gingiva as a source of stem cells with therapeutic potential
TRANSCRIPT

1
1
Gingiva as a Source of Stem Cells with Therapeutic Potential
Fournier BPJ1,2,3
, Larjava H1, Häkkinen L
1.
1Department of Oral Biological and Medical Sciences, Faculty of Dentistry, University of British
Columbia, Vancouver, Canada; 2
Paris Diderot University, Dental School, Rotschild Hospital,
AP-HP, Paris, 3UMRS872, Team 5, Molecular Oral Physiopathology, CRC Les Cordeliers, Paris,
75006, INSERM UMRS872, Pierre et Marie Curie University, Paris Descartes University,
France.
Dr. Benjamin Fournier, Paris Diderot University, UMRS872, Team 5, Molecular Oral
Physiopathology, 15 Rue de l'Ecole de Médecine, Paris 75006, France; Email:
[email protected]; Tel: 331-4427-5587; Fax: 331-4427-5591.
Dr. Hannu Larjava, University of British Columbia, Faculty of Dentistry, Department of Oral
Biological and Medical Sciences, 2199 Wesbrook Mall, Vancouver, BC, V6T 1Z3 Canada;
Email: [email protected]; Tel: 604-822-6822; Fax: 604-822-3562
Dr. Lari Häkkinen, University of British Columbia, Faculty of Dentistry, Department of Oral
Biological and Medical Sciences, 2199 Wesbrook Mall, Vancouver, BC, V6T 1Z3 Canada;
Email: [email protected]; Tel: 604-822-0096; Fax: 604-822-3562
Running Title: Therapeutic Potential of Gingival Stem Cells
Corresponding Author: Dr. Lari Häkkinen, University of British Columbia, Faculty of
Dentistry, Department of Oral Biological and Medical Sciences, 2199 Wesbrook Mall,
Vancouver, BC, V6T 1Z3 Canada; Email: [email protected]; Tel: 604-822-0096.
Page 1 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

2
2
Abstract
Postnatal connective tissues contain phenotypically heterogeneous cells populations that include
distinct fibroblast subpopulations, pericytes, myofibroblasts, fibrocytes and tissue-specific
mesenchymal stem cells (MSCs). These cells play key roles in tissue development, maintenance
and repair and contribute to various pathologies. Depending on the origin of tissue, connective
tissue cells, including MSCs, have different phenotypes. Understanding the identity and specific
functions of these distinct tissue-specific cell populations may allow researchers to develop better
treatment modalities for tissue regeneration and find novel approaches to prevent pathological
conditions. Interestingly, MSCs from adult oral mucosal gingiva possess distinct characteristics,
including neural crest origin, multipotent differentiation capacity, fetal-like phenotype and potent
immunomodulatory properties. These characteristics and an easy, relatively noninvasive access to
gingival tissue and fast tissue regeneration after tissue biopsy make gingiva an attractive target
for cell isolation for therapeutic purposes aiming to promote tissue regeneration and fast, scar-
free wound healing. The purpose of this review is to discuss the identity, phenotypical
heterogeneity and function of gingival MSCs and summarize what is currently known about their
properties, role in scar-free healing and their future therapeutic potential.
Page 2 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

3
3
Introduction
Accumulating evidence has shown that the cells that reside in postnatal connective tissue are not
all alike but compose different phenotypic subpopulations with distinct properties and functions.
Among connective tissue cells and within fibroblast cultures established from connective tissues,
several tissue-specific mesenchymal stem cell (MSC) populations have been identified that
represent a distinct cell type from fibroblasts and other connective tissue cells (Figure 1) (1-4).
Depending on the origin of tissue, MSCs have different phenotypes (4-9). Understanding the
identity and specific functions of MSCs from various tissues may allow researches to develop
better treatment modalities for tissue regeneration and find novel approaches to prevent
pathological conditions. Interestingly, human oral mucosal cells have a distinct neural crest origin,
high regeneration potential, share some traits with fetal cells and show potent immunoregulatory
properties suggesting that they may possess specific therapeutic potential. The purpose of this
review is to discuss the identity, phenotype and function of MSCs from oral mucosal gingiva and
specifically summarize what is currently known about their therapeutic potential.
Strategies to isolate and identify MSCs from oral tissues
Practically all postnatal connective tissues, including bone marrow, skin, oral mucosa, dental and
periodontal tissues and various parenchymal tissues, contain stem or progenitor cell populations
(5). Nomenclature to describe these cells has been variable in the literature, but most commonly
they have been termed mesenchymal stromal cells (MSCs) or mesenchymal stem cells (also
MSCs) (5). Recently, a term connective tissue stem cells (CTSs) was also introduced to
specifically describe corresponding postnatal orofacial cells (8). For simplicity, we will use the
term MSCs to describe these cells in this review.
Page 3 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

4
4
By definition, MSCs are described as plastic-adherent fibroblast-like cells that are clonogenic and
have the capacity for self-renewal, i.e. making copies of themselves (daughter cells) that have the
same potential as the parent cell (Figure 1). Furthermore, they express a certain set of cell surface
markers, are multipotent and when appropriately stimulated differentiate into mature type of cells
that make up different connective tissues (2-4). These may include ability for osteo-, chondro-
and adipodifferentiation (2-5, 9). In addition, some MSCs may have a capacity for differentiation
into endothelial-like, muscle (myodifferentiation) or neuronal-like cells (neuronal and glial
differentiation). Conclusive evidence of functional neuronal and glial differentiation is still,
however, lacking in vivo (5, 10).
Adult MSCs can be propagated from tissue biopsies of connective tissue with identical protocols
as has been used to culture fibroblasts as they are both plastic adherent and grow in the same cell
culture medium (Figure 1). Therefore, MSCs are likely present in every standard fibroblast
culture although they appear to encompass only a small subpopulation, often representing less
than 1% of the total cell population. MSCs have also a similar morphology with fibroblasts and
share most of the molecular markers that have been traditionally used to identify fibroblasts
(Figure 1 and Table 1). Therefore, there has been some uncertainty about the relationship and
identity of these two cell types (5, 11, 12). The situation has been complicated further by
discoveries that in general connective tissue cells between different individuals and sexes, in
different connective tissues within the same individual or within the given connective tissue are
not alike but consist of subpopulations of adherent, fibroblast-like cells that show considerable
phenotypic heterogeneity (Figure 2) and share similar growth requirements in vitro (12-14).
Page 4 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

5
5
These fibroblast-like cells can include phenotypically distinct fibroblast subsets, pericytes,
fibrocytes, myofibroblasts and MSCs (Figures 2) (6, 11, 12). In any case, currently used
characterization of MSCs rely on properties present in cultured cells as identification of these
cells in their natural tissue niche is challenging largely due to lack of specific markers.
There are basically two strategies to isolate MSCs from adult tissues for cell culture that have
also been widely used to isolate MSCs from oral tissues, including gingiva. In the first approach,
cells are grown from a tissue biopsy as for establishing cultures of plastic-adherent fibroblast. To
this end, cells are either allowed to grow out of a connective tissue biopsy placed onto a plastic
cell culture dish (explant culture method) or they are first enzymatically released from the
connective tissue before seeding into the culture plate (enzymatic digestion method). From these
cultures, MSC colonies can be isolated or enriched by the limited dilution or colony-forming
unit-fibroblastic cells (CFU-F) techniques or by separating them based on expression of MSC
signature molecules using fluorescence-activated cell sorting (FACS) or other separation
techniques (4, 5). In the limited dilution or CFU-F techniques, cells are seeded in a very low
density to a culture dish allowing colonies to form from a single cell. In the enzymatic digestion
method, cells released from tissue can also be directly seeded at low density to isolate stem cells
without a preceding cell culture step. The largest colonies that develop are then considered
candidate MSC populations as they represent cells with the most growth (i.e. self-renewal)
potential. The colony-forming efficiency varies depending on origin of the cells but remains
usually below 10%, although colony CFU-F efficiency of up to about 23 % has been described
for gingival cells (15). However, not all of these colonies comply with the MSC criteria and,
therefore, only a proportion of the colonies contain in fact MSCs. To verify MSC characteristics
Page 5 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

6
6
of the colonies, presence or absence of certain key MSC molecular markers are screened and
their differentiation potential to various mesenchymal lineages is analyzed (6). When compared
to the parental cultures (i.e. mixed cultures containing mostly fibroblasts), the limited dilution-
derived or CFU-F colonies usually have an increased capacity for multipotent differentiation.
Experiments employing repeated enrichments from existing colonies have provided further
evidence that these populations can contain cells that maintain self-renewal and differentiation
potential over several repeated selections (4, 5). However, it has turned out that not all colonies
(i.e. putative stem cell populations) are alike, thus potentially reflecting phenotypic heterogeneity
in the cells of the parental culture or tissue. Furthermore, cells within the given clonal colony
have different characteristics possible reflecting phenotypic modulation, differentiation of the
cells during the colony formation and/or asymmetric cell division (5, 16). Therefore, cell
populations derived from limited dilution or CFU-F methods are phenotypically heterogeneous
and appear to enrich MSCs, although at a variable extent. However, with the current methods it is
not possible to identify which cells in these populations represent the true stem cells. In fact, it is
unclear whether the stem cell properties are characteristic of a specific “stem cell” per se or if
they are a more general characteristic of the given cell colony containing different cell
subpopulations that include subsets of fibroblasts or other fibroblast-like cells and MSCs. It is
possible that the communication between fibroblast subpopulations and MSCs is important for
the MSC phenotype and this can only be achieved in mixed cell populations. Therefore, the niche
created by the nearby fibroblasts, whether in culture or in tissue, maybe critical to determine the
MSC characteristics, fate and functions (17, 18). This niche may be distinct in different tissues
and cells cultures derived from them ultimately leading to a distinct MSC phenotype.
Page 6 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

7
7
In the second approach that have been used to isolate MSCs from oral and other tissues, MSCs
are sorted from the parental fibroblast cultures or from a cell population enzymatically released
directly from the connective tissue biopsy based on a panel of preselected cell surface markers.
This prospective isolation procedure relies on the utility of the selected markers to accurately
identify MSCs (2-5, 9). The challenge is that most of the markers that have been used are usually
present also in other connective tissue cells, including fibroblasts, pericytes, fibrocytes and
myofibroblasts (Table 1) (5, 9, 19). To at least partially overcome this problem, several different
markers should be analyzed in a single cell at the same time, which is technically challenging.
Furthermore, the presence or absence of the surface markers do not necessarily correlate with the
phenotype of these cells in vivo (9).
Recently, another approach was developed to identify the so-called skin-derived precursor cells
(SKPs) from human and mouse skin. In this approach, cells released from skin connective tissue
by enzymatic digestion are grown in suspension in a defined serum-free medium containing EGF
and FGF-2 that supports neural stem cell growth (20, 21). Unlike in the two approaches described
above where MSCs are grown in the presence of serum as plastic adherent cells, this technique
results to selective clustering of cells to form floating neurospheres. Cells in these neurospheres
express nestin, a neural crest cell marker, and vimentin and fibronectin, fibroblast markers. Cells
from the neurospheres can be subcultured and induced to differentiate into neuronal, glial, and
mesenchymal phenotypes that can include muscle cells, osteoblasts, chondrocytes and adipocytes
(20, 22, 23). The best-characterized origin for SKPs in adult tissues is neural crest-derived dermal
papilla and dermal sheath cells of the hair follicles of facial skin, but they also appear to have a
non-neural crest, extra-follicular niche because they can also be isolated from neonatal foreskin
Page 7 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

8
8
that is exclusively of mesodermal origin (21, 24, 25). It remains to be confirmed, however,
whether the SKP-like cells isolated from foreskin are, in fact, of non-neural crest origin and
identical to neural crest-derived SKPs (24). In any case, lineage-tracing experiments in
genetically engineered mice have also supported findings that SKPs can derive from both neural
crest and non-neural crest origins (24, 26). In the oral cavity, SKP-like cells have been isolated
from the nerve endings in rat hard palatal rugae, human and rat periodontal ligament, dental pulp
and buccal mucosa. Similar to SKPs, they express neural crest markers, form neurospheres and
differentiate efficiently into neuronal-like and mesenchymal phenotypes in culture (27-32). To
date, no information about the presence or absence of similar cells in oral mucosal gingiva has
been reported.
Properties of oral mucosal and dental MSC
Oral mucosal and dental cells originate from the neural crest and have distinct properties
Lineage-tracing and other experiments suggest that most oral mucosal connective tissues and
tooth-associated tissues originate embryonically from the cranial neural crest (24). In contrast, in
many other connective tissues, including trunk skin, cells derive from the mesoderm. In
craniofacial skin, however, the dermal papilla cells of hair follicles are also from the neural crest
while stromal cells originate from the mesoderm (27, 33-36). Likely due to the distinct
developmental origin, cells present in the oral mucosal connective tissue have a distinct gene
expression profile compared to skin cells (36, 37). Therefore, it is possible that the embryonic
origin and patterning may underlie some of the phenotypic differences between oral mucosal and
skin cells contributing to the different functional outcomes, including wound-healing responses
(see below) (Table 2). However, whether dermal papilla cells from craniofacial skin and oral
Page 8 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

9
9
mucosal cells, both likely derived from the neural crest, share similar cell phenotype remains to
be shown.
Oral mucosal cells have also specific functional properties. For instance, heterotopic
recombination studies have shown that oral mucosal connective tissues or cells differently
regulate epithelial phenotype as compared to corresponding skin connective tissue or cells (37-
39). As compared to skin, tissue response to injury is also distinct in oral mucosa (see below). In
addition, resulting from the different developmental origin and distinct epithelial-mesenchymal
interactions during organogenesis, oral mucosa harbors salivary glands and teeth while skin has
hair follicles and sweat glands (40). Hair follicles in skin and teeth in oral cavity are formed by
mechanisms involving similar epithelial-mesenchymal interactions. Therefore, it may not be
surprising that while hair follicles in skin are important niches for adult tissue-specific MSCs also
tissues associated with teeth, including gingiva, periodontal ligament and dental apical papilla,
pulp and follicle contain such stem cells (41, 42). In general, MSCs from oral tissues have similar
multipotent differentiation potential as MSCs isolated from other connective tissues, including
skin and bone marrow (41). However, they also have an increased capacity for neurogenic-like
differentiation (27) and may produce tissues specific for teeth and periodontal tissues when
placed in an extra-oral niche (43, 44). Thus, possibly due to their neural crest origin, oral cells
appear preprogrammed for neurogenic and dental and periodontal tissue formation. Interestingly,
cultured vibrissae dermal papilla-derived MSCs have also neurogenic potential and can undergo
odontogenic differentiation when placed in the microenvironment of a developing tooth (44, 45).
Therefore, also other cells may have similar differentiation capacity when exposed to an
appropriate niche. Similar to oral cells, vibrissae dermal papilla-derived MSCs derive from the
Page 9 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

10
10
neural crest (24, 27). It remains to be shown, however, whether the dual neurogenic-like and
odontogenic differentiation property is restricted only to cells that have the neural crest origin.
Characteristics of oral mucosal and dental MSCs
Using the above mentioned culture methods, MSC-like cells have been isolated in the oral cavity
from periodontal ligament (PDLSCs), periodontal granulation tissue, dental pulp (DPSCs,
DPPSCs and SHEDs), dental follicle (DFPCs), apical papilla of a developing tooth (SCAPs),
buccal oral mucosal lamina propria (OMLP-PCs) and gingiva (GMSCs) (Table 3) (46-76). In
general, these cells express various neural crest markers and are, like many neural crest-derived
stem cells, highly multipotent (44) and may possess capacity for both mesenchymal
(cementogenic, odontogenic, osteogenic, chondrogenic, adipogenic) and neural-like (neuronal
and glial) differentiation in vitro (Table 3) (8, 27, 44, 48, 49, 55). As mentioned above, dental-
derived MSCs can also maintain their dental-specific differentiation potential when placed to a
heterotopic tissue niche in vivo. For instance, when PDLSCs, DPSCs or SHEDs were mixed with
a hydroxyapatite carrier and placed subcutaneously in an immunocompromised mouse, they
formed periodontal ligament and tooth root cementum-like (PDLSCs) or dentin and dental pulp-
like (DPSCs and SHEDs) tissue (43, 50, 52). In addition, in their natural niche (i.e. in a tooth
extraction socket) in minipigs in vitro-generated and appropriately induced constructs of
allogenic or autogenic dental pulp MSCs surrounded by sheets of dental follicle MSCs generated
a functional mineralized, dentin-like tooth root and a periodontal ligament (73). On the other
hand, the non-dental oral mucosal lamina propria progenitor cells (OMLP-PCs) have an excellent
neurogenic potential, in addition to osteogenic and chondrogenic capacity and are strongly
immunosuppressive. This last property depends on a mechanism that is at least in part distinct
Page 10 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

11
11
from other adult MSCs being HLA II-independent and mediated by release of
immunosuppressive mediators by the cells (56) mimicking fetal-derived MSCs (77). This is
interesting keeping in mind that human gingival cells also share properties with fetal skin cells
(see below). The more primitive or fetal-like phenotype of oral cells is further supported by
findings showing that DPPSCs cultured from the pulp of human third molars share characteristics
with pluripotent embryonic and induced pluripotent stem cells (iPSCs), including similar gene
expression profile, ability to form embryonic bodies and differentiation into meso-, endo- and
ectoderm-like layers in vitro and teratoma formation in vivo (78).
Gingiva – a distinct tissue of oral mucosa with a fetal-like fast and scarless wound healing
response
While dental tissues are a promising source of MSCs with neural crest properties, their isolation
requires usually an invasive procedure involving tooth extraction. Therefore, oral mucosal soft
tissues may provide a more attractive and practical source of such MSCs. Although in general
oral tissues share similar developmental origin (27) there are several important structural and
functional differences even between various anatomical locations in the oral mucosa. For
example, while oral cavity is covered with a stratified squamous epithelium, in certain locations
the epithelium is non-keratinized (the so-called oral lining mucosa found in cheeks, vestibules
and floor of the mouth) while in others it is keratinized (the so-called masticatory mucosa of
gingiva and hard palate) or is composed of a specialized gustatory epithelium having areas of
both keratinized and non-keratinized epithelium and specialized taste buds (surface of the tongue)
(79, 80). The oral lining and masticatory mucosa cover approximately 60% and 25% of the total
oral mucosa surface area, respectively (81). Recombination experiments using separated
Page 11 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

12
12
epithelial and connective tissues or fibroblasts and epithelial cells from keratinized and non-
keratinized parts of the mucosa have shown that the epithelial phenotypes are determined by
signals from the underlying connective tissue (82-84). Thus, the connective tissue cells and niche
produced by these cells are functionally distinct also in different areas of the oral mucosa.
Accordingly, the structure of the oral mucosal connective tissue also shows regional variations.
The lining mucosa contains an elastic, loosely organized connective tissue with a submucosa
while the masticatory mucosa of gingiva and hard palate contain a dense connective tissue that
directly attaches to the underlying bone (80).
Among oral mucosal tissues, gingiva has further specific features. It is easily accessible for tissue
biopsy and subsequent wound healing is fast and complete with very little morbidity.
Anatomically, gingiva surrounds the teeth and is directly attached to the underlying bone
(alveolar bone) and to the teeth (Figure 3). Its primary function is to provide a dynamic seal
between the teeth and oral mucosa and participate in immune defense (85). Functionally, gingiva
is highly dynamic characterized by one of the fastest tissue turnover rates in body (80, 86).
Unlike in lining oral mucosa or most parts of the skin, the epithelium covering the gingiva forms
long rete ridges that protrude into the connective tissue. The keratinized gingival epithelium
shows also distinct expression of certain cytokeratins (CK), including CK6/CK16, that are not
normally present in skin or oral lining mucosal epithelium (87-89), but that are strongly induced
in these tissues during wound healing (90).
As implied above, gingival tissues and cells seem to be primed for fast tissue regeneration that
leads to a functional advantage over other adult tissues. For instance, wound healing in the
Page 12 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

13
13
keratinized masticatory mucosa of palate and gingiva results to a significantly reduced clinical
and histological scar formation as compared to similar skin wounds (91, 92). In addition, wound-
healing speed is accelerated in both gingiva and other parts of the oral mucosa compared to skin
(91-96). The fast wound healing in oral mucosa is associated with a mild and short-lived
inflammatory response, including a reduced recruitment of neutrophils, mast cells, macrophages
and T-cells as compared to skin wounds (91-93, 96). Furthermore, the level of
immunomodulatory and anti-fibrotic TGF-3 relative to pro-fibrotic TGF-1 is elevated (97, 98).
Although presence of saliva containing several factors that promote wound closure may in part
contribute to the fast and scarless wound healing response also other mechanisms are likely
involved (99). For instance, mechanical signals transmitted from the ECM to the cells are
powerful modulators of cell function (100). Direct adhesion of gingiva to the underlying bone
and tooth surface, low abundance of elastin and organization of collagen into densely packed
collagen fibers (so-called supra-alveolar fiber apparatus) that are thicker than in the oral lining
mucosa (101-104) results to significantly higher tensile strength and stiffness of the gingiva
compared to the oral lining mucosa (105). Interestingly, mechanosignaling and increased ECM
stiffness promotes cell migration (100, 106) and regulates MSC function and fate (107)
potentially promoting wound healing in gingiva. Both the mild inflammatory response and high
relative expression of TGF-3 in oral mucosal wounds are similar to fetal skin where wounds
close fast and do not form scars (93, 108, 109). Furthermore, distinct composition of gingival
ECM and phenotype of gingival fibroblasts, resembling fetal skin cells (13, 93), may provide a
significant functional advantage in wound healing (see below). Whether the preferential wound
healing response in gingiva depends specifically on increased abundance and/or distinct function
of MSC has not been studied in detail. However, several key features of gingival cells in general
Page 13 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

14
14
may contribute to this property.
Distinct phenotype of gingival cells
Developmental origin
Based on the neural crest origin of other oral and dental tissues (27, 33, 36), it has also been
assumed that gingival connective tissue cells have the same origin, but experimental evidence has
been provided only recently. For instance, subpopulations of human oral mucosal fibroblasts
have been reported to express neural crest markers nestin, III-tubulin and GFAP in culture (74).
Most recently, a study utilizing neural-crest specific Wnt-1 reporter mice showed that most cells
in the mouse gingiva in vivo are of neural crest origin but that a smaller proportion of cells are
likely of mesodermal origin. When gingival cells from these mice were propagated in culture as
plastic-adherent cells, about 90% of MSCs generated from these cultures were positive for the
neural crest reporter gene while the rest were from mesoderm (75). Therefore, it appears that
gingival MSCs are heterogeneous and the specific developmental programs may result to
different properties of the MSC subpopulations.
Extracellular matrix niche
In addition to developmental origin, the local tissue niche that the cells reside in determines their
phenotype and function (110). There is some evidence that gingival fibroblasts and likely also
MSCs secrete and interact with a specific ECM niche. For instance, there are quantitative and
qualitative differences between molecules that compose the fibrillar and non-fibrillar ECM,
including type III collagen, sulphated glycosaminoglycans, heparan, chondroitin, dermatan and
keratan sulphate, hyaluronan, and tenascin-C in adult gingiva and skin (92, 111-113). These
Page 14 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

15
15
differences in the ECM composition are likely due to a distinct phenotype of gingival cells as
cultured gingival fibroblasts produce larger dermatan sulphate proteoglycans and higher levels of
hyaluronan than dermal fibroblasts (93, 114). Furthermore, while elastin is abundantly expressed
by skin fibroblasts, gingival connective tissue shows elastin accumulation only in association
with blood vessels and gingival fibroblasts express very low levels of this molecule in vitro (101-
103). In spite of this, many of the microfibrillar components of elastic fiber system, including
fibulin-5 and fibrillin-1 and -2, are expressed in the gingiva (115-117). Interestingly, while skin
elastic fibers show age related changes, in gingiva they remain morphologically relatively stable
(118). A survey based on analysis of 44 genes by RT-PCR in cultured fibroblasts showed that,
although gingival and skin cells expressed comparable levels of major ECM proteins, including
type I collagen and fibronectin, the expression of certain ECM proteins, such as periostin,
osteopontin and chains for type III and V collagen, was significantly lower in gingival cells. In
addition, expression of several cell adhesion-related molecules and integrin-family ECM
receptors for collagens, laminin, fibronectin and teanscin-C was significantly different from skin
fibroblasts (119-121). Furthermore, gingival cells showed reduced adhesion and spreading on
collagen and fibronectin as compared to skin cells, suggesting that the repertoire and function of
ECM receptors in gingival cells is distinct (120).
The cellular niche contains also growth factors that are bound and stored in the ECM in an
inactive form, including TGF-1, vascular-endothelial growth factor (VEGF) and fibroblast
growth factor-2 (FGF-2) (122). Very little is known about the abundance of these molecules in
gingival MSC niche in vivo or in vitro. However, gingival fibroblasts produce distinct amounts of
ECM molecules, including various proteoglycans, glycosaminoglycans, fibrillin-1 and tenascin-C,
Page 15 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

16
16
that bind growth factors. Furthermore, gingival cells show differences to skin cells with respect to
levels of various growth factors produced (see below). Thus, gingival cells produce a specific
ECM niche that likely harbors distinct levels of growth factors.
In addition to ECM composition, mechanical signals mediated from the cellular
microenvironment to the cells are important modulators of cell phenotype and differentiation fate.
For instance, cells sense the stiffness of the ECM via integrin receptors that impacts their gene
expression and directs their lineage specifications and differentiations (123). Interestingly, elastic
modulus measurements from pig tissues indicate that stiffness in gingiva is higher (elastic
modulus of about 20 kPa) as compared to oral lining mucosa (about 5-10 kPa) or skin (about 5-
10 kPa) (124-125).
Taken together, the gingival ECM niche (composition, organization and mechanical properties)
and cell interactions with it appear distinct and may determine and maintain gingival MSC
phenotype and function (Table 2).
Cell heterogeneity within gingiva
As mentioned above, cells within a given connective tissue in vivo and in vitro, including gingiva,
are phenotypically heterogeneous. In gingiva, part of this may be explained by heterogeneous
neural crest and mesodermal origin of the cells (75), but other mechanisms may also be involved.
Structurally, gingival tissue is not uniform further suggesting that functionally distinct fibroblast
population and possibly MSCs also exist in different anatomical locations. For instance, the
molecular composition of the connective tissue in the interdental papilla (part of the gingiva that
Page 16 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

17
17
resides between teeth) and in the attached gingiva (facial or oral side of the gingiva) is different
(126). Other studies have shown that fibroblasts isolated from the connective tissue papilla of
gingiva (region of connective tissue residing between long epithelia rete ridges typical for gingiva)
show higher saturation density, smaller cell size and higher production of the migration
stimulating factor (MSF) as compared to the cells isolated from the deeper reticular connective
tissue (13). MSF is found only in fetal but not in adult skin fibroblasts (127, 128). While both
adult human gingiva and fetal skin are characterized by scarless wound healing the importance of
MSF in this context and specific molecular characteristics of MSF has not been studied in detail
(93).
In addition to phenotypic differences between different anatomical locations, gingival cells show
functional heterogeneity between individuals (129) and within the given cell culture originating
from the same biopsy (Figure 2). For instance, early studies by Hassell and Stanek (130)
demonstrated that distinct subpopulations of gingival fibroblasts can be isolated from the same
biopsy, in this case from the interdental papilla. These cell subpopulations showed differences in
doubling times, cell size and collagen and glycosaminoglycan synthesis. These properties were
stable during cell culture indicating that they were an inherent property of the cells and not
affected by the cell culture (130). Other studies have shown further phenotypic heterogeneity in
cell morphology, total protein synthesis, production of different collagens (type I, III, V) and
fibronectin, proliferation, response to cytokines, growth factors, C1q component of complement
and PGE2 and in enzymatic activities (131-140). Furthermore, gingival cells can be separated
into different subpopulations based on the expression of stem cell factor (SCF) and its receptor c-
Kit (CD117) (141), although gingival cells have also been reported to be negative for CD117 (49).
Page 17 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

18
18
Sorting of a gingival cell population based on the expression of the high affinity receptor for C1q
yields a subpopulation with short doubling time and an increased protein synthesis, particularly
type III and type V collagen. These properties are stable over various passages in culture and
reminiscent of cells that are activated for a wound healing response (134). Further studies showed
that these cell subpopulations had also a different property to interact with either the C1q
collagen-like or globular domains resulting to differential activation of intracellular signaling
pathways (140). Whether any of the above cell subpopulations present in the cultures or tissues
represent MSCs or if they result from distinct mesodermal or neural crest origin has not been
explored in detail.
Gingival cell heterogeneity and phenotype may also have importance for gingival pathology.
Drug-induced gingival fibromatosis affects about 10 to 50 % of individuals taking systemic
medications, including certain Ca2+
-channel blockers, anticonvulsants and immunosuppressants,
and results to an expansive enlargement of the gingival tissue due to excessive accumulation of
ECM produced by gingival fibroblasts. This condition appears to be limited to the gingiva
suggesting that only gingival cells are susceptible to these drugs (142). Interestingly, cultured
gingival fibroblasts are heterogeneous in their response to these drugs and only some
subpopulations are activated to accumulate excessive ECM. In addition, fibroblasts isolated from
the gingival overgrowth tissue maintain their high ECM producing phenotype in culture,
suggesting that use of the medication has selectively favored the expansion of the responding cell
population in vivo (143-145). The role of MSCs in the pathogenesis of gingival overgrowth has
not been addressed. However, MSCs can be isolated from gingival overgrowth tissue and have
comparable expression of cell surface markers, self-renewal, osteo-, adipo- and chondrogenic
Page 18 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

19
19
differentiation capacity and immunomodulatory properties as MSCs from normal gingiva (146).
Gingival multipotent MSCs
The fetal-like cell phenotype with potency to support tissue regeneration suggests that gingiva
may contain an abundant source of MSC that promote tissue regeneration. Consequently, cells
that comply with the classical MSC definitions have been isolated from human gingiva from
established gingival fibroblast explant cultures (49) as well as from cells released from the tissue
by enzymatic digestion (58, 59, 146, 147). In general, gingival stem cells express cell surface
markers typical to MSCs (Table 1), show self-renewal capacity and variable colony forming
efficiency of about 3-23% in culture. Comparisons of gingival MSCs with corresponding cells
from other sources have shown variable results. In one study, gingival MSCs were similar to
MSCs found in bone marrow (59) while in another study they expressed different cell surface
markers and differentiation capabilities (49). A more consistent finding seems to be that gingival
MSC proliferate faster in culture and maintain their characteristics, karyotype and telomerase
activity better after higher passages than bone marrow MSCs (59, 60, 146).
Perivascular tissue – a niche for gingival MSCs?
In general, the tissue niche for MSCs has remained undefined but it seems likely that this niche is
different in various tissues. Maybe because of this difference, MSCs from various origins also
possess distinct phenotypes (5). The tissue location of the gingival MSCs is elusive.
Immunostaining of cells expressing the classical MSC/embryonic stem cell markers STRO-1,
OCT-4 and SSEA-4 have indicated presence of a low number of single or double
immunopositive cells in subepithelial human gingival connective tissue (49, 59, 146). In addition,
Page 19 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

20
20
undefined chord-like structures in the subepithelial gingival connective tissue show positive
staining for p75NTR, a neural crest cell marker, along with positive staining for the embryonic
stem cell markers OCT4 and SOX-2 (74). However, as the cell surface markers used to identify
MSCs are not specific for these cells the exact tissue niche for putative MSCs still remains to be
clarified.
Certain populations of MSCs have been located previously in perivascular tissue in many tissues.
Therefore, it is likely that abundant vascular network may also provide such a MSC niche in
gingiva. In addition to the tooth and hair follicle-related niches described above, earlier studies
had suggested that some of the progenitor cells involved in wound healing and tissue
regeneration in various tissues, including skin, periodontal ligament and gingiva, reside at the
perivascular location (100, 148-151). This has been subsequently confirmed in several reports
showing that perivascular cells resembling “undifferentiated fibroblasts” can migrate into the
wound and produce collagen to repair the wound (99, 152-154). More recent findings from
immunolocalization and cell culture studies have also provided evidence that some cells that
reside in perivascular tissues have characteristics of MSCs (150). Association with vasculature
would provide an ideal tissue niche for MSCs as blood vessels form a vast network expanding
throughout the entire body allowing fast access for progenitor cells into various tissue sites and
into the circulation. Perivascular location can harbor various cells, including circulating cells
passing through the endothelium into the connective tissue, fibroblasts, vascular smooth muscle
cells and pericytes. In particular pericytes, which surround endothelial cells in capillaries and
microvessels throughout the body, possesses MSC properties. These cells can be identified by
expression of NG2, CD146, -SMA and PDGFRβ (151). Cultured pericytes share, however,
Page 20 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

21
21
several key properties with fibroblasts, including similar cell morphology, expression of several
of the cell markers (Table 1) and like fibroblasts they can be propagated as plastic adherent cells
with standard fibroblast growth medium. Pericytes have also similar properties with MSCs as
they express many of the same surface markers (Table 1) and are able to differentiate into
myofibers, adipocytes, chondroblasts and osteoblasts (151, 155, 156). Thus, pericytes maybe
present in fibroblast and/or MSC cultures. Whether pericytes can be considered as MSCs and
whether some of the MSC properties of fibroblast cultures or MSCs purified from them can be
attributed to pericytes needs further experimental clarification. In any case, perivascular tissue
appears to provide a distinct niche for MSCs or MSC-like cells throughout the body, including
periodontal tissues (155).
Therapeutic potential of gingival MSCs
Widely accepted key properties of MSCs to promote tissue regeneration include multipotent
differentiation, immunomodulation and stimulation of vasculogenesis and epithelialization (157).
While differentiation of MSCs into cells of the target tissue may occur in vivo, studies have
indicated that only very low numbers survive and engraft in the tissue for long term. Therefore, it
is likely that paracrine factors released by these cells play a key role in their potency to promote
regeneration by the host cells (69). In the following paragraphs we will review the regenerative
potential of gingival cells based on evidence about the above properties.
Differentiation capacity
Similar to other MSCs, gingival MSCs selected by CFU-F or limited dilution techniques also
display the classical mesodermal tri-lineage osteo-, adipo- and chondrogenic differentiation in
Page 21 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

22
22
vitro (15, 49, 59, 146). In addition, differentiation of gingival MSCs into endodermal-like and
neural-like cells has been described but these findings need further verification (Table 3) (59, 74,
75). Interestingly, when the two gingival MSC populations derived from the neural crest and
mesoderm were compared, the neural crest-derived cells showed a better potential for
chondrogenic differentiation and expressed higher levels of neural/neural-crest markers –tubulin,
nestin and neurofilament M under neurogenic differentiation conditions in vitro (75). When
compared to periodontal ligament MSCs, gingival cells show significantly reduced doubling time
and better cloning efficiency, but periodontal ligament MSCs appear to have better differentiation
potential (15).
Studies of differentiation of gingival MSCs in vivo have also shown promising but variable
results. In general, when gingival MSCs are implanted subcutaneously, without prior incubation
in any differentiation-inducing medium, they produce connective tissue reminiscent of gingiva
(158). When cultured in an osteogenic induction medium and incorporated into a
hydroxyapatite/tricalcium phosphate carrier, fibrin gel, or other carriers and then transplanted
subcutaneously into immunocompromised mice, gingival MSCs have been reported to either
produce (15) or not to produce (49, 59) mineralized tissue. When gingival MSCs grown inside a
collagen carrier were treated as above and transplanted into critical size bone defects created in
rat mandibles or calvaria they regenerated the bone defects (Table 3) (58). In a recent study,
STRO-1+ immunoselected gingival MSCs in deproteinized bovine cancelleous bone or collagen
carrier was applied into experimental periodontal defects in mini pigs. Results showed that MSCs
containing scaffolds induced regeneration of periodontal tissues including bone, cementum and
periodontal ligament (159. However, recent findings have shown that inflammatory environment
Page 22 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

23
23
in vivo or treatment with inflammatory cytokines in vitro may suppress the differentiation
capacity of gingival MSCs (15, 158, 160).
Interestingly, similar to MSC, also unselected gingival fibroblast cultures can be induced to
differentiate into osteo-, adipo- and chondrogenic lineages as well as to myofibroblasts in vitro
(60, 72, 74). In addition, differentiation into an endodermal and neurogenic pathway has been
described but needs further confirmation (74) (Table 3). It is somewhat surprising that non-
selected gingival fibroblast cultures may have similar properties as MSCs. There is some
evidence, however, that unselected gingival fibroblast cultures may loose their differentiation
potential during successive passaging in culture faster than gingival MSCs generated by CFU-F
method (49). This preliminary finding needs further verification but implies that the proportion of
MSCs in the fibroblast cultures decreases over time. Therefore, in addition to gingival MSCs,
non-selected gingival fibroblast cultures may possess potential for therapeutic applications, but
they may need to be used as primary cultures or in their early passages. Interestingly, in a recent
study, human gingival or lining mucosal cell cultures were established using a fibroblast explant
culture technique (76). Although no clonal selection was performed, about 90% of the cells
expressed typical MSC markers and about 40-70 % pluripotency genes Oct4, Sox2 and Nanog. In
addition, when subjected to appropriate induction media cultures showed osteogenic, adipogenic
and chondrogenic differentiation in vitro. Cells were then cultured in fibrin gels and transplanted
between skin and calvaria in immunocompromized mice which resulted to mineralized tissue
formation that contained proteins typical to bone, cementum and dentin. Thus, cultures generated
from gingiva and oral lining mucosa using typical fibroblast culture protocols maybe enriched
with MSC-like cells providing an attractive and practical source of cells for therapeutic
Page 23 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

24
24
applications.
Immunomodulation
In the oral cavity, gingiva protects underlying tissues from effects of constant build-up of
microbial biofilm at the tooth-gingiva interface. Therefore, gingival cells are adapted to the first
line of defense against the biofilm and interact with microbes, microbial-derived molecules and
immune cells. Gingival stromal cells communicate with immune cells directly by establishing
cell-cell contacts or indirectly by secreting and responding to inflammatory mediators from
immune cells (157, 161). Like other MSCs, gingival MSCs have potent immunosuppressive
properties that may contribute to their regenerative and other functions (75, 162). For instance,
when injected into circulation they suppress development of allogeneic rejection, contact
hypersensitivity, experimental colitis and collagen-induced arthritis in mouse models (75, 146,
162, 163). Cell culture findings have indicated that similar to bone marrow MSCs, gingival cells
can suppress T-cell proliferation and activation, and modulate function of innate immune cells,
including dendritic cells, macrophages and mast cells. Important for tissue regeneration, gingival
cells are able to induce repolarization of inflammatory M1 macrophages to anti-inflammatory,
reparative M2 macrophages (75, 146, 164, 165). Interestingly, the neural crest-derived gingival
MSC population appears to have better immunosuppressive properties than the mesodermal-
derived subpopulation as the former were more potent to induce apoptosis of activated T-cell in
vitro and to ameliorate experimental inflammatory colitis in a mouse model in vivo (75). These
anti-inflammatory and immunomodulatory functions can likely be attributed to cellular cross-talk
of gingival cells with immune cells and release of soluble factors, including indoleamine 2.3-
dioxygenase (IDO), IL-10 and PGE2, which is similar to MSCs from different tissues (162, 165).
Page 24 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

25
25
In addition to studies about immunomodulatory effects of gingival MSC, there is vast literature
about immunoregulation by non-selected gingival fibroblast populations that may also be
relevant in general to the utility of gingival cells in therapy. Most of this research has focused on
immunomodulatory effects of gingival cells on immune defense against acute or chronic
microbial infection and have provided evidence that gingival cells in general, like gingival MSC,
have immunosuppressive properties in vitro as they reduce peripheral blood monocyte
proliferation (72, 166, 167). In gingival fibroblasts, also similar to MSCs, IFN-stimulates type
II HLA expression but the cells are not able to stimulate alloreactive T-cells (3, 168-170).
Subsequent studies have shown that IFN- induces secretion of IDO in gingival fibroblasts
similar to MSCs and that this is at least in part responsible for suppressing T cell proliferation
(171, 172). In this context it is interesting to note that transplanted gingival fibroblasts have a
better potential than skin fibroblasts to down regulate inflammation in vivo (173, 174).
Furthermore, oral mucosal wounds show a milder and shorter inflammatory reaction in response
to wounding as compared to skin wounds (91, 92, 96). This may depends, at least partially, on
immunomodulatory functions of gingival cells as it is well established that MSCs and fibroblasts
take part in regulation of inflammation during wound healing (100). This is further supported by
findings from a mouse model where systemic infusion of gingival cells resulted to accelerated
skin wound repair with rapid re-epithelialization and angiogenesis. This effect was associated
with a local inflammatory response at the wound site (164). However, it was not completely clear
whether these effects were achieved with clonally selected MSC populations or by non-selected
parental cultures (i.e. composed mostly of fibroblasts).
Page 25 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

26
26
Stimulation of angiogenesis and epithelialization
Role of gingival cells in promoting angiogenesis has not been investigated extensively. However,
autologous gingival connective tissue grafts and free gingival grafts (composed of both gingival
epithelium and connective tissue) have been used extensively in periodontal therapy and they
revascularize readily in vivo as determined by histological methods (175-177). In addition,
gingival cell cultures are able to produce pro-angiogenic factors, including VEGF, CXCL12, IL8,
MCP-1 and CXCR1 and promote endothelial cell growth in vitro (178-180). However, whether
these properties depend on fibroblasts or MSCs present in the mixed populations of cells in these
experiments needs further clarification. In addition, whether these cells have comparable ability
to promote also vasculogenesis in regenerative therapy as has been reported for other MSCs (16)
remains to be shown.
As discussed above, gingival connective tissue or cells define the epithelial phenotype in the oral
cavity. This has been evidenced by grafting procedures where gingival connective tissue induces
formation of a keratinized epithelium that corresponds to the one that is present in the normal
gingiva (82). In addition, gingival connective tissue cells appear to have a distinct property to
regulate epithelialization also in extra-oral sites. For instance, bioengineered tracheal constructs
containing autologous gingival fibroblasts induced a functional tracheal epithelial regeneration in
a rat model (181, 182). This property was similar to adipose-derived MSCs but differed from skin
or nasal fibroblasts that were not able to induce an appropriate morphological and functional
epithelial phenotype (182). Whether these properties depend on the fibroblasts and/or MSCs
needs further clarification.
Page 26 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

27
27
Use of gingival cells in future cell therapy
Use of gingival cells to promote tissue regeneration
While stem cell therapies are still largely at an experimental stage, particularly skin fibroblasts in
various applications, including living skin tissue grafts or manufactured skin substitutes, have
been used to treat dermal deficiencies and to promote skin wound healing for decades. In general,
these modalities have improved treatment outcomes and benefited the patients by improving their
quality of life, but they have not been able to consistently regenerate the skin architecture and
function (183-186). However, animal studies have indicated that use of MSCs from various
sources, including bone marrow, adipose tissue or skin, have a better potential in skin and
possibly also in other tissues (187). Most recent findings have indicated that novel technologies,
including injection of allogeneic fibroblasts or MSCs or use of iPSCs generated from skin cells,
may also be beneficial to treat certain conditions (188, 189).
As discussed above, gingival cell cultures possess abundantly MSCs with distinct capacity for
multilineage differentiation and that promote re-epithelialization, support angiogenesis and
modulate inflammation. To what extent this property depends on presence of distinct MSC or
other cell subpopulations in the gingival tissues or cell cultures, developmental origin, specific
phenotype of the cells and tissue niche that they create or other factors remains to be shown. In
any case, due to their unique properties discussed above and easy access, gingival cells could
have certain advantages over skin cells and be considered for therapeutic applications also in
various areas of the body other than the oral cavity. Particularly, these would include conditions
that require connective tissue regeneration, healing without scarring or mucosal type epithelial
keratinization.
Page 27 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

28
28
Information about utility of gingival MSCs for tissue regeneration or clinical use of gingival cells
in general in tissue engineering in other anatomical locations than the oral cavity is currently
scarce. However, full thickness oral soft tissue autografts that contain, among other cells, viable
fibroblasts and likely MSCs, have been successfully used for decades. In particular, human
palatal tissue autografts are routinely used as free gingival (containing both gingival epithelium
and connective tissue) or connective tissue grafts to augment gingival tissue deficiencies. Both
graft types induce epithelial differentiation typical to the donor tissue (i.e. keratinized stratified
squamous epithelium) (82, 83). In addition, gingival fibroblasts have been recently applied in
vascular therapy in an animal model of a carotid aneurysm (173). The results showed that,
remarkably, grafting gingival fibroblasts into the arterial walls caused reduction of the lesions
while those treated with medium control or dermal fibroblasts continued to grow. Gingival
fibroblasts also supported the elastin repair in the arterial wall (173). Potential use of gingival
cells to improve wound-healing outcome in situations that have been traditionally treated with
skin cell-derived grafts, including burn wound and other skin trauma, is another attractive future
application. Wound healing of large skin defects, even with the best current therapeutic
modalities, usually results to excessive scar formation (190). Scar-free wound healing in gingiva
suggests that grafts of gingival cells maybe used to reduce scar-forming response in these
situations. Furthermore, ability of gingival cells to promote fast wound healing maybe utilized to
treat chronic non-healing skin wounds, a major clinical problem (191).
Gingiva as a source for induced pluripotent stem cells
Page 28 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

29
29
A novel therapeutic applications for gingival cells could be their use to generate iPSCs. iPSCs
were initially produced from non-pluripotent somatic cells by forced expression of four genes,
Oct4, Sox2, Klf4 and c-Myc by using viral vectors (192). More recently it was shown that iPSCs
can also be generated using corresponding recombinant proteins or by combined expression of
Oct4, Sox2 and Klf4 only or by a combination of Oct4, Sox2, Nanog and Lin28 (193, 194).
iPSCs share key properties, including gene expression, chromatin methylation, embryonic body
and teratoma formation, viable chimera formation after injection into blastocyst and ecto-, meso-
and endodermal differentiation potency, with embryonic stem cells (192). Interestingly, it appears
that iPSC-like cells can also be generated from human skin fibroblasts by overexpression of a
single gene encoding an ECM protein fibromodulin (195), but the mechanisms involved and
potential therapeutic potential of this finding remains to be shown and confirmed.
While skin fibroblasts from various animals and humans have been the most common source to
generate iPSCs, theoretically, they can be established from any somatic cell type. Accordingly,
they have also been produced from various mesenchymal, epithelial and blood-derived cells from
different animals or humans, but in general the generation efficiency is low (often less than 1%)
and varies greatly depending on the cell type (193, 196). Recently, iPSCs were produced from
cells of oral origin, namely from human dental pulp cells, buccal mucosa and periodontal
ligament fibroblasts (197-200). Recently, iPSCs were also produced and characterized from
human and mouse gingival fibroblasts (200, 201). Interestingly, generation of iPSCs was seven
times more efficient from mouse gingival fibroblasts as compared to fibroblasts obtained from
tail skin (201). Generation of patient-specific and disease-specific gingiva-derived iPSCs would
provide tools for disease modeling and drug screening in conditions related to oral cavity. These
Page 29 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

30
30
could include syndromes involving oral or dental tissues, conditions involving congenital dental
and/or oral malformations, hereditary gingival overgrowth or certain forms of periodontal disease
(202). Although promising, iPSC-based therapies are still in their initial stages of development
and several technical issues, including further improving efficacy of iPSC generation,
understanding the properties of iPSCs and safety concerns, such as tendency of iPSCs to form
tumors and adverse effects of the viral vectors, need to be addressed before iPSCs from human
gingiva or from any other source can be considered for human therapy (193, 194).
Due to their neural-crest origin, gingival cells may be suitable for the so-called direct somatic cell
fate reprogramming (induced transdifferentiation) where one differentiated cell type is converted
to another distinct cell type without a reversion to iPSCs or use of oncogenes. Recently, this
technique has been used to generate functional induced neurons (iNs) and induced neural stem
cells (iNSCs) from various cells (203), but there is no experimental evidence about the efficiency
of gingival cells in this application.
Conclusions and future directions
Connective tissue cells form a phenotypically heterogeneous group of cells that play key roles in
tissue development, maintenance and repair and contribute to various pathologies. These cells
can be relatively easily isolated and expanded in cell culture. Depending on tissue origin,
connective tissue cell cultures contain cells, most notably MSCs, that possess distinct ability for
multipotent differentiation and immunomodulation and promote vasculogenesis/angiogenesis and
epithelialization (Table 4). Identification of distinct phenotypic properties of these cells from
different tissues may provide novel opportunities to selectively use different phenotypic traits for
Page 30 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

31
31
various tissue regeneration modalities. Adult oral mucosal connective tissue cells, and in
particular MSCs, can be easily harvested with little morbidity and possess distinct characteristics,
including neural crest origin, multipotent differentiation capacity, fetal-like phenotype and potent
immunomodulatory properties, that may be utilized to promote tissue regeneration and fast, scar-
free wound healing. Future studies should be focused on understanding the phenotypic
differences among connective tissue cell populations and MSCs, to understand the specific
properties of neural crest-derived cells, to develop better molecular markers to identify and enrich
appropriate cell populations and in detail characterize their properties important for tissue
regeneration applications in vivo. In particular, studies systematically comparing gingival cells to
corresponding cells from other tissues and sources should be conducted to further verify the
distinct potential of gingival cells in vitro and in vivo. Furthermore, developing efficient cell
delivery modalities that accurately target the cells to the area of interest and provide an
appropriate cellular niche important for their engraftment and function need further development.
Finally, assessing putative risks and adverse effects of cell-based therapies will be critical.
Acknowledgements: We thank Dr. L. Loison-Robert for technical assistance. Supported by
Canadian Institutes of Health Research.
Author Disclosure Statement: Drs. B. Fournier, H. Larjava and L. Häkkinen have no competing
financial interests.
Page 31 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

32
32
References
1. Robert L. (1992). The fibroblast, definition of its phenotype by its "programme" of
biosynthesis of the extracellular matrix. Pathol Biol (Paris) 40:851-858.
2. Keating A. (2012). Mesenchymal stromal cells: new directions. Cell Stem Cell 14:709-
716.
3. Dominici M, K Le Blanc, I Mueller, I Slaper-Cortenbach, F Marini, D Krause, R Deans,
A Keating, D Prockop and E Horwitz. (2006). Minimal criteria for defining multipotent
mesenchymal stromal cells. The International Society for Cellular Therapy position statement.
Cytotherapy 8:315-317.
4. Bianco P, X Cao, PS Frenette, JJ Mao, PG Robey, PJ Simmons and CY Wang. (2013).
The meaning, the sense and the significance: translating the science of mesenchymal stem cells
into medicine. Nat Med 19:35-42.
5. Augello A, TB Kurth and C De Bari. (2010). Mesenchymal stem cells: a perspective from
in vitro cultures to in vivo migration and niches. Eur Cell Mater 20:121-133.
6. Pevsner-Fischer M, S Levin and D Zipori. (2011). The origins of mesenchymal stromal
cell heterogeneity. Stem Cell Rev 7:560-568.
7. Bieback K, P Wuchter, D Besser, W Franke, M Becker, M Ott, M Pacher, N Ma, C
Stamm, H Kluter, A Muller and AD Ho. (2012). Mesenchymal stromal cells (MSCs): science and
f(r)iction. J Mol Med 90:773-782.
8. Mao JJ and DJ Prockop. (2012). Stem cells in the face: tooth regeneration and beyond.
Cell Stem Cell 11:291-301.
9. Hematti P. (2012). Mesenchymal stromal cells and fibroblasts: a case of mistaken identity?
Page 32 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

33
33
Cytotherapy 14:516-521.
10. Chen L, DM He and Y Zhang. (2009). The differentiation of human placenta-derived
mesenchymal stem cells into dopaminergic cells in vitro. Cell Mol Biol Lett 14:528-536.
11. Sorrell JM and AI Caplan. (2004). Fibroblast heterogeneity: more than skin deep. J Cell
Sci 117:667-675.
12. Haniffa MA, MP Collin, CD Buckley and F Dazzi. (2009). Mesenchymal stem cells: the
fibroblasts' new clothes? Haematologica 94:258-263.
13. Irwin CR, M Picardo, I Ellis, P Sloan, A Grey, M McGurk and SL Schor. (1994). Inter-
and intra-site heterogeneity in the expression of fetal-like phenotypic characteristics by gingival
fibroblasts: potential significance for wound healing. J Cell Sci 107 (Pt 5):1333-1346.
14. Phipps RP, MA Borrello and TM Blieden. (1997). Fibroblast heterogeneity in the
periodontium and other tissues. J Periodontal Res 32:159-165.
15. Yang H, LN Gao, Y An, CH Hu, F Jin, J Zhou, Y Jin Y and FM Chen. (2013).
Comparison of mesenchymal stem cells derived from gingival tissue and periodontal ligament in
different incubation conditions. Biomaterials 34(29):7033-7047.
16. Phinney DG and DJ Prockop. (2007). Concise review: mesenchymal stem/multipotent
stromal cells: the state of transdifferentiation and modes of tissue repair--current views. Stem
Cells 25:2896-2902.
17. Kuhn NZ and RS Tuan. (2010). Regulation of stemness and stem cell niche of
mesenchymal stem cells: implications in tumorigenesis and metastasis. J Cell Physiol 222:268-
277.
18. Lu W, MO Casanueva, AP Mahowald, M Kato, D Lauterbach and EL Ferguson. (2012).
Niche-associated activation of rac promotes the asymmetric division of Drosophila female
Page 33 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

34
34
germline stem cells. PLoS Biol 10:e1001357.
19. Alt E, Y Yan, S Gehmert, YH Song, A Altman, S Gehmert, D Vykoukal and X Bai.
(2011). Fibroblasts share mesenchymal phenotypes with stem cells, but lack their differentiation
and colony-forming potential. Biol Cell 103:197-208.
20. Toma JG, M Akhavan, KJ Fernandes, F Barnabe-Heider, A Sadikot, DR Kaplan and FD
Miller. (2001). Isolation of multipotent adult stem cells from the dermis of mammalian skin. Nat
Cell Biol 3:778-784.
21. Toma JG, IA McKenzie, D Bagli and FD Miller. (2005). Isolation and characterization of
multipotent skin-derived precursors from human skin. Stem Cells 23:727-737.
22. Hunt DP, C Jahoda and S Chandran. (2009). Multipotent skin-derived precursors: from
biology to clinical translation. Curr Opin Biotechnol 20:522-530.
23. Kang HK, SK Min, SY Jung, K Jung, H Jang da, OB Kim, GS Chun, ZH Lee and BM
Min. (2011). The potential of mouse skin-derived precursors to differentiate into mesenchymal
and neural lineages and their application to osteogenic induction in vivo. Int J Mol Med 28:1001-
1011.
24. Achilleos A and PA Trainor. (2012). Neural crest stem cells: discovery, properties and
potential for therapy. Cell Res 22:288-304.
25. Uchugonova A, J Duong, N Zhang, K Konig and RM Hoffman. (2011). The bulge area is
the origin of nestin-expressing pluripotent stem cells of the hair follicle. J Cell Biochem
112:2046-2050.
26. Jinno H, O Morozova, KL Jones, JA Biernaskie, M Paris, R Hosokawa, MA Rudnicki, Y
Chai, F Rossi, MA Marra and FD Miller. (2010). Convergent genesis of an adult neural crest-like
dermal stem cell from distinct developmental origins. Stem Cells 28:2027-2040.
Page 34 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

35
35
27. Kaltschmidt B, C Kaltschmidt and D Widera. (2011). Adult Craniofacial Stem Cells:
Sources and Relation to the Neural Crest. Stem Cell Rev 8:658-671.
28. Arnold WH, S Becher, A Dannan, D Widera, T Dittmar, M Jacob, HG Mannherz, T
Dittmar, B Kaltschmidt, C Kaltschmidt and WD Grimm. (2010). Morphological characterization
of periodontium-derived human stem cells. Ann Anat 192:215-219.
29. Sasaki R, S Aoki, M Yamato, H Uchiyama, K Wada, T Okano and H Ogiuchi. (2008).
Neurosphere generation from dental pulp of adult rat incisor. Eur J Neurosci 27:538-548.
30. Techawattanawisal W, K Nakahama, M Komaki, M Abe, Y Takagi and I Morita. (2007).
Isolation of multipotent stem cells from adult rat periodontal ligament by neurosphere-forming
culture system. Biochem Biophys Res Commun 357:917-923.
31. Widera D, WD Grimm, JM Moebius, I Mikenberg, C Piechaczek, G Gassmann, NA
Wolff, F Thevenod, C Kaltschmidt and B Kaltschmidt. (2007). Highly efficient neural
differentiation of human somatic stem cells, isolated by minimally invasive periodontal surgery.
Stem Cells Dev 16:447-460.
32. Widera D, C Zander, M Heidbreder, Y Kasperek, T Noll, O Seitz, B Saldamli, H Sudhoff,
R Sader, C Kaltschmidt and B Kaltschmidt. (2009). Adult palatum as a novel source of neural
crest-related stem cells. Stem Cells 27:1899-1910.
33. Dupin E and L Sommer. (2012). Neural crest progenitors and stem cells: from early
development to adulthood. Dev Biol 366:83-95.
34. Fernandes KJ, JG Toma and FD Miller. (2008). Multipotent skin-derived precursors:
adult neural crest-related precursors with therapeutic potential. Philos Trans R Soc Lond B Biol
Sci 363:185-198.
35. Shakhova O and L Sommer. Neural crest-derived stem cells. (2010). StemBook,
Page 35 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

36
36
Cambridge, MA, Harvard Stem Cell Institute, p. 2008.
36. Chai Y, X Jiang, Y Ito, P Bringas, Jr., J Han, DH Rowitch, P Soriano, AP McMahon and
HM Sucov. (2000). Fate of the mammalian cranial neural crest during tooth and mandibular
morphogenesis. Development 127:1671-1679.
37. Rinn JL, JK Wang, H Liu, K Montgomery, M van de Rijn and HY Chang. (2008). A
systems biology approach to anatomic diversity of skin. J Invest Dermatol 128:776-782.
38. Hill MW and IC Mackenzie. (1989). The influence of subepithelial connective tissues on
epithelial proliferation in the adult mouse. Cell Tissue Res 255:179-182.
39. Okazaki M, K Yoshimura, Y Suzuki and K Harii. (2003). Effects of subepithelial
fibroblasts on epithelial differentiation in human skin and oral mucosa: heterotypically
recombined organotypic culture model. Plast Reconstr Surg 112:784-792.
40. Mikkola ML and SE Millar. (2006). The mammary bud as a skin appendage: unique and
shared aspects of development. J Mammary Gland Biol Neoplasia 11:187-203.
41. Huang GT, S Gronthos and S Shi. (2009). Mesenchymal stem cells derived from dental
tissues vs. those from other sources: their biology and role in regenerative medicine. J Dent Res
88:792-806.
42. Yang CC and G Cotsarelis. (2010). Review of hair follicle dermal cells. J Dermatol Sci
57:2-11.
43. Seo BM, M Miura, S Gronthos, PM Bartold, S Batouli, J Brahim, M Young, PG Robey,
CY Wang and S Shi. (2004). Investigation of multipotent postnatal stem cells from human
periodontal ligament. Lancet 364:149-155.
44. Hynes K, D Menicanin, S Gronthos and PM Bartold. (2012). Clinical utility of stem cells
for periodontal regeneration. Periodontol 2000 59:203-227.
Page 36 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

37
37
45. Wu G, ZH Deng, XJ Fan, ZF Ma, YJ Sun, DD Ma, JJ Wu, JN Shi and Y Jin. (2009).
Odontogenic potential of mesenchymal cells from hair follicle dermal papilla. Stem Cells Dev
18:583-589.
46. Atari M, J Caballe-Serrano, C Gil-Recio, C Giner-Delgado, E Martinez-Sarra, DA
Garcia-Fernandez, M Barajas, F Hernandez-Alfaro, E Ferres-Padro and L Giner-Tarrida. (2012).
The enhancement of osteogenesis through the use of dental pulp pluripotent stem cells in 3D.
Bone 50:930-941.
47. Davies LC, M Locke, RD Webb, JT Roberts, M Langley, DW Thomas, CW Archer and P
Stephens. (2010). A multipotent neural crest-derived progenitor cell population is resident within
the oral mucosa lamina propria. Stem Cells Dev 19:819-830.
48. Estrela C, AH Alencar, GT Kitten, EF Vencio and E Gava. (2011). Mesenchymal stem
cells in the dental tissues: perspectives for tissue regeneration. Braz Dent J 22:91-98.
49. Fournier BP, FC Ferre, L Couty, JJ Lataillade, M Gourven, A Naveau, B Coulomb, A
Lafont and B Gogly. (2010). Multipotent Progenitor Cells in Gingival Connective Tissue. Tissue
Eng Part A 16:2891-2899.
50. Gronthos S, M Mankani, J Brahim, PG Robey and S Shi. (2000). Postnatal human dental
pulp stem cells (DPSCs) in vitro and in vivo. Proc Natl Acad Sci (USA) 97:13625-13630.
51. Hung TY, HC Lin, YJ Chan and K Yuan. (2012). Isolating stromal stem cells from
periodontal granulation tissues. Clin Oral Investig 16:1171-1180.
52. Miura M, S Gronthos, M Zhao, B Lu, LW Fisher, PG Robey and S Shi. (2003). SHED:
stem cells from human exfoliated deciduous teeth. Proc Natl Acad Sci (USA) 100:5807-5812.
53. Morsczeck C, W Gotz, J Schierholz, F Zeilhofer, U Kuhn, C Mohl, C Sippel and KH
Hoffmann. (2005). Isolation of precursor cells (PCs) from human dental follicle of wisdom teeth.
Page 37 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

38
38
Matrix Biol 24:155-165.
54. Sonoyama W, Y Liu, D Fang, T Yamaza, BM Seo, C Zhang, H Liu, S Gronthos, CY
Wang, S Wang and S Shi. (2006). Mesenchymal stem cell-mediated functional tooth regeneration
in swine. PLoS One 1:e79.
55. Hsu SH, GS Huang and F Feng. (2012). Isolation of the multipotent MSC subpopulation
from human gingival fibroblasts by culturing on chitosan membranes. Biomaterials 33:2642-
2655.
56. Davies LC, H Lonnies, M Locke, B Sundberg, K Rosendahl, C Gotherstrom, K Le Blanc
and P Stephens. (2012). Oral mucosal progenitor cells are potently immunosuppressive in a dose-
independent manner. Stem Cells Dev 21:1478-1487.
57. Strioga M, S Viswanathan, A Darinskas, O Slaby and J Michalek. (2012). Same or not the
same? Comparison of adipose tissue-derived versus bone marrow-derived mesenchymal stem and
stromal cells. Stem Cells Dev 21:2724-2752.
58. Wang F, M Yu, X Yan, Y Wen, Q Zeng, W Yue, P Yang and X Pei. (2011). Gingiva-
derived mesenchymal stem cell-mediated therapeutic approach for bone tissue regeneration. Stem
Cells Dev 20:2093-2102.
59. Zhang Q, S Shi, Y Liu, J Uyanne, Y Shi, S Shi and AD Le. (2009). Mesenchymal stem
cells derived from human gingiva are capable of immunomodulatory functions and ameliorate
inflammation-related tissue destruction in experimental colitis. J Immunol 183:7787-7798.
60. Tomar GB, RK Srivastava, N Gupta, AP Barhanpurkar, ST Pote, HM Jhaveri, GC Mishra
and MR Wani. (2010). Human gingiva-derived mesenchymal stem cells are superior to bone
marrow-derived mesenchymal stem cells for cell therapy in regenerative medicine. Biochem
Biophys Res Commun 393:377-383.
Page 38 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

39
39
61. Dezawa M, M Hoshino, Y Nabeshima and C Ide. (2005). Marrow stromal cells:
implications in health and disease in the nervous system. Curr Neuropharmacol 5:723-732.
62. Arthur A, G Rychkov, S Shi, SA Koblar and S Gronthos. (2008). Adult human dental
pulp stem cells differentiate toward functionally active neurons under appropriate environmental
cues. Stem Cells 26:1787-1795.
63. Ferro F, R Spelat, F D'Aurizio, E Puppato, M Pandolfi, AP Beltrami, D Cesselli, G Falini,
CA Beltrami and F Curcio. (2012). Dental pulp stem cells differentiation reveals new insights in
Oct4A dynamics. PLoS One 7:e41774.
64. Sakai K, A Yamamoto, K Matsubara, S Nakamura, M Naruse, M Yamagata, K Sakamoto,
R Tauchi, N Wakao, S Imagama, H Hibi, K Kadomatsu, N Ishiguro and M Ueda. (2012). Human
dental pulp-derived stem cells promote locomotor recovery after complete transection of the rat
spinal cord by multiple neuro-regenerative mechanisms. J Clin Invest122:80-90.
65. Zhang W, XF Walboomers, TH Van Kuppevelt, WF Daamen, PA Van Damme, Z Bian
and JA Jansen. (2008). In vivo evaluation of human dental pulp stem cells differentiated towards
multiple lineages. J Tissue Eng Regen Med 2:117-125.
66. Sakai VT, Z Zhang, Z Dong, KG Neiva, MA Machado, S Shi, CF Santos and JE Nor.
(2010). SHED differentiate into functional odontoblasts and endothelium. J Dent Res 89:791-796.
67. Seo BM, W Sonoyama, T Yamaza, C Coppe, T Kikuiri, K Akiyama, JS Lee and S Shi.
(2008). SHED repair critical-size calvarial defects in mice. Oral Dis 14:428-434.
68. Sonoyama W, Y Liu, T Yamaza, RS Tuan, S Wang, S Shi and GT Huang. (2008).
Characterization of the apical papilla and its residing stem cells from human immature permanent
teeth: a pilot study. J Endod 34:166-171.
69. d'Aquino R, V Tirino, V Desiderio, M Studer, GC De Angelis, L Laino, A De Rosa, D Di
Page 39 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

40
40
Nucci, S Martino, F Paino, M Sampaolesi and G Papaccio. (2011). Human neural crest-derived
postnatal cells exhibit remarkable embryonic attributes either in vitro or in vivo. Eur Cell Mater
21:304-316.
70. Yokoi T, M Saito, T Kiyono, S Iseki, K Kosaka, E Nishida, T Tsubakimoto, H Harada, K
Eto, T Noguchi and T Teranaka. (2007). Establishment of immortalized dental follicle cells for
generating periodontal ligament in vivo. Cell Tissue Res 327:301-311.
71. Dong R, X Liu, M Fan, L Yang, L Peng and L Zhang. (2010). Isolation and
differentiation of nestin positive cells from rat oral mucosal lamina propria. Differentiation 79:9-
14.
72. Mitrano TI, MS Grob, F Carrion, E Nova-Lamperti, PA Luz, FS Fierro, A Quintero, A
Chaparro and A Sanz. (2010). Culture and characterization of mesenchymal stem cells from
human gingival tissue. J Periodontol 81:917-925.
73. Wei F, T Song, G Ding, J Xu, Y Liu, D Liu, Z Fan, C Zhang, S Shi and S Wang. (2013).
Functional tooth restoration by allogeneic mesenchymal stem cell-based bio-root regeneration in
swine. Stem Cells Dev 22:1752-1762.
74. Marynka-Kalmani K, S Treves, M Yafee, H Rachima, Y Gafni, MA Cohen and S Pitaru.
(2010). The lamina propria of adult human oral mucosa harbors a novel stem cell population.
Stem Cells 28:984-995.
75. Xu X, C Chen, K Akiyama, Y Chai, AD Le, Z Wang Z and S Shi. (2013). Gingivae
Contain Neural-crest- and Mesoderm-derived Mesenchymal Stem Cells. J Dent Res Jul 18. [Epub
ahead of print]
76. Treves-Manusevitz S, L Hoz , H Rachima , G Montoya , E Tzur , A Vardimon , AS
Narayanan , S Amar , H Arzate and S Pitaru. (2013) Stem cells of the lamina propria of human
Page 40 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

41
41
oral mucosa and gingiva develop into mineralized tissues in vivo. J Clin Periodontol 40:73-81.
77. Gotherstrom C, O Ringden, C Tammik, E Zetterberg, M Westgren and K Le Blanc.
(2004). Immunologic properties of human fetal mesenchymal stem cells. Am J Obstet Gynecol
190:239-245.
78. Atari M, C Gil-Recio, M Fabregat, D Garcia-Fernandez, M Barajas, MA Carrasco, HS
Jung, FH Alfaro, N Casals, F Prosper, E Ferres-Padro and L Giner. (2012). Dental pulp of the
third molar: a new source of pluripotent-like stem cells. J Cell Sci 125:3343-3356.
79. Squier CA. (1991). The permeability of oral mucosa. Crit Rev Oral Biol Med 2:13-32.
80. Winning TA and GC Townsend. (2000). Oral mucosal embryology and histology. Clin
Dermatol 18:499-511.
81. Collins LM and C Dawes. (1987). The surface area of the adult human mouth and
thickness of the salivary film covering the teeth and oral mucosa. J Dent Res 66:1300-1302.
82. Karring T, NP Lang and H Loe. (1975). The role of gingival connective tissue in
determining epithelial differentiation. J Periodont Res 10:1-11.
83. Karring T, E Ostergaard and H Loe. (1971). Conservation of tissue specificity after
heterotopic transplantation of gingiva and alveolar mucosa. J Periodont Res 6:282-293.
84. Schweizer J, H Winter, MW Hill and IC Mackenzie. (1984). The keratin polypeptide
patterns in heterotypically recombined epithelia of skin and mucosa of adult mouse.
Differentiation 26:144-153.
85. Schroeder H. Oral Structural Biology. (1991). Thieme Medical Publisher, New York,
USA.
86. Page RC and WF Ammons. (1974). Collagen turnover in the gingiva and other mature
connective tissues of the marmoset Saguinus oedipus. Arch Oral Biol 19:651-658.
Page 41 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

42
42
87. Ouhayoun JP, F Gosselin, N Forest, S Winter and WW Franke. (1985). Cytokeratin
patterns of human oral epithelia: differences in cytokeratin synthesis in gingival epithelium and
the adjacent alveolar mucosa. Differentiation 30:123-129.
88. Ermich T, J Schulz and D Schumann. (1988). Pattern of oral cytokeratins: I. SDS-
electrophoretic analysis of the frequency of cytoskeletal keratins in the normal human mucosa of
mouth. Biomed Biochim Acta 47:737-742.
89. Presland RB and BA Dale. (2000). Epithelial structural proteins of the skin and oral
cavity: function in health and disease. Crit Rev Oral Biol Med 11:383-408.
90. Paladini RD, K Takahashi, NS Bravo and PA Coulombe. (1996). Onset of re-
epithelialization after skin injury correlates with a reorganization of keratin filaments in wound
edge keratinocytes: defining a potential role for keratin 16. J Cell Biol 132:381-397.
91. Mak K, A Manji, C Gallant-Behm, C Wiebe, DA Hart, H Larjava and L Hakkinen. (2009).
Scarless healing of oral mucosa is characterized by faster resolution of inflammation and control
of myofibroblast action compared to skin wounds in the red Duroc pig model. J Dermatol Sci
56:168-180.
92. Wong JW, C Gallant-Behm, C Wiebe, K Mak, DA Hart, H Larjava and L Hakkinen.
(2009). Wound healing in oral mucosa results in reduced scar formation as compared with skin:
evidence from the red Duroc pig model and humans. Wound Repair Regen 17:717-729.
93. Schor SL, I Ellis, CR Irwin, J Banyard, K Seneviratne, C Dolman, AD Gilbert and DM
Chisholm. (1996). Subpopulations of fetal-like gingival fibroblasts: characterisation and potential
significance for wound healing and the progression of periodontal disease. Oral Dis 2:155-166.
94. Sciubba JJ, JP Waterhouse and J Meyer. (1978). A fine structural comparison of the
healing of incisional wounds of mucosa and skin. J Oral Pathol 7:214-227.
Page 42 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

43
43
95. Szpaderska AM, CG Walsh, MJ Steinberg and LA DiPietro. (2005). Distinct patterns of
angiogenesis in oral and skin wounds. J Dent Res 84:309-314.
96. Szpaderska AM, JD Zuckerman and LA DiPietro. (2003). Differential injury responses in
oral mucosal and cutaneous wounds. J Dent Res 82:621-626.
97. Schrementi ME, AM Ferreira, C Zender and LA DiPietro. (2008). Site-specific
production of TGF-beta in oral mucosal and cutaneous wounds. Wound Repair Regen 16:80-86.
98. Eslami A, CL Gallant-Behm, DA Hart, C Wiebe, D Honardoust, H Gardner, L Hakkinen
and HS Larjava. (2009). Expression of integrin alphavbeta6 and TGF-beta in scarless vs scar-
forming wound healing. J Histochem Cytochem 57:543-557.
99. Hakkinen L, VJ Uitto and H Larjava. (2000). Cell biology of gingival wound healing.
Periodontol 2000 24:127-152.
100. Häkkinen L, H Larjava and L Koivisto. (2012). Granulation Tissue Formation and
Remodeling. In: Oral Wound Healing: Cell Biology and Clinical Management. H Larjava, ed.,
John Wiley & Sons, Inc., Ames, pp. 125-173.
101. Chavrier C. (1990). The elastic system fibres in healthy human gingiva. Arch Oral Biol
35 Suppl:223S-225S.
102. Hsieh PC, YT Jin, CW Chang, CC Huang, SC Liao and K Yuan. (2010). Elastin in oral
connective tissue modulates the keratinization of overlying epithelium. J Clin Periodontol
37:705-711.
103. Kielty CM, MJ Sherratt and CA Shuttleworth. (2002). Elastic fibres. J Cell Sci 115:2817-
2828.
104. Lozdan J. (1970). Studies on the mucogingival junction. Dent Pract Dent Rec 20:379-384.
105. Goktas S, JJ Dmytryk and PS McFetridge. (2011). Biomechanical behavior of oral soft
Page 43 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

44
44
tissues. J Periodontol 82:1178-1186.
106. Lee-Thedieck C, N Rauch, R Fiammengo, G Klein and JP Spatz. (2012). Impact of
substrate elasticity on human hematopoietic stem and progenitor cell adhesion and motility. J
Cell Sci 125:3765-3775.
107. Tschumperlin DJ, F Liu and AM Tager. (2013). Biomechanical regulation of
mesenchymal cell function. Curr Opin Rheumatol 25:92-100.
108. Bielefeld KA, S Amini-Nik and BA Alman. (2012). Cutaneous wound healing: recruiting
developmental pathways for regeneration. Cell Mol Life Sci Oct 4. [Epub ahead of print]
109. Namazi MR, MK Fallahzadeh and RA Schwartz. (2011). Strategies for prevention of
scars: what can we learn from fetal skin? Int J Dermatol 50:85-93.
110. Brizzi MF, G Tarone and P Defilippi. (2012). Extracellular matrix, integrins, and growth
factors as tailors of the stem cell niche. Curr Opin Cell Biol 24:645-651.
111. Bronson RE, JG Argenta, EP Siebert and CN Bertolami. (1988). Distinctive fibroblastic
subpopulations in skin and oral mucosa demonstrated by differences in glycosaminoglycan
content. In Vitro Cell Dev Biol 24:1121-1126.
112. Narayanan AS, RC Page and DF Meyers. (1980). Characterization of collagens of
diseased human gingiva. Biochemistry 19:5037-5043.
113. Pedlar J. (1984). Biochemistry of glycosaminoglycans in the skin and oral mucosa of the
rat. Arch Oral Biol 29:591-597.
114. Larjava H, J Heino, T Krusius, E Vuorio and M Tammi. (1988). The small dermatan
sulphate proteoglycans synthesized by fibroblasts derived from skin, synovium and gingiva show
tissue-related heterogeneity. Biochem J 256:35-40.
115. Tsuruga E, K Irie, Y Sakakura and T Yajima. (2002). Tropoelastin expression by
Page 44 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

45
45
periodontal fibroblasts. J Dent Res 81:198-202.
116. Tsuruga E, K Irie, Y Sakakura and T Yajima. (2002). Expression of fibrillins and
tropoelastin by human gingival and periodontal ligament fibroblasts in vitro. J Periodont Res
37:23-28.
117. Tsuruga E, K Irie and T Yajima. (2002). Gene expression and accumulation of fibrillin-1,
fibrillin-2, and tropoelastin in cultured periodontal fibroblasts. J Dent Res 81:771-775.
118. Gogly B, G Godeau, S Gilbert, JM Legrand, C Kut, B Pellat and M Goldberg. (1997).
Morphometric analysis of collagen and elastic fibers in normal skin and gingiva in relation to age.
Clin Oral Investig 1:147-152.
119. Guo F, DE Carter and A Leask. (2011). Mechanical tension increases CCN2/CTGF
expression and proliferation in gingival fibroblasts via a TGFbeta-dependent mechanism. PLoS
One 6:e19756.
120. Guo F, DE Carter, A Mukhopadhyay and A Leask. (2011). Gingival fibroblasts display
reduced adhesion and spreading on extracellular matrix: a possible basis for scarless tissue repair?
PLoS One 6:e27097.
121. Ebisawa K, R Kato, M Okada, T Sugimura, MA Latif, Y Hori, Y Narita, M Ueda, H
Honda and H Kagami. (2011). Gingival and dermal fibroblasts: their similarities and differences
revealed from gene expression. J Biosci Bioeng 111:255-258.
122. Macri L, D Silverstein and RA Clark. (2007). Growth factor binding to the pericellular
matrix and its importance in tissue engineering. Adv Drug Deliv Rev 59:1366-81.
123. Reilly GC and AJ Engler. (2010). Intrinsic extracellular matrix properties regulate stem
cell differentiation. J Biomech 43:55-62.
124. Goktas S, JJ Dmytryk and PS McFetridge. (2011). Biomechanical behavior of oral soft
Page 45 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

46
46
tissues. J Periodontol 82:1178-86.
125. Balestrini JL, S Chaudhry, V Sarrazy, A Koehler and B Hinz. (2012). The mechanical
memory of lung myofibroblasts. Integr Biol (Camb) 4:410-21.
126. Csiszar A, C Wiebe, H Larjava and L Hakkinen. (2007). Distinctive molecular
composition of human gingival interdental papilla. J Periodontol 78:304-314.
127. Grey AM, AM Schor, G Rushton, I Ellis and SL Schor. (1989). Purification of the
migration stimulating factor produced by fetal and breast cancer patient fibroblasts. Proc Natl
Acad Sci (USA) 86:2438-2442.
128. Schor SL, AM Schor, AM Grey and G Rushton. (1988). Foetal and cancer patient
fibroblasts produce an autocrine migration-stimulating factor not made by normal adult cells. J
Cell Sci 90 ( Pt 3):391-399.
129. Cockey GH, JA Boughman, EL Harris and TM Hassell. (1989). Genetic control of
variation in human gingival fibroblast proliferation rate. In Vitro Cell Dev Biol 25:255-258.
130. Hassell TM and EJ Stanek, 3rd. (1983). Evidence that healthy human gingiva contains
functionally heterogeneous fibroblast subpopulations. Arch Oral Biol 28:617-625.
131. Hakkinen L and H Larjava. (1992). Characterization of fibroblast clones from periodontal
granulation tissue in vitro. J Dent Res 71:1901-1907.
132. Ko SD, RC Page and AS Narayanan. (1977). Fibroblast heterogeneity and prostaglandin
regulation of subpopulations. Proc Natl Acad Sci (USA) 74:3429-3432.
133. Connor NS, JE Aubin and J Sodek. (1983). Independent expression of type I collagen and
fibronectin by normal fibroblast-like cells. J Cell Sci 63:233-244.
134. Bordin S, RC Page and AS Narayanan. (1984). Heterogeneity of normal human diploid
fibroblasts: isolation and characterization of one phenotype. Science 223:171-173.
Page 46 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

47
47
135. Hassell TM, DV Provenza and RA Foster. (1986). Synthetic activities of mass cultures
and clones of human gingival fibroblasts. Experientia 42:66-69.
136. Hou LT and JA Yaeger. (1993). Cloning and characterization of human gingival and
periodontal ligament fibroblasts. J Periodontol 64:1209-1218.
137. Fries KM, T Blieden, RJ Looney, GD Sempowski, MR Silvera, RA Willis and RP Phipps.
(1994). Evidence of fibroblast heterogeneity and the role of fibroblast subpopulations in fibrosis.
Clin Immunol Immunopathol 72:283-292.
138. Sempowski GD, MA Borrello, TM Blieden, RK Barth and RP Phipps. (1995). Fibroblast
heterogeneity in the healing wound. Wound Repair Regen 3:120-131.
139. Lekic PC, N Pender and CA McCulloch. (1997). Is fibroblast heterogeneity relevant to
the health, diseases, and treatments of periodontal tissues? Crit Rev Oral Biol Med 8:253-268.
140. Bordin S, LG Costa and X Tan. (1998). Fibroblast heterogeneity of signal transduction
mechanisms to complement-C1q. Analyses of calcium mobilization, inositol phosphate
accumulation, and protein kinases-C redistribution. J Periodontol 69:642-649.
141. Gagari E, MK Rand, L Tayari, H Vastardis, P Sharma, PV Hauschka and PD Damoulis.
(2006). Expression of stem cell factor and its receptor, c-kit, in human oral mesenchymal cells.
Eur J Oral Sci 114:409-415.
142. Trackman PC and A Kantarci. (2004). Connective tissue metabolism and gingival
overgrowth. Crit Rev Oral Biol Med 15:165-175.
143. Hassell TM and GH Gilbert. (1983). Phenytoin sensitivity of fibroblasts as the basis for
susceptibility to gingival enlargement. Am J Pathol 112:218-223.
144. Hassell TM, E Romberg, S Sobhani, L Lesko and R Douglas. (1988). Lymphocyte-
mediated effects of cyclosporine on human fibroblasts. Transplant Proc 20:993-1002.
Page 47 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

48
48
145. Stabellini G, C Calastrini, G Mariani, M Gioia, E Caramelli, N Gagliano, C Dellavia, C
Moscheni, L Vizzotto and EM Calvi. (2004). Extracellular glycosaminoglycan changes in healthy
and overgrown gingiva fibroblasts after cyclosporin A and cytokine treatments. J Oral Pathol
Med 33:346-353.
146. Tang L, N Li, H Xie and Y Jin. (2011). Characterization of mesenchymal stem cells from
human normal and hyperplastic gingiva. J Cell Physiol 226:832-842.
147. Mensing N, H Gasse, N Hambruch, JD Haeger, C Pfarrer and C Staszyk. (2011). Isolation
and characterization of multipotent mesenchymal stromal cells from the gingiva and the
periodontal ligament of the horse. BMC Vet Res 7:42.
148. Ross R and G Odland. (1968). Human wound repair. II. Inflammatory cells, epithelial-
mesenchymal interrelations, and fibrogenesis. J Cell Biol 39:152-168.
149. Helpap B, H Breining, S Cappel, KW Sturm and S Lymberopoulos. (1976). Wound
healing of the brain of rats after cryonecrosis. Autoradiographic investigations with 3H-
thymidine. Virchows Arch B Cell Pathol 22:151-161.
150. Crisan M, M Corselli, CW Chen and B Peault. (2011). Multilineage stem cells in the adult:
a perivascular legacy? Organogenesis 7:101-104.
151. Dulmovits BM and IM Herman. (2012). Microvascular remodeling and wound healing: a
role for pericytes. Int J Biochem Cell Biol 44:1800-1812.
152. Diaz-Flores L, R Gutierrez, JF Madrid, H Varela, F Valladares, E Acosta, P Martin-
Vasallo and L Diaz-Flores, Jr. (2009). Pericytes. Morphofunction, interactions and pathology in a
quiescent and activated mesenchymal cell niche. Histol Histopathol 24:909-969.
153. Lau K, R Paus, S Tiede, P Day and A Bayat. (2009). Exploring the role of stem cells in
cutaneous wound healing. Exp Dermatol 18:921-933.
Page 48 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

49
49
154. Feng J, A Mantesso, C De Bari, A Nishiyama and PT Sharpe. (2011). Dual origin of
mesenchymal stem cells contributing to organ growth and repair. Proc Natl Acad Sci (USA)
108:6503-6508.
155. Crisan M, S Yap, L Casteilla, CW Chen, M Corselli, TS Park, G Andriolo, B Sun, B
Zheng, L Zhang, C Norotte, PN Teng, J Traas, R Schugar, BM Deasy, S Badylak, HJ Buhring, JP
Giacobino, L Lazzari, J Huard and B Peault. (2008). A perivascular origin for mesenchymal stem
cells in multiple human organs. Cell Stem Cell 3:301-313.
156. Crisan M, B Deasy, M Gavina, B Zheng, J Huard, L Lazzari and B Peault. (2008).
Purification and long-term culture of multipotent progenitor cells affiliated with the walls of
human blood vessels: myoendothelial cells and pericytes. Methods Cell Biol 86:295-309.
157. Murakami S and H Okada. (1997). Lymphocyte-fibroblast interactions. Crit Rev Oral
Biol Med 8:40-50.
158. Ge S, KM Mrozik, D Menicanin, S Gronthos and PM Bartold. (2012). Isolation and
characterization of mesenchymal stem cell-like cells from healthy and inflamed gingival tissue:
potential use for clinical therapy. Regen Med 7(6):819-832.
159. Fawzy El-Sayed KM, S Paris , ST Becker , M Neuschl , W De Buhr , S Sälzer , A Wulff ,
M Elrefai , MS Darhous , M El-Masry , J Wiltfang and CE Dörfer . (2012). Periodontal
regeneration employing gingival margin-derived stem/progenitor cells: an animal study. J Clin
Periodontol 39:861-70.
160. Li N, N Liu, J Zhou, L Tang, B Ding, Y Duan and Y Jin. (2013). Inflammatory
environment induces gingival tissue-specific mesenchymal stem cells to differentiate towards a
pro-fibrotic phenotype. Biol Cell 105(6):261-275.
161. Takashiba S, K Naruishi and Y Murayama. (2003). Perspective of cytokine regulation for
Page 49 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

50
50
periodontal treatment: fibroblast biology. J Periodontol 74:103-110.
162. Zhang QZ, AL Nguyen, WH Yu and AD Le. (2012). Human oral mucosa and gingiva: a
unique reservoir for mesenchymal stem cells. J Dent Res 91:1011-8.
163. Chen M, W Su, X Lin, Z Guo J Wang, Q Zhang, D Brand, B Ryffel, J Huang, Z Liu, X
He, AD Le and SG Zheng. (2013). Adoptive transfer of human gingiva-derived mesenchymal
stem cells ameliorates collagen-induced arthritis VIA suppressing Th1 and Th17 and enhancing
regulatory T cell differentiation. Arthritis Rheum 65:1181-1193.
164. Zhang QZ, WR Su, SH Shi, P Wilder-Smith, AP Xiang, A Wong, AL Nguyen, CW Kwon
and AD Le. (2010). Human gingiva-derived mesenchymal stem cells elicit polarization of m2
macrophages and enhance cutaneous wound healing. Stem Cells 28:1856-68.
165. Su WR, Zhang QZ, Shi SH, Nguyen AL and Le AD. (2011). Human gingiva-derived
mesenchymal stromal cells attenuate contact hypersensitivity via prostaglandin E2-dependent
mechanisms. Stem Cells 29:1849-1860.
166. Cappellesso-Fleury S, B Puissant-Lubrano, PA Apoil, M Titeux, P Winterton, L Casteilla,
P Bourin and A Blancher. (2010). Human fibroblasts share immunosuppressive properties with
bone marrow mesenchymal stem cells. J Clin Immunol 30:607-619.
167. Wada N, D Menicanin, S Shi, PM Bartold and S Gronthos. (2009). Immunomodulatory
properties of human periodontal ligament stem cells. J Cell Physiol 219:667-676.
168. Shimabukuro Y, S Murakami and H Okada. (1992). Interferon-gamma-dependent
immunosuppressive effects of human gingival fibroblasts. Immunology 76:344-347.
169. Shimabukuro Y, S Murakami and H Okada. (1996). Antigen-presenting-cell function of
interferon gamma-treated human gingival fibroblasts. J Periodont Res 31:217-228.
170. Zou Z, Y Zhang, L Hao, F Wang, D Liu, Y Su and H Sun. (2010). More insight into
Page 50 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

51
51
mesenchymal stem cells and their effects inside the body. Expert Opin Biol Ther 10:215-230.
171. Mahanonda R, N Sa-Ard-Iam, P Montreekachon, A Pimkhaokham, K Yongvanichit, MM
Fukuda and S Pichyangkul. (2007). IL-8 and IDO expression by human gingival fibroblasts via
TLRs. J Immunol 178:1151-1157.
172. Nisapakultorn K, J Makrudthong, N Sa-Ard-Iam, P Rerkyen, R Mahanonda and O
Takikawa. (2009). Indoleamine 2,3-dioxygenase expression and regulation in chronic
periodontitis. J Periodontol 80:114-121.
173. Durand E, B Fournier, L Couty, M Lemitre, P Achouh, P Julia, L Trinquart, JN Fabiani, S
Seguier, B Gogly, B Coulomb and A Lafont. (2012). Endoluminal gingival fibroblast transfer
reduces the size of rabbit carotid aneurisms via elastin repair. Arterioscler Thromb Vasc Biol
32:1892-1901.
174. Gogly B, A Naveau, B Fournier, N Reinald, E Durand, C Brasselet, B Coulomb and A
Lafont. (2007). Preservation of rabbit aorta elastin from degradation by gingival fibroblasts in an
ex vivo model. Arterioscler Thromb Vasc Biol 27:1984-1990.
175. Guiha R, S el Khodeiry, L Mota and R Caffesse. (2001). Histological evaluation of
healing and revascularization of the subepithelial connective tissue graft. J Periodontol 72:470-
478.
176. Janson WA, MP Ruben, GM Kramer, AA Bloom and H Turner. (1969). Development of
the blood supply to split-thickness free ginival autografts. J Periodontol 40:707-716.
177. Oliver RC, H Loe and T Karring. (1968). Microscopic evaluation of the healing and
revascularization of free gingival grafts. J Periodont Res 3:84-95.
178. Suthin K, K Matsushita, M Machigashira, S Tatsuyama, T Imamura, M Torii and Y Izumi.
(2003). Enhanced expression of vascular endothelial growth factor by periodontal pathogens in
Page 51 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

52
52
gingival fibroblasts. J Periodont Res 38:90-96.
179. Hosokawa Y, I Hosokawa, K Ozaki, H Nakae and T Matsuo. (2005). Increase of CCL20
expression by human gingival fibroblasts upon stimulation with cytokines and bacterial
endotoxin. Clin Exp Immunol 142:285-291.
180. Okada Y, M Meguro, H Ohyama, S Yoshizawa, K Takeuchi-Hatanaka, N Kato, S
Matsushita, S Takashiba and F Nishimura. (2009). Human leukocyte histocompatibility antigen
class II-induced cytokines from human gingival fibroblasts promote proliferation of human
umbilical vein endothelial cells: potential association with enhanced angiogenesis in chronic
periodontal inflammation. J Periodont Res 44:103-109.
181. Kobayashi K, T Suzuki, Y Nomoto, Y Tada, M Miyake, A Hazama, I Wada, T Nakamura
and K Omori. (2010). A tissue-engineered trachea derived from a framed collagen scaffold,
gingival fibroblasts and adipose-derived stem cells. Biomaterials 31:4855-4863.
182. Kobayashi K, T Suzuki, Y Nomoto, Y Tada, M Miyake, A Hazama, T Nakamura and K
Omori. (2007). Potential of heterotopic fibroblasts as autologous transplanted cells for tracheal
epithelial regeneration. Tissue Eng 13:2175-2184.
183. Nolte SV, W Xu, HO Rennekampff and HP Rodemann. (2008). Diversity of fibroblasts--a
review on implications for skin tissue engineering. Cells Tissues Organs 187:165-176.
184. Felder JM, 3rd, E Hechenbleikner, M Jordan and J Jeng. (2012). Increasing the options
for management of large and complex chronic wounds with a scalable, closed-system dressing
for maggot therapy. J Burn Care Res 33:e169-175.
185. Metcalfe AD and MW Ferguson. (2007). Bioengineering skin using mechanisms of
regeneration and repair. Biomaterials 28:5100-5113.
186. Wong T, JA McGrath and H Navsaria. (2007). The role of fibroblasts in tissue
Page 52 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

53
53
engineering and regeneration. Br J Dermatol 156:1149-1155.
187. Jackson WM, LJ Nesti and RS Tuan. (2012). Mesenchymal stem cell therapy for
attenuation of scar formation during wound healing. Stem Cell Res Ther 3:20.
188. Nagy N, N Almaani, A Tanaka, JE Lai-Cheong, T Techanukul, JE Mellerio and JA
McGrath. (2011). HB-EGF induces COL7A1 expression in keratinocytes and fibroblasts:
possible mechanism underlying allogeneic fibroblast therapy in recessive dystrophic
epidermolysis Bullosa. J Invest Dermatol 131:1771-1774.
189. Tolar J, L Xia, MJ Riddle, CJ Lees, CR Eide, RT McElmurry, M Titeux, MJ Osborn, TC
Lund, A Hovnanian, JE Wagner and BR Blazar. (2011). Induced pluripotent stem cells from
individuals with recessive dystrophic epidermolysis bullosa. J Invest Dermatol 131:848-856.
190. Tziotzios C, C Profyris and J Sterling. (2012). Cutaneous scarring: Pathophysiology,
molecular mechanisms, and scar reduction therapeutics Part II. Strategies to reduce scar
formation after dermatologic procedures. J Am Acad Dermatol 66(1):13-24.
191. Greer N, N Foman J Dorrian, P Fitzgerald, R MacDonald, I Rutks I, and T Wilt.
Advanced Wound Care Therapies for Non-Healing Diabetic, Venous, and Arterial Ulcers: A
Systematic Review. (2012). Department of Veterans Affairs, Washington, DC, USA.
192. Takahashi K and S Yamanaka. (2006). Induction of pluripotent stem cells from mouse
embryonic and adult fibroblast cultures by defined factors. Cell 126:663-676.
193. Chun YS, K Byun and B Lee. (2011). Induced pluripotent stem cells and personalized
medicine: current progress and future perspectives. Anat Cell Biol 44:245-255.
194. Drews K, J Jozefczuk, A Prigione and J Adjaye. (2012). Human induced pluripotent stem
cells--from mechanisms to clinical applications. J Mol Med 90:735-745.
195. Zheng Z, J Jian, X Zhang, JN Zara, W Yin, M Chiang, Y Liu, J Wang, S Pang, K Ting
Page 53 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

54
54
and C Soo. (2012). Reprogramming of human fibroblasts into multipotent cells with a single
ECM proteoglycan, fibromodulin. Biomaterials 33:5821-5831.
196. Streckfuss-Bomeke K, F Wolf, A Azizian, M Stauske, M Tiburcy, S Wagner, D Hubscher,
R Dressel, S Chen, J Jende, G Wulf, V Lorenz, MP Schon, LS Maier, WH Zimmermann, G
Hasenfuss and K Guan. (2012). Comparative study of human-induced pluripotent stem cells
derived from bone marrow cells, hair keratinocytes, and skin fibroblasts. Eur Heart J. Jul 12.
[Epub ahead of print]
197. Miyoshi K, D Tsuji, K Kudoh, K Satomura, T Muto, K Itoh and T Noma. (2010).
Generation of human induced pluripotent stem cells from oral mucosa. J Biosci Bioeng 110:345-
350.
198. Tamaoki N, K Takahashi, T Tanaka, T Ichisaka, H Aoki, T Takeda-Kawaguchi, K Iida, T
Kunisada, T Shibata, S Yamanaka and K Tezuka. (2 010). Dental pulp cells for induced
pluripotent stem cell banking. J Dent Res 89:773-778.
199. Yan X, H Qin, C Qu, RS Tuan, S Shi and GT Huang. (2010). iPS cells reprogrammed
from human mesenchymal-like stem/progenitor cells of dental tissue origin. Stem Cells Dev
19:469-480.
200. Wada N, B Wang, NH Lin, AL Laslett, S Gronthos and PM Bartold. (2011). Induced
pluripotent stem cell lines derived from human gingival fibroblasts and periodontal ligament
fibroblasts. J Periodont Res 46:438-447.
201. Egusa H, K Okita, H Kayashima, G Yu, S Fukuyasu, M Saeki, T Matsumoto, S
Yamanaka and H Yatani. (2010). Gingival fibroblasts as a promising source of induced
pluripotent stem cells. PLoS One 5:e12743.
202. Srijaya TC, PJ Pradeep, RB Zain, S Musa, NH Abu Kasim and V Govindasamy. (2012).
Page 54 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

55
55
The promise of human induced pluripotent stem cells in dental research. Stem Cells Int
2012:423868.
203. Abdullah AI, A Pollock and T Sun. (2012). The path from skin to brain: generation of
functional neurons from fibroblasts. Mol Neurobiol 45:586-595.
Page 55 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

56
56
Figure legends
Page 56 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

57
57
Page 57 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

58
58
Figure 1. Comparison of fibroblasts and MSCs
A. Representative phase contrast microscope images of human gingival fibroblasts (GFBL) and
gingival stem cells (GSC) established from the fibroblast culture by CFU-F method. Images show
cells cultured at low and high density at passage 2. Morphologically, GFBLs and GSCs are
indistinguishable from each other. B. Key criteria for defining fibroblasts and mesenchymal stem
cells (MSCs). See 1-4 for references.
Page 58 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

59
59
Figure 2. Heterogeneity of connective tissue cells.
Plastic-adherent connective tissue cells form a group of cells that are phenotypically
heterogeneous between individuals, between different tissues within an individual and within a
given tissue. The phenotypic heterogeneity may result from presence of phenotypically different
fibroblast subpopulations, MSCs, myofibroblasts, pericytes and fibrocytes within a given cell
population. The different subsets of the cells share several morphological, molecular and
functional characteristics complicating their identification. However, relative proportions and
Page 59 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

60
60
distinct functions of these cell subsets may play important roles in tissue development,
maintenance, regeneration, repair and many disease processes.
Page 60 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

61
61
Figure 3. Clinical and histological features of gingiva within oral mucosa.
Page 61 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

62
62
A. Gingiva (G) represents the area of oral mucosa that surrounds teeth, is covered by a
keratinized epithelium and is demarcated from the non-keratinized oral lining mucosa (LM) by
the mucogingival junction (dotted line). B. Oral mucosa in the palate is covered by a similar
keratinized mucosa as gingiva. There is no apparent border between gingiva (G) that is closest to
the teeth and the palatal tissue proper. Characteristic features in the palate are the so-called rugae
(RG) that form through similar developmental program as other ectodermal appendages,
including teeth and hair (338). They can also contain so-called palatal neural crest-derived stem
cells (222). C. Representative histological section of human gingiva stained with hematoxylin
and eosin. Typical to the gingiva, epithelium forms long projections (rete pegs or rete ridges) that
extend deep into the connective tissue. D. Representative human gingival tissue section stained
with a monoclonal antibody that recognizes tenascin-C. Typically, gingiva displays strong
accumulation of tenascin-C at the subepithelial connective tissue. E. Collagen producing cells
populate gingival connective tissue. Representative image of immunostaining of type I
procollagen in human gingival tissue section. Type I collagen is most abundantly produced by
fibroblasts but can also be synthesized by other fibroblast-like cells. F. Immunostaining of CD90
(Thy-1) in human gingiva showing high density of immunopositive cells. In connective tissue,
CD90 can typically be expressed by fibroblasts, MSC, pericytes, myofibroblasts and fibrocytes. E:
epithelium; CT: connective tissue.
Page 62 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.
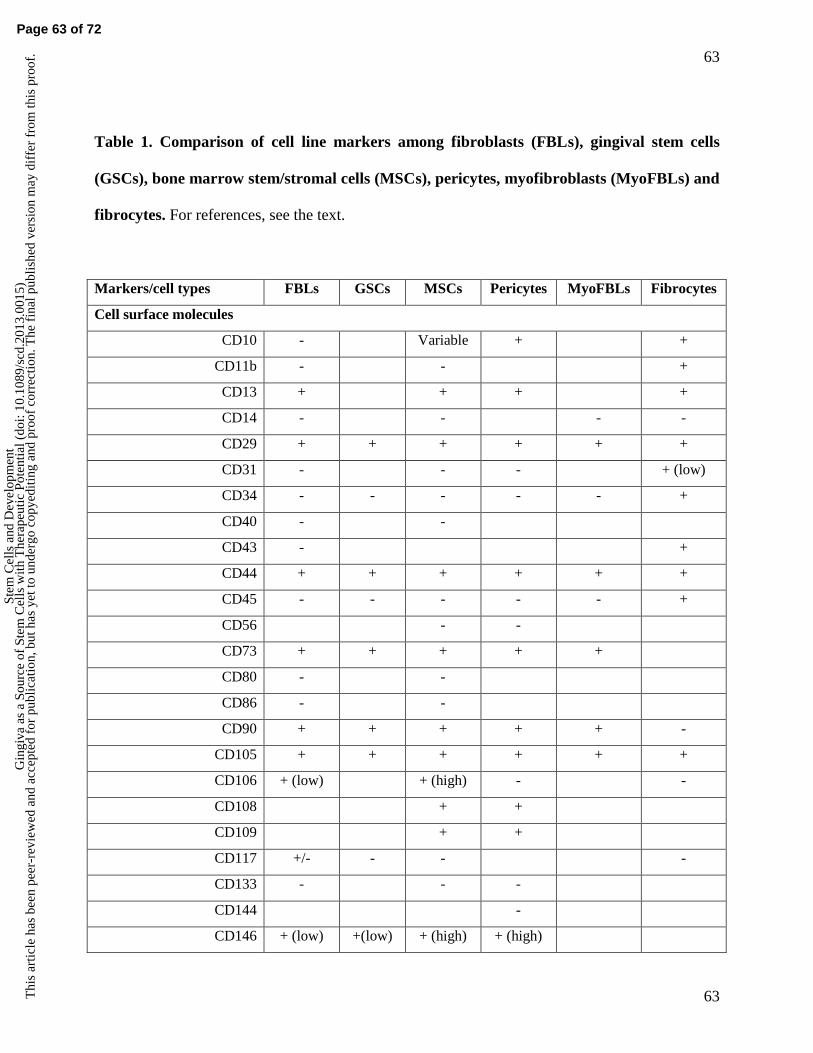
63
63
Table 1. Comparison of cell line markers among fibroblasts (FBLs), gingival stem cells
(GSCs), bone marrow stem/stromal cells (MSCs), pericytes, myofibroblasts (MyoFBLs) and
fibrocytes. For references, see the text.
Markers/cell types FBLs GSCs MSCs Pericytes MyoFBLs Fibrocytes
Cell surface molecules
CD10 - Variable + +
CD11b - - +
CD13 + + + +
CD14 - - - -
CD29 + + + + + +
CD31 - - - + (low)
CD34 - - - - - +
CD40 - -
CD43 - +
CD44 + + + + + +
CD45 - - - - - +
CD56 - -
CD73 + + + + +
CD80 - -
CD86 - -
CD90 + + + + + -
CD105 + + + + + +
CD106 + (low) + (high) - -
CD108 + +
CD109 + +
CD117 +/- - - -
CD133 - - -
CD144 -
CD146 + (low) +(low) + (high) + (high)
Page 63 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

64
64
CD164 + (low) + + +
CD166 Variable + + -
CD200 - - - (low)
CD271 Variable - + +
CD318 + +
CD324 - - - -
CD326 - -
CD340 + +
CD344 - -
CD349 + +
CCR2 Variable
CRCX4 +
HLA I + +
HLA II - - - -
HLA-ABC + +
HLA-DR - - - +
NG2 + +
PDGF-Rβ + +
PPARg -
STRO-1 + (low) + + -
STRO-3 + (low) + (high)
SSEA-4 + (low) + + +
3G5 - + +
Cytoskeletal and cytoplasmic proteins and enzymes
-SMA Variable - + + + Variable
Fibronexus +
FSP-1 (S100A4) + +
LSP-1 - +
MyoD -
Vimentin + (high) + + variable variable + (low)
Prolyl-4-hydroxylase + + + +
Alkaline phosphatase + - + +
Secreted molecules
Page 64 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

65
65
Bone sialoprotein (BSP) -
FGF-2 (bFGF) + +
Fibronectin + + + -
Collagen I + (high) + + + + + (low)
Collagen II - - - -
Collagen III + (high) + + + + (low)
MMP-9 - +
Neurofilament -
Osteocalcin (OC) - -
Osteonectin (SPARC) + +
Page 65 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

66
66
Table 2. Comparison of key phenotypic differences of gingival and skin connective tissue
cells. See text for references.
Adult Gingival Cells Adult Skin Cells
Originate mostly from neural crest Originate mostly from mesoderm
Different gene expression profile and site-specific HOX gene expression
Quantitative and qualitative differences in production of ECM molecules
Differences in integrin-family ECM receptors
Different interactions with ECM proteins
Different production and response to inflammatory mediators
Fast remodeling of a fibrin lattice Slower remodeling of a fibrin lattice
Fast invasion into a collagen matrix Slower invasion into a collagen matrix
Fetal-like phenotype that associates with fast
wound healing without scar formation
Phenotype associates with slower wound
healing and scar formation
Page 66 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

67
67
Table 3. Comparison of human bone marrow (MSCs) and various oral stem cells.
Origin Isolation technique Positive
markers
Negative
markers
Differentiation
potential
Reference
MSCs Human
bone
marrow
Adhesion properties
and
CFU-F
STRO-1
SSEA-4
CD29
CD44
CD73
CD90
CD105
CD146
VCAM1C
Coll1
Coll3
ON
FGF2
CD11b
CD14
CD34
CD45
HLA-DR
Myod
NF
Coll2
OC
BSP
OP
PPARg
In vitro: osteo-, adipo-,
and chondrogenic,
hepatocyte, skeletal
muscle, myocardial,
tendon, smooth muscle
and endothelial cell,
neuronal and glial
differentiation.
In vivo: Osteo- and
chondrogenic,
hepatocyte, muscle and
endothelial cell
differentiation.
Clinical trials for
treatment in humans:
Osteogenesis
Imperfecta, urinary
incontinence (smooth
muscle), erectile
dysfunction
(endothelium), cartilage
regeneration,
myocardial infarction,
liver regeneration.
(49, 50, 57,
61)
DPSCs
Dental
pulp stem
cells
Human
tooth dental
pulp
Enzymatic digestion
and CFU-F
SSEA-4
STRO-1
CD10
CD29
CD44
CD49a
CD14
CD34
CD45
MyoD
NF
Coll2
In vitro: Osteo- and
adipogenic, hepatocyte,
myocyte and neuronal
differentiation.
In vivo: dentine-pulp
complex formation,
50, 62, 65,
73)
Page 67 of 72 Page 68 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

68
68
CD73
CD90
CD105
CD117
CD146
VCAM1
Coll1
Coll3
OC
ON
FGF2
BSP
OP
PPARg
osteogenic and
adipogenic
differentiation,
oligodendrocyte and
neuronal differentiation.
No chondrogenic
differentiation.
SHEDs
Stem cell
from
human
exfoliated
deciduous
tooth
Human
exfoliated
deciduous
tooth pulp
Enzymatic digestion STRO-1
CD146
ALP
MEPE
FGF2
VEGFR1
Endostatin
In vitro: adipo-,
dentino-, osteogenic and
neuronal differentiation.
In vivo: bone formation,
odontoblasts and
endothelial
differentiation.
(52, 66, 67)
SCAPs
Stem cells
from apical
papilla
Human
apical
papilla of
immature
permanent
teeth
Enzymatic digestion
and CFU-F
STRO-1
ALP
CD24
CD29
CD73
CD90
CD105
CD106
CD146
CD166
CD18
CD34
CD45
CD150
In vitro: osteo-, adipo-,
dentogenic and neuronal
differentiation.
In vivo: tooth root and
periodontal ligament
formation.
(54, 68)
DFSCs
Dental
follicle
stem cell
Human
dental
follicle of
impacted
3rd
molars
Enzymatic digestion
and CFU-F or
SSEA4 sorting
SSEA-4
OCT4
CD90
CD56
Tuj1
TRAI-60
CD133 In vitro: osteo-, adipo-,
myo-, cementogenic and
neuronal differentiation.
In vivo: periodontal
ligament, skeletal and
adipose tissue
(53, 69, 70,
73)
Page 69 of 72 St
em C
ells
and
Dev
elop
men
tG
ingi
va a
s a
Sour
ce o
f St
em C
ells
with
The
rape
utic
Pot
entia
l (do
i: 10
.108
9/sc
d.20
13.0
015)
Thi
s ar
ticle
has
bee
n pe
er-r
evie
wed
and
acc
epte
d fo
r pu
blic
atio
n, b
ut h
as y
et to
und
ergo
cop
yedi
ting
and
proo
f co
rrec
tion.
The
fin
al p
ublis
hed
vers
ion
may
dif
fer
from
this
pro
of.
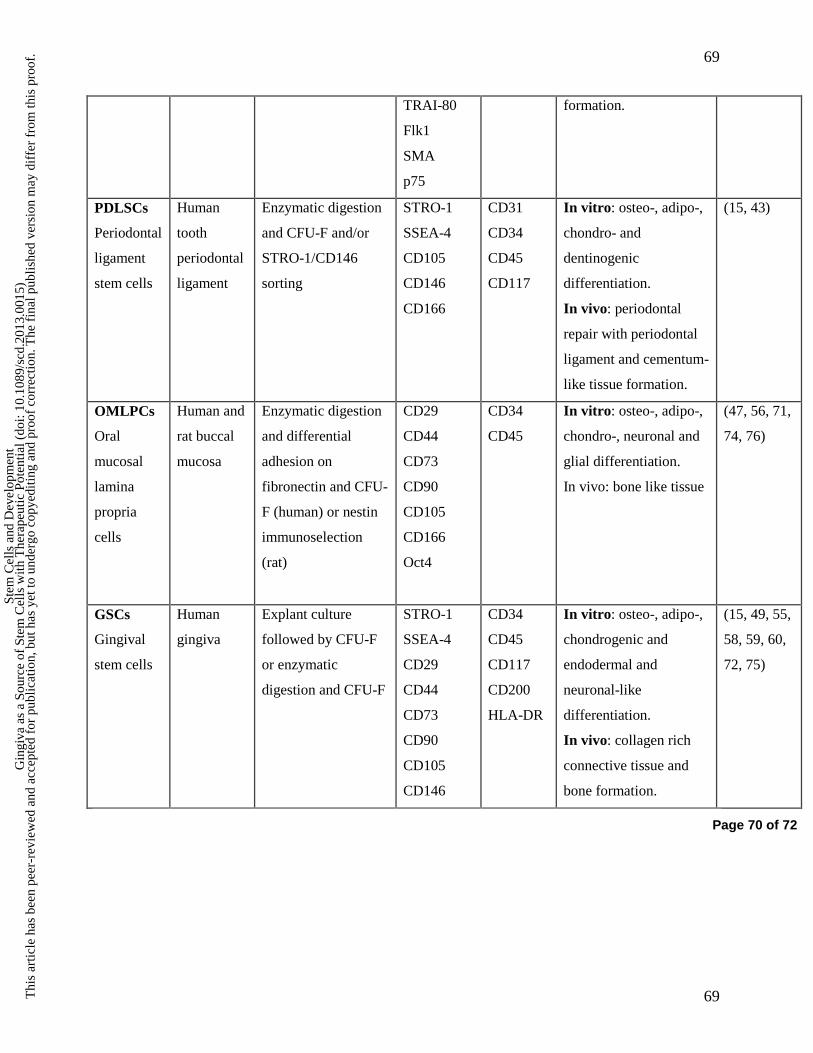
69
69
TRAI-80
Flk1
SMA
p75
formation.
PDLSCs
Periodontal
ligament
stem cells
Human
tooth
periodontal
ligament
Enzymatic digestion
and CFU-F and/or
STRO-1/CD146
sorting
STRO-1
SSEA-4
CD105
CD146
CD166
CD31
CD34
CD45
CD117
In vitro: osteo-, adipo-,
chondro- and
dentinogenic
differentiation.
In vivo: periodontal
repair with periodontal
ligament and cementum-
like tissue formation.
(15, 43)
OMLPCs
Oral
mucosal
lamina
propria
cells
Human and
rat buccal
mucosa
Enzymatic digestion
and differential
adhesion on
fibronectin and CFU-
F (human) or nestin
immunoselection
(rat)
CD29
CD44
CD73
CD90
CD105
CD166
Oct4
CD34
CD45
In vitro: osteo-, adipo-,
chondro-, neuronal and
glial differentiation.
In vivo: bone like tissue
(47, 56, 71,
74, 76)
GSCs
Gingival
stem cells
Human
gingiva
Explant culture
followed by CFU-F
or enzymatic
digestion and CFU-F
STRO-1
SSEA-4
CD29
CD44
CD73
CD90
CD105
CD146
CD34
CD45
CD117
CD200
HLA-DR
In vitro: osteo-, adipo-,
chondrogenic and
endodermal and
neuronal-like
differentiation.
In vivo: collagen rich
connective tissue and
bone formation.
(15, 49, 55,
58, 59, 60,
72, 75)
Page 70 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

70
70
Table 4. Special characteristics of human gingiva and gingival fibroblasts and MSCs that
can be beneficial for their use in various tissue regeneration modalities.
Gingiva in vivo
Easy access and low morbidity after tissue biopsy
Mild and short inflammatory response to wounding
Fast wound re-epithelialization and closure
Scarless wound healing response
Distinct composition of ECM
Distinct regulation of epithelial phenotype by connective tissue
Majority of the connective tissue cells originate from the neural crest
Gingival cells
Neural crest origin
Fetal-like phenotype
Distinct phenotypical properties
Potent immunomodulatory properties
Produce factors that promote vasculogenesis
Produce factors that promote epithelialization
Fibroblast cultures contain MSCs
Possess capacity for multipotent differentiation into mesodermal lineages and neural-like
differentiation
Efficient source to produce iPSCs
Page 71 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.

71
71
Page 72 of 72
Stem
Cel
ls a
nd D
evel
opm
ent
Gin
giva
as
a So
urce
of
Stem
Cel
ls w
ith T
hera
peut
ic P
oten
tial (
doi:
10.1
089/
scd.
2013
.001
5)T
his
artic
le h
as b
een
peer
-rev
iew
ed a
nd a
ccep
ted
for
publ
icat
ion,
but
has
yet
to u
nder
go c
opye
ditin
g an
d pr
oof
corr
ectio
n. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r fr
om th
is p
roof
.