functional morphology and development of veliger larvae of
TRANSCRIPT

Functional Morphology andDevelopment of Veliger Larvae
of the European Oyster,Ostrea edulis Linne
THOMAS R. WALLER
SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY • NUMBER 328

SERIES PUBLICATIONS OF THE SMITHSONIAN INSTITUTION
Emphasis upon publication as a means of "diffusing knowledge" was expressedby the first Secretary of the Smithsonian. In his formal plan for the Institution, JosephHenry outlined a program that included the following statement: "It is proposed topublish a series of reports, giving an account of the new discoveries in science, andof the changes made from year to year in all branches of knowledge." This themeof basic research has been adhered to through the years by thousands of titles issuedin series publications under the Smithsonian imprint, commencing with SmithsonianContributions to Knowledge in 1848 and continuing with the following active series:
Smithsonian Contributions to Anthropo/ogySmithsonian Contributions to Astrophysics
Smithsonian Contributions to BotanySmithsonian Contributions to the Earth Sciences
Smithsonian Contributions to PaleobiologySmithsonian Contributions to ZoologySmithsonian Studies in Air and Space
Smithsonian Studies in History and Technology
In these series, the Institution publishes small papers and full-scale monographsthat report the research and collections of its various museums and bureaux or ofprofessional colleagues in the world cf science and scholarship. The publications aredistributed by mailing lists to libraries, universities, and similar institutions throughoutthe world.
Papers or monographs submitted for series publication are received by theSmithsonian Institution Press, subject to its own review for format and style, onlythrough departments of the various Smithsonian museums or bureaux, where themanuscripts are given substantive review. Press requirements for manuscript and artpreparation are outlined on the inside back cover.
S. Dillon RipleySecretarySmithsonian Institution

S M I T H S O N I A N C O N T R I B U T I O N S T O Z O O L O G Y • N U M B E R 3 2 8
Functional Morphology andDevelopment of Veliger Larvae
of the European Oyster,Ostrea edulis Linne
Thomas R. Waller
SMITHSONIAN INSTITUTION PRESS
City of Washington
1981

A B S T R A C T
Waller, Thomas R. Functional Morphology and Development of VeligerLarvae of the European Oyster, Ostrea edulis Linne. Smithsonian Contributions toZoology, number 328, 70 pages, 152 figures, 1 table, 1981.—Veliger larvae ofthe European oyster taken seven days and 15 days after release from parentwere anesthetized, fixed, critical-point dried, and examined with scanningelectron microscopes. Study of body and shell in a growth series revealedmany structures and patterns of development previously poorly known orunknown: (1) the presence of four ciliary bands on the velum, not three; (2)the heel-first development of the foot, with medial ciliation of the toe precedingcomplete ciliation; (3) the order of appearance and structure of major ductopenings on the foot and body wall; (4) the beginning of larval eyes ashemispherical tufts of elongate microvilli; (5) the early development of gillprimordia; and (6) the early appearance of a prominent gill bridge, whichoriginates by cross-contact of cilia on the mantle margins followed by migra-tion of epithelial cells and tissue fusion.
A striking posterodorsal notch and growth track on the left valve ofprodissoconch II of many Ostreidae and Gryphaeidae is formed by postanalcilia, which deform the fragile calcifying edge of the left valve but are unableto deform the more massive margin of the right valve.
The transition from shell gland to mantle occurs very early in the pro-dissoconch-I stage, and increasing specialization of the mantle edge producesa three-layered shell well before the prodissoconch-I/II boundary, whichmerely marks the moment when the valves first completely enclose the bodyand close against one another. Shell microstructure does not change untilmetamorphosis, at which time the aragonitic microstructures disappear andare succeeded by far coarser calcitic structures. The postmetamorphic myos-tracum is an exception. It remains aragonitic and is possibly a continuation ofthe inner layer of the larval shell. The prodissoconch ligament is formed byperiostracum, the fibrous resilium appearing later near the time of metamor-phosis. The larval shells of oviparous and larviparous oysters differ, those ofthe former having a prodissoconch I that is smaller compared to prodissoconch
OFFICIAL PUBLICATION DATE is handstamped in a limited number of initial copies and is recordedin the Institution's annual report, Smithsonian Year. SERIES COVER DESICN: The coral Montastreacavemosa (Linnaeus).
Library of Congress Cataloging in Publication DataWaller, Thomas R.Functional morphology and development of veliger larvae of the European oyster, Ostrea edulis
Linne(Smithsonian contributions to zoology ; no. 328)Bibliography: p.1. European oyster—Development. 2. Mollusks—Larvae. 3. Mollusks—Development. I.
Title. II. Title: Veliger larvae of the European oyster, Ostrea edulis Linne. III. Series:Smithsonian Institution. Smithsonian contributions to zoology ; no 328
QL1.S54 no. 328 [QL430.7.09] 591s [594M1] 80-23129

Contents
Page
Introduction 1Acknowledgments 1
Background 2Review of Larval Development 3Materials and Methods 9Terms, Measurements, and Limitations 10Observations and Functional Interpretations 11
Behavior 11Morphology 13
Velum 14Foot 17Structures in Gill Cavities 18
Static-Duct Openings 18Photoreceptors 20Protonephridial-Duct Openings 20
Gills 20Anal Region 22Mantle Edge 23Shell 24
Shape and Sculpture 24Shell Microstructure 42Hinge and Margin 45
Shell Morphology of Other Oyster Larvae 47Discussion 48Summary and Conclusions 62
Velum 62Foot 63Structures in Gill Cavities 63Gill Primordia 63Anal Region and Posterodorsal Shell Notch 63Mantle Edge 64Shell 64
Literature Cited 66


Functional Morphology andDevelopment of Veliger Larvae
of the European Oyster,Ostrea edulis Linne
Thomas R. Waller
Introduction
Two groups of oysters may be distinguished onthe basis of their breeding habits (Morton, 1958;Galtsoff, 1964; Stenzel, 1971). An "oviparous" ornonincubatory group, which includes the Amer-ican oyster, Crassostrea virginica (Gmelin), the Jap-anese oyster, C. gigas (Thunberg), and other spe-cies, discharges eggs and sperm directly into thesurrounding seawater. Fertilization is thus exter-nal, and the developing planktonic larvae areafforded no parental protection. In contrast, "lar-viparous" or incubatory oysters, such as the Eu-ropean oyster, Ostrea edulis Linne, and the Pacificoyster, 0. lurida Carpenter, cast only sperm intothe surrounding water. Eggs are retained withinthe shell of the female and are fertilized internallyby sperm drawn into the mantle cavity with theincurrent stream. Early development occurswithin the shell of the parent, and when thelarvae are finally released (some 6 to 18 days afterfertilization depending on temperature) they pur-sue a short planktonic life (generally 0 to 17 days,also depending on temperature) before perma-nently settling on a suitable substrate (Erdman,
Thomas R. Waller, Department of Paleobiology, National Museumof Natural History, Smithsonian Institution, Washington, D.C.20560.
1935; Korringa, 1941; Chanley, pers. comm.,1979).
The present study is concerned primarily withlarval development in the larviparous group, spe-cifically in the European oyster, Ostrea edulis. De-scriptions deal with larval morphology from timeof release from parent to just before eyespotsbecome pigmented and the foot becomes fullyfunctional. Emphasis is on structure of the velum,development of organs in the mantle cavity, ul-trastructure and development of shell and man-tle, and relationship between features of shell andbody. For comparison, larval shells of some ovi-parous species, particularly of Crassostrea gigas andC. virginica, are discussed briefly.
ACKNOWLEDGMENTS.—I thank Paul Chanley,Fundacion Chile, Coquimbo, whose shipment ofliving larvae from the Shelter Island Oyster Com-pany launched this project. Warren Blow, WalterR. Brown, Mary-Jaque Mann, Duane Hope,John Harshbarger, and Kenneth M. Towe of theSmithsonian Institution, Ruth D. Turner of Har-vard University, and Melbourne R. Carriker ofthe University of Delaware provided technicalassistance. Critical reviews were provided byChanley, Turner, and Carriker, as well as FrankR. Bernard and Daniel B. Quayle, NanaimoBiological Station, British Columbia, Dale Bonar
1

SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
and Robert D. Burke, University of Maryland,Richard A. Lutz, Yale University, and P. Dina-mani, Fisheries Research Division, Wellington,New Zealand. The artwork is by Lawrence B.Isham, Department of Paleobiology, SmithsonianInstitution.
Background
In the vast literature on the biology of oysterlarvae that has accumulated since Brach (1690)and Leeuwenhoeck (1695) first trained their earlycompound microscopes on these tiny organisms,there are still relatively few detailed studies ofbivalve larval development and morphology (seereviews by Horst, 1883-1884, Korringa, 1941,and Galtsoff, 1964). The most complete descrip-tion of cell lineage and early development of thepreveliger stages of oyster larvae remains that ofHorst (1883-1884), which describes the develop-ment of larvae of Ostrea edulis from fertilizationup to the moment of release from parent. Otherstudies of these early stages are more limited inscope or lack detail, e.g., Davaine (1853) andFernando (1931) on 0. edulis, Hori (1933) on 0.lurida, Brooks (1880) and Galtsoff (1964) on Crass-ostrea virginica,;, and Fujita (1929, 1934) on C.gigas. More detailed information on early devel-opment can be obtained from studies of unrelatedbivalves, particularly the work of Meissenheimer(1900) on Dreissena, Hatschek (1880) on Teredo,and Allen (1961) on Pandora.
The gross anatomy of more advanced oysterlarvae is known primarily through the exhaustivestudy of Erdmann (1935), which is again con-cerned with the European oyster. Erdmann beganhis study with a brief description of the oldestgrowth stage considered by Horst (1883-1884),the newly released larva, and finished with anextensive description of the oldest free-swimmingstage, the 'ansatzreife' (attachment-ready) larva,now generally referred to as the pediveliger (Car-riker, 1961). Millar (1955) studied the mechanismof food movement in the gut of larval Ostrea edulisof unspecified growth stage, but presumably thepediveliger stage, and updated information pre-
viously furnished by Yonge (1926) and Erdmann(1935). Cole (1938) and Hickman and Gruffydd(1971) concentrated on growth stages beyondthose studied by Erdmann and dealt with the fateof larval organs of 0. edulis through metamorpho-sis. Cranfield (1973a, b, c, 1974) described theultrastructure and histochemistry of the foot andmantle edge of the pediveliger of Ostrea edulis andthe function of these organs during settlementand cementation.
Until recently investigation of the larval shellof the oyster has lagged far behind the study ofsoft tissue. Most papers dealing with larval shellsof oysters have been concerned primarily withidentification and hence have concentrated onshell outline and the configuration of hinge teeth,both of which are within the range of light mi-croscopy. Most of these studies deal only withlate larval stages and present the hinge in a highlydiagrammatic fashion (e.g., Rees, 1950; Ranson,1960; Stenzel, 1971). Pascual (1971, 1972), how-ever, published exceptionally high-quality lightmicrographs showing the development of thehinge of both oviparous and larviparous species,including Ostrea edulis, and Dinamani (1973) usedlight microscopy to show hinge development inthe oviparous species, Saccostrea glomerata. Dina-mani (1976) also used scanning electron micros-copy to demonstrate differences in the develop-ment of the provinculum (larval hinge) of oystersin the genera Crassostrea, Saccostrea, and Ostrea.Carriker and Palmer (1979a) used scanning elec-tron microscopy to study the shell of Crassostreavirginica, including microsculpture, microstruc-ture, and a peculiar posterodorsal notch. Thenotch was discussed from a functional point ofview by Waller (1979) in abstract and will beexamined in detail in the present study.
In 1975 Turner and Boyle published the firstscanning electron micrographs of whole, anesthe-tized bivalve larvae prepared by critical-pointdrying, a technique first developed by Anderson(1951, 1956) and now widely applied in biologyand medicine (see Becker and Johari, 1978, forextent of application). Boyle and Turner (1976)later used this technique to illustrate the velar

NUMBER 328
and pedal ciliation of a pholad, Martesia striata,and Waller (1980) demonstrated the extent towhich critical-point drying could be used in stud-ies of cell surfaces, micro-anatomy, and mantle-shell relationships in mature bivalves in the OrderArcoida.
Review of Larval Development
The diameter of the spherical egg of Ostreaedulis is about 150 /tm when it is released fromthe gonad (Orton, 1937; Korringa, 1941; Yonge,1960). This size is at least two or three times thatof the eggs of oviparous species (Amemiya, 1926;Hori, 1933).
Differing accounts of cell lineage and gastru-lation were given by Horst (1883-1884) and Fer-nando (1931). Basically, cleavage follows the typ-ical molluscan spiral pattern. Micromeres spreadfrom the animal pole over the surface of themacromeres toward the vegetal pole, and gastru-lation is intermediate between epiboly and em-boly. It is unclear whether the blastopore remainsopen to become the mouth at a later stage(Horst's view and also Fujita's, 1934) or closesand subsequently becomes the site of a stomo-daeum and later the mouth (Fernando's view).The morphological outcome in either case is anembryo that is spheroidal, with the animal poledomed and the vegetal pole concave and bearingthe presumptive mouth opening (Figure la). Nocilia have been reported on the surface of theembryo at this point in the development of Ostreaedulis, although the embryo of Crassostrea virginicaat a comparable stage has been shown with acovering of fine cilia (Galtsoff, 1964). Amemiya(1926) mentioned that the blastula of C. virginicarotates, and Seno (1929) described the gastrula ofOstrea denselamellosa as being ciliated and motile.
Two important structures, a crown of motilecilia (prototroch) and an ectodermal invagination(shell gland), disrupt the outline of the embryo asit enters the trochophore stage. The prototroch iscentered on the animal pole; the shell glandappears on one side, nearer to the animal polethan to the vegetal pole (Figure \b,c). Authors do
pt
app
m
FIGURE 1.—Early development of Ostrea edulis, redrawn fromHorst (1883-1884) with presumptive dorsal region (shellgland and later the hinge) at top and anterior to right: a,gastrula; b, early trochophore; c, middle trochophore; d, latetrochophore; e, early veliger. (a = anus, aa = anterioradductor, ap = animal pole, app = apical pit, m = mouth,pmc = presumptive mantle cavity, pmo = presumptivemouth opening, pt = prototroch, s = shell, sg = shell gland,v = velum, vp = vegetal pole.)
not agree, however, on the order of appearanceof these structures. In Ostrea edulis, Davaine (1853)observed that the shell gland appears at the sametime as the earliest prototroch. Horst (1883-1884)showed the shell gland appearing even beforegastrulation is completed and well before theearliest appearance of the prototroch (Figure Ib),implying that the first appearance of the shellgland in Ostrea edulis is earlier than in otherbivalve larvae. Finally, Fernando (1931) showedthe prototroch appearing shortly after gastrula-tion and the shell gland definitely following theearliest appearance of the prototroch. In Crass-

SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
ostrea gigas, Fujita (1934) found that the shell-gland invaginates about the same time the blas-topore begins to close and before development ofthe prototroch.
Descriptions of the early prototroch also differ.Horst (1883-1884) showed that in Ostrea edulis theprototrochal cilia stem from a single ring of cells(trochoblasts), the center of the ring being asimple dome (Figure \c). Neither Davaine (1853)nor Horst observed any cilia on the dome,whereas Fernando (1931) illustrated an apicaltuft of cilia from the earliest appearance of theprototroch onward. In Crassostrea gigas, Fujita(1934) described an apical "flagellum" six orseven times as long as the surrounding velar ciliain veliger larvae that were in the early D-shapedphase of shell development (probably earliest pro-dissoconch II). This "flagellum" gradually be-came shorter relative to surrounding cilia as de-velopment proceeded, but Fujita did not specifywhether or not it eventually disappeared duringthe veliger stage. It is not yet known whetherthese developmental patterns represent real vari-ation due to environmental or genetic differencesor whether they are artifacts of techniques.
Subsequent changes in the external morphol-ogy of the trochophore and veliger involve eva-gination of the shell gland, secretion and enlarge-ment of a shell, formation of the anal tuft andanus, gradual elaboration of the prototroch intothe velum, and formation of the mantle cavity inwhich the foot and other structures develop (Fig-ures 1 d,e, 2, 3).
The structure of the shell gland and the earliestformed shell are among the most poorly knownaspects of larval morphology. Early authors de-bated whether the initial shell of a bivalve is asingle structure or a paired structure. These dif-ferences are readily resolved, however, when it isnoted that authors referring to a single structure,such as Horst (1883-1884), were referring to thefirst-formed uncalcified cuticle, whereas those re-ferring to a paired structure, such as Lacaze-Duthiers (1854) and Brooks (1880), were describ-ing the first calcified structures, which are indeedpaired (Raven, 1958). Obviously the organic
structure must precede calcification, for, as dis-cussed below and as noted by Carriker and Pal-mer (1979a) and Kniprath (1979), it is the per-iostracum that separates the microenvironmentfor initial calcification from the surrounding sea-water.
Secretion of the initial organic periostracum isby cells that border the nearly closed opening ofthe shell-gland invagination (Kniprath, 1979),and it seems that some trace of this earliest por-tion of the shell should be detectable on thesurface of prodissoconch I. Unfortunately, de-scriptions of the form of the invagination and itsopening are incomplete. Horst (1883-1884)showed the invagination as a transverse groove inOstrea edulis, attenuating at its ends, with theplane of the groove inclined to the surface of theembryo and passing inward toward the posterior(Figure Ib). Lacaze-Duthiers (1854) referred tothe calcified valves as being formed from twothickenings of the epiblast on each side of a dorsaldepression, and Brooks (1880) observed a similarorigin in the American oyster. The first-formedshell, including the initial organic cuticle (or per-iostracum) and the paired calcified valves, musttherefore be a transversely elongate structure,which indeed it is as revealed by scanning electronmicroscope.
When the shell gland, which is the rudiment ofthe mantle, actually becomes the mantle is amatter of definition made difficult by the factthat the change is gradual. It is implicit in theterms "gland" and "mantle" that the former isspecialized epithelium and the latter is a sheet-like fold of epithelium enclosing a mantle cavity.It is also generally understood that a "mantle"has a specialized edge having either a periostracalgroove or a commarginal band of specialized cellscapable of secreting a periostracum. Unfortu-nately, however, there are no detailed studies ofthe histology of the transition between shell glandand mantle in the Bivalvia, and it is necessary tolook to studies of gastropod larvae for a clearerunderstanding of the changes that take place.
Cather (1967), who extended the previous workof Raven (1958), found that in the gastropod

NUMBER 328
Ilyanassa, secretion of a thin membrane (initialperiostracum) by the shell gland begins in theinvaginated stage (see also Demian and Yousif,1973a, b). The membrane is at first attached bymicrovilli to the apical ends of all of the cells inthe invagination but subsequently lifts from thecentral cells and closes the opening of the invag-ination. From this point on, the periostracum isattached only to the leading edge of the shellgland. Calcification begins after the shell glandevaginates. The calcium carbonate is aragonite,which, as noted by Cather, is formed in a viscousmatrix that underlies the periostracum. After se-cretion of the first calcified shell, the central cellsof the shell gland change from columnar to squa-mous and no longer show evidence of secretoryactivity. The marginal cells, however, remain intheir original columnar, active state. It is at thispoint that Cather prefers to use the term "man-tle" rather than "shell gland," because an initialphase of calcification has been completed andthere is a clear differentiation in the cells whichcomprise the structure. Subsequent growth of themantle edge in the larva is by incorporation ofsurrounding ectodermal cells as well as by prolif-eration of the cells in the mantle.
Kniprath (1977) found a different ontogeny ofshell gland and mantle in the gastropod Lymnaeastagnalis. The initial periostracum is not secretedby the invaginated cells of the shell gland, butonly by the rosette of cells surrounding the invag-ination. Functional activation of other cells pro-ceeds from the peripheral rosette inward. Theinitial periostracum is therefore a ring consistingof the same type of material that in subsequentstages will form the outermost layer of the perios-tracum. More centrally located cells secrete innerlayers and close the central hole in the initialperiostracal ring. After evagination of the shellgland, the free mantle edge forms when the pe-ripheral cells of the gland arch upward and asecond arched ring of cells differentiates on theoutside of the first arched ring. These two ridgesbecome opposed to one another, forming theperiostracal groove between them.
Both of these patterns of development in gas-
tropods demonstrate that commarginal differen-tiation of secretory cells occurs very early in thedevelopment of the shell gland and that thechange from shell gland to mantle is gradual andtransitional. In contrast, students of bivalve lar-vae have long considered the shell-gland phase toextend through the prodissoconch-I stage of shelldevelopment and have assumed that the pro-dissoconch-I/II boundary represents an abruptchange from calcification by the shell gland tocalcification by the mantle (e.g., Bernard, 1896;Ansell, 1962; Ockelmann, 1965; Kume and Dan,1968; Calloway and Turner, 1979). This assump-tion, however, has been based on a change inexternal sculpture rather than on actual obser-vation of histological or shell ultrastructuralchanges. It will be argued in the present studythat the prodissoconch-I/II boundary representsnothing more than the onset of valve closure.
In oyster larvae, the calcified shell valves ex-tend laterally as the shell gland or mantle in-creases in area, and at the stage represented byFigure \d the valves cover nearly half the dorso-ventral dimension of the trochophore. The ciliaof the prototroch increase in size, but the pretro-chal region remains dome-shaped. According toHorst (1883-1884), it is at about this stage thatthe intestine first communicates with the exteriorby way of the anus.
At the point where the anus forms, a tuft ofcilia also appears on the epiderm. In fact, ciliationprobably precedes formation of the anus, asshown by Hatschek (1880) in Teredo, Meisenhei-mer (1900) in Dreissena, and Allen (1961) in Pan-dora. This ciliary tuft was unknown to Horst(1883-1884) and therefore is absent from FiguresId and \e. Dantan (1916) first noted its presencein the larva of the European oyster and describedit in detail, noting that short cilia surround theanus and that longer cilia occur just dorsal to theanus. He considered this "postanal ciliary crown"to be homologous with the telotroch generallypresent in trochophores of the mollusk-annelidline, and Fernando (1931) actually called the tuftthe "telotroch."
Elaboration of the prototroch to a velum in-

SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
volves five main events: (1) doubling of the ringof trochoblasts to produce a double row of cilia,(2) elongation and clustering ("compounding")of the cilia, (3) subsidence in the center of theapical dome to form an apical pit, (4) enlarge-ment of the tissue in the apical pit to produce anapical organ and eventually a cerebral ganglion,and (5) the formation of velar retractor musclesinserted into the shell and complexly ramified inthe velum. Aside from the apical cilia, authorsdiffer on whether or not cilia are present on theepiderm central to the preoral ring of locomotorycilia in the developing prototroch. Horst (1883-1884) and Fernando (1931) found none, but Dan-tan (1916) showed fine cilia present over theentire apical dome except at the center, where hemaintained that cilia are absent.
Authors have also differed in their ideas onhow the mantle cavity develops. Meisenheimer(1900) thought that it formed by an actual in-folding of the body wall. However, Erdmann(1935) maintained, probably correctly, that theprecursor of the mantle cavity is the flattenedposteroventral region of the trochophore lyingbetween mouth and anus. This area becomes themantle cavity not through any infolding but sim-ply because it is overgrown by advancing edgesof the mantle and shell that are curved ratherthan flattened.
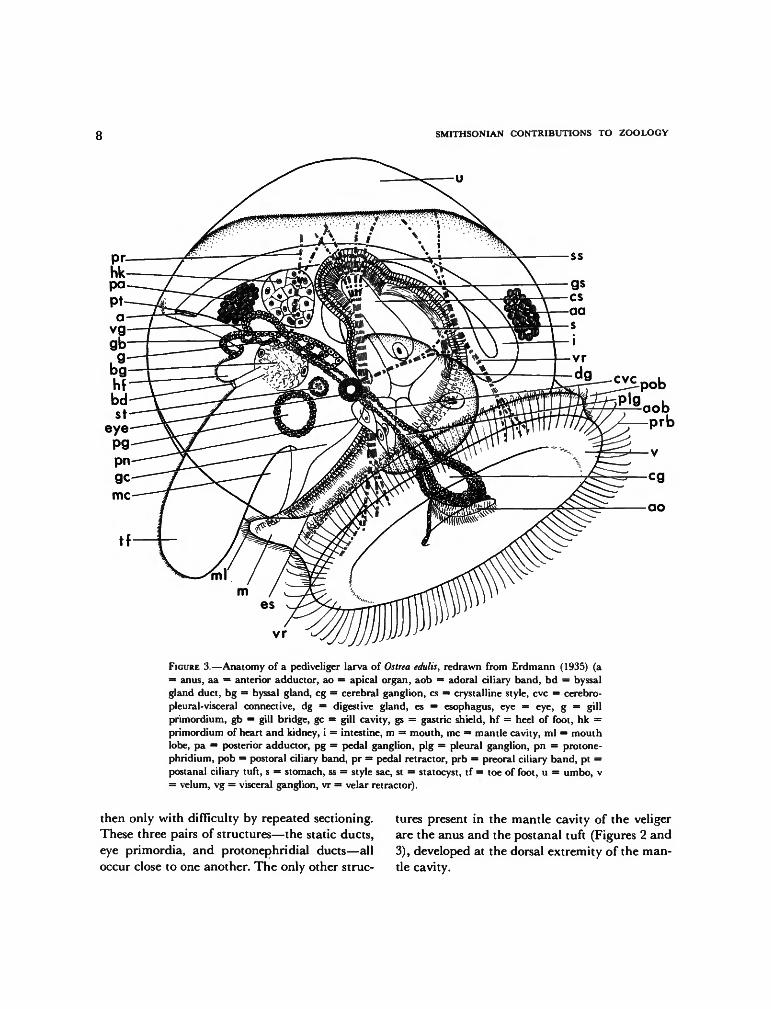
With the ontogenetic developments describedabove, the trochophore of the European oysterbecomes a veliger and is ready for release from itsparent. From this stage up to the end of larvaldevelopment, Erdmann's (1935) reconstructions,reproduced in Figures 2 and 3, are the mostinformative. External features of interest in thepresent study are the velum, mouth region, struc-tures in the mantle cavity, and the anal region.
In the newly released larva (Figure 2), Erd-mann distinguished only one clearly delimitedciliary band, preoral in position and containinglong, locomotory cilia stemming from a doublerow of large cells. The entire subvelar area wassaid to be covered with fine cilia that moveparticles indiscriminately toward the mouth. Inthe pediveliger stage (Figure 3), however, he was
able to distinguish three ciliary bands separatedfrom one another by narrow non-ciliated spaces:a preoral locomotory band, a subvelar adoralband of short cilia, and a second subvelar bandbearing longer cilia and said to be postoral inposition. Erdmann did not specifically refer tothe long cilia in the preoral band as being com-pound but clearly stated that they occur in tuftslying parallel to one another.
Erdmann also found that in both early andlate veligers a conical apical pit occurs slightlyposterior to the center of the velum. The posteriorslope of the pit is steep and without cilia, whereasthe anterior slope is shallow and bears short cilia.During development the epithelium underlyingthe anterior slope of the pit thickens and projectsfurther into the lumen of the velum, where it isunderlain by the cerebral (cephalic) ganglion.According to Erdmann, throughout the veligerstage the mouth is actually a part of the velarmass, occurring on a lobe on the posterior sub-velar region (Figures 2, 3). Some investigators(Horst, 1883-1884; Yonge, 1926, 1960) have con-fused this mouth lobe with the rudiment of thefoot.
There has been no general agreement on thenature of ciliation about the mouth. Dantan(1916) described only a postoral ciliary tuft,which he considered to be a remnant of thepostoral band found in some other bivalves. Erd-mann (1935), on the other hand, found no tuftbut considered the mouth to be surrounded bycilia of the adoral and postoral velar bands. Dan-tan (1916) also observed invaginations on theborders of the mouth which he interpreted to bebuccal glands.
Erdmann (1935) showed that the foot developsmedially from an outgrowth of the ectoderm ofthe body wall. In the newly released larva (Figure2), there is little external expression of the footprimordium other than an invagination markingthe site of the byssal gland. An elevated, ciliatedpapilla, the heel or metapodium, then appearsdorsal to the byssal-gland invagination. Later,the epithelium ventral to the byssal gland prolif-erates and grows outward to produce the toe or

NUMBER 328
ms
ss
aa
ao
FIGURE 2.—Anatomy of a young, newly released, six-day-old veliger larva of Ostrea edulis,redrawn from Erdmann (1935) (a = anus, aa = anterior adductor, ao = apical organ, bg —primordial byssal gland, dg = digestive gland, es = esophagus, m = mouth, mew = mantle-cavity wall, me = mouth embayment, ml = mouth lobe, ms = free mesenchymal cell, pn =protonephridium, s = stomach, ss = style sac, v = velum, vr = velar retractor).
propodium, which becomes a prominent struc-ture by the end of the veliger stage.
Soon after release from parent, deep gill cavi-ties or "kiemenhohlen" (Hatschek, 1880) form oneach side of the foot by infolding of the bodywall. A single gill primordium also forms on eachside as an ectodermal ridge of the mantle. As inall bivalve larvae thus far studied, this is theprimordium of the inner demibranch (e.g., La-caze-Duthiers, 1854; Rice, 1908; Raven, 1958;Knight-Jones, 1954b). Later, papillae which arethe primordia of the gill filaments form on theridges, and the ridges themselves elongate poste-riorly, nearly to the edge of the mantle. At thispoint in Ostrea edulis, the ridges on each side join
to one another by a bridge of epithelium (Erd-mann, 1935, and Figure 3). The outer demi-branchs do not develop until after metamorphosis(Yonge, 1926).
Medially, on each side of the base of the pri-mordial foot, tiny invaginations form the pairedstatocysts, which, according to Erdmann (1935),maintain openings into the mantle cavity onlyfor a brief time before being closed off. Theprimordia of the eyes also appear early as out-growths from the body wall in the gill cavitieslateral and slightly anterior to the base of thefoot. Lastly, paired protonephridia also appearvery early, but Erdmann could detect openingsto the exterior only late in the veliger stage and
SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
tf
ss
vr
FIGURE 3.—Anatomy of a pediveliger larva of Ostrea edulis, redrawn from Erdmann (1935) (a= anus, aa = anterior adductor, ao = apical organ, aob = adoral ciliary band, bd = byssalgland duct, bg = byssal gland, eg = cerebral ganglion, cs = crystalline style, eve = cerebro-pleural-visceral connective, dg = digestive gland, es = esophagus, eye = eye, g = gillprimordium, gb = gill bridge, gc = gill cavity, gs = gastric shield, hf = heel of foot, hk =primordium of heart and kidney, i = intestine, m = mouth, me = mantle cavity, ml = mouthlobe, pa = posterior adductor, pg = pedal ganglion, pig = pleural ganglion, pn = protone-phridium, pob = postural ciliary band, pr = pedal retractor, prb = preoral ciliary band, pt =postanal ciliary tuft, s = stomach, ss = style sac, st = statocyst, tf = toe of foot, u = umbo, v= velum, vg = visceral ganglion, vr = velar retractor).
then only with difficulty by repeated sectioning.These three pairs of structures—the static ducts,eye primordia, and protonephridial ducts—alloccur close to one another. The only other struc-
tures present in the mantle cavity of the veligerare the anus and the postanal tuft (Figures 2 and3), developed at the dorsal extremity of the man-tle cavity.

NUMBER 328
The structure of the shell of the veliger stage issomewhat better known than that of the pre-veliger shell. As is generally the case with bivalvelarvae having planktonic planktotrophic stages,a boundary line separates a pitted and radiallystriate prodissoconch I from a commarginallystriate prodissoconch II (Werner, 1939; Ockel-mann, 1965). As previously mentioned anotherprominent sculptural feature of the prodissoconchII of oysters is a posterodorsal notch and resultantspiral growth track (Tanaka, 1960; Waller, 1979;Carriker and Palmer, 1979a). The shell is oval inlateral view, with a shape somewhat like that ofa small Nuculacean clam. The hinge line is shortbut with strong interlocking hinge teeth in theprodissoconch-II stage. Some authors (e.g., Ran-son, 1960; Stenzel, 1971) have described a centralresilium (or internal fibrous ligament) as beingpresent between the teeth, but this is now knownto be incorrect (Pascual, 1971, 1972; Carriker andPalmer, 1979a). The resilium does not appearuntil at or near metamorphosis, and it is then atthe anterior edge of the anterior hinge teeth, notin the center.
The ultrastructure of the larval shell of Ostreaedulis has not been described, although Carrikerand Palmer (1979a) have dealt with the ultra-structure of the prodissoconch II of another spe-cies, Crassostrea virginica.
In Ostrea edulis, metamorphosis involves per-manent cementation of shell to substrate, disap-pearance of the velum, foot, anterior adductormuscle, and eyespots, appearance of labial palps,and elaboration of gills and mantle edges. Theseevents and changes have been thoroughly de-scribed by Cole (1938), Hickman and Gruffydd(1971), and Cranfield (1973a, b, c, 1974) andneed not be reviewed here. Relatively little atten-tion has been given to the abrupt changes in shellfabric and mineralogy that occur at metamor-phosis. Carriker and Palmer (1979a) showed thatin Crassostrea virginica the finely textured aragoniteof the prodissoconch changes abruptly to coarselyprismatic and foliated calcite at metamorphosis.Descriptions of dissoconch shell mineralogy and
fabric can be found in Galtsoff (1964), Taylor,Kennedy, and Hall (1969), and Stenzel (1971).
Materials and Methods
Two samples of living veliger larvae of Ostreaedulis were received from the Shelter Island OysterCompany, Greenport, Long Island, New York.The specimens of one sample were anesthetizedand fixed when they were still in an early pro-dissoconch-II stage, seven days from time ofrelease from parent. The specimens of the secondsample were anesthetized and fixed when theyhad reached an advanced prodissoconch-II stage,15 days after release.
Specimens were anesthetized by transferringthem to a 50-ml culture dish half filled withseawater and adding one or two drops of 2-phenoxyethanol (Eastman Kodak Co.) to the sea-water in the dish. These specimens, which werestill capable of swimming but incapable of re-tracting their velum, were then transferred to a2-ml dish and were killed by adding a drop offixative consisting of 2% glutaraldehyde bufferedwith sodium cacodylate (0.025 M in seawater)prepared according to the method of Turner andBoyle (1975). When the beating of velar ciliaceased, the seawater was drawn off and replacedby the fixative. Specimens were dehydrated witha graded series of ethanols and amyl acetate andcritical-point dried in a Denton DCP-1 unit.
Short pieces of No. 39 coated copper wire wereshaped so that each piece would stand on itscoiled base with an upright stalk. The critical-point dried specimens were placed on a cleanglass surface; the tip of the wire stalk was dippedin glue (polyvinyl acetate), which was allowed todry to a "tacky" consistency; and this glued tipwas then touched to a larva at a point whichwould allow the velum and gaping mantle cavityto be viewed with the scanning electron micro-scope. The wires bearing the larvae were then seton their bases on a glass coverslip and fastened tothe coverslip with a dilute water-soluble glue(trade name, "Elmer's Glue-all," Borden, Inc.).The cover-slip was then fastened to a standard

10 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
aluminum stub by means of a carbon suspension.Mounting the specimens on wires permitted sub-sequent reorientation on the stub simply by bend-ing the wire. The specimens were sputter-coatedunder vacuum first with a thin layer of carbonand then with gold-palladium and were studiedwith Coates and Welter 106B and CambridgeMark-IIA scanning electron microscopes.
Some of the fresh larval shells of Ostrea edulis,ethanol-preserved shells of Crassostrea gigas (Thun-berg, 1793), and dried postlarval museum speci-mens of Neopycnodonte cochlear (Poli, 1795) weremechanically fractured, cleaned ultrasonically inneutral distilled water, and mounted directly onglass coverslips. Other shells were etched in anoxygen plasma in a Tegal "Plasmod" low-tem-perature asher or were treated with commercialgrade laundry bleach (5% NaOCl) to remove theperiostracum. During the period of nearly twoyears during which this study was in progress,critical-point dried specimens were stored in aglass silica-gel dessicator and showed no evidenceof deterioration or change in appearance at thelevels of magnification of the scanning electronmicroscopes.
The illustrated specimens of larval shells ofOstrea edulis and Crassostrea gigas were numberedto permit the reader to determine whether or notthe micrographs are of the same or differentindividuals. Each microscope stub was given aseparate number, and if the stub contained morethan one specimen, each specimen was lettered.These specimens will not become part of thepermanent collection of the Museum because oftheir small size, fragile mounts, and likelihoodthat the dried tissues will deteriorate. Specimensof Neopyonodonte cochlear from the museum collec-tions bear USNM (National Museum of NaturalHistory, formerly United States National Mu-seum) catalog numbers.
Terms, Measurements, and Limitations
The terms "prodissoconch I" and "pro-dissoconch II," first used by Werner (1939), referto successive stages of development of the larval
shell as well as to the shell itself. The pro-dissoconch-I stage extends in time from the be-ginning of calcification to the end of developmentof prodissoconch I. The end of this stage on theshell exterior is indicated by a more or less abruptchange in sculpture from faintly radially striatewithout prominent commarginal growth ridgesto commarginally ridged without significant ra-dial striae. In oyster larvae, this boundary alsocoincides with the origin of the posterodorsalnotch and its growth track. The prodissoconch-IIstage extends in time from the end of the pro-dissoconch-I stage to metamorphosis and the be-ginning of the postmetamorphic shell, the "dis-soconch." The prodissoconch-II shell includes theentire prodissoconch-I shell plus whatever newgrowth has been added, both to the peripheryand to the interior surfaces. The dissoconch stagespans the entire time from metamorphosis todeath. The relationships of these terms to otherscurrently in use are shown in Figure 4.
Directional terms are with respect to the larvalshell. "Dorsal" is toward the larval hinge, "ven-tral" away from the hinge; "anterior" and "pos-terior" refer to directions parallel to the hingeand toward the sites of the anterior and posteriorlarval adductors. "Proximal" and "distal" havebeen used in the customary anatomical sense,meaning toward earlier formed parts and towardlater formed parts, respectively. "Apical" and"basal" are used as in histology, meaning awayfrom or toward the basal lamina of an epithelialcell. "Commarginal" refers to sculptural or struc-tural features of the shell that are parallel to theshell margins or to previous traces of the shellmargin; the term is being increasingly used inmolluscan morphology in place of the geometri-cally inaccurate term "concentric." "Radial" re-fers to elements of the shell that radiate from theumbo and are approximately perpendicular tothe shell margin. "Length" is the anterior-poste-rior dimension of the shell parallel to the hingeline; "height" is the dorsal-ventral dimension per-pendicular to the hinge. "Convexity" (= depth)is the maximum transverse width perpendicularto the plane of commissure.

NUMBER 328 11
TERM
EGG
BLASTULA
GASTRULA
TROCHOPHORE
VELIGER
D-SHAPED STAGE
UMBO STAGE
PEDIVELIGER
SPAT
PRE-SHELLEDSTAGE
(LESS THAN1 DAY)
—
PRODISSOCONCH 1(ABOUT 7 DAYS)
PRODISSOCONCH II(10-15 DAYS)
DISSOCONCH(REMAINDER OF LIFE)
FIGURE 4.—Relationship of stages of development of the prodissoconch to other common termsused to describe the larval shell and body of Ostrea edulis (dashed lines indicate uncertainty ortransition; durations of stages may be highly variable).
Measurements based on scanning electron mi-crographs are subject to inaccuracies due to thetremendous foreshortening of depth of field andthe difficulty of knowing whether the end-pointsof a measurement are in the plane of focus. Theaccuracy of observation may also be affected bythe state of preservation of the specimen. Chem-icals used in preservation and preparation maycause slight etching or rearrangement of shellmicrostructural components. In general, however,calcified surfaces described herein have similarappearances regardless of mode of preparation(e.g., air-dried without preservation in alcohol,air-dried from alcohol, critical-point dried,plasma etched). An additional inaccuracy maybe introduced by the shrinkage of soft tissuesduring dehydration and critical-point drying (es-timated by Boyde, 1978, to be up to 25% linear).As noted in a previous study (Waller, 1980), thisproblem is not serious at the levels of interpreta-tion involved, and shrinkage-induced fracturingalong cell boundaries may be useful in that itreveals the outlines of cells and the relationshipbetween cell surfaces and adjacent structures.
Finally, cultured specimens may differ fromindividuals living under natural conditions, andboth genetic and environmental factors may af-fect larval morphology (Culliney, Boyle, andTurner, 1972). In the present study, the larvae of
Ostrea edulis are from the same parents, and thedegree to which morphological features and de-velopmental patterns in different breeding stocksvary has not been investigated.
Observations and Functional Interpretations
BEHAVIOR
Erdmann (1935) described the behavior of ve-liger larvae of Ostrea edulis, but certain observa-tions relevant to functional morphology and therelationship of body and shell are repeated here.Swimming larvae seven days after release fromparent were in an early prodissoconch-II stage,with straight dorsal margin unobscured by um-bones, valves of about equal convexity and 200to 210 /im in length, no apparent foot, and noeyespots (Figure 5).
When the valves were closed, the larvae weredistinctly negatively buoyant and lay on the bot-tom of the container with their infolded velarcilia quiescent. Because valves were transparentat this stage, the velum, velar retractors, androtating stomach contents were visible. Rotationof stomach contents in all specimens observedwas clockwise when viewed from the anterior. Aspointed out by Millar (1955), anti-clockwisemovement is inhibited by the internal profile ofthe stomach floor.

12 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
FIGURE 5.—Transmitted-light micrograph of living Ostreaedulis in early predissoconch-II stage (about X 350, bar = 20Urn).
When swimming was about to begin, the valvesgaped slightly, and velar cilia began to vibrateand protrude from between the valves. Mostspecimens extruded and unfolded their velumvery rapidly. Other specimens, possibly sick orinjured, extruded their velum but did not unfoldit, which caused them to spiral on the bottom ofthe container. When the velum unfolded com-pletely and became an oval disk, the velar (ante-roventral) side of the larva rotated upward andthe larva swam upward from the bottom. Thedirection of movement was with the anterior endfirst, with the plane of the velar crown slightlydeclined toward the anterior. An apt analogy isthe flight of a helicopter, which is pulled upwardand forward by its rotor. As noted by Knight-Jones (1954a), larvae of Ostrea edulis viewed fromabove spiral in a clockwise direction as they moveforward.
When the velum was extended but still folded,so that the shell was still on its side, it could beseen that the gut and stomach were pushed an-teroventrally with the velum to a position in the
ventral half of the shell. The dorsal half was thena void except for the posterior portion of the gut,which remained relatively fixed in position. Infact the anus showed no movement at all relativeto the shell margin during extrusion of the velum.Velar retractors crossing the void converged totheir insertions in the velar mass and branched totheir insertions near the dorsal margins of thevalves.
On the posterior side of the velar crown, thelobe containing the mouth was visible in someliving specimens. The ventral side of the mouthwas bordered by a tuft of relatively inactive andstiff cilia, and a fine stream of fluid interpreted asmucus could be seen exiting posteriorly from thistuft. Yonge (1926) noted that surplus food whichis not ingested leaves by the same route as thisstream, "so that a larva swimming through athick suspension of food leaves behind it a trail ofparticles embedded in a long string of mucus."
All views of swimming larvae were necessarilyventral because of their consistent orientationwith the velar crown upward. By focusing belowthe velum on a point along the posterodorsal shellmargin, however, it was possible to observe defe-cation by active specimens. Feces were in theform of tiny rods or strings, and their forcefulexpulsion appeared to be directly from the anus.No rolling of fecal material in the mantle cavityby cilia, which was suggested by Yonge (1926),was observed. The trajectory of expelled feces didnot lie in the plane of commissure but rather wasin a posterior direction slanting from the left sideof the veliger toward the right side.
Larvae 15 days beyond release from parentreached 280 fim in shell length and had valvesthat were coiled sufficiently so that umbones werebeginning to obscure the hinge. The left valvehad also surpassed the right in convexity. Nosignificant behavioral modifications were ob-served, except that when the velum was with-drawn and the valves were shut, the larva wasmore likely to land on its larger, left valve. Thesespecimens were not yet in the pediveliger stage.Although the foot was present in most specimens,it was not yet sufficiently developed for crawling.

NUMBER 328 13
TABLE 1.—Transitional stages in development of observed specimens of Ostrea edulis from neartime of release from parent (early prodissoconch II) to near time of appearance of pigmentedeyespots (late prodissoconch II)
Growth
stage
1
23
4
5
6
7
8
9
Approximate
shell length(in microns)
190
200250
250
265
280
290
300
310
Foot
heel only, no toeheel only, no toefaint toe primordium
toe primordium is aprominent bulge
toe beginning toproject from mantle
toe of moderatelength, unciliatedand without cement-gland opening
toe of moderatelength, ciliatedalong midline, withcement-gland ductopening
elongate toe,medially ciliated
toe furtherelongated, ciliatedon all surfaces andcapable of lateralmovement
Gill bridge
absentabsentciliary tufts on
posterior mantleedge, unfused
ciliary tufts, unfused
ciliary tufts, unfused
ciliary tufts, unfused
weak ciliary fusion
weak tissue fusion inaddition to ciliaryfusion
epithelial bridge
Few of the specimens had pigmented eyespots,but black pigment was present around the baseof the velum and on the ventral mantle edges.
MORPHOLOGY
Length of time from spawning is not an accu-rate predictor of developmental stage. Erdmann(1935) found that rate of development is stronglyinfluenced by temperature, and Culliney, Boyle,and Turner (1972) stressed the variability of cul-tured populations. It is not surprising, therefore,that the two samples of larvae seven and fifteendays from release contained a variety of growthstages and that these stages actually overlapped
between the samples. Because there is little vari-ation in relative rates of development of oneorgan with respect to another, these stages rep-resented a continuum of development. To facili-tate description in the present study, however,this continuum was divided arbitrarily into ninestages, which are outlined in Table 1. Figures 9through 84 are arranged in sequence accordingto these stages.
The following observations are presented in theform of a travel with the scanning electronmicroscope, starting with the velum, plungingdorsally into the mantle cavity, then emerging onthe posterior side of the hinge and passing acrossthe exterior of the shell. Next, the microstructureof the shell is viewed by means of fractures, and

14 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
then the relationship between mantle and shelland the inner shell surface, particularly the hingeapparatus, are examined after the removal of softtissues. The final section deals with changes ob-servable in the early dissoconch following meta-morphosis.
Velum
During growth of the veliger, the velar crownretains its oval shape but increases in size, ranging(in micrographs of fixed specimens) from a lengthof about 150 jum and width of 110 jum in Stage 1(Figure 19) to about 200 by 140 /im by Stage 3(Figures 33, 34). (Fully extended vela were notobserved in later stages of fixed specimens.)
The central region of the velar disk bears adeep apical pit, which in surface view is nearlycircular with a diameter of 10 to 15 ftm (Figures21, 30, 37). In all cases the pit slopes toward theposterior so that its anterior slope is shallow andposterior slope steep, as shown in Figure 30 andalso by Erdmann (1935, Figures 2 and 3 herein)in longitudinal section. A tuft of from 20 to 100fine cilia 6 to 8 /an in length stems from theanterior floor of the pit. These cilia barely projectabove the sides of the pit, which is itself in thecenter of the broadly concave central region ofthe velar disk. No ontogenetic trends in the size,depth, or degree of ciliation of the pit could bedetected from the surface views, nor was it possi-ble to observe the floor of the pit. According toHickman and Gruffydd (1971), the very center ofthe floor of the pit is occupied by the apicalorgan, which differs in cellular arrangement fromthe surrounding velar tissue.
In some bivalve groups, particularly the Ve-neridae, a long apical flagellum or tuft of verylong cilia is present throughout larval life and issaid to be sensory (Ansell, 1962; see also Boyleand Turner, 1976). It is tempting to postulate asimilar function for the short apical cilia of Ostreaedulis, because they are unsuited for locomotionand food-gathering and are underlain by thecerebral ganglion (Hickman and Gruffydd, 1971).
The epithelium surrounding the apical pit isdevoid of cilia and bears microvilli, some 5 or 6per micron, over its entire surface (Figures 19, 21,30, 31, 34, 37). In general, the apical surfaces ofthe cells comprising the epithelium of the disk areflat, except immediately surrounding the apicalpit, where they are commonly strongly tumescent.A mucus-like substance coats the tips of the mi-crovilli, and where microvillous surfaces of adja-cent cells come into contact the microvilli appearto adhere to one another (Figure 21).
The periphery of the velar crown is girdled byfour bands of cilia (Figures 6, 31, 33), not three asdescribed by Erdmann (1935). Two of the bandsare preoral in position, one is adoral, and thefourth is postoral.
The inner preoral band (Figure 31), describedhere for the first time, is about 8 to 10 jum inwidth and forms a complete oval separated fromthe outer preoral band by a nonciliated space
opc
aoc
FIGURE 6.—Diagrammatic cross section of right half of velumof Ostrea edulis viewed from anterior (outer preoral cilia andpostoral cilia are at bottom of effective beat; arrows showdirections of metachronal waves; ac = apical cilia, aoc =adoral cilia, ipc = inner preoral cilia, opc = outer preoral(locomotory) cilia, poc = postoral cilia, sh = shell).

NUMBER 328 15
that varies in width depending on the degree ofextension of the velum. The dense cilia of theband are no longer than about 20 jum and ran-domly arranged, showing no clumping or fusionnor any indication of coordinated metachronalmovement. The band is indistinct and possiblyincompletely developed in Stage 1, fairly distinctby Stage 2, and apparently fully developed byStage 3.
The function of the inner preoral band is un-known. Both the size and arrangement of its ciliaresemble those of the adoral band, which receivesfood particles from the compound cilia of theouter preoral band and transports them to themouth (Yonge, 1926). Although the inner preoralband is in a position to receive food particlesdirectly from the water that impinges on thesurface of the velum during swimming, there isno connection with the mouth because the outerpreoral band intervenes. If the inner preoral bandis involved in food-gathering, it would have topass the food particles to the compound cilia ofthe outer preoral band, which would then transferthe particles to the adoral band, whence theywould pass to the mouth. A more likely functionalinterpretation is that the band is an upcurrenttactile receptor.
The outer preoral ciliary band, which is con-cerned with locomotion and feeding, dominatesthe velum because of its width and the size andcomplexity of its cilia (Figures 9, 19, 28, 31-34).The cilia are clustered in a manner that has ledsome investigators to refer to them as "compoundcilia", consisting of numerous single cilia whicharise from a limited area of the same cell, contactone another a short distance above their bases,and remain in contact for nearly their entirelength before splaying apart at their tips. Totallength of a cluster is in the neighborhood of 50 to70 jum, and the number of individual cilia in onecluster can only be guessed from the scanningelectron micrographs—perhaps 20 to 80. Erd-mann (1935) determined from sections that infact the outer preoral band is a double row ofciliary clusters stemming from a double row oflarge cells.
Knight-Jones (1954a, b) found a common planin the locomotory bands of many molluscan ve-ligers, including both bivalves and gastropods. Inswimming position the effective beat of the loco-motory cilia is downward over the edge of thevelum. The beats are coordinated so that metach-ronal waves are generated along the band ap-proximately at a right angle to the plane of beatof each cilium. These "diaplectic" waves aretermed "dexioplectic" if the effective beat of thecilia is to the right of the direction of propagationof the metachronal wave and "laeoplectic" if thebeat of the cilia is to the left. Knight-Jones ap-plied the same terms to the orientation of therows of cilia in each compound cilium, which hefound were always aligned parallel ("orthoplec-tic" rows) or perpendicular ("diaplectic" rows) tothe plane of beat. The functional outcome of thisarrangement is that diaplectic metachronism per-mits elongation of cilia which, because of closecontact or fusion, can flex more strongly. Thespacing pattern of the cilia relative to the direc-tion of propagation of metachronal waves mini-mizes interference between cilia, and the effectivebeat of the ciliary clusters generates a thrustwhich propels the veliger upward with the velarcrown leading.
Metachronal waves, which pass continuouslyaround the oval locomotory band, cause the ve-liger to rotate in a direction opposite to thedirection of travel of the waves (Knight-Jones,1954b). As noted by Knight-Jones, this rotationis countered in two ways. First, the shape of theveliger itself may produce a drag that results ina speed of rotation which is slower than the speedof the metachronal waves. Secondly, the beat ofthe locomotory cilia may not be perpendicular tothe velar margin and to the direction of move-ment of the metachronal waves, but rather thecilia may beat in a plane oblique to the marginsuch that rotation is slowed or even completelycountered (Figure 7).
Scanning electron microscopy revealed that inOstrea edulis the pattern of compound cilia in theouter preoral band is as predicted by Knight-Jones (1954b). In each compound cilium the

16 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
V- DIRECTION OF PROROGATION OF METACHRONAL WAVES
^ " * k DIRECTION OF ROTATION OF LARVA
FIGURE 7.—Theoretical relationships of laeoplectic meta-chronal waves, obliquity of locomotory cilia, and rotation oflarva during swimming: a, cilia strictly diaplectic, rotationof larva produced by metachronal waves; b, obliquity of ciliareinforcing rotation produced by metachronal waves; c,obliquity of cilia countering rotation produced by metach-ronal waves.
constituent cilia are arranged in long orthoplecticand short diaplectic rows (Figure 32). The num-bers of cilia involved cannot be determined with-out sectioning, but it appears that 10 to 15 arealigned in the orthoplectic rows. The numbers ofcilia in the diaplectic rows appear to increasefrom the inner edge of the band toward the outeredge, suggesting that the orthoplectic rows di-verge in that direction. Contact between ciliaappears to be closest within the diaplectic rows.
Knight-Jones (1954a, b) observed that in Ostreaedulis the metachronal waves are laeoplectic.When the velum of swimming larvae is viewedfrom above, metachronal waves move in a clock-wise direction, but the entire specimen rotates inthe same direction. It can therefore be predictedthat the effective stroke of the cilia lies in a planeoblique to the velar margin as shown in Figure7c. This is exactly the pattern shown by thelocomotory cilia of each of 12 specimens fixedwith velum extended (Figure 19). It appears thatthe obliquity and intensity of the beat of thelocomotory cilia thus more than counter the ro-tational force imparted by metachronal waves.
The outer preoral band is separated by a nar-row nonciliated space, 4 or 5 /im wide, from theadoral band, which leads to and surrounds themouth (Figures 11, 31, 33, 39). Like the innerpreoral band, the adoral band in fixed specimensconsists of randomly arranged short cilia approx-
imately 8 jum in length that show no evidence ofcoordinated movement. Even though the adoralband approaches 20 p in width, it is completelyoverlapped by the compound cilia of the outerpreoral band when they are at the bottom of theireffective beat. Yonge (1926) found that food par-ticles are thrown by the large cilia of the innerpreoral band onto the adoral band where theyare embedded in mucus and carried back to themouth. Thus, the direction of the currents gen-erated by adoral cilia would run from anterior toposterior on each side of the velum.
The postoral band of cilia, first described byErdmann (1935), is actually a single row of com-pound cilia along the dorsal edge of the adoralband (Figures 6, 28). Each compound cilium is15 to 20 jum long and consists of a single ortho-plectic row of four or five cilia. Some of thescanning electron micrographs (e.g., Figure 28)suggest that the effective beat of these cilia istoward the adoral band and counter to the beatof the outer preoral, locomotory cilia (Figure 6).Strathmann, Jahn, and Fonseca (1972) foundthat such "opposed band" systems are commonciliary feeding mechanisms in molluscan veligers.By means of high-speed cinefilms, they deter-mined that both the preoral and postoral bandsare essential for feeding. The postoral cilia permitincreased movement of the preoral cilia relativeto the water in the latter half of their effectivestroke, and it is at this point that most of theclearance of particles from the water occurs. Theyalso found that some veligers could continueswimming without feeding, presumably by stop-ping the beat of the postoral cilia.
If the idea of Knight-Jones (1954b) that thedirection of diaplectic metachronal waves is con-stant in an individual regardless of function iscorrect, then other functions for the postoral bandare suggested. Metachronal waves passing alongthis band would be laeoplectic and would run ina direction opposite to that of the metachronalwaves of the outer preoral band because thedirections of effective beat are opposite. Thisopposition of wave patterns may facilitate clean-ing of the preoral cilia by the postoral cilia. It is

NUMBER 328 17
also possible that cilia of the postoral band mayserve to counter and adjust the rate of rotationimparted by the outer preoral band.
At the posterior end of the velar disk, thepostoral band passes posterior to the mouth andappears to merge with the postoral tuft. The ciliacomprising this tuft (Figures 11, 20, 25, 33, 39)tend to be more densely clustered and appear tobe more rigid than those of the postoral band.Also, a portion of the tuft is out of line with thepostoral band, being distinctly more posterior inposition. These observations suggest that the pos-toral tuft is a distinct structure, a view also heldby Meisenheimer (1900) and Dantan (1916). Itdoes not appear to be merely the posterior edgeof the postoral band as described by Erdmann(1935). The relative rigidity of the cilia in thepostoral tuft suggest that it may serve a sensoryfunction. It may also facilitate the posteriorwardstreaming of excess mucus and food, because it isat the point of egress of this stream as observedby Yonge (1926) and in the preceding section onbehavior.
The cells that comprise the epithelium of theentire subvelar region are large and devoid ofcilia. Like the epithelium of the apical disk, how-ever, the surface is microvillous, the tips of themicrovilli appear mucus-covered, and the micro-villi of adjacent cell surfaces adhere to one an-other where these surfaces are brought into con-tact in folds or between tumescent cells (Figure27). No sensory structures nor any glandularducts were detected. The anterior (Figures 18, 26,29) and lateral (Figures 43, 44, 47, 48) subvelarsurfaces merge with the mantle, and the posteriorsurface of the stalk (Figures 11, 33, 39) passesdorsally into the mantle cavity.
Foot
The first structure to be encountered in follow-ing the body wall dorsally from the mouth is theprimordium of the foot. Basically, the mode ofdevelopment, with the heel first and then the toe,is as described by Erdmann (1935), but scanningelectron microscopy allows the pattern of devel-
opment of pedal cilia and some of the secretoryducts to be seen. (See Table 1 for the nine stagesof development.)
In Stage 1, the heel of the foot is alreadypresent as a small protuberance from the bodywall midway along a medial line passing fromthe mouth to the hinge (Figures 11, 14). It istransversely elongated and dorsoventrally com-pressed, about 26 by 9 jum, and is densely ciliatedon its medial, ventral surface. The nonciliateddorsal surface of the heel rises sharply from thebody wall. The ciliated ventral surface is moregently sloping and at its base bears a pair of largeciliated duct openings (Figure 15). The totaltransverse diameter of this double opening is 13/un, each duct opening being about 7 jum. Theseare the openings of the byssal gland complex,which Erdmann (1935) showed to be the first ofmany organs to form on the ventral side of thelarva. Although Erdmann referred only to a singlegland, Cranfield (1973a) demonstrated that bythe pediveliger stage several types of glands thatshare a common ciliated duct in this location arepresent. The double opening may represent anearly stage in the development of the complexbefore a common duct is formed.
By Stage 3 the primordium of the toe hasappeared on the ventral side of the heel, and bothstructures push out together from the body wall.The process is due to proliferation of epithelialcells as shown by Erdmann (1935), not to infold-ing of the body wall on each side as supposed byMeisenheimer (1900). Like the heel, the toe is atransversely elongated ridge, but it is larger (50by 25 /tm), has more gently sloping dorsal andventral surfaces, and lacks cilia (Figures 40, 42).
In Stage 4, the heel and toe are more stronglyelevated from the body wall, and the byssal-ductopening is clearly single, measuring 7 /im in trans-verse diameter. There are still no cilia on the toeprimordium. In Stages 5 and 6, the ventral slopeof the toe steepens as it begins to bulge ventrally,but there are as yet no cilia (Figure 52). By Stage7, the toe of the foot has become a ventrallyprojecting, somewhat flattened cylindrical struc-ture bearing cilia along its midline (Figure 58).

18 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
This band of cilia is narrow adjacent to the heel,broadens toward the tip of the toe, and remainsbroad as it rounds the tip onto the newly formedanterior surface that faces the inner wall of themantle cavity. By this stage a second duct open-ing, about 1 /im in diameter, has appeared some-what to the left of the midline of the foot (Figure58). This is the opening for gland types Cl andC2 of Cranfield (1973a), which will be involvedin secretion of cement after the larva settles(Cranfield, 1975).
By Stage 8 the toe has lengthened into astrongly ventrally projecting tapered cylinderwith a bluntly rounded tip (Figure 68). The entirefoot measures about 75 /im in length and about53 jum in width near the heel. Cilia are stillrestricted to a medial band on the heel andproximal portion of the toe, but the band hasbecome wider near the tip of the toe, coveringnearly the entire width. The opening of gland-types C1 and C2, which has widened to a diam-eter of about 4 jum, is located at about one fourththe distance from the byssal-duct opening to thetip of the toe. The axis of the toe is still in themedian plane, and there is no evidence of com-plex foot torsion or extension in the micrographs.
By Stage 9, the foot has about the same diam-eter as in Stage 8 but has lengthened to about 90/im1 and shows evidence in the micrographs ofbeing able to pivot on its base (Figure 78). Densecilia now cover nearly the entire sole. Althoughthe size of the foot suggests that it can nowprobably be extended outward between thevalves, observations of living specimens from thesame sample, 15 days from release, revealed nonewhich had entered the pediveliger stage with afully functional foot. All of the cell surfaces of thefoot bear microvilli like those of the velum.
Structures in Gill Cavities
Lateral to the foot, the inner wall of the mantlecavity is deeply recessed to produce a pair of
1 Because the velum of the specimen shown in Figure 78is partially retracted, it is likely that the foot is also.
ectodermal pockets. These pockets were called"kiemenhohlen" (gill cavities) by Erdmann(1935), because they contain the anterior ends ofthe primordial gills. In addition to the gill pri-mordia, which are described below, three pairedstructures develop in each gill cavity: (1) theopenings of static ducts, (2) photoreceptors (calledeyespots after they become pigmented in moreadvanced larvae), and (3) structures which arepossibly the openings of protonephridial ducts(Figures 8, 43-50). The relative positions of thesestructures are shown diagramatically in Figure 8,which is drawn from a specimen in developmen-tal Stage 3, where the structures are best exposed.(See Table 1 for an outline of developmentalstages.) The structures as they appear in thisspecimen will be described first, followed by de-scriptions of other stages of development.
STATIC-DUCT OPENINGS.—The interpretation ofthe first pair of structures as openings of static
FIGURE 8.—Diagrammatic posterior view of body wall inmantle cavity of Ostrea edulis veliger, Stage 3 (a = anus, bp= boundary between prodissoconch I and II, bdo = byssal-gland duct opening, eye = microvillous eye primordium, hp= heel primordium, pdn = posterodorsal notch, pdo =protonephridial duct opening, pt = postanal tuft, sdo =static duct opening, tp = toe primordium).

NUMBER 328 19
ducts, which connect statocysts to the exterior, isbased on correspondence in size and position tostructures so described by Erdmann (1935) insectional views. These openings are the closest ofthe three structures to the midline, each openingbeing a circle 7 to 9 jum in diameter lying on themedial side of the gill cavity at the lateral edge ofthe toe primordium. Short cilia 2 to 4 jum inlength originating in the walls of the duct occupyits lumen and extend across its opening, meetingat the center. The cilia must transport fluid alongthe axis of the duct. The majority of micrographsshow that the concave side of curved cilia istoward the interior of the duct; straight cilia thatare probably fixed in their effective-beat positionalso show slight flexure at their base toward theinterior of the duct (Figures 16, 45, 46, 49, 50, 54,57). In view of the different forms of cilia ineffective and recovery beats outlined by Gray(1928, 1930; see also the reviews by Sleigh, 1962,1974), these observations suggest that fluid istransported through the duct from the outsideinward toward the statocysts.
In the Stage-3 specimen, a second incompletering of long vibratile organelles, interpreted ascilia rather than flagella (see following interpre-tation), borders the ventral edge of the ductopening. These cilia extend about 100° of arcaround the periphery of the left duct and onlyabout 45° around the periphery of the right.They reach about 14 jum in length and are there-fore much longer than the cilia in the duct open-ing. Rather than extending over the duct open-ing, the long cilia extend ventrally along theepithelium of the mantle-cavity wall.
The question arises whether these long vibratileorganelles are cilia or flagella. Although the in-ternal 9 + 2 structure is known to be the same inboth types of organelles, the direction of move-ment of a cilium is perpendicular to its long axisand parallel to the surface of the cell that bearsit, whereas a flagellum generally moves in a seriesof waves that pass from its base upward along itslong axis, thus moving fluids parallel to its length(Sleigh, 1962). In the Stage-3 specimen, the po-sition of the long organelles is not diagnostic but
suggests flagellate movement parallel to the cellsurface, thereby pulling water from over the ductopening and transporting it ventrally toward theposterior edge of the mouth lobe. Specimens indevelopmental Stage 8, however, show these or-ganelles in a flexed position, bent over the static-duct opening (Figure 74). Assuming that thisposition is not an artifact of preparation, then itappears that the long organelles are cilia. Ratherthan pulling water over the duct openings, theywould then push water over and possibly into theopenings.
Ontogenetic variation in the static-duct open-ings entails an apparent increase in diameter,ranging from less than 6 [im in Stage 1 (Figure16) to about 11 jum in Stage 7 (Figures 56, 57).The presence and absence of the long marginalcilia suggest individual variation or possibly lossof cilia during fixation. In Stage 3 describedabove, long cilia are present on both duct open-ings (Figures 45, 46, 49, 50); in Stages 1 (Figure16) and 7 (Figures 56, 57) they are lacking onboth openings; in Stage 5 (Figures 53, 54) theyare present on the left opening but not on theright; and in Stage 8 (Figures 73, 74) they arepresent on the right opening but not on the left.In Stages 4 and 9 long cilia are present on theright side, but the left side was not observable.
In the present study, static-duct openings canbe demonstrated to be present from the time ofrelease from parent to a stage just before thepediveliger in which there is still no evidence ofclosing. The openings thus appear to persistlonger than observed by Erdmann (1935), whothought that they constrict early in the veligerstage and finally close as the statocysts migrateinward from the epithelium. Only closed stato-cysts are present in mature oysters (Carazzi,1902).
A common assumption is that the static ductsof invertebrates are excretory (e.g., Barber, 1968).However, the configuration of the cilia borderingthe openings and lining the walls of the staticducts of Ostrea edulis suggest that particles maypass inward from the exterior toward the stato-cysts. This may also be the case in some other

20 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
species. Cragg and Nott (1977) found inwardlypointing cilia in the static ducts of pediveligerlarvae of Pecten maximus and suggested that sta-toconia of variable appearance may be intro-duced into the statocyst from the exterior via thestatic duct (see also Buddenbrock, 1915, andPlate, 1924).
PHOTORECEPTORS.—The second structure in thegill cavity is interpreted as an early developmen-tal stage of the larval eye, because its position (onthe medial wall of the gill cavity anterior to thestatic-duct opening and ventral and medial to theanterior extremity of the primordial gill axis)exactly corresponds to the position of the pig-mented eyespot in the later pediveliger stageillustrated by Erdmann (1935). The flower-likestructure is raised, hemispherical, approximately6 /im in diameter, and is covered with hundredsof hair-like organelles. These organelles are inter-peted as microvilli rather than cilia because theyare only about 0.1 jum in diameter and no morethan 1 jum long. The stem of the structure isobscured but is clearly narrower than the hemi-spherical head.
The primordial eye seems to show a slightontogenetic increase in size, ranging from lessthan 6 /im in diameter in Stage 1 (Figure 16) tonearly 9 /im in Stage 7 (Figure 59), and alsobecomes more widely separated from the static-duct opening. By Stage 8, however, the structureappears submerged in the epithelium so that itsapparent diameter at the epithelial surface issmaller (about 4 ftm), and its microvilli are nolonger as strongly projecting.
These changes are in accord with Erdmann's(1935) finding that the eye primordium in aseven-day old veliger consists of a thickening ofthe epithelium of the gill-cavity wall which pro-duces a bulge on the epithelial surface. The sub-mergence noted herein in Stage 8 seems to signifya trend toward the conditions found by Cole(1938) in the later, pediveliger stage. By that timethe eye becomes an almost spherical cup of pig-mented epithelium filled with a gelatinous matrixand having its aperture closed by a lens-like body.The early microvillous nature of the eye primor-
dium suggests that the larval eyes are of rhabdom-eric rather than ciliary origin (Eakin, 1965). Therhabdomeric type is prevalent in the Mollusca.
PROTONEPHRIDIAL-DUCT OPENINGS.—The thirdstructure, which is possibly the opening of aprotonephridial duct, is exceedingly difficult toresolve in the specimens observed, because it liesdeep within the gill cavity and is always partiallyobscured by the primordial eye. Basically, thestructure appears to be a small tuft of eight toten cilia, which are 3 to 4 jum in length, lying onthe dorsolateral side of and nearly in contact withthe eye (Stage 3, Figures 45, 49). The cilia arisefrom a small depression about 2 pirn in diameter,but it is not possible to determine whether it isthe opening to a pit or duct. The bases of the ciliaare perpendicular to the subjacent epithelial sur-face, and it appears that their effective beat isdorsally directed. These structures could only bedetected in Stages 1 and 3. They are probablyabsent in Stage 5, definitely absent in Stage 7,and probably absent in Stage 8, suggesting thattheir duration may be variable or that they maydisappear during fixation.
The interpretation of these paired structures asthe openings of protonephridial ducts is specula-tive not only because of the obscure morphologyof the structures themselves, but also becauseErdmann (1935) was unable to find the preciseposition of these ducts even after repeated sec-tioning. Examination of Erdmann's figured sec-tions reveals that the protonephridia developearly in the veliger stage and by the pediveligerstage open to the exterior through a tiny pore inthe vicinity of the eye.
Gills
By means of scanning electron microscopy, thethree-dimensional form of the developing gill pri-mordia of the oyster can be described in detail.The most remarkable aspect of this developmentis the early formation of the bridge of tissue thatin mature oysters connects the posterior ends ofthe left and right gills. This bridge arises by cross-contact and possibly cross-fusion of cilia in Stage

NUMBER 328 21
7 followed by cross-fusion of epithelium in Stage8.
In Stage 1, the only element of the gill primor-dia which can be observed is a cluster of cilia atthe mid-posterior edge of each mantle lobe (Fig-ures 9, 10). It will be seen later that these cilia arethe precursors of the gill bridge. They stem fromthe first three rows of cells along the mantle edge,whereas other marginal cilia are generally re-stricted to the first row. Prepared specimens inthis stage and also in Stage 2 do not permitobservation of gill primordia on the epitheliumdeeper in the mantle cavity.
In Stage 3 the right gill primordium is a low,broad epithelial ridge about 20 jum wide tendingdirectly anteriorly along the dorsal side of the gillcavity from the mid-posterior edge of the mantle(Figures 47, 48). The ridge is low and barelydiscernible at the mantle edge but increases inheight, possibly to about 7 /im, as it approachesthe body wall. Cell boundaries are clearly dis-cernible. The cells are variable in size, the largestapproximately 3 /im in diameter; all have curved,microvillous apical surfaces. Crossing the anteriorthird of the ridge at a right angle are two minutetransverse ridges, the primordia of the gill fila-ments. Each of these ridges is only one cell wideand is bordered anteriorly and posteriorly by adepression of about the same width as the ridge.The primordium of the left gill in Stage 3 presentsbasically the same form (Figures 43, 44), but it isnot possible to determine whether it has the samenumber of transverse ridges. A few widely scat-tered cilia, no more than 3 fim in length, arepresent on each primordium. As in Stage 1, acluster of cilia is present on the mid-posteriormargin of each mantle lobe.
Little change occurs in the basic arrangementof the gill primordia until Stage 7. In specimensat this stage of development, only the posteriorextremities of the gill ridges could be observed,but these are instructive. A few cilia stemmingfrom the posterior extremity of each gill ridgecontact one another across the midline to producean exceedingly fine ciliary bridge (Figures 63-65).So tenuous is this connection that the electron
beam of the scanning electron microscope causedsome of the ciliary connections to break, a processthat could be observed on the cathode-ray view-ing screen. On each side the cilia arise from amound of epithelium at the end of the gill ridge(Figure 65). The mantle edge itself has by thistime extended distally from the posterior extrem-ity of the gill ridge and still bears a cluster of longcilia on its mid-posterior margin (see Figure 71,Stage 8). Apparently the clusters of cilia describedas occurring on the mantle margin in Stages 1through 3 have lost their marginal position asnew cells are added along the mantle edge. Newcilia continue to form at the same location on themargin, but the original cilia have now cross-fused, and cells proliferating around the base ofthe ciliary bridge are beginning to push upwardalong the bridge.
By Stage 8, elongate epithelial cells are presentacross the entire bridge on its anterior side, butthe bridge remains largely ciliary on its posteriorside (Figures 69, 70). The diameter of the bridgeat its narrowest point on the midline is scarcelymore than 2 /im, but it is surely stretched andattenuated when the valves are gaping. Cellspresent on the bridge are elongated along thebridge axis and appear to be growing over thefused cilia while new cilia form from the newcells. The bridge widens gradually toward eachside into the expanded posterior extremities of thegill ridges (Figures 71, 72).
Also by Stage 8, the gill ridges have undergoneconsiderable change and are beginning to showunequal development, the right ridge being re-tarded with respect to the left. Height of the ridgehas increased to about 20 jum, and in a sectiontransverse to the ridge axis, height now exceedswidth. The right primordial gill still has twotransverse ridges on its anterior half. Each trans-verse ridge is a cluster of apically convex cells ofvarious sizes. The anterior end of the main gillridge has become separated from the mantle ep-ithelium and is now the anterior end of the gillaxis (Figures 71, 72). The anterior insertion pointof this axis is slightly dorsal and lateral to thestatic-duct opening and slightly posterior and

22 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
dorsal to the primordial eye. Scattered cilia arepresent along the crest of the gill. The left gillridge at this stage is about the same size as theright, but possesses an additional transverse ridgeand depression.
In Stage 9, the gill bridge has increased itsminimum diameter on the midline to nearly 4/im, and the epithelial portion of the bridge looksstronger relative to the fused ciliary portion,which is still present (Figures 81, 82). The gillprimordia have basically the same configurationas in Stage 8, but the transverse ridges thatcomprise the primordial filaments are even moreprominent, and the anterior gill axis has increasedin length. The number of primordial filaments isthe same as in Stage 8, two on the right gill ridgeand three on the left. Close examination of thedepression between the two primordial filamentson the right gill ridge (Figures 83, 84) reveals thatit contains a mucus-gland opening (identified bya comparison with such structures in critical-point dried gill filaments of mature pectinidsshown by Owen and McCrae, 1976). It is likelythat these gland openings are already present inthe interfilamental depressions in Stage 8.
It is doubtful that the gills are functional inany of these early stages, except as the increasedsurface area which they contribute aids in respi-ration. The basic incurrent-excurrent pattern ofthe adult, however, is probably already present.As suggested by the direction of beat inferredfrom micrographs of the marginal cilia of thestatic-duct opening and the cilia adjacent to thepresumed protonephridial duct openings, waterflows dorsally past the primordial gills into thepresumptive suprabranchial region.
Anal Region
Dorsal to the foot and gills, the mantle cavitybecomes shallower and its walls converge on theshell margin at the posterior end of the hinge. Inthis anal region, three structures are presentthroughout the veliger stage: the anus, a postanalciliary tuft, and a distinct concavity, referred to
here as the posterodorsal notch, in the margin ofthe left valve.
The anus lies to the left of the midline. Inspecimens in Stages 1 and 3, where it is in aconstricted stage, it is a tiny oval structure ap-proximately 3.5 by 6 /im with the short axistransverse (Figures 17, 33, 41) and with short ciliaprojecting from its lumen. In Stage 8, one speci-men shows the anus in a dilated condition (Fig-ures 67, 69). In this state the anus is nearlycircular (26 by 29 /im) and is filled with fecalmaterial that obscures the cilia of the lumen.Specimens fixed in the act of defecating (Figure75) show that waste material is compacted intofeces within the rectum and is not rolled into fecesin the mantle cavity as suggested by Yonge(1926). Compaction is probably by way of ciliaryaction wihin the rectum at times when abundantfood is passing through. Peristalsis, said to bepresent by Yonge (1926), is unlikely, because nomuscles are known to occur in the walls of thelarval intestine or rectum (Erdmann, 1935).
The postanal tuft is raised and separated fromthe anus by a low step in the body wall (Figures12, 33, 40, 41, 43, 47, 69). Cilia in the tuftapproach 15 /im in length and are densely clus-tered ventrally, just dorsal to the anus; dorsally,the cilia are confined to a narrow medial band.The dense clustering of the cilia prevents deter-mining their direction of beat from an analysis oftheir curvature, except for a few cilia at theventral edge of the tuft. These appear to have adorsally directed effective beat. As mentioned inthe earlier description of behavior, living larvaepropel fecal material outward from the region ofthe postanal tuft along a trajectory which is tothe left of the plane of commissure of the valves.Presumably, some of the postanal cilia beat inthat direction.
The margin of the left valve is strongly de-flected adjacent to the dense postanal tuft andforms a distinct notch in the lip of the shellmargin and a slight outward deflection of theouter surface of the valve. In contrast, the marginof the right valve is deflected only very slightly ornot at all at this location (Figures 89-92, 94, 95,

NUMBER 328 23
137). In specimens that are fixed with the valvesnearly or completely closed (Figures 22, 51, 76),it can be seen that the cilia of the postanal tuftproject between the valves adjacent to the notch.The short cilia that lie in a medial band dorsal tothe tuft align with the plane of commissure of thevalves and barely project. When the valves arecompletely closed, an opening at the site of theshell notch may or may not be present. In allcases where an opening is present, it is a thin slitnot exceeding 7 /xm in maximum dimension.Removal of the periostracum by bleaching orplasma etching enlarges the opening (Figures 88,89), indicating that the periostracum of the op-posing valves at the site of the notch contributesto its closing.
Growth and extension of the shell marginspreserve the locus of positions of the notch toproduce a sharply delimited raised growth trackon the exterior of the left valve and in some casesa very low, barely discernible ridge on the rightvalve (Figures 88, 90, 91). The beginning of thegrowth track indicates that the notch first formsat the prodissoconch-I/II boundary; the end ofthe track demonstrates that the notch disappearsat metamorphosis, at the prodissoconch-disso-conch boundary (Figure 120). At its earliest pointon the left-valve surface, the growth track iscommonly narrower or unevenly constricted com-pared to its appearance slightly later (Figures 22,91, 95). The width of the growth track increasesvery gradually from about 10 jum to about 16 /im,suggesting that the size of the postanal ciliary tuftalso increases during development. This gradualincrease in width of the track, however, is notcompletely uniform. At each growth line thegrowth track constricts, and between each growthline it widens.
Although waste material is ejected from themantle cavity by the postanal cilia adjacent tothe notch, the notch itself provides too small anopening for this ejection to occur when the valvesare closed. Many specimens fixed with the valvesclosed have trapped waste material between thevalves in the notch, and feces are far too large topass by the notch even when the valves are agape
(Figure 75). Furthermore, the notch is a far morecommon site for injury to the margin of the shellthan elsewhere (Figures 12, 51, 76), suggestingthat foreign particles lodged between closedvalves at the notch produce the injury.
An explanation of the function of the notch,how it forms, why it is restricted largely to theleft valve, and why it is present only during theprodissoconch-II stage requires an examination ofthe nature of the mantle edge, the configurationof the shell margin, and the mode of formation ofthe prodissoconch-I/II boundary. This explana-tion will therefore be deferred until the finaldiscussion.
Mantle Edge
The epithelium forming the stalk of the velumand the body wall merges without change in itssurface appearance into the mantle epitheliumlining the inner surfaces of the valves. Microvilliare present everywhere on the surface; cilia areabsent except on the mantle margin or on struc-tures, such as the gill primordia, which form fromthe mantle.
The configuration of the mantle edge of veligerStages 1 through 9 is similar to that examined indetail by Cranfield (1974) in the pediveliger stage.Two marginal folds are present, separated by theperiostracal groove (Figures 36, 61, 71, 72).Widely scattered cilia on the mantle margin stemfrom what appears to be the first row of cells ofthe inner fold proximal to the periostracal groove(Figures 10, 35, 36, 38). These cells also bearelongate microvilli which appear to be swollen byand covered with secretion. The width of themicrovillous band is about 3 fim. At the distaledge of this band, which forms the sharp leadingedge of the inner mantle fold, microvilli as longas 1.5 jum lie against the surface of the perios-tracum and appear to be partially embedded init (Figure 36). On the basis of Cranfield's (1974)description, the marginal cilia appear to representthe cilia that he shows as occurring at the apex ofthe inner fold. A prominent band of cilia that hedescribed as running along the margin of the

24 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
inner fold and stemming from the third row ofcells inward from the margin does not appear tobe present in the stages examined in the presentstudy. A third row of cilia, which Cranfieldshowed stemming from the outer surface of theinner fold and extending toward the newly gen-erated periostracum, also could not be detectedor perhaps was not distinguished from the distalband.
Radial muscles inserted in the shell and ex-tending into the inner mantle fold were alsoshown to be present in the veliger by Cranfield(1974). I infer that they are already present atleast by Stage 5, because at this stage the mantleedge behaved differently than in earlier stagesduring preparation of the specimens. In Stages 1through 4, the mantle margin remained attachedto the periostracum (Figures 10, 35). Where frac-turing of tissue due to dessication occurred, it wasproximal to the periostracal groove and followedno particular pattern. In Stages 5 through 9,however, the entire mantle margin pulled awayfrom the periostracum, severing the periostracumnear where it emerged from the periostracalgroove so that a narrow band of periostracumremained in the groove (Figures 60, 61, 72, 79).The mantle edge then lifted up from the innersurface of the shell along a line of separation thatis probably the line of attachment of the radialmuscles (the pallial line). In Figures 72 and 79,radially aligned stringers of tissue that are prob-ably muscles can be seen meeting the shell alongthis line.
This separation of the mantle edge from theperiostracum and the inner shell surface permit-ted inspection of parts of the outer fold of themantle which otherwise would have been shieldedfrom view by the periostracum and shell. It canbe seen in Figures 60-62 that the distal edge ofthe outer fold attenuates to a thin edge andconsists of what appear to be long, densely mattedmicrovilli or cell processes. The edge is consis-tently scalloped around the entire periphery ofthe shell, the width of the projections being 12 to15 /im (Figures 60, 61). It is likely that the reen-trants in this scalloped margin are adjacent to
cell boundaries, which are not visible in the mi-crographs. The salients would then representmaximal growth of microvilli from centers ofcells. The outer surface of the outer fold, from itsdistal edge to the inferred pallial line, is entirelycovered by very long microvilli, many of whichremain embedded in the shell and appear to beintimately connected with its secretion (Figures72, 79, 83). The outer surface of the mantleproximal to the pallial line was not examined.
Shell
SHAPE AND SCULPTURE.—The few fully formedlarval shells (prodissoconch II) of Ostrea edulis(Figure 118-120) observed in the present studyare variable in size, ranging from 265 to 360 /imin length and 250 to 300 jam in height (measure-ments from the prodissoconchs of three specimensthat had cemented to the bottom of a laboratorybeaker). The valves are unequal in convexity, theleft being more convex than the right and theconvexity of closed valves being on the order of250 jum. In contrast, prodissoconch I is less vari-able in size, typically measuring 170 /an long and160 /im high with the valves equal in convexity(measurements from 17 veligers). Although noneof the specimens examined permit accurate mea-surement of convexity, it appears that this dimen-sion of prodissoconch I is on the order of 90 to100/tm.
The two prodissoconch stages can generally bedistinguished on the basis of three features: (1)prodissoconch I is separated from prodissoconchII by the first prominent commarginal growthline; (2) the surface of prodissoconch I is nearlysmooth or very faintly radially and commargin-ally striate, whereas that of prodissoconch II isdistinctly commarginally striate but lacks radialstriae; and (3) the posterodorsal notch and itsgrowth track begin at the prodissoconch-I/IIboundary. A fourth, less consistent criterion is achange in the angle of spiral coiling at the bound-ary between the two stages, generally from ahigher angle to a lower angle. In lateral view,prodissoconch I is D-shaped, but so also are the

NUMBER 328 25
11FIGURES 9-12.—Ostrea edulis, specimen 372A, early prodissoconch II, stage 1: 9, left posteriorside (X 350, bar = 20 Jim); 10, detail of Figure 9, mantle edge (M), ciliary tuft on mantle (C),and periostracum (P) (X 2500, bar = 5 /xm); 11, posterior view (X 370, bar = 20 /am); 12, detailof Figure 11, posterodorsal shell notch (N) with injury (I) and postanal ciliary tuft (AT) (X1750, bar = 5 pm).

26 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
FIGURES 13-17.—Ostrea edulis, specimen 372A, early prodissoconch II, stage 1: 13, gill cavity(GC) on right side of foot primordium (F) (X 2500, bar = 5 pm); 14, heel primordium (H) (X2400, bar = 5 fim); 15, detail of Figure 14, double byssus-duct opening (BD) (X 5800, bar = 2fim); 16, gill cavity on left side of foot primordium with oblique view of static-duct opening(SD), side views of cilia of possible prontonephridial-duct opening (PD), and eye primordium(E) (X 8200, bar == 1 /wn); 17, anus (arrow) and postanal ciliary tuft (AT) separated by fracture(FR) in the body wall (X 2000, bar = 5 fim).

NUMBER 328 27
FIGURES 18-21.—Ostrea edulis, specimen 372A, early prodissoconch II, stage 1: 18, left anteriorside (X 340, bar = 20 jam); 19, apical view of slightly retracted velum (X 520, bar = 20 /im); 20,detail of Figure 11, mouth (arrow) and postoral ciliary tuft (OT) (X 1600, bar = 5 fim); 21,detail of Figure 19, apical pit and surrounding tumescent epithelial cells (X 3000, bar = 5 fim).

28 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
26FIGURES 22-27.—Ostrea edulis, early prodissoconch II, between stages 1 and 2: 22, specimen369B, posterodorsal notch (N) and postanal ciliary tuft (AT) (X 3000, bar = 5 /un); 23,specimen 369C, left side (X 300, bar = 50 /un); 24, detail of Figure 23, disturbed periostracumon prodissoconch I (arrow) and repaired injury (I) on prodissoconch II (X 1800, bar = 5 fun);25, specimen 369C, folded velum, mouth (arrow), and postoral ciliary tuft (OT), viewed fromposterior side (X 800, bar = 10 /un); 26, specimen 373D, anterior side of velar stalk (V) (X 670,bar = 20 /un); 27, specimen 372B, tumescent cells on side of folded velum (X 2300, bar = 5fun).

NUMBER 328 29
FIGURES 28-32.—Ostrea edulis: 28, specimen 373D, early prodissoconch II, between stages 1 and2, detail of Figure 26, compound cilia of post oral band (X 5500, bar = 2 /im); 29 specimen373D, anterior (X 470, bar = 20 jam); 30, specimen 373D, apical pit of velum (X 2000, bar =5 /im); 31, specimen 372C, early prodissoconch II, between stages 2 and 3, slightly folded velumviewed obliquely from left side, inner preoral band (arrow) (X 410, bar = 20 /im); 32, detail ofFigure 31, compound cilia of outer preoral band (X 7000, bar = 2 /im).

30 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
FIGURE 33.—Oslrea edulis, specimen 373B, mid-prodissoconch II, stage 3, posterior view (X 620,bar = 20 /un).

NUMBER 328 31
FIGURES 34-38.—Ostrea edulis, specimen 373B, mid-prodissoconch II, stage 3: 34, slightly foldedvelum (X 300, bar = 50 /im); 35, posterior edge of mantle (M) viewed obliquely from over edgeof shell (X 2000, bar = 5 jum); 36, detail of Figure 35, periostracum (P) at contact with secretion-laden periostracal groove (G) (X 7500, bar = 2 Jim); 37, detail of Figure 34, apical pit of velum(X 3500, bar = 2 /im); 38, detail of Figure 36, periostracal groove (G) (X 15000, bar = 1 jum).

32 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
FIGURES 39-42.—Ostrea edulis, specimen 373B, mid-prodissoconch II, stage 3: 39, posterior view(X 330, bar = 50 /tm); 40 detail of Figure 39 (X 650, bar = 20 /zm); 41, detail of Figure 40,postanal ciliary tuft (AT) adjacent to posterodorsal notch (N), and anus (arrow) separated fromtuft by fracture (FR) in body wall (X 1700, bar = 5 fim); 42, detail of Figure 40, footprimordium with well-developed heel (H) and incipient toe (T) (X 1700, bar = 5 pm).

NUMBER 328 33
FIGURES 43-46.—Ostrea edulis, specimen 373B, mid-prodissoconch II, stage 3, stereomicrographs:43 and 44, left gill cavity (X 650, bar = 20 fun); 45 and 46, detail of Figures 43 and 44, openingof static duct with long fringing cilia (SD), primordial microvillous eye (E), and possibleprotonephridial-duct opening (arrow) (X 3000, bar = 5 jtim).
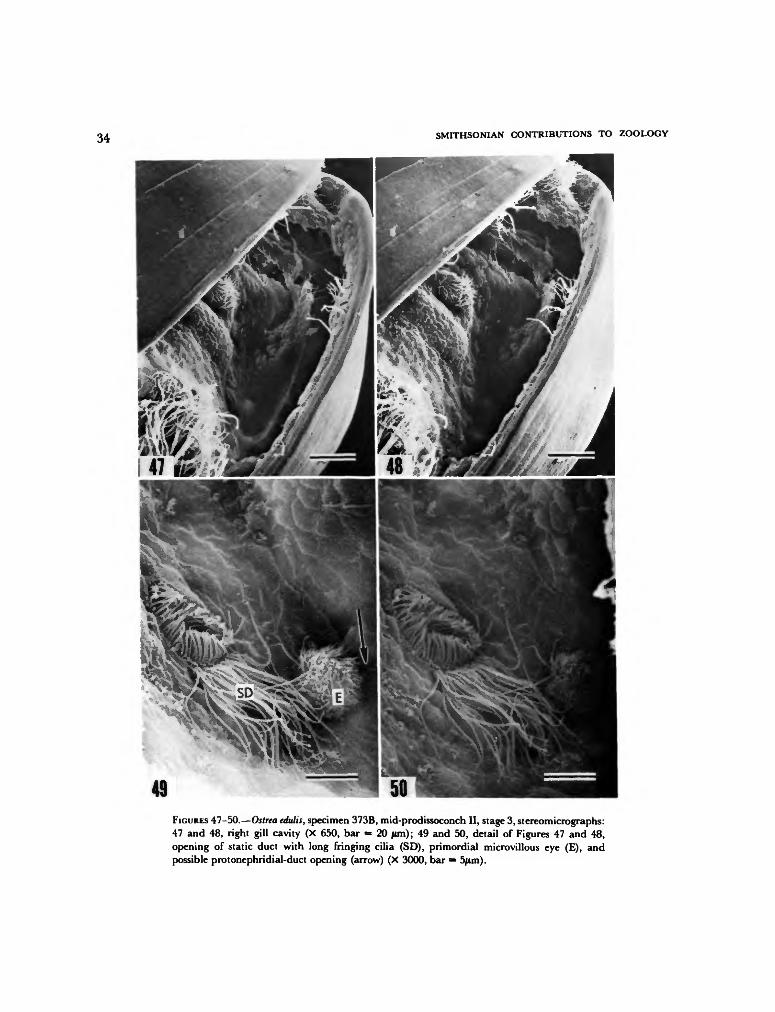
34 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
49FIGURES 47-50.—Ostrea edulis, specimen 373B, mid-prodissoconch II, stage 3, stereomicrographs:47 and 48, right gill cavity (X 650, bar = 20 jwn); 49 and 50, detail of Figures 47 and 48,opening of static duct with long fringing cilia (SD), primordial microvillous eye (E), andpossible protonephridial-duct opening (arrow) (X 3000, bar = 5/xm).

NUMBER 328 35
53FIGURES 51-54.—Ostrea edulis, specimen 375C, mid-prodissoconch II, stage 5: 51, prodissoconchI-II boundary (black arrows), notch and growth track (N) with repaired injury (white arrow)in left valve, and postanal ciliary tuft (AT) (X 780, bar = 10 /im); 52, foot (X 1500, bar = 10/im); 53, right gill cavity with opening of static duct (SD) and microvillous eye (E) (X 2470, bar= 5 /im); 54, detail of Figure 53, opening of static duct (X 8000, bar = 1 /im).

36 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
58FIGURES 55-59.—Ostrea edulis, specimen 376B, late prodissoconch II, stage 7: 55, ventral view,velum (V) folded and partially retracted (X 280, bar = 50 jum); 56, right gill cavity, opening ofstatic duct (SD) beside foot and microvillous eye (E) partially obscured by mantle edge (X2100, bar = 5 |im); 57, detail of opening of right static duct, viewed obliquely from ventral side(X 5400, bar = 2 /im); 58, detail of foot viewed obliquely, opening of cement-gland duct (arrow)(X 1150, bar = 10 /im); 59, detail of Figure 56, microvillous eye (X 4500, bar = 2 jam).

NUMBER 328 37
64FIGURES 60-65.—Ostrea edulis, specimen 376B, late prodissoconch II, stage 7: 60, posteroventralmantle edge (M) contracted and pulled away from periostracum (P) (X 1050, bar = 10 /im);61, detail of Figure 60, scalloped edge of outer fold of mantle (M) with periostracum (P)adhering to its inner surface (X 3600, bar = 2 ftm); 62, outer surface of outer fold of mantle inanterodorsal region (X 7900, bar = 1 /im); 63, posterior view of cross-fused mantle ciliacomprising primordial gill bridge (X 3100, bar = 2 |im); 64, detail of primordial gill bridge(X 3900, bar = 2 /im); 65, protrusion of epithelial cells (EP) at right end of primordial gillbridge (X 5000, bar = 2 Mm).

38 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
FIGURES 66-69.—Ostrea edulis, specimen 373A, late prodissoconch II, stage 8: 66, obliqueposterior view (X 275, bar = 50/mi); 67, posteroventral view, velum (V) retracted (X 270, bar= 50 fim); 68, detail of Figure 67, foot (F) and gill bridge (arrow) (X 1000, bar = 10 /im); 69,posterodorsal region viewed from right ventral side, dilated anus (A) containing excrement, gillbridge (arrow), and heel of foot (H) (X 1000, bar = 10 jtim).

NUMBER 328 39
73FIGURES 70-74.—Ostrea edulis, specimen 373A, late prodissoconch II, stage 8: 70, posterior viewof gill bridge consisting of cross-fused cilia (CC) and epithelial cells (EP) (X 7000, bar = 2 /am);71, left gill cavity viewed obliquely from posteroventral side, primordial gill (arrow) and gillbridge, mantle edge (M) with periostracal groove withdrawn from edge of shell (X 1100, bar= 10 /im); 72, right gill cavity (X 1270, bar = 10 fim); 73, oblique view of opening of static ducton left side of foot (X 5600, bar = 2 /xm); 74, opening of static duct (SD) beneath long fringingcilia on right side of foot (F) (X 3400, bar = 5 /un).

40 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
FIGURES 75-79.—Ostrea edulis, specimen 373C, late prodissoconch II, stage 9: 75, posterior view,velum retracted (X 270, bar = 50 /mi); 76, detail of Figure 75, posterodorsal notch (N) andgrowth track with injury, postanal ciliary tuft (X 750, bar = 20 /im); 77, detail of Figure 78,opening of cement-gland duct (CD) (X 3600, bar = 2 jtim); 78, foot, bordered on dorsal side bygill bridge partially obscured by feces (FE) (X 950, bar = 10 fim); 79, oblique view of outersurface of right mantle lobe withdrawn from shell, periostracal groove (G), and possible radialmuscle fibers (MF) (X 2000, bar = 5 jtim).

NUMBER 328 41
13FIGURES 80-84.—Ostrea edulis, specimen 373C, late prodissoconch II, stage 9: 80, left gillprimordium (GP) viewed from posteroventral side (X 900, bar = 10 /im); 81, gill bridge viewedfrom posterior, edge of outer fold of mantle and edge of shell on each side (X 1650, bar = 5/im); 82, detail of Figure 81, left end of gll bridge with elongate epithelial cells and cilia (X5340, bar = 2 pm); 83, right gill primordium (GP) viewed from posteroventral side (X 1200,bar = 1 0 /tm); 84, detail of Figure 83, probable mucus-gland opening (MG) between primordiaof gill filaments (GF) (X 7500, bar = 2 fun).

42 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
early stages of prodissoconch II. Prodissoconch-IIgrowth along the margins of prodissoconch Igradually pushes the prodissoconch-I valves awayfrom the plane of commissure and causes them torotate on the hinge until they project dorsal tothe hinge line, finally eliminating the D shape(compare Figure 11 to Figure 75).
The exterior surface of the earliest formed partof prodissoconch I has a sculptural pattern likethat on the prodissoconch I of Crassostrea virginicadescribed by Carriker and Palmer (1979a) andcalled by them the "punctate-stellate pattern"(Figures 85, 90, 92-94, 96, 97). In detail thepattern consists of two intergrading zones on eachvalve, a small, central "pitted zone" surroundedby a much larger "stellate-radial zone."
The pitted zone is oval in shape, roughly 50 by30 pim, with its long diameter parallel to thedorsoventral axis of the shell and its dorsal edgeseparated from the hinge by a distance of about15 to 20 fim. The pits are very shallow and rangein diameter from less than 1 jum in the center ofthe zone to over 2 jLim at the periphery of thezone. The entire pitted area tends to be somewhatdepressed relative to the surrounding pro-dissoconch-I surface and generally tends to beplanar or only very slightly convex. In somespecimens, however, the surface of the pitted zoneis thrown into relatively coarse folds which appearto involve both the periostracum and the outersurface of the outer calcified layer (Figure 98).
The stellate-radial zone begins at the peripheryof the pitted zone with a series of 10 to 15 low,obscure, radially oriented ridges separated by V-shaped depressed areas which point into the pit-ted zone (Figures 85, 90, 96, 97). The ridgesrapidly subside in elevation distally and disap-pear, the width of the ridged band being no morethan about 25 /im and generally considerably less.The remaining surface of the stellate-radial zoneis nearly smooth except for numerous exceedinglyfine radial striae which diverge in V-shaped clus-ters. Fine commarginal growth lines (about twoor three per micron) are also present (Figures 91,94, 97), and which feature, radial or commar-ginal, shows in the micrograph depends on the
angle of incidence of the electron beam. Theearliest commarginal lines begin near the begin-ning of the stellate-radial zone, and the patternthat they trace on the shell surface suggests thatthe early calcified shell is dumbbell shaped andtransversely elongate (Figure 85). Both the finecommarginal lines and the divaricate radial linesappear to persist through the prodissoconch-IIstage but are obscured in the later stage by muchcoarser commarginal ridges and by the somewhatthicker periostracum (Figures 11, 23, 24, 39, 66,92).
SHELL MICROSTRUCTURE.—The prodissoconchis covered by a periostracum so thin that it isdifficult to examine and measure by scanningelectron microscopy. Fracture sections and viewsof broken edges of periostracum suggest that itsthickness is generally less than 0.1 /im in the earlystages of prodissoconch I and that it may thickenslightly during the prodissoconch-II stage.
The outer surface of the calcified portion of theprodissoconch was examined after gentle removalof the periostracum by molecular bombardment(low-temperature plasma ashing) in oxygen (Fig-ures 99-105). In the pitted zone of prodissoconchI, tiny grains or needles, which are assumed to bearagonite2, are clustered in a polygonal pattern.In planar views of the surface, the grains orneedles are on the order of 0.1 jum in diameter,and the polygons are from 1 to 3 /im in diameter.In the stellate-radial zone, the same type of gran-ules or needles are present but are arranged inradially directed divergent rows that correspondto and probably produce the divergent radialpattern previously described on the periostracalsurface. Approaching the prodissoconch-I/IIboundary, the radial rows of granules become
2 Stenzel (1964) determined by X-ray diffraction that thelarval shell of Crassostrea virginica is aragonitic in composition;Carriker and Palmer (1979a) determined by the same meth-ods that both the prodissoconch-I and prodissoconch-IIstages of C. virginica are aragonite. There remains a possibil-ity, however, that the earliest calcification adjacent to theperiostracum in the pitted zone may be different. Watabe(1956) reported the presence of dahllite in the prodissoconchI of a pteriacean, Pinctada martensii (Dunker).

NUMBER 328 43
50 Aim
FIGURE 85.—Diagram of early prodissoconch I oiOstrea edulisshortly after evagination of the shell gland and before foldingalong the presumptive hinge. Pitted zones represent the twoinitial centers of calcification beneath a single, dumbbell-shaped periostracal sheet.
closer until on prodissoconch II the entire surface,including the growth track of the posterodorsalnotch, consists of a thin, dense layer of closelypacked granules (Figures 103, 143). Immediatelybeneath this granular surface are declined gran-ular laths (Figures 106-108), described below.
Fractures through the prodissoconch of Ostreaedulis reveal a more complex structure than pre-viously reported in bivalve larvae and demon-strate a gradual change in shell microstructurefrom the earliest portion of prodissoconch I to theend of prodissoconch II. As shown in Figures 109through 116, three layers are present:
1. The outer layer, which underlies the perios-tracum, is generally about 0.7 /im in thickness inthe early prodissoconch I but thins toward theprodissoconch-I margin and appears to be absentover the greater portion of prodissoconch II. Thelayer consists of exceedingly fine, irregular prismsthat in the inner part of the layer vary from 0.05to 0.1 /im in diameter. Next to the periostracum,the prisms attenuate into the fine needles de-scribed above. In scanning electron micrographs,the prisms comprising the outer layer appear tobe columns of granules, as described by Carrikerand Palmer (1979a) in the prodissoconch ofCrassostrea virginica.
2. The inner layer, which forms the inner sur-face of the larval shell except at the free marginsof the valves, is distinctly prismatic in structure.In scanning electron micrographs, the prisms ap-
pear to be uniform in texture rather than granularas in the outer layer. They are also coarser thanthe outer prisms, reaching diameters of about 0.3/im. In fully developed prodissoconchs, the innerlayer is thickest in the umbonal region, beneaththe prodissoconch-I portion of the shell, where itmay exceed 3 /im in thickness. The layer thinsdistally to less than 1 /im beneath the pro-dissoconch-II portion of the shell and finally tozero near the shell margin. The exact line ofdisappearance has not been determined, but itappears to be close to the line of attachment ofmantle to shell, about 15 to 20 /im from the shellmargin.
3. Beneath the pitted zone of prodissoconch Ia third, middle layer is little more than a discon-tinuity between the inner and outer prismaticlayers. This discontinuity thickens beneath theearly portion of the stellate-radial zone and be-comes a layer of randomly arrayed granules, afabric that is generally referred to as "homoge-neous." The increase in thickness is particularlynoticeable in the dorsal region (Figure 111),where this layer comprises the hinge teeth andhinge plate. In the vicinity of the prodissoconch-I/II boundary, the middle layer increases greatlyin thickness and, with the disappearance of theouter, prismatic layer, becomes the outer layeritself. At the shell margin, distal to the line ofattenuation of the inner prismatic layer, the ho-mogeneous layer is the only one present. In pro-dissoconch II the homogeneous layer also changesin character in that granules are no longer ran-domly arrayed but rather are clustered into lathsthat decline toward the shell margin from theouter surface to the inner surface (Figures 106-108, 115, 116). On the growth surface of thislayer, which is the inner surface of the shell alongthe free margins, the granular laths are alignedin commarginal rows (Figure 117; see also Figure148 of Crassostrea gigas).
These sculptural and textural changes duringgrowth of the prodissoconch suggest a pattern ofdifferentiation of epithelial cells during the tran-sition from shell gland to mantle (Figure 86). Aspreviously described, the first commarginal

44 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
FIGURE 86.—Hypothetical relationship between layers ofdistinctive microstructure in the larval shell of Ostrea edulisand cellular differentiation during the transition from shellgland to mantle: a, shell-gland phase of calcification at thebeginning of the prodissoconch-I stage; b, beginning of tran-sition from shell gland to mantle, early to middle pro-dissoconch I; c, early phase of calcification by the mantle,middle to late prodissoconch I; and d, calcification by man-tle, prodissoconch II. P and H represent cells secretingperiostracum and homogeneous shell structure respectively.Cells bearing a dot (•) and an "x" represent successivesecretory phases of cells originally present in the shell-glandphase.
growth line to form on prodissoconch I circum-scribes a transversely elongate, somewhat dumb-bell-shaped figure as shown in Figure 85. Thepitted zones in the expanded lateral lobes of thisinitial shell probably represent two initial centersof calcification on the inner surface of a single,dumbbell-shaped periostracal sheet. Calcificationprobably begins simultaneously throughout thearea of each pitted zone, as evidenced by thepolygonal distribution pattern of crystals de-scribed above. It also seems likely that thesecenters of calcification as well as the periostracalsheet occupy a transverse depression with en-larged ends on the dorsal surface of the trochop-hore. The folded relief on some pitted zones sug-gests that distortion may occur during evagina-tion of the shell gland, whereas the uniformlycurved surface of the stellate-radial zone suggests
that calcification in this region occurs after eva-gination.
Secretion of the stellate-radial zone of pro-dissoconch I is incremental, as evidenced bygrowth lines, and proceeds outward from thepitted zones in all directions including toward thehinge, as evidenced by the presence of radialelements between the pitted zones and the hinge(Figures 96, 137). Initial folding of the embryonicshell can occur only through the last remnant ofuncalcified periostracum, which becomes the lig-ament at the site of the hinge (described below).
As previously described, the homogeneous shellultrastructure that makes up nearly the entirethickness of prodissoconch II can be traced backto an origin in the discontinuity between theouter and inner prismatic layers present beneaththe umbones. The beginning of the middle layerpossibly indicates differentiation of a new corn-marginal band of cells near the periphery of theshell gland and a simultaneous change in secre-tory function of the older, more centrally locatedcells (Figure 86b). This is the kind of change thatCather (1967) used to mark the change from shellgland to mantle in a gastropod larva, and it seemsdesirable to use the same system in describing theontogeny of bivalve larvae. Subsequent thicken-ing of the middle shell layer thus may representincrease in size of a band of special secretory cellswhich persists until metamorphosis. Because thisspecialized band, and hence the mantle, beginswell before the prodissoconch-I/II boundary, theboundary itself cannot be explained as a result ofthe change from shell gland to mantle.
At metamorphosis, the form, microstructure,and mineralogy of the shell suddenly change. Theuniformly coiled, gibbous form of the pro-dissoconch is replaced by the irregular, flattenedform of the dissoconch, which is now cementedto the substrate (Figures 118, 119). Both valves ofthe prodissoconch change abruptly from entirelyaragonitic (Stenzel, 1964; Carriker and Palmer,1979a) to nearly entirely calcitic (Taylor, Ken-nedy, and Hall, 1969), and the final, homogene-ous shell microstructure of the prodissoconchchanges to much coarser prisms and folia (Figures

NUMBER 328 45
121-131). The mature shell of Ostrea edulis iscomposed primarily of foliated calcite, with athin outer layer of simple prismatic calcite oneach valve. The only aragonite known to bepresent in the dissoconch is restricted to areas ofmuscle attachment (the myostraca) and to theligament area (Carriker and Palmer, 1979b).
The prismatic calcitic outer layer of each valveof the dissoconch is initially no more than about1 jum in thickness, consisting of tabular polygonalprisms bounded exteriorly by periostracum (Fig-ures 121-129, 131). Scanning electron micro-graphs of the outer ends of prisms from which theperiostracum has been removed show that initialgrowth of each prism is accretionary, beginningat a nucleation point that is offset from the centerof the outer face of the prism toward the proximalside of the shell (Figure 123). As calcificationproceeds concentrically around each nucleationpoint, the centers of calcification impinge uponone another and crowding produces the polygo-nal form. Once the close-packed polygonal con-dition has been reached, the prisms can only growperpendicularly or at high angle to the periostra-cal surface. The diameters of prisms, measured ata distance of 1.5 jum from the edge of the pro-dissoconch, range from about 5 to 15 /an, with asomewhat larger size on the right valve than onthe left.
Vertical growth of the calcitic prisms is termi-nated by the advancing front of an inner shelllayer a short distance inside of the distal marginof the dissoconch (Figures 128, 129). This innerlayer is foliated calcite (Taylor, Kennedy, andHall, 1969), consisting of pointed laths inclinedat a very low angle to the plane of the shell. Eachlath has a thickness ranging from less than 1 /imto about 2 jum, a width from 0.5 to 3 fim, and anunknown length. Transition between prismaticand foliated microstructure is apparent both infracture sections (Figures 127, 131) and on thegrowth surface of the shell (Figure 129).3 Not only
3 A similar transition has been reported in the Pectinaceaand has contributed to a hypothesis that the prismaticmicrostructure is the evolutionary precursor of foliated mi-crostructure (Waller, 1972).
does foliated calcite cover the inner surface of theprismatic calcitic outer layer; it also extends in-ward over the inner surface of the prodissoconchand eventually completely fills the prodissoconchinterior.
In addition to prismatic and foliated calcite, athird microstructure is also present in the earliestdissoconch. It is irregularly prismatic and appar-ently is associated with the attachment of theposterior adductor to the shell (Figures 126, 130).As would be expected by its limited area ofsecretion, the microstructure forms a thin, lat-erally restricted layer which forms on the innersurface of previously secreted foliated calcite andin turn is covered by later secreted foliated calcite.Such irregularly prismatic microstructures asso-ciated with muscle attachments were termed"myostraca" by Oberling (1964) and are knownto be aragonitic in composition (Taylor, Ken-nedy, and Hall, 1969). What is of particularinterest here is the very early appearance of themyostracum, actually beneath the rim of theprodissoconch but separated from this rim by athin layer of foliated calcite (Figure 130).
The ultrastructure of this earliest myostracumis nearly identical to that of the inner shell layerof the prodissoconch (Figures 109-112, 114, 115).Although continuity between the two microstruc-tures cannot be demonstrated in the scanningelectron micrographs taken for this study, it istempting to postulate that they are intercon-nected and that the mantle epithelium that givesrise to the inner layer of the prodissoconch mayhave features in common with the adhesive epi-thelium in areas of muscle attachment (Tompaand Watabe, 1976; Waller, 1980). If so, the com-bination of anatomical and biochemical featuresthat produce the structure may be deeply rootedin the genetic program governing ontogeny of theshell.
HINGE AND MARGIN.—Ranson (1939) showedthat the prodissoconch-I hinge of Ostrea edulis islinear and lacks teeth, and Werner (1939) dem-onstrated that this is also the case in other biv-alves. In the present study, all of the specimensexamined are beyond the prodissoconch-I stage,

46 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
but information on the early development of thehinge is provided by commarginal growth lines.The growth line that separates prodissoconch Ifrom prodissoconch II can be traced to the hingeline and shows that there is little increase inlength of hinge during the prodissoconch II stage.The prominent hinge teeth of prodissoconch IIdevelop at the ends of the prodissoconch-I hinge(Figures 94, 96, 97).
The only ligament present in prodissoconch Iis likely the same band of thickened periostracumthat can be seen in early prodissoconch II (Figure139). The valve margins of prodissoconch I areprobably simple and not interlocking, as indi-cated by the presence of a largely prismatic mi-crostructure during this stage. It is unlikely thatthe coarse, parallel prisms of this microstructurecould be secreted on the interlocking shell margin.
Changes in number, spacing, shape, and sizeof hinge teeth during the development of pro-dissoconch II have been described by Pascual(1972) and will not be repeated here except torestate two points: (1) The posterior hinge teethare larger than the anterior from the beginningof their development at the start of prodissoconchII. Early in prodissoconch-II development (at ashell length of about 230 /im in Pascual's sam-ples), the anterior teeth begin to be filled inrapidly by new secretion of shell (Figure 138).Finally, at the end of the prodissoconch-II stage,only the posterior teeth remain in most specimens.(2) The central portion of the hinge between thehinge teeth is about equally developed in eachvalve throughout the prodissoconch-II stage.When the valves are closed, the central part ofthe hinge of one valve meets that of the othervalve approximately along the plane of commis-sure (Figures 137, 138), although tiny denticula-tions or rugosities may be present (Figure 136).
The central region of the hinge is not the siteof a resilium or tiny internal ligament, such asthat illustrated by Stenzel (1971)4, nor is there
4 Figure J37 in Stenzel (1971), showing a tiny central,internal ligament in the prodissoconch of Ostrea edulis, is saidto be from Ranson (1939), but Ranson's original figureshowed no such ligament.
any indication of a fibrous inner layer, such asthat indicated by Trueman (1951). The onlyligament present during the prodissoconch-IIstage is external, consisting of a band of thickperiostracum (also noted by Calloway andTurner, 1979, and Lutz, 1979). Growth lines onthe prodissoconch-II surface converge at the endsof this band and indicate that ligament lengthincreases only slightly during the prodissoconch-II phase. Scanning electron microscopy of thehinge plates of young dissoconchs of Ostrea edulisshow that the resilium first appears at or nearmetamorphosis at a point just anterior to theanterior hinge teeth (Figure 140). Pascual (1972)reached the same conclusion for Ostrea stentina,but Carriker and Palmer (1979a) found that inCrassostrea virginica the resilium appears earlier inthe prodissoconch-II stage.
The anterior and posterior sides of the hingeteeth of prodissoconch II bear curved ridges (Fig-ure 139) that fit into corresponding curvedgrooves on the sides of the corresponding socketsof the opposite valve. The curvature of thesefeatures is invariably concave toward the hingeline, and in fact the center of curvature liesapproximately on the axis of rotation of thevalves. Similar features are found on the larvalhinge teeth of Crassostrea virginica (Carriker andPalmer, 1979a), C. gigas (herein, Figure 149),Mytilus edulis and Modiolus modiolus (Lutz andHidu, 1979), and Argopecten irradians (Waller, un-published data). As shown by Stanley (1978),such a ridge and groove system on the hinge teethof mature bivalves serves to reduce shear betweenpartially opened valves and produces a tighterinterlocking system when the valves are in theprocess of opening and closing. The same functionappears to be served in larval shells.
As described by Pascual (1972), the configura-tion of the hinge of species of Ostrea becomesmuch more variable in larval shells approachingmetamorphosis than in younger shells. The an-terior teeth may or may not disappear completely,and the rugosity produced by secondary denti-culations on the primary teeth and across thecentral portion of the hinge may become extreme.

NUMBER 328 47
10/Lrrn
FIGURE 87.—Diagrammatic cross section of interlockingvalve margins in Ostrea edulis.
In all of the prodissoconch-II stages of Ostreaedulis examined in the present study, the valvesfit together in tongue-in-groove fashion by wayof margins that differ between valves (Figures 87,117, 132-135). Although these features are some-what variable, they can be traced around theentire extent of the free margins of the shell exceptfor the area of the posterodorsal notch of the leftvalve (described above). In the notch, the thinleading edge is either absent or folded back (Fig-ures 22, 89). The tongue-in-groove arrangementprovides a tight seal and prevents shear betweenclosed valves.
SHELL MORPHOLOGY OF OTHER
OYSTER LARVAE
A number of features of the larval shell ofOstrea edulis are present in the shells of other oysterspecies of both the oviparous and larviparoustypes.
1. The punctate-stellate pattern present on theexterior surface of prodissoconch I of Ostrea edulisis also present in Crassostrea virginica (Carriker andPalmer, 1979a) and Crassostrea gigas (Waller, un-published data).
2. The larval shell microstructure of Crassostreagigas (Figures 145-147) is similar to that of Ostreaedulis. Although Carriker and Palmer (1979a) didnot discuss microstructural change during growthof the prodissoconch, their figures indicate thatshell microstructure in Crassostrea virginica is of the
same type and undergoes the same kind of changeduring growth.
3. The prodissoconch-II shell margins of Crass-ostrea gigas (Figures 143, 144) are like those ofOstrea edulis. Scanning electron micrographs inCarriker and Palmer (1979a) indicate that thesame interlocking arrangement also exists inCrassostrea virginica.
4. The posterodorsal notch and growth trackin the prodissoconch II of Ostrea edulis are presentin the left valves of many species of oyster larvaein this stage of development. In the Family Os-treidae, the presence of the notch has been doc-umented in Saccostrea echinata, Ostrea edulis, 0.circumpicta, 0. denselamellosa, 0. futamiensis, Crass-ostrea virginica, C. ariakensis, and C. gigas by Tanaka(1960), in C. virginica by Carriker and Palmer(1979a), in Saccostrea glomerata and Ostrea corrugataby P. Chanley (pers. comm., 1979), and in C.gigas by the present study (Figure 141-144). Fur-thermore, close inspection of light micrographspublished by Ranson (1939) and Pascual (1971,1972) reveals that a posterodorsal notch is prob-ably present also in Crassostrea angulata and Ostreastentina. In the Family Gryphaeidae, larval shellsof Neopycnodonte cochlear shown herein (Figures 151,152) also have a posterodorsal notch in pro-dissoconch II. According to P. Chanley (pers.comm., 1979), the notch is not present in Ostrealutaria and 0. chilensis.
5. Although patterns of hinge dentition differin detail between species, the basic configurationof massive anterior and posterior teeth separatedby a central space devoid of hinge teeth is similarin early stages of prodissoconch II in Ostrea stentina(Pascual, 1972), Crassostrea angulata (Pascual,1972; Dinamani, 1976), Saccostrea glomerata (Din-amani, 1973, 1976), C. gigas (Dinamani, 1976;herein, Figure 150), and C. virginica (Dinamani,1976; Carriker and Palmer, 1979a). In specimensexamined with scanning electron microscope,curved ridges on the sides of teeth and sockets arepresent (Dinamani, 1976; Carriker and Palmer,1979a; herein, Figure 149).
In contrast to these similarities between species,

48 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
important differences separate the larval shells ofthe oviparous and larviparous groups.
1. Although the sizes attained by the pro-dissoconch at metamorphosis may be similar be-tween these two groups, there are great differencesin the dimensions of the prodissoconch-I stageand hence in the ratio of size of prodissoconch Ito size of prodissoconch II. Prodissoconch I is verysmall in the oviparous group, generally on theorder of 70 fim in length, which ranges from 20%to 25% of the length of the fully formed pro-dissoconch II. In contrast, prodissoconch I lengthsin larviparous species, including Ostrea edulis and0. stentina (herein and Pascual, 1972), range from140 to 190 jum and therefore occupy a muchgreater proportion of the length of the total larvalshell (50% to 60%).
2. Because the onset of asymmetry in the con-vexity of the two valves appears to begin at theprodissoconch-I/II boundary, the great differ-ences in the proportions of these stages betweenoviparous and larviparous oysters lead to greatdifferences in shape. In the fully developed pro-dissoconch II of species ofCrassostrea, the left valvegreatly exceeds the right valve in convexity, andtherefore its umbo extends farther over the hingethan in species of Ostrea (Pascual, 1971, 1972;Carriker and Palmer, 1979a; herein, Figure 142).As a consequence of the relatively greater coilingof the left valve in species of Crassostrea, the trackof its posterodorsal notch is longer and moreobviously spiral than in species of Ostrea (Carrikerand Palmer, 1979a; herein, Figure 142).
3. In both Ostrea edulis and 0. stentina, the hingeteeth remain separated during prodissoconch-IIdevelopment, while the anterior teeth graduallydiminish in prominence. The central portion ofthe hinge between the teeth is equally developedin both valves. In contrast, in Crassostrea angulata(Pascual, 1971; Dinamani, 1976); C. gigas (Dina-mani, 1976), and C. virginica (Dinamani, 1976;Carriker and Palmer, 1979a) the anterior andposterior teeth develop at the expense of thecentral portion of the hinge, and it is the posteriorteeth that decrease in prominence. The centralportion of the hinge is unequally developed be-
tween valves. The central area is a projection onthe right valve and a depression on the left, sothat the area itself serves as a sort of massive toothand socket.
4. Although incremental growth is present inthe prodissoconch-I phase because of the enlarge-ment of the secretory epithelium of the shell-gland/mantle, there is a difference in the promi-nence of incremental lines between species. Theyare prominent on the prodissoconch-I surface ofOstrea edulis but nearly lacking in the same areaof Crassostrea gigas (Figure 141) and C. virginica(Carriker and Palmer, 1979a).
Discussion
The prodissoconch-I/II boundary in larvalshells of bivalves was shown by Ockelmann (1965)to be an important indicator of type of larvaldevelopment and capability of dispersal. How-ever, he offered no detailed explanation of howthe boundary forms other than that it representsa change from calcification by the shell gland tothat by the mantle edge, an explanation also putforth by many others.
Two lines of evidence suggest that a change inmode of calcification is not the cause of theprodissoconch-I/II boundary. First, if the shellgland is defined as the specialized glandular epi-thelium in the transverse dorsal invagination ofthe trochophore, then there is no correspondencein position between the boundary on the pro-dissoconch and this feature. As shown above, onlythe earliest stages of calcification of prodissoconchI, inside the first discernible growth line, aretransversely elongate with respect to the larvalbody. The only feature that seems to suggest bothevagination and a change in mode of calcificationis the boundary between the central pitted zoneon the umbo of prodissoconch I and the surround-ing stellate-radial zone. Secondly, although theearliest phases of calcification in the oyster larvahave never been described in detail, evidencefrom other groups of mollusks (see above reviewof early development) suggests that the changefrom shell gland to mantle is not accompanied by
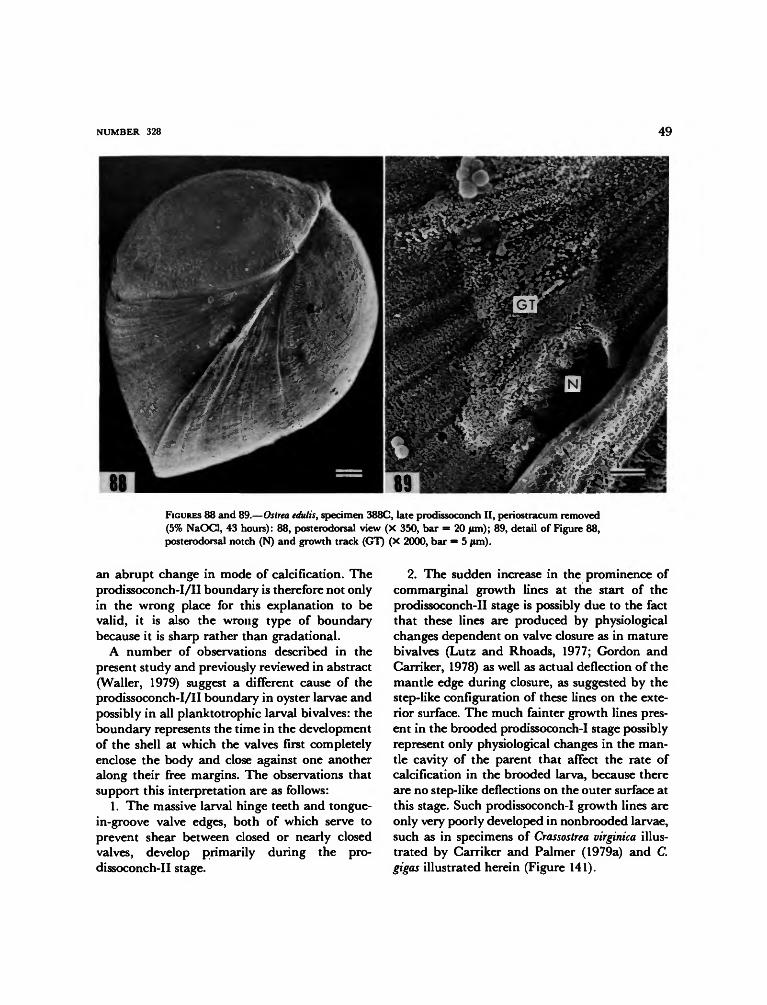
NUMBER 328 49
FIGURES 88 and 89.—Ostrea edulis, specimen 388C, late prodissoconch II, periostracum removed(5% NaOCl, 43 hours): 88, posterodorsal view (X 350, bar = 20 jum); 89, detail of Figure 88,posterodorsal notch (N) and growth track (GT) (X 2000, bar = 5 ftm).
an abrupt change in mode of calcification. Theprodissoconch-I/II boundary is therefore not onlyin the wrong place for this explanation to bevalid, it is also the wrong type of boundarybecause it is sharp rather than gradational.
A number of observations described in thepresent study and previously reviewed in abstract(Waller, 1979) suggest a different cause of theprodissoconch-I/II boundary in oyster larvae andpossibly in all planktotrophic larval bivalves: theboundary represents the time in the developmentof the shell at which the valves first completelyenclose the body and close against one anotheralong their free margins. The observations thatsupport this interpretation are as follows:
1. The massive larval hinge teeth and tongue-in-groove valve edges, both of which serve toprevent shear between closed or nearly closedvalves, develop primarily during the pro-dissoconch-II stage.
2. The sudden increase in the prominence ofcommarginal growth lines at the start of theprodissoconch-II stage is possibly due to the factthat these lines are produced by physiologicalchanges dependent on valve closure as in maturebivalves (Lutz and Rhoads, 1977; Gordon andCarriker, 1978) as well as actual deflection of themantle edge during closure, as suggested by thestep-like configuration of these lines on the exte-rior surface. The much fainter growth lines pres-ent in the brooded prodissoconch-I stage possiblyrepresent only physiological changes in the man-tle cavity of the parent that affect the rate ofcalcification in the brooded larva, because thereare no step-like deflections on the outer surface atthis stage. Such prodissoconch-I growth lines areonly very poorly developed in nonbrooded larvae,such as in specimens of Crassostrea virginica illus-trated by Carriker and Palmer (1979a) and C.gigas illustrated herein (Figure 141).

50 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
92FIGURES 90-93.—Ostrea edulis, critical-point dried: 90, specimen 368B, early prodissoconch II,dorsal view (X 450, bar = 20/im); 91, detail of Figure 90, posterodorsal notch and growth track(GT) and prodissoconch-I/II boundary (arrows) (X 1160, bar = 10 jum); 92, specimen 375B,middle prodissoconch II, left dorsal view (X 310, bar = 50 /im); 93, detail of Figure 92,prodissoconch-I region (X 820, bar = 10 fun).

NUMBER 328 51
FIGURES 94-99.—Ostrea edulis: 94, specimen 387G, air dried, middle prodissoconch II, dorsalview of left valve, prodissoconch-I/II boundary (arrow) (X 310, bar = 50 pan); 95, detail ofFigure 94, posterodorsal notch and growth track (X 1740, bar = 5 /im); 96, detail of Figure 94,pitted and stellate-radial zones (X 760, bar = 20 /un); 97, specimen 387E, early prodissoconchII, dorsal view of prodissoconch-I sculpture of right valve (X 740, bar = 20 /im); 98, specimen385B, early dissoconch, right valve, folds in pitted zone of prodissoconch, hinge toward bottom(X 4000, bar = 2 /im); 99, specimen 388A, middle prodissoconch II, dorsal view of outer surfaceof prodissoconch-I region of left valve, periostracum removed (5% NaOGI, 43 hours) (X 680,bar = 20 /im).

52 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
104FIGURES 100-105.—Ostrea edulis, periostracum removed (5% NaOCl, 43 hrs.): 100, specimen388A, mid-prodissoconch II, left valve, outer surface of pitted zone (PZ) and early stellate-radial zone (SR) (X 2060, bar = 5 /tm); 101, detail of Figure 100, pitted zone (X 7500, bar =2 (im); 102, detail of Figure 100, stellate radial zone, direction of growth toward upper left (X5000, bar = 2 /im); 103, specimen 388A, outer surface of prodissoconch-II stage (lower left)adjacent to prodissoconch-I/II boundary (arrow) (X 5000, bar = 2 nm); 104, specimen 388B,
prodissoconch 4t,—left-valve* ^mter - surface-of pitted zone (PZ) and early stellate-iadialzone (SR) (X 1700, bar = 5 fun); 105, specimen 388B, outer surface of middle stellate-radialzone, direction of growth toward lower right (X 2000, bar = 5 /im).

NUMBER 328 53
110FIGURES 106-111.—Ostrea edulis: 106, specimen 384, left valve, periostracum and outermost shelllayers partially removed by decay, prodissoconch-I/II boundary (arrow) (X 310, bar = 50 fim);107, detail of Figure 106, granular laths of prodissoconch II, direction of growth to lower left(X 1000, bar = 10 /im); 108, detail of Figure 107 (X 5000, bar = 2 /im); 109, specimen 376C,mid-prodissoconch II, right valve, critical-point dried, radial fracture through pitted zone (X20500, bar = 0.5 /im); 110, specimen 376C, radial fracture through early stellate-radial zone(X 15500, bar = 1 |im); 111, specimen 376C, fracture from early stellate-radial zone (lowerright) to late stellate-radial zone (X 3800, bar = 2 /un) (OP = outer prismatic layer, GH =granular homogeneous layer, IP = inner prismatic layer).

54 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
FIGURES 112-117.—Ostrea edulis: 112, specimen 387D, air dried, mid-prodissoconch II, rightvalve, commarginal fracture on distal side of prodissoconch-I/II boundary passing obliquelythrough vestige of outer prismatic layer (OP), granular homogeneous layer (GH), and innerprismatic layer (IP) (X 7600, bar = 2 fun); 113, detail of Figure 112 (X 41000, bar = 0.2 jum);114, specimen 387D, fracture through prodissoconch-II stage perpendicular to shell surfacesbut 45° to growth lines, outer granular homogeneous layer (GH), glue contamination (GL),inner prismatic layer (IP) (X 6800, bar = 2 tun); 115, specimen 387D, radial fracture throughearly prodissoconch-II stage, direction of growth to right (X 10500, bar = 1 /im); 116, specimen377B, critical-point dried, late prodissoconch II, radial fracture through ventral margin of rightvalve, outer side toward upper right (X 10500, bar = 1 /im); 117, specimen 387E, air dried,mid-prodissoconch II, inner surface of anterior margin of right valve, periostracum (P) presentnear outer edge and removed elsewhere (X 7360, bar = 2 |im).

NUMBER 328 55
FIGURES 118-123.—Ostrea edulis, specimen 383, air dried, early dissoconch: 118, left side (X 27,bar = 500 urn); 119, dorsal view (X 38, bar = 200 /im); 120, detail of Figure 119, prodissoconchvalves separated by dissoconch ligament area (L) (X 120, bar = 100 |im); 121, detail of Figure118, near ventral edge, prismatic calcitic outer shell layer and underlying foliated calcite,direction of growth to lower left (X 1000, bar = 10/im); 122, detail of Figure 121 (X 2760, bar= 5 /im); 123, outer surface of prismatic calcite of right valve showing accretionary lines onprism faces, direction of growth toward upper right (X 3900, bar =

56 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
FIGURES 124-129.—Ostrea edulis, early dissoconchs, air dried: 124, specimen 384, outer surfaceof right valve at contact (arrow) between prodissoconch and dissoconch (X 840, bar = 10/im);125, detail of Figure 124 (X 4200, bar = 2 /im); 126, specimen 385B, radial fracture throughearly dissoconch of left valve, outer prismatic calcite (PC) underlain by foliated calcite (FC)and lens of adductor myostracum (arrow) (X 5500, bar = 2 /im); 127, specimen 385B, leftvalve, radial fracture, distal to lower right, outer prismatic calcite (PC) transitional to innerfoliated calcite (FC) (X 4200, bar = 2 /xm); 128, specimen 385B, oblique view of inner surfaceof right valve of dissoconch hear ventral margin, prismatic calcitic outer layer (PC) and foliatedinner layer (FC), direction of growth to right (X 300, bar = 50 /im); 129, detail of Figure 128(X 1000, bar = 10/im).

NUMBER 328 57
FIGURES 130-135.—Ostrea edulis, air dried: 130, specimen 385A, left valve of early dissoconch,commarginal fracture through dissoconch along margin of prodissoconch, myostracum (M)separated by foliated calcite (FC) from prodissoconch (PR) (X 5000, bar = 2 fun); 131,specimen 385A, radial fracture through early dissoconch, left valve, outer prismatic calcite (PC)underlain by foliated calcite (FC) (X 5000, bar = 2 /mi); 132, specimen 387C, late prodissoconchII, inner side of right ventral margin, granular laths showing through thin periostracum (X6200, bar = 2 |im); 133, specimen 387F, middle prodissoconch II, inner side of left ventralmargin (X 6500, bar = 2 /im); 134, specimen 389L, middle prodissoconch II, radial fracturethrough right ventral margin, inner side at top (X 4450, bar = 2 urn); 135, specimen 387G,middle prodissoconch II, oblique view along ventral margin (X 7400, bar = 2 /im).
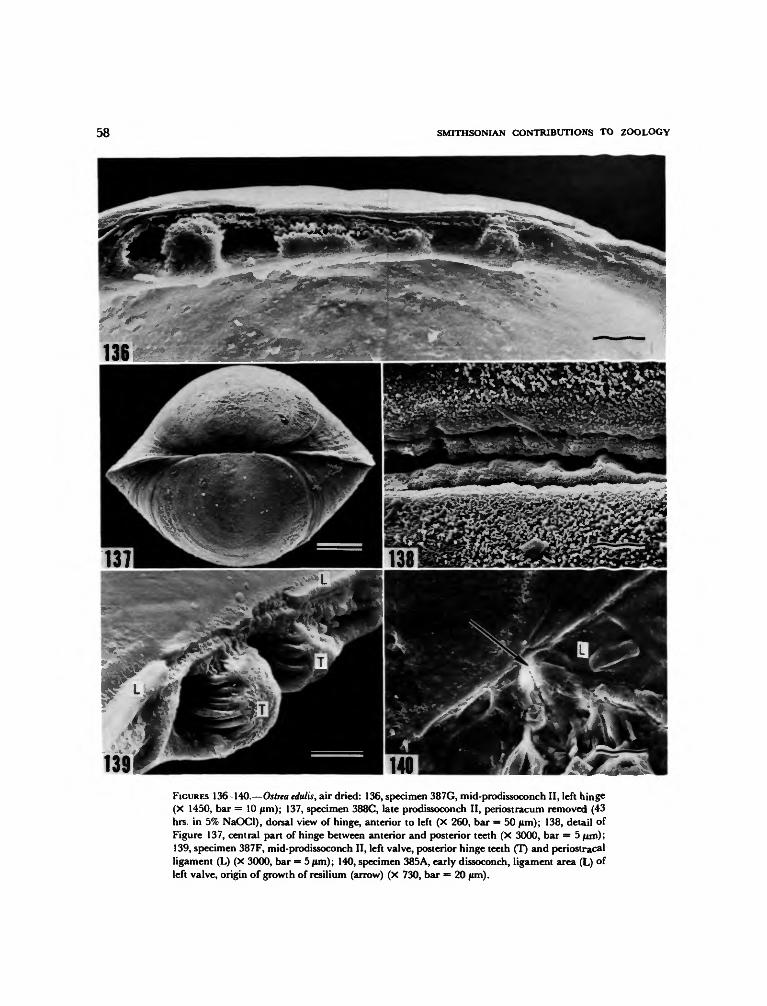
58 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
139FIGURES 136-140.—Ostrea edulis, air dried: 136, specimen 387G, mid-prodissoconch II, left hinge(X 1450, bar = 10 /tm); 137, specimen 388C, late prodissoconch II, periostracum removed (43hrs. in 5% NaOCl), dorsal view of hinge, anterior to left (X 260, bar = 50 fun); 138, detail ofFigure 137, central part of hinge between anterior and posterior teeth (X 3000, bar = 5/um);139, specimen 387F, mid-prodissoconch II, left valve, posterior hinge teeth (T) and periostracalligament (L) (X 3000, bar = 5 |im); 140, specimen 385A, early dissoconch, ligament area (L) ofleft valve, origin of growth of resilium (arrow) (X 730, bar = 20 /im).

NUMBER 328 59
143FIGURES 141-144.—Crassostrea gigas, late prodissoconch II: 141, specimen 362B, air dried, dorsalview, anterior toward lower left, prodissoconch-I/II boundary (arrows) (X 350, bar = 20 /«n);142, specimen 362A, air dried, posterior view, prodissoconch-I/II boundary (arrows) (X 325,bar = 20 fim); 143 and 144, specimen 367G, periostracum removed by plasma ashing,stereoscopic view of posterodorsal notch and interlocking valve margins (X 3400, bar = 2 jam).
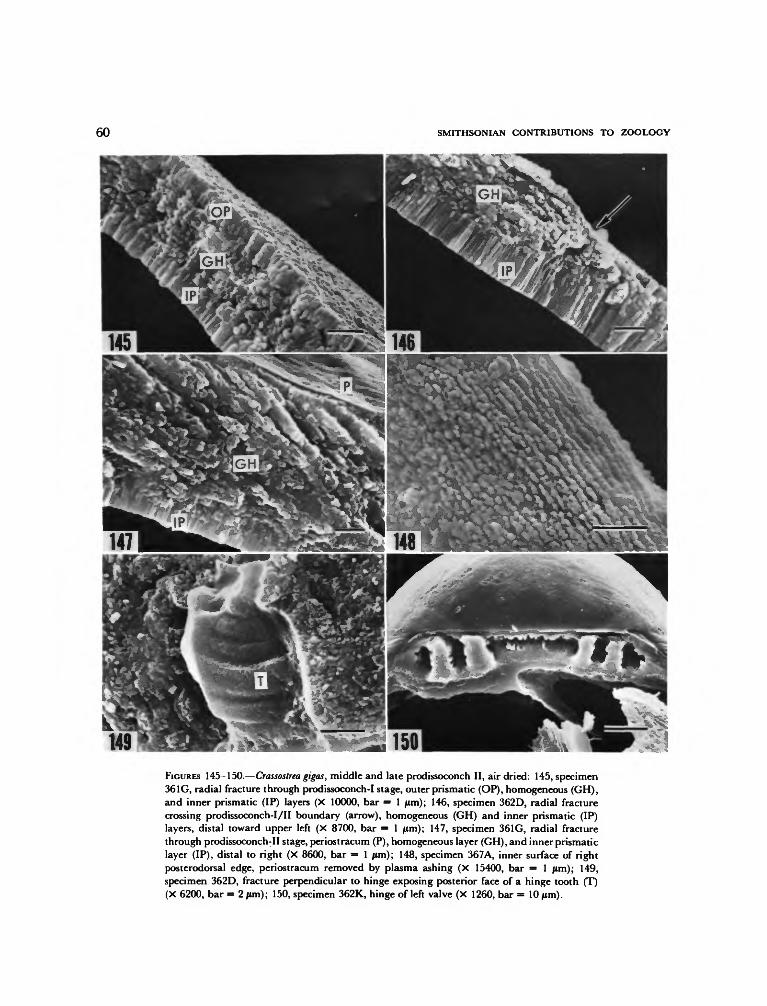
60 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
149FIGURES 145-150.—Crassostrea gigas, middle and late prodissoconch II, air dried: 145, specimen361G, radial fracture through prodissoconch-I stage, outer prismatic (OP), homogeneous (GH),and inner prismatic (IP) layers (X 10000, bar = 1 jam); 146, specimen 362D, radial fracturecrossing prodissoconch-I/II boundary (arrow), homogeneous (GH) and inner prismatic (IP)layers, distal toward upper left (X 8700, bar = 1 jtim); 147, specimen 361G, radial fracturethrough prodissoconch-II stage, periostracum (P), homogeneous layer (GH), and inner prismaticlayer (IP), distal to right (X 8600, bar = 1 jum); 148, specimen 367A, inner surface of rightposterodorsal edge, periostracum removed by plasma ashing (X 15400, bar = 1 fim); 149,specimen 362D, fracture perpendicular to hinge exposing posterior face of a hinge tooth (T)(X 6200, bar = 2 urn); 150, specimen 362K, hinge of left valve (X 1260, bar = 10/im).

NUMBER 328 61
FIGURES 151-152.—Neopycnodonte cochlear, prodissoconchs preserved on beaks of mature shells:151, USNM 196422, left valve (X 200, bar = 50 /un); 152, USNM 196422, left valve of anotherspecimen, posterodorsal view of growth track (GT) of posterodorsal notch extending betweenprodissoconch-I/II boundary and prodissoconch-dissoconch boundary (arrows) (X 530, bar =20/um).
3. Injuries affecting both valve margins at thesame point, which indicate that the valves closedon a foreign object or were injured simultaneouslyon both valve edges after being closed, are rare inthe prodissoconch-I stage but common in theprodissoconch-II stage.
4. The prodissoconch-I/II boundary in Ostreaedulis appears to coincide approximately withtime of release from parent, as indicated by spec-imens examined herein and by the material ex-amined by Erdmann (1935). It is at this monentthat complete valve closure would first be neces-sary for protection and survival.
The concept that the prodissoconch-I/IIboundary represents the onset of valve closurefacilitiates understanding the functional signifi-cance of some of the differences between ovipa-rous and larviparous larvae. As pointed out byOckelmann (1965), the size of prodissoconch I iscorrelated with size of egg. Oviparous species,which have a relatively small prodissoconch I,
produce vast quantities of small eggs; larviparousspecies, with a relatively large prodissoconch I,produce relatively small quantities of much largereggs. The prodissoconch I of larviparous bivalvesis larger simply because an embryo of greatervolume must be surrounded before valve closureis possible.
Although cell sizes may differ between theoviparous and larviparous groups and size itselfis not an adequate indicator of rate of growth,evidence suggests that completion of the pro-dissoconch-I stage of larviparous individuals maytake considerably longer than the correspondingstage of oviparous individuals [e.g., compare theseven-day brooding period of Ostrea edulis to theless-than-24-hour prodissoconch I of Argopectenirradians, an oviparous scallop (Waller, 1976; seealso Amemiya, 1926)]. It is conceivable that inoviparous species small egg size evolved not onlyto balance high mortality with high numbers ofpotential progeny, but also to reduce the time

62 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
needed to achieve protection from a closed shellto a minimum by reducing the volume to becovered by the initial closure.
A striking feature of the larval oyster shell isthe posterodorsal notch in the left-valve marginand the spiral track that this notch leaves on theexterior surface of the shell during its migrationwith shell growth. Observations reported abovedemonstrate that the notch forms exactly adja-cent to the postanal ciliary tuft and also that themantle edge and shell microstructure in the areaof the notch are no different from those on eitherside of it. It thus appears that the beating of thepostanal cilia actually deflects the thin periostra-cal envelope of the margin at this point, and thatthe deflection is made permanent by subsequentcalcification.
Although the postanal tuft is medial in posi-tion, it is not likely that the cilia beat morestrongly against one valve than the other. Why,then, is the notch much more prominent on theleft valve than on the right? The answer seems tolie in the fact that the cross-sectional configura-tions of the valve margins differ, a thin sharpedge being present on the left side, a thickrounded edge on the right. Deflection of the left-valve margin probably requires less mechanicalforce than does deflection of the right.
As stated in the review of early development,the postanal tuft is a continuation of the telotrochpresent during the trochophore stage and thus ispresent throughout the prodissoconch-I phase oflarval shell development. The sudden appearanceof the notch at or near the prodissoconch-I/IIboundary can readily be explained if the abovecause of this boundary—that it represents theonset of valve closure—is valid. It is at this timethat the valve margins would first be within reachof the beating postanal cilia. The notch disap-pears at the prodissoconch/dissoconch boundary,because the postanal tuft disappears at metamor-phosis and the newly formed rectum extends theanus ventrally, away from the inner, dorsal wallof the mantle cavity as in mature oysters.
It appears that the posterodorsal notch is aprominent feature throughout the Ostreidae. It is
unclear, however, why this feature is so stronglydeveloped in oysters but absent or only verypoorly developed in the plankototrophic larvalshells of other bivalve groups. Thus far, a com-parable but less well-developed feature has beenobserved only on a member of the Pholadacea(observable in Figure 2 of Calloway and Turner,1979). It is not known whether absence of aposterodorsal notch results from differences in theposition of the postanal tuft, in its degree ofciliation, or in the configurations of the shellmargins.
Another striking feature of oyster larvae is theappearance of the bridge between the posteriorextremities of the left and right gills well beforemetamorphosis and before the development ofthe gills themselves. As demonstrated above, thisbridge may arise by fusion of cilia present onopposite mantle lobes of the veliger. Cells thengrow medially from the mantle edge along theciliary bridge, and a bridge of tissue is completedat a time when the gill primordia are first becom-ing differentiated from the mantle epithelium.Superficially, this gill bridge has the appearanceof an adductor muscle and arises by the veryprocess by which adductor muscles are thoughtto have originated during the course of molluscanevolution (Yonge, 1957). Here, however, it isunlikely that any muscle fibers are present in thebridge, and the actual adductors arise during thecourse of larval development in the mesoderm bythe aggregation of free mesenchymal cells (Erd-mann, 1935).
Summary and Conclusions
Scanning electron microscopy of anesthetized,critical-point dried veliger larvae of Ostrea edulisL. revealed many anatomical features of bodyand shell not previously observed by other tech-niques. In summary, these features are as follows.
Velum
1. The apical pit contains a tuft of fine, shortcilia that barely project above the sides of the pitand are possibly sensory.

NUMBER 328 63
2. The apical pit is separated from the periph-ery of the velar crown by an expanse of non-ciliated, microvillous epithelium. The peripheryof the velum is girdled by four ciliary bands: aninner preoral band of fine cilia of unknown func-tion, an outer preoral band of long compoundcilia that function in locomotion and food gath-ering, an adoral band of fine cilia concerned withmovement of food particles to the mouth, and apostoral band of compound cilia that appear toassist the cilia of the outer preoral band in trap-ping food and to adjust locomotory forces.
3. The locomotory cilia of the outer preoralband beat in a plane that is oblique to the velarmargin. The obliquity and intensity of this beatmore than counter the opposing rotational forceimparted by laeoplectic metachronal waves, andthe larva thus rotates in a clockwise direction.
4. The postoral tuft is a distinct structure ofrelatively rigid cilia and is not merely the poste-rior edge of the postoral ciliary band. The tuft ispossibly sensory and may facilitate streaming ofexcess mucus and food posteriorly.
Foot
The heel and opening of the byssal-gland com-plex develop before the toe. The toe graduallybulges from the body wall and gradually becomesciliated along its midline and finally over itsentire surface. The opening of the cement-glandduct appears before the foot is fully formed.
Structures in Gill Cavities
1. Static-duct openings are continuously pres-ent throughout all stages studied and probablyremain open until metamorphosis. Analysis ofcurvature of fixed cilia suggest that cilia in thelumen of a static duct beat inward toward thestatocyst; long cilia fringing the ventral side ofeach duct opening beat dorsally, over the open-ing. These patterns support the idea that earlystatoliths and statoconia may originate exter-nally.
2. Cerebral eyes, which in Ostrea edulis become
pigmented late in the veliger stage, originate veryearly, probably before release from parent. Inearly veligers they are hemispherical tufts of mi-crovilli. Later, these tufts submerge in the epithe-lium, eventually becoming cup-shaped eyes.
3. A pitlike ciliated structure near each eye ispossibly an opening of a protonephridial duct.
Gill Primordia
The gill bridge, which in mature Ostrea edulisconnects the posterior ends of left and right gillsto one another, appears early in veliger develop-ment. It originates by cross-fusion of tufts of ciliaformed at the edge of opposite mantle lobes.Epithelial cells gradually migrate along thebridge and fuse at the midline, producing abridge that is at first both ciliary and epithelialand later completely epithelial. The mode offormation is precisely like that hypothesized forbivalve adductors, but here there are no musclecells involved, and the larval adductors originateduring development by an altogether differentprocess.
The gill primordia begin as anterior-posteriorridges of epithelium; filament primordia in theform of transverse ridges on the initial ridgedevelop later. Asymmetry begins in the mid-ve-liger stage, with the right gill primordium havingfewer primordial filaments than the left. Struc-tures interpreted as mucus glands are alreadypresent between the filament primordia. Al-though it is doubtful that the gills are functionalat this early stage, the basic incurrent-excurrentpattern through the mantle cavity seems to beestablished.
Anal Region and Posterodorsal Shell Notch
Feces are formed by compaction of waste inthe lumen of the gut and rectum and are movedby ciliary action through the anus, which is offsetto the left of the midline. Fecal material is force-fully ejected to the exterior in a trajectory thatcrosses the plane of commissure toward the rightside of the shell. At least part of the expulsive

64 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
force is generated by the postanal ciliary tuft,which lies dorsal to the anus along the postero-dorsal margin of the shell. The action of cilia ofthis tuft also deforms the shell margin, particu-larly the margin of the left valve, which is moredeformable than the margin of the right valve.The result is a distinct posterodorsal notch andgrowth track on the left valve. The notch origi-nates at the prodissoconch-I/II boundary, be-cause this boundary marks the onset of valveclosure and contact between valve margins andthe postanal ciliary tuft. The notch disappears atthe prodissoconch-dissoconch boundary, becausethe postanal ciliary tuft disappears at metamor-phosis.
Mantle Edge
The mantle edge is two-fold through all veligerstages. The leading edge of the inner fold haslong microvilli, some of which appear to beembedded in the periostracum. This fold also hasscattered cilia. The outer fold, which lies beneaththe newly generated periostracum, attenuates toa sharp edge consisting of long, densely mattedmicrovilli or cell processes. The outer surface ofthe outer fold is covered with long microvilli,many of which remain embedded in the shellafter the mantle is removed. Mantle contractionin dried specimens suggests that radial muscles inthe mantle are not present until the mid-veligerstage. These muscles appear to insert into theshell along a commarginal pallial line, as in ma-ture bivalves.
Shell
Exterior microsculptural patterns and micro-structure of the shell both reveal a progression ofchanges during the ontogeny of the prodissoconchthat can be explained in terms of a graduallyenlarging and differentiating shell gland andmantle. The prodissoconch begins at two centersof calcification on a single periostracal sheet afterthe invaginated stage of the shell gland. Thesecenters have a pitted appearance on the exterior;
gentle removal of the periostracum by plasmaashing reveals a polygonal pattern of tiny crystalsbeneath the periostracum. Thereafter, beneath astellate-radial external sculpture, calcification ismarginal and incremental, the increments possi-bly reflecting physiological irregularities in themantle cavity of the parent in which the larvadevelops.
Fracture sections show that the shell becomesthree-layered during the prodissoconch-I stage,with the middle layer beginning as little morethan a discontinuity between outer and innerprismatic layers. This middle layer increases inprominence in prodissoconch I and finally domi-nates the structure of the shell in prodissoconchII.
An hypothesis of cellular differentiation of themantle edge, advanced to explain the changingshell microstructures, leads to the conclusion thatthe change from shell gland to mantle is gradualand that it occurs well before the prodissoconch-I/II boundary.
The fine aragonitic microstructures of the pro-dissoconch change abruptly to relatively coarseprismatic and foliated calcitic microstructures inthe dissoconch. The foliated structure extendsinward and covers the inner surface of the pro-dissoconch. Myostracal layers appear in the ear-liest dissoconch and may be continuous with theinner prismatic layer of the larval shell. Thissuggests that attachment of mantle to larval shellmay have features in common with postlarvaladhesive mantle.
Hinge teeth develop primarily during the pro-dissoconch-II stage and have curved ridges ontheir sides that offset shear between partiallyclosed valves as in mature bivalves. The earlyligament is formed by the periostracum; the re-silium does not appear until near or possibly aftermetamorphosis. When the resilium appears, it isanterior to the anterior larval hinge teeth, not inthe center of the larval hinge as shown in someprevious studies. The margins of prodissoconch IIfit together in tongue-in-groove fashion to providea tight seal and to prevent shear between closedvalves.

NUMBER 328 65
On the basis of a limited comparison, many ofthe features of the larval shell of Ostrea edluis arealso present in the larval shells of other oysterspecies: pitted and stellate-radial sculptural pat-terns of prodissoconch I, configuration and ontog-eny of shell microstructures, interlocking valvemargins, posterodorsal notch and growth trackand its ontogeny, and basic pattern of hinge teethand ligament. Important differences, however,separate species in oviparous and larviparousgroups. Compared to the larval shells of larvipa-rous species, those of oviparous species have amuch smaller prodissoconch I compared to pro-dissoconch II. As a result of greater coiling duringthe prodissoconch-II stage, oviparous larval shellsalso have a more convex shape, with umbonesoverhanging the hinge line further. Fine incre-mental growth lines, present on prodissoconch Iof 0. edulis, appear to be absent from pro-dissoconch I of oviparous species. Important dif-ferences between species also occur in the ontog-eny of hinge teeth.
The prodissoconch-I/II boundary has been ex-plained in the past as the result of a change inmode of calcification from that by shell gland tothat by the mantle. However, the actual differ-entiation of the mantle edge appears to precedethe boundary, and the sharpness of the boundaryis not at all reflected in the transitional changefrom shell gland to mantle. Instead, evidencedetailed here for the first time suggests that theboundary marks the onset of valve closure.
Precisely how these observations should enterinto an interpretation of the evolution of oystersis unclear at present, because as yet there are nocomparable details on larvae of other bivalves. Itis likely, however, that as study of other groupsboth by scanning and transmission electron mi-croscopy progresses, a comparative anatomy willeventually develop that will allow, for the firsttime, assessment of the role of larvae in the evo-lutionary diversification of the Bivalvia.

Literature Cited
Allen, J. A.1961. The Development of Pandora inaequivalvis (Linne).
Journal of Embryology and Experimental Morphology,9(2):252-268, 5 figures, 2 tables.
Amemiya, I.1926. Notes on Experiments on the Early Developmental
Stages of the Portuguese, American and EnglishNative Oysters, with Special Reference to theEffect of Varying Salinity. Journal of the MarineBiological Association of the United Kingdom, 14(1):161-175, 1 figure, 6 tables.
Anderson, T. F.1951. Techniques for the Preservation of Three-dimen-
sional Structures in Preparing Specimens for theElectron Microscope. Transactions of the New YorkAcademy of Science, series 2, 13(3): 130-133.
1956. Electron Microscopy of Micro-organisms. In G.Oster and A. Pollistor, editors, Physical Techniquesin Biological Research, volume 3 (Cells and Tissues),chapter 5, pages 177-240, 22 figures, 1 table. NewYork: Academic Press.
Ansell, A. D.1962. The Functional Morphology of the Larva, and the
Post-larval Development of Venus striatula (DaCosta). Journal of the Marine Biological Association ofthe United Kingdom, 42(2):419-443, 13 figures.
Barber, V. C.1968. The Structure of Mollusc Statocysts, with Partic-
ular Reference to Cephalopods. Symposia of theZoological Society of London, number 23:37-62, 15figures.
Becker, R. P., and O. Johari1978. Scanning Electron Microscopy—1978. Volume 2, 1134
pages. AMF O'Hare, Illinois: Scanning ElectronMicroscopy, Inc. [Post Office Box 66507].
Bernard, F.1896. Deuxieme Note sur le developpement et la mor-
phologie de la coquille chez les Lamellibranches(Taxodontes). Bulletin de la Societe Geologique deFrance, series 3, 24:54-82, 15 figures.
Boyde, A.1978. Pros and Cons of Critical Point Drying and Freeze
Drying for SEM. In R. P. Becker and O. Johari,editors, Scanning Electron Microscopy—1978, 2:303-314. AMF O'Hare, Illinois: Scanning ElectronMicroscopy, Inc. [Post Office Box 66507].
Boyle, P. J., and R. D. Turner1976. The Larval Development of the Wood Boring
Piddock Martesia striata (L.) (Mollusca: Bivalvia:Pholadidae)./ounwi/ of Experimental Marine Biologyand Ecology, 22:55-68, 4 figures, 2 tables.
Brach, J.1690. Observ. CCIII, De ovis Ostreorum. Ephemeridum
Academia Leopoldina Naturae Curiosorum, VIII: 506-508.
Brooks, W. K.1880. The Development of the Oyster (Ostrea virginiana
[sic] List). Johns Hopkins University, Studies from theBiological Laboratory, 4:1-83, 10 plates.
Buddenbrock, W. von1915. Die Statocyste von Pecten, ihre Histologie und
Physiologic Zoologisches Jahrbuch Abteilung fur all-gemeine Zoologie und Physiologie, 35:301-356, 15 fig-ures, plates 7-8.
Calloway, C. B., and R. D. Turner1979. New Techniques for Preparing Shells of Bivalve
Larvae for Examination with the Scanning Elec-tron Microscope. Bulletin of the American Malacolog-ical Union for 1978, pages 17-24, 3 plates.
Carazzi, D.1902. Contributo all'istologia e alia fisiologia dei Lamel-
libranchi. Internationale Monatsschriftfur Anatomie undPhysiologie, 20:57-87, 5 figures, plates 3 and 4.
Carriker, M. R.1961. Interrelation of Functional Morphology, Behavior,
and Autecology in Early Stages of the BivalveMercenaria mercenaria. Journal of the Elisha MitchellScientific Society, 77(2): 168-241, 39 figures, 13 ta-bles.
Carriker, M. R., and R. E. Palmer1979a. Ultrastructural Morphogenesis of Prodissoconch
and Early Dissoconch Valves of the Oyster Crass-ostrea virginica. Proceedings of the National ShellfisheriesAssociation, 69:103-128, 63 figures.
1979b. A New Mineralized Layer in the Hinge of theOyster. Science, 206:691-693, 2 figures, 1 table.
Cather, J. N.1967. Cellular Interactions in the Development of the
Shell Gland of the Gastropod, llyanassa. The Journalof Experimental Zoology, 166(2):205-223, 24 figures.
Cole, H. A.1938. The Fate of the Larval Organs in the Metamor-
phosis ofOstrea edulis. Journal of the Marine Biological
66

NUMBER 328 67
Association of the United Kingdom, 22:469-484, 8
figures.Cragg, S. M., and J. A. Nott
1977. The Ultrastructure of the Statocysts in the Pedi-veliger Larvae of Pecten maximus (L.) (Bivalvia).
Journal of Experimental Marine Biology and Ecology,
27(l):23-36, 14 figures, 1 table.Cranfield, H. J.
1973a. A Study of the Morphology, Ultrastructure, andHistochemistry of the Foot of the Pediveliger ofOstrea edulis. Marine Biology, 22(3): 187-202, 16 fig-
ures, 3 tables.1973b. Observations on the Behaviour of the Pediveliger
of Ostrea edulis during Attachment and Cementing.Marine Biology, 22(3):203-2O9, 2 figures, 2 tables.
1973c. Observations on the Function of the Glands of theFoot of the Pediveliger of Ostrea edulis duringSettlement. Marine Biology, 22(3):211-223, 13 fig-ures, 4 tables.
1974. Observations on the Morphology of the MantleFolds of the Pediveliger of Ostrea edulis L. andTheir Function during Settlement. Journal of theMarine Biological Association of the United Kingdom,
54:1-12, 4 figures, 2 plates, 1 table.1975. The Ultrastructure and Histochemistry of the Lar-
val Cement of Ostrea edulis L. Journal of the Marine
Biological Association of the United Kingdom, 55 :497-
503, 1 figure, 3 plates, 1 table.Culliney, J. L., P. J. Boyle, and R. D. Turner
1972. New Approaches and Techniques for StudyingBivalve Larvae. In W. L. Smith and M. H. Chan-ley, editors, Culture of Marine Invertebrate Animals,
pages 257-271, 2 figures. New York: Plenum Pub-lishing Corp.
Dantan, J.-L.1916. La Larve de VOstrea edulis (L.). Annales de L'Institut
Oceanographique (Foundation Albert Ier, Prince de Mon-
aco). 7(6): 20 pages, 3 figures, 2 plates.Davaine, C.
1853. Recherches sur la generation des huitres. ComptesRendus des Se'ances et Memoires de la Socie'te de Biologie,
Pans, series 1, 4(1852):297-339, 2 plates.Demian, E. S., and F. Yousif
1973a. Embryonic Development and Organogenesis inthe Snail Marisa cornuanetis (Mesogastropoda: Am-pullariidae), I: General Outlines of Development.Malacologia, 12(1): 123-150, 24 figures, 1 table.
1973b. Embryonic Development and Organogenesis inthe Snail Marisa cornuanetis (Mesogastropoda: Am-pullariidae), IV: Development of the Shell Gland,Mantle and Respiratory Organs. Malacologia,12(2): 195-211, 13 figures.
Dinamani, P.1973. Embryonic and Larval Development in the New
Zealand Rock Oyster, Crassostreaglomerata (Gould).The Veliger, 15(4): 295-299, 19 figures, 1 table.
1976. The Morphology of the Larval Shell of Saccostreaglomerata (Gould, 1850) and a Comparative Studyof the Larval Shell in the Genus Crassostrea Sacco,1897 (Ostreidae). Journal of Molluscan Studies, 42:
95-107, 7 figures.Eakin, R. M.
1965. Evolution of Photoreceptors. In Cold Spring HarborSymposia on Quantitative Biology, 30:363-370, 2 fig-
ures. Cold Spring Harbor, Long Island, N. Y.:Cold Spring Harbor Laboratory of QuantitativeBiology.
Erdmann, W.1935. Untersuchungen iiber die Lebensgeschichte der
Auster, 5: Uber die Entwicklung und die Anato-mie der "Ansatzreifen" Larve von Ostrea edulis mitBemerkungen iiber die Lebensgeschichte der Aus-ter. Wissenschaftliche Meeresuntersuchungen Kommission
zur Wissenschaftlichen Untersuchung der Deutschen
Meere in Kiel und der Bwlogischen Anstalt auf Helgoland,
new series, 19(6): 25 pages, 5 figures, 8 plates.Fernando, W.
1931. The Origin and Development of the Pericardiumand Kidneys in Ostrea. Proceedings of the Royal Society
of London, series B, 107(752):391-397, plates 29-32.
Fujita, T.1929. On the Early Development of the Common Jap-
anese Oyster. Japanese Journal ofZoology, 2(3) :353-
358, plate 7.1934. Note on the Japanese Oyster Larva. In Proceedings
of the Fifth Pacific Sciences Congress, Canada 1933, 5:
4111-4117, 5 figures, 1 table. Toronto: Universityof Toronto Press.
Galtsoff, P. S.1964. T h e American Oyster. United States Department of
the Interior, Fish and Wildlife Service, Bureau of Com-
mercial Fisheries, Fishery Bulletin, 164: iii + 480
pages, 400 figures, 46 tables.Gordon, J., and M. R. Carriker
1978. Growth Lines in a Bivalve Mollusk: SubdailyPatterns and Dissolution of the Shell. Science, 202:519-521,4 figures.
Gray, J.1928. Ciliary Movement, viii -I- 162 pages, 105 figures, 16
tables. Cambridge: Cambridge University Press.1930. The Mechanism of Ciliary Movement, VI: Pho-
tographic and Stroboscopic Analysis of CiliaryMovement . Proceedings of the Royal Society of London,
series B, 107(751):313-332, 10 figures, plates 20-22.

68 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
Hatschek, B.1880. Ueber Entwicklungsgeschichte von Teredo. Arbeiten
aus dem Zoologiscken Institute der Universitat Wien und
der Zoologischen Station in Tnest, 3(1): 1-44, 3 plates.
Hickman, R. W., and LI. D. Gruffydd1971. The Histology of the Larva of Ostrea edulis during
Metamorphosis . In Fourth European Marine Biology
Symposium, Bangor, Wales, 1969, pages 281-294, 4
figures. Cambridge: Cambridge University Press.Hori,J.
1933. On the Development of the Olympia Oyster, Ostrealunda Carpenter, Transplanted from United Statesto J a p a n . Bulletin of the Japanese Society of Scientific
Fisheries, l(6):269-276, 17 figures.Horst, R.
1883-1884. The Development of the Oyster (Ostrea edulisL). [Translation from Dutch by Herman Jacob-son.] United States Commission of Fish and Fisheries,
Report of the Commissioner for 1884, part 12 [1886]:
891-910, 2 plates.Knight-Jones, E. W.
1954a. Notes on Invertebrate Larvae Observed at Naplesduring May and June. Pubblicaziom della StazwneZoologica di Napoli, 25(1): 135-144, 2 figures, 1 ta-ble.
1954b. Relations between Metachronism and the Direc-tion of Ciliary Beat in Metazoa. Quarterly Journal ofMicroscopical Science, 95:503-521.
Kniprath, E.1977. Zur Ontogenese des Schalenfeldes von Lymnaea
stagnahs. Wilhelm Roux's Archives of Developmental
Biology, 181:11-30, 7 figures.1979. The Functional Morphology of the Embryonic
Shell-Gland in the Conchiferous Molluscs. Mala-cologia, 18:549-552, 2 figures.
Korringa, P.1941. Experiments and Observations on Swarming, Pe-
lagic Life and Setting in the European Flat Oyster,Ostrea edulis L. [Thesis, University of Amsterdam,1940.] Archives Se'erlandaises de Zoologier, 5: 249
pages, 24 figures.Kume, M., and K. Dan
1968. Invertebrate Embryology. [Translation from Japaneseby J. C. Dan.] xvi + 605 pages, illustrated. Bel-grade: NOLIT Publishing House. [Published forthe National Library of Medicine, Public HealthService. United States Department of Health, Ed-ucation, and Welfare, and the National ScienceFoundation, Washington, D. C. Originally pub-lished by Bai Fu Kan Press, Tokyo, 1957.]
Lacaze-Duthiers, M. H.1854. Nouvelles Observations sur le developpment des
hint res. Comptes Rendus Hebdomadaires des Seances de
I'Academte des Sciences, 39:1197-1200.
Leeuwenhoek, A. van1695. Arcana Naturae detecta. Volume II, letters 92 and 94.
Lutz, R. A.1979. The Bivalve "Larval Ligament Pit" as an Exclu-
sively Post-larval Feature (Abstract). Proceedings ofthe National Shellfisheries Association, 69:197.
Lutz, R. A., and H. Hidu1979. Hinge Morphogenesis in the Shells of Larval and
Early Post-Larval Mussels (Mytilus edulis L. andModiolus modwlus (L.). Journal of the Marine Biological
Association of the United Kingdom, 59:111 121, 1
figure, 1 plate, 1 table.Lutz, R. A., and D. C. Rhoads
1977. Anaerobiosis and a Theory of Growth Line For-mation. Science, 198:1222-1227, 2 figures.
Meisenheimer, J.1900. Entwicklungsgeschichte von Dreissensia polymorpha
Pall. Zeilschnft fiir ivissenschaftliche Zoologie, 6 9 : 1 -
137, 18 figures, 13 plates.Millar, R. H.
1955. Notes on the Mechanism of Food Movement inthe Gut of the Larval Oyster, Ostrea edulis. Quarterly
Journal of Microscopical Science, 96(4): 539 544, 3 fig-
ures.Morton, J. E.
1958. Mollusca. First edition, 232 pages, 23 figures. Lon-don: Hutchinson and Company.
Oberling, J. J.1964. Observations on Some Structural Features of the
Pelecypod Shell. Mitteilungen der NaturforschendenGesellschaft in Bern, new series, 20: 60 pages, 3figures, 6 plates.
Ockelmann, K. W.1965. Developmental Types in Marine Bivalves and
Their Distribution along the Atlantic Coast ofEurope. In L. R. Cox and J. F. Peake, editors,Proceedings of the First European Malacological Congress
{London, September 17-21, 1962), pages 25-35, 5
figures. London: Conchological Society of GreatBritain and Ireland and the Malacological Societyof London.
Orton,J. H.1937. Oyster Biology and Oyster-Culture, Being the Buckland
Lectures for 1935. 211 pages. London: Edward Ar-nold and Co.
Owen, G., and J. M. McCrae1976. Further Studies on the Latero-frontal Tracts of
Bivalves. Proceedings of the Royal Society of London,
series B, 194:527-544, 28 figures, plates 1-7.Pascual, E.
1971. Morfologia de la charnela larvaria de Crassostreaangulata (Lmk.) en diferentes fases de su desarrollo.

NUMBER 328 69
Investigaaon Pesquera, 35(2):549-563, 10 figures.1972. Estudio de la conchas larvarias de Ostrea stentina,
Payr. y Ostrea edulis L. Investigacion Pesquera, 36(2):297-310, 8 figures.
Plate, L.
1924. Gleichgewichtserhaltung und Schwerkraftorgane.In L. Plate, editor, Allgemeine Zoologie und Abstam-mungslehre, part 2, pages 92-131, figures 88-130.Jena: Gustav Fischer.
Ranson, G.1939. Le Provinculum de la prodissoconque de quelques
Ostreides. Bulletin du Museum d'Histoire Naturelle(Paris), series 2, 11:318-332, 4 figures, 1 plate.
1960. Les prodissoconques (coquilles larvaires) des Os-treides vivants. Bulletin de I'Institut Oceanographique(Foundation Albert Ier, Prince de Monaco), 1183:41 pages, 136 figures.
Raven, C. P.
1958. Morphogenesis: The Analysis of Molluscan Development.311 pages, 66 figures. New York: Pergamon Press.
Rees, C. B.1950. The Identification and Classification of Lamelli-
branch Larvae. Hull Bulletins of Marine Ecology,3(19): 73-104, 4 figures, 5 plates.
Rice, E. L.1908. Gill Development in Mytilus. Biological Bulletin,
14(2):61-77, 9 figures.Seno, H.
1929. A Contribution to the Knowledge of the Devel-opment of Ostrea denselamellosa Lischke. Journal ofthe Imperial Fisheries Institute, 24(5): 129-135, 3plates.
Sleigh, M. A.1962. The Biology of Cilia and Flagella. xiii + 223 pages, 53
figures, 18 plates, 14 tables. Oxford: PergamonPress.
1974. Cilia and Flagella. xi + 500 pages, illustrated. Lon-don and New York: Academic Press.
Stanley, S. M.1978. Aspects of the Adaptive Morphology and Evolu-
tion of the Trigoniidae. Philosophical Transactions ofthe Royal Society of London, series B, 284(1OO1):247-258, figures 29-32, 2 plates.
Stenzel, H. B.1964. Oyster: Composition of the Larval Shell. Science,
145(3628): 155 156, 2 figures.1971. Oysters. In R. C. Moore, editor, Treatise on Inverte-
brate Paleontology, part N, volume 3, Mollusca 6,Bivalvia, pages 953 1224. Lawrence, Kansas:Geological Society of America and University ofKansas Press.
Strathmann, R. R., T. L. John, and J. R. C. Fonseca1972. Suspension Feeding by Marine Invertebrate Lar-
vae: Clearance of Particles by Ciliated Bands of aRotifer, Pluteus, and Trochophore. Biological Bul-letin, 142:505-519, 5 figures.
Tanaka, Y.
1960. Identification of Larva of Saxostrea echinata (Quoyet Gaimard). Venus [Japanese Journal of Malacology),21(l):32-38, 7 figures, 2 plates.
Taylor, J. D., W. J. Kennedy, and A. Hall1969. The Shell Structure and Mineralogy of the Biv-
alvia, Introduction: Nuculacea-Trigonacea. Bul-letin of the British Museum {Natural History), Zoology,supplement 3: 125 pages, 77 figures, 29 plates.
Tompa, A. S., and N. Watabe1976. Ultrastructural Investigation of the Mechanisms
of Muscle Attachment to the Gastropod Shell.Journal of Morphology, 149(3) :339 351, 1 figure, 3plates.
Trueman, E. R.
1951. The Structure, Development, and Operation ofthe Hinge Ligament of Ostrea edulis. Quarterly Jour-nal of Microscopical Science, 92(2): 129-140, 8figures.
Turner, R. D., and P. J. Boyle1975. Studies of Bivalve Larvae Using the Scanning
Electron Microscope and Critical Point Drying.Bulletin of the American Malaculogical Union, Inc.,1974:59-65, 4 plates, 2 tables.
Waller, T. R.1972. The Functional Significance of Some Shell Micro-
structures in the Pectinacea (Mollusca: Bivalvia).In International Geological Congress, 24th Session, Mon-treal Canada, 7(Paleontology):48-56, 3 figures, 1table.
1976. The Development of the Larval and Early Post-larval Shell of the Bay Scallop, Argopeclen irradians(Abstract). Bulletin of the American Malacological Un-wn, Inc., 1976:46.
1979. Formation of a Posterodorsal Notch in LarvalOyster Shells and the Prodissoconch-I/II Bound-ary in the Bivalvia (Abstract). Bulletin of the Amer-ican Malacological Union, Inc., 1978:55-56.
1980. Scanning Electron Microscopy of Shell and Man-tle in the Order Arcoida (Mollusca: Bivalvia).Smithsonian Contributions to Zoology, 313: 58 pages,46 figures, 1 table.
Watabe, N.1956. Dahllite Identified as a Constituent of Pro-
dissoconch I of Pmctada martensn (Dunker). Science,124(3223) :630, 1 table.

70 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
Werner, B.1939. Uber die Entwicklung und Artunterscheidung von
Muschellarven des Nordseeplanktons, unter be-suderer Berucksichtigung der Schalen-entwick-lung. Zoologische Jahrbucher, 66(1): 1-54, 22 figures,3 tables.
Yonge, C. M.1926. Structure and Physiology of the Organs of Feeding
and Digestion in Ostrea edulis. Journal of the MarineBiological Association, United Kingdom, 14(2):295-386, 42 figures, 20 tables.
1957. Mantle Fusion in the Lamellibranchia. Pubblica-zioni del la Stazione Zoologica di Napoli, 29:151-171,11 figures.
1960. Oysters. 207 pages, 72 figures, 17 plates. London:Collins.



REQUIREMENTS FOR SMITHSONIAN SERIES PUBLICATION
Manuscripts intended for series publication receive substantive review within theiroriginating Smithsonian museums or offices and are submitted to the Smithsonian InstitutionPress with approval of the appropriate museum authority on Form SI-36. Requests forspecial treatment—use of color, foldouts, casebound covers, etc.—require, on the sameform, the added approval of designated committees or museum directors.
Review of manuscripts and art by the Press for requirements of series format and style,completeness and clarity of copy, and arrangement of all material, as outlined below, willgovern, within the judgment of the Press, acceptance or rejection of the manuscripts and art.
Copy must be typewritten, double-spaced, on one side of standard white bond paper,with i v i " margins, submitted as ribbon copy (not carbon or xerox), in loose sheets (notstapled or bound), and accompanied by original art. Minimum acceptable length is 30 pages.
Front matter (preceding the text) should include: title page with only title and authorand no other information, abstract page with author/title/series/etc., following the establish-ed format, table of contents with indents reflecting the heads and structure of the paper.
First page of text should carry the title and author at the top of the page and an unnum-bered footnote at the bottom consisting of author's name and professional mailing address.
Center heads of whatever level should be typed with initial caps of major words, withextra space above and below the head, but with no other preparation (such as all caps orunderline). Run-in paragraph heads should use period/dashes or colons as necessary.
Tabulations within text (lists of data, often in parallel columns) can be typed on the textpage where they occur, but they should not contain rules or formal, numbered table heads.
Formal tables (numbered, with table heads, boxheads, stubs, rules) should be sub-mitted as camera copy, but the author must contact the series section of the Press for edito-rial attention and preparation assistance before final typing of this matter.
Taxonomic keys in natural history papers should use the alined-couplet form in thezoology and paleobiology series and the multi-level indent form in the botany series. Ifcross-referencing is required between key and text, do not include page references within thekey, but number the keyed-out taxa with their corresponding heads in the text.
Synonymy in the zoology and paleobiology series must use the short form (taxon,author, yearpage), with a full reference at the end of the paper under "Literature Cited."For the botany series, the long form (taxon, author, abbreviated journal or book title, volume,page, year, with no reference in the "Literature Cited") is optional.
Footnotes, when few in number, whether annotative or bibliographic, should be typedat the bottom of the text page on which the reference occurs. Extensive notes must appear atthe end of the text in a notes section. If bibliographic footnotes are required, use the shortform (author/brief title/page) with the full reference in the bibliography.
Text-reference system (author/year/page within the text, with the full reference in a"Literature Cited" at the end of the text) must be used in place of bibliographic footnotes inall scientific series and is strongly recommended in the history and technology series:"(Jones, 1910:122)" or ".. . . Jones (1910:122)."
Bibliography, depending upon use, is termed "References," "Selected References," or"Literature Cited." Spell out book, journal, and article titles, using initial caps in all majorwords. For capitalization of titles in foreign languages, follow the national practice of eachlanguage. Underline (for italics) book and journal titles. Use the colon-parentheses systemfor volume/number/page citations: "10(2):5-9." For alinement and arrangement ofelements, follow the format of the series for which the manuscript is intended.
Legends for illustrations must not be attached to the art nor included within the text butmust be submitted at the end of the manuscript—with as many legends typed, double-spaced, to a page as convenient.
Illustrations must not be included within the manuscript but must be submitted sepa-rately as original art (not copies). All illustrations (photographs, line drawings, maps, etc.)can be intermixed throughout the printed text. They should be termed Figures and shouldbe numbered consecutively. If several "figures" are treated as components of a single largerfigure, they should be designated by lowercase italic letters (underlined in copy) on the illus-tration, in the legend, and in text references: "Figure 9JD." If illustrations are intended to beprinted separately on coated stock following the text, they should be termed Plates and anycomponents should be lettered as in figures: "Plate 9b_." Keys to any symbols within anillustration should appear on the art and not in the legend.
A few points of style: (1) Do not use periods after such abbreviations as "mm, ft,yds, USNM, NNE, AM, BC." (2) Use hyphens in spelled-out fractions: "two-thirds." (3)Spell out numbers "one" through "nine" in expository text, but use numerals in all othercases if possible. (4) Use the metric system of measurement, where possible, instead ofthe English system. (5) Use the decimal system, where possible, in place of fractions.(6) Use day/month/year sequence for dates: "9 April 1976." (7) For months in tabular list-ings or data sections, use three-letter abbreviations with no periods: "Jan, Mar, Jun," etc.
Arrange and paginate sequentially EVERY sheet of manuscript—including ALL frontmatter and ALL legends, etc., at the back of the text—in the following order: (1) title page,(2) abstract, (3) table of contents, (4) foreword and/or preface, (5) text, (6) appendixes,(7) notes, (8) glossary, (9) bibliography, (10) index, (11) legends.

i