gene expression analyses of human mesenchymal stem …
TRANSCRIPT

J Oral Tissue Engin 2007;5(1) 35-47
Gene Expression Analyses of Human Mesenchymal Stem Cells Cultured in
Osteogenic Differentiation Medium for 3, 7, 14 and 21 Days by Genome Focus
DNA Microarray and Real -time PCR
Masayuki TAIRA1, Naoyuki CHOSA2, Kaori SASAKI1, Setsuo SAITOH1, Takashi NEZU1,
Nobuko SATO2 and Yoshima ARAKI1
1Department of Dental Materials Science and Technology, 2Department of Biochemistry,
Iwate Medical University School of Dentistry, Iwate, Japan. SYNOPSIS The purpose of this study was to evaluate gene expressions of human mesen-chymal stem cells (hMSC) cultured in osteogenic differentiation medium (OM)which contained ascorbic acid, β-glycerophosphate and dexamethasone for 0(control), 3, 5, 7, 14 and 21 days by 8.5k Genome Focus DNA microarray andreal-time PCR. It was confirmed by the DNA microarray analysis that 327 genes ofhMSC were significantly up-regulated by culture in OM especially at 14 and 21days while 156 genes were down-regulated. Up-regulated genes included os-teoblast-related genes such as secreted phosphoprotein 1 (osteopontin) gene andhypothetical protein expressed in osteoblast gene, along with angiogene-sis-related genes and cell cycle arrest-related genes, while down-regulated genescontained stroma-related genes and keratin-related genes. Expressions of severalosteogenic differentiation marker genes such as (down-regulated) osteonectingene and (up-regulated) BMP2 gene were also evaluated by real-time PCR. Geneexpression database identified here might contribute to dental tissue engineering. Key words: Mesenchymal stem cells, Osteogenic differentiation medium, Geneexpression, DNA microarray, Real-time PCR
ORIGINAL ARTICLE
INTRODUCTION In dental tissue engineering therapy, culture of mesenchymal stem cells (MSC) collected from iliac-crest-derived bone marrow is regarded as one im-portant clinical technique1 because MSC can be multiplied, seeded in bio-ab-sorbable scaffold materials and differentiated into osteoblasts2,3. Osse-ous defects could be restored using the ex vivo formed complex of scaffold and osteoblasts4.
Osteogenic differentiation of stem cells and osteoblastic cells has often been inducted by osteogenic differentia-tion medium (OM), namely, α-minimum essential medium supplemented with ascorbic acid, Na β-glycerophosphate and dexamethasone5,6. As for gene ex-pression by stimulation of OM, several osteogenic differentiation marker genes such as osteopontin and bone sialo protein were examined by RT-PCR or
35
TAIRA et al., Gene Expressions of hMSC in Osteogenic Medium
36
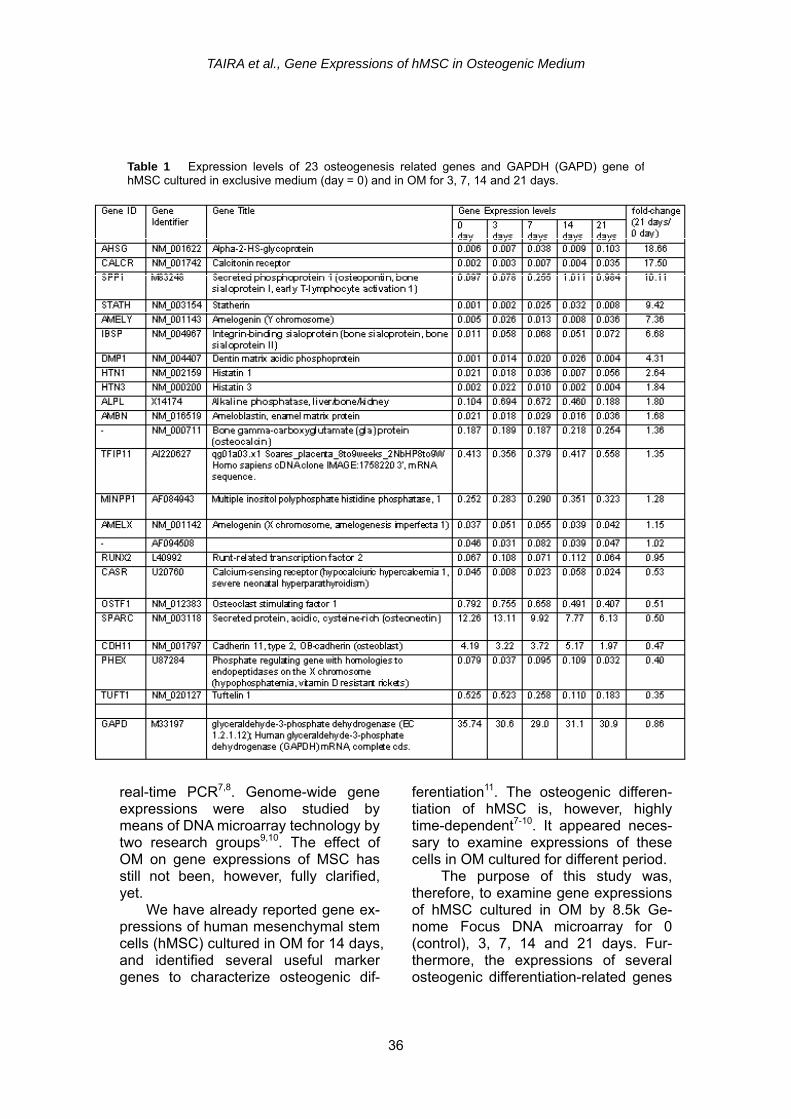
Table 1 Expression levels of 23 osteogenesis related genes and GAPDH (GAPD) gene ofhMSC cultured in exclusive medium (day = 0) and in OM for 3, 7, 14 and 21 days.
real-time PCR7,8. Genome-wide gene expressions were also studied by means of DNA microarray technology by two research groups9,10. The effect of OM on gene expressions of MSC has still not been, however, fully clarified, yet.
We have already reported gene ex-pressions of human mesenchymal stem cells (hMSC) cultured in OM for 14 days, and identified several useful marker genes to characterize osteogenic dif-
ferentiation11. The osteogenic differen-tiation of hMSC is, however, highly time-dependent7-10. It appeared neces-sary to examine expressions of these cells in OM cultured for different period.
The purpose of this study was, therefore, to examine gene expressions of hMSC cultured in OM by 8.5k Ge-nome Focus DNA microarray for 0 (control), 3, 7, 14 and 21 days. Fur-thermore, the expressions of several osteogenic differentiation-related genes

J Oral Tissue Engin 2007;5(1) 35-47
were evaluated and confirmed by real-time PCR. MATERIALS AND METHODS hMSC (PT-2501, Takara, Tokyo, Japan) (cell passage number = 5) were cultured in six polystyrene dishes 125mm in di-ameter (168351, Nunc, Rochester, NY, U.S.A.) with 20 mL exclusive α-minimum essential medium supple-mented with 10% exclusive mesen-chymal cell growth supplement (serum portion, PT-4106, Takara, Tokyo, Japan) and 1% penicillin/streptomycin (PT-3238, Takara, Tokyo, Japan) in a 5% CO2 in-cubator at 37℃. At sub-confluence, the cells (about 107 cells) were collected from one dish by a plastic cell scraper (as a control, date = 0 day) following twice cell wash with PBS (-) solution. The cells on five dishes were further cultured with fresh OM consisting of control medium and three additives such as 50 μg /mL L (+) ascorbic acid (012-04802, Wako Chemical, Osaka, Japan), 10mM Na β-glycerophosphate (Sigma Chemical, St. Louis, MO, U.S.A.) and 10-8 M dexamethasone (Sigma Chemical, St. Louis, MO, U.S.A.). After 3, 7, 14 and 21 days cul-ture with OM, the cells were collected by a cell scraper following twice cell wash with PBS (-) solution. For the entire cell cultures, the medium exchange was conducted every three days.
Total RNAs (at least 10 μg) were extracted from the cells using TRIZOL reagent (Invitrogen, Carlsbad, CA, U.S.A.). Genomic DNA was removed by DNase treatment (DNase I, 2215A, Ta-kara, Tokyo, Japan). A company (Ku-rabo, Osaka, Japan) conducted DNA microarray analyses; namely, reverse transcription, labeling, microarray hy-bridization, scanning and raw data analyses using a software (GeneSpring, GX, Agilent Technologies Japan, Tokyo, Japan). For the hybridization, six human
8.5k Genome Focus GeneChips (Affy-metrix, Santa Clara, U.S.A.) were used. The expressions of genes in the time course and in categories of biological functions were further analyzed by a web-based DNA microarray data analy-sis system (Genesifter Net, VizXlabs, Seattle, WA, U.S.A.).
Real-time PCR analyses were per-formed to quantitatively identify the amounts (copy numbers) of GAPDH, collagen 1A1 (type I collagen, α 1), os-teonectin (SPARC), osteopontin, os-teocalcin and BMP2 mRNAs of hMSC cultured in exclusive control medium (day 0) and OM for 3, 5, 7, 14 and 21 days, using a machine (Light Cycler, Roche Diagnostics, Tokyo, Japan) and primer/probe sets (410964 for GAPDH mRNA, 4651456 for collagen 1A1 mRNA, 4410688 for osteonectin mRNA, 4410530 for ostepontin mRNA, 4410521 for osteocalcin mRNA and 4391594 for BMP2 mRNA; Nippon Gene Research Lab., Sendai, Miyagi, Japan) mixed with FastStart DNA master hybridization probes (3 003 248, Light Cycler, Roche Diagnostics, Tokyo, Japan). The thermal heating (denature, amplification and cool) conditions followed the instructions of each primer/probe sets. RESULTS Table 1 shows expression levels of 23 osteogenesis-related genes and GAPDH (GAPD) gene of hMSC cultured in exclusive medium (day = 0) and in OM for 3, 7, 14 and 21 days. Well- known osteogenic differentiation-related genes such as alkaline phosphatase (ALPL) and osteonectin (SPARC) are included. The most right column ex-presses the fold change defined by the gene expression level at 21 days di-vided by that at 0 day. Early-stage os-teogenic differentiation marker genes such as ALPL and SPARC displayed the highest expression at 7 days, followed by gradual decline at 14 and 21 days.
37

TAIRA et al., Gene Expressions of hMSC in Osteogenic Medium
38
The medium-stage osteogenic differen-tiation gene such as secreted phospho-protein 1 (osteopontin) (SPP1) exhibited the highest expression at 14 days while the last-stage osteogenic differentiation marker gene such as bone gamma- carboxyglutamate (gla) protein (osteo-calcin) showed the highest expression at 21 days. With the addition of the cul-ture period in OM, the expression of osteoclast stimulating factor 1 (OSTF1) and Tuftelin 1 (TUFT1) declined. The expressions of other genes were quite low, or altering.
The medium-stage osteogenic differen-tiation gene such as secreted phospho-protein 1 (osteopontin) (SPP1) exhibited the highest expression at 14 days while the last-stage osteogenic differentiation marker gene such as bone gamma- carboxyglutamate (gla) protein (osteo-calcin) showed the highest expression at 21 days. With the addition of the cul-ture period in OM, the expression of osteoclast stimulating factor 1 (OSTF1) and Tuftelin 1 (TUFT1) declined. The expressions of other genes were quite low, or altering.
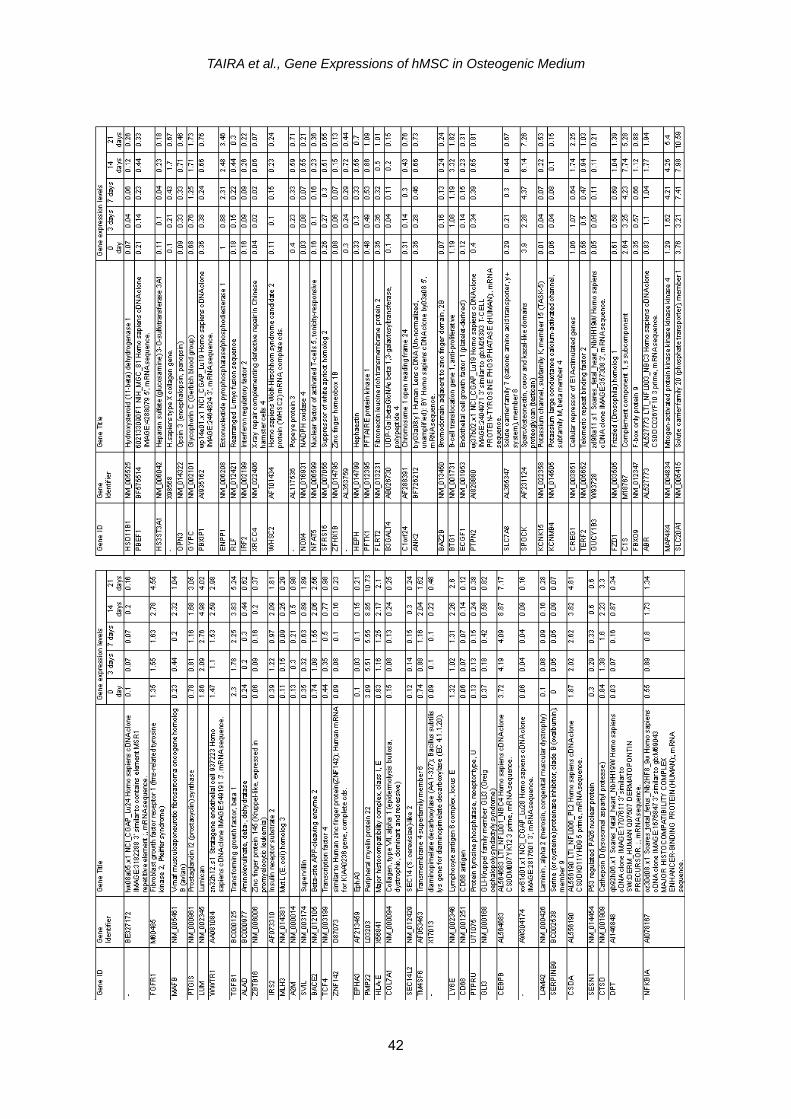
We also found 327 genes of hMSC whose expressions were significantly up-regulated by OM especially at 14 and 21 days (Table 2). Although many genes are listed, it was noted that OM caused hMSC to up-regulate (1) osteoblast- related genes such as secreted phos-phoprotein (osteopontin) (SPP1) gene and hypothetical protein expressed in osteoblast gene (IFI44L); (2) angio-genesis-related genes such as vascular endothelial growth factor B (VEGFB) gene and vascular cell adhesion mole-
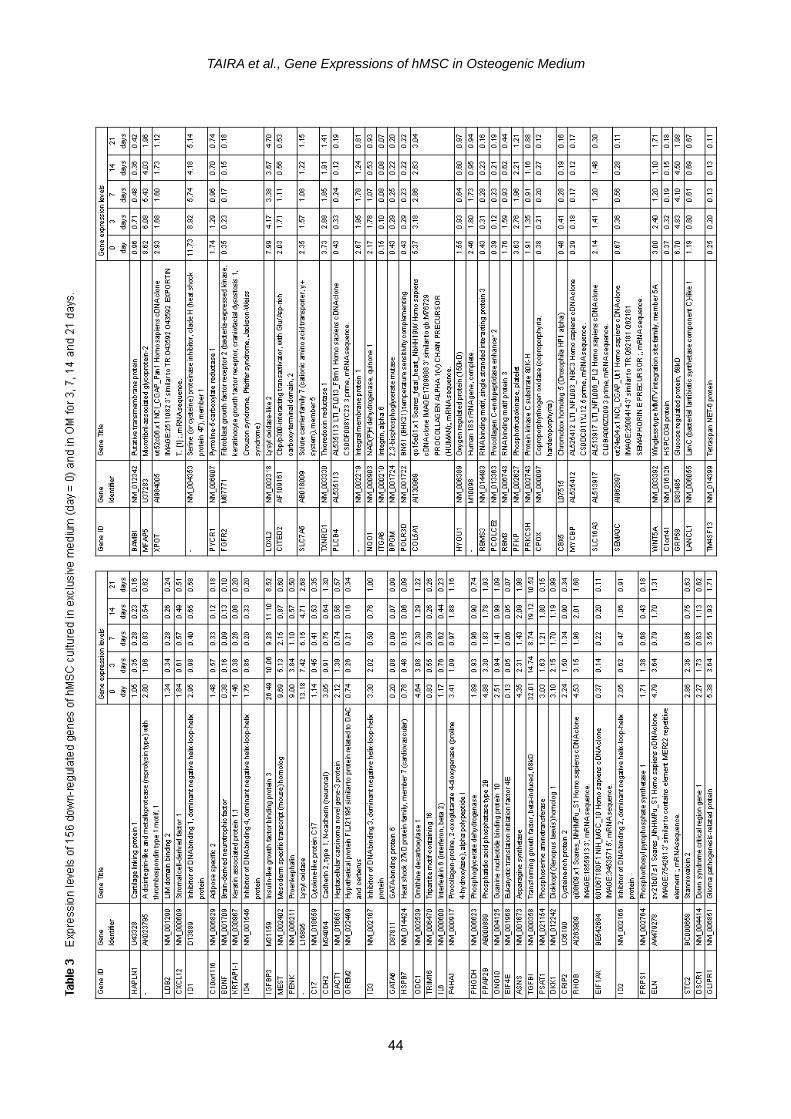
cule 1 (VCAM1) gene; and (3) cell cycle arrest genes such as growth ar-rest-specific 7 gene and cyclin- dependent kinase inhibitor 2A (CDKN2A) gene. Other interesting up-regulated genes included those of phospholipid transfer protein (PLTP), glycoprotein (transmembrane) nmb (GPNMB) and transforming growth fac-tor, beta 1 (TGFB1). On the other hand, we identified 156 genes whose expres-sions were significantly down-regulated especially at 14 and 21 days (Table 3). it was noticed that OM caused hMSC to down-regulate (1) stromal cell-related genes such as stromal cell-derived fac-tor (CXCL12) gene and bone marrow stromal cell antigen 1 (BST1) gene; and (2) keratin related genes such as keratin 18 (KRT18) gene and keratin associated protein 1.1 (KRTAP1-1) gene; and (3) antioxidant related genes such as NAD(P)H dehydrogenase, quinone 1 (NQO1) gene and catalase (CAT) gene. Other interesting down-regulated genes included those of cartilage linking pro-tein 1 (HPPLN1), adipose specific 2
(C10orf116), hyaluronan synthase 1 (HAS1) and cysteine-rich protein 1 (CRIP1). Many other up-regulated and down- regulated genes are listed in Tables 2 and 3, respec-tively, but they cannot be systematically related to the osteogenic differentia-tion in this study.
We also found 327 genes of hMSC whose expressions were significantly up-regulated by OM especially at 14 and 21 days (Table 2). Although many genes are listed, it was noted that OM caused hMSC to up-regulate (1) osteoblast- related genes such as secreted phos-phoprotein (osteopontin) (SPP1) gene and hypothetical protein expressed in osteoblast gene (IFI44L); (2) angio-genesis-related genes such as vascular endothelial growth factor B (VEGFB) gene and vascular cell adhesion mole-
cule 1 (VCAM1) gene; and (3) cell cycle arrest genes such as growth ar-rest-specific 7 gene and cyclin- dependent kinase inhibitor 2A (CDKN2A) gene. Other interesting up-regulated genes included those of phospholipid transfer protein (PLTP), glycoprotein (transmembrane) nmb (GPNMB) and transforming growth fac-tor, beta 1 (TGFB1). On the other hand, we identified 156 genes whose expres-sions were significantly down-regulated especially at 14 and 21 days (Table 3). it was noticed that OM caused hMSC to down-regulate (1) stromal cell-related genes such as stromal cell-derived fac-tor (CXCL12) gene and bone marrow stromal cell antigen 1 (BST1) gene; and (2) keratin related genes such as keratin 18 (KRT18) gene and keratin associated protein 1.1 (KRTAP1-1) gene; and (3) antioxidant related genes such as NAD(P)H dehydrogenase, quinone 1 (NQO1) gene and catalase (CAT) gene. Other interesting down-regulated genes included those of cartilage linking pro-tein 1 (HPPLN1), adipose specific 2
(C10orf116), hyaluronan synthase 1 (HAS1) and cysteine-rich protein 1 (CRIP1). Many other up-regulated and down- regulated genes are listed in Tables 2 and 3, respec-tively, but they cannot be systematically related to the osteogenic differentia-tion in this study.
Fig. 1 shows real-time PCR analyses results (sample copy number / GAPDH copy number) of five osteogenic differentia-tion-related genes of hMSC such as COL1A1 (type I collagen, α 1), SPARC (osteonectin), os-teopontin, osteocalcin and BMP2 when cultured in the
Fig. 1 shows real-time PCR analyses results (sample copy number / GAPDH copy number) of five osteogenic differentia-tion-related genes of hMSC such as COL1A1 (type I collagen, α 1), SPARC (osteonectin), os-teopontin, osteocalcin and BMP2 when cultured in the
Fig. 1 Real-time PCR analyses results (sample copynumber / GAPDH copy number) of five osteogenic differen-tiation-related genes of hMSC such as COL1A1 (type I col-lagen alpha 1), SPARC (osteonectin), osteopontin, osteo-calcin and BMP2 when cultured in the exclusive medium(day 0) and in osteogenic differentiation medium (OM) for 3,7, 14 and 21 days.

J Oral Tissue Engin 2007;5(1) 35-47
exclusive medium (day 0) and in OM for 3, 7, 14 and 21 days. It was pointed out that OM up-regulated BMP 2 gene at 14 and 21 days; little affected osteopontin and osteocalcin genes; and down-regulated COL1A1 and SPARC (osteonectin) genes with increasing the culture period of more than three days. As for osteopontin and osteocalcin genes, expression levels declined from 0 to 7 days in OM, but increased from 7 to 14 days in OM. DISCUSSION DNA microarray analyses using Ge-neChip provides a database from which many researchers can draw new ideas such as pathway analyses12. We wel-come such approach taken by other in-vestigators. There existed little informa-tion concerning genome-wide gene ex-pressions of hMSC when cultured in OM for different period up to 21 days9,10.
In this study, Genome Focus Ge-neChip tested the expression levels of about 8.5k (8500) genes but did not cover entire human genes (about 47k13). The repetition number of DNA microar-ray analysis was one at each condition (culture periods = 0, 3, 7, 14 and 21 days, respectively) due to the high cost of DNA microarray analyses conducted by a company. Withal such drawback, we still believe that results obtained here could contribute to dental tissue engineering. There is an opinion that gene expression data by GeneChip is quite reliable even when the test run is once14, although the test repetition is quite desirable.
hMSC is multi-potent, and can dif-ferentiate into several different pheno-types such as osteoblast, chondrocyte and adipocyte, etc15. As reported before, OM appeared to cause hMSC to pro-gressively induce osteochondral- ossification with angiogenesis11. There-fore, osteoblast-related genes gradually intensified (Table 2) while cartilage,
adipose and keratin related genes tended to slowly declined (Table 3). Osteogenic differentiation marker genes sequentially peaked in the manner that initial-stage osteogenic differentiation marker genes such as alkaline phos-phatase and osteonectin genes first peaked, followed by middle-stage os-teogenic differentiation marker gene such as osteopontin gene, while last-stage osteogenic differentiation marker gene such as osteocalcin peaked last7,8 (Table 1).
It appears that dentin matrix acidic phosphoprotein (DMP1) gene16 can be used as another osteogenic differentia-tion marker, which peaked at 14 days in OM culture (Table 1). In this study, its expression level is too low to make a conclusive statement, and research to verify this assumption is highly expected. Another new potential osteogenic dif-ferentiation marker gene was hypo-thetical protein expressed in osteoblast gene (IFI44L)17, which peaked at 14 days in OM with moderate signal inten-sities (Table 2). Research to confirm this idea is also highly probable.
Real-time PCR analyses (Fig. 1) further demonstrated that BMP2 gene18 could be an additional osteogenic dif-ferentiation marker gene. BMP2 gene was unfortunately contained in Genome Focus DNA microarray. Real-time PCR analyses (Fig. 1) succeeded in confirm-ing the down-regulation of SPARC (os-teonectin) of hMSC by OM, but failed in proving the up-regulation of osteopontin and osteocalcin genes of hMSC by OM. We plan to conduct DNA microarray analyses of hMSC in OM using com-plete human gene-labeled GeneChips; and to repeat real-time PCR analyses, especially those of osteopontin and os-teocalcin genes whose signals were too low in this study (Fig. 1). Larger sample (total RNA) amount might be helpful for next real-time PCR analyses.
39

TAIRA et al.,
Gene Expressions of hMSC in Osteogenic Medium
40

J Oral Tissue Engin 2007;5(1) 35-47
41
TAIRA et al., Gene Expressions of hMSC in Osteogenic Medium
42

J Oral Tissue Engin 2007;5(1) 35-47
43
TAIRA et al., Gene Expressions of hMSC in Osteogenic Medium
44

J Oral Tissue Engin 2007;5(1) 35-47
45

TAIRA et al., Gene Expressions of hMSC in Osteogenic Medium
46
Due to advance of osteochondra-
ossification with angiogenesis, it also appeared reasonable to see up-regulation of angiogenesis-related genes19, cell cycle arrest-related genes20 and transforming growth factor beta 1 gene12 in addition to up-regulation of osteogenesis-related genes, as mentioned in Results. Due to advance of osteochondral-ossification with angiogenesis, it seemed natural to observe down-regulation of other phe-notype-related genes such as stromal cell-related genes, keratin related genes, antioxidant related genes, cartilage linking protein 1 gene, adipose specific 2 gene, hyaluronan synthase 1 gene and cysteine-rich protein 1 gene, as mentioned in Results.
We cannot, however, refer to many other genes listed in Tables 2 and 3, which might have meaningful influence on osteogenic differentiation of hMSC caused by OM. We are looking forward other researchers to utilize the database presented here. Time-course analyses of expressions of hMSC genes cultured in OM appear to be quite useful for di-agnosis (recovery check) and clinical (gene therapy) treatment of damaged dental oral hard tissues in the future. It should be cautioned here that genes identified in this study considerably differ from those8,9 listed in pre-existing pa-pers except well-known genes such as alkaline phosphatase and BMP2 genes. ACKNOWLEDGEMENTS This study was supported in part by (1, 2) Grant-in-Aids (B) 18390522 and (Ex-ploratory Research) 19659511 by Japan Society for the Promotion of Science and (3, 4) Grant-in-Aids for High-performance Biomedical Materials Research from 2005 to 2009 and for the Open Research project from 2007 to 2011 from Ministry of Education, Culture, Sports, Science and Technology, Japan.
REFERENCES 1) Kinoshita Y, Kobayashi M, Hidaka T,
Ikada Y. Reconstruction of mandibular continuity defects in dogs using poly(L-lactide) mesh and autogenic par-ticuclate cancellous bone and marrow: preliminary report. J Oral Maxillofac Surg. 1997;55;718-723.
2) Gao J, Dennis JE, Solchaga LA, Awa-dallah AS, Goldberg VM, Caplan AI. Tissue-engineered fabrication of an os-teochondral composite graft using rat bone marrow-derived mesenchymal stem cells. Tissue Eng. 2001;7: 363- 371.
3) Donzeli E, Salvade A, Mimo P, Vigano M, Morrone M, Papagna R, Carini F, Zaopo A, Miloso M, Baldoni M, Tredici G. Mesenchymal stem cells cultured on a collagen scaffold: In vitro osteogenic differentiation. Arch Oral Biol. 2007;52: 64-73.
4) Yoshikawa T, Ohgushi H. Autogenous cultured bone graft-bone reconstruction using tissue engineered approach. Ann Chir Gynaecol. 1999;88:186-192.
5) Coelho MJ, Cabral AT, Fernandes MH. Human bone cell cultures in biocom-patibility testing. Part I.: osteoblastic differentiation of serially passaged hu-man bone marrow cells cultured in α-MEM and in DMEM. Biomaterials 2000;21:1087-1094.
6) Bielby RC, Boccaccini AR, Polak JM, Buttery LD. In vitro differentiation and in vivo mineralization of osteogenic cells derived from human embryonic stem cells. Tissue Eng. 2004;10:1518-1525.
7) Frank O, Heim M, Jakob M, Barbero A, Schafer D, Bendik I, Dick W, Heberer M, Martin I. Real-time quantitative analysis of human bone marrow stromal cells during osteogenic differentiation in vitro. J Cell Biochem. 2002;85:737-746.
8) Friedman M, Long MW, Hankenson KD. Osteogenic differentiation of human mesenchymal stem cells is regulated by bone morphogenetic protein-6. J Cell Biochem. 2006;98:538-554.
9) Doi M, Nagano A, Nakamura Y. Ge-nome-wide screening by cDNA mi-croarray of genes associated with ma-trix mineralization by human mesen-chymal stem cells in vitro. Biochem Biophys Res Commun. 2002;290:381- 390.
10) Kulterer B, Friedl G, Jandrositz A, San-chez-Cabo F, Prokesch A, Paar C, Scheideler M, Windhager R, Preisegger K-H, Trajanoski Z. Gene expression

J Oral Tissue Engin 2007;5(1) 35-47
profiling of human mesenchymal stem cells derived from bone marrow during expansion and osteoblast differentiation. BMC Genomics 2007;8;70 doi 10.1186 / 1471-2164 (15 pages) (Open Access Net artcle).
11) Taira M, Chosa N, Sasaki K, Saitoh S, Nezu T, Sato N, Araki Y. DNA microar-ray analyses of gene expression in human mesenchymal stem cells cul-tured in osteogenic differentiation me-dium for 14 days. J Oral Tissue Engin. 2005;3:25-53.
12) Zamurovic N, Cappellen D, Rohner D, Susa M. Coordinated activation of notch, Wnt, and transforming growth fac-tor-beta signaling pathways in bone morphogenic protein 2-induced osteo-genesis. Notch target gene Hey1 inhib-its mineralization and Runx2 transcrip-tional activity. J Biol Chem. 2004; 279: 37704-37715.
13) Schenke-Braun M, Corget JA. Expres-sion profiling using affymetrix genechip probe arrays. Methods Mol Biol. 2007;366:13-40.
14) Ogura N, Akutsu M, Tobe M, Sakamaki H, Abiko Y, Kondoh T. Microarray analy-sis of IL-beta-stimulated chemokine genes in synovial fibroblasts from hu-man TMJ. J Oral Pathol Med. 2007;36: 223-228.
15) Jackson L, Jones DR, Scotting P, Sottile V. Adult mesenchymal stem cells: dif-ferentiation potential and therapeutic applications. J Postgrad Med. 2007;53: 121-127.
16) Lu Y, Zhang S, Xie Y, Pi Y, Feng JQ. Differential regulation of dentin matrix protein 1 expression during odonto-genesis. Cells Tissues Oragans 2005; 181:241-247.
17) Toukap AN, Galant C, Theate I, Mau-doux AL, Lories RJ, Houssiau FA, Lauwerys BR. Identification of distinct gene expression profiles in the syno-vium of patients with systemic lupus erythematosus. Arthritis Rheum. 2007; 56:1579-1588.
18) Yamagiwa H, Endo N, Tokunaga K, Hayami T, Hatano H, Takahashi HE. In vivo bone-forming capacity of human bone marrow-derived stromal cells is stimulated by recombinant human bone morphogenetic protein-2. J Bone Miner Metab. 2001;19:20-28.
19) Deckers MM, Karperien M, van der Bent C, Yamashita T, Papapoulos SE, Lowik CW. Expression of vascular endothelial growth factors and their receptors dur-ing osteoblast differentiation. Endocri-nology 2000;141:1667-1674.
20) Ogasawara T, Kawaguchi H, Jinno S, Itaka K, Takato T, Nakamura K, Oka-yama H. Bone morphogenetic protein 2-induced osteoblast differentiation re-quires Smad-mediated down-regulation of Cdk6. Mol Cell Biol. 2004;24:6560- 6568.
(Received, August 27, 2007/ Accepted, September 25, 2007)
Corresponding author: Masayuki TAIRA, Ph.D. Department of Dental Materials Science and Technology, Iwate Medical University School of Dentistry, 1-3-27, Chuo-dori, Morioka, Iwate, Japan, 020-8505 Tel: +81-19-651-5111(Ext4217) Fax:+81-19-651-8407 Email: [email protected]
47