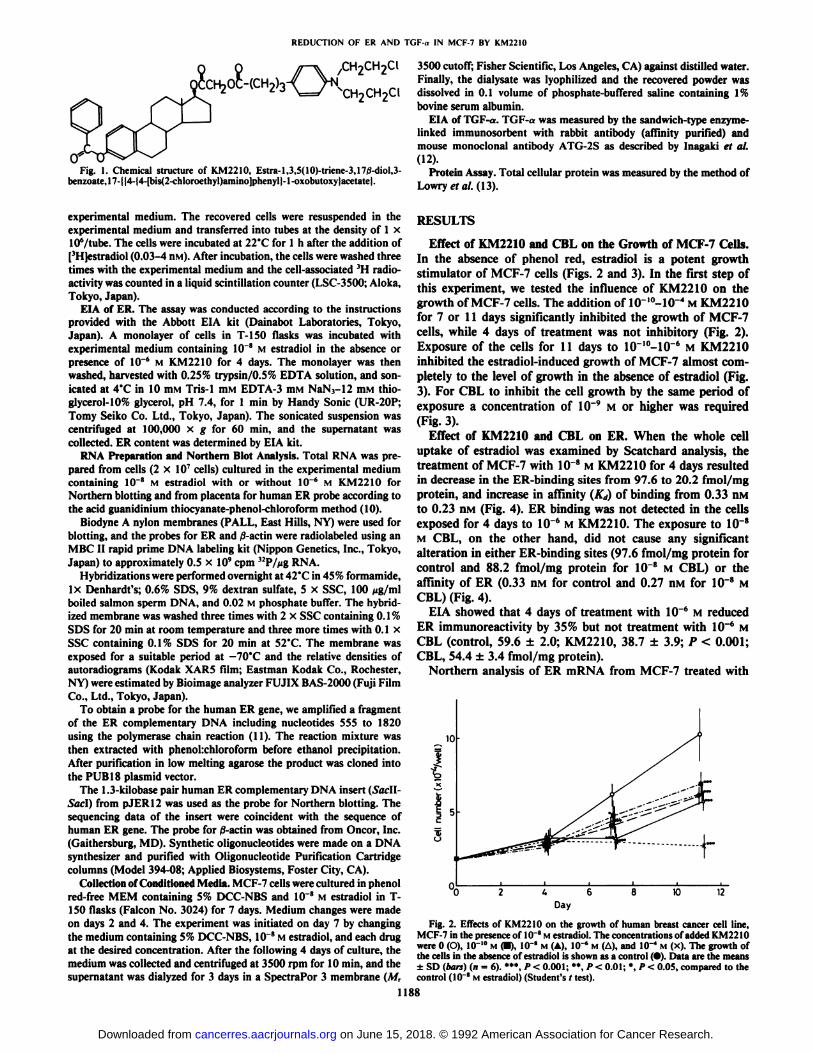
Fig. I. Chemical structure of KM2210, Estra-1,3,5(10)-triene-3,17/3-diol,3-benzoate,17-||4-|4-[bis(2-chloroethyl)amino]phenyl|-l-oxobutoxy|acetate|.
3500 cutoff; Fisher Scientific, Los Angeles, CA) against distilled water.Finally, the dialysate was lyophilized and the recovered powder was
\ / ^HoCHoCl dissolved in 0.1 volume of phosphate-buffered saline containing 1%bovine serum albumin.
EIA of TGF-a. TGF-a was measured by the sandwich-type enzyme-linked immunosorbent with rabbit antibody (affinity purified) andmouse monoclonal antibody ATG-2S as described by Inagaki et al.(12).
Protein Assay. Total cellular protein was measured by the method ofLowry et al. (13).
experimental medium. The recovered cells were resuspended in theexperimental medium and transferred into tubes at the density of 1 x106/tube. The cells were incubated at 22°Cfor l h after the addition of| '11|i-stradidl (0.03-4 HM).After incubation, the cells were washed threetimes with the experimental medium and the cell-associated 'II radio
activity was counted in a liquid scintillation counter (LSC-3500; Aloka,Tokyo, Japan).
EIA of ER. The assay was conducted according to the instructionsprovided with the Abbott EIA kit (Dainabot Laboratories, Tokyo,Japan). A monolayer of cells in T-1SO flasks was incubated withexperimental medium containing 10 * M estradici in the absence orpresence of 10~6 M KM2210 for 4 days. The monolayer was then
washed, harvested with 0.25% trypsin/0.5% EDTA solution, and sonicated at 4'C in 10 HIMTris-1 HIM EDTA-3 HIMNaN3-12 HIMthio-
glycerol-10% glycerol, pH 7.4, for 1 min by Handy Sonic (UR-20P;Tomy Seiko Co. Ltd., Tokyo, Japan). The sonicated suspension wascentrifuged at 100,000 x g for 60 min, and the supernatant wascollected. ER content was determined by EIA kit.
RNA Preparation and Northern Blot Analysis. Total RNA was prepared from cells (2 x 10" cells) cultured in the experimental mediumcontaining IO"8 M estradici with or without 10~6 M KM2210 for
Northern blotting and from placenta for human ER probe according tothe acid guanidinium thiocyanate-phenol-chloroform method (10).
Biodyne A nylon membranes (PALL, East Hills, NY) were used forblotting, and the probes for ER and /3-actin were radiolabeled using anMBC II rapid prime DNA labeling kit (Nippon Genetics, Inc., Tokyo,Japan) to approximately 0.5 x 10' cpm "P/ng RNA.
Hybridizations were performed overnight at 42°Cin 45% formamide,ix Denhardt's; 0.6% SDS, 9% dextran sulfate, 5 x SSC, 100 fig/ml
boiled salmon sperm DNA, and 0.02 M phosphate buffer. The hybridized membrane was washed three times with 2 x SSC containing 0.1%SDS for 20 min at room temperature and three more times with 0.1 xSSC containing 0.1% SDS for 20 min at 52°C.The membrane wasexposed for a suitable period at —¿�70°Cand the relative densities of
autoradiograms (Kodak XAR5 film; Eastman Kodak Co., Rochester,NY) were estimated by Bioimage analyzer FUJIX BAS-2000 (Fuji FilmCo., Ltd., Tokyo, Japan).
To obtain a probe for the human ER gene, we amplified a fragmentof the ER complementary DNA including nucleotides 555 to 1820using the polymerase chain reaction (11). The reaction mixture wasthen extracted with phenohchloroform before ethanol precipitation.After purification in low melting agarose the product was cloned intothe PUB 18 plasmid vector.
The 1.3-kilobase pair human ER complementary DNA insert (Sac\\-Saci) from pJFR 12 was used as the probe for Northern blotting. Thesequencing data of the insert were coincident with the sequence ofhuman ER gene. The probe for /3-actin was obtained from Oncor, Inc.(Gaithersburg, MD). Synthetic oligonucleotides were made on a DNAsynthesizer and purified with Oligonucleotide Purification Cartridgecolumns (Model 394-08; Applied Biosystems, Foster City, CA).
Collection of Conditioned Media. MCF-7 cells were cultured in phenolred-free MEM containing 5% DCC-NBS and IO"8 M estradiol in T-
150 flasks (Falcon No. 3024) for 7 days. Medium changes were madeon days 2 and 4. The experiment was initiated on day 7 by changingthe medium containing 5% DCC-NBS, IO"8M estradiol, and each drug
at the desired concentration. After the following 4 days of culture, themedium was collected and centrifuged at 3500 rpm for 10 min, and thesupernatant was dialyzed for 3 days in a SpectraPor 3 membrane (M,
RESULTS
Effect of KM2210 and CBL on the Growth of MCF-7 Cells.In the absence of phenol red, estradiol is a potent growthstimulator of MCF-7 cells (Figs. 2 and 3). In the first step ofthis experiment, we tested the influence of KM2210 on thegrowth of MCF-7 cells. The addition of 10-'0-10-1 M KM2210
for 7 or 11 days significantly inhibited the growth of MCF-7cells, while 4 days of treatment was not inhibitory (Fig. 2).Exposure of the cells for 11 days to 10-IO-10-6 M KM2210
inhibited the estradiol-induced growth of MCF-7 almost completely to the level of growth in the absence of estradiol (Fig.3). For CBL to inhibit the cell growth by the same period ofexposure a concentration of 10~9 M or higher was required
(Fig. 3).Effect of KM2210 and CBL on ER. When the whole cell
uptake of estradiol was examined by Scatchard analysis, thetreatment of MCF-7 with 10~8 M KM2210 for 4 days resulted
in decrease in the ER-binding sites from 97.6 to 20.2 fmol/mgprotein, and increase in affinity (Kd) of binding from 0.33 nivito 0.23 nM (Fig. 4). ER binding was not detected in the cellsexposed for 4 days to IO'6 M KM2210. The exposure to 10~8
M CBL, on the other hand, did not cause any significantalteration in either ER-binding sites (97.6 fmol/mg protein forcontrol and 88.2 fmol/mg protein for 10~8 M CBL) or theaffinity of ER (0.33 nM for control and 0.27 nivi for 10~8 M
CBL) (Fig. 4).EIA showed that 4 days of treatment with IO"6 M reduced
ER immunoreactivity by 35% but not treatment with 10~6 M
CBL (control, 59.6 ±2.0; KM2210, 38.7 ±3.9; P < 0.001;CBL, 54.4 ±3.4 fmol/mg protein).
Northern analysis of ER mRNA from MCF-7 treated with
Fig. 2. Effects of KM2210 on the growth of human breast cancer cell line,MCF-7 in the presence of 10~*Mestradiol. The concentrations of added KM2210were 0 (O), 10~10M (•),IO"8 M (A), IO"6 M (A), and 10"* M (x). The growth of
the cells in the absence of estradiol is shown as a control (•).Data are the means±SD (bars) (n = 6). *",/>< 0.001; «*,/»<0.01; *, P< 0.05, compared to thecontrol (10~8 M estradiol) (Student's t test).
1188
on June 15, 2018. © 1992 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

REDUCTION OF ER AND TGF-n IN MCF-7 BY KM22IO
-12 -11 -10 -9 -8 -7 -6 -5 -4
Log concentration (M)
Fig. 3. Growth inhibition of MCF-7 as a function of the concentration ofKM2210 and CBL. The cells were incubated for 11 days at 37'C with various
concentrations of KM22IO (O) and CBL (•).KM2210 and CBL were added tocultures containing 10~* M estradici (£¡).Data are the means ±SD (bars)
(n = 6).
0.08
0.04
offl
20Bound (pM)
tÃ)
Fig. 4. Scatchard analysis of estradici binding to MCF-7. Prior to the saturationbinding experiment, the cells were plated at 10 cells/bottle with the experimentalmedium supplemented with 10 " M estradiol and cultured for 1 day. KM2210and CBL were then added and incubated for 4 more days at 37*C. The values
plotted are the means of one representative experiment in triplicate. •¿�control(IO'1 M estradiol): O, IO"' M KM2210; •¿�,IO'8 M CBL.
secretion increased by estradiol was almost completely negatedby l(T6MKM2210(Table 1).
Effect of Exogenous TGF-a on the Growth of MCF-7. In theabsence of estradiol, exogenous TGF-a added at 10-1000 pg/ml promoted the growth of MCF-7 dose dependently, and theeffect at 100 to 1000 pg/ml was equivalent to the growth-promoting effect of IO'8 M estradiol. The addition of TGF-a
mostly restored the inhibition of estradiol-stimulated growthby IO"8 M KM2210 but restored the inhibition by 10~6 M
K.M2210 only partially (Table 2).
DISCUSSION
KM2210, a benzoate of an estradiol-chlorambucil conjugate,was initially synthesized as a target-oriented anticancer drugwith estradiol as the vehicle targeting CBL to ER-positivetumors. The mode of action of KM2210 is little understood,and because of its wide variety of actions, it appears essentialto investigate its mode of action with the use of a well-characterized cell line. MCF-7 is a human breast cancer cell line whichcontains a high concentration of ER (14). Its growth is inducedby estrogen and is dependent on several autocrine growthfactors such as TGF-a (3, 15) and IGF-I (5). Growth is specifically inhibited by antiestrogens and the inhibition is rescuedby estradiol (16, 17). Thus, in the present study using MCF-7,
Kb
9.49-
6.20-!
4.40-
2.37-
1.35-i
-ER mRNA
- 28S ribosomal RNA
- /î-actinmRNA
10~6 M KM2210 for 4 days demonstrated that KM2210 de
creased ER mRNA by approximately 88% (Fig. 5).Irreversibility of the Action of KM2210 on the Estradiol-
induced Growth of MCF-7. Inhibition of estradiol-inducedgrowth of MCF-7 by 4 days of pretreatment with 10~8and 10~6
M KM2210 was not reversed by removal of the drug andaddition of 10~8M estradiol, while the inhibitory effect of CBL
was reversible (Fig. 6).Effect of KM2210 on Secretion of TGF-a in MCF-7. The
effects of pretreatment with KM2210 and CBL on the amountof TGF-a secreted from MCF-7 into medium are shown inTable 1. The secretion of TGF-a from MCF-7 was suppressedsignificantly by 4 days of treatment with 10~8 and 10~6 M
KM2210, namely from 2.02 ±0.09 to 1.68 ±0.06 ng/mgprotein (83% of control) at 10~8 M or to 1.40 ±0.1 ng/mgprotein (69% of control) at IO"6 M. The portion of TGF-a
0.24-
Ml
Fig. 5. Effect of KM2210 on ER mRNA expression. MCF-7 was treated with(Lanes 2 and 4) or without (lanes I and 3) 1(T6 M KM22IO and 10"' M estradiol
for 4 days and mRNA was probed with human ER complementary DNA (LanesI and 2) by Northern blotting as described in "Materials and Methods." Thefilters were then washed and rehybridized for human ,>'artin determination (Lanes
3 and 4). This blot is representative of 3 experiments. Autoradiographs wereobtained after 2 days exposure. Kb, kilobases.
1189
on June 15, 2018. © 1992 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

REDUCTION OF ER AND TGF-a IN MCF-7 BY KM2210
Table 1 Amount ofTGF-a secreted from MCF-7 into the conditioned mediumcultured for 4 days with various concentrations ofKM2210 and CBL
Fig. 6. Irreversibility of the action of KM2210 on the estrogen-stimulatedgrowth of MCF-7 cells. The cells were plated (n = 6) and grown in the experimental medium supplemented with Id * M estradici and 10 * M KM2210 (B),IO'4 M KM2210 (O, 10-' M CBL (D), or 10'' M CBL (E). Control (A) wascultured with estradici (/;'?) alone. The medium was changed every 2 days. Onday 4, the drug was removed and either vehicle (•)or 10 * M estradici (O) was
added as indicated by the arrow. Data shown are the means ±SD (bars)(n = 6). •¿�*,P < 0.001; *, P < 0.01, compared to the absence of 10"" M estradici(Student's r test).
we investigated whether or not KM221(1has a growth-inhibitory effect on the cell and, if so, whether a change in ER orTGF-a secretion is associated with its effects.
KM2210 at Kr'0-l(r6 M inhibited the estrogen-dependentgrowth of MCF-7 after 7 days of exposure. For CBL to inhibitthe cell growth by the same period of exposure, a concentrationof IO"9 M or higher was required. Metabolism of KM2210
appeared to be very slow under this experimental condition assuggested from the observation by Kubota et al. (2) that even 2weeks after the administration of KM22 IO,KM2210, but notCBL, is detected in the blood of tumor-bearing mice.
Although the growth-inhibitory action of KM2210 was notevident until the 7th day of treatment, a significant suppressionof the ER-binding activity was observed by the 4th day ofexposure. It is suggested that the early event, namely thereduction of ER-binding sites, may play a role in the growthinhibition. It may be argued that the treatment by KM2210affects the cellular uptake of | 'H]fstradini and the whole cellassay results in a spuriously decreased ER-binding activity.However, the reduction of ER-binding sites in MCF-7 treatedwith KM2210 was observed also by Fluoro-Cep staining (ZeusScientific Inc.),3 a procedure which make the cell membranepermeable to allow the cell to uptake the fluorescein-estradiolconjugate. In addition, decreases of ER immunoreactivity andER mRNA were demonstrated. These changes as well as therather long exposure period (4 days) required to suppress ER-binding sites may indicate that KM2210 inhibited ER-synthe-sis. Another mechanism may be involved in the decreased ER-binding sites, as it was disproportionally greater as comparedwith the decrease of ER immunoreactivity. If the primary siteof action is on the ER synthesis, KM2210 is a unique ant¡estrogen unlike drugs such as tamoxifen. Unlike inhibition by la-mu\ iten, which is readily rescued by estradici (16, 17), inhibÃ-
DrugControl
No estradici10"' MestradiolKM2210IO"8
MIO"6
MTotal
protein(mg/flask)0.62
0.630.66
0.65TGF-ang/mg
protein"1.32±0.08*-f
2.02 ±0.091.68
±0.06''1.40±0.10f%
of control65
10083
69CBL, 10-*M 0.56 1.93 ±0.11 87
" MCF-7 was cultured in phenol red-free MEM containing 5% DCC-NBS and10 " M estradici in T-150 flask for 7 days. The experiment was started on day 7(logarithmic growth phase) by changing the medium containing 5% DCC-NBS,IO* M estradici, and each drug at the desired concentration. After the subsequent
4 days of culture, the medium and cells were collected and measured.'' Mean ±SD of triplicate determinations.' P < 0.001 compared to the control (10~" M estradiol) (Student's t test).* P < 0.01 compared to control (10~" M estradiol) (Student's t test).
Table 2 Effect of exogenous TGF-a on the growth of MCF-7 in the presence ofestradiol along with IO'' M or IO'* M KM2210
Cell no. (x 10-4/well)°
ExogenousTGF-a(pg/ml)0
10100
1000ControlNo
estradiol*10.6
±2.0f13.0± 1.7''15.4 ±2.1'16.2 ±2.8'Estradiol(IO"'
M)15.9
±0.817.4 ±1.817.7 ±\.f>a17.5 ±0.9'KM2210IO"'
M11.1
±1.113.4 ±1.6'13.9± 1.8'15.3 ±2.4'IO'6
M11.1
±0.811.1 ±1.611.3 ±1.313.1 ±1.1'
' T. Kubota, personal communication.
°The cells (2.5 x 104/well) were exposed to KM2210 in the presence of 10~'M estradici for 4 days and then TGF-a was added to the wells and cultured for 7more days. Cell number was counted on day 10.
* Endogenous TGF-a secreted was less than 7 pg/ml/2.5 x IO4 cells as
calculated by the data shown in Table 1.' Mean ±SD (n = 6).' P < 0.05 compared to TGF-a (0 pg/ml) (Student's t test).' P < 0.01 compared to TGF-a (0 pg/ml) (Student's / test).
tion by KM2210 was irreversible, or at least long-lasting. Thisdifference may either be a reflection of the longer half-life ofthe latter drug or a difference in its mode of action, the latteraction being more than functional and affecting cellularcomponent(s).
In using the estrogen conjugate, the growth-promoting effectofestrogen is of some concern. In the case of KM2210, however,decrease of ER prevents response of the cells to the action ofany estradiol which might be liberated during the metabolismof KM2210. Although KM2210 is an estradiol-CBL conjugate,CBL itself did not affect ER-binding sites and showed muchweaker growth-inhibitory activity as compared with K.M2210on a molar base. It appears that the effect of KM2210 on MCF-7 is caused by KM2210 per se and not by CBL liberated in theprocess of KM2210 degradation.
It has been shown that TGF-a is induced in MCF-7 byestradiol and is regarded as one of the representative mediatorsof growth-promoting action of estradiol on the cell (18). Exposure of MCF-7 for 4 days to 10~8or 10~6MKM2210 but notto 10~6MCBL inhibited the secretion of TGF-a from the cell.Secretion of TGF-a during 4-day culture of MCF-7 was 1.32ng/mg protein in the absence of estradiol and 2.01 ng/mgprotein in the presence of estradiol. Because the increment byestradiol was almost completely abolished by the addition of10 '' M KM2210, it is reasonable to attribute the inhibition of
estradiol stimulated cell growth to negation by KM2210 ofestradiol-stimulated TGF-a secretion. KM2210 suppressed the
1190
on June 15, 2018. © 1992 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

REDUCTION OF ER AND TGF-o IN MCF-7 BY KM2210
secretion of TGF-a within 4 days at concentrations of 10 8and10~6M.This suggested that the suppression of TGF-a secretion
occurred earlier as a cellular event than a reduction in cellgrowth rate (7-11 days). Although further study is needed, it ishighly possible that the suppression of ER synthesis caused byKM2210 resulted in reduction of TGF-a secretion from MCF-7. We have previously reported that a selectiveanticancer agent,alkyl-lysophospholipid, inhibited the secretion of TGF-a alongwith the reduction of ER-binding sites (6).
The growth of MCF-7 in the absence of estradiol was stimulated by exogenous TGF-a and the effect was equal to that of10~8Mestradiol; the simultaneous addition of TGF-a and 10~8
M estradiol did not show additive effect on the growth. Thesedata support the view that TGF-a is one of the most potentgrowth-promoting factors induced by estradiol. The growth-inhibitory effect of 10~8MKM2210 in the presence of estradiol
was generally restored by the addition of TGF-a, while theinhibition by 10~6Mof KM2210 was restored only partially. It
may be that KM2210 at high concentrations also affects themechanism which is required for TGF-a to exhibit its fullgrowth-stimulatory effect.
This study demonstrated that KM2210 is an inhibitor ofestradiol-stimulated growth of MCF-7 and that inhibition ofthe estradiol-stimulated TGF-a secretion, probably through thereduction of ER synthesis, is associated with the growth inhibitory effect. This is interesting from the recent clinical findingsthat activity of the EGF receptor is one of the most importantindices for predicting relapse-free primary breast cancer andoverall survival (19). TGF-a acts on breast cancer cells via aninteraction with the EGF receptor (20). Thus, inhibition ofTGF-a secretion is expected to result in a similar biologicalconsequence as the low level of EGF receptor. For this reason,KM2210 and alkyl-lysophospholipid (6), which reduce ER sitesand inhibit secretion of TGF-a in the breast cancer cell, areexpected to constitute a new type of anticancer drug.
ACKNOWLEDGMENTS
We are grateful to Kiroh Asano of Kureha Industry Co., Ltd., forproviding us with KM 2210 and chlorambucil.
REFERENCES
1. Ohsawa, N.. Yamazaki, /.. Wagatsuma, T., and Isurugi, K. Bestrabucil; apossible target-oriented anticancer agent (in Japanese). Jpn. J. Cancer (hemother., 11: 2115-2124, 1984.
2. Kubota, T., Kawamura, E., Suzuki, T., Yamada, T., Toyoda, H., Miyagawa,
T., and Kurokawa T. Antitumor activity and pharmacokinetics of estra-1,3,5( 10)-triene-3,17/3-diol, 3-benzoate, 17-((4-(4-(bis(2-chloroethyl)amino)-phenyl)-l-oxobutoxy)acetate) (bestrabucil) in human tumor xenografts serially transplanted into nude mice. Jpn. J. Clin. Oncol., 16: 357-364, 1986.
3. Dickson, R. B. Bates, S. E., McManaway, M. E., and Lippman, M. E.Characterization of estrogen responsive transforming activity in humanbreast cancer cell lines. Cancer Res., 46: 1707-1713, 1986.
4. Knabbe, C, Lippman, M. E., Wakefield, L. M., Flanders, K. C., Kasid, A.,Dm nek, R., and Dickson, R. B. Evidence that transforming growth factor-ßis a hormonal!} regulated negative growth factor in human breast cancercells. Cell, 48: 417-428, 1987.
5. Huff, K. K., Kaufman, D., Gabbay, K. H., Spencer, M. E., Lippman, M. E.,and Dickson, R. B. Secretion of an insulin-like growth factor-I-related proteinby human breast cancer cells. Cancer Res., 46:4613-4619, 1986.
6. Kosano, H., Yasutomo, Y., Kugai, N., Nagata, N., Inagaki, H., Tanaka, S,.and Takatani, O. Inhibition of estradiol uptake and transforming growthfactor a secretion in human breast cancer cell line MCF-7 by an alkyl-lysophospholipid. Cancer Res., 50: 3172-3175, 1990.
7. Kosano, H., and Takatani, O. Reduction of epidermal growth factor bindingin human breast cancer cell lines by an alkyl-lysophospholipid. Cancer Res.,¥«.•6033-6036,1988.
8. Horwitz, K. B.. and McGuire, W. L. Nuclear mechanisms of estrogen action:effects of estradiol and antiestrogens on estrogen receptor and nuclear receptor processing. J. Biol. Chem., 253: 8185-8198, 1978.
9. Taylor, C. M., Blanchard, B., and Zava, D. T. A simple method to determinewhole cell uptake of radiolabelled oestrogen and progesterone and theirsubcellular localization in breast cancer cell lines in monolayer culture. J.Steroid Biochem., 20:1083-1088, 1984.
10. Chomoczynski, P., and Sacelli, N. Single step method of RNA isolation byacid guanidinium thiocyanate-phenol-chloroform extraction. Anal.Biochem., 162: 156-159, 1987.
11. Marchuk, D., Drumm, M., Saulino, A., and Collins, F. C. Construction ofT vectors, a rapid and general system for direct cloning of unmodified PCRproducts. Nucleic Acid Res., 19: 1154, 1991.
12. Inagaki, H., Katoh, M., Kurosawa-Ohsawa, K., and Tanaka, S. A newsandwich enzyme-linked immunosorbent assay (ELISA) for transforminggrowth factor-a (TGF-a) based upon conformational modification by antibody binding. J. Immunoi. Methods, 128: 27-37, 1990.
13. Lowry, O. H., Rosebrough, N. J., Farr, A. L., and Randall, R. J. Proteinmeasurement with the Polin phenol reagent. J. Biol. Chem. 193: 265-275,1951.
14. Brooks, S. C., Locke, E. R., and Soule, H. D. Estrogen receptor in humancell line (MCF7) from breast carcinoma. J. Biol. Chem., 248: 6251-6253,1973.
15. Dickson, R. B., Huff, K. K., Spencer, E. M., and Lippman, M. E. Inductionof epidermal growth factor-related polypeptides by 17/3-estradiol in MCF-7human breast cancer cells. Endocrinology, 118:138-142, 1986.
16. Lippman, M. E., Bolán,G., and Huh, K. The effects of estrogens andantiestrogens on hormone responsive human breast cancer in long-term tissueculture. Cancer Res., 36:4595-4601, 1976.
17. Coezy, E., Borgna, J. L., and Rochefort, H. Tamoxifen and metabolites inMCF7 cells: correlation between binding to estrogen receptor and inhibitionof cell growth. Cancer Res., 42: 317-323, 1982.
18. Dickson, R. B., and Lippman, M. E. Estrogenic regulation of growth andpolypeptide growth factor secretion in human breast carcinoma. Endocr.Rev., 8: 29-43, 1987.
19. Sainsbury, J. R. C., Farndon, J. R., Needham, G. K., and Malcolm, A. J.Epidermal-growth-factor receptor status as predictor of early recurrence ofand death from breast cancer. Lancet, /: 1398-1402, 1987.
20. Pike, L. J., Marquardt, H., Todaro, G. J., Gallis, B., Casnellie, J. E.,Bornslein. P., and Krebs, E. G. Transforming growth factor and epidermalgrowth factor stimulate the phosphorylation of a synthetic, tyrosine-contain-ing peptide in a similar manner. J. Biol. Chem., 257; 14628-14631, 1982.
1191
on June 15, 2018. © 1992 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

1992;52:1187-1191. Cancer Res Hiroshi Kosano, Tetsuro Kubota, Nakaaki Ohsawa, et al.
SecretionαTransforming Growth Factor-Its Relation to Reduction of Estrogen Receptor andConjugate (KM2210) in Human Breast Cancer Cell Line MCF-7: Growth-inhibitory Action of an Estrogen-Chlorambucil
Updated version
http://cancerres.aacrjournals.org/content/52/5/1187
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/52/5/1187To request permission to re-use all or part of this article, use this link
on June 15, 2018. © 1992 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from