gopherus agassizii
TRANSCRIPT

CHANGING THE SURVIVAL FORMULA FOR THE MOJAVE DESERT TORTOISE
(GOPHERUS AGASSIZII) THROUGH HEAD-STARTING
by
PEARSON A. MCGOVERN
(Under the Direction of Tracey D. Tuberville and Clinton T. Moore)
ABSTRACT
Mojave desert tortoise populations are in decline and improving juvenile survival pre-
release (head-starting) is being evaluated to augment populations. We released three
treatment groups to evaluate the potential of combination head-starting. Treatment groups
consisted of tortoises reared outdoors for 6-7 years (n = 30), outdoors for two years (n =
24), or indoors for one year followed by outdoors for one year (‘combination head-
started’; n = 24). Combination head-starts were smaller than 6-7-year-old outdoor reared
animals at release, and both groups were significantly larger than animals reared solely
outdoors for two years. All treatment groups had nearly identical body conditions, while
two-year-old outdoor animals had significantly softer shells than either of the other
treatments pre-release. Combo head-starts exhibited strong post-release site-fidelity in
comparison to the solely outdoor reared treatments. Size was a significant predictor of
survival, with combo head-starts and 6-7-year-old outdoor head-starts exhibiting
particularly high survival rates 10-months post-release.
INDEX WORDS: Reptile, turtle, desert tortoise, Gopherus agassizii, population
augmentation, head-starting, applied conservation, survivorship,
recruitment

CHANGING THE SURVIVAL FORMULA FOR THE MOJAVE DESERT TORTOISE
(GOPHERUS AGASSIZII) THROUGH HEAD-STARTING
by
PEARSON A. MCGOVERN
B.S., Texas A&M University, 2017
A Thesis Submitted to the Graduate Faculty of The University of Georgia in Partial
Fulfillment of the Requirements for the Degree
MASTER OF SCIENCE
ATHENS, GEORGIA
2019

© 2019
PEARSON A. MCGOVERN
All Rights Reserved

CHANGING THE SURVIVAL FORMULA FOR THE MOJAVE DESERT TORTOISE
(GOPHERUS AGASSIZII) THROUGH HEAD-STARTING
by
PEARSON A. MCGOVERN
Major Professors: Tracey D. Tuberville
Clinton T. Moore
Committee: Kurt A. Buhlmann
Jeff Hepinstall-Cymerman
Electronic Version Approved:
Ron Walcott
Interim Dean of the Graduate School
The University of Georgia
December 2019

iv
ACKNOWLEDGEMENTS
Any use of trade, firm, or product names is for descriptive purposes only and does not
imply endorsement by the United States Government. Funding for this research was
provided by the National Park Service (PSAC‐CESU Cooperative Agreement numbers
P08AC00193 and P17AC01606), California Energy Commission (Agreement numbers
500‐10‐20 and EPC-16-038), and by the U.S. Department of Energy under award DE-
FC09-07SR22506 and DE-EM0004391 to the University of Georgia Research
Foundation. All procedures were conducted in accordance with permits issued by the
U.S. Fish and Wildlife Service (TE-17838A-3), U.S. National Park Service (MOJA-
2018-SCI-0016 [under study MOJA-00258]), and California Department of Fish and
Wildlife (SC-0011221). Animal protocols were approved by the University of Georgia
under Animal Use Permits A2013 02-017-Y1-A0 and A2017 01-021-Y3-A3. I thank the
Warnell School of Forestry and Natural Resources at the University of Georgia for
funding through the Warnell Assistantship program. Roy C. Averill-Murray (USFWS -
Desert Tortoise Recovery Office) and Rebecca Jones (California Department of Fish and
Wildlife) provided assistance with permitting. Drs. Brian Todd and Mark Peaden
provided immeasurable support and commitment to the completion of this project. Jacob
Daly was instrumental in providing both field and analytical support. I also want to thank
Melia Nafus, Max Steele, Carmen Candal, and Collin Richter for their valuable
contributions in both husbandry and field work.

v
It was my choice of committee that made this project and my entire master’s
experience such an unforgettable time in my life. To Dr. Tracey Tuberville, a huge thank
you for the time and effort you put into making this thesis a valuable addition to the
conservation of the desert tortoise. The thoughtfulness and care that you place into each
detail of the scientific process is something I will draw from for the rest of my career. To
Dr. Kurt Buhlmann, I will always remember the many talks we shared on the excitement
and joy that all things nature bring to us both. There is still so much to be seen and
discovered, and I look forward to pursuing these countless opportunities with the
unceasing excitement with which you live life. You and Tracey have solidified my love
for turtles while expanding my skills and knowledge to really make a change! To Dr.
Clint Moore, I have been impressed with you since the first day I walked into your office,
and I will be forever thankful that you were willing to serve as my co-advisor. This
project benefitted greatly from your perspective and unrivaled skill. To Dr. Jeff
Hepinstall-Cymerman, incorporating your expertise was a crucial component of not only
this study but my growth as a scientist.
Dr. Mark Peaden (‘Marky mark’), you already know how much you’ve done for
me and this project, so instead my thanks is for the friend you have been since day one.
Having you and Hankster around made the field feel like home. A huge thanks to my lab
mates, all of whom embraced me into their lab and always had my back. I also want to
thank Adam Clause for showing me how to be a true professional in this field.
When contemplating the pursuit of this degree, it was the support of loved ones
that made the decision a no-brainer. My parents have supported me from day one and for

vi
that I will be forever grateful. Cam, your love and endless patience deserve so much
credit in the completion of this project.

vii
TABLE OF CONTENTS
Page
ACKNOWLEDGEMENTS ............................................................................................... iv
LIST OF TABLES ............................................................................................................. ix
LIST OF FIGURES .............................................................................................................x
CHAPTER
1 INTRODUCTION AND LITERATURE REVIEW .........................................1
Literature Cited ..........................................................................................10
2 COMPARING OUTDOOR ONLY REARING VERSUS A COMBINATION
OF INDOOR-OUTDOOR REARING FOR HEAD-STARTING THE
MOJAVE DESERT TORTOISE (GOPHERUS AGASSIZII) .........................19
Introduction ................................................................................................19
Methods......................................................................................................23
Results ........................................................................................................34
Discussion ..................................................................................................38
Literature Cited ..........................................................................................45
3 SIZE AS A PREDICTOR OF POST-RELEASE SURVIVAL FOR HEAD-
STARTED MOJAVE DESERT TORTOISES (GOPHERUS AGASSIZII) ....65
Introduction ................................................................................................65
Methods......................................................................................................69
Results ........................................................................................................78

viii
Discussion ..................................................................................................80
Literature Cited ..........................................................................................85
4 SUMMARY AND CONCLUSIONS ..............................................................99
Literature Cited ........................................................................................102

ix
LIST OF TABLES
Page
Table 2.1: Summary statistics of growth metrics for head-started Mojave desert tortoises
(Gopherus agassizii) reared under two different husbandry
treatments………………………………………………………………………...56
Table 2.2: Post-release movement metrics for head-started Mojave desert tortoises
(Gopherus agassizii) from two treatment groups (Combination head-started and
Outdoor head-started) released in September 2018 in the Mojave National
Preserve, CA, USA ................................................................................................57
Table 3.1: Candidate models used to evaluate which predictors (Age, MCL, Total
Displacement, Surface activity, Burrow switching) have the greatest effect on the
post-release fate of head-started Mojave desert tortoises released into the Mojave
National Preserve, CA, USA. ................................................................................94
Table 3.2: Predicted 10-month survival rates and 95% confidence intervals for juvenile
Mojave desert tortoises derived using a generalized linear model with fate as a
binary response variable and both MCL at release (mm) and proportion of surface
activity as predictor variables ................................................................................95

x
LIST OF FIGURES
Page
Figure 2.1: Midline carapace length (MCL) of 2016 cohort juvenile Mojave desert
tortoises (Gopherus agassizii) in two treatment groups ........................................58
Figure 2.2: Mass of 2016 cohort juvenile Mojave desert tortoises (Gopherus agassizii) in
two treatment groups..............................................................................................59
Figure 2.3: Body condition of 2016 cohort juvenile Mojave desert tortoises (Gopherus
agassizii) in two treatment groups .........................................................................60
Figure 2.4: Shell hardness index (SHI) versus midline carapace length (MCL) for two
treatment groups of juvenile Mojave desert tortoises (Gopherus agassizii)
compared after both one year and two years of pre-release head-starting. ............61
Figure 2.5: Shell hardness index (SHI) versus midline carapace length (MCL) for 2011,
2012, and 2016 cohort juvenile Mojave Desert Tortoises (Gopherus agassizii) in
September 2018 .....................................................................................................62
Figure 2.6: Mean distance moved from release site during pre-dormancy dispersal (left)
and mean distance moved from dormancy burrow after 10 months post-release
(right) by juvenile Mojave desert tortoises (Gopherus agassizii) from two
treatment groups.....................................................................................................63
Figure 2.7: Kaplan-Meier survival curves for head-started Mojave desert tortoises
(Gopherus agassizii) in two treatment groups released in the Mojave National
Preserve, CA, USA. ...............................................................................................64

xi
Figure 3.1: Map of release site with 78 release points for head-started Mojave desert
tortoises released in the Mojave National Preserve, CA ........................................96
Figure 3.2: Kaplan-Meier survival curve for head-started Mojave desert tortoises
(Gopherus agassizii) released into the Mojave National Preserve, CA, USA. ......97
Figure 3.2: Observed survival fates and predicted survival probabilities based on size
(MCL) at release for juvenile Mojave desert tortoises (Gopherus agassizii)
released into the Mojave National Preserve, CA, USA. ........................................98

1
CHAPTER 1
INTRODUCTION AND LITERATURE REVIEW
Wildlife species worldwide are experiencing severe population declines (Gibbons et al.
2000, Vie et al. 2009). Particularly at risk are turtles and tortoises, with nearly half of all
species threatened with extinction (Turtle Taxonomy Working Group 2017). Threats
include habitat loss and degradation (Dodd 1990, Gibbons et al. 2000, Gibbs and Shriver
2002), poaching for consumption and the illegal pet trade (Shiping et al. 2006), invasive
species (Buhlmann and Coffman 2001, Cadi and Joly 2004, Bolton and Brooks 2010),
subsidized predators (Browne and Hecnar 2007, Esque et al. 2010), and a changing
climate (Ihlow et al. 2012). The effects of these widespread threats are often exacerbated
by the suite of life history traits that characterize most chelonians (Congdon et al.
1993,1994, Tucker et al. 2001). Chelonians typically follow a pattern of slow growth,
delayed sexual maturity, high adult survivorship, and low rates of hatchling recruitment
(Iverson 1991, Turner et al. 1987, Congdon et al. 1993), traits which greatly increase
vulnerability to continued losses and contribute to markedly slow population recovery
(Congdon et al. 1994, Hall et al. 1999, Dorcas et al. 2007). Studies have shown that turtle
populations are predominantly reliant on and sustained by high rates of adult survival
(Doak et al. 1994, Heppell 1998). However, if strategies for mitigating losses to the adult
population are combined with efforts to bolster multiple life-stages, population recovery
may be expedited (Crawford et al. 2014, Spencer et al. 2017). In response, turtle
conservation programs have begun promoting and implementing management techniques

2
aimed at improving neonate and juvenile survival alongside efforts that reduce adult
mortality (Burke 2015, USFWS 2011, Crawford et al. 2014, Buhlmann et al. 2015).
Head-starting - the process of rearing offspring in captivity through their most
vulnerable life-stages to increase survivorship (Burke 2015) - is experiencing a recent
resurgence in turtle conservation after a period of discordant skepticism (Frazer 1992,
Heppell et al. 1996, Seigel and Dodd 2000). Head-starting programs have been
implemented for a variety of turtle species, including western pond turtles (Actinemys
marmorata; Haegen et al. 2009), redbelly turtles (Pseudemys rubriventris; Haskell et al.
1996), Blanding’s turtles (Emydoidea blandingii; Buhlmann et al. 2015), European pond
turtles (Emys orbicularis; Mitrus 2005), gopher tortoises (Gopherus polyphemus;
Tuberville et al. 2015), and Galapagos giant tortoises (Chelonoidis spp.; Milinkovitch et
al. 2013, Jensen et al. 2015). The controversial history of head-starting stems from its
expensive implementation, its concentrated focus in which only one element (juvenile
survival) of the larger conservation problem is addressed, and the limited post-release
monitoring data available to formally evaluate its effectiveness (Frazer 1992, Morafka et
al. 1997, Seigel and Dodd 2000). However, studies have shown that using head-starting
concurrently with other management initiatives can expedite population stability as well
as improve recovery (Crawford et al. 2014, Spencer et al. 2017). Solely using head-
starting to combat population declines without first ameliorating the causes of the
declines is ineffective; however, if used as part of a toolbox for managing population
viability it may play a vital role in conserving imperiled populations.
As part of the process of developing and refining species-specific head-starting
programs, evaluations should consider the effects of both captive and release conditions

3
on head-started animals. When augmenting wildlife populations through the release of
head-started individuals, habitat quality at the release site must be a priority (Griffith et
al. 1989). Habitat quality considers not only habitat requirements such as availability of
refugia, nest sites, and forage, but also more broadly whether predator abundances and
levels of intra- and interspecific competition approximate natural conditions. Through
ensuring that animals are released into optimal habitat, practitioners encourage fidelity to
the release site (Griffith et al. 1989, Larkin et al. 2004). In turn, increasing site fidelity
diminishes energetically costly movements away from the intended site, which is the
single greatest reason for failure in reptile release projects (Germano and Bishop 2009).
Long bouts of searching or dispersal from the release site, most often observed
immediately post-release, can also increase risk of environmental exposure and predation
(Hazard and Morafka 2002, Attum et al. 2010). Thus, post-release monitoring should
address both movement and survival following release.
Another important metric to consider when evaluating head-starting is individual
health. Individual health can be used to assess the quality of husbandry treatments but
may also be an important predictor of post-release performance and success (Booth and
Hixon 1999, Schmutz and Ely 1999, Garner et al. 2011). Evaluating pre-release health is
vital in avoiding disease transfer to previously unaffected populations (Woodford and
Rossiter 1994), as well as improving post-release survival by releasing individuals with
high body condition (Booth and Hixon 1999). Body condition, calculated as a ratio of
mass to some measure of length or volume, has been used extensively as a metric of
individual health (Stevenson et al. 2006). In tortoises specifically, a high body condition
translates to greater water balance and storage capacity, which in turn increases an

4
individual’s ability to withstand periods of drought (Nagy et al. 2002, Loehr et al. 2007).
For species inhabitating resource limited environments such as deserts, conservation of
limited water resources is integral for continued survival (Averill-Murray 2002).
The Mojave desert tortoise (Gopherus agassizii, hereafter ‘desert tortoise’ or
‘tortoise’) is a fossorial species endemic to the Mojave and Colorado deserts in the
southwest U.S.A. (Murphy et al. 2011). Desert tortoises confront a myriad of threats,
including habitat loss (Berry 1984), road mortality (Boarman and Sazaki 1996, Peaden et
al. 2015), anthropogenically subsidized predators (Esque et al. 2010), disease (USFWS
2011), and a changing climate (Lovich et al. 2014), all of which may act independently or
in concert (Foreman 1986, USFWS 2011). These threats have caused severe and
continued population declines (Allison and McLuckie 2018), leading to the development
and implementation of strategies aimed at recovering populations. Specific strategies
include fencing of roadways, habitat restoration, translocation of displaced wild
individuals, and head-starting (USFWS 2011). Head-starting, as previously mentioned,
focuses on improving survival of the vulnerable hatchling age-class (Burke 2015).
Annual survivorship and recruitment of hatchlings in the wild is naturally low, while
likely highly episodic (Gibbons 1987, Morafka 1994). Thus, management techniques that
focus on improving survival of this naturally at-risk age-class may help contribute to
population recovery (Heppell 1998, Tuberville et al. 2009).
Most formal head-starting programs for the desert tortoise have focused on
outdoor rearing in predator-proof enclosures in natural habitat (Morafka et al. 1997).
These initial projects sought to determine whether outdoor predator-proof enclosures
could be used to successfully rear juvenile desert tortoises (Morafka et al. 1997) and if

5
supplemental rain had an effect on growth or survivorship of tortoises while in captivity
(Nagy et al. 2015a). Annual survivorship of captive-reared juveniles has been shown to
be high (>90%) in outdoor predator-proof pens (Nagy et al. 2015a, Spangenberg 1996,
Tuberville et al. 2019). Supplemental rain increased growth during captivity and was
especially important in improving survival of older juveniles during drought conditions
(Nagy et al. 2015a). However, because desert tortoises living outdoors undergo both
winter dormancy and summer estivation, growth was still relatively slow, requiring 5-9
years of outdoor rearing for juveniles to reach a size considered to be predator-resistant
(Hazard et al. 2015, Nagy et al. 2015b). Post-release monitoring of juveniles found that a
minimum release size of 84-110 mm midline carapace length (MCL) conferred an
advantage in avoiding predation. These initial studies recommended use of supplemental
rain during captive rearing to increase growth and selecting release sites at least 546 m
from the head-start facility to preclude homing (Hazard et al. 2015, Nagy et al. 2015a,
Nagy et al 2015b).
In 2011, University of Georgia and University of California, Davis initiated a
joint long-term project in the Mojave National Preserve, CA to experimentally evaluate
head-starting as a recovery tool for the desert tortoise. Initial experiments at our site
focused on quantifying survival and growth differences between directly released
hatchling tortoises and tortoises head-started for 6-18 months under two different levels
of supplemental rain (Tuberville et al. 2019). Doubling the level of supplemented rain
significantly increased growth, although the modest improvement underscored the
lengthy head-starting period required for juveniles reared solely outdoors to attain
recommended release sizes (Tuberville et al. 2019). However, annual survival during

6
captivity averaged 94% across rain treatments (Tuberville et al. 2019), corroborating
findings from previous studies that annual survival of juvenile desert tortoises in outdoor
predator-proof pens is high (Nagy et al. 2015a, Spangenberg 1996).
Tuberville et al. (2019) monitored post-release survival of animals released at age
0-18 months, providing baseline estimates for future comparisons at our site. Although
the release sizes (40.8-61.5 mm MCL) were smaller than the release sizes recommended
in previous studies (Nagy et al. 2015b, Hazard et al. 2015), the goal was to incorporate an
experimental control (by including directly released hatchlings) and provide pilot data for
refining head-starting protocols. None of the factors examined - including size at release,
time of year released, maternal effects, or season - were found to have a significant effect
on post-release survival, with annual survival estimates averaging 48-49% across all
animals (Tuberville et al. 2019). Collectively, these results suggest that unmeasured
factors play an important role in survival of head-starts reared outdoors for only short
periods in captivity (6-18 mo). Alternatively, the effects of these factors may not have
been evident at the range of sizes compared.
Due to the considerable investment necessary to rear animals outdoors to
recommended release sizes, Tuberville et al. (2019) suggested that indoor rearing should
be investigated as a potential head-starting technique for desert tortoises. Indoor head-
starting has been used successfully with both aquatic and terrestrial turtle species
(Buhlmann et al. 2015, Green 2015, Quinn et al. 2018), and has the potential to greatly
increase the efficiency of head-starting programs. Animals reared indoors remain active
and feeding year-round and thus exhibit accelerated growth, leading to a decrease in the

7
duration of the head-start period required for animals to attain the recommended release
size.
A recent study by Daly et al. (2018) implemented this recommendation by
evaluating the effects of indoor head-starting on growth, body condition and shell
hardness in desert tortoises. The study compared direct released hatchlings to both
outdoor-reared and indoor-reared tortoises head-started for seven months, all from the
same year cohort, making it the first study to include an indoor husbandry component for
head-starting the Mojave desert tortoise. Indoor-reared tortoises grew over 16 times faster
than direct-released animals and over eight times faster than those reared in outdoor
predator-proof enclosures, clearly showing the growth advantage of foregoing
hibernation in a stable indoor environment (Daly et al. 2018). The study also evaluated
body condition and shell hardness for all released individuals. Body condition values for
solely indoor head-started tortoises were lower than those of their solely outdoor-reared
siblings, although still within the range of values calculated for wild desert tortoises
(Nagy et al. 2002). Shell hardness was compared between seven-month-old indoor-reared
tortoises and similarly sized but older (3-4yr old) outdoor-reared tortoises. Tortoises
reared indoors had softer shells than older tortoises raised outdoors, although this may be
the result of the shell ossification process needing to develop over a period greater than
that of the study (Daly et al. 2018, Nagy et al. 2011). In fact, shell hardness values for all
indoor animals were within the normal range for this species when compared to estimates
for wild tortoises of this age class (Daly et al. 2018, Nagy et al. 2011). Shell hardness,
which increases with both size and age, is a key metric in head-starting due to its
presumed correspondence with vulnerability to predation (Nagy et al. 2011), particularly

8
ravens (Kristan & Boarman 2003). Daly et al. (2019) evaluated the post-release survival
of these indoor head-starts, comparing them to both directly released hatchlings and
animals head-started solely outdoors for seven months. Treatment groups exhibited no
difference in post-release survival (Daly et al. 2019). In contrast to a previous release at
the same site, in which only one mortality (2.2% of animals released) was attributed to
avian predation (Tuberville et al. 2019), common ravens (Corvus corax) were found to be
the main cause of death (26.5% of animals released) for head-starts released by Daly et
al. (2019). To reduce the risk of raven predation, Daly et al. (2019) recommended
releasing head-starts in the fall to avoid brooding ravens and at least 1.6 km from possible
raven nesting structures (powerlines, billboards, etc.).
To follow up on the findings of these prior studies and to continue evaluating
head-starting as a viable management technique for the Mojave desert tortoise, this study
incorporated the use of combination head-starting. Combination head-starting (hereafter
“combo” head-starting), as defined in this study, is the combination of initial rearing
indoors followed by outdoor rearing prior to release. This afforded “combo” head-started
tortoises the advantage of increased growth during the indoor phase while also allowing
them time outdoors for increased shell ossification, as well as physical and behavioral
acclimatization to their natural environment. The objectives of my thesis research are to
evaluate this novel head-starting method by comparing pre- and post-release metrics of
success between two year old “combo” head-started tortoises, tortoises of their same age
but smaller size reared solely outdoors, and older tortoises of the same size reared solely
outdoors. In Chapter 2, I assigned animals from the same cohort to one of two treatments:
1) “combo” head-started animals reared indoors for one year followed by outdoors for

9
one year, and 2) animals reared solely outdoors for two years. I compare their pre-release
morphometrics, body condition, and shell hardness as well as their post-release
movement and survival for 10 months following release. In Chapter 3, I compare survival
and release size between animals reared solely outdoors for different head-starting
durations (two years or 6-7 years) to refine the recommended size at release for this
threatened species. In the concluding chapter (Chapter 4), I synthesize the results of my
research, place them in the context of previous findings, and make recommendations for
their implementation in future head-starting programs for desert tortoises. Given the dire
condition of turtle populations worldwide (Turtle Taxonomy Working Group 2017),
contributing to the knowledge of turtle population augmentations is critical for future
conservation efforts.

10
LITERATURE CITED
Attum, O., W.E. Farag, S.M.B. El Din, and B. Kingsbury. 2010. Retention rate of hard-
released translocated Egyptian tortoises Testudo kleinmanni. Endangered Species
Research 12:11-15.
Averill-Murray, R. C. 2002. Effects on survival of desert tortoises (Gopherus agassizii)
urinating during handling. Chelonian Conservation and Biology 4:430-435.
Berry, K.H. 1984. The status of the desert tortoise (Gopherus agassizii) in the United
States. Desert Tortoise Council Report to the U.S. Fish and Wildlife Service,
Sacramento, California, USA.
Boarman, W.I., and M. Sazaki. 1996. Highway mortality in desert tortoises and small
vertebrates: success of barrier fences and culverts. Pages 169-173 in G.J. Evink,
P. Garrett, D. Zeigler, and J. Berry (eds.), Trends in addressing transportation
related wildlife mortality seminar. Environmental Management Office, U.S.
Department of Transportation, Tallahassee, Florida.
Bolton, R. M., and R.J., Brooks. 2010. Impact of the seasonal invasion of Phragmites
australis (common reed) on turtle reproductive success. Chelonian Conservation
and Biology 9:238-243.
Booth, D. J., and M.A., Hixon. 1999. Food ration and condition affect early survival of
the coral reef damselfish, Stegastes partitus. Oecologia 121:364-368.
Browne, C. L., and S.J., Hecnar. 2007. Species loss and shifting population structure of
freshwater turtles despite habitat protection. Biological Conservation 138:421-
429.

11
Buhlmann, K.A. and G. Coffman. 2001. Fire ant predation of turtle nests and
implications for the strategy of delayed emergence. Journal of the Elisha Mitchell
Scientific Society 94-100.
Buhlmann, K. A., S. L. Koch, B. O. Butler, T. D. Tuberville, V. J. Palermo, B. A.
Bastarache, and Z. A. Cava. 2015. Reintroduction and head-starting: tools for
Blanding’s Turtle (Emydoidea blandingii) conservation. Herpetological
Conservation and Biology 10:436-454.
Burke, L. R. 2015. Head-starting turtles: learning from experience. Herpetological
Conservation and Biology 10:299–308.
Cadi, A., and P., Joly. 2004. Impact of the introduction of the red-eared slider
(Trachemys scripta elegans) on survival rates of the European pond turtle (Emys
orbicularis). Biodiversity and Conservation 13:2511–2518.
Congdon, J.D., A.E. Dunham, and R.C. van Loben Sels. 1993. Delayed sexual maturity
and demographics of Blanding’s turtles (Emydoidea blandingii): implications for
conservation and management of long-lived organisms. Conservation Biology
7:826-833.
Congdon, J.D., A.E. Dunham, and R.C. van Loben Sels. 1994. Demographics of common
snapping turtles (Chelydra serpentina): implications for conservation and
management of long-lived organisms. American Zoologist 34:397–408.
Crawford, B.A., J.C. Maerz, N.P. Nibbelink, K.A. Buhlmann, and T.M Norton. 2014.
Estimating the consequences of multiple threats and management strategies for
semi‐aquatic turtles. Journal of Applied Ecology 51:359-366.
Daly, J.A., K.A. Buhlmann, B.D. Todd, C.T. Moore, J.M. Peaden, and T.D. Tuberville.

12
2018. Comparing growth and body condition of indoor-reared, outdoor-reared,
and direct-released juvenile Mojave desert tortoises. Herpetological Conservation
and Biology 13:622-633.
Daly, J.A., K.A. Buhlmann, B.D. Todd, C.T. Moore, J.M. Peaden, and T.D. Tuberville.
2019. Survival and movements of head-started Mojave desert tortoises. The
Journal of Wildlife Management. In review.
Dodd Jr, C. K. 1990. Effects of habitat fragmentation on a stream-dwelling species, the
flattened musk turtle Sternotherus depressus. Biological Conservation 54:33-45.
Dorcas, M.E., J.D. Willson, and J.W. Gibbons. 2007. Crab trapping causes population
decline and demographic changes in diamondback terrapins over two
decades. Biological Conservation 137:334-340.
Esque, T.C., K.E. Nussear, K.K. Drake, A.D. Walde, K.H. Berry, R.C. Averill-Murray,
A.P. Woodman, W.I. Boarman, P.A. Medica, J. Mack, and J.S. Heaton. 2010.
Effects of subsidized predators, resource availability, and human population
density on desert tortoise populations in the Mojave Desert, USA. Endangered
Species Research 12:167-177.
Foreman, L.D., J.M. Brode, R. Haussler, and K. Kramer. 1986. The responsibilities of
federal and state agencies for protection of the desert tortoise in California.
Herpetologica 1:59.
Frazer, N.B. 1992. Sea turtle conservation and halfway technology. Conservation
Biology 6:179-184.

13
Garner, T.W., J.M. Rowcliffe, and M.C. Fisher. 2011. Climate change, chytridiomycosis
or condition: an experimental test of amphibian survival. Global Change
Biology 17:667-675.
Germano, J. M., and P.J., Bishop. 2009. Suitability of amphibians and reptiles for
translocation. Conservation Biology 23:7-15.
Gibbons, J.W., 1987. Why do turtles live so long? BioScience 37:262-269.
Gibbons, J. W., D.E. Scott, T.J. Ryan, K.A. Buhlmann, T.D. Tuberville, B.S. Metts, J.L.
Greene, T. Mills, Y. Leiden, S. Poppy, and C.T. Winne. 2000. The global decline
of reptiles, déjà vu amphibians. BioScience 50:53-666.
Gibbs, J.P. and W.G. Shriver. 2002. Estimating the effects of road mortality on turtle
populations. Conservation Biology 16:1647-1652.
Griffith, B., J.M. Scott, J.W. Carpenter, and C. Reed. 1989. Translocation as a species
conservation tool: status and strategy. Science 245:477-480.
Hall, R.J., P.F. Henry, and C.M. Bunck. 1999. Fifty-year trends in a box turtle population
in Maryland. Biological Conservation 88:165-172.
Haskell, A., T.E. Graham, C.R. Griffin, and J.B. Hestbeck. 1996. Size related survival of
headstarted redbelly turtles (Pseudemys rubriventris) in Massachusetts. Journal of
Herpetology 30:524-527.
Hazard, L., and D. Morafka. 2002. Comparative dispersion of neonate and headstarted
juvenile desert tortoises (Gopherus agassizii): A preliminary assessment of age
effects. Chelonian Conservation and Biology 44:135-147.
Hazard, L.C., D.J. Morafka, and L.S. Hillard. 2015. Post-release dispersal and predation
of head-started juvenile desert tortoises (Gopherus agassizii): effect of release site

14
distance on homing behavior. Herpetological Conservation and Biology 10:504-
515.
Heppell, S.S., L.B. Crowder, and D.T. Crouse. 1996. Models to evaluate head-starting as
a management tool for long-lived turtles. Ecological Applications 6:556-565.
Heppell, S.S. 1998. Application of life-history theory and population model analysis to
turtle conservation. Copeia 1998:367-375.
Ihlow, F., J. Dambach, J.O. Engler, M. Flecks, T. Hartmann, S. Nekum, and D. Rödder.
2012. On the brink of extinction? How climate change may affect global
chelonian species richness and distribution. Global Change Biology 18:1520-
1530.
Iverson, J.B. 1991. Patterns of survivorship in turtles (order Testudines). Canadian
Journal of Zoology 69:385-391.
Jensen, E.L., W. Tapia, A. Caccone, and M.A. Russello. 2015. Genetics of a head-start
program to guide conservation of an endangered Galápagos tortoise (Chelonoidis
ephippium). Conservation Genetics 16:823-832.
Kristan, W.B., and W.I. Boarman. 2003. Spatial pattern of risk of common raven
predation on desert tortoises. Ecology 84:2432-2443.
Larkin, J.L., J.J. Cox, M.W. Wichrowski, M.R. Dzialak, and D.S. Maehr. 2004.
Influences on release‐site fidelity of translocated elk. Restoration Ecology 12:97-
105.
Loehr, V.J.T., M.D. Hofmeyr, and B.T. Henen. 2007. Annual variation in the body
condition of a small, arid-zone tortoise, Homopus signatus signatus. Journal of
Arid Environments 71:337–349.

15
Lovich, J. E., C.B. Yackulic, J. Freilich, M. Agha, M. Austin, K.P. Meyer, and S.A. Root.
2014. Climatic variation and tortoise survival: has a desert species met its
match? Biological Conservation 169:214-224.
Lovich, J.E., J.R. Ennen, C.B. Yackulic, K. Meyer-Wilkins, M. Agha, C. Loughran, C.
Bjurlin, M. Austin, and S. Madrak. 2015. Not putting all their eggs in one basket:
bet-hedging despite extraordinary annual reproductive output of desert
tortoises. Biological Journal of the Linnean Society 115:399-410.
Milinkovitch, M.C., R. Kanitz, R. Tiedemann, W. Tapia, F. Llerena, A. Caccone, J.P.
Gibbs, and J.R. Powell. 2013. Recovery of a nearly extinct Galápagos tortoise
despite minimal genetic variation. Evolutionary Applications 6:377–383.
Mitrus, S. 2005. Headstarting in European pond turtles (Emys orbicularis): Does it
work? Amphibia-Reptilia 26:333-341.
Morafka, D.J., K.H. Berry, and E.K. Spangenberg. 1997. Predator-proof field enclosures
for enhancing hatchling success and survivorship of juvenile tortoises: a critical
evaluation. Pages 147–165 in J. Van Abbema, editor. Proceedings: Conservation,
Restoration, and Management of Tortoises and Turtles: An International
Conference Conservation. Wildlife Conservation Society Turtle Recovery
Program and the New York Turtle and Tortoise Society, New York, New York,
USA.
Murphy, R.W., K.H. Berry, T. Edwards, A.E. Leviton, A. Lathrop, and J.D. Riedle. 2011.
The dazed and confused identity of Agassiz’s land tortoise, Gopherus agassizii
(Testudines, Testudinidae) with the description of a new species, and its
consequences for conservation. ZooKeys 113:39–71.

16
Nagy, K.A., B.T. Henen, D.B. Vyas, and I.R. Wallis. 2002. A condition index for the
desert tortoise (Gopherus agassizii). Chelonian Conservation and Biology 4:425–
429.
Nagy, K., M. Tuma, and L. Hillard. 2011. Shell hardness measurements in juvenile desert
tortoises Gopherus agassizii. Herpetological Review 42:191–195.
Nagy, K.A., L.S. Hillard, S. Dickson, and D.J. Morafka. 2015a. Effects of artificial rain
on survivorship, body condition, and growth of head-started desert tortoises
(Gopherus agassizii) released to the open desert. Herpetological Conservation and
Biology 10(Symposium):535-549.
Nagy, K.A., L.S. Hillard, M.W. Tuma, and D.J. Morafka. 2015b. Head-started desert
tortoises (Gopherus agassizii): movements, survivorship and mortality causes
following their releases. Herpetological Conservation and Biology 10:203-215.
Peaden, J.M., T.D. Tuberville, K.A. Buhlmann, M.G. Nafus, and B.D. Todd. 2015.
Delimiting road-effect zones for threatened species: implications for mitigation
fencing. Wildlife Research 42:650–659.
Schmutz, J.A., and C.R. Ely. 1999. Survival of greater white-fronted geese: effects of
year, season, sex, and body condition. The Journal of wildlife management
63:1239-1249.
Seigel, R.A., and C.K. Dodd, Jr. 2000. Manipulation of turtle populations for
conservation: halfway technologies or viable options? Pp. 218-238 In Turtle
Conservation. Klemens, M.W. (Ed.). Smithsonian Institution Press, Washington,
DC, USA.
Shiping, G., W. Jichao, S. Haitao, S. Riheng, and X. Rumei. 2006. Illegal trade and

17
conservation requirements of freshwater turtles in Nanmao, Hainan Province,
China. Oryx, 40:331-336.
Spencer, R. J., J.U. Dyke, and M.B. Thompson. 2017. Critically evaluating best
management practices for preventing freshwater turtle extinctions. Conservation
Biology 31:1340-1349.
Stevenson, R.D., and W.A. Woods Jr. 2006. Condition indices for conservation: new uses
for evolving tools. Integrative and comparative biology 46:1169-1190.
Tuberville, T.D., M.T. Norton, K.A. Buhlmann, and V. Greco. 2015. Head-starting as a
management component for gopher tortoises (Gopherus polyphemus).
Herpetological Conservation and Biology 10:455–471.
Tuberville, T.D., K.A. Buhlmann, R. Sollmann, M.G. Nafus, J.M. Peaden, J.A. Daly, and
B.D. Todd. 2019. Effects of short-term head-starting on growth and survival in
the Mojave desert tortoise (Gopherus agassizii). Herpetological Conservation and
Biology 14:171–184.
Tucker, A.D., J.W. Gibbons, and J.L. Greene. 2001. Estimates of adult survival and
migration for diamondback terrapins: conservation insight from local extirpation
within a metapopulation. Canadian Journal of Zoology 79:2199-2209.
Turner, F., P.A. Medica, and R.B. Bury. 1987. Age-size relationships of desert tortoises
(Gopherus agassizii) in southern Nevada. Copeia 4:974-979.
Turtle Taxonomy Working Group. 2017. Turtles of the world, 7th edition: annotated
checklist of taxonomy, synonymy, distribution with maps, and conservation
status. Pages 329–479 in Rhodin, A.G.J., Pritchard, P.C.H., van Dijk, P.P.,
Saumure, R.A., Buhlmann, K.A., Iverson, J.B., and R.A. Mittermeier, R.A.,

18
editors. Conservation Biology of Freshwater Turtles and Tortoises: A
Compilation Project of the IUCN/SSC Tortoise and Freshwater Turtle Specialist
Group. Chelonian Research Monographs.
USFWS 2011. Revised recovery plan for the Mojave population of the desert tortoise
(Gopherus agassizii). U.S. Fish and Wildlife Service, Pacific Southwest Region,
Sacramento, California, USA.
Vié, J.C., C. Hilton-Taylor, and S.N. Stuart. 2009. Wildlife in a Changing World–An
Analysis of the 2008 IUCN Red List of Threatened Species. International Union
for Conservation of Nature, Gland, Switzerland.
Woodford M.H., and P.B. Rossiter. 1994. Disease risks associated with wildlife
translocation projects. In: Olney P.J.S., Mace G.M., Feistner A.T.C. (eds)
Creative Conservation 178-200. Springer, Dordrecht

19
CHAPTER 2
COMPARING OUTDOOR ONLY REARING VERSUS A COMBINATION OF
INDOOR-OUTDOOR REARING FOR HEAD-STARTING THE MOJAVE DESERT
TORTOISE (GOPHERUS AGASSIZII)
INTRODUCTION
Many wildlife populations are in severe decline globally due to anthropogenic
threats (Gibbons et al. 2000, Loh et al. 2008, Vie et al. 2009). In response,
conservationists and management agencies have implemented diverse management
strategies to recover populations (Myers et al. 2000, Lindenmayer et al. 2006, USFWS
2011, Haag et al. 2014). While strategies aimed at acquiring and protecting natural
habitat remain paramount in conserving wildlife, species with depleted populations often
require additional interventions (Ellis et al. 2000, Jachowski et al. 2009, Milinkovitch et
al. 2013). Head-starting is an approach in which offspring are protected through their
early life stages before release at a size presumed to be less prone to mortality (USFWS
2011, Burke 2015). When used alongside management techniques that mitigate the
original causes of decline, head-starting has the potential to expedite population recovery
(Tomillo et al. 2008, Crawford et al. 2014, Spencer et al. 2017). Head-starting has been
used in the recovery of a variety of taxa (e.g., Mona Island iguanas [Cyclura cornuta
stejnegeri], Pérez-Buitrago et al. 2008; Philippine crocodiles [Crocodylus mindorensis],
van de Ven et al. 2009; mangrove finches [Camarhynchus heliobates], Cunninghame et
al. 2015; and hellbenders [Cryptobrancus alleganiensis], Crane and Mathis 2011), but to
increase success and efficiency, it is important to continue evaluating, improving, and
customizing this management technique for each at-risk species.

20
The Mojave desert tortoise (Gopherus agassizii) is a keystone species of the
Mojave and Colorado deserts of the southwest U.S.A. that has experienced severe
population declines (Luckenbach 1982, Murphy et al. 2011, USFWS 2011, Allison and
McLuckie 2018) and has been classified as threatened under the U.S. Endangered Species
Act. Populations of this desert endemic are affected by habitat loss, road mortality,
subsidized predators, and the direct and indirect effects of changing climate (Berry 1986,
Barrows 2011, USFWS 2011, Peaden et al. 2015). To combat these threats and recover
depleted populations, a wide range of management strategies has been suggested and
employed (USFWS 2011), including head-starting.
The Mojave desert tortoise has been the subject of head-starting efforts for nearly
30 years (Morafka et al. 1997). Initial projects involved the rearing of hatchlings in
outdoor predator-proof enclosures situated in natural habitat to evaluate the effects of
captive rearing enclosures on nest success and hatchling survival and to gather insight
into unknown aspects of juvenile tortoise ecology (Morafka et al. 1997). Juvenile survival
in predator-proof enclosures was high (>90%; Morafka et al. 1997, Spangenberg 1996),
leading to additional head-starting projects (Nagy et al. 2015a, Tuberville et al. 2019).
These subsequent projects incorporated the use of supplemental rain in outdoor
enclosures to increase growth and improve survival during periods of natural drought.
Recent studies continue to build on these earlier findings to identify possible avenues for
further improvement of these metrics.
Slow growth rates have burdened conventional outdoor captive-rearing efforts for
the desert tortoise (Hazard et al. 2015, Nagy et al. 2015a). Desert tortoises spend roughly
95% of their lives below ground during the thermal extremes of winter and summer

21
(Woodbury and Hardy 1948, Ernst and Lovich 2009). These prolonged bouts of inactivity
during inhospitable ambient conditions result in restricted opportunities for foraging and
growth (Medica et al. 2012). Previous studies have found that 5–9 years of outdoor
captive-rearing are required to raise juveniles to a size that confers a survival advantage
upon release into the wild (84-110 mm midline carapace length (MCL); Nagy et al. 2011,
Hazard et al. 2015, Nagy et al. 2015a). To reduce this extended captivity, indoor captive-
rearing has recently been used to enhance growth by allowing head-starts to remain
active and foraging year-round (Daly et al. 2018). The expedited growth exhibited by
tortoises reared under this technique was significant, producing 7-month-old captive-
reared tortoises equivalent in size to 5-6-year-old wild animals (Daly et al. 2018).
However, indoor captive-reared tortoises had softer shells and slightly lower body
condition compared to older animals (3-4 yrs old) of similar size reared solely outdoors
(Daly et al. 2018). Rearing hatchlings solely indoors also holds the potential of raising
tortoises naïve to their surroundings upon release into the wild (Grueber et al. 2017).
Incorporating an outdoor rearing component may serve as pre-release enrichment by
providing head-starts with native forage, opportunities for burrowing, and exposure to
natural environmental cues. Environmental enrichment has been shown to improve the
survival of captive reared animals released to the wild (Tetzlaff et al. 2019). In this study,
we introduce the use of combination head-starting, defined as initial captive-rearing
indoors followed by conventional outdoor captive-rearing prior to release. This novel
approach maximizes tortoise growth during indoor rearing and exposes tortoises to
environmental cues during outdoor rearing. The outdoor rearing period should also
promote shell ossification through exposure to natural UV light (Acierno et al. 2006).

22
We use shell hardness and body condition as metrics for monitoring the possible
effects of accelerated growth during the captive period in desert tortoises. Shell hardness
has been found to increase with both body size and age in juvenile desert tortoises (Nagy
et al. 2011, Daly et al. 2018). Although we did not find studies showing a direct
correlation between shell hardness and increased survival, an increase in shell hardness
presumably reduces the vulnerability to certain predators, such as ravens (Corvus corax;
Kristan and Boarman 2003, Nagy et al. 2015b). Body condition, an estimate of body
density, has been used as a physiological metric of health in previous tortoise studies
(Nagy et al. 2002, Loehr et al. 2007, McCoy et al. 2011). Nagy et al. (2015a) found that
increased body condition from supplemental watering resulted in higher survival in
outdoor predator proof enclosures during a natural drought. Body condition values may
also increase concurrently with the shell ossification process (as harder shells weigh
more), highlighting the value of monitoring both metrics.
Long-distance dispersal away from the release site is the primary reason for
failure in herpetofaunal release programs (Germano and Bishop 2009). Thus, as a
possible indicator of short-term success, we assessed post-release movement. For head-
starting to ultimately succeed, released animals must survive to maturity and reproduce
successfully (Pritchard 1981). However, monitoring a released juvenile to reproductive
age may take over 10 years post-release. As a more immediate metric of this ultimate
goal, we monitored survival during the first 10 months post-release.
In this study, we compared two head-starting treatments for the Mojave desert
tortoise: 1) solely outdoor captive-rearing for two years; and 2) combination head-
starting, consisting of one year of indoor captive-rearing followed by one year of outdoor

23
captive-rearing. We compared size, body condition and shell hardness between the two
treatment groups using animals from the same year cohort. We also compared movement
and survival from release to dormancy and during the first 10 months post-release.
Ultimately, by reducing the duration of the captive rearing period and decreasing the cost
of implementing this technique, combination head-starting may increase the efficiency
and success of population augmentation efforts.
METHODS
Study Site
This study was conducted in the Ivanpah Valley in the Mojave National Preserve
(MNP), San Bernardino County, California, which is within the Eastern Mojave recovery
unit of the Mojave desert tortoise (Nussear et al. 2009, USFWS 2011). The husbandry
portions of this study were conducted at the Ivanpah Desert Tortoise Research Facility
(IDTRF) at 820 m elevation. The release site, located approximately 15 km away,
occurred at 1000-1100 m elevation. The release site was dominated by Yucca woodland
habitat composed primarily of creosote (Larrea tridentata) and white bursage (Ambrosia
dumosa), with Yucca (Y. brevifolia, Y. shidigera), pencil cholla (Cylindropuntia
ramosissima), and littleleaf ratany (Krameria erecta) heavily intermixed (Todd et al.
2016). Limited anthropogenic disturbances at the release site included abandoned cattle
corrals, a powerline, and seldom-used dirt roads.
Obtaining hatchlings
In April 2011, we initiated the IDTRF head-starting program by capturing,
attaching transmitters (RI-2B, Holohil Systems Ltd., Ontario, Canada), and radio-tracking
30 adult female desert tortoises in the Ivanpah Valley of the MNP. We tracked females at

24
least monthly using a 3-element Yagi antenna (AF Antronics, Inc., Urbana, IL) and a
R1000 receiver (Communications Specialists, Inc., Orange, CA). In April 2016, we
brought all radio-tracked females to the IDTRF and radiographed them to check for the
presence of calcified eggs (Diagnostic Imaging Systems, Poskam, Colorado, USA; 60
kvp, 0.8 mAS, 74 cm focal length; Gibbons and Greene 1979). We immediately released
all non-gravid females at their point of capture. We placed gravid females in individual
nesting enclosures and allowed them to nest naturally. These predator-proof nesting
enclosures measured 5 x 9 m and included pre-made burrows, natural cover and forage,
as well as sprinkler systems for supplemental watering (Tuberville et al. 2019). After
initial placement in the enclosures, we radiographed females weekly to monitor for egg
deposition. If nesting was confirmed, we immediately released females at their point of
capture. In accordance with our permits, we released all females that did not nest within
30 days at their last known burrow location.
We allowed eggs to incubate naturally in the predator-proof enclosures.
Approximately 80 days after the estimated nesting date, we began searching pens daily
for emerged hatchlings. We removed hatchings as they emerged from nests and marked
them by notching unique combinations of marginal scutes using numerical codes
assigned by USFWS (modified from Cagle 1939). We also searched nests for any
unhatched eggs or unemerged hatchlings. After all hatchlings from the 2016 cohort had
emerged and been uniquely marked, we used a stratified random design to assign all
healthy neonates (evenly separating siblings between treatments; Daly et al. 2018,
Tuberville et al. 2019) from our 2016 cohort to one of two treatment groups: solely
outdoor head-started (hereafter Outdoor HS) or combination (indoor + outdoor) head-

25
started (hereafter Combo HS). Both treatment groups were reared in captivity for two
years prior to release.
Outdoor HS
On 07 September 2016, we moved the 38 hatchlings assigned to the Outdoor HS
treatment to outdoor predator-proof enclosures at the IDTRF. Captive rearing enclosures
measured 9 m x 9 m and included pre-made starter burrows (53-cm long, 10-cm diameter
perforated plastic pipes buried at 20o angles), natural cover and forage, as well as rotating
sprinkler systems (Nafus et al. 2015b, Daly et al. 2018, Tuberville et al. 2019). We placed
a maximum of 10 animals into each pen (0.125 tortoises/m2), and siblings from the same
clutch were separated into different pens. We provided supplemental food as well as
artificial rain once every 1-2 weeks for 30-min during the active season (March-October).
Rain supplementation was provided using rotating garden sprinklers to sustain native
plant growth (Beatley 1974), allow hatchlings to drink, and to encourage tortoises to feed
on supplemental food.
Tortoises were fed a mixture of leafy greens and water-soaked Mazuri® Tortoise
Diet (Mazuri Exotic Animal Nutrition, St. Louis, MO). The greens included dandelion
(Taraxacum officinale), mustard greens (Brassica juncea), turnip greens (Brassica rapa),
collards (a cultivar of Brassica oleracea), endive (Cichorium endivia), and escarole (C.
endivia latifolia). The weekly combination of greens varied by availability, though
always consisted of at least three different types in an attempt to approximate the
nutritional properties of the desert tortoise’s natural diet (Jarchow et al. 2002). We
monitored the amount provided to each pen to avoid attracting ants and rodents with
excess food.

26
Combo HS
We reared 30 Combo HS tortoises in mesocosms in the indoor climate-controlled
IDTRF for the first year (September 2016-September 2017) before moving them to an
outdoor predator-proof enclosure for the second year (September 2017-September 2018).
We set ambient temperature inside the IDTRF to a constant 24.4 °C. Indoor mesocosms
were constructed from 50-gallon (189 L) Rubbermaid (Atlanta, Georgia, USA) stock
tanks (132 x 79 x 30.5 cm) and lined with natural substrate (Daly et al. 2018). We
prepared six identical tanks, each of which held five tortoises (n = 30). Each tank
contained three hides made from longitudinally halved plastic pipe (11.5 cm in diameter
and 12-cm in length), a paper feeding plate, and a humid hide box. Humid hide boxes
have been shown to promote smooth shell growth in tortoises (Wiesner and Iben 2003).
Hide boxes were made from Rubbermaid Roughneck (Atlanta, Georgia, USA) tote boxes
(40 x 26 x 18 cm) and filled with approximately 4 cm of peat moss, which we moistened
every 3-4 days and replaced every two weeks. Tortoises were able to enter the hide box
through a burrow-shaped entrance hole cut into each tote. We suspended Mini Combo
Deep Dome Dual Lamp Fixtures (ZooMed Laboratories Inc., San Luis Obispo,
California) 30 cm over each tank with a 50 W ZooMed Repti Basking Spot Lamp bulb
for daytime basking (37 °C) and a ZooMed 50 W Infrared Basking Spot bulb for night-
time heat (32 °C). In addition to the dual basking lights, we provided each tub with a 26
W Exo-Terra Reptile UVB150 bulb (45 cm above substrate; Rolf C. Hagen Corp.,
Mansfield, MA., U.S.A) for optimal calcium metabolism and D3 conversion. We
connected all lights to automatic timers, regulated monthly to reflect the natural

27
photoperiod. Indoor conditions were held constant to allow juveniles to remain active and
forage year-round (Diez et al. 2009).
We fed the Combo HS group ad libitum 4-5 times per week during the indoor
rearing period (Diez et al. 2009), using the same food mixture as described above for the
Outdoor HS treatment. Twice per week, we supplemented their food with a light dusting
of Rep-Cal Calcium with Vitamin D3 (Rep-Cal Research Labs, Los Gatos, California).
We soaked Combo HS tortoises weekly for 15-30 minutes in 1-2 cm of water. In
September 2017, we moved all Combo HS tortoises outdoors to a single 10 m x 30 m
predator-proof enclosure at the IDTRF for their second year of rearing. This larger
enclosure contained all of the previously mentioned enclosure components (refugia,
natural vegetation, sprinklers) while maintaining a similar tortoise density (0.11
tortoises/m2) to the Outdoor HS treatment. Supplemental rain was provided weekly
during the activity season (March – October) in this pen.
Morphometrics
We weighed and measured (hereafter ‘measured’) all hatchlings immediately after
emergence from the nest. We then measured Outdoor HS tortoises once each spring and
fall during the captive rearing period. We measured Combo HS tortoises monthly while
indoors (September 2016-September 2017) and then on the same cycle as the Outdoor
HS treatment thereafter. We recorded mass to the nearest 0.1 g (Model HH320, OHAUS
Corporation, Parsippany, NJ, USA); when tortoise mass exceeded 300 g, we recorded
mass to the nearest 1 g (My Weigh 6001, HBI Technologies, Phoenix, AZ, USA). We
recorded midline carapace length (MCL), maximum shell height, and maximum shell

28
width on the bridge to the nearest 0.1 mm using dial calipers (Series 505, Mitutoyo,
Aurora, IL, USA).
Body Condition
We calculated body condition (BC) for all animals from both treatment groups
using measurements taken at hatching (‘initial’), one-year post-hatching (‘Year 1’; when
the Combo HS group was transitioned outdoors), and immediately prior to release
(‘release’; 2 years post-hatching) based on the formula described by Nagy et al. (2002):
𝐵𝑜𝑑𝑦 𝑐𝑜𝑛𝑑𝑖𝑡𝑖𝑜𝑛 (𝑔
𝑐𝑚3) =
𝑤𝑒𝑖𝑔ℎ𝑡 (𝑔)
𝑠ℎ𝑒𝑙𝑙 𝑣𝑜𝑙𝑢𝑚𝑒 (𝑐𝑚3)
𝑆ℎ𝑒𝑙𝑙 𝑣𝑜𝑙𝑢𝑚𝑒 (𝑐𝑚3) = 𝑀𝐶𝐿(𝑐𝑚) × 𝑤𝑖𝑑𝑡ℎ(𝑐𝑚) × ℎ𝑒𝑖𝑔ℎ𝑡(𝑐𝑚)
Shell Hardness
We measured shell hardness of all juveniles (n = 48) after one year of captive
rearing (September 2017) and again just prior to release (September 2018). To calculate
shell hardness index (SHI) we first used a 4-inch (10.2 cm) tension-calibrated micrometer
(L.S. Starrett Company, Athol, Massachusetts, USA; model: 3732XFL-4) to measure
each tortoise’s uncompressed shell height (UCSH) at the center of the third vertebral
scute (Nagy et al. 2011). We then measured compressed shell height (CSH) by turning
the micrometer spindle and compressing each tortoise’s shell between the two measuring
faces until the spindle ratchet slipped continually for approximately 270 degrees (Daly et
al. 2018). We used these measurements to calculate a shell hardness index as described
by Nagy et al. (2011), in which a SHI value of 100 corresponds to maximum hardness
(i.e., an incompressible shell):
𝑆ℎ𝑒𝑙𝑙 𝐻𝑎𝑟𝑑𝑛𝑒𝑠𝑠 𝐼𝑛𝑑𝑒𝑥 (𝑆𝐻𝐼) =𝐶𝑆𝐻
𝑈𝐶𝑆𝐻 × 100

29
Experimental Releases
We released the 2016 cohort Outdoor HS (n = 24) and Combo HS animals (n = 24)
concurrently and in the same area as another release group of older solely outdoor-reared
juveniles (n = 30; 2011-2012 cohorts), discussed separately in Chapter 3. Our release
location was chosen based on the 1.6 km minimum recommended distance from raised
structures such as powerlines that could serve as perching sites for ravens (Daly et al.
2019), accessibility for frequent tracking, and habitat quality (Baxter 1988, Nafus et al.
2015a, Todd et al. 2016). At our study site, juvenile desert tortoises select for
microhabitats with sandy soils, high density perennial shrubs, abundant rodent burrows,
and washes (Todd et al. 2016). Washes have been shown to influence movement (Nafus
et al. 2017, Peaden et al. 2017) and provide increased forage (Jennings and Berry 2015).
Using ArcGIS (version 10.5; ESRI, Redlands, CA), we first created a 0.6-km2 rectangular
release plot (300 m X 2000 m) parallel to an access road. We then used the ‘Fishnet’
function to generate release points every 50 m throughout the plot. From there, we set an
interior buffer (buffer in from plot edge) of 50 m and removed all points within the
buffered area. We then selectively removed interior points to create three equally spaced
blocks 300 m apart and each containing 26 release points for a total of 78 release points.
We buffered 10 m around each release point to maximize our ability to select a release
refugium consisting of a large perennial shrub and an intact kangaroo rat (Dipodomys
spp.) burrow (Nafus et al. 2015a, Todd et al. 2016, Daly et al. 2019). Prior to release, we
enlarged the burrow to provide a refuge large enough to fully conceal the released
animal. Under this release protocol the minimum possible distance between release
burrows was 30 m. Final release points ranged between 150-350 m from the access road

30
and between 1.9-2.5 km to the closest powerline. We chose to separate release blocks to
allow us to track blocks individually, facilitating the tracking of this many animals, and to
minimize our activity in each block.
Post-release monitoring
We attached Advanced Telemetry Systems (ATS; Advanced Telemetry Systems, MN,
U.S.A) radio-transmitters to the fifth vertebral scute with gel epoxy (Devcon 5-minute
epoxy gel, ITW Engineered Polymers, County Clare, Ireland). We fitted Outdoor HS
with R1670 transmitters (3.1 g) and Combo HS with the slightly larger R1680 model (3.6
g). Before the epoxy hardened, we applied a thin coat of sand to camouflage the
transmitter package (Kazmaier et al. 2002). We checked each animal within 24 hours of
release to make sure their transmitters were working. After the initial 24-hour post-
release check, we tracked animals within each release block twice per week for the first
three weeks (25 September 2018-15 October 2018) before reducing our tracking
frequency to once per week until dormancy. During the dormancy period (November
2018-February 2019) we tracked juveniles every 10-14 days before returning to a weekly
tracking schedule in March for the remainder of the study (March 2019-July 2019). We
continued tracking during the winter dormancy period, though at a reduced rate, because
juvenile desert tortoises may be surface active when weather is favorable in winter
months and because we were interested in monitoring winter mortalities (Morafka 1994,
Wilson et al. 1999). At each tracking occasion, we recorded the tortoise’s location to the
nearest ±3 m using a handheld GPS (Garmin model GPSmap 76CSx, Olathe, Kansas).

31
Statistical methods
All statistical tests were performed in Program R (R Core Team 2017), and all inferences
were supported at a threshold type I error rate (alpha) of 0.05. Data are presented as
group means ± 1 standard error (SE). Graphical visualization of model residuals and
Shapiro-Wilk tests were used to test model assumptions of normally-distributed residuals.
When necessary, we transformed the data to meet model assumptions.
Morphometrics
We used linear mixed effects models (LME; ‘nlme’ package, ‘lme’ function) to
test for differences between treatment groups in MCL, mass, BC, and SHI. We compared
these response variables at hatching (‘initial’; except SHI), after 1 year of captive rearing
(‘Year 1’), and at the end of the two-year captive-rearing period (‘release’). We set the
metric of interest as the response variable and included mother identification number (ID)
as a random effect in all models to account for maternal effects (Steyermark and Spotila
2001, Nafus et al. 2015). We included MCL as a continuous covariate in models testing
the effect of treatment on SHI, allowing us to evaluate whether treatments varied at
similar sizes. We also tested for interaction between MCL and SHI in both (‘Year 1’ and
‘release’) SHI models. In any model in which the interaction was not significant, we
retained MCL as a covariate but removed the interactive term from the final model.
Movement
We used the Spherical Law of Cosines (Movable Type Ltd. 2015) to calculate
straight-line distances between successive tracking locations. For each individual, we
calculated mean distance moved per tracking event (excluding dormancy tracking events
from 01 November 2018 – 05 March 2019), final displacement at dormancy (31 October

32
2018) from the original release burrow, final distance from the dormancy burrow at the
end of the study (23 July 2019; for all animals surviving through dormancy), and final
displacement from release burrow until the animal died, went missing or until the end of
the study (23 July 2019; for all animals surviving >25 days post-release). We chose to
include all animals surviving at least 25 days post-release in the latter analyses because
animals in both treatment groups reached a clear asymptote for displacement from the
release site within 25 days. We did not include the dormancy period (01 November 2018
– 05 March 2019) in our movement analyses due to the fact that no movement was
recorded during this time (also seen in Rautenstrauch et al. 1998), though we did
document scat on burrow aprons throughout the winter which can be presumed to be
evidence of winter basking activity during sunny winter days.
We used a gamma hurdle model (Mullahy 1986) to compare distance moved per
tracking event (log link for nonzero distance component, logit link for binary outcome of
moved/not moved) between treatment groups (for all animals that emerged post-
dormancy; Daly et al. 2019). We fit this model under a Bayesian estimation framework
using the ‘brms’ package in R (R version 3.5, www.r-project.org, accessed 05 Sep 2019)
which implements Markov chain Monte Carlo (MCMC) sampling of the posterior
distributions of model parameters. To account for the non-independence of repeated
measures on the same individuals, we included individual identification number (ID) as a
random effect in our model. We used noninformative improper flat priors (uniform
distribution on the real line) to sample treatment group effects. We ran 4 chains of the
MCMC sampler, each with 10,000 iterations and a thinning rate of 1, while discarding
the first 1,000 iterations as burn-in (Daly et al. 2019). We considered treatment group

33
differences to be significant if 95% Bayesian credible intervals (BCIs) of model
predictions did not overlap.
To compare pre-dormancy movement distance, post-dormancy movement
distance, and final displacement between treatment groups, we used a generalized linear
model (Gamma family, log link) with the movement metric as the response variable and
treatment group as a categorical predictor variable. For both pre-dormancy movement
distance and final displacement, we included only those animals surviving >25 days post-
release (21 October 2019; n = 46). To be included in the post-dormancy movement
comparison, animals had to survive the dormancy period and emerge in Spring 2019 (n =
45). We added 0.1 m to all pre-dormancy and post-dormancy movement data to eliminate
zeros contained in the data. We report data as raw means ± 1 SE in text and tables and
figures for ease of interpretation, unless otherwise noted.
Survivorship
Upon finding a deceased tortoise, we thoroughly examined the surrounding area
for clues of cause of death. We photographed each carcass, including any signs of
predation, and removed carcass remnants for further analysis. We attributed deaths to one
of five sources of mortality: mammal depredated, bird depredated, unidentified predator,
exposure, or unknown fate. Signs of predation included digging, tracks, scat, and
condition of the carcass or transmitter. We classified carcasses that were found near clear
digging activity or that had obvious tooth marks as “mammal predated.” Remains near
perch sites or with clear signs of being punctured by a beak were classified as “bird
depredated.” Remains found without tooth marks or lacking evidence of avian predation
were considered depredated by an “unidentified predator.” Animals found intact on the

34
surface or dead in their burrow were presumed to have died due to “exposure.” An
“unknown fate” classification was given to animals lost due to radio failure (faulty
batteries or the antenna became damaged), and these animals were right censored (no
assumption of fate) in our survival analysis. We estimated survival rates in the field from
release until dormancy (25 September 2018 – 31 October 2018) and from release until the
end of the tracking period (September 2018 – July 2019) using the Kaplan-Meier
estimator in the R package ‘survival’ for each treatment group.
RESULTS
Sixteen females collectively laid 92 eggs, resulting in 68 successfully emerged hatchlings
(73.9% emergence) after an average incubation period of 87 days (range 74-106 days).
We randomly selected 48 hatchlings (which represented 15 of the 16 females that
successfully nested) for inclusion in this study.
Morphometrics
Neither mean MCL (45.2 ± 0.3 mm) nor mean mass (20.8 ± 0.3 g) at hatching
(‘initial’) differed between treatment groups (MCL: F1,32 = 0.442, p = 0.51; mass: F1,32 =
0.235, p = 0.63; Table 2.1). After one year of captive rearing (‘Year 1’; September 2017),
when the Combo HS treatment was transitioned to outdoor rearing pens, Combo HS
tortoises (MCL: 102.5 ± 1.5 mm; mass: 205.8 ± 7.2 g) were significantly longer and
heavier than Outdoor HS tortoises (MCL: 66.5 ± 1.5 mm, F1,32 = 291.9, p < 0.001; mass:
62.6 ± 7.4 g, F1,32 = 194.0, p < 0.001; Table 2.1). Combo HS tortoises (MCL: 117.2 ± 1.7
mm, mass: 300.6 ± 10.7 g) retained their size advantage through release (September
2018; Outdoor HS MCL: 83.7 ± 1.7 mm, F1,32 = 193.9, p < 0.001; Outdoor HS mass:
115.9 ± 10.7 g, F1,32 = 148.6, p < 0.001; Table 2.1; Figures 2.1, 2.2), with Combo HS

35
tortoises weighing on average nearly three times that of Outdoor HS tortoises and
achieving the shell length of an 8-year-old wild tortoise (Medica et al. 2012).
Body condition
Using the formula from Nagy et al. (2002), average initial BC was 0.53 ± 0.004
g/cm3 (n = 48) and did not differ between treatment groups (F1,32 = 2.965, p = 0.10; Table
2.1). Body condition values at both Year 1 (September 2017; Combo HS: 0.59 ± 0.007;
Outdoor HS: 0.57 ± 0.007; F1,32 = 2.267, p = 0.14; Table 2.1) and at release (September
2018; Combo HS: 0.57 ± 0.007; Outdoor HS: 0.57 ± 0.007; F1,32 = 0.008, p = 0.93; Table
2.1; Figure 2.3) remained similar between treatment groups. We also found no difference
when comparing mean BC at release for animals found dead prior to spring emergence (n
= 3; 0.58 ± 0.009) to those known to survive dormancy (n = 45; 0.57 ± 0.007; F1,32 =
0.523, p = 0.40).
Shell hardness
Year 1 SHI did not vary between Combo HS (x̄ = 95.9 ± 0.6%) and Outdoor HS
tortoises (x̄ = 94.8 ± 1.31%; t = -0.81, df = 30, p = 0.43; Table 2.1; Figure 2.4). Two
Outdoor HS animals were unavailable for measurements in Year 1 due to early
hibernation. After two years of captive rearing, Combo HS tortoises (x̄ = 97.9 ± 0.4%)
had significantly harder shells than Outdoor HS tortoises (x̄ = 95.7 ± 0.6%; t = -3.86, df =
31, p = 0.0005; Table 2.1, Figure 2.4) at release. Size (MCL) was not a significant
predictor of SHI in the models (p > 0.77 for both ‘Year 1’ and ‘release’), nor was the
interaction between treatment and MCL (p > 0.34 for both ‘Year 1’ and ‘release’). After
two years, Combo HS animals had the shell hardness values of 6-7-year-old outdoor-
reared tortoises at our site (Figure 2.5).

36
Movement
Combo HS and Outdoor HS tortoises did not differ in probability of moving or in
distance moved per movement event during the 10-month study period based on
overlapping Bayesian Credible Intervals [95% BCI]. Probability of moving between
tracking events was 0.46 (95% BCI = 0.42 - 0.50) for the Combo HS group, and 0.42
(95% BCI = 0.38 - 0.47; Table 2.2) for the Outdoor HS group. During movement events,
Combo HS tortoises moved an average of 36.2 m (95% BCI = 29.7 - 44.7 m) per event,
compared to 43.4 m (95% BCI = 34.1 - 54.6 m; Table 2.2) by Outdoor HS tortoises.
Between release and dormancy (‘pre-dormancy’; 25 September 2018 – 31
October 2018), Outdoor HS tortoises dispersed farther from their release burrows (n=23;
x̄ = 255 ± 89 m; t = 1.63, df = 45, p = 0.11), moving nearly twice the distance moved by
Combo HS tortoises (n=23; x̄ = 114 ± 40 m; Table 2.2; Figure 2.6). However, between
emerging from dormancy and the end of the study (‘post-dormancy’; 05 March 2019 – 25
July 2019), Outdoor HS tortoises reduced their movement away from their release
location (n=22; x̄ = 92 + 34 m) and exhibited similar displacement distances between
their dormancy location and final location when compared to Combo HS tortoises (n=23;
x̄ = 54 + 19 m; t = 1.05, df = 44, p = 0.30; Table 2.2; Figure 2.6). However, when the
two largest post-dormancy movers from the Outdoor HS group (896 m and 534 m) were
removed from the data, mean post-dormancy movement distance was reduced to 30 m for
the Outdoor HS treatment group. Total displacement from release burrow until the end of
the study (25 September 2018 – 23 July 2019) was heavily influenced by pre-dormancy
dispersal away from the release burrow, leading to a significant difference in final
displacement between treatment groups (t = 2.05, df = 45, p = 0.047; Table 2.2). Outdoor

37
HS tortoises (n=22; x̄ = 326 ± 93 m) dispersed over two times the distance of Combo HS
tortoises (n=23; x̄ = 143 ± 41 m) during the 10-month post-release monitoring period.
Notably, within treatment groups, there was large variation in final displacement, ranging
from 9–1178 m (Combo HS) and 11–1041 m (Outdoor HS) based on all animals that
survived dormancy.
Post-release survivorship
Survival from release to dormancy (5 weeks post-release; 25 September 2018 - 31
October 2019) was equally high for both treatment groups at 96% (n = 23/24 Combo HS
and n = 23/24 Outdoor HS; Figure 2.7). During dormancy, one Outdoor HS tortoise was
found dead after having been removed from its winter burrow by a mammalian predator,
as evidenced by signs of digging at the burrow. At the end of the 10-month post-release
monitoring period (25 September 2018 - 25 July 2019), 38 (79.2%) of the 48 released
head-starts were confirmed alive. All ten mortalities were attributed to predation; no
animals died due to exposure. Six (12.5% of released tortoises) head-starts were mammal
predated, three (6.2% of released tortoises; all from Outdoor HS group) were killed by
avian predators, and one (2.1% of released tortoises) was killed by an unidentified
predator. The head-start killed by an unidentified predator was found dead and buried in
its burrow. The burrow entrance showed signs of mammalian digging, while both the
burrow and tortoise carcass were covered in harvester ants (Veromessor spp.). Overall
estimated survival for the 48 head-starts during the 10-month post-release monitoring
period (September 2018 - July 2019) was 0.79 (95% CI, 0.69-0.92; Figure 2.7). Ten-
month survival was 0.88 (95% CI, 0.75-1.0) for the Combo HS treatment and 0.71 (95%

38
CI, 0.55-0.92; Figure 2.7) for the Outdoor HS treatment, with overlapping 95%
confidence intervals indicating no difference between treatment groups.
DISCUSSION
After two years of captive rearing, Combo HS tortoises were both larger and had
harder shells than Outdoor HS tortoises from the same cohort. Combo HS tortoises were
the size of an 8-year-old wild juvenile with the shell hardness of an 11-year-old wild
tortoise (Nagy et al. 2011, Medica et al. 2012), and weighed nearly three times more than
same-aged Outdoor HS tortoises. However, Outdoor HS tortoises were also large for their
age when compared to wild counterparts, attaining the size of 4-5-year-old wild tortoises
with just two years of captive rearing (Medica et al. 2012). Outdoor HS animals in this
study grew 19.3 mm MCL/yr, an enhanced rate that can be attributed to the supplemental
food and water they were provided weekly during the activity season (March-October).
Tuberville et al. (2019) reported growth rates of 12.7 mm MCL/yr for juvenile tortoises
reared outdoors without supplemental food at the IDTRF. Collectively, these results lend
support for providing supplemental food to desert tortoises during outdoor captive
rearing, particularly as vegetation in the pens becomes depleted over time.
The growth rate of Combo HS tortoises during their year indoors (4.7 mm/month)
was similar to that found by Daly et al. (2018) for tortoises reared indoors under nearly
the same protocols (4.3 mm/month). Collectively, the findings of Daly et al. (2018) and
those of this study highlight the significant growth advantage of indoor captive rearing
for head-starting efforts for desert tortoises. Combo HS tortoises surpassed the Nagy et
al. (2015) recommended release size of 100 mm MCL after just one year of indoor
rearing (x̄ = 102.5 mm MCL), while Outdoor HS tortoises reached a mean size equivalent

39
to the Hazard et al. (2015) recommendation of 84 mm MCL after two years outdoors (x̄ =
83.7 mm MCL; Figure 2.1).
Throughout the two-year captive rearing period, both treatment groups exhibited
similar body conditions. In contrast, Daly et al. (2018) found that tortoises reared indoors
for seven months had lower body condition than same-aged outdoor-reared individuals
(although this relationship diminished when comparing larger individuals). Daly et al.
(2018), the only other study to incorporate an indoor captive-rearing component for the
desert tortoise, posited that the decreased body condition exhibited by indoor head-starts
may have been a result of decreased bone density in relation to body size. By
incorporating UV lighting and a calcium supplement for the duration of the indoor
captive-rearing period, our husbandry protocols may have enhanced calcium assimilation
during the indoor rearing phase of the Combo HS treatment in comparison to tortoises
reared indoors by Daly et al. (2018), a possibility corroborated by our shell hardness
measurements (see below).
Body condition of head-started tortoises ranged from 0.47-0.65 g/cm3 across
treatments and during the captive rearing period. Nagy et al. (2002) determined prime
body condition values for wild desert tortoises to be between 0.6-0.7 g/cm3, which 21-
27% of individuals in our study attained. However, prime body condition values
represent peak nutritional status and thus do not correspond with the post-summer
(September) body condition values obtained in this study. In fact, it has been shown that
desert tortoises often emerge from summer aestivation weighing less than they did the
previous spring (Nagy et al. 1986). Nonetheless, after initial measurements, all
individuals in our study had body condition values above the Nagy et al. (2002) threshold

40
value for a dehydrated desert tortoise (0.45 g/cm3). Likewise, using the body condition
formula from Loehr et al. (2004), the tortoises in our study were well within the range of
body condition values for both adult speckled tortoises (Chersobius signatus; Loehr et al.
2004) and western pond turtles (Actinemys marmorata; Ashton et al. 2015).
The slow growth and delayed maturity exhibited by chelonians stem from their
investment in maintenance and growth of a protective shell (Klemens 2000). The level of
protection, and presumably the ability to withstand a predatory attack, can best be
measured by quantifying shell hardness, which increases with both size and age in
healthy turtles (Hill 1999, Nagy et al. 2011). Though significantly larger, Combo HS
tortoises at Year 1 had shell hardness values similar to Outdoor HS tortoises at Year 1. It
seems that one year of indoor conditions (i.e. UV lighting, calcium supplementation, and
accelerated growth) or one year of outdoor conditions (i.e. supplemental food and water
as well as natural sunlight) at our site both result in nearly equal shell hardness values.
However, Combo HS tortoises had significantly harder shells at release than 2-year old
Outdoor HS tortoises. Two-year old Combo HS tortoises had shell hardness values equal
to those calculated for an 11-year-old wild desert tortoise (Nagy et al. 2011). Through
combining the accelerated growth of indoor captive rearing and the natural shell
ossification of outdoor captive rearing, we were able to significantly increase the shell
hardness of two-year-old captive reared desert tortoises relative to both wild counterparts
as well as tortoises captive reared solely outdoors.
Extreme post-release dispersal can remove individuals from optimal habitat, result
in higher incidence of exposure and predation, and ultimately impede translocation
efforts (Stamps and Swaisgood 2007, Germano and Bishop 2009, Roe et al. 2010). In our

41
study, with the exception of 2 individual outliers (> treatment mean + 2 SD), released
head-starts did not move far from their release burrows. Both treatment group means
were within the range (~100-400 m) of mean initial (< 2mo) dispersal distances reported
for head-started or translocated juvenile desert tortoises (Nafus et al. 2017, Germano et
al. 2017). However, in our study, pre-dormancy dispersal distance of Outdoor HS
tortoises was twice that of Combo HS tortoises. This increased movement by Outdoor HS
tortoises may be related to their smaller release size, allowing them to heat up faster each
morning (Berry and Turner 1986, Geffen and Mendellssohn 1988, Harris et al. 2015) and
possibly take advantage of a longer period of favorable surface temperatures each day to
disperse farther than the larger Combo HS tortoises.
Roughly 80% of the total dispersal by both treatment groups occurred in the first
three weeks post-release, supporting previous studies of post-release movement of head-
started desert tortoises (Hazard et al. 2015, Nagy et al. 2015, Germano et al. 2017, Daly
2017) and gopher tortoises (Quinn et al. 2018). Germano et al. (2017) also reported a
secondary dispersal event after the first dormancy period, during which 13% of their
juveniles engaged in movements away from the release site. Similarly, four tortoises
(9%) in our study dispersed >100 m (range 104-896 m) away from their dormancy
burrows during the spring activity period. Post-dormancy dispersal was slightly less (~20
m) for Outdoor HS tortoises, when two extreme movers were removed from the data,
possibly due to the larger size of Combo HS tortoises necessitating a slightly larger home
range to acquire all necessary resources (Lindstedt et al. 1986, though see Harless et al.
2009). However, total displacement during the 10-month study period was predominantly
driven by pre-dormancy dispersal from the release burrow. Combination head-starting

42
significantly decreased this initial dispersal, resulting in Combo HS tortoises settling
nearly 200 m closer to their release burrows than Outdoor HS tortoises after ten months.
Future studies should investigate the replicability of this result, alongside the potential
physiological explanations for its manifestation. Reducing initial dispersal allows refined
management within conservation areas as well as continued monitoring of population
recovery.
When compared to previous releases of juvenile desert tortoises (Morafka et al.
1997, Hazard and Morafka 2002, Nagy et al. 2015, Germano et al. 2017, Nafus et al.
2017), including those from our release area (Tuberville et al. 2019, Daly et al. 2019),
tortoises in our study exhibited particularly high survival rates. This finding was
especially pronounced when considering the survival of our Combo HS group, whose
88% 10-month survival marks the highest survivorship ever reported for any size-class of
juvenile desert tortoises. Mortality risks for juvenile tortoises released at our sites in the
Ivanpah valley have changed over time. Original releases (Tuberville et al. 2019) focused
on locating appropriate microhabitats for release as well as the effects of short-term
outdoor captive rearing. Short-term (outdoor) captive rearing resulted in the release of
smaller head-starts (40.8–61.5 mm MCL), a possible explanation for the significant
losses as a result of exposure in that study. Smaller juveniles experience increased rates
of water loss and are thus at a greater risk for desiccation during post-release activities
(Wilson et al. 2000). Over 15% of head-starts released by Tuberville et al. (2019) died
due to suspected exposure, greatly outnumbering the losses attributed to avian predators
(n = 1, 2.2%). Ravens became a significant source of mortality in the subsequent release
at our site by Daly et al. (2019), with nearly 27% of all released tortoises lost to these

43
opportunistic predators in the first year post-release (Sherman 1993, Boarman 2003). In
turn, Daly et al. (2019) recommended releasing tortoises over 1.6 km away from
powerlines, a known nesting structure for ravens. Incorporating this recommendation as
part of our release protocols coupled with the larger size of our combo HS tortoises may
account for the higher survival rates documented in our study. Building on previous
efforts has increased the success and efficiency of our head-starting program.
Management implications
In just two years of captive rearing, combination head-starting produced tortoises
larger than all previously published recommendations for minimum size at release for the
desert tortoise (Nagy et al. 2015, Hazard et al. 2015). Combo HS tortoises, equivalent in
size to 8-year-old wild tortoises (Medica et al. 2012), had nearly prime body condition
values (Nagy et al. 2002) and a shell hardness index similar to an 11-year-old wild
tortoise (Nagy et al. 2011). In addition, Combo HS tortoises dispersed shorter distances
than Outdoor HS tortoises.
Both post-release site fidelity and survival were high for both treatment groups.
By releasing tortoises in late September, we were able to minimize the amount of time
available for dispersal prior to dormancy, increasing the probability that animals would
settle near the release area. An early fall release also allowed tortoises sufficient time to
establish burrows and become familiar with areas for resource acquisition before the
especially vulnerable spring activity period documented in this and in other desert
tortoise post-release studies (Nagy et al. 2015a, Daly et al. 2019). The combination of a
fall release when raven activity is lower and selecting release sites >1.6 km from
potential perching sites (Daly et al. 2019), likely contributed to the high post-release

44
survival exhibited by both treatment groups. In agreement with previous studies (Hazard
et al. 2015, Daly et al. 2019), the initial results of our study support releasing head-starts
during the fall activity period. To determine the effect of annual climatic variation on our
findings, replication as well as continued monitoring should be prioritized.

45
LITERATURE CITED
Acierno, M. J., M.A. Mitchell, M.K. Roundtree, and T.T. Zachariah. 2006. Effects of
ultraviolet radiation on 25-hydroxyvitamin D3 synthesis in red-eared slider turtles
(Trachemys scripta elegans). American Journal of Veterinary Research 67:2046-
2049.
Ashton, D.T., J.B. Bettaso, and H.H. Welsh Jr. 2015. Changes across a decade in size,
growth, and body condition of western pond turtle (Actinemys marmorata)
populations on free-flowing and regulated forks of the Trinity River in northwest
California. Copeia 103:621-633.
Barrows, C. W. 2011. Sensitivity to climate change for two reptiles at the Mojave–
Sonoran Desert interface. Journal of Arid Environments 75:629-635.
Baxter, R.J. 1988. Spatial distribution of desert tortoises (Gopherus agassizii) at
Twentynine Palms, California: implications for relocations. Pages 180–189 in
Szaro R.C., Severson K.E., Patton D.R., technical coordinators. Management of
amphibians, reptiles, and small mammals in North America: proceedings of the
symposium. Fort Collins, Colorado: U.S. Department of Agriculture, Forest
Service, Rocky Mountain Forest and Range Experiment Station. General
Technical Report RM-166
Beatley, J.C. 1974. Phenological events and their environmental triggers in Mojave
Desert ecosystems. Ecology 55:856-863.
Berry, K. 1986. Desert tortoise (Gopherus agassizii) research in California, 1976-1985.
Herpetologica 42:62-67.
Berry, K. H., and F B. Turner. 1986. Spring activities and habits of juvenile desert

46
tortoises, Gopherus agassizii, in California. Copeia 1986:1010-1012.
Bertolero, A., J.Ll. Pretus, and D. Oro. 2018. The importance of including survival
release costs when assessing viability in reptile translocations. Biological
Conservation 217:311-320.
Boarman, W. I. 2003. Managing a subsidized predator population: reducing common
raven predation on desert tortoises. Environmental Management 32:205-217.
Cagle, F.R. 1939. A system of marking turtles for future identification. Copeia 1939:170-
173.
Congdon, J.D., A.E. Dunham, and R.C. Van Loben Sels. 1993. Delayed sexual maturity
and demographics of Blanding’s turtles (Emydoidea blandingii): implications for
conservation and management of long-lived organisms. Conservation Biology
7:826-833.
Congdon, J.D., A.E. Dunham, and R.C. van Loben Sels. 1994. Demographics of common
snapping turtles (Chelydra serpentina): implications for conservation and
management of long-lived organisms. American Zoologist 34:397-408.
Crane, A.L., and A. Mathis. 2010. Predator-recognition training: a conservation strategy
to increase post-release survival of hellbenders in head-starting programs. Zoo
Biology 30:611-622.
Crawford, B.A., J.C. Maerz, N.P. Nibbelink, K.A. Buhlmann, and T.M. Norton. 2014.
Estimating the consequences of multiple threats and management strategies for
semi-aquatic turtles. Journal of Applied Ecology 51:359-366.

47
Cunninghame, F., R. Switzer, B. Parks, G. Young, A. Carrión, P. Medranda, and C.
Sevilla. 2015. Conserving the critically endangered mangrove finch: head-starting
to increase population size. Galapagos Report 2013-2014:151-157.
Daly, J.A. 2017. Indoor rearing as a component of head-starting the Mojave Desert
Tortoise (Gopherus agassizii). M.S. Thesis. University of Georgia, Athens.
Daly, J.A., K.A. Buhlmann, B.D. Todd, C.T. Moore, J.M. Peaden, and T.D. Tuberville.
2018. Comparing growth and body condition of indoor-reared, outdoor-reared,
and direct-released juvenile Mojave Desert Tortoises. Herpetological
Conservation and Biology 13:622-633.
Daly, J.A., K.A. Buhlmann, B.D. Todd, C.T. Moore, J.M. Peaden, and T.D. Tuberville.
2019. Survival and movements of head-started Mojave desert tortoises. Journal of
Wildlife Management 83:1700-1710. DOI:
Diez, M., B. Vanstrazeele, D. Justet, J. Detilleux, P. Dortu, L. Grolet, and C. Remy.
2009. Effects of different levels of energy allowances and of hibernation on
growth in hatchling Testudo hermanni boettgeri (Mojsisovic, 1889). Zoo Animal
Nutrition 4:118-130.
Doak, D., P. Kareiva, and B. Klepetka. 1994. Modeling population viability for the desert
tortoise in the western Mojave Desert. Ecological Applications 4:446-460.
Ellis, D. H., G.F. Gee, S.G. Hereford, G.H. Olsen, T.D. Chisolm, J.M. Nicolich, and J.S.
Hatfield. 2000. Post-release survival of hand-reared and parent-reared Mississippi
sandhill cranes. The Condor 102:104-112.
Ernst, C. H., and J.E. Lovich. 2009. Turtles of the United States and Canada. Johns
Hopkins University Press, Baltimore, Maryland.

48
Fischer, J., and D.B. Lindenmayer. 2000. An assessment of the published results of
animal relocations. Biological Conservation 96:1-11.
Foreman, L.D., J.M. Brode, R. Haussler, and K. Kramer. 1986. The responsibilities of
federal and state agencies for protection of the desert tortoise in California.
Herpetologica 1:59.
Geffen, E., and H. Mendolssohn. 1989. Activity patterns and thermoregulatory behavior
of the Egyptian tortoise Testudo kleinmanni in Israel. Journal of Herpetology
23:404-409.
Grueber, C.E., E.E. Reid-Wainscoat, S. Fox, K. Belov, D.M. Shier, C.J. Hogg, and D.
Pemberton. 2017. Increasing generations in captivity is associated with increased
vulnerability of Tasmanian devils to vehicle strike following release to the wild.
Scientific Reports 7:2161.
Haag, W. R., and J.D. Williams. 2014. Biodiversity on the brink: an assessment of
conservation strategies for North American freshwater
mussels. Hydrobiologia 735:45-60.
Harless, M.L., A.D. Walde, D.K. Delaney, L.L. Pater, and W.K. Hayes. 2009. Home
range, spatial overlap, and burrow use of the desert tortoise in the West Mojave
Desert. Copeia 378-389.
Harris, B.B., T.M. Norton, N.P. Nibbelink, and T.D. Tuberville. 2015. Overwintering
ecology of juvenile gopher tortoises (Gopherus polyphemus). Herpetological
Conservation and Biology 10:645-653.
Hazard, L.C., D.J. Morafka, and L.S. Hillard. 2015. Post-release dispersal and predation
of head-started juvenile desert tortoises (Gopherus agassizii): effect of release site

49
distance on homing behavior. Herpetological Conservation and Biology 10:504-
515.
Hill, R.A. 1999. Size-dependent tortoise predation by baboons at de Hoop Nature
Reserve, South Africa. South African Journal of Science 95:123-124.
Jachowski, D. S., and J.M. Lockhart. 2009. Reintroducing the black-footed ferret Mustela
nigripes to the Great Plains of North America. Small Carnivore
Conservation 41:58-64.
Jarchow, J.L., H.E. Lawler, T.R. Van Devender, and C.S. Ivanyi. 2002. Care and diet of
captive Sonoran desert tortoises. Pages 289–311 in T. R. Van Devender, editor.
The Sonoran Desert Tortoise. The University of Arizona Press, Tucson, AZ.
Jennings, W.B., and K.H. Berry. 2015. Desert tortoises (Gopherus agassizii) are selective
herbivores that track the flowering phenology of their preferred food plants. PLoS
ONE 10:e0116716.
Kazmaier, R.T., E.C. Hellgren, and D.C. Ruthven, III. 2002. Home range and dispersal of
Texas tortoises, Gopherus berlandieri, in a managed thornscrub ecosystem.
Chelonian Conservation Biology 4:488-496.
Kelly, E.C., B.L. Cypher, and D.J. Germano. 2019. Temporal variation in foraging
patterns of Desert Kit Foxes (Vulpes macrotis arsipus) in the Mojave Desert,
California, USA. Journal of Arid Environments 167:1-7.
Klemens, M.W. 2000. Turtle Conservation. Edited by M.W. Klemens. Smithsonian
Institution Press. Washington D.C., USA.
Lindenmayer, D. B., J.F. Franklin, and J. Fischer. 2006. General management principles
and a checklist of strategies to guide forest biodiversity conservation. Biological

50
conservation 131:433-445.
Lindstedt, S.L., B.J. Miller, and S.W. Buskirk. 1986. Home range, time, and body size in
mammals. Ecology 67:413-418.
Loehr, V., B. Henen, and M. Hofmeyr. 2004. Reproduction of the smallest tortoise, the
Namaqualand speckled padloper, Homopus signatus signatus. Herpetologica
60:759-761.
Loehr, V.J.T., M.D. Hofmeyr, and B.T. Henen. 2007. Annual variation in the body
condition of a small, arid-zone tortoise, Homopus signatus signatus. Journal of
Arid Environments 71:337-349.
Loh, J., B. Collen, L. McRae, T.T. Carranza, F.A. Pamplin, R. Amin, and J.E.M. Baillie.
2008. Living Planet Index. In: C. Hails (ed.) Living Planet Report 2008, pp. 6-20.
WWF International, Gland, Switzerland.
Luckenbach, R. A. 1982. Ecology and management of the desert tortoise (Gopherus
agassizii) in California. Pages 1–38 in North American tortoises: conservation
and ecology (R. B. Bury, editor). United States Fish and Wildlife Service,
Washington, D.C., Wildlife Research Report 12:1-126.
Masin, S., G.F. Ficetola, and L. Bottoni. 2015. Head starting European pond turtle (Emys
orbicularis) for reintroduction: patterns of growth rates. Herpetological
Conservation and Biology 10:516-524.
McCoy, E. D., R.D. Moore, H.R. Mushinsky, and S.C. Popa. 2011. Effects of rainfall and
the potential influence of climate change on two congeneric tortoise
species. Chelonian Conservation and Biology 10:34-41.

51
Medica, P. A., K.E. Nussear, T.C. Esque, and M.B. Saethre. 2012. Long-term growth of
desert tortoises (Gopherus agassizii) in a southern Nevada population. Journal of
Herpetology 46:213-220.
Milinkovitch, M. C., R. Kanitz, R. Tiedemann, W. Tapia, F. Llerena, A. Caccone, and
J.R. Powell. 2013. Recovery of a nearly extinct Galápagos tortoise despite
minimal genetic variation. Evolutionary Applications 6:377-383.
Morafka, D.J. 1994. Neonates: missing links in the life histories of North American
tortoises. In: Bury, R.B., Germano D.J. (Eds.), Biology of North American
Tortoises. National Biological Survey, Fish and Wildlife Research 13,
Washington, DC, pp. 161–173.
Morafka, D.J., K.H. Berry, and E.K. Spangenberg. 1997. Predator-proof field enclosures
for enhancing hatchling success and survivorship of juvenile tortoises: a critical
evaluation. Conservation, Restoration, and Management of Tortoises and Turtles,
An International Conference. 147-165.
Movable Type Ltd. 2015. Calculate distance, bearing and more between
latitude/longitude points. Movable Type Scripts.
http://www.movabletype.co.uk/scripts/latlong.html. Accessed 01 Dec 2015.
Murphy, R.W., K.H. Berry, T. Edwards, A.E. Leviton, A. Lathrop, and J.D. Riedle. 2011.
The dazed and confused identity of Agassiz’s land tortoise, Gopherus agassizii
(Testudines, Testudinidae) with the description of a new species, and its
consequences for conservation. ZooKeys 113:39–71.
Myers, N., R.A. Mittermeier, C.G. Mittermeier, G.A. Da Fonseca, and J. Kent. 2000.
Biodiversity hotspots for conservation priorities. Nature 403:853.

52
Nafus, M.G., J.M. Germano, J.A. Perry, B.D. Todd, A. Walsh, and R.R. Swaisgood.
2015a. Hiding in plain sight: a study on camouflage and habitat selection in a
slow-moving desert herbivore. Behavioral Ecology 26:1389-1394.
Nafus, M. G., B.D. Todd, K.A. Buhlmann, and T.D. Tuberville. 2015b. Consequences of
maternal effects on offspring size, growth and survival in the desert
tortoise. Journal of Zoology 297:108-114.
Nafus, M. G., T.C. Esque, R.C. Averill‐Murray, K.E. Nussear, and R.R. Swaisgood.
2017. Habitat drives dispersal and survival of translocated juvenile desert
tortoises. Journal of Applied Ecology 54:430-438.
Nagy, K., B. Henen, B. Devesh, and I. Wallis. 2002. A condition index for the desert
tortoise (Gopherus agassizii). Chelonian Conservation and Biology 4:425–429.
Nagy, K., M. Tuma, and L. Hillard. 2011. Shell hardness measurements in juvenile desert
tortoises Gopherus agassizii. Herpetological Review 42:191–195.
Nagy, K.A., S. Hilliard, S. Dickson, and D.J. Morafka. 2015a. Effects of artificial rain on
survivorship, body condition, and growth of head-started desert tortoises
(Gopherus agassizii) released to the open desert. Herpetological Conservation and
Biology 10:535–549.
Nagy, K.A., L.S. Hillard, M.W. Tuma, and D.J. Morafka. 2015b. Head-started desert
tortoises (Gopherus agassizii): movements, survivorship and mortality causes
following their release. Herpetological Conservation and Biology 10:203-215.
Olsson, M., and R. Shine. 2002. Growth to death in lizards. Evolution 56:1867-1870.
Peaden, J.M., T.D. Tuberville, K.A. Buhlmann, M.G. Nafus, and B.D. Todd. 2015.
Delimiting road-effect zones for threatened species: implications for mitigation

53
fencing. Wildlife Research 42:650–659.
Pérez-Buitrago, N., M.A. Garcia, A. Sabat, J. Delgado, A. Alvarez, O. McMillan, and
S.M. Funk. 2008. Do head-start programs work? Survival and body condition in
head-started Mona Island iguanas Cyclura cornuta stejnegeri. Endangered
Species Research 6:55-65.
Platenberg, R.J. and R.A. Griffiths. 1999. Translocation of slow-worms (Anguis fragilis)
as a mitigation strategy: a case study from south-east England. Biological
Conservation 90:125-132.
Pritchard, P. C. H. 1981. Criteria for scientific evaluation of head-starting. Marine Turtle
Newsletter 19:3-4.
Randel III, C.J. 2015. Home Range, Reproduction, and Survival of the Desert Kit Fox,
Southeastern, California. PhD Dissertation, Texas A&M University, College
Station, Texas, USA. 59 p.
Rautenstrauch, K.R., A.L. Rager, and D.L. Rakestraw. 1998. Winter behavior of desert
tortoises in southcentral Nevada. The Journal of wildlife management pp:98-104.
R Core Team. 2017. R: a language and environment for statistical computing. Foundation
for Statistical Computing, Vienna, Austria.
Seigel, R.A., and C.K. Dodd, Jr. 2000. Manipulation of turtle populations for
conservation: halfway technologies or viable options? Pp. 218-238 In Turtle
Conservation. Klemens, M.W. (Ed.). Smithsonian Institution Press, Washington,
DC, USA.
Sherman, M. W. 1993. Activity patterns and foraging ecology of nesting ravens in the
Mojave Desert, California. Thesis, Colorado State University, Fort Collins, USA.

54
Spencer, R.J., J.U. Van Dyke, and M.B. Thompson. 2017. Critically evaluating best
management practices for preventing freshwater turtle extinctions. Conservation
Biology 31:1340-1349.
Steyermark, A.C. and J.R. Spotila. 2001. Effects of maternal identity and incubation
temperature on snapping turtle (Chelydra serpentina) growth. Functional Ecology
15:624-632.
Tetzlaff, S.J., J.H Sperry, and B.A. DeGregorio. 2019. Effects of antipredator training,
environmental enrichment, and soft release on wildlife translocations: A review
and meta-analysis. Biological Conservation 236:324-331.
Todd, B.D., B. Halstead, L.P. Chiquoine, J.M. Peaden, K.A. Buhlmann, T.D. Tuberville,
and M.G. Nafus. 2016. Habitat selection by juvenile Mojave desert tortoises.
Journal of Wildlife Management 80:720–728.
Tomillo, P.S., V.S. Saba, R. Piedra, F.V. Paladino, and J.R. Spotila. 2008. Effects of
illegal harvest of eggs on the population decline of leatherback turtles in Las
Baulas Marine National Park, Costa Rica. Conservation Biology 22:1216-1224.
Tuberville, T.D., M.T. Norton, K.A. Buhlmann, and V. Greco. 2015. Head-starting as a
management component for gopher tortoises (Gopherus polyphemus).
Herpetological Conservation and Biology 10:455–471.
Tuberville, T.D., K.A. Buhlmann, R. Sollmann, M.G. Nafus, J.M. Peaden, J.A. Daly, and
B.D. Todd. 2019. Effects of short-term head-starting on growth and survival in
the Mojave Desert Tortoise (Gopherus agassizii). Herpetological Conservation
and Biology 14:171–184.
USFWS 2011. Revised recovery plan for the Mojave population of the desert tortoise

55
(Gopherus agassizii). U.S. Fish and Wildlife Service, Pacific Southwest Region,
Sacramento, California, USA.
van de Ven, W.A.C., J.P. Guerrero, D.G. Rodriguez, S.P. Telan, M.G. Balbas, B.A.
Tarun, M. van Weerd, J. van der Ploeg, Z. Wijtten, F.E. Lindeyer, and H.H. de
Iongh. 2009. Effectiveness of head-starting to bolster Philippine crocodile
Crocodylus mindorensis populations in San Mariano municipality, Luzon,
Philippines. Conservation Evidence 6:111-116.
Vié, J.C., C. Hilton-Taylor, and S.N. Stuart. 2009. Wildlife in a Changing World–An
Analysis of the 2008 IUCN Red List of Threatened Species. International Union
for Conservation of Nature, Gland, Switzerland.
Wiesner, C.S., and C. Iben. 2003. Influence of environmental humidity and dietary
protein on pyramidal growth of carapaces in African spurred tortoises
(Geochelone sulcata). Journal of Animal Physiology and Animal Nutrition 87:66–
74.
Wilson, D.S., K.A. Nagy, C.R. Tracy, D.J. Morafka, and R.A. Yates. 2001. Water
balance in neonate and juvenile desert tortoises, Gopherus
agassizii. Herpetological Monographs (2001):158-170.
Woodbury, A., and R. Hardy. 1948. Studies of the desert tortoise, Gopherus agassizii.
Ecological Monographs 18:145–200.

56
TABLES
Table 2.1. Summary statistics of growth metrics for juvenile Mojave desert tortoises
(Gopherus agassizii) reared under two different husbandry treatments (Outdoor head-
started, Combination head-started) including measurements taken shortly after hatching
in Fall 2016 (‘initial’), measurements taken after one year of captive rearing in Fall 2017
(‘Year 1’; when Combination head-starts were transitioned outdoors), and measurements
taken before release in Fall 2018 (‘Release’). Intervals are reported at 95% confidence. P-
values (significant values in bold) correspond to linear mixed effects models (with
mother ID as random effect). MCL= midline carapace length.
Metric Treatment n Mean SE Lower CI Upper CI Min Max p-value
Initial MCL (mm) Outdoor HS 24 45.4 0.4 44.6 46.1 41.4 47.6 0.51
Combo HS
24 45.0 0.4 44.2 45.8 41.0 47.7
Initial mass (g) Outdoor HS 24 21.1 0.5 20.1 22.1 16.5 23.6 0.63
Combo HS
24 20.9 0.5 19.8 21.9 16.8 25.0
Initial BC (g/cm3) Outdoor HS 24 0.54 0.01 0.53 0.55 0.49 0.60 0.10
Combo HS
24 0.53 0.01 0.51 0.54 0.45 0.58
Year 1 MCL (mm) Outdoor HS 24 66.5 1.5 63.4 69.5 52.2 76.9 <0.0001
Combo HS
24 102.5 1.5 99.6 105.5 84.9 116.6
Year 1 mass (g) Outdoor HS 24 62.6 7.4 47.9 77.5 34.3 85.5 <0.0001
Combo HS
24 205.8 7.2 191.4 220.3 124 302.0
Year 1 BC (g/cm3) Outdoor HS 24 0.57 0.01 0.56 0.58 0.51 0.63 0.14
Combo HS
24 0.59 0.01 0.57 0.60 0.50 0.65
Year 1 SHI Outdoor HS 24 94.8 1.31 92.1 97.4 90.2 97.2 0.39
Combo HS
24 95.9 0.6 94.8 97.1 92.9 98.2
Release MCL (mm) Outdoor HS 24 83.7 1.7 80.2 87.1 68.6 97.8 <0.0001
Combo HS
24 117.2 1.7 113.7 120.6 102.3 136.7
Release mass (g) Outdoor HS 24 115.9 10.7 94.3 137.4 67.3 175.9 <0.0001
Combo HS
24 300.6 10.7 279.0 322.2 200.0 421.0
Release BC (g/cm3) Outdoor HS 24 0.57 0.01 0.56 0.58 0.46 0.61 0.93
Combo HS
24 0.57 0.01 0.56 0.58 0.52 0.63
Release SHI Outdoor HS 24 95.7 0.6 94.5 96.8 92.5 97.2 0.0005
Combo HS 24 97.9 0.4 97.0 98.7 94.9 99.1

57
Table 2.2. Post-release movement metrics for 48 juvenile Mojave desert tortoises
(Gopherus agassizii) from two treatment groups (Combination head-started and Outdoor
head-started) released in September 2018 in the Mojave National Preserve, CA.
Displacement pre-dormancy represents dispersal from release burrow until dormancy for
all animals surviving to dormancy (n = 46; 25 September 2018 – 31 October 2019).
Displacement post-dormancy represents movement from dormancy burrow until the end
of the study for all animals that emerged post-dormancy (n = 45; 05 March 2018 – 23
July 2019). Total displacement represents the distance from release burrow until the end
of the study for all animals that emerged post-dormancy (n = 45; 25 September 2018 – 23
July 2019). Movement probability is defined as the probability of moving during any
weekly tracking event. BCI = Bayesian credible interval. We accepted significance at
alpha = 0.05 or if BCIs did not overlap. Significant p values are in bold.
Metric Treatment n Mean SE Min Max p-value
Displacement pre-
dormancy
Outdoor HS
Combo HS
23
23
255
114
89
40
26
0
1084
1183
0.11
Displacement post-
dormancy
Outdoor HS
Combo HS
22
23
92
53
34
19
0
0
896
231
0.299
Displacement 10-
months
Outdoor HS
Combo HS
22
23
326
143
93
41
11
9
1041
1178
0.047
n Estimate Lower
BCI
Upper Result
BCI
Movement probability
Outdoor HS
Combo HS
22
23
.42
.46
.38
.42
.47
.50
Not
different
Distance moved
during movement
events
Outdoor HS
Combo HS
22
23
43.4
36.2
34.1
29.7
54.6
44.7
Not
different

58
FIGURES
Figure 2.1. Midline carapace length (mm) at release for 2016 cohort juvenile Mojave
desert tortoises (Gopherus agassizii) from two treatment groups: Combination head-
started (“Combo HS”) and Outdoor head-started (“Outdoor HS”) in September 2018.
Dashed horizontal lines represent previously published release size recommendations
(Nagy et al. 2015, Hazard et al. 2015). The “whiskers” correspond to + 1.5 times the
interquartile range (IQR).
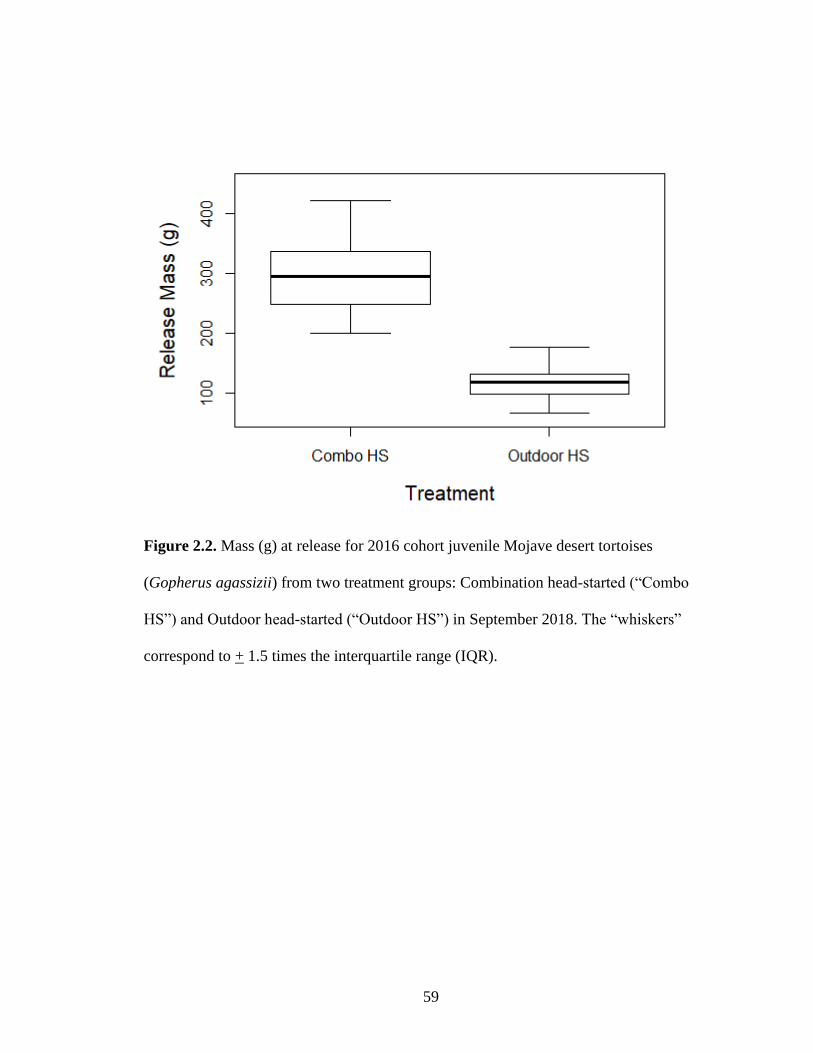
59
Figure 2.2. Mass (g) at release for 2016 cohort juvenile Mojave desert tortoises
(Gopherus agassizii) from two treatment groups: Combination head-started (“Combo
HS”) and Outdoor head-started (“Outdoor HS”) in September 2018. The “whiskers”
correspond to + 1.5 times the interquartile range (IQR).

60
Figure 2.3. Body condition (g/cm3) at release for 2016 cohort juvenile Mojave desert
tortoises (Gopherus agassizii) from two treatment groups: Combination head-started
(“Combo HS’) and Outdoor head-started (“Outdoor HS”) in September 2018. Body
condition calculated using Nagy et al. (2002) formula. The “whiskers” correspond to +
1.5 times the interquartile range (IQR).

61
Figure 2.4. Shell hardness index (SHI) versus midline carapace length (MCL) at Year 1
(left) and Year 2 (‘Release’; right) for 2016 cohort juvenile Mojave desert tortoises
(Gopherus agassizii) from two treatment groups: Combination head-started (“Combo
HS”) and Outdoor head-started (“Outdoor HS”) in September 2017 and September 2018.
Shell hardness index calculated using Nagy et al. (2011) protocols.

62
Figure 2.5. Shell hardness index (SHI) versus midline carapace length (MCL) for 2011,
2012, and 2016 cohort juvenile Mojave desert tortoises (Gopherus agassizii) from three
treatment groups: Combination head-started (“Combo HS”), 2016 Outdoor head-started
(“2016 Outdoor HS”), and 2011-2012 Outdoor head-started (“2011-2012 Outdoor HS”)
in September 2018. All three treatment groups were released concurrently in this study
and “2011-2012 Outdoor HS” animals are compared in Chapter 3. Shell hardness index
calculated using Nagy et al. (2011) protocols.
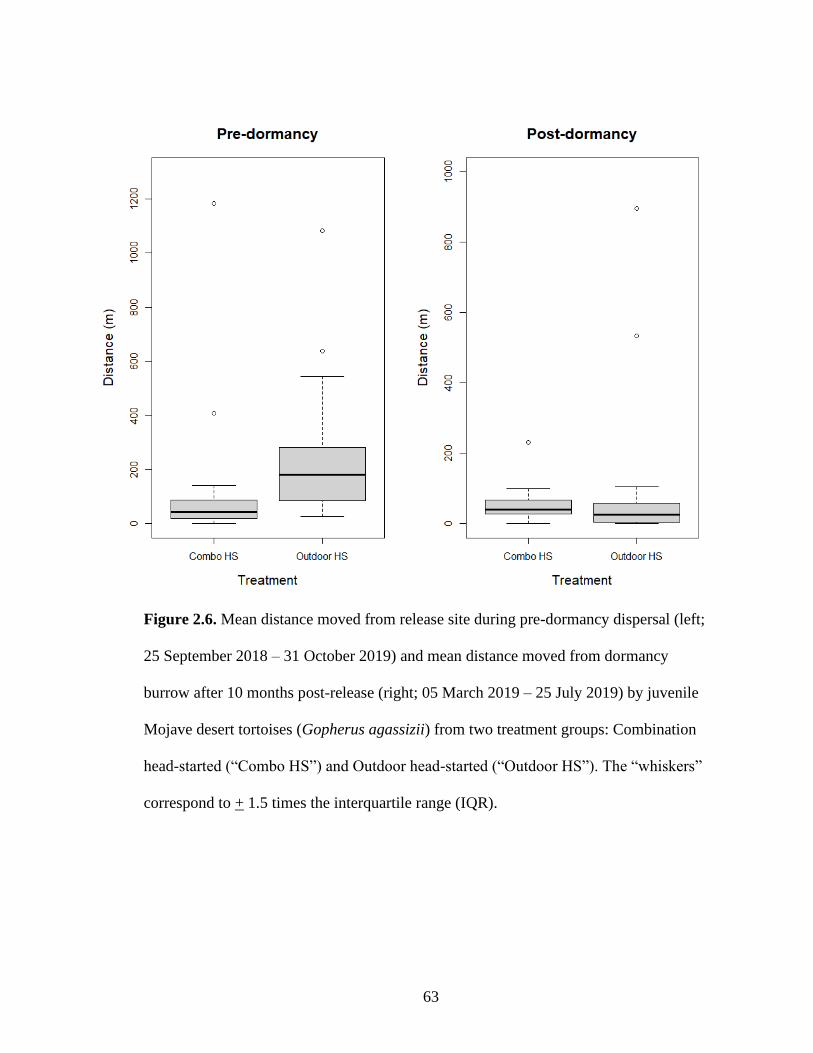
63
Figure 2.6. Mean distance moved from release site during pre-dormancy dispersal (left;
25 September 2018 – 31 October 2019) and mean distance moved from dormancy
burrow after 10 months post-release (right; 05 March 2019 – 25 July 2019) by juvenile
Mojave desert tortoises (Gopherus agassizii) from two treatment groups: Combination
head-started (“Combo HS”) and Outdoor head-started (“Outdoor HS”). The “whiskers”
correspond to + 1.5 times the interquartile range (IQR).

64
Figure 2.7. Kaplan-Meier survival curves for 48 Mojave desert tortoises (Gopherus
agassizii) in two treatment groups: Combination head-started (“Combo HS”; n = 24) and
Outdoor head-started (“Outdoor HS”; n = 24). Estimates are shown from 25 September
2018 until 23 July 2019 (43 weeks). Data were right-censored, meaning no assumptions
were made about the fate of lost or missing animals. Dashed vertical lines represent the
beginning and end of the dormancy period (31 October 2018 – 05 March 2019). Shaded
bands are 95% confidence intervals.

65
CHAPTER 3
SIZE AS A PREDICTOR OF POST-RELEASE SURVIVAL FOR MOJAVE DESERT
TORTOISES (GOPHERUS AGASSIZII)
INTRODUCTION
Positive relationships between size and survival have been demonstrated in a variety of
taxa (Wilbur and Collins 1973, Swingland et al. 1979, Murie and Boag 1984, Williams et
al. 1993). This relationship is especially evident in long-lived organisms exhibiting type
III survivorship, where mortality is inversely related with age and thus often, size
(Iverson 1991). Chelonians, generally characterized by low hatchling survival, slow
growth, delayed maturity, and high adult survival, are a prime example of an organism
with type III survivorship (Iverson 1991, Congdon et al. 1993, Heppell 1998). The
elevated survivorship of adult chelonians is a result of ontogenetic advances in size,
thermal and hydric homeostasis, mobility, and shell hardness (Wilson 1991, Haskell et al.
1996, Wilson et al. 2001, Nagy et al. 2011). Though the highest rates of survival in
chelonians are typically exhibited by adults, survival increases with each successive size-
class (Iverson 1991). This relationship presumably leads to differential rates of survival
across the size range of the juvenile life-stage (Haegen et al. 2009, Nagy et al. 2015a).
Growth rates, which affect the age at which juveniles reach a given size, can vary
widely as a result of environmental, climatic, demographic, and genetic factors (Ernst
1975, Rhen and Lang 1995, Steyermark and Spotila 2001, Mack et al. 2018). Ernst
(1975) measured the growth rates of a northern population of spotted turtles (Clemmys
guttata), determining that growth was limited by the length of the active season and the

66
corresponding water temperatures. Koper and Brooks (2000) measured growth rates of
juvenile painted turtles (Chrysemys picta) and corroborated previous findings that
warmer, longer active seasons increase growth (Parmenter 1980, Frazer et al. 1993).
Studies have also found that snapping turtles (Chelydra serpentina) grow faster in
habitats with high productivity (Brown et al. 1994), and that rapid growth in gopher
tortoises (Gopherus polyphemus) may be attributed to high quality habitat achieved
through active management (Mushinsky et al. 1994). These studies suggest that certain
factors, such as habitat quality and food availability, could potentially be managed to
improve growth in wild populations. In turn, captive reptile populations were studied to
determine whether husbandry conditions could be manipulated to accelerate growth rates
(Reiber et al. 1999, Jarvie et al. 2015, Daly et al. 2018).
Head-starting, the rearing of offspring in captivity until they are presumably more
likely to survive in the wild (Burke 2015), is gaining prominence among chelonian
recovery efforts and is predicated on the assumption that juvenile size at release is
positively correlated with post-release survival (Haegen et al. 2009, Buhlmann et al.
2015, Quinn et al. 2018). In addition to attempting to increase post-release survival by
releasing larger individuals, head-starting can increase the number of juveniles recruited
into the population at the release site. This is achieved by removing the threat of
predation on the vulnerable hatchling size class while providing optimal conditions for
growth during captivity. Although prioritizing habitat protection and high adult
survivorship remains paramount in conserving populations, head-starting may help
stabilize declining populations if implemented alongside other conservation measures
(Tomillo et al. 2008, Crawford et al. 2014, Spencer et al. 2017). This multi-faceted

67
approach, where head-starting is used as a short-term augmentation tool together with
more broadly focused conservation initiatives, is being applied in the recovery of the
Mojave desert tortoise (Gopherus agassizii; USFWS 2011).
The Mojave desert tortoise (hereafter ‘desert tortoise’), federally listed as
threatened in 1990, has experienced concerning population declines for the past half-
century (USFWS 2011). Contributing to these declines are multiple factors, including
habitat destruction, increased incidence of drought, and growing populations of
subsidized predators (Berry 1986, Foreman 1986, USFWS 2011). In 1989, researchers
began evaluating head-starting as a potential addition to the wide range of management
strategies used to mitigate desert tortoise population declines (Morafka et al. 1997).
Early head-starting projects focused on evaluating the health and survival of head-
starts during captivity (Morafka et al. 1997, Nagy et al. 2015a), but researchers soon
transitioned to assessing which factors most affected post-release success (Nagy et al.
2015b, Hazard et al. 2015). Based on three months of monitoring post-release
survivorship, Hazard et al. (2015) recommended the release of head-starts measuring at
least 84 mm midline carapace length (MCL). Subsequently, Nagy and colleagues (2015b)
found that annual survivorship reached 100% during non-drought years at their study site
for animals >100 mm MCL at time of release. Drought years induced prey-switching
behavior in coyotes (Canis latrans), resulting in losses of the larger individuals (>100
mm MCL) as well (Nagy et al. 2015b). Though these recommended release sizes may
seem similar, a 16-mm difference in MCL represents a minimum of one additional year
of growth in captivity under conventional outdoor rearing conditions (for growth rates see
Tuberville et al. 2019, Daly et al. 2018, Nagy et al. 2015b). As a result, 5-9 years of

68
outdoor rearing are required to reach previously recommended release sizes (Hazard et al.
2015, Nagy et al. 2015b). Therefore, refining the recommended release size has the
potential to both increase post-release success and maximize the efficiency of head-
starting desert tortoises.
In addition to the direct effects of size on survival, certain post-release behaviors
may affect survival of released head-starts, and these behaviors might be expected to vary
depending on an individual’s size or age. Long distance dispersal from the release site is
the primary elevator of mortality risk in herpetofaunal translocations (Germano and
Bishop 2009). Dispersal increases risk of exposure to both thermal extremes and
predation (Hazard and Morafka 2002, Stamps and Swaisgood 2007), thus distance moved
from release site may negatively correlate with survival probability. Increased exposure
and predation may also result from longer or more frequent bouts of surface activity.
Daly et al. (2019) found that all surviving tortoises released in their study were found out
of their burrows on less than 30% of tracking events, and decreased surface activity had a
positive effect on survival probability. Desert tortoises spend a large proportion of their
time inactive in their burrows (Zimmerman et al. 1994) but individuals vary in the
number of burrows used in an activity season (Harless et al. 2009). Because moving
between or relocating to a different burrow requires tortoises to be surface active, the
number of burrows used by an individual may be negatively associated with survival.
Alternatively, burrow switching could also be regarded as a form of predator avoidance
(Henen et al. 2017), with greater survival probability associated with individuals that
employ this strategy. Thus, both the tendency to be surface active and variation in burrow
switching may be important predictors of post-release survival.

69
Since 2011, our team has been evaluating the potential role of head-starting in
contributing to the recovery of the desert tortoise. During that time, we have conducted a
series of experimental studies (Nafus et al. 2015, Todd et al. 2016, Daly et al. 2018,
Tuberville et al. 2019, Daly et al. 2019) focused on identifying combinations of both pre-
and post-release conditions that maximize tortoise survival and the efficiency of head-
starting. The goal of our study is to evaluate the effect of release size on survival of head-
started desert tortoises during their first year following release. We examined this effect
by releasing captive-reared tortoises spanning a wide range of body sizes (68-145 mm
MCL). Along with size at release, we also assessed the effect of time spent in captivity,
post-release dispersal distance, surface activity, and burrow switching behavior on post-
release survival.
METHODS
Study site
Both the captive husbandry and field components took place within the Mojave
National Preserve (MNP), a 650,000 ha preserve in San Bernardino County, California.
Specifically, our captive-rearing facility, the Ivanpah Desert Tortoise Research Facility
(IDTRF), and our release site were in the Ivanpah Valley in the northeastern most corner
of the MNP. The primary habitat at our release site was “Yucca Woodland,” dominated
by creosote bush (Larrea tridentata), white bursage (Ambrosia dumosa), littleleaf ratany
(Krameria erecta), and heavily intermixed Yucca (Y. schidigera, and Y. brevifolia;
Turner et al. 1984; Todd et al. 2016). Although habitat suitability remains relatively high
in the Ivanpah Valley (Nussear et al., 2009), current tortoise densities (2.8 per km2 in
2012; Allison and McLuckie 2018) are much lower than they were historically (77 – 85

70
per km2 in 1977 to 1980; Turner et al. 1984). The captive-rearing portion of the study,
carried out at the IDTRF, made use of indoor rearing facilities and outdoor predator-proof
enclosures (further details below) located 15 km from our release site.
Obtaining hatchlings
In April 2011, we began tracking 30 adult female desert tortoises from our field
site in the Ivanpah Valley. We tracked females monthly using a 3-element Yagi antenna
(AF Antronics, Inc., Urbana, IL) and a R1000 receiver (Communications Specialists,
Inc., Orange, CA). Each April (except 2014) we brought all females to the IDTRF and
radiographed them to check for the presence of calcified eggs (Diagnostic Imaging
Systems, Poskam, Colorado, USA; 60 kvp, 0.8 mAS, 74 cm focal length; Gibbons and
Greene 1979). We placed gravid females in individual nesting enclosures (5 x 9 m) and
allowed them to nest naturally. Nesting enclosures were constructed to deter both avian
and mammalian predators and contained natural vegetation for shade and forage. We
provided females with artificial burrows and supplemental water. Weekly, we monitored
for egg deposition by radiographing females in the pens. If nesting was confirmed, we
immediately released the female at her point of capture. Accordant with our permits, we
released, at their last known burrow location, any females that had not nested within 30
days.
Eggs incubated naturally in the enclosures. After approximately 80 days, we
began searching the pens daily for emerged hatchlings. We marked all hatchlings by
notching unique combinations of marginal scutes using codes assigned by USFWS
(Cagle 1939). We carefully excavated all nests with un-emerged hatchlings to recover
any unhatched eggs or entombed hatchlings. We randomly assigned all healthy neonates

71
to rearing enclosures at the conclusion of the hatching season. This study includes
juveniles from the 2011, 2012 and 2016 cohorts.
Pre-release husbandry
Each year we placed all (prior to 2015) or a subset (2015 and later) of the available
hatchlings in predator-proof enclosures at the IDTRF to be reared solely outdoors. The
enclosures contained natural substrate and vegetation (Alberts 2007) which we
supplemented with rocks, downed Yucca logs, and “starter burrows” as shelter sites. We
also outfitted each enclosure with rotating garden sprinklers to provide artificial rain
every 1-2 weeks during the tortoise active season (March – October; for more
information see Tuberville et al. 2019, Daly et al. 2018). Husbandry protocols within the
enclosures have been modified over the course of the head-starting program, as noted
below, to respond to changing habitat conditions in enclosures and different research
objectives.
Initially, tortoises from the 2011-2015 cohorts received only supplemental rain
and relied solely on natural vegetative growth in the pens for forage (Beatley 1974).
Starting in March 2016, due to the depletion of native forage within the pens, we began
providing supplemental food during each artificial watering event. Supplemental food, a
measured mix of leafy greens and Mazuri® Tortoise Diet (Mazuri Exotic Animal
Nutrition, St. Louis, MO; see Chapter 2 and Daly et al. 2018), approximated the
nutritional properties of the desert tortoise’s natural diet (Jarchow et al. 2002). Thus, the
2011-2015 cohorts initially received only supplemental water but starting in Spring 2016
also received supplemental food, whereas the 2016 cohort received both supplemental
water and food throughout their outdoor rearing.

72
Beginning in 2015, we began rearing a subset of each cohort indoors in an attempt
to accelerate their growth while in captivity (Daly et al. 2018). Daly et al. (2018)
compared indoor captive-rearing to conventional outdoor captive-rearing, finding that
indoor-reared tortoises (7 mo) grew significantly faster (8x) than their outdoor-reared
siblings but lacked the shell hardness of similar sized, yet older (3-4 yrs old) outdoor-
reared individuals. To combine the increased growth of indoor rearing and the increased
shell hardness of outdoor rearing, in fall 2016 we initiated a novel method of combination
rearing. Combination rearing consisted of one year of indoor rearing followed by one
year of outdoor rearing in predator-proof enclosures (Chapter 2), with the outdoor rearing
portion including both supplemental water and food as previously described.
As part of the current study, we released 78 tortoises from three different cohorts,
representing a wide range of release sizes. We released 30 tortoises from our 2011-2012
cohorts. We released 48 tortoises from our 2016 cohort – half of which were reared solely
outdoors while the other half was combination reared. By releasing animals from these
three cohorts (2011, 2012, 2016) reared under the protocols described above, we were
able to release tortoises at sizes that bracketed the previously recommended release sizes
(84 mm MCL, Hazard et al. 2015; 100 mm MCL, Nagy et al. 2015b). We did not
consider husbandry treatment in analyzing individual fate because previous releases at
our site have failed to detect differences in post-release survival and behavior based on
husbandry treatment groups (Tuberville et al. 2019, Daly et al. 2019, Chapter 2).
Morphometrics and growth
We measured all hatchlings immediately after they emerged from nests. After
these initial measurements, we measured animals each fall (September), including

73
immediately prior to release (25 September 2018). At each measuring period, we
recorded midline carapace length (MCL, straight-line distance from the anterior edge of
the nuchal scute to the inside of the natural notch in the supracaudal scute) to the nearest
0.1 mm using vernier calipers. We compared mean MCL at hatching among cohorts
using linear mixed effects models (‘nlme’ package, ‘lme’ function in R) with mother
identification number as a random effect. Similarly, we compared mean MCL at release
among cohorts. When a significant difference was detected (α = 0.05 rate of type I error),
we used Tukey’s post-hoc comparisons (‘glht’ function in the ‘multcomp’ package) to
investigate pairwise differences.
To calculate mean annual growth rates for each individual, we separated the
captive-rearing period into three husbandry stages; 1) outdoor rearing with no
supplemental food, 2) outdoor rearing with supplemental food, and 3) indoor rearing.
Thus, for tortoises from our 2011-2012 cohorts, we calculated separate mean annual
growth rates for their time spent in pens with no supplemental food and for their time
spent in pens with supplemental food. Additionally, we calculated mean annual growth
rates for their first two years of life, allowing us to compare growth of same-aged
tortoises reared outdoors with (2016 cohort) and without (2011-2012 cohorts)
supplemental food. For the combination-reared tortoises, we calculated separate annual
growth rates for the indoor and outdoor rearing stages. To calculate annual growth rates,
we divided the change in MCL between initial and last fall measurements by the number
of years reared under that husbandry stage. For the 2011-2012 cohorts, for which we
modified outdoor husbandry protocols to include supplemental feeding during spring
2016, we used fall 2015 measurements as final measurements for calculating growth for

74
outdoor rearing with no supplemental feeding and used those same measurements as
initial sizes to calculate growth resulting from outdoor rearing with supplemental food.
We felt comfortable using fall 2015 measurements despite the March 2016 change in
husbandry, as previous studies have found that desert tortoises exhibit almost no growth
before April and after July each year (Medica et al. 1975). Data are presented as means ±
1 SE.
Experimental releases
In fall 2018, we released all juveniles into Yucca woodland habitat at our site. The
specific release site was chosen based on habitat quality, its accessible yet remote
location, and previously published recommendations on minimum distance from potential
perch sites for avian predators, namely ravens (Nafus et al. 2015, Todd et al. 2016, Daly
et al. 2019). Using ArcGIS (version 10.5; ESRI, Redlands, CA), we first delineated a 0.6
km2 rectangular release plot (300 m X 2000 m) parallel to our lone sandy access road,
with the nearest corner >1.9 km from a powerline near our site. We then used the
‘Fishnet’ function to generate release points throughout the plot at 50-m intervals. Next,
we removed points to create three equally spaced blocks of release points within the
rectangular plot. Points were then labeled 1–78 to correspond with a release location for
each juvenile tortoise. Each block, separated from each other by approximately 350 m,
comprised 26 points. Release points were buffered 10 m to maximize our ability to select
a release refugium for each juvenile that contained a large perennial shrub and an intact
kangaroo rat burrow (Dipodomys spp.; Nafus et al. 2015, Todd et al. 2016) at each
generated release location. Each juvenile tortoise was released head-first into a kangaroo
rat burrow that we had previously enlarged as needed to safely conceal the released

75
animal. Under this release protocol, taking into account the buffered limit of each release
point, the minimum possible distance between release burrows was 30 m. We conducted
bouts of tracking effort by replicate release block to most efficiently track the large
number of released animals.
Prior to release, we held all juveniles indoors for 3–5 days at the IDTRF, allowing
us to soak and feed tortoises, visually assess their health, and attach transmitters. We
randomly assigned each juvenile, irrespective of treatment, to one of the 78 release
centers (Figure 3.1). We released all juveniles on 25 September 2018. We released
animals in their pre-selected refugium between 0600 and 1000 h to avoid extreme
temperatures.
Post-release monitoring
We attached radio-transmitters to juveniles prior to their release. We affixed
transmitters (3.1 g R1670 on 2016 animals reared solely outdoors; 3.6 g R1680
transmitters on all others; Advanced Telemetry Systems, MN, USA) to the fifth vertebral
scute with 5-minute epoxy (Devcon 5-minute epoxy gel, ITW Engineered Polymers,
County Clare, Ireland). Transmitters weighed no more than 5% of the body mass at time
of release of the recipient tortoise (apart from the three smallest 2016 outdoor animals for
which the transmitter weighed 6% of the pre-attachment body mass). To help camouflage
the transmitter bundle, we coated all transmitters with a thin layer of sand before the
epoxy hardened (Kazmaier et al. 2002).
We tracked each animal 24 hours after initial release and then twice per week for
the next three weeks. We then tracked juveniles weekly until dormancy (31 October
2018), then every 10-14 days during dormancy (November 2018 – February 2019). As

76
tortoises became active in the spring (March 2019), we resumed weekly tracking for the
remainder of the study (March 2019 – July 2019). At each tracking occasion, we recorded
the tortoise’s location to the nearest ±3 m using a handheld GPS (Garmin model
GPSMAP 76, Olathe, Kansas). We used these tracking occasions to monitor post-release
movement and behavior (surface activity, burrow switching), which were subsequently
used as predictors in our survival models (details below).
We used the Spherical Law of Cosines (Movable Type Ltd. 2015) to calculate, for
each individual tortoise, the straight-line distance between its release burrow and its final
location (total displacement). We set its final location as where it was found on the last
day of the study (23 July 2019) for all individuals surviving the study or, for those that
died, their last known live location. We calculated surface activity as the number of times
a tortoise was found outside a burrow or pallet (burrow with width longer than length)
divided by the total number of tracking occasions. We defined burrow switching as the
number of unique burrows used by an individual divided by the total number of tracking
occasions that individual was found in a burrow; the proportion thus accounted for
differences in number of tracking occasions among individuals due to mortalities during
the study.
Survivorship
We estimated post-release survival rates through the end of the monitoring period
(September 2018–July 2019) using the Kaplan-Meier estimator in the R package
‘survival’ (R Core Team 2017). When a deceased tortoise was found, we carefully
inspected the surrounding area for any signs of what may have caused the death of the
individual. Signs of predation included digging, tracks, scat, and teeth marks or beak

77
punctures on the carcass or transmitter. We photographed each carcass in situ and
collected them for further analysis. We attributed mortalities to one of five
classifications: mammal depredated, bird depredated, unidentified predator, exposure, or
unknown fate. Clear digging activity or obvious tooth marks suggested mammal
predation. We classified remains near perch sites with clear signs of being struck by a
beak as bird depredated. If the carcass was damaged but remains showed no evidence of
tooth marks or avian predation, we considered it to be depredated by an unidentified
predator. We presumed dead tortoises found intact either on the surface or in their
burrows to have been killed by exposure. We classified animals lost due to radio failure
(faulty batteries or the unit fell off) as unknown fate, and we right censored (no
assumption of fate) these animals in our survival analysis.
We evaluated the effect of several factors on individual fate, using only the
individuals with a known fate at the end of the study. We included time spent in captivity,
MCL at release, total displacement, surface activity, and burrow switching as potential
predictors in a combination of generalized linear models (GLMs) with a binary response
(alive = 1 and dead = 0). Coefficients of correlation (r) among predictor variables were
all <0.65, with time in captivity and MCL at release being the most highly correlated (r =
0.63). We then used Akaike information criterion (AIC) values to compare among the
competing models and evaluate model fit. To account for uncertainty in model selection,
we used multi-model averaging (R package ‘AICcmodavg’). We generated model-
averaged predictions and 95% confidence intervals (CIs) of survival across a range (60-
150 mm) of MCL values, holding all other predictor variables to their average values. We

78
kept all predictor variables other than MCL constant because MCL at release is the sole
predictor under a managers control.
RESULTS
Morphometrics and growth
Mean MCL at hatching was 45.1 ± 0.2 mm (n = 78), though it differed among
cohorts (F2,48 = 9.022, p < 0.0001). Mean hatching MCL for the 2011 cohort (45.9 ± 0.3
mm) and 2016 cohort (45.2 ± 0.2 mm) was significantly larger than for the 2012 cohort
(43.9 ± 0.2 mm; p < 0.032 in both cases). For the 2011-2012 cohorts, growth during
outdoor rearing averaged 11.4 ± 0.4 mm MCL/yr with no supplemental food (2011–
2015) and 13.5 ± 0.6 mm MCL/yr with the addition of supplemental food (2016–2018).
During their first two years of life (2011-2013 and 2012-2014 for our 2011 and 2012
cohorts, respectively), 2011-2012 animals grew 11.0 ± 0.4 mm MCL/yr. Juveniles from
our 2016 cohort reared solely outdoors with supplemental food grew 19.2 ± 0.7 mm
MCL/yr. Tortoises reared using combination rearing grew 57.5 ± 2.0 mm MCL/yr while
indoors (September 2016 – September 2017) and 14.7 ± 1.3 mm MCL/yr while outdoors
with supplemental food (September 2017 – September 2018). The overall combination of
rearing conditions and cohorts used in this study resulted in release sizes ranging from
68–145 mm MCL. Mean time spent in captivity among released animals was 3.64 years
(range = 2 – 7).
Surface activity and burrow switching
Mean total displacement over the course of the 10-month post-release monitoring
period was 295 ± 55 m (n = 76; range = 9 – 3062 m). Proportion of surface-active
tracking events averaged 0.43 ± 0.03 (n = 76; range = 0.13 – 1.0) among all released

79
head-starts with a known fate at the end of the study. The mean number of unique
burrows used by each individual over the monitoring period (25 September 2018 – 23
July 2019), or until the animal died, was 5.3 ± 0.21 (range = 0 – 9). Mean burrow
switching by tortoises in our study was 0.35 ± 0.03 (range 0.0 – 1.0).
Survivorship
Of the 78 released tortoises, 63 (80.8%) were confirmed alive at the end of the
study (23 July 2019). Based on analysis of each deceased animal in the field as well as
the surroundings of each carcass, all mortalities were attributed to predation. Mammal
predators were responsible for the largest percentage of mortality (12.8% of released
tortoises). Three (3.8%) head-starts were bird predated and two (2.6%) were lost due to
radio failure. Notably, all tortoises depredated by avian predators were 2016 solely
outdoor reared animals (68-98 mm MCL at release). Based on the 78 tortoises released in
this study, total 10-month post-release survival was estimated at 0.83 (95% CI, 0.75-0.92;
Figure 3.2).
No model for survival in the candidate set gathered >0.25 of AIC model weight,
indicating high uncertainty in model selection (Table 3.1). In each of the 4 most
competitive models in the set (△AIC < 2.0), only MCL was a significant predictor of fate
(p 0.009). Model-averaged predictions over the range of MCL (with all other measured
covariates held constant at their respective means) showed that survival probability
increased with size at release (Table 3.2; Figure 3.3). Ten-month predicted survival
probability ranged from 0.65 (0.42 – 0.83) to 0.93 (0.78 – 0.98) at the MCL limits used in
this study (60 – 150 mm MCL). Head-starts released at our study site had predicted
survival probabilities of 0.76 (0.64 – 0.86) and 0.82 (0.71 – 0.89) at the previously

80
recommended release sizes of 84 mm MCL and 100 mm MCL (Hazard et al. 2015, Nagy
et al. 2015b, respectively).
DISCUSSION
Among chelonians, smaller individuals often have higher rates of mortality than larger
conspecifics (Haskell et al. 1996, O’Brien et al. 2005). The mortalities documented in our
study followed this trend. Over half (7/13; 54%) of the mortalities we documented were
of animals smaller than 91 mm MCL, a disproportionate amount given that only 27%
(21/78) of the animals released were below this size threshold. Furthermore, the three
smallest individuals released in this study were all found depredated. Though we
evaluated a suite of possible predictors on individual fate, size was the sole significant
predictor among those we examined of whether an animal lived or died in our study.
A positive relationship between size and survival has been previously reported for
desert tortoises (Hazard et al. 2015, Nagy et al. 2015b). Nagy et al. (2015b) found that
survivorship reached 100% for tortoises > 100 mm MCL in their first-year post-release,
adding that mortality was highest amongst the smallest individuals (<80 mm MCL), none
of which survived longer than four years post-release. Another study at that same site in
the western Mojave Desert found that no animal above 84 mm MCL died during the short
three-month monitoring period following release (Hazard et al. 2015). Notably, both
studies were predominantly affected by raven predation. In fact, Hazard et al. (2015)
reported ravens as the sole source of mortality, whereas only three (23% of mortalities)
individuals in our study were concluded to have been killed by avian predators
(presumably ravens). The size of our largest raven-depredated individual (90 mm MCL)
was similar to that reported by both Nagy et al. (2015b; 80 mm MCL) and Hazard et al.

81
(2015; 84 mm MCL) for raven predation at their site. Notably, Nagy et al. (2015b) found
that raven predation was likely the cause for the near complete eradication of tortoises
below 80 mm MCL from their study over a three-year period. The three raven-depredated
juveniles in our study ranged between 68-90 mm MCL, the two smallest of which
showed clear signs of being punctured by a beak while the largest individual lacked
puncture wounds. Our results, coupled with previous findings of size thresholds for avian
predation (Hazard et al. 2015, Nagy et al. 2015b), suggest that risk of raven-related
mortality is reduced as tortoises grow and their shells harden (Nagy et al. 2011).
Age and size are often too correlated to reliably differentiate their relative effects
on response variables. However, as a result of overlapping sizes among cohorts due to
differential husbandry practices used in this study, we were able to separately evaluate
size and age (time spent in captivity) as predictors of post-release fate. We found that
size, rather than age, was the sole significant predictor of fate at our site, with larger
juveniles exhibiting greater survival than same-aged, smaller individuals. This finding
suggests that accelerating growth of hatchling head-starts holds the potential to maximize
the success and efficiency of head-starting programs. For example, simply by
incorporating both an indoor and outdoor component in the pre-release husbandry of
head-started desert tortoises, our study shows that head-starting programs can rear
tortoises equivalent in size to six-year-old outdoor reared animals in just two years. This
reduction in the captive period equates to four years of savings on husbandry- and
personnel-related expenditures, greatly increasing the potential for programs to expand
the number of head-starts that can be reared each year or result in cost-savings that can be
invested in additional management practices for the benefit of the species (i.e. restoration,

82
habitat protection). Importantly, based on our releases to date, we have found no evidence
that the accelerated growth rates associated with indoor rearing, at least under the
husbandry practices we used, result in physiological impairment or differences in post-
release behavior or survival (Chapter 2, Tuberville et al. 2019, Daly et al. 2018, 2019).
Reducing time spent in captivity may also hold physiological benefits, in addition
to saving limited conservation dollars (Warkentin et al. 1990, Hermes et al. 2004, de
Assis et al. 2015). A study on the effects of long-term captivity on the stress response of
the Cururu toad (Rhinella icterica), found that prolonged captivity led to elevated stress
hormone levels over the duration of the captive period as well as decreased immune
response (de Assis et al. 2015). Long-term captivity has also been implicated in increased
basal metabolic rate and decreased antimicrobial fighting ability in birds (Warkentin et al.
1990, Buehler et al. 2008). In reptiles specifically, time spent in captivity has been found
to alter feeding behaviors and negatively affect post-release survival in rat snakes
(Pantherophis obsoletus; DeGregorio et al. 2013, 2017). Our finding – that time in
captivity can be reduced by incorporating an indoor rearing component – limits the
potential for manifestation of any negative consequences that might be associated with
prolonged captivity.
Tortoise growth responds to husbandry manipulations, as seen by the variable
growth rates of tortoises reared under the differing husbandry protocols implemented in
this study. Maximum growth was observed when using indoor rearing, during which
head-starts grew nearly three times faster (57.5 mm MCL/yr) than the fastest growing
cohort of solely outdoor reared juveniles (19.2 mm MCL/yr) at our site. Alternatively, for
programs focused on or logistically constrained to solely using outdoor rearing, we found

83
that growth was vastly improved in same-aged juvenile tortoises by the addition of
supplemental food, coupled with supplemental watering. While using indoor rearing is
clearly the most efficient way to accelerate growth, outdoor rearing allows head-starts to
acclimate to outdoor conditions and develop natural thermoregulatory behaviors. Given
that we did not release solely indoor-reared tortoises as part of this study, we cannot be
certain how lack of outdoor conditioning would have affected post-release survival
relative to the head-starts released in our study. Solely indoor-reared tortoises released by
Daly et al. (2019) at our study site exhibited survival rates similar to both outdoor reared
and direct released siblings, although any potential differences may have been masked by
high predation rates associated with a single brooding pair of ravens that became
established shortly after the tortoises were released. We were able to minimize this source
of mortality in our study by releasing animals in the fall after raven nesting was
completed and by releasing animals > 1.9 km away from powerline pylons that serve as
raven perch structures (Daly et al. 2019).
In summary, our study lends further support for the positive relationship between
size and post-release survival in head-started juvenile desert tortoises, while highlighting
how the higher growth rates associated with the indoor phase of combination rearing can
improve the efficiency of head-starting programs. We found that size, not age, best
predicts post-release survival and corroborated that indoor rearing allows for significantly
increased growth in comparison to conventional outdoor rearing (Daly et al. 2018). We
include estimated survival probabilities over a range of juvenile desert tortoise sizes,
providing managers with an increased ability to predict outcomes of subsequent head-
start releases based on release size. At our study site, raven predation only affected

84
animals < 90 mm MCL, corroborating previous size thresholds for raven predation at a
study site in the western Mojave Desert (Hazard et al. 2015, Nagy et al. 2015b). Though
raven predation ceased above 90 mm MCL, predicted survival continued to increase
incrementally over the entire MCL range used in this study. We provide survival
estimates over a wide range of potential release sizes because recommending a single
specific size at release for future desert tortoise head-starting programs fails to recognize
the complexity of variable release site conditions and logistical constraints faced by
individual projects. In producing our survival estimates, we acknowledge that survival
will vary depending on habitat quality, predator abundance, climatic conditions, and other
sources of variability. Practitioners must factor in their own site-specific conditions and
program-specific objectives when determining the ideal size at release and the most
efficient, logistically feasible method of attaining that size.

85
LITERATURE CITED
Alberts, A.C. 2007. Behavioral considerations of headstarting as a conservation strategy
for endangered Caribbean rock iguanas. Applied Animal Behaviour Science
102:380–391.
Allison, L.J., and A.M. McLuckie. 2018. Population trends in Mojave Desert tortoises
(Gopherus agassizii). Herpetological Conservation and Biology 13:433–452.
Beatley, J.C. 1974. Phenological events and their environmental triggers in Mojave
Desert ecosystems. Ecology 55:856–863.
Belthoff, J.R. and A.M. Dufty Jr. 1998. Corticosterone, body condition and locomotor
activity: a model for dispersal in screech-owls. Animal Behaviour 55:405–415.
Berry, K. 1986. Desert tortoise (Gopherus agassizii) research in California, 1976-1985.
Herpetologica 42:62–67.
Bertolero, A., J.L Pretus, and D. Oro. 2018. The importance of including survival release
costs when assessing viability in reptile translocations. Biological
Conservation 217:311–320.
Boulon, R.H., 1994. Growth rates of wild juvenile hawksbill turtles, Eretmochelys
imbricata, in St. Thomas, United States Virgin Islands. Copeia 811–814.
Brown, G.P., C.A. Bishop, and R.J. Brooks. 1994. Growth rate, reproductive output, and
temperature selection of snapping turtles in habitats of different productivities.
Journal of Herpetology 28:405–410.
Buehler, D.M., T. Piersma, and B. Irene Tieleman. 2008. Captive and free‐living red
knots Calidris canutus exhibit differences in non‐induced immunity that suggest

86
different immune strategies in different environments. Journal of Avian
Biology 39:560–566.
Buhlmann, K. A., S. L. Koch, B. O. Butler, T. D. Tuberville, V. J. Palermo, B. A.
Bastarache, and Z. A. Cava. 2015. Reintroduction and head-starting: tools for
Blanding’s Turtle (Emydoidea blandingii) conservation. Herpetological
Conservation and Biology 10:436–454.
Burke, L. R. 2015. Head-starting turtles: learning from experience. Herpetological
Conservation and Biology 10:299–308.
Cagle, F.R. 1939. A system of marking turtles for future identification. Copeia 1939:170–
173.
Congdon, J.D., A.E. Dunham, and R.C. van Loben Sels. 1993. Delayed sexual maturity
and demographics of Blanding's turtles (Emydoidea blandingii): implications for
conservation and management of long‐lived organisms. Conservation
Biology 7:826–833.
Crawford, B.A., J.C. Maerz, N.P. Nibbelink, K.A. Buhlmann, and T.M. Norton. 2014.
Estimating the consequences of multiple threats and management strategies for
semi-aquatic turtles. Journal of Applied Ecology 51:359–366.
Daly, J.A. 2017. Indoor rearing as a component of head-starting the Mojave Desert
Tortoise (Gopherus agassizii). M.S. Thesis. University of Georgia, Athens.
Daly, J.A., K.A. Buhlmann, B.D. Todd, C.T. Moore, J.M. Peaden, and T.D. Tuberville.
2018. Comparing growth and body condition of indoor-reared, outdoor-reared,
and direct-released juvenile Mojave Desert Tortoises. Herpetological
Conservation and Biology 13:622–633.

87
Daly, J.A., K.A. Buhlmann, B.D. Todd, C.T. Moore, J.M. Peaden, and T.D. Tuberville.
2019. Survival and movements of head‐started Mojave desert tortoises. The
Journal of Wildlife Management 83:700–1710.
de Assis, V.R., S.C.M. Titon, A.M.G. Barsotti, B. Titon Jr, and F.R. Gomes. 2015.
Effects of acute restraint stress, prolonged captivity stress and transdermal
corticosterone application on immunocompetence and plasma levels of
corticosterone on the cururu toad (Rhinella icterica). PloS One 10:p.e0121005.
Ernst, C.H. 1975. Growth of the spotted turtle, Clemmys guttata. Journal of Herpetology
9:313–318.
Foreman, L.D., J.M. Brode, R. Haussler, and K. Kramer. 1986. The responsibilities of
federal and state agencies for protection of the desert tortoise in California.
Herpetologica 1:59.
Frazer, N.B., J.L. Greene, and J.W. Gibbons. 1993. Temporal variation in growth rate and
age at maturity of male painted turtles, Chrysemys picta. American Midland
Naturalist 130:314–324.
Gibbons, J. W., and J. L. Greene. 1979. X-ray photography: a technique to determine
reproductive patterns of freshwater turtles. Herpetologica 86–89.
Haegen, W.M.V., S.L. Clark, K.M. Perillo, D.P. Anderson, and H.L. Allen. 2009.
Survival and causes of mortality of head‐started western pond turtles on Pierce
National Wildlife Refuge, Washington. The Journal of Wildlife
Management 73:1402–1406.
Harless, M.L., A.D. Walde, D.K. Delaney, L.L. Pater, and W.K. Hayes. 2009. Home
range, spatial overlap, and burrow use of the desert tortoise in the West Mojave

88
Desert. Copeia 378–389.
Haskell, A., T.E. Graham, C.R. Griffin, and J.B. Hestbeck. 1996. Size related survival of
headstarted redbelly turtles (Pseudemys rubriventris) in Massachusetts. Journal of
Herpetology 30:524–527.
Hazard, L.C., D.J. Morafka, and L.S. Hillard. 2015. Post-release dispersal and predation
of head-started juvenile desert tortoises (Gopherus agassizii): effect of release site
distance on homing behavior. Herpetological Conservation and Biology 10:504–
515.
Henen, B.T., U.P. Van Bloemestein, M.D. Hofmeyr, and C.A. Weatherby. 2017.
Variation in the daily activity, movement and refugia of Critically Endangered
geometric tortoises, Psammobates geometricus, in autumn and spring. African
Journal of Herpetology 66:79–92.
Heppell, S. 1998. Application of life-history theory and population model analysis to
turtle conservation. Copeia 1998:367–375.
Hermes, R., T.B. Hildebrandt, and F. Göritz. 2004. Reproductive problems directly
attributable to long-term captivity–asymmetric reproductive aging. Animal
Reproduction Science 82:49–60.
Iverson, J.B. 1991. Life history and demography of the yellow mud turtle, Kinosternon
flavescens. Herpetologica 373–395.
Jarchow, J.L., H.E. Lawler, T.R. Van Devender, and C.S. Ivanyi. 2002. Care and diet of
captive Sonoran desert tortoises. Pages 289–311 in T. R. Van Devender, editor.
The Sonoran Desert Tortoise. The University of Arizona Press, Tucson, AZ.
Jarvie, S., A.M. Senior, S.C. Adolph, P.J. Seddon, and A. Cree. 2015. Captive rearing

89
affects growth but not survival in translocated juvenile tuatara. Journal of Zoology
297:184–193.
Kazmaier, R.T., E.C. Hellgren, and D.C. Ruthven, III. 2002. Home range and dispersal of
Texas tortoises, Gopherus berlandieri, in a managed thornscrub ecosystem.
Chelonian Conservation Biology 4:488–496.
Koper, N., and R.J. Brooks. 2000. Environmental constraints on growth of painted turtles
(Chrysemys picta) in northern climates. Herpetologica 421–432.
Loehr, V., B. Henen, and M. Hofmeyr. 2004. Reproduction of the smallest tortoise, the
Namaqualand speckled padloper, Homopus signatus signatus. Herpetologica
60:759–761.
Mack, J.S., H.E. Schneider, and K.H. Berry. 2018. Crowding Affects Health, Growth,
and Behavior in Headstart Pens for Agassiz's Desert Tortoise. Chelonian
Conservation and Biology 17:14–26.
Medica, P.A., R.B. Bury, and F.B. Turner. 1975. Growth of the desert tortoise (Gopherus
agassizi) in Nevada. Copeia 639–643.
Morafka, D.J., K.H. Berry, and E.K. Spangenberg. 1997. Predator-proof field enclosures
for enhancing hatchling success and survivorship of juvenile tortoises: a critical
evaluation. Conservation, Restoration, and Management of Tortoises and Turtles,
An International Conference. 147–165.
Murie, J.O., and Boag, D.A. 1984. The relationship of body weight to overwinter survival
in Columbian ground squirrels. Journal of Mammalogy 65:688–690.
Mushinsky, H.R., D.S. Wilson, and E.D. McCoy. 1994. Growth and sexual dimorphism
of Gopherus polyphemus in central Florida. Herpetologica 119–128.

90
Nafus, M.G., J.M. Germano, J.A. Perry, B.D. Todd, A. Walsh, and R.R. Swaisgood.
2015. Hiding in plain sight: a study on camouflage and habitat selection in a slow-
moving desert herbivore. Behavioral Ecology 26:1389–1394.
Nagy, K., M. Tuma, and L. Hillard. 2011. Shell hardness measurements in juvenile desert
tortoises Gopherus agassizii. Herpetological Review 42:191–195.
Nagy, K.A., L.S. Hillard, M.W. Tuma, and D.J. Morafka. 2015a. Head-started desert
tortoises (Gopherus agassizii): movements, survivorship and mortality causes
following their releases. Herpetological Conservation and Biology 10:203–215.
Nagy, K.A., L.S. Hillard, S. Dickson, and D.J. Morafka. 2015b. Effects of artificial rain
on survivorship, body condition, and growth of head-started desert tortoises
(Gopherus agassizii) released to the open desert. Herpetological Conservation and
Biology 10:535–549.
Nussear, K. E., T. C. Esque, R. D. Inman, L. Gass, K. A. Thomas, C. S. A. Wallace, J. B.
Blainey, D. M. Miller, and R. H. Webb. 2009. Modeling habitat of the Desert
Tortoise (Gopherus agassizii) in the Mojave and parts of the Sonoran Deserts of
California, Nevada, Utah, and Arizona. U.S. Geological Survey Open-File Report
2009-1102. 18 p.
Nussear, K.E., C.R. Tracy, P.A. Medica, D.S. Wilson, R.W. Marlow, and P.S. Corn.
2012. Translocation as a conservation tool for Agassiz's desert tortoises:
survivorship, reproduction, and movements. The Journal of Wildlife
Management 76:1341–1353.
O’Brien, S., B. Robert, and H. Tiandray. 2005. Hatch size, somatic growth rate and size-
dependent survival in the endangered ploughshare tortoise. Biological

91
Conservation 126:141–145.
Parmenter, R.R. 1980. Effects of food availability and water temperature on the feeding
ecology of pond sliders (Chrysemys s. scripta). Copeia 1980:503–514.
Quinn, D.P., K.A. Buhlmann, J.B. Jensen, T.M. Norton, and T.D. Tuberville. 2018. Post‐
release movement and survivorship of head‐started gopher tortoises. The Journal
of Wildlife Management 82:1545–1554.
Rautenstrauch, K.R., A.L.H. Rager, and D.L. Rakestraw. 1998. Winter behavior of desert
tortoises in southcentral Nevada. The Journal of Wildlife Management 62:98–
104.
R Core Team. 2017. R: a language and environment for statistical computing. Foundation
for Statistical Computing, Vienna, Austria.
Reiber, C.L., S. Malekpour, and M. McDaniel. 1999. Effects of post-hatching
maintenance temperature on desert tortoise (Gopherus agassizii) shell morphology
and thermoregulatory behavior. Journal of Herpetology 33(2):234–240.
Rhen, T. and J.W. Lang. 1995. Phenotypic plasticity for growth in the common snapping
turtle: effects of incubation temperature, clutch, and their interaction. The
American Naturalist 146:726–747.
Spencer, R. J., J.U. Dyke, and M.B. Thompson. 2017. Critically evaluating best
management practices for preventing freshwater turtle extinctions. Conservation
Biology 31:1340–1349.
Steyermark, A.C. and J.R. Spotila. 2001. Effects of maternal identity and incubation
temperature on snapping turtle (Chelydra serpentina) growth. Functional
Ecology 15:624–632.

92
Swingland, I.R., M.J. Coe, D.R. Stoddart, and T.S. Westoll. 1979. The natural regulation
of giant tortoise populations on Aldabra Atoll: recruitment. Philosophical
Transactions of the Royal Society of London. B, Biological Sciences 286:177–
188.
Todd, B.D., B. Halstead, L.P. Chiquoine, J.M. Peaden, K.A. Buhlmann, T.D. Tuberville,
and M.G. Nafus. 2016. Habitat selection by juvenile Mojave desert tortoises. The
Journal of Wildlife Management 80:720–728.
Tomillo, P.S., V.S. Saba, R. Piedra, F.V. Paladino, and J.R. Spotila. 2008. Effects of
illegal harvest of eggs on the population decline of leatherback turtles in Las
Baulas Marine National Park, Costa Rica. Conservation Biology 22:1216–1224.
Tuberville, T.D., K.A. Buhlmann, R. Sollmann, M.G. Nafus, J.M. Peaden, J.A. Daly, and
B.D. Todd. 2011. Effects of short-term, outdoor head-starting on growth and
survival in the Mojave desert tortoise (Gopherus agassizii). Herpetological
Conservation and Biology 14:171–184.
Turner, F.B., P.A. Medica, and C.L. Lyons. 1984. Reproduction and survival of the desert
tortoise (Scaptochelys agassizii) in Ivanpah Valley, California. Copeia 811–820
USFWS 2011. Revised recovery plan for the Mojave population of the desert tortoise
(Gopherus agassizii). U.S. Fish and Wildlife Service, Pacific Southwest Region,
Sacramento, California, USA.
Warkentin, I.G. and N.H. West. 1990. Impact of long-term captivity on basal metabolism
in birds. Comparative Biochemistry and Physiology. A, Comparative
Physiology 96:379–381.

93
Wilbur, H.M., and J.P. Collins. 1973. Ecological aspects of amphibian metamorphosis.
Science 182:1305–1314.
Williams, T.D., Cooch, E.G., Jefferies, R.L., and Cooke, F. 1993. Environmental
degradation, food limitation and reproductive output: juvenile survival in lesser
snow geese. Journal of Animal Ecology 62:766–777.
Wilson, D. S. 1991. Estimates of survival for juvenile gopher tortoises, Gopherus
polyphemus. Journal of Herpetology 25:376–379.
Wilson, D.S., K.A. Nagy, C.R. Tracy, D.J. Morafka, and R.A. Yates. 2001. Water
balance in neonate and juvenile desert tortoises, Gopherus agassizii.
Herpetological Monographs 15:158–170.
Zimmerman, L.C., M.P. O'Connor, S.J. Bulova, J.R. Spotila, S.J. Kemp, and C.J. Salice.
1994. Thermal ecology of desert tortoises in the eastern Mojave Desert: seasonal
patterns of operative and body temperatures, and microhabitat
utilization. Herpetological Monographs 45–59.

94
TABLES
Table 3.1. Twenty-one candidate models to evaluate which predictors (Time in captivity,
MCL, Total displacement, Surface activity, Burrow switching) have the greatest effect on
the post-release fate (1=alive, 0=dead) of head-started Mojave desert tortoises released
into the Mojave National Preserve, San Bernandino, CA, USA. The most parsimonious
model is indicated in bold. Model terms included: TIC (Time in captivity in years; 2,6, or
7), MCL (Midline carapace length in mm at release, September 2018), TDisp (Total
displacement from release burrow until death or the end of the study on 23 July 2019),
SA (surface activity, or the proportion of tracking events that an individual was on the
surface), and Burr (Burrow switching, calculated as the number of unique burrows used
divided by the total number of tracking events found in a burrow). Values presented
include log likelihood (LL), model degrees of freedom (K), Akaike Information Criteria
(AIC), delta AIC (△AIC), and Akaike weights (AICWt), which display the weight of
each model in the candidate set.
Model LL K AIC △AIC AICWt
MCL + SA
MCL
TIC + MCL + SA
MCL + Burr + SA
MCL + TDisp
MCL + Burr
TIC + MCL
TIC + MCL + Burr + SA
MCL + TDisp + Burr + SA
MCL + TDisp + Burr
TIC + MCL + TDisp
TIC + MCL + Burr
TIC + MCL + TDisp + Burr + SA
TIC + MCL + TDisp + Burr
TIC
TIC + TDisp
TIC + Burr
TIC + SA
TIC + TDisp + Burr
TIC + Burr + SA
TIC + TDisp + BuSw + SA
-31.9
-33.6
-31.7
-31.9
-33.2
-33.3
-33.6
-31.7
-31.7
-33.0
-33.0
-33.3
-31.4
-32.8
-35.9
-35.4
-35.5
-35.8
-35.0
-35.4
-34.9
2
1
3
3
2
2
2
4
4
3
3
3
5
4
1
2
2
2
3
3
4
67.9
69.2
69.5
69.9
70.3
70.7
71.1
71.4
71.4
71.9
72.0
72.6
72.8
73.6
73.7
74.9
75.0
75.7
75.9
76.7
77.6
0.0
1.3
1.6
2.0
2.4
2.8
3.2
3.5
3.5
4.0
4.1
4.7
4.9
5.7
5.8
7.0
7.1
7.8
8.0
8.8
9.7
0.25
0.13
0.11
0.09
0.07
0.06
0.05
0.04
0.04
0.03
0.03
0.02
0.02
0.01
0.01
0.008
0.007
0.005
0.004
0.003
0.002

95
Table 3.2. Predicted 10-month survival rates and 95% confidence intervals for head-
started juvenile Mojave desert tortoises (Gopherus agassizii) derived using a generalized
linear model with fate as a binary response variable (0 = “alive”, 1 = “dead”) and MCL,
time in captivity, total displacement, surface activity, and burrow switching as predictor
variables. Time in captivity, total displacement, surface activity, and burrow switching
were kept constant at their overall means when generating survival estimates over the
range of MCL values (60–150 mm). MCL = midline carapace length in mm.
MCL Survival Probability (%) Lower 95% CI Upper 95% CI
60
65
70
75
80
85
90
95
100
105
110
115
120
125
130
135
140
145
150
64.6
67.2
69.6
72.0
74.2
76.3
78.3
80.2
81.9
83.5
85.0
86.3
87.5
88.6
89.7
90.6
91.4
92.1
92.8
41.9
46.7
51.4
55.8
59.9
63.5
66.6
69.2
71.2
72.8
74.0
75.0
75.7
76.3
76.8
77.2
77.5
77.8
78.0
82.7
83.1
83.6
84.2
84.9
85.8
86.8
88.0
89.3
90.6
91.8
93.0
94.1
95.1
95.9
96.7
97.3
97.8
98.2

96
FIGURES
Figure 3.1. Map of release site with 78 release centers for juvenile Mojave desert
tortoises (Gopherus agassizii) in the Mojave National Preserve, San Bernardino, CA. All
78 tortoises were released on 25 September 2018. Animals from three different cohorts
(2011, 2012, 2016) were released.

97
Figure 3.2. Kaplan-Meier survival curve for 78 head-started Mojave desert tortoises
(Gopherus agassizii) released into the Mojave National Preserve, San Bernardino, CA,
USA. Estimates are based on monitoring from 25 September 2018 until 23 July 2019 (43
weeks). Data were right-censored, in that no assumptions were made about the fate of lost
or missing animals (n = 2). Shaded bands are 95% confidence intervals.
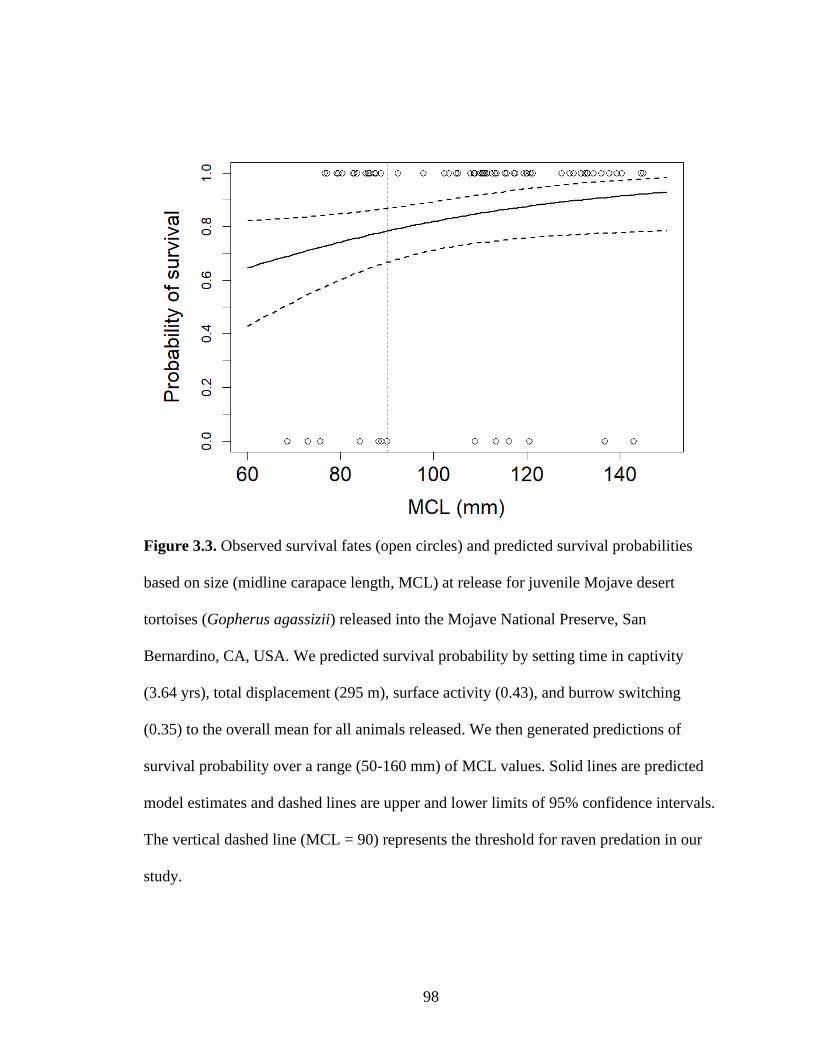
98
Figure 3.3. Observed survival fates (open circles) and predicted survival probabilities
based on size (midline carapace length, MCL) at release for juvenile Mojave desert
tortoises (Gopherus agassizii) released into the Mojave National Preserve, San
Bernardino, CA, USA. We predicted survival probability by setting time in captivity
(3.64 yrs), total displacement (295 m), surface activity (0.43), and burrow switching
(0.35) to the overall mean for all animals released. We then generated predictions of
survival probability over a range (50-160 mm) of MCL values. Solid lines are predicted
model estimates and dashed lines are upper and lower limits of 95% confidence intervals.
The vertical dashed line (MCL = 90) represents the threshold for raven predation in our
study.

99
CHAPTER 4
SUMMARY AND CONCLUSIONS
Head-starting has been evaluated by desert tortoise researchers for nearly three decades,
resulting in continued and substantial increases in knowledge. Outdoor predator-proof
enclosures were the first test of the feasibility of rearing the desert tortoise (Morafka et al.
1997), followed by the addition of supplemental water and food to encourage growth and
boost survival in the enclosures (Nagy et al. 2015, Germano et al. 2017, Tuberville et al.
2019). Growth was still slow however, which led Daly et al. (2018) to incorporate the use
of indoor rearing to bypass winter dormancy and greatly enhance the growth advantage
of being reared under captive conditions. These previous studies formed the foundation
for the continued improvement of this technique.
This thesis evaluates the potential role of combination rearing, a novel method of
indoor rearing coupled with outdoor rearing, in the short-term recovery of the Mojave
desert tortoise. To maximize the ability of head-starting to make an impact on reversing
population declines, programs must evaluate all aspects of their protocols, leading to
continued improvement and increased efficiency. Head-starting alone will not be enough
to undo the range-wide declines sustained by this species, however, as part of a holistic
conservation program (Klemens 2000), head-starting can play a vital role in expediting
population stability (Crawford et al. 2014, Spencer et al. 2017).
In Chapter 2, we experimentally evaluated the pre- and post-release effects of
combination (indoor/outdoor) rearing as compared to the conventional method of strictly
outdoor rearing. We found that two years of combination rearing resulted in juveniles

100
equivalent in size to an 8-year-old wild tortoise with the shell-hardness of an 11-year-old
wild individual (Nagy et al. 2011). For rearing conducted strictly outdoors, we confirmed
that the continued use of supplemental feeding to expedite outdoor head-start growth is
warranted and recommended during this phase. Beyond increased size and shell-
hardness, combination reared juveniles exhibited minimal dispersal tendencies and,
following release, had one of the highest annual survival rates documented in head-
started desert tortoises. All in all, combination rearing was found to be an efficient and
successful technique for both maximizing growth during captivity and subsequent
survival during the first-year post-release. However, only continued monitoring of
released head-starts, including to maturity, will provide the ultimate answers on the long-
term viability of this technique (Lindenmayer et al. 2012, Bertolero et al. 2018). In
addition to long-term monitoring, we also recommend the implementation of
combination rearing and post-release studies in other parts of the desert tortoise’s range
to test the replicability of our findings throughout the diverse habitats inhabited by this
species (USFWS 2011).
In Chapter 3, we sought to test the effect of several predictor variables on post-
release fate of head-started Mojave desert tortoises. We released juveniles ranging in size
from 68-145 mm midline carapace length (MCL) and reared under variable husbandry
protocols. Among our potential predictors (size, time in captivity, total displacement,
surface activity, and burrow switching), only size at release had a significant effect on the
survivorship of released head-starts. This finding, which suggests that two-year-old
combination reared individuals have the same survival probability as 6-7-year-old
outdoor reared tortoises, has the potential to markedly increase the utility of head-

101
starting. Even without considering the financial savings associated with expediting the
head-starting process by 4-5 years, this reduction in the time that it takes to rear robust
head-starts can increase the capacity of current head-starting programs, thereby greatly
accelerating the contribution of head-starting to the recovery of the desert tortoise. Most
importantly, by increasing the efficiency with which funds are used in head-starting,
programs will be able to devote resources to ensuring that the original causes of decline
are also ameliorated (Frazer et al. 1992).

102
LITERATURE CITED
Bertolero, A., J.Ll. Pretus, and D. Oro. 2018. The importance of including survival
release costs when assessing viability in reptile translocations. Biological
Conservation 217:311–320.
Crawford, B.A., J.C. Maerz, N.P. Nibbelink, K.A. Buhlmann, and T.M. Norton. 2014.
Estimating the consequences of multiple threats and management strategies for
semi-aquatic turtles. Journal of Applied Ecology 51:359–366.
Daly, J.A., K.A. Buhlmann, B.D. Todd, C.T. Moore, J.M. Peaden, and T.D. Tuberville.
2018. Comparing growth and body condition of indoor-reared, outdoor-reared,
and direct-released juvenile Mojave Desert Tortoises. Herpetological
Conservation and Biology 13:622–633.
Germano, J.M., M.G. Nafus, J.A. Perry, D.B. Hall, and R.R. Swaisgood. 2017. Predicting
translocation outcomes with personality for desert tortoises. Behavioral
Ecology 28:1075–1084.
Klemens, M.W. 2000. Turtle Conservation. Edited by M.W. Klemens. Smithsonian
Institution Press. Washington D.C., USA.
Lindenmayer, D.B., G.E. Likens, A. Andersen, D. Bowman, C.M. Bull, E. Burns, C.R.
Dickman, A.A. Hoffmann, D.A. Keith, M.J. Liddell, and A.J. Lowe. 2012. Value
of long‐term ecological studies. Austral Ecology 37:745–757.
Morafka, D.J., K.H. Berry, and E.K. Spangenberg. 1997. Predator-proof field enclosures
for enhancing hatchling success and survivorship of juvenile tortoises: a critical
evaluation. Conservation, Restoration, and Management of Tortoises and Turtles,
An International Conference. 147–165.

103
Nagy, K., M. Tuma, and L. Hillard. 2011. Shell hardness measurements in juvenile desert
tortoises Gopherus agassizii. Herpetological Review 42:191–195.
Nagy, K.A., S. Hilliard, S. Dickson, and D.J. Morafka. 2015a. Effects of artificial rain on
survivorship, body condition, and growth of head-started desert tortoises
(Gopherus agassizii) released to the open desert. Herpetological Conservation and
Biology 10:535–549.
Spencer, R. J., J.U. Dyke, and M.B. Thompson. 2017. Critically evaluating best
management practices for preventing freshwater turtle extinctions. Conservation
Biology 31:1340–1349.
Tuberville, T.D., K.A. Buhlmann, R. Sollmann, M.G. Nafus, J.M. Peaden, J.A. Daly, and
B.D. Todd. 2019. Effects of short-term head-starting on growth and survival in
the Mojave Desert Tortoise (Gopherus agassizii). Herpetological Conservation
and Biology 14:171–184.
USFWS 2011. Revised recovery plan for the Mojave population of the desert tortoise
(Gopherus agassizii). U.S. Fish and Wildlife Service, Pacific Southwest Region,
Sacramento, California, USA.