gelechiidae moths are capable of chemically dissolving the pollen
TRANSCRIPT

Gelechiidae Moths Are Capable of Chemically Dissolvingthe Pollen of Their Host Plants: First DocumentedSporopollenin Breakdown by an AnimalShixiao Luo1, Yongquan Li1, Shi Chen1, Dianxiang Zhang1*, Susanne S. Renner2*
1 Key Laboratory of Plant Resource Conservation and Sustainable Utilization, South China Botanical Garden, The Chinese Academy of Sciences, Guangzhou, China,
2 Department of Biology, University of Munich (LMU), Munich, Germany
Abstract
Background: Many insects feed on pollen surface lipids and contents accessible through the germination pores. Pollenwalls, however, are not broken down because they consist of sporopollenin and are highly resistant to physical andenzymatic damage. Here we report that certain Microlepidoptera chemically dissolve pollen grains with exudates from theirmouthparts.
Methodology/Principal Findings: Field observations and experiments in tropical China revealed that two species ofDeltophora (Gelechioidea) are the exclusive pollinators of two species of Phyllanthus (Phyllanthaceae) on which their larvaedevelop and from which the adults take pollen and nectar. DNA sequences placed the moths and plants phylogeneticallyand confirmed that larvae were those of the pollinating moths; molecular clock dating suggests that the moth clade isyounger than the plant clade. Captive moths with pollen on their mouthparts after 2-3 days of starvation no longer carriedintact grains, and SEM photographs showed exine fragments on their proboscises. GC-MS revealed cis-b-ocimene as thedominant volatile in leaves and flowers, but GC-MS analyses of proboscis extracts failed to reveal an obvious sporopollenin-dissolving compound. A candidate is ethanolamine, which occurs in insect hemolymphs and is used to dissolvesporopollenin by palynologists.
Conclusions/Significance: This is the first report of any insect and indeed any animal chemically dissolving pollen.
Citation: Luo S, Li Y, Chen S, Zhang D, Renner SS (2011) Gelechiidae Moths Are Capable of Chemically Dissolving the Pollen of Their Host Plants: FirstDocumented Sporopollenin Breakdown by an Animal. PLoS ONE 6(4): e19219. doi:10.1371/journal.pone.0019219
Editor: Edward Newbigin, University of Melbourne, Australia
Received February 15, 2011; Accepted March 22, 2011; Published April 28, 2011
Copyright: � 2011 Luo et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricteduse, distribution, and reproduction in any medium, provided the original author and source are credited.
Funding: This study was supported by the Chinese Academy of Sciences (Grant SCBG-200734), the National Science Foundation of China (Grant 30700044,30870367), and the Natural Science Foundation of Guangdong Province (Grant 7301182). The funders had no role in study design, data collection and analysis,decision to publish, or preparation of the manuscript.
Competing Interests: The authors have declared that no competing interests exist.
* E-mail: [email protected] (SSR); [email protected] (DZ)
Introduction
Obligate mutualisms are often supported by unique adaptations
that facilitate specific interactions between the partners. Well-
known examples are found among plants and their pollinators,
and a striking number of the most tightly co-evolved interactions
involve moths. Examples include yucca flowers and yucca moths
[1], senita cacti and senita moths [2], and Phyllantheae flowers
and Epicephala moths [3,4]. In each of these mutualisms, special
behaviors or mouthparts evolved, resulting in efficient transport of
pollen to stigmas. None of these moths feed on pollen. Moths that
actively ingest pollen belong to relatively old families with biting-
chewing mouthparts capable of crushing the walls of spores or
pollen grains, such as Agathiphagidae, Heterobathmiidae, and
Micropterigidae [5,6,7]. Higher Lepidoptera instead have mouth-
parts adapted for fluid feeding, and none are known to regularly
ingest pollen except species of Heliconius (including Laparus), which
digest pollen contents (but not pollen walls) in a mix of saliva and
nectar [8,9,10].
The reason Lepidoptera and other insects, including bee larvae
who obligately feed on pollen, all void the walls of pollen and
spores intact is that these walls consist of sporopollenin, a complex
polymer that is composed of fatty acids and phenolic compounds.
The precise structure of sporopollenin is unknown [11]. Sporo-
pollenin is resistant to nonoxidative physical, chemical, and
biological treatments and insoluble in both aqueous and organic
solvents [12,13]. This chemical inertness accounts for the
preservation of pollen grains for tens to hundreds of millions of
years [14]. There is also an extensive fossil record of insects gut
contents containing pristine pollen [15]. Yet spores and pollen
contain up to 61% protein [16] and are an important food source
for many animals whose gut enzymes enable them to digest
substances on pollen surfaces, such as pollenkitt, as well as pollen
contents, accessible through the spores’ germination pores.
Here, we report that certain gelechiid moths in the genus
Deltophora are capable of chemically dissolving the pollen walls’
sporopollenin. This is the first record of the chemical breakdown
of spore walls by any eukaryote. The Deltophora moths have highly
specialized proboscises (c. 3 mm long) and are the only pollinators
of two unrelated species of Phyllanthus in the Phyllanthaceae, a
family comprising 1750 species in 59 genera (1270 in Phyllanthus),
several with intricate phenology and pollinator interactions [4,17].
PLoS ONE | www.plosone.org 1 April 2011 | Volume 6 | Issue 4 | e19219

We carried out field observations on the Deltophora/Phyllanthus
mutualism at several sites in tropical China, with one population
studied over four years to document the phenological fit between
the partner’s life cycles and to verify that the plants had no other
pollinators.
Molecular sequence data were used to verify that moth larvae
feeding on host leaves were those of the pollinating adults and to
infer the phylogenetic placements of the two moth species and the
two plant species, given that all or in large and taxonomically
poorly known groups. The DNA data also permitted molecular
clock-based estimates of divergence times for the plant and insect
partners and thus to test whether they could have interacted over
much of their evolution. Finally, we used GC-MS to assess
whether the moths might find their oviposition sites (i.e., young
leaves of the host) and adult foraging sites (i.e., the flowers of the
host) by a specific odor. The chemical breakdown of the pollen
walls was documented photographically, using freshly caught,
instantly killed moths and moths caught at flowers but then held
captive and starved for 2–3 days. The sporopollenin-dissolving
compound exuded by the moth proboscises could not be identified
from dead moths, but studies of insect hemolymphs suggest at least
one common candidate chemical.
Results
Plant and moth ecology and behaviorDuring the 300 h of observations (at seven study sites in China
and Vietnam; Materials and Methods), the only visitor and
pollinator of Phyllanthus cochinchinensis was Deltophora sp. 1. During
the 200 h of observations of the Hainan-endemic P. rheophyticus,
the only flower visitor and pollinator was Deltophora sp. 2. No other
insects visited the flowers. Beginning in the early evening and
continuing until midnight, Deltophora moths would visit male
flowers to forage for pollen and nectar, while female flowers were
visited only for nectar (Fig. 1C and D; Fig. 2C and D). Moths
switched between flowers and plants and were also observed laying
their eggs on young leaves. Flower visits lasted 45–60 seconds
(N = 10), with single flowers being probed about 10 times, meaning
a moth would uncurl and curl its proboscis and insert it into the
same flower (probing of male flowers 10.462.7 SD; probing of
female flowers 9.662.3 SD; each with N = 5). Male and female
moths carried pollen grains on their proboscises (Fig. 1F; Fig. 2F),
and pollen grains under the microscope could unambiguously be
identified as coming from the host plants. Cross-pollination is very
effective as shown by the high fruit set in the dioecious species, P.
cochinchinensis (Supporting Information S1). The male flowers
typically produce 840062141 pollen grains (data for P. cochinch-
inensis), and the pollen of both Phyllanthus species is rich in lipids but
lacks starch.
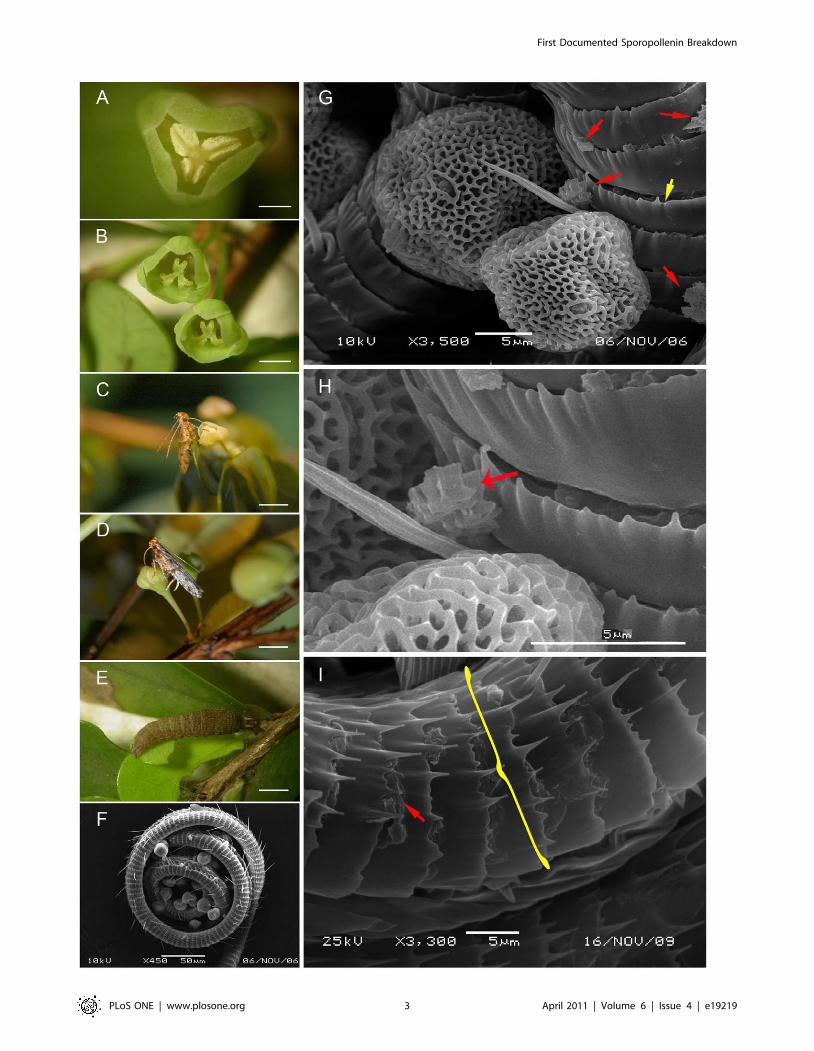
Pollen walls dissolved by a secretion from the Deltophoraproboscises
The proboscises of instantly killed Deltophora were c. 3 mm long
(N = 20 for sp. 1; both species had similar body and proboscis sizes)
and were covered with pollen grains (Fig. 1F; 2F). Their structure
appears to be unique in Gelechioidea [18; L. Kaila, Senior
Curator, Finnish Museum of Natural History, personal commu-
nication, 11 Nov. 2009]: It consists of curved integument casings
that surround the proboscis (Fig. 1G–I; Fig. 2G–K). Remains of
partially dissolved pollen grain walls on the proboscises of freshly
killed moths (N = 40) are marked by red arrows in Fig. 1G–I as
well as Fig. 2G–I. Moths caught on flowers (23 moths from P.
cochinchinensis and 21 from P. rheophyticus) and then starved for 2–3
days inside large plastic bottles no longer had any intact grains on
their proboscises. Pollen fragments lying between the circular
integument casings suggest that the sporopollenin-dissolving
chemical is produced between these casings. Older Deltophora
specimens in our collection have brittle proboscises, perhaps
because the corrosive action of this chemical attacks the proboscis
over time, in spite of its strongly armored surface.
Temporal fit between moth and plant life cyclesThe life cycles of both moth species were closely linked to the
phenology of their plant hosts, which appear to be the moths’ only
larval food plants. Experimental rearing of moth larvae in 2006,
2007, and 2008 succeeded on their host’s leaves. Adult Deltophora
appeared in mid-May and immediately mated. During May, June,
and July, moths could regularly be seen visiting flowers. No adult
moths were found before or after the hosts’ flowering seasons. The
youngest instar larvae fed internally on young leaves before
emerging on the lower leaf surface. Older caterpillars reached 8–
10 mm in length (Fig. 1E and 2E) and also fed only on young
leaves. Larvae fed only at night; during the day they rested on
lower leaf surfaces. The characteristic feeding damage is shown in
Fig. 1E and Fig. 2E and the extent of leaf damage in Supporting
Information S1.
Caterpillars of Deltophora sp. 1 placed on the leaves of Glochidion
eriocarpum (Phyllanthaceae) did not feed. On their host leaves,
however, larvae developed and pupated normally and eclosed
after 30 to 36 days. A few instead entered a 9–11 month-long
pupal diapause (under laboratory conditions) from which adults
emerged simultaneous with the next flowering period of their plant
host.
Molecular phylogenetics and divergence time estimatesfor the plant and moth species
Of the c. 1270 described species of Phyllanthus, 130 have been
sequenced for matK. In a maximum likelihood tree from these
sequences, the two Phyllanthus species studied here are only
distantly related, and a relaxed molecular clock calibrated with
Phyllanthus fossils suggests that their most recent common ancestor
lived 55 (38–54) Ma ago (Fig. 3).
For the moths, a maximum likelihood tree of 29 cytochrome c
oxidase subunit I (COI) sequences, including sequences of
Tegeticula and Galagete for calibration purposes, other Gelechioidea,
and four sequences representing the two Deltophora species, showed
that the studied species clearly belong in the Gelechiidae and that
the larvae represented the same species as the adult moths (larval
sequences are not included in Fig. 4 because redundant sequences
were removed before the clock analyses; Materials and Methods).
A relaxed clock model (Fig. 4) suggested a divergence time for the
Deltophora species of 16 (29–4) Ma. This estimate is highly
preliminary because of the extreme molecular under-sampling of
Gelechioidea and the great genetic distances of the two calibration
points. Nevertheless, the distant phylogenetic relationships of the
two Phyllanthus species and the much younger divergence time of
the moths (Figs. 3 and 4) together reject a hypothesis of co-
speciation.
Results of the GC-MS analysesDetails of the GC-MS method are given in Materials and
Methods. The dominant compound detected in fresh P.
cochinchinensis flowers and leaves was 1,3,6-octatriene (also called
cis-b-ocimene). Other compounds were present in only small
amounts and could not be identified. No unusual compounds
were present in methanol or chloroform extracts of moth
proboscises.
First Documented Sporopollenin Breakdown
PLoS ONE | www.plosone.org 2 April 2011 | Volume 6 | Issue 4 | e19219
First Documented Sporopollenin Breakdown
PLoS ONE | www.plosone.org 3 April 2011 | Volume 6 | Issue 4 | e19219

Discussion
Phyllanthus cochinchinensis and Deltophora sp. 1, and P. rheophyticus
and Deltophora sp. 2 are two pairs of obligate mutualists. No other
insects visited the minute flowers during several hundred hours of
observations, and the moths’ entire life cycle, from the larvae
feeding on the young leaves of their hosts to the adults emerging
synchronously with flowering in the hosts, is adapted to their host
plants’ phenology and chemistry. That cis-b-ocimene is the
dominant scent compound in both, the leaves and the flowers
(of P. cochinchinensis; the other species was not analyzed), suggests
that this is the key signal for host selection. Cis-b-ocimene has also
been found in the female flowers of the Phyllanthaceae Breynia vitis-
idaea [19], and is common in floral scents.
The genus Deltophora belongs to the Gelechiidae, a species-rich
group of Lepidoptera that may not be monophyletic [20]. Most of
the c. 870 genera of Gelechiidae are poorly defined, and the genus
Deltophora, originally described from South Africa and supposedly
containing 19 species on all continents except Antarctica, is no
exception [21]. Prior to our observations, host plants of larvae and
adults were unknown [21]. Without denser species sampling, it is
not possible to infer how long the Deltophora moths may have
interacted with their current Phyllanthus hosts, which are
unexpectedly distantly related to each other as shown by the
molecular phylogeny (Fig. 3).
This is the first case of an animal chemically dissolving spore
walls, i.e., the sporopollenin in the pollen exine. Among the few
substances known to dissolve sporopollenin is ethanolamine, also
called 2-aminoethanol or monoethanolamine, which palynologists
use to remove sporopollenin [12,13]. Ethanolamine is an
extremely volatile organic compound. It is an abundant head
group for phospholipids, substances found in biological mem-
branes, and is present in insect hemolymphs, usually as
phosphatidyl-ethanol-amine. Ethanolamine is among the defense
secretions exuded as droplets from glandular hairs by the pupa of
the Mexican bean beetle, Epilachna varivestis [22,23]. Given its
known ability to dissolve sporopollenin, ethanolamine is a possible
candidate for the unknown chemical produced by Deltophora.
Chemical investigation of the compounds responsible for the spore
wall breakdown will require the GC-MS analysis of freshly killed,
perhaps even living moths; our analyses using methanol or
chloroform extracts of several month-old proboscises yielded no
unusual compounds.
Pollen feeding in other groups of Lepidoptera is entirely
different than that in the gelechiid Deltophora studied here. Thus,
Heliconius (including Laparus), a group of nymphalid Lepidoptera,
produce saliva that has proteolytic activity [10,24], and light
microscopy has revealed that pollen grains exposed to the saliva
for .200 min had modified ‘‘exines,’’ which has been attributed
to proteolysis [25]. Sporopollenin, however, does not consist of
proteins (nor do other spore wall layers). The proboscises of
Heliconius butterflies bear long sensory bristles on which pollen is
accumulated [7,8]. They are entirely different from those of the
Deltophora moths studied here. Just like these moths, however,
Heliconius repeatedly insert and retract their proboscis when
visiting single flowers, and it was this behavior that first alerted
us to the moth’s active up-take of pollen grains, not just nectar
from the Phyllanthus flowers. The Deltophora larval stage lasts 30–36
days, which is relatively short; we do not know whether the moth’s
diet is supplemented with compounds from the pollen walls, but
this could be tested with radioactive labeling.
The discovery of two new obligate plant/pollinator interactions,
involving the hereto unknown complete breakdown of spore walls,
shows how little is known about arthropod chemical interactions
with their environment. Given this limited knowledge and the
ubiquity of spore walls in nature, it seems unlikely that Deltophora
moths should be the only arthropods capable of dissolving
sporopollenin.
Materials and Methods
Plant species, study sites, and study durationPhyllanthus cochinchinensis (Loureiro) Sprengel (vouchers Luo 248
and 249, deposited in the herbaria IBSC and M) is a shrub
(Fig. 2A) that occurs in India, China, Cambodia, Laos, and
Vietnam, usually in low vegetation or along forest edges at 200–
600 m alt. The species is dioecious, with axillary inflorescences.
Male flowers are borne in fascicles, but usually open one at a time.
Each has three stamens that dehisce longitudinally (Fig. 1A).
Female flowers are also borne in fascicles and have three bifid
recurved styles (Fig. 1B). Both sexes offer nectar. Phyllanthus
cochinchinensis was studied at seven sites in China and Vietnam
(precise localities and duration of observations are given in the
Supporting Information S1). Study sites were 7 to 800 km apart
from each other.
Phyllanthus rheophyticus M. G. Gilbert & P. T. Li also is a small
shrub, but different from P. cochinchinensis it is monoecious. The
species is endemic to Hainan Island [26] where is grows along
riverbeds (Fig. 2B). The axillary bisexual inflorescences usually
produce two to four male flowers and one female flower. Flowers
are similar to those of P. cochinchinensis, and again both sexes offer
nectar. Phyllanthus rheophyticus was studied at three sites on Hainan
Island.
From 2006 to 2010, observations where carried out between
May and September each year, which are the only months during
which the two Phyllanthus species produce flowers. Overall, flower
visitors and leaf feeding larvae were studied for c. 300 h in P.
cochinchinensis and for c. 200 h in P. rheophyticus, mostly at night but
occasionally also during the day. At two study sites, flowering
shrubs of P. cochinchinensis (30 staminate and 25 pistillate
individuals) and P. rheophyticus (30 monoecious individuals) were
monitored weekly during the entire reproductive season from May
to September.
Nectar production was assessed both, during the day and at
night, using 10 marked flowers of each sex on five plants of each
species. Marked flowers were covered with fine netting to exclude
nectar foragers, and nectar was sampled at 0800 h, 1200 h,
1600 h, 1900 h, 2100 h, and 2300 h. Pollen grains were tested for
the presence of lipids or starch by staining in either sudan III or
iodine (IKI) solutions, followed by examination under a micro-
Figure 1. Deltophora sp. 1 moths and Phyllanthus cochinchinensis flowers and leaves photographed in the field (A–E) or under theSEM (F–I). A. Male flower with dehisced anthers. B. Female flowers with the bifid stigmas. C. Deltophora sp. 1 moth drinking nectar from a maleflower. D. Deltophora sp. 1 moth drinking nectar from a female flower. E. Larva of Deltophora sp. 1 on a young leaf, the characteristic feeding damageis visible along the lower leaf margin. F. Pollen-coated proboscis from an instantly killed Deltophora sp. 1 moth; note that all pollen grains are intact.G. Intact and broken P. cochinchinensis pollen grains on a proboscis. Red arrows indicate exine fragments. The yellow arrow points to one of the rings(casings) sheathing the proboscis. H. An almost completely dissolved pollen wall (red arrow). I. The proboscis-surrounding casing (yellow bracket) of amoth killed following a few days of starvation. The red arrow points to a pollen wall fragment.doi:10.1371/journal.pone.0019219.g001
First Documented Sporopollenin Breakdown
PLoS ONE | www.plosone.org 4 April 2011 | Volume 6 | Issue 4 | e19219

Figure 2. Habits of the focal plant species and Deltophora sp. 2 moths interacting with Phyllanthus rheophyticus. A Habit of Phyllanthuscochinchinensis. B. Habit of P. rheophyticus. C. Deltophora sp. 2 drinking nectar from a male flower of P. rheophyticus. D. Deltophora sp. 2 visiting afemale flower of P. rheophyticus. E. Larva of Deltophora sp. 2 on young leaves of P. rheophyticus; the arrow points to the feeding damage. F. Pollen-coated recoiled proboscis of Deltophora (the arrow points to the pollen). G and H. Dissolved pollen wall fragments (red arrows) on the proboscis of aDeltophora moth killed after two days of starvation. I. Nearly dissolved sporopollenin (red arrow) and the unique integument casing (yellow brackets)on the proboscis of a Deltophora sp. 2 moth killed by starvation. J. The blue arrow marks one of the characteristic casings on the upper part of the
First Documented Sporopollenin Breakdown
PLoS ONE | www.plosone.org 5 April 2011 | Volume 6 | Issue 4 | e19219

scope. Dimethylthiazol-diphenyl-tetrazolium bromide (MTT) was
used to assess stigma receptivity. Experimental investigation of the
plants’ mating systems is described in the Supporting Information
S1.
Moth species and their behaviorMoths that visited the plants’ flowers or leaves were collected for
identification and examination of their pollen loads under a
stereoscope (Stemi DV4; Zeiss, Germany). The proboscises of
moths that were either killed immediately at flowers or following
2–3 days of captivity in large plastic bottles were examined under
an SEM (JSM-6360LV, JEOL, Japan). We also searched for
Deltophora adults or larvae outside the flowering seasons of their
Phyllanthus hosts, checking especially co-occurring Phyllanthaceae,
such as Breynia fruticosa (Mull.Arg.) Hook.f., Glochidion eriocarpum
Zipp. ex Span., Phyllanthus reticulatus Poir., and P. nanellus P.T. Li.
Adults and larvae were sent to specialists for identification
(Acknowledgments), and vouchers have been deposited in the
zoological collections of Nankai University and the Zoologische
Staatssammlung in Munich (ZSM).
Male and female flowers visited by Deltophora moths were
examined for moth eggs under a stereoscope. Both Phyllanthus
species were regularly checked for moth eggs on leaves,
inflorescences, and flower bracts, and moth larvae were brought
to the laboratory for rearing on host leaves to study the length of
larval and diapause stages. We also tried whether they would feed
on Glochidion eriocarpum leaves. Numbers of lab-reared larvae in
2006, 2007, and 2008 are specified in the Supporting Information
S1, and the quantification of leaf damage due to larval feeding is
described in Supporting Information S1.
Molecular phylogenetics and molecular clock-baseddivergence time estimation for the moths and their planthosts
For molecular phylogenetics and molecular clock dating, total
DNA was extracted from wild-caught and lab-reared Deltophora
adults and larvae using standard methods. We PCR-amplified a
part of the mitochondrial cytochrome oxidase subunit I gene
(COI) using the primers CO1-F 59-ATA ATT TTT TTT ATA
GTT ATA C-39 and CO1-R 59-GAT GGG CTC ATA CAA
TAA ATC CTA-39 [4]. Sequences have been submitted to
GenBank (GU247447-GU247452). Deltophora cytochrome c oxi-
dase subunit I (COI) sequences were aligned with GenBank
Gelechioidea sequences, with outgroup taxa chosen based on [18]
and [27]. For time calibration (below), we included Tegeticula
(Prodoxidae) and Galagete (Autostichidae). The tree was rooted on
Prodoxidae.
For each Phyllanthus species, we isolated total genomic DNA
from silica-dried leaves material using the methods described in
[28] and sequenced the plastid maturase K (matK) gene for which
over 130 other fully vouchered Phyllanthaceae sequences are
available in GenBank [28–31]. The GenBank matK sequence of
Bridelia ferruginea was renamed Bridelia sp. based on [28] and that of
Sauropus androgynus became S. garrettii based on [32]. Primers were
those described in [33]. New sequences have been submitted to
GenBank (GU263803 and GU263804). The matK matrix used to
infer closest relatives consisted of 132 sequences of Phyllanthaceae
plus a few Picrodendraceae, Euphorbiaceae, and Putranjivaceae
for rooting purposes.
Maximum likelihood (ML) analyses on the moth and plant
matrices were performed under the GTR + G model of
substitution using RAxML [34]. Bootstrap support was estimated
in RAxML with 100 replicate heuristic searches under the same
model as used in the searches. Maximum likelihood trees with
branch lengths were then used in relaxed molecular clock models
that incorporated at least one fossil-based calibration point (given
as a probability distribution). Specifically, we used the uncorrelat-
ed-rates Bayesian relaxed clock approach implemented in BEAST
v.1.4.8 [35], which uses a Markov chain Monte Carlo (MCMC)
method to co-estimate topology, substitution rates, and node ages.
Posterior probability distributions of node ages were obtained from
the cytochrome c oxidase subunit I or matK alignments, with the
number of taxa slightly reduced by taking out identical or near-
identical moth and plant sequences. Analyses used a speciation
model that followed a Yule tree prior, with rate variation across
branches lognormally distributed. MCMC chains were run for 10
million generations, with parameters sampled every 1000th step.
Whether the chains had reached stationarity was checked using
Tracer v. 1.4.1 [36]; this showed that the effective sample sizes for
all estimated parameters were well above 200.
For calibration of the moth phylogeny, we used a normal prior
distribution with a mean of 3.3 million years (Ma) and a SD of 1
Ma for the Galagete crown group and a mean of 32.4 Ma (SD 1 Ma)
for the Tegeticula crown group. The divergence time between
Tegeticula synthetica and T. yuccasella has been dated to 32.4 Ma [37].
The first radiation event on the Galapagos archipelago within the
Galagete lineage has been dated to 3.3 Ma [27).
For calibration of the plant phylogeny, the crown group age of
Phyllanthus sensu lato (including Breynia, Glochidion, and Sauropus) was
set to 55 million years (Ma) (SD = 1 Ma) based on pollen described
from the Lower Eocene Woolwich bed in Kent, England [38].
Although the pollen of Phyllanthus is heterogeneous, the Woolwich
pollen appears to be correctly assigned. As an alternative minimal
constraint, we could use fossil fruits that resemble those of
Phyllanthus from the Maastrichtian, 65–70 Ma ago [39].
GC-MS analyses of volatilesVolatiles emitted from the young leaves and from female and
male flowers were collected in the field by enclosing the leaves or
clusters of 20–50 flowers for 2 hours within a polyacetate bag.
Alternatively, 50 cut flowers were placed in 10 ml headspace glass
vials (CNW Technologies GmbH, Dusseldorf, Germany) that were
immediately sealed. We used solid phase microextraction (SPME;
Sigma-Aldrich) and SPME fibre assemblies (65 mm PDMS/DVB,
Sigma-Aldrich) to adsorb odour from the headspace. To analyze
the proboscis chemistry of Deltophora sp. 1 moths, we placed up to
ten proboscises in either methanol or chloroform solutions. Scent
samples and proboscis extracts were analyzed by thermal
desorbtion on a gas chromatograph/mass spectrometer (GC-
MS) QP2010 Plus Shimadzu QP2010 instrument. The conditions
used were column oven temperature 50.0uC, injection tempera-
ture 250.00uC, flow control mode linear velocity, pressure
53.5 kPa, total flow 12.0 mL/min, column flow 1.00 mL/min,
linear velocity 36.3 cm/sec, and purge flow 6.0 mL/min. The
GC-MS data were processed using GC Solution software release
2.5 SU3 (Shimadzu Co., Japan).
proboscis of Deltophora sp. 2. K. Longitudinal section of a proboscis, showing the thick integument and the food canal through which nectar issucked up. Dissolved pollen material might enter the food canal through the circular openings in the integument (yellow arrow).doi:10.1371/journal.pone.0019219.g002
First Documented Sporopollenin Breakdown
PLoS ONE | www.plosone.org 6 April 2011 | Volume 6 | Issue 4 | e19219

Figure 3. Chronogram for the Phyllantheae obtained under a Bayesian relaxed clock. Bars around node ages indicate the 95% highestposterior density intervals for nodes with a posterior probability .0.7. Phyllanthus cochinchinensis and P. rheophyticus, the species pollinated by theDeltophora moths studied here, are highlighted.doi:10.1371/journal.pone.0019219.g003
First Documented Sporopollenin Breakdown
PLoS ONE | www.plosone.org 7 April 2011 | Volume 6 | Issue 4 | e19219
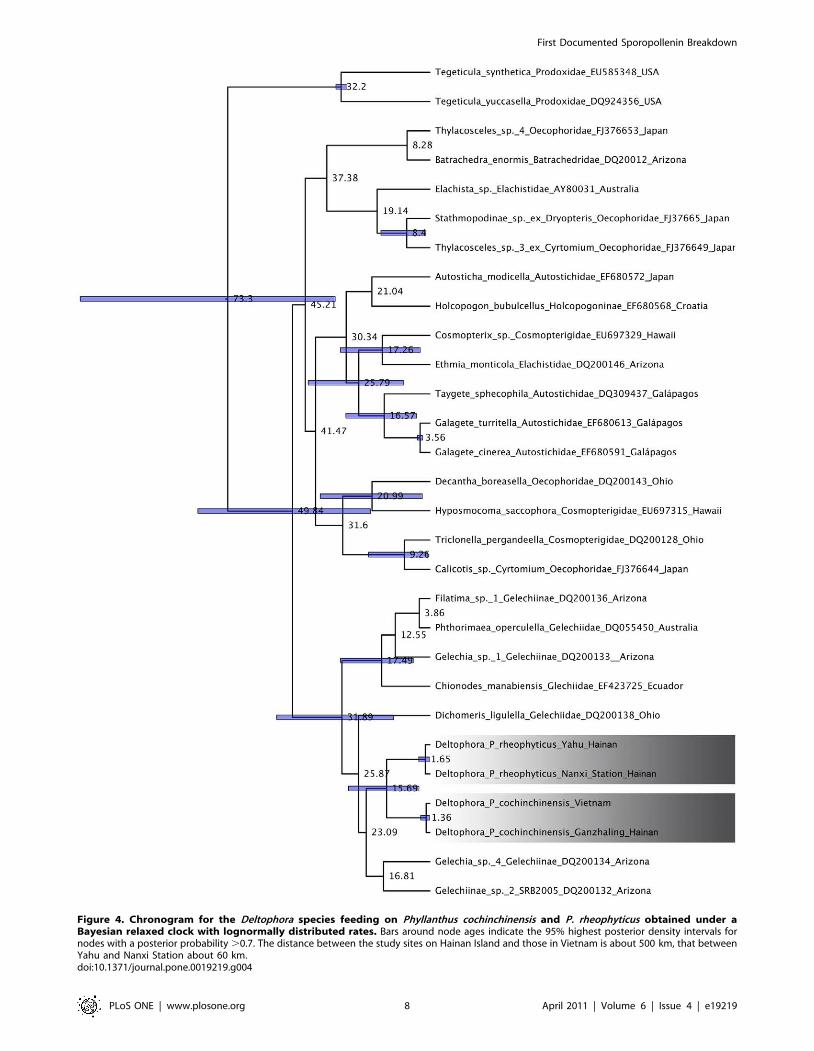
Figure 4. Chronogram for the Deltophora species feeding on Phyllanthus cochinchinensis and P. rheophyticus obtained under aBayesian relaxed clock with lognormally distributed rates. Bars around node ages indicate the 95% highest posterior density intervals fornodes with a posterior probability .0.7. The distance between the study sites on Hainan Island and those in Vietnam is about 500 km, that betweenYahu and Nanxi Station about 60 km.doi:10.1371/journal.pone.0019219.g004
First Documented Sporopollenin Breakdown
PLoS ONE | www.plosone.org 8 April 2011 | Volume 6 | Issue 4 | e19219

Supporting Information
Supporting Information S1 Additional supporting information
may be found in the online version of this article, specifically
details on localities, observation times, quantification of leaf
damage caused by the Deltophora larvae, and experiments on plant
mating systems.
(DOC)
Acknowledgments
We thank L. V. Lam, P. Zhu, H. J. Yang, and R. T. Li for assistance in the
field; W. Heller, Institute for Biochemical Plant Pathology, Munich, for
help with the interpretation of GC-MS data; and H. H. Li, Nankai
University for confirming moth identifications. Ron Hodges and Lauri
Kaila helped with the interpretation the Deltophora proboscis structure.
Author Contributions
Conceived and designed the experiments: SL YL. Performed the
experiments: SL YL SC. Analyzed the data: SL SSR. Contributed
reagents/materials/analysis tools: SSR DZ. Wrote the paper: SSR SL.
References
1. Pellmyr O, Thompson JN (1992) Multiple occurrences of mutualism in the
yucca moth lineage. Proc Natl Acad Sci U S A 89: 2927–2929.2. Fleming TH, Holland JN (1998) The evolution of obligate pollination
mutualisms: senita cactus and senita moth. Oecologia 114: 368–375.3. Kato M, Takimura A, Kawakita A (2003) An obligate pollination mutualism and
reciprocal diversification in the tree genus Glochidion (Euphorbiaceae). Proc Natl
Acad Sci U S A 100: 5264–5267.4. Kawakita A, Kato M (2009) Repeated independent evolution of obligate
pollination mutualism in the Phyllantheae–Epicephala association. Proc Roy Soc B276: 417–426.
5. Pellmyr O, Thien LB, Bergstrom G, Groth I (1990) Pollination of New
Caledonian Winteraceae: opportunistic shifts or parallel radiation with theirpollinators. Pl Syst Evol 173: 143–157.
6. Zeller-Lukashort HC, Kurz ME, Lees DC, Kurz MA (2007) A review ofMicropterix Hubner, 1825 from northern and central Europe (Micropterigidae).
Nota lepid 30: 235–298.
7. Krenn HW (2010) Feeding mechanisms of adult Lepidoptera: Structure,function, and evolution of the mouthparts. Ann Rev Entomol 55: 307–327.
8. Gilbert LE (1972) Pollen feeding and reproductive biology of Heliconius
butterflies. Proc Natl Acad Sci U S A 69: 1403–1407.
9. O’Brien DM, Boggs CL, Fogel ML (2003) Pollen feeding in the butterflyHeliconius charitonia: isotopic evidence for essential amino acid transfer from
pollen to eggs. Proc Roy Soc B 270: 2631–2636.
10. Eberhard SH, Hikl AL, Boggs CL, Krenn HW (2009) Saliva or regurgitatednectar? What do Heliconius butterflies (Lepidoptera: Nymphalidae) use for pollen
feeding. Ann Entomol Soc Am 102: 1105–1108.11. Dobritsa AA, et al. (2009) CYP704B1 is a long-chain fatty acid v-hydroxylase
essential for sporopollenin synthesis in pollen of Arabidopsis. Plant Physio 151:
574–589.12. Bailey IW (1960) Some useful techniques in the study and interpretation of
pollen morphology. J Arn Arb 41: 141–148.13. Southworth D (1974) Solubility of pollen exines. Am J Bot 61: 36–44.
14. Doyle JA, Hickey LJ (1976) Pollen and leaves from the mid-Cretaceous PotomacGroup and their bearing on early angiosperm evolution. In: Beck CB, ed. Origin
and Early Evolution of Angiosperms. New York: Columbia University Press. pp
139–206.15. Krassilov VA, Rasnitsyn AP, Afonin SA (2007) Pollen eaters and pollen
morphology: Coevolution through the Permian and Mesozoic. Afr Invert 48:3–11.
16. Roulston TH, Cane JH, Buchmann SL (2000) What governs protein content of
pollen: Pollinator preferences, pollen-pistil interactions, or phylogeny? EcolMonogr 70: 617–643.
17. Luo SX, Zhang DX, Renner SS (2007) Duodichogamy and androdioecy in theChinese Phyllanthaceae Bridelia tomentosa. Am J Bot 94. 260–265.
18. Kaila L (2004) Phylogeny of the superfamily Gelechioidea (Lepidoptera:Ditrysia): An exemplar approach. Cladistics 20: 303–340.
19. Svensson GP, Okamoto T, Kawakita A, Goto R, Kato M (2010) Chemical
ecology of obligate pollination mutualisms: testing the ‘private channel’hypothesis in the Breynia-Epicephala association. New Phytol 186: 995–1004.
20. Bucheli SR, Wenzel J (2005) Gelechioidea (Insecta: Lepidoptera) systematics: Areexamination using combined morphology and mitochondrial DNA data. Mol
Phyl Evol 35: 380–394.
21. Sattler K (1979) A taxonomic revision of the genus Deltophora Janse, 1950(Lepidoptera: Gelechiidae). Bull Brit Mus (Nat Hist), Entom Ser 38: 263–322.
22. Timmermans M, Randoux T, Daloze D, Braekman JC, Pasteels JM, et al.
(1992) The chemical defense of Doryphorina beetles (Coleoptera: Chrysomelidae).Biochem Syst Ecol 20: 343–349.
23. Attygalle AB, Blankespoor CL, Eisner T, Meinwald J (1994) Biosynthesis of adefensive insect alkaloid: Epilachnene from oleic acid and serine. Proc Natl Acad
Sci U S A 91: 12790–12793.
24. Eberhard SH, Hrassnigg N, Crailsheim K, Krenn HW (2007) Evidence ofprotease in the saliva of the butterfly Heliconius melpomene (L.) (Nymphalidae,
Lepidoptera). Insect Physio 53: 126–131.25. Krenn HW, Eberhard M, Eberhard S, Hikl AL, Huber W, Gilbert L (2009)
Mechanical damage to pollen aids nutrient acquisition in Heliconius butterflies
(Nymphalidae). Arthropod-Plant Interactions 3: 203–208.26. Li PT, Qiu H, Ma J, Zhu H, Gilbert MG, et al. (2009) Phyllanthus. In Wu ZY,
Raven PH, Hong DY, eds. Flora of China vol. 11. Beijing: Science Press/StLouis: Missouri Botanical Garden Press. pp 180–190.
27. Schmitz P, Cibois A, Landry B (2007) Molecular phylogeny and dating of an
insular endemic moth radiation inferred from mitochondrial and nuclear genes:The genus Galagete (Lepidoptera: Autostichidae) of the Galapagos Islands. Mol
Phyl Evol 45: 180–192.28. Li YQ, Dressler S, Zhang DX, Renner SS (2009) More Miocene dispersal
between Africa and Asia — the case of Bridelia (Phyllanthaceae). Syst Bot 34:521–529.
29. Kathriarachchi H, Hoffmann P, Samuel R, Wurdack KJ, Chase MW (2005)
Molecular phylogenetics of Phyllanthaceae inferred from five genes (plastid atpB,matK, 39 ndhF, rbcL and nuclear PHYC). Mol Phyl Evol 36: 112–134.
30. Kathriarachchi H, Samuel R, Hoffmann P, Mlinarec J, Wurdack KJ, et al.(2006) Phylogenetics of the tribe Phyllantheae (Phyllanthaceae; Euphorbiaceae
sensu lato) based on nrITS and plastid matK DNA sequence data. Am J Bot 93:
637–655.31. Kawakita A, Kato M (2006) Assessment of the diversity and species specificity of
the mutualistic association between Epicephala moths and Glochidion trees. MolEcol 15: 3567–3582.
32. Pruesapan K, Telford IRH, Bruhl JJ, Draisma SGA, Welzen PC van (2008)Delimitation of Sauropus (Phyllanthaceae) based on plastid matK and nuclear
ribosomal ITS DNA sequence data. Ann Bot 102: 1007–1018.
33. Samuel R, Kathriarachchi H, Hoffmann P, Barfuss MHJ, Wurdack KJ, et al.(2005) Molecular phylogenetics of Phyllanthaceae: Evidence from plastid matK
and nuclear PHYC sequences. Am J Bot 92: 132–141.34. Stamatakis A, Hoover P, Rougemont J (2008) A rapid bootstrap algorithm for
the RAxML web-servers. Syst Biol 75: 758–771. http://phylobench.vital-it.ch/
raxml-bb/.35. Drummond AJ, Rambaut A (2007) BEAST: Bayesian evolutionary analysis by
sampling trees. BMC Evolutionary Biology 7: 214. doi:10.1186/1471–2148–7–214.
36. Rambaut A, Drummond A (2003) Tracer: MCMC trace analysis tool. Oxford,UK: University of Oxford.
37. Gaunt MW, Miles MA (2002) An insect molecular clock dates the origin of the
insects and accords with palaeontological and biogeographic landmarks. MolBiol Evol 19: 748–761.
38. Gruas-Cavagnetto C, Kohler E (1992) Pollens fossils d’Euphorbiacees del’Eocene francais. Grana 31: 291–304.
39. Nambudiri EMV, Binda PL (1989) Dicotyledonous fruits associated with
coprolites from the Upper Cretaceous (Maastrichtian) Whitemud Formation,southern Saskatchewan, Canada. Rev Pal Palyn 59: 57–66.
First Documented Sporopollenin Breakdown
PLoS ONE | www.plosone.org 9 April 2011 | Volume 6 | Issue 4 | e19219