genetic toxicology of the photosensitization of chinese hamster cells by phthalocyanines
TRANSCRIPT

Photochemistry and Photobiology Vol. 45, No. 2, pp. 227-230, 1987 Printed in Great Britain. All rights reserved
OO31-8655/87 $03 .OO + 0 .OO Copyright 0 1987 Pergamon Journals Ltd
GENETIC TOXICOLOGY OF THE PHOTOSENSITIZATION OF CHINESE HAMSTER CELLS
BY PHTHALOCYANINES E. BEN-HuR*, T. FUJIHARA?, F. SUZUKI* and M. M. ELKIND'.'
Department of Radiology and Radiation Biology and lCellular and Molecular Biology Interdisciplinary Program Graduate Faculty, Colorado State University. Fort Collins, CO 80523,
USA
(Received 30 May 1986; accepted 25 Augusr 1986)
Abstract-Chloroaluminum phthalocyanine (CAPC) was recently shown to photosensitize cell killing in culture and tumor destruction in vivo. Because this compound is potentially useful in the pho- todynamic therapy of cancer, its properties as a genotoxic agent were evaluated. Applying the technique of alkaline elution to study DNA integrity, it was found that CAPC could produce single- strand breaks in the DNA of Chinese hamster cells after exposure to white fluorescent light. At equicytotoxic doses, the number of DNA strand breaks produced by CAPC photosensitization was about three times lower than that induced by X-irradiation. During incubation in growth medium after exposure to CAPC-plus-fluorescent light, cells rejoined DNA strand breaks at a rate similar to that observed after X-irradiation. Resistance to 6-thioguanine (6-TG') or to ouabain (OUA') were used as end points of mutagenic potential. Following a treatment that caused -90% cell killing, there was a slight mutagenic effect, i.e. the frequencies were increased by -40% above the background or spontaneous mutations. However, this enhancement was not statistically significant. Taken together, the foregoing, plus an earlier observation that there is no variation in the sensitivity of cells to CAPC + light through the cell cycle, lead to the inferences that DNA damage does not play a major role in cell killing and that the mutagenic potential of this treatment is small.
INTRODUCTION
Photodynamic therapy (PDT)B is a modality for the experimental treatment of cancer generally utilizing the photosensitization of solid tumors to red light by treatment with H P D (see Wilson and Jeeves, 1986, for a recent review). Recently PC were shown to be efficient photosensitizers for cultured mam- malian cells (Ben-Hur and Rosenthal, 1985a-q Brasseur et a l . , 1986; Chan et a / . , 1986) as well as for the in vivo eradication of transplanted tumors in experimental animals (Ben-Hur et al., 1986; Sel- man et al., 1986).
Metallophthalocyanines are porphyrin-like com- pounds that strongly absorb red light (A,,, = 675 nm). Consequently, they are expected to utilize red light more efficiently than H P D because the latter compound has only a minor absorption peak at 630 nm. Also, 675 nm light penetrates more deeply into tissues than 630 nm. Another advantage of PC is that pure compounds may be used instead of the
*Present address: Nuclear Research Center-Negev. P.O. Box 9001, Beer-Sheva, Israel.
tPresent address: Department of Pathology, Kobe Uni- versity, Chuo-ku, Kobe 650, Japan.
$Present address: Division of Radiation Biology, Kan- azawa University, Takara-Machi, Kanazawa 920, Japan.
&To whom correspondence should be addressed. YAbbreviafions: CAPC, chloroaluminurn phthal-
ocyanine; FL, fluorescent light; HGPRT, hypoxanthine- guanine phosphoribosyl transferase; HPD, hemato- porphyrin derivative; OUA, ouabain; PBS, phosphate buffered saline; PC, phthalocyanines; PDT, photodynamic therapy; 6-TG. 6-thioguanine.
mixture comprising HPD. PC's have important tech- nological applications as heterogenous catalysts, dyes, and pigments. As a result, their toxicity has been extensively tested. Various PC derivatives were shown to be nontoxic when given in high doses for long periods to rabbits, mice, guinea pigs, cats, and dogs (Moser and Thomas, 1963). However, copper PC sulfonate used in the above tests is a poor photosensitizer (Ben-Hur et a/. , 1985). When the potent photosensitizer aluminum PC sulfonate is used, phototoxicity in rat liver can be demonstrated when the animals are exposed to 675 nm red laser light of intensity as low as 15 J cm-* (Ben-Hur et a / . , 1986).
Photosensitization by PC can cause membrane damage (Ben-Hur and Rosenthal, 1986). However, it is not yet known whether such damage is the basis for its cytotoxicity. DNA could also be a target. In the latter event, we would also expect PC pho- tosensitization to be mutagenic and possibly car- cinogenic as well.
In this work, the induction of DNA damage and its repair was studied using cultured Chinese ham- ster cells exposed to CAPC + FL. In addition, measurements of the induction of mutations at two different loci were made. The genotoxic potential of PC + FL appears to be small based on these criteria.
MATERIALS AND METHODS
Cells. Chinese hamster fibroblasts, clone V79-B310H. were grown in 10 cm plastic Petri dishes using culture techniques that have been described (Utsumi and Elkind,
327
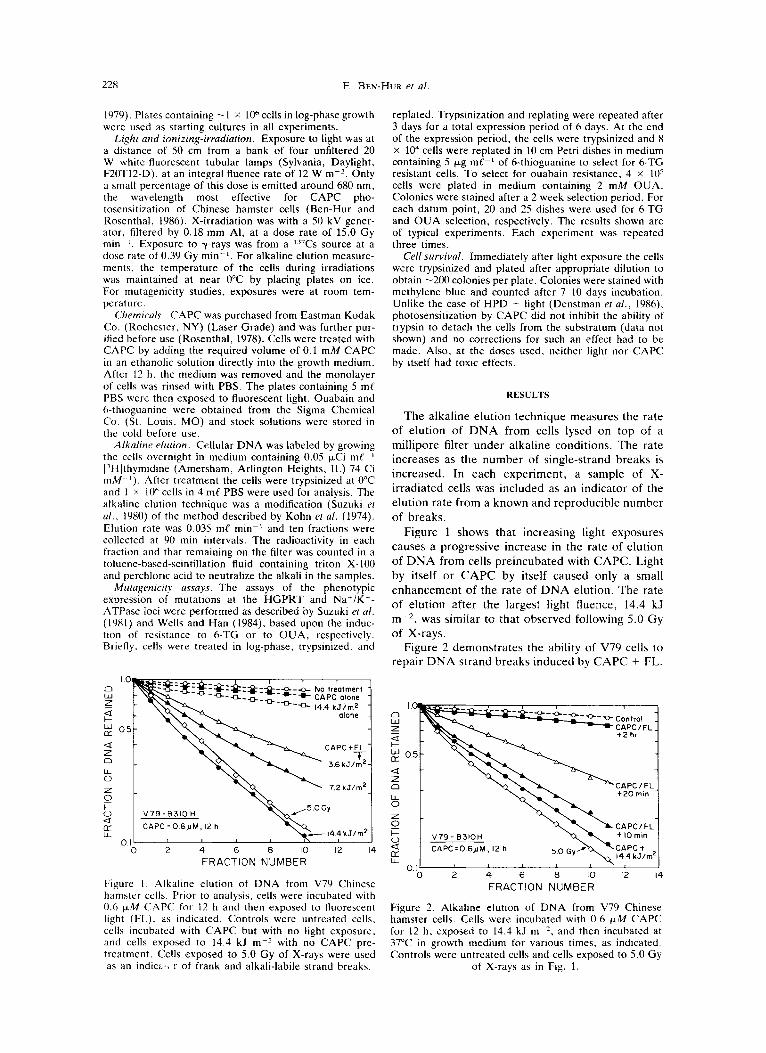
228 E. BEN-HUR et a/
1979). Plates containing - 1 x lo6 cells in log-phase growth were used as starting cultures in all experiments.
Light and ionizing-irradiation. Exposure to light was at a distance of 50 cm from a bank of four unfiltered 20 W white-fluorescent tubular lamps (Sylvania, Daylight, F20T12-D). at an integral fluence rate of 12 W m-2. Only a small percentage of this dose is emitted around 680 nm, the wavelength most effective for CAPC pho- tosensitization of Chinese hamster cells (Ben-Hur and Rosenthal, 1986). X-irradiation was with a 50 kV gener- ator, filtered by 0.18 mm Al, at a dose rate of 15.0 Gy min-I. Exposure to y-rays was from a 13'Cs source at a dose rate of 0.39 Gy min-I. For alkaline elution measure- ments, the temperature of the cells during irradiations was maintained at near 0°C by placing plates on ice. For mutagenicity studies, exposures were at room tem- perature.
Chemicals. CAPC was purchased from Eastman Kodak Co. (Rochester, NY) (Laser Grade) and was further pur- ified before use (Rosenthal, 1978). Cells were treated with CAPC by adding the required volume of 0.1 mM CAPC i n an ethanolic solution directly into the growth medium. After 12 h, the medium was removed and the monolayer o f cells was rinsed with PBS. The plates containing 5 mP PBS were then exposed to fluorescent light. Ouabain and 6-thioguanine were obtained from the Sigma Chemical Co. (St. Louis. MO) and stock solutions were stored in the cold before use.
Alkaline elution. Cellular D N A was labeled by growing the cells overnight in medium containing 0.05 pCi me- ' ['Hlthymidine (Amersham, Arlington Heights, IL) 74 Ci mM ~ ' ) . After treatment the cells were trypsinized at 0°C and 1 X 10" cells in 4 mP PBS were used for analysis. The alkaline elution technique was a modification (Suzuki et ul., 1980) of the method described by Kohn el al. (1974). Elution rate was 0.035 mY min-l and ten fractions were collected at 90 min intervals. The radioactivity in each fraction and that remaining on the filter was counted in a toluene-based-scintillation fluid containing triton X-100 and perchloric acid to neutralize the alkali in the samples.
Mutagenicity assays. The assays of the phenotypic expression of mutations at the H G P R T and Na+/K+- ATPase loci were performed as described by Suzuki et a/. (1981) and Wells and Han (1984), based upon the induc- tion of resistance to 6-TG or to O U A , respectively. Briefly. cells were treated in log-phase, trypsinized. and
n No treatment - W CAPC alone - z 14.4 kJ/m2 -
alone - 2 & 0.5
a C A P C + F L - z 3.6hJ/ rn2-
LL 0
\ - + - n
7 . 2 h J / m 2 ] 1
- . 0 2 4 6 8 1 0 1 2 1 4
FRACTION NUMBER
Figure I Alkaline elution of DNA from V79 Chinese hamster cells. Prior to analysis, cells were incubated with 0.6 p M CAPC for 12 h and then exposed to fluorescent light (FL). as indicated. Controls were untreated cells, cells incubated with CAPC but with no light exposure. and cells exposed to 14.4 kJ m-z with no CAPC pre- treatment. Cells exposed to 5.0 Gy of X-rays were used as an indici,> r of frank and alkali-labile strand breaks.
replated. Trypsinization and replating were repeated after 3 days for a total expression period of 6 days. At the end of the expression period, the cells were trypsinized and 8 X lo4 cells were replated in 10 cm Petri dishes in medium containing 5 pg me-' of 6-thioguanine to select for 6-TG resistant cells. To select for ouabain resistance. 4 x lo5 cells were plated in medium containing 2 mM O U A . Colonies were stained after a 2 week selection period. For each datum point, 20 and 25 dishes were used for 6-TG and O U A selection, respectively. The results shown are of typical experiments. Each experiment was repeated three times.
Cell survival. Immediately after light exposure the cells were trypsinized and plated after appropriate dilution to obtain -200 colonies per plate. Colonies were stained with methylene blue and counted after 7-10 days incubation. Unlike the case of H P D + light (Denstman et al., 1986). photosensitization by CAPC did not inhibit the ability of trypsin t o detach the cells from the substratum (data not shown) and no corrections for such an effect had to be made. Also, at the doses used, neither light nor CAPC by itself had toxic effects.
RESULTS
The alkaline elution technique measures the rate of elution of D N A from cells lysed on top of a millipore filter under alkaline conditions. The rate increases as the number of single-strand breaks is increased. In each experiment, a sample of X- irradiated cells was included as an indicator of the elution rate from a known and reproducible number of breaks.
Figure 1 shows that increasing light exposures causes a progressive increase in the rate of elution of DNA from cells preincubated with CAPC. Light by itself or CAPC by itself caused only a small enhancement of the rate of DNA elution. The rate of elution after the largest light fluence, 14.4 kJ m-2, was similar to that observed following 5.0 Gy of X-rays.
Figure 2 demonstrates the ability of V79 cells to repair D N A strand breaks induced by CAPC + FL.
, Y , 0.11 I
0 2 4 6 8 1 0 1 2 1 4 FRACTION NUMBER
Figure 2. Alkaline elution of DNA from V79 Chinese hamster cells. Cells were incubated with 0.6 p M CAPC for 12 h, exposed to 14.4 kJ m-?, and then incubated at 37°C in growth medium for various times, as indicated. Controls were untreated cells and cells exposed to 5.0 Gy
of X-ravs as in Fig. 1.

Genetic toxicology of phthalocyanines 229
Table 1 . Mutation induction by 0.6 pM chloroaluminum phthalocyanine followed by exposure to fluorescent light
Exposure Induction of 6-TG' cells Induction of OUA' cells (kJ m-z) per lo6 survivors (S.E.) per 10" survivors (S.E.)
0 17.4 t 2.7 2.15 t 0.34 2 17.5 2 2.5 4 26.5 2 4.3 1.43 2 0.31 6 22.4 ? 3.1 2.71 t 0.25
6-TG' = resistant to 6-TG. OUAr = resistant to OUA
10 0 5 0 2 01 SURVIVING FRACTION
! 0.6
Figure 3. Mutation induction by CAPC + FL and by y- rays in V79 Chinese hamster cells. For details see Materials and Methods. FL stands for fluorescent light. Control cells were exposed to light only or CAPC in the dark (A) , 0.6 pM for 12 h. The inset shows the survival curve of cells exposed to CAPC + FL. Note that mutation frequencies are observed values minus background fre-
quencies.
This is reflected in slower rates of elution when cells exposed to 14.4 kJ mP2 were subsequently incubated in growth medium for increasing time intervals at 37°C. After a 2-h repair interval, the rate of elution is almost the same as that of untreat- ed cells.
The data for mutagenicity of CAPC + FL, using resistance to 6-TG and to O U A as end points, are shown in Table 1 and in Fig. 3 . As a positive control, a typical induction curve of 6-TG' cells by y-rays was also measured. To compare the mutagenic potential of CAPC + FL to that of y-rays, in Fig. 3 the induction of 6-TG' and OUA' cells for these two treatments is plotted against surviving fraction. The inset in the figure shows the survival curve for CAPC + FL. It is evident that the latter treatment, for a given level of survival, is inefficiet in inducing phenotypic changes at a locus which responds to point mutations, OUA', as well as one which can be induced by major chromosomal changes as well as point mutations, 6-TG'. Neither light nor CAPC by itself increased the number of mutants above background.
DISCUSSION
The results with alkaline elution, Figs. 1 and 2, clearly show that photosensitization by CAPC can cause D N A strand breakage and that cells are able to repair this type of DNA damage. The breaks observed may not be frank breaks, however, but may reflect D N A damage that is expressed as breaks under alkaline conditions. This inference comes from the observation that 3-aminobenzamide does not enhance the photocytotoxicity induced by CAPC (Ben-Hur and Rosenthal, 1985b). 3-Amino- benzamide is a potent inhibitor of poly(ADP4bose) synthesis and as such it enhances the cytotoxicity of agents which produce D N A strand breaks, such as ionizing radiation (Ben-Hur, 1984; Elkind, 1985). Quantitatively, the amount of D N A damage that is detected after CAPC + 14.4 kJ m-' is about the same as after 5.0 Gy of X-rays (Figs. 1 and 2). At the latter dose, about 30% of V79 cells survive. After incubation in CAPC, an equitoxic exposure to FL is - 5 kJ mP2 (Ben-Hur and Rosenthal, 1985~) . Assuming a linear dependence of D N A damage on FL exposure after CAPC incubation, X- rays are about three times more efficient in pro- ducing D N A damage, expressible under alkaline conditions, than CAPC + FL.
Although D N A damage is produced after CAPC + FL treatment (Fig. l ) , the lack of an effect on survival due to the 3-aminobenzamide after treat- ment suggests that frank breaks d o not result because of the evidence that frank scissions in DNA due to single- and double-strand damage is required to activate the synthesis of poly(ADP-ribose) (Halldorson et al . , 1978; Benjamin and Gill, 1980). Alternatively, the cytotoxicity resulting from CAPC + FL may not involve DNA as a principal target, thus the lack of an effect of the inhibition of the synthesis of the polymer.
The small, statistically not significant changes in the frequencies of 6-TG' and OUA' cells with CAPC + FL treatment (Table 1 and Fig. 3 ) further support the inference that the DNA damage which results has little biological effect. Because for equal killing treatments, CAPC + FL is about one-third as effective in producing alkaline labile lesions as is

230 E. BEN-HUR ei al.
the case for ionizing radiation, it would not be likely that OUA' cells would be induced by CAPC + FL. The reason is that if the target for cell killing were to be D N A it would be very much too large com- pared to that corresponding to a target of o n e o r a few base pairs in the gene coding for N a + , K'- ATPase. For a cell t o be OUA' and t o survive, only a small change in the structural gene of this essential enzyme is possible. A t a killing treatment of , for example, C A P C + 14.4 kJ m-2 (i.e. survival G 0.01, Fig. 3), which has an effect o n DNA integrity under high p H conditions similar t o a 5.0 Gy dose of X-rays (Fig. l), o n average o n e alkaline labile lesion would have been registered in every 2 x lo6 base pairs, i .e . , a DNA target much too large to result in an induction of an altered but still func- tional Na+ ,K+-ATPase gene significantly evident above background.
In respect t o 6-TG resistance, t he lesser effec- tiveness of CAPC + FL treatment compared to y- rays (Fig. 3) suggests either that t he D N A damage due to CAPC + FL is ineffective or that t he repair, which is evident in Fig. 2 , is faithful and essentially complete in surviving cells.
Taken together, t he evidence discussed above, plus a lack of a variation in survival response through the cell cycle (Ben-Hur and Rosenthal, 1985c), suggests that cell killing d u e t o CAPC + FL treatment probably does not principally involve D N A and that the damage registered in D N A is only weakly mutagenic
Acknow'ledgements-We thank the United States Public Health Service, Department of Health and Human Services, for support for this research from the National Cancer Institute, under Grant Number CA 33701.
REFERENCES Ben-Hur. E. (1984) Involvement of poly(ADP-ribose)
in the radiation response of mammalian cells. Inr. J.
Ben-Hur. E.. A. Carmichael. P. Riesz and I. Rosen- thal (1985) Photochemical generation of superoxide radical and the cytotoxicity of phthalocyanines. In[. J. Radial. Biol. 48. 837-846.
Ben-Hur, E. and I. Rosenthal (1985a) The phthal- ocyanines: A new class of mammalian cell pho- tosensitizers with a potential for cancer phototherapy. Inr. 1. Radial. Biol. 47, 145-147.
Ben-Hur. E. and I. Rosenthal (1985b) Factors affect- ing the photokilling of cultured Chinese hamster cells by phthalocyanines. Radial. Res. 103, 403-409.
Ben-Hur, E. and I. Rosenthal (198%) Photosensitized inactivation of Chinese hamster cells by phthal- ocyanines. Photochem. Phoiobiol. 42. 129-133.
Ben-Hur. E. and I. Rosenthal (1986) Photohemolysis of human erythrocytes induced by aluminum phthalo- cyanine tetrasulfonate. Cancer Lerr. 30, 321-327.
Radiar. Biol. 46, 659-671.
Ben-Hur, E. and I. Rosenthal (1986) Action spectrum (600-700 nm) for chloroaluminum phthalocyanine- induced phototoxicity in Chinese hamster cells. Lasers Life Sci. 1. In press.
Ben-Hur, E., I. Rosenthal, S. G. Bown and D. Phil- lips (1986) The phthalocyanines: sensitizers with a potential for photodynamic therapy of cancer. In Pho- iomedicine (Edited by E. Ben-Hur and I. Rosenthal), Vol. 2, CRC Press, Boca Raton, FL. In press.
Benjamin, R. C. and D. M. Gill (1980) Poly(ADP- ribose) synthesis in vitro programmed by damaged DNA: a comparison of DNA molecules containing dif- ferent types of strand breaks. J . Biol. Chem. 255,
Brasseur, N . , H. Ah, D. Autenrieth, R. Langlois and J . E. van Lier (1985) Biological activities of phthal- ocyanines-111. Photoinactivation of V-79 Chinese ham- ster cells by tetrasulfophthalocyanines. Photochem. Phoiobiol. 42, 515-521.
Chan, W. S . , R. Svensen, D. Phillips and I. R. Hart (1986) Cell uptake, distribution and response to alumi- num chloro sulphonated phthalocyanine, a potential anti-tumor photosensitizer. Br. J. Cancer 53, 255-263.
Denstman, S. C., L. E. Dillehay and J. R. Wil- liams (1986) Enhanced susceptibility to HPD-sen- sitized phototoxicity and correlated resistance to trypsin detachment in SV-40 transformed IMR-90 cells. Pho- iochem. Phoiobiol. 43, 145-147.
Elkind, M. M. (1985) DNA damage and cell killing, cause and effect? Cancer 56, 2351-2363.
Halldorson, H., D. A. Gray and S. Shall (1978) Po- ly(ADP-ribose) polymerase activity in nucleotide per- meable cells. FEBS Lett. 85, 349-352.
Kohn, K. W., C. A. Friedman, R. A. G. Ewig and Z . M. Iqbal (1974) DNA chain growth during replication of L1210 cells. Alkaline elution of large DNA segments from cells lysed on filters. Biochemistry 13, 4134-4139.
Moser, F. H. and A. C. Thomas (1963) In Phihal- ocyanine Compounds. Reinhold, New York.
Rosenthal, I. (1978) Photochemical stability of Rho- damine 6G in solution. Opt. Commun. 24; 164-166.
Selman, S. H., M. Kreimer-Birnbaum, K. Chaudhuri, G. M. Garbo, D. A. Seaman, R. W. Kerk, E. Ben-Hur and I. Rosenthal (1986) Photodynamic treatment of transplantable bladder tumors in rodents after pre- treatment with chloroaluminum tetra- sulfophthalocyanine. J . Urol. 136, 141-145.
Suzuki, F., A. Han, G . R. Lankas, H. Utsumi and M. M. Elkind (1981) Spectral dependencies of killing, mutation and transformation in mammalian cells and their relevance to hazards caused by solar ultraviolet radiation. Cancer Res. 41, 4916-4924.
Suzuki, F., E . Watanabe and M. Horikawa (1980) Repair of X-ray-induced DNA damage in aging human diploid cells. Expl. Cell Res. 127, 299-307.
Utsumi, H . and M. M. Elkind (1979) Potentially lethal damage versus sublethal damage: independent repair processes in actively growing Chinese hamster cells. Radial. Res. 77, 346-360.
Wells, R. L. and A. Han (1984) Action spectra for killing and mutation of Chinese hamster cells exposed to mid- and near-ultraviolet monochromatic light. Muiai. Res. 129, 251-258.
Wilson, B. C. and P. Jeeves (1986) Photodynamic Therapy of Cancer. In Phoromedicine (Edited by E. Ben-Hur and I . Rosenthal), Vol. 2. CRC Press. Boca Raton, FL. In press.
10502-10508.