genetic recomeination in synchronized cultures of … · 2003-07-22 · genetic recomeination in...
TRANSCRIPT

GENETIC RECOMEINATION IN SYNCHRONIZED CULTURES OF SACCHAROMYCES CEREVISIAE
ROCHELLE E. ESPOSITO
Department of Genetics, Uniuersity of Washington, Seattle
Received October W . 1967
XPERIMENTAL evidence from studies in several fungi has demonstrated E that intragenic recombination is predominantly non-reciprocal whereas inter- genic recombination is primarily reciprocal. These observations have led to studies to determine whether intragenic and intergenic recombination derive from the same or different mechanisms. In recent years experimental data sup- porting both interpretations have been obtained. The frequent association of intragenic recombination with reciprocal recombination of outside markers in yeast (FOGEL and HURST 1967), Neurospora (MITCHELL 1955), and Aspergillus (PRITCHARD 1960), suggests that both events are the results of a single mecha- nism. However, the observations that intragenic and intergenic recombination respond differently to heat shock (MITCHELL 1957), changes in genetic back- ground ( STADLER 1959a; CATCHESIDE, JESSOP, and SMITH 1964), chemical muta- gens (EBBS 1967; ROMAN personal communication), and display different inter- ference patterns (STADLER 1959b), favor the view that different mechanisms are involved.
The existence of at least two recombination mechanisms was proposed by ROMAN and JACOB (1958), following their observation in yeast that outside marker exchange among ultraviolet-induced intragenic recombinants declines with increasing doses of irradiation, i.e. that intragenic and intergenic recom- binaton need not be obligatorily associated with one another. They suggested that intragenic recombination might occur during the period of DNA replication by a copying error whereas intergenic recombination might occur after replication by a breakage-reunion process. If it were possible to demonstrate that agents which induce recombination stimulate intragenic and intergenic events at different points in the cell cycle this would provide evidence in favor of their interpreta- tion. However, recent studies in Ustilago maydis (HOLLIDAY 1965, 1966), indi- cate that both recombinational events are maximally induced during the period of DNA synthesis.
The purpose of the following study was to inquire whether intragenic and intergenic recombination are maximally stimulated at the same or different points during the mitotic cell cycle of yeast. In the experiments reported below
Supported by Grants 5 R01 AI00328 and 5 T01-GM00328 of the National Insbtutes of Health,
Present address, Molecular Biology Laboratory, Unweisrty of Wisconsin, Madison, Wisconsin Public Health Service
Genetics 59: 191-210 June 1968

192 ROCHELLE E. ESPOSITO
ultraviolet light and X rays were used to stimulate recombination in synchro- nized cultures of Saccharomyces cerevisiae.
MATERIALS AND METHODS
Strains: The following strains of Saccharumyces cerevisiae were used during the course of this work. Diploids Z65 and Z69 were derived from stocks provided by Dr. D. C. HAWTHORNE, University of Washington, Seattle. Diploid XS380 was synthesized by Dr. S. NAKAI, National Institute of Radiological Sciences, Chiba-shi, Japan. The first number following the symbol designates the locus, the second, the allele. The linkage relationships of the relevant markers are given in centimorgans (MORTIMER and HAWTHORNE 1966).
265
0 b 2 - 1 ty1-1 h& ad, + + -- 0 60 35 38 5
17-2-2 ty1-2 + ad, url
ad1 + + 3- ad,
Z69
0 _ _ - + + o + h 2 - 2 ty1-2 hi7-l
XS380
0 [el-13 ty3-3 1Y 5-2 ad,,,-,
le1-1 ty3-1 lY5-I d 5 - 1 5 80 20 10 60
SYMBOLS: ad = adenine, hi = histidine, le = leucine, ly = lysine, met = methionine, t y = tyrosine, ur = uracil.
Media The amounts of the various ingredients indicated below are those required for the preparation of 1 liter. Solid medium was obtained by the addition of 15 g of Difco agar to 1 liter of medium.
YMA-I0 g dextrose, 5 g peptone, 3 g yeast extract, 3 g malt extract. Minimal-20 g dextrose, 6.7 g Difco yeast nitrogen base. Sporulation medium-9.8 g potassium acetate, 2.5 g yeast extract, 1 g dextrose, 10 mg histidine, 20 mg lysine, 50 mg tyrosine. Pregrowth and syn- chrony-feeding medium-the composition of these media varies depending upon the particular strain. For each diploid the pregrowth medium is its particular complex growth medium in double strength. The synchrony-feeding medium is identical to the complex growth medium. Starvation medium-$75 g potassium chloride, 0.34 g calcium chloride, 0.5 g magnesium chloride.
Media for 265: Complex growth medium-YMA plus 10 mg adenine, 10 mg histidine, 20 mg lysine, 10 mg methionine, 10 mg tryptophane, 50 mg tyrosine, 10 mg uracil. Defined growth medium for FUDR experiments-13.4 g Difco yeast nitrogen base, 30 mg adenine, 30 mg histidine, 60 mg lysine, 30 mg methionine, 30 mg tryptophane, 150 mg tyrosine, 30 mg uracil, brought to 800 ml with distilled water. 200 ml of sterile 10% dextrose and 1 g of 5-fluorodeoxyuridine (FUDR) were added after the above preparation had been autoclaved and cooled to room temperature. The FUDR was a gift from Hoffman La Roche, Nutley, New Jersey. Synthetic completeMinima1 plus 10 mg adenine, 10 mg histidine, 20 mg lysine, 10 mg methionine, 10 mg tryptophan, 50 mg tyrosine, 10 mg uracil. Histidineless, tyrosineless, and lysineless media-Synthetic complete minus the amino acid indicated .2 pg/ml tyrosine medium -Synthetic complete with 2 pg/ml tyrosine instead of 50 pg/ml tyrosine.
Media for Z69: Complex growth medium, synthetic complete, lysineless, tyrosineless, and histidineless media are the same as described for Z65. Magdala Red medium-Minimal plus 20 mg adenine, 20 mg histidine, 4Q mg lysine, 20 mg methionine, 20 mg tryptophan, 100 mg tyrosine, 20 mg uracil, 20 mg Magdala Red. The Magdala Red was made up as a solution of

RECOMBINATION IN SYNCHRONIZED YEAST 193
1 mg/ml in distilled water. The solution was sterilized at 100'C for 1 hour, and was added to the medium after the medium had hem autoclaved.
Media for XS380: Complex growth medium-YMA plus 25 mg adenine, 150 mg leucine, 50 mg lysine, 25 mg methionine, 125 mg phenlyalanine, 25 mg tryptophan, 125 mg tyrosine, 25 mg uracil. Synthetic complete-Minimal plus 10 mg adenine, 60 mg leucine, 40 mg lysine, 10 mg methionine, 50 mg phenylalanine, 10 mg tryptophan, 50 mg tyrosine. Adenineless, leucineless, lysineless, methionineless, and tyrosineless-phenylalanineless media-Synthetic com- plete minus the nutrilite indicated.
Procedure for synchronizing yeast cells: Synchronized cultures of Saccharomyces cereuisiae were obtained according to the methods described by WILLIAMSON and SCOPES (1962). The technique consists of alternately feeding and starving stationary phase cultures. Greater than 97% of the cells were single (unbudded) before the feeding-starvation treatment and remained unbudded during synchronization. The synchrony of a culture was measured in two ways. The culture was suspended in a complex or defined growth medium, aerated at 25"C, and the increase in the number of budding cells in time was observed with a phase contrast microscope. The synchrony was also measured by counting the number of cell units in the culture by means of a Coulter Counter. Samples were counted following a ten second exposure to ultrasonication with a Bronwell Biosonic 11 needle probe ultrasonication unit. The treatment was given to enhance separation of daughter cells at the completion of the first cell division. Under these cmditions, cell number rises with the appearance of the second generation of buds. Samples were removed at various points during synchronous growth and stored in formalin (two drops of 33% formaldehyde per ml of culture), for convenience, before being scored for cell number and the percentage of budding cells. Cultures were routinely grown at 25"C, and aerated on a reciprocating water bath shaker. A more detailed review of the synchronization techniques has been published by WILLIAMSON (1964.).
DNA measurements: DNA was extracted from samples of approximately 2 x 108 whole cells according to the method of OCUR and ROSEN (1950). The cells were washed in cold 70% ethanol and a solution of 0.1% perchloric acid in 70% ethanol. They were extracted twice in a mixture of 3 parts of 95% ethanol to 1 part of ether at 65°C for 3 minutes. After two further washes in cold 0 . 2 ~ perchloric acid and cold I M perchloric acid, the nucleic acid was extracted in l~ perchloric acid at 70°C for 20 minutes. DNA was assayed colormetrically with the diphenylamine reagent (BURTON 1956). Highly polymerized calf thymus DNA from Mann Chemical Company served as a standard.
Ultrauiolet irradiation: Samples were removed at intervals during synchronous growth and resuspended in 7 ml of sterile water at a concentration of 5 x 106 to 1 x 1 0 7 cells per ml. The irradiations were performed in dim light in open 5 cm Petri dishes with two Hanovia germi- cidal lamps emitting 16 ergs per mm? per second at a target distance of 35 cm.
X-ray irradiation: Cells were spread on tbe surface of agar plates and irradiated at a distance of 19 cm with soft X rays delivered from a Picker control rectifier unit (60 kvp, 50 ma) and Machlett OEG-60 X-ray unit. The instrument, set at 50' kvp and 20 ma. delivered 72 roentgens per second at the surface of the agar plate.
The detection of recombinatiom T o detect intragenic recombination in diploid 265, cells were plated on mzdia lacking either lysine, tyrosine, or histidine. After several preliminary experiments it mas discovered that the detection of tyrosine recombinants could be dramatically improved if small amounts of tyrosine were added to otherwise tyrosineless media. Subse- quently, Z65 was assayed for tyrosine recombinants on media containing 2 pg/ml tyrosine. a level of tyrosine which permitted maximum recovery of tyrosine prototrophs. Intragenic recom- bination at the le,, met,,, tys, ly5, and ad, loci of strain XS380 was assayed on media lacking either leucine, methionine, tyrosine and phenylalanine, lysine, or adenine.
Recombination involving heterozygous loci was estimated by means of the adenine color system in yeast (ROMAN 1956), or by the Magdala Red auxotrophic staining technique by HORN and WILKIE (1966). Recombination of the ad, and ad, loci of strain Z65 was measured by the number of red/white sectored colonies observed when diploid cells were plated on com- plete medium, since homozygosis of ad, or ad, leads to the production of a red pigment. Magdala

194 ROCHELLE E. ESPOSITO
Red was used to detect recombination in diploid ZM, heterozygous for ly,, ty,, hi,, ur,, ad,, and ad,. Homozygosis was indicated by the formation of light pink/dark pink sectored colonies. The dark pink sectors contained prototrophic cells which were either heterozygous or homozygous dominant for all of the loci.
Three hundred to nine hundred prototrophic colonies of a particular type were counted for each estimate of intragenic recombination. Fifteen hundred to four thousand colonies were counted for each estimate of intergenic recombination. In experiments in which non-lethal doses of irradiation were employed, the prototroph frequencies were expressed as prototrophs per 106 cells plated. Since there was no loss in viability the number of cells plated was estimated from Coulter counts of approximately fifty thousand cells per sample. In experiments in which lethal doses of irradiation were used the prototroph frequencies were expressed as prototrophs per 106 viable cells. The number of viable cells was based on counts of approximately one thousand colonies per sample.
Tests for allelism in heteroallelic diploids: Tyrosine prototrophs derived from strain 265 were analyzed for the ly2, ty, , and hi, alleles that were present in 265. The prototrophs were sporu- lated on solid sporulation medium and the asci were dissected by micromanipulation. The segre- gants were crossed to tester stocks containing 1yZ-?, ty,-,, hi7-,, o r lyz-l, ty,_,, hi,-,. The diploids from these crosses were suspended in distilled water and drops containing approximately 106 cells were plated on lysineless, 2 pg/ml tyrosine, or histidineless media. The plates were irradiated with ultraviolet light for thirty seconds and incubated at 30°C for three to four days. Segregants that gave a low frequency of prototrophs (less than ten) when crossed to the tester
, 1
20 L
T i m e in m i n u t e s FIGURE 1 .-Cell growth, DNA synthesis and UV induced intragenic recombination in diploid
265. Growth medium: YMA. UV dose: 75 scconds/sample.

RECOMBINATION I N SYNCHRONIZED YEAST 195
strain on a given selective medium were scored as being homoallelic to the tester strain. Segre- gants that gave a high frequency (greater than thirty) were scored as being heteroallelic to the tester strain. Double mutant segregants homoallelic to both testers were not found.
Tests of prototrophic sectors from heterozygous diploids: Sectored colonies derived from strain 269 were streaked on Magdala Red medium. Single light pink colonies (heterozygous or homo- zygous dominant) were sporulated and asci dissected by micromanipulation. The segregants were tested for growth on lysineless, 2 pg/ml tyrosine, and histidineless media to determine if the prototrophic sector was heterozygous o r homozygous dominant at the Zy,, ty,, and hi, loci.
EXPERIMENTAL RESULTS
Intragenic recombination: Intragenic recombination was studied primarily in diploid Z65, heteroallelic at the Zy2, tyl, and hi7 loci on chromosome 11. Samples were removed from synchronously dividing cultures of this strain at ten minute intervals for approximately one and one half cell divisions. Each sample was irradiated with a constant non-lethal dose of ultraviolet light and plated to de- termine viability and the I'requency of intragenic recombination leading to lysine, tyrosine, or histidine prototrophy. The results are shown in Figure 1.
The degree of synchrony of the culture is measured by the increase in the
UJ
100.2 \ G
50 2 2 2 : [ * z o L ; r y 1 \
2 3 0 0 60 I20 180 240
Time in m inu tes
FIGURE 2.-Cell growth, DNA synthesis, and X-ray induced intragenic recombination in diploid Z65. Growth medium: YMA. X-ray dose: 1500 r/saniple.

196 ROCHELLE E. ESPOSITO
frequency of budding cells and the rise in cell number at the completion of the first cell division. Prototroph values are given as prototrophs per IO6 cells plated since there is no loss of viability.
The data indicate that ultraviolet light is more effective in inducing intragenic recombination at specific points in the cell cycle than at others. The frequency of ultraviolet-induced intragenic recombination rises before the initiation of DNA synthesis, reaches a peak, and declines as replication begins. The highest prototroph values, observed prior to replication are approximately three times greater than the lowest values observed after replication. The spontaneous fre- quencies remain constant and are approximately thirty times lower than the minimum ultraviolet-induced frequencies shown in Figure 1. These experiments have been repeated several times with the same results.
A similar set of experiments were performed using X rays instead of ultraviolet light as the inducing agent. The purpose was to determine whether the fluctua- tion in ultraviolet-induced recombination was associated with the particular
200[*v LYf
100
- -100 2 % b ~ d d ~ n g , , ~ - o - o ~ o ~ o ~ o - o a- P - 5 0 3
P Q
0 60 120 180 240 I 4 0 s -Ow---:
T i m e in minutes
FIGURE 3.-Cell growth, DNA synthesis, and UV induced intragenic recombination in diploid XS380. Growth medium: 4YMA. UV dose: 60 seconds/sample.
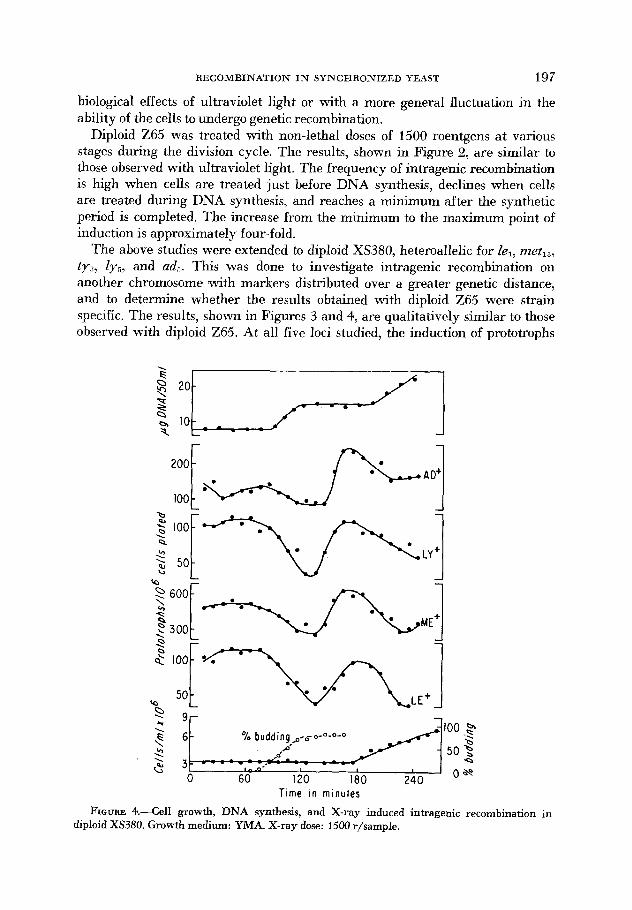
RECOMBINATION IN SYNCHRONIZED YEAST 197
biological effects of ultraviolet light or with a more general fluctuation in the ability of the cells to undergo genetic recombination.
Diploid Z65 was treated with non-lethal doses of 1500 roentgens at various stages during the division cycle. The results, shown in Figure 2, are similar to those observed with ultraviolet light. The frequency of intragenic recombination is high when cells are treated just before DNA synthesis, declines when cells are treated during DNA synthesis, and reaches a minimum after the synthetic period is completed. The increase Prom the minimum to the maximum point of induction is approximately four-fold.
The above studies were extended to diploid XS380, heteroallelic for le,, metls, ty3, ly,, and ad,. This was done to investigate intragenic recombination on another chromosome (with markers distributed over a greater genetic distance, and to determine whether the results obtained with diploid Z65 were strain specific. The results, shown in Figures 3 and 4, are qualitatively similar to those observed with diploid Z65. At all five loci studied, the induction of prototrophs
U
100.3 \F ' 5 3 Q
h budding p-o- 0-O-O-O
2 OS?
0 60 120 180 240 T i m e in minutes
FIGURE 4.--Cell growth, DNA synthesis, and X-ray induced intragehic recombination in diploid XS380. Growth medium: YMA. X-ray dose: 1500 r/sampIe.

198 ROCHELLE E. ESPOSITO
begins at a high level, declines during DNA synthesis, reaches a minimum and rises again as synthesis is completed. This relationship is the same as that ob- served in diploid Z65 despite a thirty-five minute difference in the generation time of the two strains.
Previous studies have indicated that intragenic recombination in mitotic and meiotic yeast cells is primarily non-reciprocal (ROMAN 1956, 1963; HURST and FOGEL 1964; FOGEL and HURST 1967). To determine whether the events under investigation in the present study were also non-reciprocal a sample of tyrosine prototrophs from the experiment shown in Figure 1 was analyzed with respect to the alleles present in the recombinant diploids (Table 1). The tyrosine locus was arbitrarily chosen for investigation. Nineteen prototrophs were examined from a high point of recombination induction (160 minutes of growth) and eighteen prototrophs from a low point (120 minutes of growth). The prototrophic diploids were sporulated and the asci dissected by micromanipulation. The geno- types of the segregants were inferred from the results of allelism tests.
The tyrosine-requiring segregants of each diploid were uniformly either ty,-, or tyl-2. No tyrosine dependent segregants of the double mutant genotype tyl-l, ty,-, were recovered. Assuming random assortment of centromeres this segregant would have been expected from half of the diploids in which tyrosine prototrophy
TABLE 1A
Analysis of ty+ recombinants at 160 minutes
lY2-2 tYl.2 hi7-1 lY2-1 ty1-1hi7-2
Z65
tY+ Spore A Spore B Spore C Spore D No. Zy t y hi ly zy hi Ly ty hi ZY t y hi
1 2-1 1-1 7-1 2-2 + 7-2 2-2 1-1 7-2 2-1 + 7-1 2' 2-2 + 7-1 2-1 + 7-1 2-1 1-1 7-1 2-2 1-1 7-1 3 2-1 1-1 7-2 2-2 + 7-1 2-2 + 7-2 2-1 1-1 7-1 4* 2-1 1-1 7-2 2-1 + 7-2 2-2 1-1 7-2 2-2 + 7-2 5 2-2 1-1 7-1 2-2 + 7-1 2-1 1-1 7-2 2-1 f 7-2 6 2-2 + 7-1 2-1 + 7-2 2-1 1-1 7-1 2-2 1-1 7-2 7 2-1 + 7-1 2-2 1-2 7-1 2-1 f 7-2 2-2 1-2 7-2 8 2-2 1-1 7-2 2-1 + 7-2 2-1 1-1 7-1 2-2 + 7-1 9 2-2 f 7-2 2-1 + 7-1 2-2 1-1 7-1 2-1 1-1 7-2
10 2-1 + 7-1 2-1 + 7-2 %2 1-2 7-1 2-2 1-2 7-2 11 2-2 1-2 7-1 2-1 1-2 7-2 2-1 f 7-1 2-2 + 7-2 12 2-2 + 7-1 2-1 f 7-2 2-2 1-1 7-2 2-1 1-1 7-1 13* 2-2 + 7-2 2-1 1-1 7-2 2-2 + 7-2 2-1 1-1 7-2 14 2-1 + 7-2 2-2 1-1 7-1 2-2 1-1 7-1 2-1 + 7-2 15 2-2 + 7-1 2-1 1-1 7-1 2-1 1-1 7-2 2-2 + 7-2 16 2-1 + 7-2 2-1 1-2 7-1 2-2 + 7-2 2-2 1-2 7-1 17 2-1 + 7-2 2-1 1-1 7-2 2-2 + 7-1 2-2 1-1 7-1
19 2-2 + 7-1 2-2 1-1 7-2 2-1 1-1 7-1 2-1 + 7-2 18 2-1 1-2 7-1 2-1 1-2 7-2 2-2 f- 7-2 2-2 + 7-1
* Homoallelic for histidine.

RECOMBINATION IN SYNCHRONIZED YEAST
TABLE 1B
Analysis of ty+ recombinants ut 120 minutes
199
I V + No.
Spore A Is t s hi
1 2 3 4 5 6 7'
9 IO* 11 12 13 14 15 16 17 18
a
2-2 1-2 7-2
2-1 + 7-2
2-1 1-1 7-1 2-2 1-2 7-1 2-2 + 7-1 2-1 + 7-1
2-2 1-2 7-2 2-1 1-2 7-2 2-1 f 7-2 2-2 + 7-1
2-2 1-1 7-1 2-2 + 7-1 2-2 1-1 7-1 2-2 1-1 7-1
2-2 + 7-1
2-2 1-1 7-1
2 4 + 7-1
2-2 + 7-2
2-1 + 7-1 2-1 + 7-1 2-2 1-2 7-2 2-1 1-2 7-2 2-1 + 7-2 2-2 1-2 7-1
2-2 + 7-1 2-1 + 7-2 2-1 1-1 7-2 2-2 + 7-2 2-1 + 7-1 2-2 1-1 7-2 2-2 1-2 7-2 2-1 + 7-1 2-1 + 7-2 2-1 1-1 7-1 2-1 1-1 7-1 2-2 + 7-1 2-2 1-1 7-1 2-1 1-1 7-2 2-2 + 7-2
2-2 + 7-1 2-1 1-2 7-1 2-2 1-2 7-2
2-2 + 7-2 2-2 1-2 7-2 2-1 1-2 7-1 2-1 1-2 7-2 2-1 + 7-2 2-2 + 7-2 2-1 + 7-2 2-2 + 7-1 2-2 1-2 7-1 2-1 1-1 7-2 2-2 + 7-1 2-2 1-1 7-1 2-1 + 7-1 2-1 1-1 7-2 2-2 1-1 7-2 2-1 1-1 7-2 2-1 7-1 2-2 1-1 7-1 2-1 1-1 7-2 2-1 + 7-1 2-2 + 7-2 2-1 1-1 7-2 2-2 1-1 7-2 2-1 + 7-1 2-1 1-1 7-2 2-2 + 7-1 2-1 + 7-2 2-1 + 7-2 2-1 1-1 7-2 2-2 + 7-1
* Homoallelic for histidine.
was the result of a reciprocal exchange at the tyrosine locus. These results show that the intragenic events observed in the present study are primarily non- reciprocal.
Intergenic recombination: Genetic exchange between genes, or genes and their respective centromeres, was studied in heterozygous diploid cells. In the two experiments shown in Figures 5 and 6, the diploid used was heterozygous for Zy2, tyl , hi,, ad,, ad,, and url. Cells 'were irradiated with non-lethal doses of either ultraviolet light (Figure 5 ) , or X rays (Figure 6), at intervals during synchronous growth, and plated on Magdala Red medium to stain the auxotrophic portion of sectored colonies. The total frequency of sectoring involving homozy- gosis at all of the loci is shown in the center portion of each graph. These data are strikingly similar to the intragenic recombination results described in Figures 1 through 4. Following an initial decline, the maximum sectoring frequency occurs just prior to or at the start of replication. The minimum occurs just after synthesis has been completed. The total sectoring frequency at all six loci varies in the ultraviolet experiment from approximately two percent at the low point to seven percent at the high point, and in the X-ray experiment from slightly less than one percent to four to five percent.
As a control, a sample of sectored colonies from high and low points of induc- tion, in the experiment shown in Figure 5 , was analyzed to determine whether the sectoring was caused by reciprocal or non-reciprocal mitotic recombination.

200 ROCHELLE E. ESPOSITO
.
Coulter counts
\ LILil 60 I20 I80 e o * 5 9 Time in minutes
FIGURE 5.-Cell growth, DNA synthesis, and UV induced intergenic recombination in diploid Z69. Growth medium YMA. UV dose: 60 seconds/sample. Sectors scored on Magdala Red com- plete medium to detect homozygosis at Zyz, tyl, hi,, UT1, ad, and ad,.
O F I
Percent budding
. o s
0 60 120 180 240 8
Percent budding 1oo.p
0-4 o s 0 60 120 180 240 8
1; - - - -(yxT*/={]f~O 2 . 5
Time in minutes
FIGURE 6.-Cell growth, DNA synthesis, and X-ray induced intergenic recombination in diploid Z69. Growth medium: YMA. X-ray dose: 1500 r/sample. Sectors were scared on Magdala Red complete medium to detect homozygosis at Zyz, ty l , hi,, UT,, ad,, and ad,.

RECOMBINATION IN SYNCHRONIZED YEAST 201
TABLE 2
Genetic analysis of seciored colonies resulting from inftergenic recombindon in diploid Z69
Class* Auxotrophic sector Prototrophic sector Number
R
N R
R
NR
R
N R
R
NR
R
NR
R NR
Total
lY2 tY, hi, lY, f Y , hi,
1Yz tY1 hi, LY, fY1 hi,
ty , hi, ty , hi,
ty, hi, ty , hi,
hi, hi,
hi, hi,
-
-
b 2 tY, IY, tY,
IYZ tY, l r , fY1
lY2 IY,
lY2 1Y 9
-
-
+ + + + + + + + + lY, Y 1 hi,
+ + + lY, + + + + + 1Yz tY, hi,
+ + + lY, Y 1 + + + + IY, tYl hi,
+ + + + + + + + + ly2 y 1 hi,
+ + + + t y , hi,
+ + + IY? ty , hi ,
11
3
14
0
7
7
2
0
1
2
35 12 47
* R = genotype of colony indicating reciprocal recombination. NR = genotype of colony indicating nx-reciprocal recombination.
The results are shown in Table 2. Among exchanges resulting in homozygosis for ly2, tyl, and hi,, eleven out of fourteen cases were due to reciprocal events between the 2yz locus and the centromere. Among those involving the ty, and hi, loci, all fourteen cases tested were the products of reciprocal exchange between lyz and ty,. Among sectors involving the hi, locus alone, half of the cases tested resulted from reciprocal exchange between ty, and hi,, the other half from non- reciprocal exchange at the locus. The reason for the increased proportion of non- reciprocal events among exchanges involving the hi, locus alone is not known.

202 ROCHELLE E. ESPOSITO
It has been suggested that in mitotic cells reciprocal exchange may be restricted in regions of the chromosome distal to the centromere leading to an apparent increase in the proportion of non-reciprocal events in these regions. From the data presented in Table 2, it was concluded that the majority of cases studied were the result of reciprocal mitotic crossing over. These results are in agr-. ,ement with other studies of ultraviolet and X-ray induced mitotic recombination (ROMAN personal commtmication; MORTIMER personal communication) .
The experiments cited above compare intragenic and intergenic recombination in different strains. To test the validity of the results intragenic and intergenic events were compared in the same strain utilizing diploid Z65. Intragenic re- combination was measured by the frequency of tyrosine prototrophy as pre- viously described. Since the diploid was also heterozygous for ad, and ad, inter- genic recombination was estimated from the frequency of homozygosis at these loci. Recombinants were readily detected as rcd/white sectored colonies since adl/adl and ad2/ad2 cells accumulate a red pigment (no attempt was made to distinguish the two types of red adenine requiring sectors). In the experiment shown in Figure 7, samples 'were treated with sixty seconds of ultraviolet light
L J
CO 2 8 1 . t.-Budd;n' /-Itt! 4 p-es-*
\ \ ---a' 2
\
$ 4 9,
9, 60 I20 180 240 c, Time in minutes
FIGURE 7.-Cell growth, DNA synthesis, UV induced intragenic recombination at fyl, and intergenic recombination at the ad, and ad, loci in diploid Z65. Growth medium: YMA. U\' dose: 60 seconds/sample.

RECOMBINATION IN SYNCHRONIZED YEAST 203
at fifteen minute intervals during synchronous growth. The pattern of ultra- violet-induced intragenic and intergenic recombination is similar to that pre- viously described.
Dosage response studies: The high induction of genetic recombination at par- ticular periods in the mitotic cell life cycle could be explained by changes in the radiosensitivity of cells during the division cycle. To test this possibility a syn- chronized culture of diploid Z65 was irradiated with lethal doses of ultraviolet light at a low and high point of recombination induction. The results, shown in Figure 8, indicate that the cells are in fact more sensitive to the lethal effects of ultraviolet light at the high point of recombination induction. If the variations in radiosensitivity which account for differential lethality during the cell cycle were directly responsible for the cycle in recombination frequency then one would expect the frequency of recombination to be the same at high and low points of
I I I I
I I 1 I I I I I I I I
!
Histidine'
150 min.
I U ~ L 0 I 2 3 4 5 0 I 2 3 4 50-
Minutes of UV
U loo I 2 3 4 5
Minutes of UV FIGURE 8.-UV dosage response of intragenic recombination and survival in diploid Z65.
Growth medium: YMA. Samples treated at 150 (A) and 105 (A) minutes of synchronized growth with 0-5 minutes of UV. indicates 50% survival.

204 ROCHELLE E. ESPOSITO
induction among cells compared at the same survival value. When recombination frequencies at these two times are compared at the same survival, however, the high point of recombination induction still yields a greater number of recom- binants than the low point. Thus, the lethal effects of irradiation do not appear to be the immediate cause of the fluctuation in recombination induction.
The recovery of intragenic recombinants: Suboptimal enrichment of the selec- tive media or post-irradiation incubation in nutrient medium prior to plating, both of (which permit additional cell division, do not eliminate the periodic fluc- tuation observed in the experiments described (Table 3) . Thus, it is unlikely that a differential recovery or expression of recombinants induced at different times in the cell cycle is responsible for the fluctuation in induced recombination.
The relationship of DNA synthesis to induced recombination: If the decline in induced recombination is obligatorily associated with the occurrence of DNA synthesis. inhibition of DNA synthesis should prolong the period of high induc- tion. The effect of FUDR, a known inhibitor of DNA replication (COHEN, FLAKS, BARNER, LOEB, and LICHTENSTEIN 1958), on a synchronously dividing culture of Z65 is shown in Figure 9. There is no effect on the first budding cycle regard- less of the time of addition of the inhibitor. The second budding cycle correspond- ing to the completion of the first cell division is temporarily inhibited when FUDR is added at 0, 20, 40, or 60 minutes of growth. Addition of FUDR at 80, 100, 120 or 140 minutes oE growth has progressively less effect on cell division indicating that between 60 and 100 minutes the cells have participated in an activity that permits them to divide for at least one additional division in the presence of FUDR. Previous studies have shown that DNA synthesis occurs dur- ing this interval in non-FUDR treated cultures (Figures 1 and 2). The results
TABLE 3
The recovery of UV induced intragenic recombinants after post-irradiation incubation in Y M A ~ ~ ~~~ ~~ ~ ~
Minut- of Prototrophs per 108 cells plated
of growth incubation In YMA lY+ W+ hi+ Minutes post irradiation
75 0 23 60 51 high 30 24 77 58
induction 60 25 65 56 90 38 70 76
120 41 66 74 255 22 55 85
120 low
induction
0 30 60 90
150 210
8 22 16 10 16 18 11 21 20 14 U) 19 14 20 22 14 15 16
~~~
Diploid Z65 was treated with 60 seconds of UV at 75 or 1 2 0 minutes of synchronized growth, resuspended on YMA and plated at intervals to detect recombination.

I -
o - . . . A - -
I - U 0 - n - O , , *
n f i U ”
120 min. 4 0 0 0
I - -
a
n 0
n - n n - n 100 min. 0
I - b
‘ Q
\
F \
(3“ * \
7
” - - e u v -
a n - O I
80 min
5- =
Y U U 0 ” 0 - - c U - 0
60 min I -
5- 4 1 - 40 min
O ~ O O O ~ ~ 0
- . U 0 0 0 - 0 0 0 0 0 ” O 0
5- “ 4
5- 4 , n n n - - * - - e - 0 - 0 , o] . . n - r r r 0 - o _
20 min I -
0
- I -
O
. “ O O - - * a - * -
0 -

ROCHELLE E. ESPOSITO
- FUDR
+ FUDR
-
T i m e i n minutes FIGURE 10.-X-ray, and X-ray + FUDR induced intragenic recombination in d i p l ~ d 265.
Growth medium: complete or complete + 1 mg/ml FUDR added at 0 minutes of growth. X-ray dose: 1500 r/sample. X-ray + FUDR curves are corrected for prototroph induction by FUDR alone.

R E C O M B I N A T I O N I N S Y N C H R O N I Z E D YEAST 207
therefore suggest that F'UDR added before 60 minutes inhibits the round of replication that usually accompanies the first budding cycle, and that this repli- cation is necessary for the completion of the first cell division. The effect of FUDR is a temporary one since cell division resumes at about 300 minutes.
The induction of lysine, tyrosine, and histidine prototrophy in strain Z65 by X rays alone, and by X rays in the presence of FUDR is shown in Figure 10. The latter curve has been corrected for the frequency of prototrophs formed in FUDR alone since FUDR also stimulates allelic recombination (ESPOSITO 1967). In the FUDR-treated culture the hequency of X-ray induced recombination remains high at the tyrosine and histidine loci for a period of time substantially past the time at which the frequency drops in a non-F'UDR medium. This is expected if DNA synthesis is associated with the decline in induced recombina- tion. However, X-ray induced lysine prototrophy is not maintained at a high level during the FUDR inhibition of DNA synthesis. The lysine locus may escape the FUDR inhibition and replicate before the other loci, from reserve pools of thymine. Alternatively, there may be more factors involved in reducing the susceptibility of cells to radiation-induced recombination than the initiation of DNA synthesis.
DISCUSSION
The purpose of this investigation was to determine whether intragenic and intergenic recombination could be separated by their pattern of induction during the mitotic cell life cycle of yeast.
Studies of X-ray and ultraviolet-induced recombination indicate that both types of exchange are maximally induced at similar points in the mitotic cell life cycle. The frequency of both intragenic and intergenic recombination is at a high level when cells are treated prior to the period of DNA replication. The frequency declines \when cells are treated during the period of replication and reaches a minimum when treatment is given at the completion of replication. The recombination cycle of yeast differs from the cycle described in Ustilago ( HOLLIDAY 1965, 1966) wherein both intragenic and intergenic recombination are maximally stimulated in cells undergoing DNA replication.
The failure to detect a significant difference in the pattern of intragenic and intergenic recombination (in yeast and Ustilago) may be interpreted in favor of one mechanism generating both events at particular stages of growth. Since other evidence (MITCHELL 1957; ROMAN and JACOB 1958; STADLER 1959a, 1959b; CATCHESIDE, JESSOP, and SMITH 1964; EBBS 1967; ROMAN personal communica- tion), suggests a dissociation of intragenic and intergenic recombination, it seems more plausible that both types of exchange share an event(s) in common which fluctuates during the mitotic cell cycle resulting in the patterns of recombination described above. The differences in the recombination cycles of yeast and Usti- lago may represent differences in the timing of these events during division. The fact that the point of maximal recombination induction is not obligatorily asso- ciated with DNA synthesis, indicates that DNA synthesis per se is not the basis of the cycle. While the pattern of recombination induction in the two organisms

208 ROCHELLE E. ESPOSITO
differs in relation to DNA replication, the recombination cycles are more con- cordant in their timing relative to ultraviolet sensitivity with respect to lethality. Both in yeast and in Ustilago, it has been found that periods of high recombina- tion induction coincide with periods of high sensitivity to the lethal effects of ultraviolet light.
The dosage response experiments reported here, demonstrate that the recom- bination frequency of cells from a point of low recombination induction is less than that of cells from J point of high induction even when these frequencies are compared at the same survival value. This result indicates that the changes responsible for increased recombination induction and lethality are coincident but not necessarily identical.
The pattern of induced recombination in yeast and Ustilago may reflect, in part, changes during the cell cycle in the activity or synthesis of enzymes asso- ciated with genetic recombination. In both yeast and bacteria (GORMAN, TAURO, LA BERGE, and HALVORSON 1964; MASTERS, KUEMPEL, and PARDEE 1964), it has been found that the synthesis of a number of enzymes occurs at discrete periods during the cell cycle. The timing of recombination events in yeast and Ustilago may similarily represent changes in the synthesis or activity of enzymes asso- ciated with recombination. Since dark repair enzymes have been implicated in the recombination process (CLARK and MARGULIES 1965; HOWARD-FLANDERS and THERIOT 1966), and in ultraviolet sensitivity regarding lethality ( HANAWALT 1966), both the fluctuation in recombination induction and in survival following irradiation of yeast and Ustilago may be due to periodic synthesis or activation of these dark repair enzymes during the cell cycle.
In the experiments reported here no reproducible polarity of recombination events proceeding sequentially along a chromosome arm, for any of the diploids investigated, was observed. This failure to detect a polarity in the timing of recombination events reflects a further difference in the recombination cycles of yeast and Ustilago. In Ustilago, HOLLIDAY (1965) has reported polarized inter- genic recombination proceeding sequentially along a chromosome arm. NO sim- ilar polarity of intragenic events has been observed (HOLLIDAY 1966). This addi- tional difference in the recombination cycles of yeast and Ustilago remains to be explained.
Previous studies have indicated a non-random coincidence of intragenic and intergenic recombination in asynchronous populations of yeast ( FOGEL and HURST 1963). In addition, WILKIE and LEWIS (1963), have observed non- random simultaneous homozygosis of several unlinked heterozygous loci. These results may be explained by the findings of the present study which indicate that intragenic and intergenic recombination are maximally induced at the same discrete period of the cell life cycle. An asynchronous population could be ex- pected to contain a subset of sensitive cells (which would show maximum induc- tion of both intragenic and intergenic recombination resulting in a non-random coincidence of events.
It is a pleasure to thank PROFFSSOR H. ROMAN for his guidance and interest during the course of this work. I would also like to express my sincere appreciation to DRS. M. S. ESPOSITO, R.
-

RECOMBINATION IN SYNCHRONIZED YEAST 209
HOLLIDAY, L. SANDLER, D. H. WILLIAMSON, D. C. HAWTHORNE, and D. R. STADLER for many helpful and stimulating discussions, and to MRS. J. MILLER and MRS. M. MILLS for excellent technical assistance.
SUMMARY
When synchronized cultures of Saccharomyces cerevisiae are irradiated dur- ing the mitotic cell life cycle with non-lethal doses of ultraviolet light or X rays to stimulate recombination, a characteristic induction pattern is observed. The ability of the cells to recombine in response to treatment rises or begins at a high level prior to DNA synthesis; the response is at a maximum at the start of replication, it declines during replication, and reaches a minimum just after the completion of replication. This cycle applies to the induction of both intragenic and intergenic recombination. Variations in the recoverability of recombinants or in the radiosensitivity of the cells with respect to lethality, do not appear to be a direct cause of the fluctuation in induced recombination. The initiation of DNA replication may be involved in the cycle, since the maximum intervals for recombination can be shifted and extended to a later time by inducing a tempo- rary delay in DNA replication with the inhibitor FUDR. There is no obvious polarity of recombination events proceeding sequentially along a chromosome arm during the mitotic cell cycle.
LITERATURE CITED
BURTON, K., 1956 A study of the conditions and mechanism of the diphenylamine reaction for
CATCHESIDE, D. G., A. P. JESSOP, and B. R. SMITH, 1964 Genetic controls of allelic recombina-
CLARK, A. J., and A. D. MARGULIES, 1965 Isolation and characterization of recombination-
COHEN, S . S., J. G. FLAKS, H. D. BARNER, M. R. LOEB, and J. LICHTENSTEIN, 1958 The mode of action of 5-fluorouracil and its derivatives. Proc. Natl. Acad. Sci. U.S. 44: 1004-1012.
EBBS, S., 1967 The effect of chemical mutagens on mitotic recombination in yeast. M.S. Thesis, University of Washington, Seattle.
ESPOSITO. R. E., 1967 Genetic recombination in synchronized cultures of Saccharomyces cere- uisiae. Ph.D. Thesis, University of Washington, Seattle.
FOGEL, S., and D. D. HURST, 1963 Coincident relations between gene conversion and mitotic recombination in Saccharomyces. Genetics 48: 321-328. __ 1967 Meiotic gene con- version in yeast tetrads and the thesry of recombination. Genetics. 57: 455-481.
Timing of enzyme synthesis during synchronous division in yeast. Biochem. Biophys. Res. Commun. 15: 43-49.
The UV sensitivity of bacteria: Its relation to the DNA replication cycle. Photochem. Photobiol. 5 : 1-12,
Induced mitotic crossing-over in relation to genetic replication in syn- chronously dividing cells of Ustilago maydis. Genet. Res. 6: 104-120. __ 1966 Studies on mitotic gene conversion in Ustilago. Genet. Res. 8 : 323-337.
Use of Magdala Red for the detection of auxotrophic mutants of Saccharomyces cereziisiae. J. Bacteriol. 91 : 1388.
the colorimetric estimation of deoxyribonucleic acid. Biochem. J. 62 : 315-322.
tion in Neurospora. Nature 202 : 1242-1243.
deficient mutants of Escherichia coli K12. Proc. Natl. Acad. Sci. U.S. 53: 451-459.
GORMAN, J., P. TAURO, M. LA BERGE, and H. 0. HALVORSON, 1964
HANAWALT, P. C., 1966
HOLLIDAY, R., 1965
HORN, P., and D. WILKIE, 1966

21 0 ROCHELLE E. ESPOSITO
HOWARD-FLANDERS, P., and L. THERIOT, 1966 Mutants of Escherichia coli K12 defective in DNA repair and genetic recombination. Genetics 53: 1137-1 150.
HURST, D. D., and S. FOGEL, 1964 Mitotic recombination and heteroallelic repair in Saccharo- myces cereuisiae. Genetics 50: 435-458.
MASTERS, M., P. L. KUEMPEL, and A. PARDEE, 1964 Enzyme synthesis in synchronous cultures of bacteria. Biochem. Biophys. Res. Commun. 15: 38-42.
MITCHELL, H. K., 1957 Crosssing over and gene conversion in Neurospora. In, The Chemical Basis of Heredity. Edited by W. D. MCELROY and B. GLASS. Johns Hopkins Press, Baltimore, pp. 94-113.
MITCHELL, M. B., 1955 Aberrant recombinant of pyridoxine mutants of Neurospora. Proc. Natl. Acad. Sci. U.S. 41 : 215-220.
MORTIMER, R. K., and D. C . HAWTHORNE, 1966 Genetic mapping in Saccharomyces. Genetics 53: 165-173.
OGUR, M., and G. ROSEN, 1950 The nucleic acids of plant tissues. The extraction and estimation of deoxypentose nucleic acid and pentose nucleic acid. Arch. Biochem. Biophys. 25 : 262-276.
PRITCHARD, R. H., 1960 Localized negative interference and its bearing on models of gene recombination. Genet. Res. 1 : 1-24.
ROMAN, H., 1956 Studies of gene mutation in Saccharomyces. Cold Spring Harbor Symp. Quant. Biol. 23: 111-118. - 1963 Gene conversion in fungi. In, Methodology in Basic Genetics. Edited by W. J. BURDETTE. Holden-Day, San Francisco, pp. 209-227.
A comparison of spontaneous and ultraviolet-induced allelic recombination with reference to the recombination of outside markers. Cold Spring Harbor Symp. Quant. Biol. 23: 155-160.
Gene conversion of cysteine mutants in Neurospora. Genetics 44: 647- 655. - 195913 The relationship of gene conversion to crossing over in Neurospora. Proc. Natl. Acad. Sci. U.S. 45: 1625-1629.
WILKIE, D., and D. LEWIS, 1963 Genetics 4.8: 1701-1716.
ROMAN, H., and F. JACOB, 1958
STADLER, D. R., 1959a
The effect of ultraviolet light on recombination in yeast.
Division synchrony in yeasts. In, Synchrony in Cell Division and
A rapid method for synchronizing division in the
WILLIAMSON, D. H., 1%
WILLIAMSON, D. H., and A. W. SCOPES, 1962 Growth. Edited by E. ZEUTHEN. Wiley, New York, pp. 351-360,589-591.
yeast Saccharomyces cereuisiae. Nature 193 : 256-257.