gaba induces gabaergic mscs in cultured embryonic rat thalamic neurons
TRANSCRIPT

GABA Induces GABAergic MSCsin Cultured Embryonic Rat
Thalamic NeuronsQI-YING LIU,* JEAN VAUTRIN, ANNE E. SCHAFFNER, WU MA, AND JEFFERY L. BARKER
Laboratory of Neurophysiology, National Institute of Neurological Disorders and Stroke, National Institutes of Health,Bethesda, Maryland 20892
KEY WORDS GABAA receptors; thalamus; neurotransmission; neuronal develop-ment; synapse
ABSTRACT Application of 0.1–10 µMGABAin the vicinity of cultured embryonic ratthalamic neurons recorded with patch pipettes in the presence of 2 µM TTX induced orincreased the frequency of miniature synaptic currents (MSCs) that reversed polarity atthe Cl2 equilibrium potential. These MSCs were blocked by the GABAA receptorantagonist bicuculline and exhibited exponential decay kinetics that closely paralleledthose estimated from fluctuation analysis of Cl2 channels activated pharmacologically byapplying 1–10 µM GABA to the same cells. We conclude that the MSCs are mediated byGABA. Application of the GABAA receptor agonist muscimol activated Cl2 current butfailed to induce GABAergic MSCs while submicromolar concentrations of GABA evokedGABAergic MSCs but did not activate Cl2 channels. The GABAB receptor agonist(2)baclofen did not mimic GABA in inducing MSCs. Induction of GABAergic MSCs byGABA required extracellular Ca21. Verapamil and Co21, which block voltage-dependentcalcium channels, completely blocked GABA-induced MSCs independent of their effectson the direct activation of a Cl2 current response. The results indicate that GABA cantrigger GABAergic Cl2-dependent MSCs in a Cao21-dependent manner. The mechanismmay involve a novel receptor and/or signal transduction pathway. Synapse 25:15–23,1997. r 1997 Wiley-Liss, Inc.
INTRODUCTION
g-aminobutyric acid (GABA) is well-established as afast-acting neurotransmitter throughout adult verte-brate central nervous systems (CNS), activating Cl2-and K1-selective ion channels and modulating voltage-dependent Ca21 channels (Bormann, 1988; Macdonaldand Olson, 1994; Rabow et al., 1995; Sivilotti andNistri, 1991). Recent evidence indicates that GABAemerges during the embryonic period throughout theCNS where it may initially function as a morphogenicsignal rather than as a fast-acting neurotransmitter(Behar et al., 1994; Lauder, 1993; Lauder et al., 1986).Pharmacological application of GABA to embryonic andearly postnatal neurons at all levels of the CNS testedthus far in different in vitro preparations depolarizesthem via activation of Cl2 channels (Ben-Ari et al.,1989; Cherubini et al., 1990, 1991; Mandler et al., 1990;Reichling et al., 1994; Walton et al., 1993; Wu et al.,1992). This depolarization may be sufficient to activatevoltage-dependent Ca21 channels, thereby elevatingcytosolic Ca21 (Reichling et al., 1994;Wang et al., 1994).However, GABA-induced changes in Ca21 that appear
to be independent of membrane depolarization via Cl2
channel activation have also been reported (Connor etal., 1987; Segal, 1993). Recently, synaptic-like transientsignals evoked by GABAhave been recorded in culturedpostnatal mouse retinal ganglion cells (Bahring et al.,1994) and granule cells in thin postnatal rat cerebelarslices (Kaneda et al., 1995). Here we show that GABAinduces miniature GABAergic synaptic-like Cl2 cur-rents in cultured embryonic rat thalamic neurons in aCao21-dependent manner at concentrations below thoserequired to activate postsynaptic Cl2 channels directlyand that this effect cannot be mimicked by muscimol,which directly activates postsynaptic GABAA receptor/Cl2 channels.
MATERIALS AND METHODS
Thalamic neurons were dissociated from embryonicday 17 rat embryos by papain digestion according to the
Received 15 December 1995; Accepted 27 March 1996.
*Correspondence to: Qi-Ying Liu,M.D., Laboratory of Neurophysiology, NINDS,NIH, Bldg. 36/Rm. 2C02, 9000 Rockville Pike, Bethesda, MD 20892. E-mail:liuqy @ codon.nih.gov.
SYNAPSE 25:15–23 (1997)
r 1997 WILEY-LISS, INC.

method of Huettner and Baughman (1986). Briefly, thethalamic tissues were carefully cut into small pieces,transferred into 5 ml Earl’s Balanced Salt Solution(EBSS) containing 20 U/ml papain, 0.005%DNase (bothfrom Boehringer Mannheim Co., Indianapolis, IN),0.5 mM EDTA, and 1 mM L-cysteine, and then kept inan incubator at 37°C for 50 min. Single neurons, ob-tained by triturating the tissue with a Pasteur pipette,were washed twice with culture medium containing90%Minimal Essential Medium (MEM, GIBCO, GrandIsland, NY), 5% fetal bovine serum (FBS), and 5% horseserum (HS) (Biofluid, Rockville, MD), and antibiotic/antimycotic solution (Sigma, St. Louis, MO). The neu-rons were then plated in 35 mm plastic culture dishescoatedwith highmolecular weight poly-d-lysine (Sigma)at a density of about 5 3 105 cells/dish. After severaldays, the differentiating neurons formed visible net-works. Cells in culture for 3 days or more were patch-clamped in the whole-cell configuration with pipettescontaining (in mM): 140 CsCl, 0.1 CaCl2, 2 MgCl2,1.1 EGTA, 5 ATP, 5 phosphocreatine, and 5 HEPES(pH 7.2, osmolarity 290 mOsm). The recording pipetteswere pulled from 1.5 mm o.d. capillary tubes (WPI,Sarasota, FL) with a computer-controlled microelec-trode puller (BB-CH-PC, Mecanex SA, Nyon, Switzer-land). Electrical resistances of the pipettes in recordingsaline ranged between 10 and 15 MV when filled withintracellular solution. Shortly before the experiment,the culture medium was replaced with extracellularrecording solution composed of (in mM): 140 NaCl,5.4 KCl, 0.8 MgCl2, 1.8 CaCl2, 10 HEPES, 10 Glucose(pH 7.4, osmolarity 310 mOsm). All recordings werecarried out at room temperature (22–25°C) on a Zeissinverted microscope. Pipettes for applying saline orligands were positioned within 20 µm of the cell body orone of the precesses of the recorded neuron; applica-tions were made at low pressure (#2 kpascal). Themembrane potential was held at 280 mV except other-wise specified. Membrane current was recorded using aList EPC-7 amplifier (List-Electronic, Darmstadt, Ger-many) at a gain of 5 mV/pA and stored on videotapeswith a VR-100 digital recorder (Instrutech, Great Neck,NY) and a Sharp VCR. Signals were filtered at 2 kHz(Frequency Devices 9002, 8-pole Bessel) and sampled at5 kHz for off-line analysis with a 486 PC computerusing the SCAN and SPAN program (courtesy J. Demp-ster, Strathclyde University, Glasgow, UK).
RESULTSGABA induces GABAergic miniature
synaptic-like currents in TTX
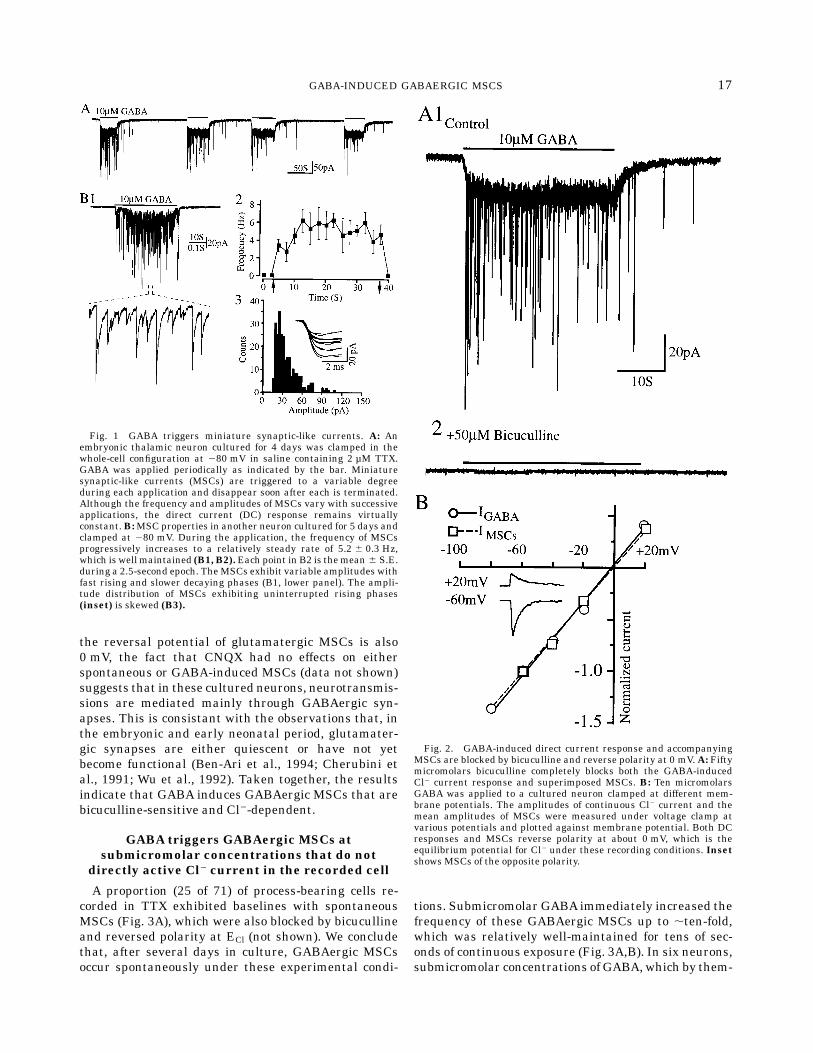
Application of 10 µM GABA by low pressure within,20 µm of the recorded cell evoked direct current (DC)responses of variable amplitude in all 71 cells tested. Inthe majority (n 5 43), these were associated with adramatic induction or increase in the frequency ofminiature synaptic currents (MSCs), as shown in Fig-
ure 1A, 1B1. These were recorded in the presence of2 µM TTX, which blocked voltage-dependent Na1 cur-rents (not shown). Repeated applications of GABA at,1 min intervals elicited virtually identical DC currentresponses while the frequency and amplitude of super-imposed MSCs were variable, both during an indi-vidual application and from exposure to exposure (Fig.1A). The episodes of MSCs evoked by GABA werehighly variable in their pattern of activity, as is evidentfrom this and the other examples illustrated. It wasapparent that the complex pattern of MSC activitieswas not precisely coupled to the period of exposurewhen the DC current response was used to monitormicromolar levels of GABA. MSC frequency was alsoquite variable during GABA exposure with evidenceboth of well-sustained rates of 4–8 Hz in some cells(n 5 26, Fig. 1B1) and progressively lower rates withrepeated (Fig. 1A) or continued exposure (Fig. 2A) inothers (n 5 17). The GABA-induced synaptic-like cur-rents sometimes out-lasted the DC current responsefollowing the termination of the application (n 5 29;Fig. 1A, also Figs. 2A1, 4A1, 5A, and Fig. 6A,B). Inother cells, GABA-evoked MSCs disappeared togetherwith the DC current immediately (Fig. 1B1) or soonafter the termination of GABA exposure (n 5 14; Fig.5B1, 5B3).The GABA-activated MSCs consisted of transients
characteristic of miniature currents recorded at syn-apses in different preparations of CNS tissue. Most ofthem exhibited fast, uninterrupted rising phases withwell-defined peaks in amplitude and considerably slowerdecay rates (Fig. 1B1, 1B3). Amplitude distributions ofMSCs with uninterrupted rising phases were consis-tently asymmetrical and skewed rather than symmetri-cal and normal (Fig. 1B3).To test if GABA-induced MSCs were GABAergic, we
added bicuculline, a GABAA receptor antagonist, to therecording chamber.As shown in Fig. 2A,MSCs superim-posed on a DC current response to GABAwere recordedunder control conditions but neither a DC currentresponse nor MSCs were detected after the addition of50 µM bicuculline (n 5 3). These effects were reversible(not shown). Hence, the MSCs were blocked by bicucul-line like the receptors activated directly by GABA. Thebaseline noise was considerably reduced by bicuculline(compare Fig. A1 with A2), suggesting that GABA wasalso tonically released in these cultured thalamic neu-rons as reported previously in cultured embryonichippocampal neurons (Valeyev et al., 1993; Vautrin etal., 1993). To further characterize the ionic basis ofGABA-induced MSCs, both the amplitude of the DCcurrent response and the mean amplitude of superim-posed MSCs were measured at different membranepotentials (Fig. 2B). The I-V plots show that both theDC current response to GABA and the MSCs reversedpolarity at about 0 mV (n 5 4), the equilibrium poten-tial for Cl2 under those recording conditions. Although
16 Q.-Y. LIU ET AL.
the reversal potential of glutamatergic MSCs is also0 mV, the fact that CNQX had no effects on eitherspontaneous or GABA-induced MSCs (data not shown)suggests that in these cultured neurons, neurotransmis-sions are mediated mainly through GABAergic syn-apses. This is consistant with the observations that, inthe embryonic and early neonatal period, glutamater-gic synapses are either quiescent or have not yetbecome functional (Ben-Ari et al., 1994; Cherubini etal., 1991; Wu et al., 1992). Taken together, the resultsindicate that GABA induces GABAergic MSCs that arebicuculline-sensitive and Cl2-dependent.
GABA triggers GABAergic MSCs atsubmicromolar concentrations that do not
directly active Cl2 current in the recorded cell
A proportion (25 of 71) of process-bearing cells re-corded in TTX exhibited baselines with spontaneousMSCs (Fig. 3A), which were also blocked by bicucullineand reversed polarity at ECl (not shown). We concludethat, after several days in culture, GABAergic MSCsoccur spontaneously under these experimental condi-
tions. Submicromolar GABA immediately increased thefrequency of these GABAergic MSCs up to ,ten-fold,which was relatively well-maintained for tens of sec-onds of continuous exposure (Fig. 3A,B). In six neurons,submicromolar concentrations of GABA,which by them-
Fig. 1 GABA triggers miniature synaptic-like currents. A: Anembryonic thalamic neuron cultured for 4 days was clamped in thewhole-cell configuration at 280 mV in saline containing 2 µM TTX.GABA was applied periodically as indicated by the bar. Miniaturesynaptic-like currents (MSCs) are triggered to a variable degreeduring each application and disappear soon after each is terminated.Although the frequency and amplitudes of MSCs vary with successiveapplications, the direct current (DC) response remains virtuallyconstant.B:MSC properties in another neuron cultured for 5 days andclamped at 280 mV. During the application, the frequency of MSCsprogressively increases to a relatively steady rate of 5.2 6 0.3 Hz,which is well maintained (B1, B2).Each point in B2 is themean 6 S.E.during a 2.5-second epoch. TheMSCs exhibit variable amplitudes withfast rising and slower decaying phases (B1, lower panel). The ampli-tude distribution of MSCs exhibiting uninterrupted rising phases(inset) is skewed (B3).
Fig. 2. GABA-induced direct current response and accompanyingMSCs are blocked by bicuculline and reverse polarity at 0 mV. A: Fiftymicromolars bicuculline completely blocks both the GABA-inducedCl2 current response and superimposed MSCs. B: Ten micromolarsGABA was applied to a cultured neuron clamped at different mem-brane potentials. The amplitudes of continuous Cl2 current and themean amplitudes of MSCs were measured under voltage clamp atvarious potentials and plotted against membrane potential. Both DCresponses and MSCs reverse polarity at about 0 mV, which is theequilibrium potential for Cl2 under these recording conditions. Insetshows MSCs of the opposite polarity.
17GABA-INDUCED GABAERGIC MSCS
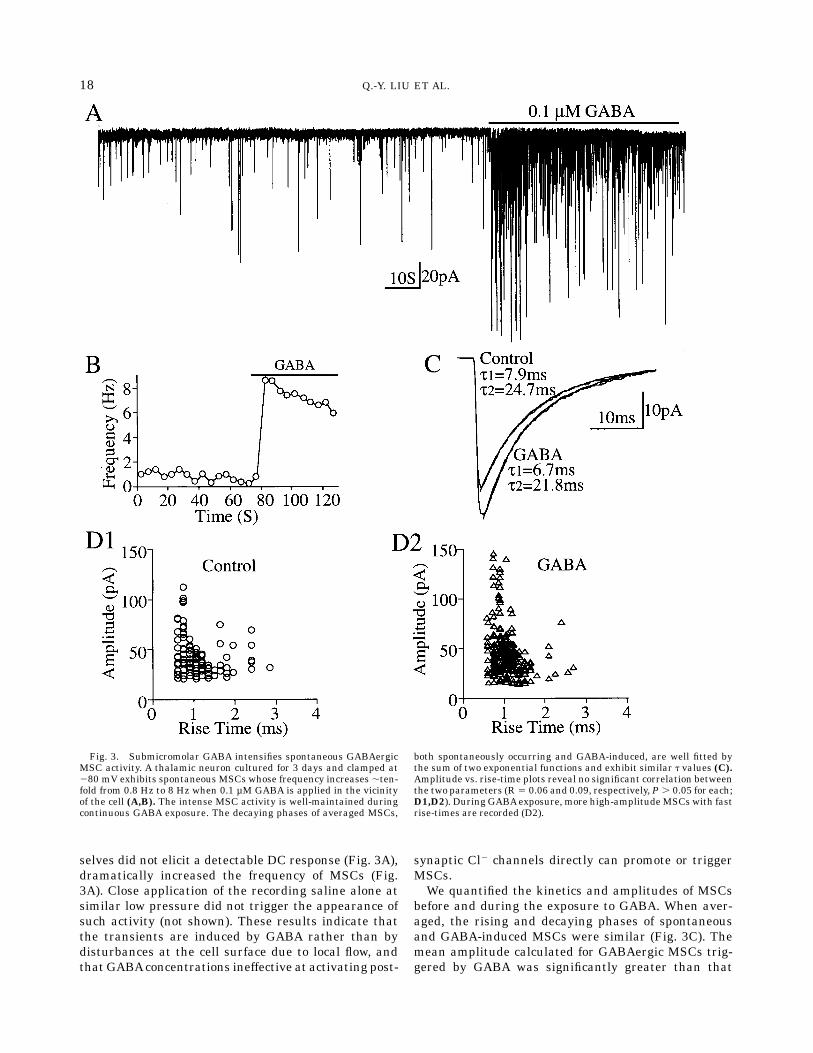
selves did not elicit a detectable DC response (Fig. 3A),dramatically increased the frequency of MSCs (Fig.3A). Close application of the recording saline alone atsimilar low pressure did not trigger the appearance ofsuch activity (not shown). These results indicate thatthe transients are induced by GABA rather than bydisturbances at the cell surface due to local flow, andthat GABAconcentrations ineffective at activating post-
synaptic Cl2 channels directly can promote or triggerMSCs.We quantified the kinetics and amplitudes of MSCs
before and during the exposure to GABA. When aver-aged, the rising and decaying phases of spontaneousand GABA-induced MSCs were similar (Fig. 3C). Themean amplitude calculated for GABAergic MSCs trig-gered by GABA was significantly greater than that
Fig. 3. Submicromolar GABA intensifies spontaneous GABAergicMSC activity. A thalamic neuron cultured for 3 days and clamped at280 mV exhibits spontaneous MSCs whose frequency increases ,ten-fold from 0.8 Hz to 8 Hz when 0.1 µM GABA is applied in the vicinityof the cell (A,B). The intense MSC activity is well-maintained duringcontinuous GABA exposure. The decaying phases of averaged MSCs,
both spontaneously occurring and GABA-induced, are well fitted bythe sum of two exponential functions and exhibit similar t values (C).Amplitude vs. rise-time plots reveal no significant correlation betweenthe two parameters (R 5 0.06 and 0.09, respectively, P . 0.05 for each;D1,D2). During GABAexposure, more high-amplitudeMSCs with fastrise-times are recorded (D2).
18 Q.-Y. LIU ET AL.

associated with spontaneous MSCs (Fig. 3C). In sixcells, the mean amplitude calculated by averaging90–400 MSCs in each cell before and during GABAincreased by 16.7 to 78.6%. MSC amplitude averaged45.2 6 2.8 pA for spontaneous and 65.2 6 5.8 pA dur-ing GABA (144.1 6 9.6%; P , 0.05). The increase inmean amplitude occurred without significant changesin the modal values of rise-time (not shown) and decay(Fig. 3C). Inspection of the amplitude vs. rise-time plotreveals that the increase in mean MSC amplitude wasdue to the more frequent occurrence of high-amplitudesignals having some of the fastest rise-times (Fig. 3D2).We quantified the amplitude and rise-time distribu-tions of individual MSCs whose rising phases wereuninterrupted. Amplitude did not correlate with rise-time either positively or negatively for spontaneous(r 5 0.06; P . 0.05; Fig. 3D1) or GABA-evoked MSCs(r 5 0.09; P . 0.05; Fig. 3D2). The results indicate thatspontaneous and GABA-induced MSCs involve trans-
mission at the same postsynaptic receptors. GABApromotes both an increase in GABAergic MSC fre-quency and amplitude.
Muscimol and baclofen do not triggerGABAergic MSCs
Muscimol, a structural analogue of GABA, readilyactivated GABAA receptor/Cl2 channels (Fig. 4A2) butfailed to elicit MSCs either at micromolar (Fig. 4A2) orsubmicromolar concentrations (Fig. 4B2) in seven cellsthat exhibited GABA-induced MSCs at the correspond-ing concentrations (Fig. 4A1,B1). (2)Baclofen, a GABAB
receptor agonist, also failed to mimic GABA in inducingGABAergic MSCs in cells where GABA was effective(Fig. 4C; n 5 4).
GABA’s induction of GABAergic MSCs isCao21-dependent and blocked by Co21
and verapamil
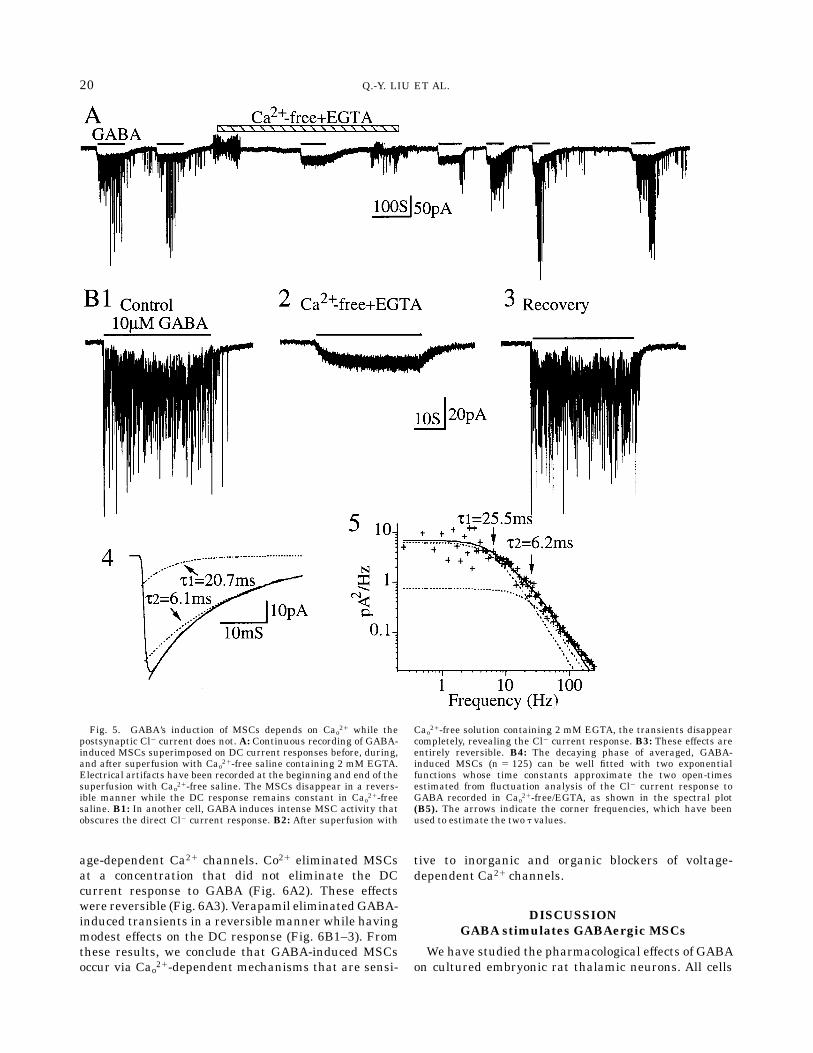
To determine if extracellular Ca21 was required forthe induction of MSCs in TTX, we applied GABA in thepresence and absence of extracellular Ca21 (Cao21). Thestimulatory effects of GABA were rapidly and revers-ibly eliminated when the cells (six of six cells tested)were superfusedwithCao21-free saline containingEGTA(Fig. 5A,B1–B3). In Cao21-free plus EGTA,GABAevokeda DC response completely devoid of MSCs (Fig. 5A,B2).These results indicate that the requirements for GABA-triggered MSCs differ from those involving exogenousactivation of GABAA receptor/Cl2 channels, with Cao21
being required for the former but not the latter effect.We quantified the decay phases of GABA-induced
MSCs and found that, when averaged, they consis-tently decayed in a bi-exponential manner with timeconstant (t) values of 5.6 6 0.7msec and 24.9 6 2.2 msecat 280 mV (n 5 10; Fig. 5B4). Since induction of MSCsby GABA required Cao21 while activation of Cl2 chan-nels in the recorded neurons did not, we were able tocompare measured t values of MSC decay with esti-mates of the exponentially-distributed open-channelkinetics directly activated by exogenous GABA (tGABA)in the same cell. Spectral analysis of the pharmacologi-cally activated Cl2 channels consistently revealed twoexponential components with tGABA values closely com-parable to those fitting the averaged bi-exponentialdecay (Figs. 5B5). The close similarity between corre-sponding pairs of t values suggests that the channelsputatively opened by endogenous transmitter havesimilar, if not identical, open-time kinetics to thosedirectly activated by GABA. The close correspondencein kinetics directly implicates GABAas the transmittermediating MSCs consistent with their Cl2-dependencyand bicuculline sensitivity described above.Since the GABA-inducedMSCs depended on extracel-
lular Ca21, we investigated the sensitivity of thisphenomenon to inorganic and organic blockers of volt-
Fig. 4. Muscimol and (2)baclofen fail to trigger MSCs. GABArapidly induces Cl2 current that is superimposed with MSCs exhibit-ing characteristic fast rising and slow decaying phases in a neuroncultured for 4 days (A1).Muscimol applied to the same neuron evokesa Cl2 current response that exhibits fluctuations characteristic ofrandomly occurring channel activity rather than transient signalscomposed of well-synchronized openings (A2). In another neuroncultured for 5 days that exhibits occasional low-amplitude transients,0.1 µMGABAincreasesMSCs dramatically (B1) but 0.1 µMmuscimolhas little or no effects on the cell (B2). Ten micromolars GABA inducesMSCs superimposed on Cl2 current in a neuron cultured for 4 days(C1) while 10 µM (2)baclofen applied to the same neuron does notinduce any obvious changes in the current trace (C2). Membranepotential was voltage-clamped at 280 mV in all recordings.
19GABA-INDUCED GABAERGIC MSCS
age-dependent Ca21 channels. Co21 eliminated MSCsat a concentration that did not eliminate the DCcurrent response to GABA (Fig. 6A2). These effectswere reversible (Fig. 6A3). Verapamil eliminatedGABA-induced transients in a reversible manner while havingmodest effects on the DC response (Fig. 6B1–3). Fromthese results, we conclude that GABA-induced MSCsoccur via Cao21-dependent mechanisms that are sensi-
tive to inorganic and organic blockers of voltage-dependent Ca21 channels.
DISCUSSIONGABA stimulates GABAergic MSCs
We have studied the pharmacological effects of GABAon cultured embryonic rat thalamic neurons. All cells
Fig. 5. GABA’s induction of MSCs depends on Cao21 while thepostsynaptic Cl2 current does not. A: Continuous recording of GABA-induced MSCs superimposed on DC current responses before, during,and after superfusion with Cao21-free saline containing 2 mM EGTA.Electrical artifacts have been recorded at the beginning and end of thesuperfusion with Cao21-free saline. The MSCs disappear in a revers-ible manner while the DC response remains constant in Cao21-freesaline. B1: In another cell, GABA induces intense MSC activity thatobscures the direct Cl2 current response. B2: After superfusion with
Cao21-free solution containing 2 mM EGTA, the transients disappearcompletely, revealing the Cl2 current response. B3: These effects areentirely reversible. B4: The decaying phase of averaged, GABA-induced MSCs (n 5 125) can be well fitted with two exponentialfunctions whose time constants approximate the two open-timesestimated from fluctuation analysis of the Cl2 current response toGABA recorded in Cao21-free/EGTA, as shown in the spectral plot(B5). The arrows indicate the corner frequencies, which have beenused to estimate the two t values.
20 Q.-Y. LIU ET AL.

tested responded to GABA, which activated Cl2 cur-rents that were completely and reversibly blocked bybicuculline. We conclude that functional GABAA recep-tor-coupled Cl2 channels are widely expressed in thispopulation during the embryonic period. AlthoughGABA is a primary inhibitory transmitter in the adultmammalian CNS, it has been reported that in embry-onic and early neonatal period, GABAacting on GABAA
receptors providesmost of the excitatory drive, whereasexcitatory glutamatergic synapses are quiescent (Ben-Ari et al., 1994; Cherubini et al., 1991; Wu et al., 1992).We found that in over half the cells tested, the Cl2
current response evoked by GABA in TTX sufficient toeliminate Na1 current and network-evoked synapticactivity was accompanied by miniature synaptic cur-rents (MSCs), suggesting that the presynaptic effect ofapplied GABA is functionally excitatory. That virtuallyall of the GABA-induced MSCs were long-lasting (tensof milliseconds), reversed polarity at ECl and wereeliminated by bicuculline (Figs. 1B; 2A2,B; 4A1,B1)identifies them as GABAergic. Faster-decaying gluta-mate-mediated mEPSCs were observed only in neuronscultured for at least 1 week (data not shown) support-ing the concept that GABA transmission may dominateat early stages of development (Gaiarsa et al., 1995).
Although submicromolar GABA did not evoke Cl2currents on any of the recorded cells, it triggeredGABAergic MSCs or increased both their frequencyand average amplitude when already present. GABA-evoked MSCs rapidly and reversibly disappeared com-pletely in Cao21-free medium while the directly acti-vated Cl2 current remained unaltered. There wasexcellent agreement between the two exponential com-ponents of GABA-activated Cl2 channel kinetics in-ferred from fluctuation analysis of Cl2 currents evokedin Cao21-free medium and the bi-exponential kinetics ofMSC decay averaged in Ca21-containing saline in thesame cell. Taken together, the results indicate that: 1)exogenousGABAcan trigger the appearance of GABAer-gic MSCs in a Cao21-dependent manner or increase thefrequency and amplitude of spontaneously occurringMSCs; and 2) the kinetic properties of GABA-activatedreceptors on the recorded cell determine the timecourse of MSC decay.
GABA-evoked GABAergic MSCs exhibitproperties characteristic of spontaneous
GABAergic MSCs
We quantified the amplitudes of all MSCs withuninterrupted rising phases, which we interpret to
Fig. 6. Inorganic and organic blockers of voltage-dependent Ca21 channels eliminate GABA-inducedMSCs. A1–3: GABA evokes a direct current response superimposed with MSCs. Addition of 1 mM Co21
completely eliminates theMSCs and depresses the direct current response. TheMSCs recover completely,as doesmost of the direct current response.B1–3:Verapamil (50 µM) eliminates theMSCs completely andreversibly while having only modest effects on the direct current response.
21GABA-INDUCED GABAERGIC MSCS

reflect all-or-none episodic discharges of GABA. Ampli-tude vs. rise-time plots of GABA-evokedMSCs virtuallysuperimposed with those derived from analysis of spon-taneously occurring GABAergicMSCs recorded in thesecultured thalamic neurons (e.g., Fig. 3). This suggeststhat similar processes underlie the evoked and sponta-neous GABAergic MSCs. Furthermore, GABAergicMSCs with amplitude and rise-time characteristicsthat generate similar amplitude vs. rise-time plots havebeen reported for embryonic hippocampal neuronsmain-tained in dissociated cell culture (Vautrin et al., 1992,1993), and for hippocampal neurons recorded in slicepreparations of postnatal and adult tissues (Edward etal., 1990; Ropert et al., 1990). Hence, GABAergic MSCsrecorded in different cells and physiological prepara-tions of CNS tissue may all be generated by a similarset of processes.
GABA-evoked GABAergic MSCs may involvenovel GABA receptor functions
GABA’s induction or intensification of GABAergicMSCs may involve a novel receptor and/or signaltransductionmechanism.We found that GABA’s stimu-lation of GABAergic MSCs occurred at submicromolarconcentrations, which were consistently ineffective atdirectly activating Cl2 current in these cells. Thissuggests that unless there are GABAA receptors coupledto Cl2 channels that respond to submicromolar GABA,GABA’s activation of Cl2 channels may not be requiredfor triggering MSCs. Consistent with this idea, wefound that muscimol readily activated Cl2 channels inall cells tested but never elicited MSCs even in thosewhere GABA triggered MSCs. In addition, baclofen, aGABAmimetic at GABAB receptors, failed to evokeMSCs. The negative results with these GABAmimeticsand with application of saline eliminate simple distur-bances at the cell surface as a contributing factor in thephenomenology. The effectiveness of submicromolarGABA, coupled with the ineffectiveness of the GABAmi-metics at GABAA and GABAB receptors, suggest thatGABA may interact with a novel receptor and signaltransduction mechanism to trigger the MSCs.These pharmacological effects of exogenous GABA
indicate that GABA receptor-coupled mechanisms existat or near sites of GABA release, which can serve toinitiate or intensify spontaneous transmission at GABAreceptor/Cl2 channels. Since this phenomenology oc-curred when embryonic neurons had differentiatedvisible networks of processes, we presume that thetransmission takes place at sites of functional synapticcontacts. In fact, simultaneous recordings from synapti-cally-coupled pairs of cultured embryonic thalamicneurons reveal spontaneous and evoked, bicuculline-sensitive, Cl2-dependent synaptic currents (Qi-YingLiu, unpublished observations). The spontaneous syn-aptic currents in these pair-recordings possessed ampli-tudes and kinetics similar to those described here for
spontaneous and GABA-evoked MSCs (unpublishedobservation). These results are consistent with the ideathat the GABAergic MSCs triggered by GABA applica-tion arise at synaptic contacts that were already func-tional.The pharmacological effects of submicromolar to mi-
cromolar GABA to stimulate MSC activity requiredextracellular Ca21 and were sensitive to well-estab-lished inorganic (Co21) or organic (verapamil) blockersof voltage-dependent Ca21 channels, implicating Ca21
entry via Ca21 channels in the phenomenology. Presum-ably, GABA receptor-coupled Ca21 entry sites co-localize with those of GABArelease so that submicromo-lar GABA triggers or intensifies transient discharge ofGABA, which momentarily activates the receptor-coupled Cl2 channels that underlie GABAergic MSCs.As noted in the Introduction, activation of GABAA
receptor-coupled Cl2 channels depolarizes developingnerve cells recorded electrically with sharp microelec-trodes in slice preparations of hippocampal (Ben-Ari etal., 1989; Cherubini et al., 1990, 1991) and spinal cordcells (Wu et al., 1992). In these preparations, thedepolarizing effects are sufficient to trigger action poten-tials since EGABA (ECl) is supra-threshold for activationof voltage-dependent, inwardly directed currents.Micro-molar-supramicromolar muscimol or GABA depolarizecultured spinal cord cells recorded with potentiometricdyes (Walton et al., 1993) or patch pipettes (Wang et al.,1994). Flow cytometric recordings of spinal, hippocam-pal, and striatal cells in suspension with potentiometricdyes shows that during the embryonic period, themajority of embryonic neurons depolarize in responseto GABA and muscimol (Fiszman et al., 1990, 1993;Mandler et al., 1990). Activation of GABA receptor-coupled Cl2 channels near release sites may depolarizecells, triggering extracellular Ca21 entry via voltage-gated Ca21 channels, which in turn could elevatecytoplasmic free Ca21 (Lin et al., 1994; Reichling et al.,1994). If this is true, then submicromolar GABA actingat a highly efficient GABAA receptor/Cl2 channel maystimulate transient release of GABA via depolarizingeffects near release sites, yet be ineffective at directlyactivating Cl2 current recorded at the cell body. Musci-mol did not mimic GABA in inducing MSCs, yet readilyactivated Cl2 currents recorded at the cell body in allcells. Similar results were found for isoguvacine (datanot shown). If all cells have depolarizing responses toGABA at the level of the cell body, then depolarizingstimuli acting via GABAA receptor/Cl2 channels at thecell body are, per se, not capable of triggering GABArelease at release sites wherever they are located. Inaddition, the results with muscimol and isoguvacinedemonstrate that the GABA receptor-coupled mecha-nism that triggers MSCs has very low or no affinity formuscimol and isoguvacine. The underlying mecha-nisms for its activation by GABA are a challenge forfuture study.
22 Q.-Y. LIU ET AL.

In conclusion, in embryonic rat thalamic neuronsdifferentiating in culture we have found that submicro-molar GABA triggers pulsatile release of micromolarGABA, resulting in a variable period of fast transmis-sion at Cl2 channels. The variabilities in onset andoffset of the phenomenon and its dependency on extra-cellular Ca21 remain to be explained.
REFERENCES
Bahring, R., Standhardt, H., Martelli, E.A., and Grantyn, R. (1994)GABA-activated chloride currents of postnatal mouse retinal gan-glion cells are blocked by acetylcholine and acetylcarnitine: howspecific are ion channels in immature neurons? Eur. J. Neurosci.,6:1089–1099.
Behar, T.N., Schaffner, A.E., Colton, C.A., Somogyi, R., Olah, Z., Lehel,C., and Barker, J.L. (1994) GABA-induced chemokinesis and NGF-induced chemotaxis of embryonic spinal cord neurons. J. Neurosci.,14:29–38.
Ben-Ari, Y., Cherubini, E., Corradetti, R., and Gaiarsa, J.L. (1989)Giant synaptic potentials in immature rat CA3 hippocampal neu-rons. J. Physiol. (Lond.), 416:303–325.
Ben-Ari, Y., Tseeb, V., Raggozzino, D., Khazipov, R., Gaiarsa, J.L.(1994) g-Aminobutyric acid (GABA): a fast excitatory transmitterwhich may regulate the development of hippocampal neurones inearly postnatal life. Prog. Brain Res., 102:261–273.
Bormann, J. (1988) Electrophysiology of GABAA and GABAB receptorsubtypes. Trends Neurosci., 11:112–116.
Cherubini, E., Rovira, C., Gaiarsa, J.L., Corradetti, R., and Ben-Ari, Y.(1990) GABAmediates excitation in immature rat CA3 hippocampalneurons. Int. J. Dev. Neurosci., 8:481–490.
Cherubini, E., Gaiarsa, J.L., and Ben-Ari, Y. (1991) GABA: anexcitatory transmitter in early postnatal life. Trends Neurosci.,14:515–519.
Connor, J.A., Tseng, H.Y., and Hockberger, P.E. (1987) Depolarization-and transmitter-induced changes in intracellular Ca21 of rat cerebel-lar granule cells in explant cultures. J. Neurosci., 7:1384–1400.
Edwards, F.A., Konnerth, A., and Sakmann, B. (1990) Quantal analy-sis of inhibitory synaptic transmission in the dentate gyrus of rathippocampal slices: a patch-clamp study. J. Physiol. (Lond.), 430:213–249.
Fiszman, M.L., Novotny, E.A., Lange, G.D., and Barker, J.L. (1990)Embryonic and early postnatal hippocampal cells respond to nano-molar concentrations of muscimol. Dev. Brain Res., 53:186–193.
Fiszman, M.L., Behar, T., Lange, G.D., Smith, S.V., Novotny, E.A., andBarker, J.L. (1993) GABAergic cells and signals appear together inthe early post-mitotic period of telencephalic and striatal develop-ment. Dev. Brain Res., 73:243–251.
Gaiarsa, J.L., McLean, H., Congar, P., Leinekugel, X., Khazipov, R.,Tseeb, V., Ben-Ari, Y. (1995) Postnatal maturation of gamma-aminobutyric acidA and B-mediated inhibition in the CA3 hippocam-pal region of the rat. J. Neurobiol., 26:339–349.
Huettner, J.E., and Baughman, R.W. (1986) Primary culture ofidentified neurons from visual cortex of postnatal rats. J. Neurosci.,6:3044–3060.
Kaneda, M., Farrant, M., and Cull-Candy, S.G. (1995) Whole-cell andsingle-channel currents activated by GABA and glycine in granulecells of the rat cerebellum. J. Physiol. (Lond.), 485:419–435.
Lauder, J.M. (1993) Neurotransmitters as growth regulatory signals:role of receptors and second messengers. Trends Neurosci., 16:233–240.
Lauder, J.M., Han, V.K., Henderson, P., Verdoorn, T., and Towle, A.C.(1986) Prenatal ontogeny of the GABAergic system in the rat brain:an immunocytochemical study. Neuroscience, 19:465–493.
Lin, M.H., Takahashi, M.P., Takahashi, Y., and Tsumoto, T. (1994)Intracellular calcium increase induced by GABA in visual cortex offetal and neonatal rats and its disappearance with development.Neurosci. Res., 20:85–94.
Macdonald, R.L., and Olsen, R.W. (1994) GABAA receptor channels.Ann. Rev. Neurosci., 17:569–602.
Mandler, R.N., Schaffner, A.E., Novotny, E.A., Lange, G.D., Smith,S.V., and Barker, J.L. (1990) Electrical and chemical excitabilityappear one week before birth in the embryonic rat spinal cord. BrainRes., 522:46–54.
Rabow, L., Russek, S.J., and Farb, D.H. (1995) From ion currents togenomic analysis: recent advances in GABAA receptor research.Synapse, 21:189–274.
Reichling, D.B., Kyrozis, A., Wang, J., and MacDermott, A.B. (1994)Mechanisms of GABA and glycine depolarization-induced calciumtransients in rat dorsal horn neurons. J. Physiol. (Lond.), 476:411–421.
Ropert, N., Miles, R., and Korn, H. (1990) Characteristics of miniatureinhibitory postsynaptic currents in CA1 pyramidal neurons of rathippocampus. J. Physiol. (Lond.), 428:707–722.
Segal, M. (1993) GABA induces a unique rise of [Ca]i in cultured rathippocampal neurons. Hippocampus, 3:229–238.
Sivilotti, L., and Nistri, A. (1991) GABA receptor mechanisms in thecentral nervous system. Prog. Neurobiol., 36:35–92.
Valeyev, A.Y., Cruciani, R.A., Lange, G.D., Smallwood, V.S., andBarker, J.L. (1993) Cl2 channels are randomly activated by continu-ous GABA secretion in cultured embryonic rat hippocampal neu-rons. Neurosci. Lett. 155:199–203.
Vautrin, J., Schaffner, A.E., and Barker, J.L. (1992) Two classes ofspontaneous GABA-mediated miniature synaptic currents in cul-tured rat hippocampal neurons. Neurosci. Lett., 138:67–71.
Vautrin, J., Schaffner, A.E., and Barker, J.L. (1993) Quantal andsubquantal GABAergic transmission in cultured rat hippocampalneurons. Hippocampus, 3:93–102.
Walton, M.K., Schaffner, A.E., and Barker, J.L. (1993) Sodium chan-nels, GABAA receptors, and glutamate receptors develop sequen-tially on embryonic rat spinal cord cells. J. Neurosci., 13:2068–2084.
Wang, J., Reichling, D.B., Kyrozis, A., and MacDermott, A.B. (1994)Developmental loss of GABA- and glycine-induced depolarizationand Ca21 transients in embryonic rat dorsal horn neurons inculture. Eur. J. Neurosci., 6:1275–1280.
Wu, W.L., Ziskind-Conhaim, L., and Sweet, M.A. (1992) Early develop-ment of glycine- and GABA-mediated synapses in rat spinal cord. J.Neurosci., 12:3935–3945.
23GABA-INDUCED GABAERGIC MSCS