fumarase deficiency causes protein and metabolite ... chemical biology article fumarase deficiency...
TRANSCRIPT

Article
Fumarase Deficiency Causes Protein andMetabolite
Succination and Intoxicates MycobacteriumtuberculosisGraphical Abstract
Highlights
d Fumarase is essential for growth of Mtb
d Fumarase deficiency inMtb is bactericidal in vitro and during
mouse infection
d Fumarate accumulation leads to metabolite and protein
succination
d Fumarase-deficient Mtb is hypersusceptible to oxidative
stress
Ruecker et al., 2017, Cell Chemical Biology 24, 1–10March 16, 2017 ª 2017 Elsevier Ltd.http://dx.doi.org/10.1016/j.chembiol.2017.01.005
Authors
Nadine Ruecker, Robert Jansen,
Carolina Trujillo, ..., Henrik Molina,
Kyu Y. Rhee, Sabine Ehrt
In Brief
Ruecker et al. report that Mycobacterium
tuberculosis is uniquely vulnerable to
fumarase deficiency. Fumarase depletion
perturbed essential metabolic pathways
and led to intracellular fumarate
accumulation. Fumarate caused
metabolite and protein succination, a
covalent chemical modification of
cysteine thiol residues. This affected two
ofMtb’s major antioxidants, catalase and
mycothiol, causing hypersusceptibility to
oxidative stress.

Please cite this article in press as: Ruecker et al., Fumarase Deficiency Causes Protein and Metabolite Succination and Intoxicates Mycobacteriumtuberculosis, Cell Chemical Biology (2017), http://dx.doi.org/10.1016/j.chembiol.2017.01.005
Cell Chemical Biology
Article
Fumarase Deficiency Causes Protein and MetaboliteSuccination and IntoxicatesMycobacterium tuberculosisNadine Ruecker,1 Robert Jansen,2 Carolina Trujillo,1 Susan Puckett,1,5 Pradeepa Jayachandran,1,6 Gerardo G. Piroli,3
Norma Frizzell,3 Henrik Molina,4 Kyu Y. Rhee,2 and Sabine Ehrt1,7,*1Department of Microbiology and Immunology2Department of MedicineWeill Cornell Medical College, New York, NY 10065, USA3Department of Pharmacology, Physiology & Neuroscience, School of Medicine, University of South Carolina, Columbia, SC 29209, USA4Proteomics Resource Center, Rockefeller University, New York, NY 10065, USA5Present address: Department of Medical Microbiology, University Medical Center Utrecht, 3584 CX Utrecht, the Netherlands6Present address: Department of Biological Sciences and RNA Institute, University at Albany, Albany, NY 12222, USA7Lead Contact
*Correspondence: [email protected]
http://dx.doi.org/10.1016/j.chembiol.2017.01.005
SUMMARY
Enzymes of central carbon metabolism are essen-tial mediators of Mycobacterium tuberculosis(Mtb) physiology and pathogenicity, but are oftenperceived to lack sufficient species selectivity tobe pursued as potential drug targets. Fumarase(Fum) is an enzyme of the canonical tricarboxylicacid cycle and is dispensable in many organisms.Transposon mutagenesis studies in Mtb, however,indicate that Fum is required for optimal growth.Here, we report the generation and characteriza-tion of a genetically engineered Mtb strain in whichFum expression is conditionally regulated. This re-vealed that Fum deficiency is bactericidal in vitroand during both the acute and chronic phases ofmouse infection. This essentiality is linked tomarked accumulations of fumarate resulting in pro-tein and metabolite succination, a covalent modifi-cation of cysteine thiol residues. These resultsidentify Mtb Fum as a potentially species-specificdrug target whose inactivation may kill Mtbthrough a covalently irreversible form of metabolictoxicity.
INTRODUCTION
Mycobacterium tuberculosis (Mtb) remains a global health threat
causing 1.5 million fatal infections annually (http://www.who.int/
tb/en). Tuberculosis (TB) treatment requires a regimen of four
drugs for a period of at least 6 months and demands
careful implementation and high patient compliance. Healthcare
mismanagement and poor patient adherence favor the occur-
rence of multidrug-resistant strains, leading to an even higher
demand for new chemotherapeutics with novel mechanisms of
action.
Cell
Mtb’s central carbon metabolism (CCM) provides not only
energy for growth, but also pathogenicity. Several studies
have highlighted the importance of CCM enzymes for Mtb’s
ability to grow and persist in mice (Marrero et al., 2010, 2013;
McKinney et al., 2000; Munoz-Elıas and McKinney, 2005;
Pandey and Sassetti, 2008; Puckett et al., 2014; Trujillo et al.,
2014; Venugopal et al., 2011). Knowledge about Mtb’s
metabolism, therefore, benefits the understanding of TB patho-
genesis and can identify potential new targets and chemothera-
peutic interventions.
Fumarase catalyzes the reversible hydration of fumarate to
malate, a reaction of the tricarboxylic acid (TCA) cycle. The
TCA cycle oxidizes acetyl coenzyme A to carbon dioxide and
generates reducing equivalents for ATP production. It also pro-
vides precursors for amino acid and nucleotide biosynthesis.
However, fumarate is not only generated in the TCA cycle, but
is also a product of argininosuccinate lyase (ArgH) in the urea cy-
cle and of adenylosuccinate lyase (PurB) in purine biosynthesis
(Figure 1A).
Two classes of fumarases can be differentiated (Woods et al.,
1988). Class I fumarases occur only in prokaryotes, are thermo-
labile, iron dependent, and form homodimers. Class II fumarases
can be found in both prokaryotes and in eukaryotic mitochon-
dria, they are thermostable, iron independent, and form homote-
tramers. Several prokaryotes possess multiple fumarases
including E. coli, which encodes fumarases of both classes.
Expression of each depends on growth rate, oxygen availability,
and carbon source (Park and Gunsalus, 1995; Tseng et al.,
2001). Mtb encodes a single class II-type fumarase gene, fum
(rv1098c). Fum shares 45% sequence identity with the human
enzyme, including a conserved active site serine residue (Cole
et al., 1998; Mechaly et al., 2012). Somewhat unexpectedly,
transposon mutagenesis studies predicted Fum to be essential
for optimal growth of Mtb in vitro (Griffin et al., 2011; Zhang
et al., 2012). This was surprising because Mtb encodes a func-
tional glyoxylate shunt that generates succinate and malate
and should, therefore, bypass the Fum reaction (Eoh and
Rhee, 2013) (Figure 1A). Mtb has also been reported to be
capable of co-catabolizing different carbon substrates, making
Chemical Biology 24, 1–10, March 16, 2017 ª 2017 Elsevier Ltd. 1

Figure 1. Fum Depletion Kills Mtb
(A) Schematic depicting the TCA cycle, the urea
cycle, and reactions leading to fumarate produc-
tion. ArgH, argininosuccinate lyase; PurB, ad-
enylosuccinate lyase.
(B and C) Growth (B) and survival (C) of WT and
Fum-DUC in 7H9 complete medium with (+) and
without (�) atc. Colony-forming units (CFU) were
determined by culturing serial dilutions on agar
plates at different time points post inoculation. See
also Figure S2.
(D) Fum amounts in Fum-DUC were determined by
immunoblot. The proteasome subunit B (PrcB)
serves as loading control.
(E) Quantitative Fum immunoblotting in Fum-DUC
extracts from cultures on day 7 after atc addition.
Serial dilutions of WT extract were used for relative
quantification. PrcB served as loading control.
(F) Impact of Fum depletion on starved Mtb. WT
and Fum-DUC were cultured in PBS with 0.05%
tyloxapol for 10 days, before atc was added (ar-
row). CFU were determined by culturing serial di-
lutions on agar plates at different time points post
inoculation.
(G) Fum depletion in Fum-DUC during PBS star-
vation was analyzed by immunoblot. PrcB serves
as loading control. All data are representative of at
least two independent experiments. See also Fig-
ures S1 and S2.
Please cite this article in press as: Ruecker et al., Fumarase Deficiency Causes Protein and Metabolite Succination and Intoxicates Mycobacteriumtuberculosis, Cell Chemical Biology (2017), http://dx.doi.org/10.1016/j.chembiol.2017.01.005
Fum dispensable in the presence of a glycolytic carbon source
and a fatty acid source (Beste et al., 2011; de Carvalho et al.,
2010). Fumarase null mutants in other bacteria, and even eukary-
otes, have been reported to be viable (Carls and Hanson, 1971;
Mercado-Lubo et al., 2009; Song et al., 2013; Xu et al., 2013).
Interestingly, a small-molecule inhibitor of Fum that potently
and selectively inhibitsMtb Fum, via binding to an allosteric reg-
ulatory site, was recently reported (Kasbekar et al., 2016). How-
ever, this compound only slowed growth and failed to kill Mtb,
leaving the phenotypic consequences of Fum inactivation in
Mtb unexplored.
We sought to address this ambiguity and generated a condi-
tional knockdown strain of Mtb Fum. We found that Fum deple-
tion not only inhibited growth, but was also bactericidal in
replicating conditions in vitro and during acute and chronic
mouse infection. Surprisingly, and in contrast to previous studies
2 Cell Chemical Biology 24, 1–10, March 16, 2017
of other CCM enzymes in Mtb, we
were unable to metabolically complement
Fum deficiency, indicating a dominant-
negative toxicity. Metabolic profiling of
a Fum-depleted strain revealed severe
accumulation of fumarate. Immunoblot
analysis, followed by protein mass
spectrometry and metabolomic profiling,
further revealed that fumarate covalently
reacted with thiol residues resulting in
succination of numerous proteins andme-
tabolites, including mycothiol. The domi-
nant-negative consequences of Fum defi-
ciency and heightened susceptibility to
peroxide stress differentiate Mtb from
many other bacteria and eukaryotes, and identify Fum as poten-
tially attractive drug target.
RESULTS
Fum Depletion in Mtb Cannot Be Metabolically RescuedIn Vitro and Is Detrimental during Acute and ChronicMouse InfectionTo study the function of Fum inMtb, we generated a conditional
knockdown mutant (Fum-DUC) in strain H37Rv, using the previ-
ously described dual control (DUC) genetic switch in which
expression of Fum is controlled by both inducible transcriptional
silencing and proteolytic degradation (Kim et al., 2013) (Fig-
ure S1). Depletion of Fum following addition of anhydrotetracy-
cline (atc) inhibited Mtb growth and led to a rapid decline in
viability in nutrient-rich 7H9 medium (Figures 1B and 1C) and
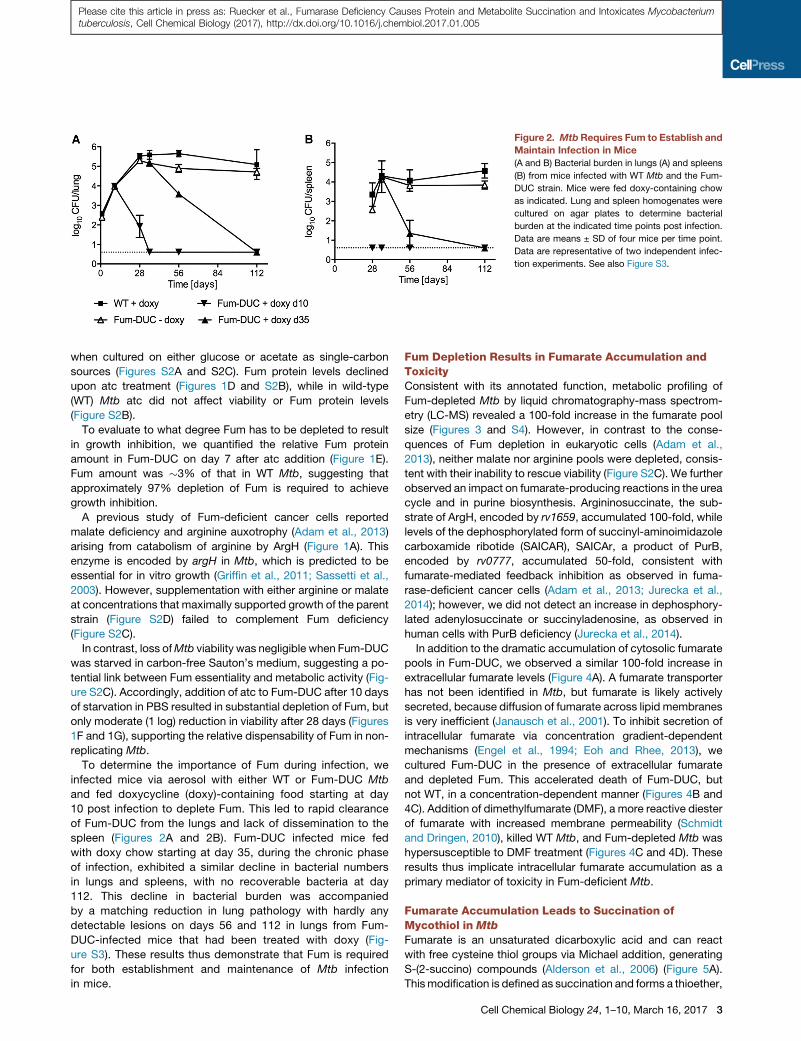
Figure 2. MtbRequires Fum to Establish and
Maintain Infection in Mice
(A and B) Bacterial burden in lungs (A) and spleens
(B) from mice infected with WT Mtb and the Fum-
DUC strain. Mice were fed doxy-containing chow
as indicated. Lung and spleen homogenates were
cultured on agar plates to determine bacterial
burden at the indicated time points post infection.
Data are means ± SD of four mice per time point.
Data are representative of two independent infec-
tion experiments. See also Figure S3.
Please cite this article in press as: Ruecker et al., Fumarase Deficiency Causes Protein and Metabolite Succination and Intoxicates Mycobacteriumtuberculosis, Cell Chemical Biology (2017), http://dx.doi.org/10.1016/j.chembiol.2017.01.005
when cultured on either glucose or acetate as single-carbon
sources (Figures S2A and S2C). Fum protein levels declined
upon atc treatment (Figures 1D and S2B), while in wild-type
(WT) Mtb atc did not affect viability or Fum protein levels
(Figure S2B).
To evaluate to what degree Fum has to be depleted to result
in growth inhibition, we quantified the relative Fum protein
amount in Fum-DUC on day 7 after atc addition (Figure 1E).
Fum amount was �3% of that in WT Mtb, suggesting that
approximately 97% depletion of Fum is required to achieve
growth inhibition.
A previous study of Fum-deficient cancer cells reported
malate deficiency and arginine auxotrophy (Adam et al., 2013)
arising from catabolism of arginine by ArgH (Figure 1A). This
enzyme is encoded by argH in Mtb, which is predicted to be
essential for in vitro growth (Griffin et al., 2011; Sassetti et al.,
2003). However, supplementation with either arginine or malate
at concentrations that maximally supported growth of the parent
strain (Figure S2D) failed to complement Fum deficiency
(Figure S2C).
In contrast, loss ofMtb viability was negligible when Fum-DUC
was starved in carbon-free Sauton’s medium, suggesting a po-
tential link between Fum essentiality and metabolic activity (Fig-
ure S2C). Accordingly, addition of atc to Fum-DUC after 10 days
of starvation in PBS resulted in substantial depletion of Fum, but
only moderate (1 log) reduction in viability after 28 days (Figures
1F and 1G), supporting the relative dispensability of Fum in non-
replicating Mtb.
To determine the importance of Fum during infection, we
infected mice via aerosol with either WT or Fum-DUC Mtb
and fed doxycycline (doxy)-containing food starting at day
10 post infection to deplete Fum. This led to rapid clearance
of Fum-DUC from the lungs and lack of dissemination to the
spleen (Figures 2A and 2B). Fum-DUC infected mice fed
with doxy chow starting at day 35, during the chronic phase
of infection, exhibited a similar decline in bacterial numbers
in lungs and spleens, with no recoverable bacteria at day
112. This decline in bacterial burden was accompanied
by a matching reduction in lung pathology with hardly any
detectable lesions on days 56 and 112 in lungs from Fum-
DUC-infected mice that had been treated with doxy (Fig-
ure S3). These results thus demonstrate that Fum is required
for both establishment and maintenance of Mtb infection
in mice.
Fum Depletion Results in Fumarate Accumulation andToxicityConsistent with its annotated function, metabolic profiling of
Fum-depleted Mtb by liquid chromatography-mass spectrom-
etry (LC-MS) revealed a 100-fold increase in the fumarate pool
size (Figures 3 and S4). However, in contrast to the conse-
quences of Fum depletion in eukaryotic cells (Adam et al.,
2013), neither malate nor arginine pools were depleted, consis-
tent with their inability to rescue viability (Figure S2C). We further
observed an impact on fumarate-producing reactions in the urea
cycle and in purine biosynthesis. Argininosuccinate, the sub-
strate of ArgH, encoded by rv1659, accumulated 100-fold, while
levels of the dephosphorylated form of succinyl-aminoimidazole
carboxamide ribotide (SAICAR), SAICAr, a product of PurB,
encoded by rv0777, accumulated 50-fold, consistent with
fumarate-mediated feedback inhibition as observed in fuma-
rase-deficient cancer cells (Adam et al., 2013; Jurecka et al.,
2014); however, we did not detect an increase in dephosphory-
lated adenylosuccinate or succinyladenosine, as observed in
human cells with PurB deficiency (Jurecka et al., 2014).
In addition to the dramatic accumulation of cytosolic fumarate
pools in Fum-DUC, we observed a similar 100-fold increase in
extracellular fumarate levels (Figure 4A). A fumarate transporter
has not been identified in Mtb, but fumarate is likely actively
secreted, because diffusion of fumarate across lipid membranes
is very inefficient (Janausch et al., 2001). To inhibit secretion of
intracellular fumarate via concentration gradient-dependent
mechanisms (Engel et al., 1994; Eoh and Rhee, 2013), we
cultured Fum-DUC in the presence of extracellular fumarate
and depleted Fum. This accelerated death of Fum-DUC, but
not WT, in a concentration-dependent manner (Figures 4B and
4C). Addition of dimethylfumarate (DMF), a more reactive diester
of fumarate with increased membrane permeability (Schmidt
and Dringen, 2010), killed WT Mtb, and Fum-depleted Mtb was
hypersusceptible to DMF treatment (Figures 4C and 4D). These
results thus implicate intracellular fumarate accumulation as a
primary mediator of toxicity in Fum-deficient Mtb.
Fumarate Accumulation Leads to Succination ofMycothiol in Mtb
Fumarate is an unsaturated dicarboxylic acid and can react
with free cysteine thiol groups via Michael addition, generating
S-(2-succino) compounds (Alderson et al., 2006) (Figure 5A).
Thismodification is defined as succination and forms a thioether,
Cell Chemical Biology 24, 1–10, March 16, 2017 3

Figure 3. Metabolic Consequences of Fum Deficiency
Intrabacterial pool sizes of selected metabolites in the indicated Mtb strains after 24 hr cultivation on filters on top of 7H9 medium with 0.2% glucose with or
without atc. Pool sizes are expressed as area under the curve normalized to protein content. Data are mean values ± SD of three biological replicates and are
representative of two independent experiments. Statistically significant differences between atc-treated and untreated cultures that were observed in two
biologically independent experiments are reported. *p% 0.05, **p% 0.005 by Student’s t test. The differences between pool sizes in atc-treated and untreated
WT were not reproducibly statistically significant. Enzymatic reactions are depicted as arrows; the dashed arrow indicates that the respective enzyme has not
been identified in Mtb; black dotted arrows indicate spontaneous dephosphorylation reactions. AICAR, 1-(5’-phosphoribosyl)-5-amino-4-imidazolecarbox-
amide; AMP, adenosine 5’-monophosphate; ArgH, argininosuccinate lyase; PurB, adenylosuccinate lyase; SAICAR, succinylaminoimidazole carboxamide
ribose-50-phosphate; SAICAr, succinylaminoimidazole carboxamide ribose. See also Figure S4.
Please cite this article in press as: Ruecker et al., Fumarase Deficiency Causes Protein and Metabolite Succination and Intoxicates Mycobacteriumtuberculosis, Cell Chemical Biology (2017), http://dx.doi.org/10.1016/j.chembiol.2017.01.005
in contrast to succinylation, which generates a thioester when
succinyl-coenzyme A reacts with lysine residues (Zhang et al.,
2011). In eukaryotes, this reactivity results in succination
of glutathione (GSH) (Sullivan et al., 2013). Accordingly, we
observed succination of mycothiol (MSH), the mycobacterial
counterpart (Figures 5B and 5C). In addition to succinated
MSH, we also detected free succinocysteine, the reaction prod-
uct of cysteine and fumarate (Figures 5D and 5E).
Fumarate Accumulation Leads to Succination ofProteinsSuccination results in a similar modification of protein cysteine
residues in eukaryotes (Alderson et al., 2006; Merkley et al.,
2014). We therefore analyzed whole-cell extracts from WT and
Fum-DUC Mtb using an antibody against S-(2-succino)cysteine
(2SC) (Nagai et al., 2007) and observed numerous 2SC-positive
signals specifically in lysates of Fum-DUC treated with atc (Fig-
ure 6A). Expression of fumarase with an active site mutation
4 Cell Chemical Biology 24, 1–10, March 16, 2017
(S318A) neither restored viability nor prevented succination,
confirming that succination was attributable to loss of Fum activ-
ity (Figure S5A). In addition, succination occurred specifically in
cells accumulating fumarate and was not just a marker of cell
death (Figure S5B).
Multiple proteins appeared succinated in Fum-DUC, with the
most intense signal at a molecular range of 75–100 kDa, which
was also present in WT treated with DMF (Figure S5C). To iden-
tify succinated proteins, we analyzed trypsin-digested gel pieces
of the corresponding molecular weight range by liquid chroma-
tography-tandem mass spectrometry (LC-MS/MS) querying
the data for the expected mass shifts corresponding to succina-
tion (H4C4O4: 116.010959 Da) and dimethylsuccination (H8C6O4:
144.042259 Da). We identified three proteins containing succi-
nated cysteine residues in both Fum-DUC-treated samples,
as well as WT Mtb-treated with DMF (Table 1). These corre-
sponded to aminopeptidase N (Rv2467), catalase-peroxidase
KatG (Rv1908c), and the 1,4-alpha-glucan branching enzyme
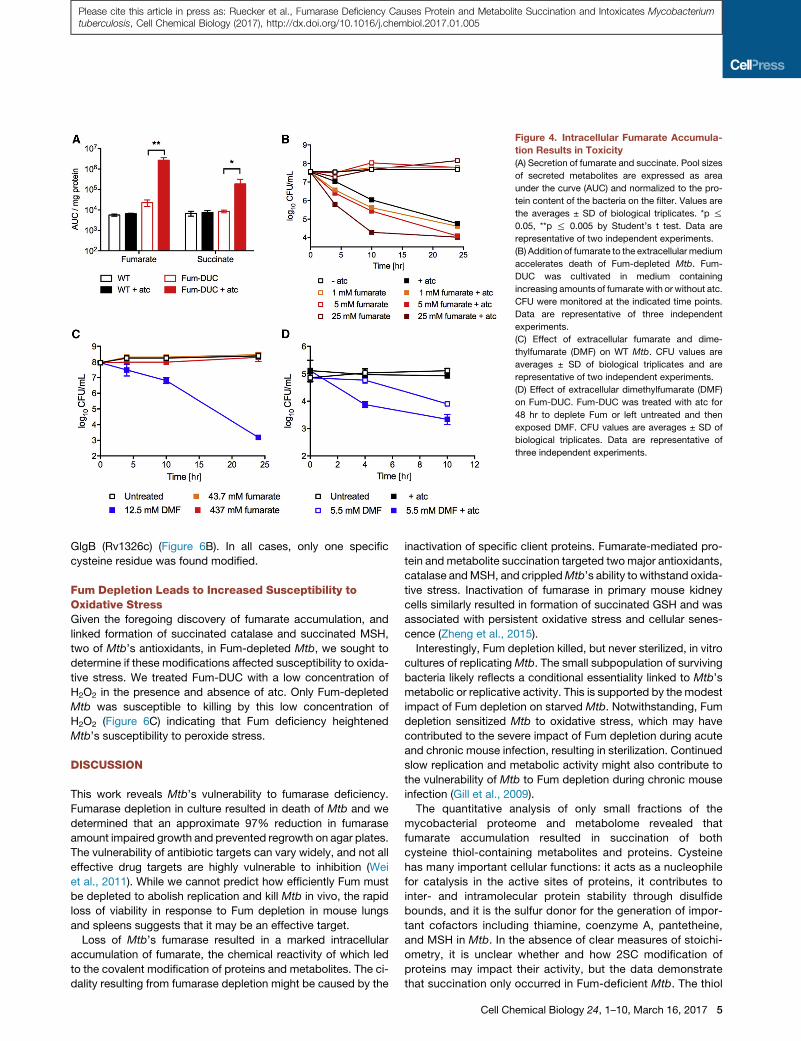
Figure 4. Intracellular Fumarate Accumula-
tion Results in Toxicity
(A) Secretion of fumarate and succinate. Pool sizes
of secreted metabolites are expressed as area
under the curve (AUC) and normalized to the pro-
tein content of the bacteria on the filter. Values are
the averages ± SD of biological triplicates. *p %
0.05, **p % 0.005 by Student’s t test. Data are
representative of two independent experiments.
(B) Addition of fumarate to the extracellular medium
accelerates death of Fum-depleted Mtb. Fum-
DUC was cultivated in medium containing
increasing amounts of fumarate with or without atc.
CFU were monitored at the indicated time points.
Data are representative of three independent
experiments.
(C) Effect of extracellular fumarate and dime-
thylfumarate (DMF) on WT Mtb. CFU values are
averages ± SD of biological triplicates and are
representative of two independent experiments.
(D) Effect of extracellular dimethylfumarate (DMF)
on Fum-DUC. Fum-DUC was treated with atc for
48 hr to deplete Fum or left untreated and then
exposed DMF. CFU values are averages ± SD of
biological triplicates. Data are representative of
three independent experiments.
Please cite this article in press as: Ruecker et al., Fumarase Deficiency Causes Protein and Metabolite Succination and Intoxicates Mycobacteriumtuberculosis, Cell Chemical Biology (2017), http://dx.doi.org/10.1016/j.chembiol.2017.01.005
GlgB (Rv1326c) (Figure 6B). In all cases, only one specific
cysteine residue was found modified.
Fum Depletion Leads to Increased Susceptibility toOxidative StressGiven the foregoing discovery of fumarate accumulation, and
linked formation of succinated catalase and succinated MSH,
two of Mtb’s antioxidants, in Fum-depleted Mtb, we sought to
determine if these modifications affected susceptibility to oxida-
tive stress. We treated Fum-DUC with a low concentration of
H2O2 in the presence and absence of atc. Only Fum-depleted
Mtb was susceptible to killing by this low concentration of
H2O2 (Figure 6C) indicating that Fum deficiency heightened
Mtb’s susceptibility to peroxide stress.
DISCUSSION
This work reveals Mtb’s vulnerability to fumarase deficiency.
Fumarase depletion in culture resulted in death of Mtb and we
determined that an approximate 97% reduction in fumarase
amount impaired growth and prevented regrowth on agar plates.
The vulnerability of antibiotic targets can vary widely, and not all
effective drug targets are highly vulnerable to inhibition (Wei
et al., 2011). While we cannot predict how efficiently Fum must
be depleted to abolish replication and kill Mtb in vivo, the rapid
loss of viability in response to Fum depletion in mouse lungs
and spleens suggests that it may be an effective target.
Loss of Mtb’s fumarase resulted in a marked intracellular
accumulation of fumarate, the chemical reactivity of which led
to the covalent modification of proteins and metabolites. The ci-
dality resulting from fumarase depletion might be caused by the
inactivation of specific client proteins. Fumarate-mediated pro-
tein and metabolite succination targeted twomajor antioxidants,
catalase andMSH, and crippledMtb’s ability to withstand oxida-
tive stress. Inactivation of fumarase in primary mouse kidney
cells similarly resulted in formation of succinated GSH and was
associated with persistent oxidative stress and cellular senes-
cence (Zheng et al., 2015).
Interestingly, Fum depletion killed, but never sterilized, in vitro
cultures of replicatingMtb. The small subpopulation of surviving
bacteria likely reflects a conditional essentiality linked to Mtb’s
metabolic or replicative activity. This is supported by the modest
impact of Fum depletion on starved Mtb. Notwithstanding, Fum
depletion sensitized Mtb to oxidative stress, which may have
contributed to the severe impact of Fum depletion during acute
and chronic mouse infection, resulting in sterilization. Continued
slow replication and metabolic activity might also contribute to
the vulnerability of Mtb to Fum depletion during chronic mouse
infection (Gill et al., 2009).
The quantitative analysis of only small fractions of the
mycobacterial proteome and metabolome revealed that
fumarate accumulation resulted in succination of both
cysteine thiol-containing metabolites and proteins. Cysteine
has many important cellular functions: it acts as a nucleophile
for catalysis in the active sites of proteins, it contributes to
inter- and intramolecular protein stability through disulfide
bounds, and it is the sulfur donor for the generation of impor-
tant cofactors including thiamine, coenzyme A, pantetheine,
and MSH in Mtb. In the absence of clear measures of stoichi-
ometry, it is unclear whether and how 2SC modification of
proteins may impact their activity, but the data demonstrate
that succination only occurred in Fum-deficient Mtb. The thiol
Cell Chemical Biology 24, 1–10, March 16, 2017 5

Figure 5. Fumarate Accumulation Leads to
Metabolite Succination
(A) The electrophile fumarate can react with
cysteine thiol residues in a Michael addition
generating S-(2-succino) compounds.
(B) Intracellular pool sizes of a metabolite with a
mass to charge ratio (m/z) of 603.158 in extracts
fromMtb Fum-DUC treated with atc for 24 hr. *p%
0.05, by Student’s t test.
(C) Comparison ofMS-MS fragmentation spectra of
603.158 with fragmentation spectra of MSH after
incubation with fumarate.
(D) Intracellular pool sizes of a metabolite with
an m/z of 238.038. *p % 0.05, by Student’s t test.
(E) Comparison of MS/MS fragmentation spectra
of 238.038 with fragmentation spectra of succino-
cysteine.
Please cite this article in press as: Ruecker et al., Fumarase Deficiency Causes Protein and Metabolite Succination and Intoxicates Mycobacteriumtuberculosis, Cell Chemical Biology (2017), http://dx.doi.org/10.1016/j.chembiol.2017.01.005
of MSH is the major redox buffer in Mtb (found in millimolar
quantities in the cytoplasm of Mtb; Buchmeier et al., 2003),
and is rendered potentially idle by succination. Loss of
MSH biosynthesis, although dispensable for viability of Mtb,
increased susceptibility to hydrogen peroxide and low pH
(Buchmeier et al., 2003, 2006). This is consistent with our
observation of increased susceptibility of Fum-depleted Mtb
to peroxide stress.
Enzymes function within the context of extensively inter-
connected pathways and inhibition of a single enzyme
can impact multiple pathways (Eoh and Rhee, 2013, 2014).
This is also the case with Fum. Moreover, the dominant-
negative toxicity of Fum deficiency may kill Mtb via effects
on multiple pathways, albeit restricted to replicating Mtb.
Mtb’s vulnerability to Fum depletion is unique and not
observed in other organisms, revealing Fum as a poten-
tially attractive, species-specific target for new and urgently
needed TB drugs.
SIGNIFICANCE
The development of novel chemotherapeutics against
tuberculosis requires a better understanding of the essential
6 Cell Chemical Biology 24, 1–10, March 16, 2017
processes in Mycobacterium tubercu-
losis (Mtb). Fumarase functions in the
tricarboxylic acid (TCA) cycle, which
plays a central role inMtb’s metabolism
and pathogenesis. Here, we demon-
strate that fumarase is essential for
growth of Mtb, in contrast to its
dispensability inmany other organisms.
Fumarase depletion perturbed essen-
tial metabolic pathways and led to
intracellular fumarate accumulation,
which in turn caused metabolite and
protein succination, a covalent chemi-
cal modification of cysteine thiol resi-
dues. This affected two of Mtb’s major
antioxidants, catalase and mycothiol,
causing hypersusceptibility to oxidative
stress. Importantly, Fumarase deple-
tion killed Mtb not only in vitro, but also during the acute
and chronic phases of infection in a mouse model of tuber-
culosis. These studies revealMtb’s vulnerability to fumarase
depletion and identify fumarase as a potentially attractive,
species-selective drug target in Mtb.
EXPERIMENTAL PROCEDURES
Culture Conditions
Mtb H37Rv WT and mutant strains were grown aerated in standing flasks at
37�C with 5% CO2 in Middlebrook 7H9 liquid medium containing 0.2%
glucose, 0.5% glycerol, 5% BSA, 0.085% sodium chloride, and 0.05% Tween
80. For agar plates Middlebrook 7H10 agar with 10% commercial OADC sup-
plement (final concentration 0.5% BSA, 0.2% glucose, 0.085% NaCl, 0.006%
oleic acid, and 0.0003% catalase) and 0.5% glycerol was used.
Carbon-defined minimal medium consisted of modified Sauton’s medium
(Allen, 1998): asparagine was replaced by 0.5 g/L ammonium sulfate and
Tween 80 was replaced by 0.05% tyloxapol. Acetate and or glucose were
added to yield a concentration of 0.2% (w/v) each. Arginine and malate were
used as indicated. To ensure adaptation of the bacteria to the medium, the
bacteria were cultured in the respective medium for 7 days before atc
was added.
H37Rv DFba was cultured in the presence of fatty acid-free BSA and
with 0.4% glycerol, 0.4% glucose, and 0.05% tyloxapol as described
previously (Puckett et al., 2014). Antibiotics were used in the following

Figure 6. Fumarate Accumulation Leads to Protein Succination
(A) Detection of 2SC proteins in lysates of Fum-DUC grown with or without atc. For each condition 5 mg protein was loaded. Succinated GAPDH (Alderson et al.,
2006) was loaded as a positive control for succination (labeled C). PrcB serves as loading control.
(B) Tandem MS spectra of the triply charged tryptic cysteine-containing peptide: LQV TYE GCE PHT VAD AYR, from 1,4-alpha-glucan branching enzyme GlgB
(Rv1326c). Fragment ion (y and b ions)-annotated spectra are shown (top to bottom) of the iodoacetamide-modified cysteine peptide (m/z of 703.6605) matched
in the ‘‘WT + atc’’ sample, the succinated peptide (m/z of 723.3422) matched in the ‘‘Fum-DUC + atc,’’ sample and the dimethylsuccinated peptide (m/z of
732.6677) matched in the ‘‘WT + DMF’’ sample, respectively. All peptides were matched with mass accuracy better than 2 ppm. See also Figure S5.
(C) Fum deficiency increases susceptibility to hydrogen peroxide (H2O2). Fum-DUC was exposed to 5 mMH2O2 in the presence or absence of atc for 30 hr. CFU
values are averages ± SD of biological triplicates. Data are representative of three independent experiments.
Please cite this article in press as: Ruecker et al., Fumarase Deficiency Causes Protein and Metabolite Succination and Intoxicates Mycobacteriumtuberculosis, Cell Chemical Biology (2017), http://dx.doi.org/10.1016/j.chembiol.2017.01.005
concentrations: hygromycin (50 mg/mL), zeocin (25 mg/mL), and kanamycin
(25 mg/mL). atc was added at 500 ng/mL to deplete Fum.
For metabolite analysis,Mtb strains at OD580 �1 were seeded onto 0.22 mM
nitrocellulose filters on top of 7H10 agar plates containing 0.5% BSA, 0.085%
NaCl, 0.2% glucose, and 0.5% glycerol. Every 3 days, filters were moved to
fresh plates, to gain biomass. After 7 days filters were transferred to a swim-
ming pool setup (Eoh and Rhee, 2013; Maksymiuk et al., 2015), using liquid
7H9 medium with or without 500 ng/mL atc.
To assess the susceptibility to peroxide, Fum-DUC was treated either with
5 mM hydrogen peroxide or atc or both. After 4 hr, 10 hr, and 24 hr, 5 mM
hydrogen peroxide was replenished.
Metabolite Extraction, Detection, Fragmentation, and Analysis
To extract metabolites bacteria were quenched in cold LC-MS-grade ace-
tonitrile:methanol:water (40:40:20), scraped off of filters, and mechanically
lysed using a bead beater prior to metabolite analysis. Lysates were ster-
ilized using a 0.22 mm filter. Secreted fumarate and succinate were quan-
tified in the medium underneath the filter. Prior to LC-MS analysis, extracts
were diluted 1:1 with mobile phase (acetonitrile with 0.5% formic acid) and
centrifuged (10 min 10,000 3 g). LC-MS analysis was performed using
an Agilent 1,200 Series Liquid Chromatography system with a Cogent
Diamond Hydride Type C column (Microsolv Technologies) coupled to
an Agilent Accurate-Mass TOF 6230, as described previously (Eoh and
Rhee, 2013).
Standards for metabolite identification were obtained from Sigma, except
S-(2-succinyl)-L-cysteine (www.polypeptide.com), SAICAr (MyBioSource),
and succinyladenosine (MyBioSource). Succinated MSH was prepared by
incubating MSH and 500 mM fumarate for 24 hr at 37�C. Targeted data anal-
ysis was performed using Profinder B.06.00 software (Agilent). Metabolite
concentrations were normalized to biomass based on measurement of
residual peptide content in individual samples using the BCA assay (Thermo
Scientific). Fragmentation spectra were obtained using the same conditions
Cell Chemical Biology 24, 1–10, March 16, 2017 7
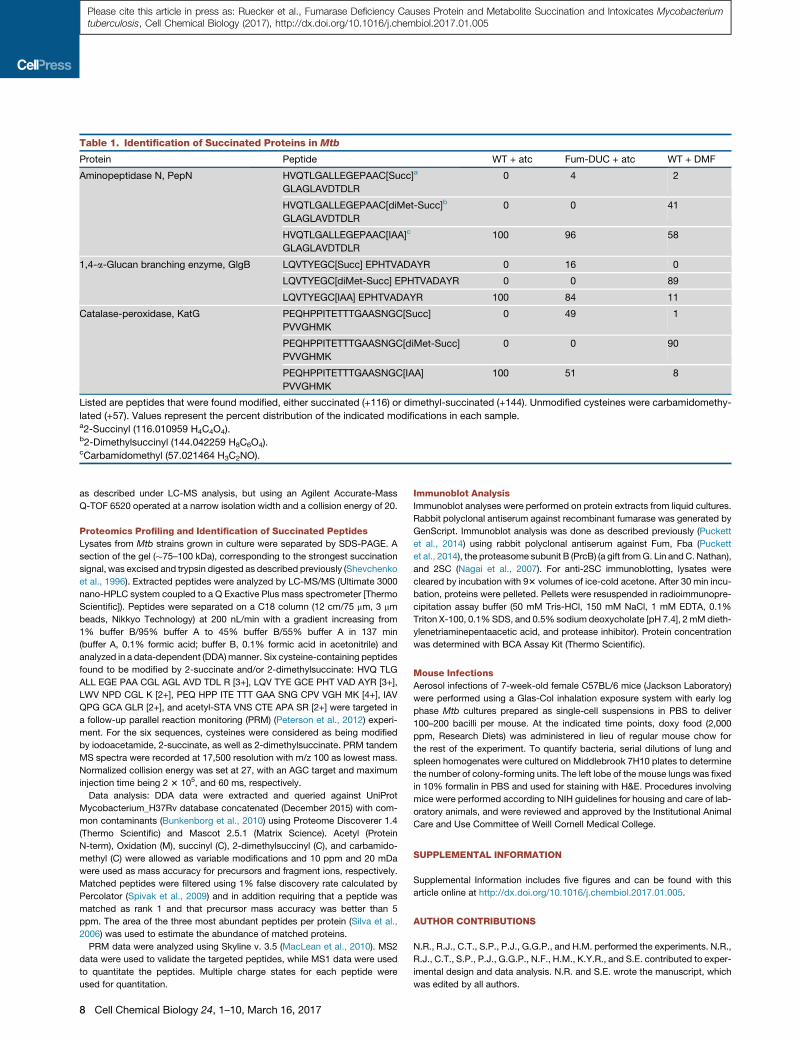
Table 1. Identification of Succinated Proteins in Mtb
Protein Peptide WT + atc Fum-DUC + atc WT + DMF
Aminopeptidase N, PepN HVQTLGALLEGEPAAC[Succ]a
GLAGLAVDTDLR
0 4 2
HVQTLGALLEGEPAAC[diMet-Succ]b
GLAGLAVDTDLR
0 0 41
HVQTLGALLEGEPAAC[IAA]c
GLAGLAVDTDLR
100 96 58
1,4-a-Glucan branching enzyme, GlgB LQVTYEGC[Succ] EPHTVADAYR 0 16 0
LQVTYEGC[diMet-Succ] EPHTVADAYR 0 0 89
LQVTYEGC[IAA] EPHTVADAYR 100 84 11
Catalase-peroxidase, KatG PEQHPPITETTTGAASNGC[Succ]
PVVGHMK
0 49 1
PEQHPPITETTTGAASNGC[diMet-Succ]
PVVGHMK
0 0 90
PEQHPPITETTTGAASNGC[IAA]
PVVGHMK
100 51 8
Listed are peptides that were found modified, either succinated (+116) or dimethyl-succinated (+144). Unmodified cysteines were carbamidomethy-
lated (+57). Values represent the percent distribution of the indicated modifications in each sample.a2-Succinyl (116.010959 H4C4O4).b2-Dimethylsuccinyl (144.042259 H8C6O4).cCarbamidomethyl (57.021464 H3C2NO).
Please cite this article in press as: Ruecker et al., Fumarase Deficiency Causes Protein and Metabolite Succination and Intoxicates Mycobacteriumtuberculosis, Cell Chemical Biology (2017), http://dx.doi.org/10.1016/j.chembiol.2017.01.005
as described under LC-MS analysis, but using an Agilent Accurate-Mass
Q-TOF 6520 operated at a narrow isolation width and a collision energy of 20.
Proteomics Profiling and Identification of Succinated Peptides
Lysates from Mtb strains grown in culture were separated by SDS-PAGE. A
section of the gel (�75–100 kDa), corresponding to the strongest succination
signal, was excised and trypsin digested as described previously (Shevchenko
et al., 1996). Extracted peptides were analyzed by LC-MS/MS (Ultimate 3000
nano-HPLC system coupled to a Q Exactive Plus mass spectrometer [Thermo
Scientific]). Peptides were separated on a C18 column (12 cm/75 mm, 3 mm
beads, Nikkyo Technology) at 200 nL/min with a gradient increasing from
1% buffer B/95% buffer A to 45% buffer B/55% buffer A in 137 min
(buffer A, 0.1% formic acid; buffer B, 0.1% formic acid in acetonitrile) and
analyzed in a data-dependent (DDA)manner. Six cysteine-containing peptides
found to be modified by 2-succinate and/or 2-dimethylsuccinate: HVQ TLG
ALL EGE PAA CGL AGL AVD TDL R [3+], LQV TYE GCE PHT VAD AYR [3+],
LWV NPD CGL K [2+], PEQ HPP ITE TTT GAA SNG CPV VGH MK [4+], IAV
QPG GCA GLR [2+], and acetyl-STA VNS CTE APA SR [2+] were targeted in
a follow-up parallel reaction monitoring (PRM) (Peterson et al., 2012) experi-
ment. For the six sequences, cysteines were considered as being modified
by iodoacetamide, 2-succinate, as well as 2-dimethylsuccinate. PRM tandem
MS spectra were recorded at 17,500 resolution with m/z 100 as lowest mass.
Normalized collision energy was set at 27, with an AGC target and maximum
injection time being 2 3 105, and 60 ms, respectively.
Data analysis: DDA data were extracted and queried against UniProt
Mycobacterium_H37Rv database concatenated (December 2015) with com-
mon contaminants (Bunkenborg et al., 2010) using Proteome Discoverer 1.4
(Thermo Scientific) and Mascot 2.5.1 (Matrix Science). Acetyl (Protein
N-term), Oxidation (M), succinyl (C), 2-dimethylsuccinyl (C), and carbamido-
methyl (C) were allowed as variable modifications and 10 ppm and 20 mDa
were used as mass accuracy for precursors and fragment ions, respectively.
Matched peptides were filtered using 1% false discovery rate calculated by
Percolator (Spivak et al., 2009) and in addition requiring that a peptide was
matched as rank 1 and that precursor mass accuracy was better than 5
ppm. The area of the three most abundant peptides per protein (Silva et al.,
2006) was used to estimate the abundance of matched proteins.
PRM data were analyzed using Skyline v. 3.5 (MacLean et al., 2010). MS2
data were used to validate the targeted peptides, while MS1 data were used
to quantitate the peptides. Multiple charge states for each peptide were
used for quantitation.
8 Cell Chemical Biology 24, 1–10, March 16, 2017
Immunoblot Analysis
Immunoblot analyses were performed on protein extracts from liquid cultures.
Rabbit polyclonal antiserum against recombinant fumarase was generated by
GenScript. Immunoblot analysis was done as described previously (Puckett
et al., 2014) using rabbit polyclonal antiserum against Fum, Fba (Puckett
et al., 2014), the proteasome subunit B (PrcB) (a gift fromG. Lin and C. Nathan),
and 2SC (Nagai et al., 2007). For anti-2SC immunoblotting, lysates were
cleared by incubation with 93 volumes of ice-cold acetone. After 30 min incu-
bation, proteins were pelleted. Pellets were resuspended in radioimmunopre-
cipitation assay buffer (50 mM Tris-HCl, 150 mM NaCl, 1 mM EDTA, 0.1%
Triton X-100, 0.1%SDS, and 0.5% sodium deoxycholate [pH 7.4], 2 mMdieth-
ylenetriaminepentaacetic acid, and protease inhibitor). Protein concentration
was determined with BCA Assay Kit (Thermo Scientific).
Mouse Infections
Aerosol infections of 7-week-old female C57BL/6 mice (Jackson Laboratory)
were performed using a Glas-Col inhalation exposure system with early log
phase Mtb cultures prepared as single-cell suspensions in PBS to deliver
100–200 bacilli per mouse. At the indicated time points, doxy food (2,000
ppm, Research Diets) was administered in lieu of regular mouse chow for
the rest of the experiment. To quantify bacteria, serial dilutions of lung and
spleen homogenates were cultured on Middlebrook 7H10 plates to determine
the number of colony-forming units. The left lobe of the mouse lungs was fixed
in 10% formalin in PBS and used for staining with H&E. Procedures involving
mice were performed according to NIH guidelines for housing and care of lab-
oratory animals, and were reviewed and approved by the Institutional Animal
Care and Use Committee of Weill Cornell Medical College.
SUPPLEMENTAL INFORMATION
Supplemental Information includes five figures and can be found with this
article online at http://dx.doi.org/10.1016/j.chembiol.2017.01.005.
AUTHOR CONTRIBUTIONS
N.R., R.J., C.T., S.P., P.J., G.G.P., and H.M. performed the experiments. N.R.,
R.J., C.T., S.P., P.J., G.G.P., N.F., H.M., K.Y.R., and S.E. contributed to exper-
imental design and data analysis. N.R. and S.E. wrote the manuscript, which
was edited by all authors.

Please cite this article in press as: Ruecker et al., Fumarase Deficiency Causes Protein and Metabolite Succination and Intoxicates Mycobacteriumtuberculosis, Cell Chemical Biology (2017), http://dx.doi.org/10.1016/j.chembiol.2017.01.005
ACKNOWLEDGMENTS
We thank Hyungjin Eoh for assistance with preliminary experiments; Dirk
Schnappinger for discussions and advice; Gang Lin and Carl Nathan for
PrcB-specific antiserum; and Weizhen Xu for help with data analysis. This
work was supported by a grant from the NIH (AI063446 to S.E.) and grants
from the Bill and Melinda Gates Foundation (grants 42848 and OPP1024065
to Dirk Schnappinger).
Received: September 11, 2016
Revised: December 7, 2016
Accepted: January 19, 2017
Published: February 16, 2017
REFERENCES
Adam, J., Yang, M., Bauerschmidt, C., Kitagawa, M., O’Flaherty, L.,
Maheswaran, P., Ozkan, G., Sahgal, N., Baban, D., Kato, K., et al. (2013). A
role for cytosolic fumarate hydratase in urea cycle metabolism and renal
neoplasia. Cell Rep. 3, 1440–1448.
Alderson, N.L., Wang, Y., Blatnik, M., Frizzell, N., Walla, M.D., Lyons, T.J., Alt,
N., Carson, J.A., Nagai, R., Thorpe, S.R., and Baynes, J.W. (2006). S-(2-
Succinyl)cysteine: a novel chemical modification of tissue proteins by a
Krebs cycle intermediate. Arch. Biochem. Biophys. 450, 1–8.
Allen, B.W. (1998). Mycobacteria: general culture methodology and safety
considerations. In Mycobacteria Protocols, T. Parish and D.M. Roberts, eds.
(Humana Press), pp. 15–30.
Beste, D.J.V., Bonde, B., Hawkins, N.,Ward, J.L., Beale, M.H., Noack, S., Noh,
K., Kruger, N.J., Ratcliffe, R.G., and McFadden, J. (2011). 13C Metabolic flux
analysis identifies an unusual route for pyruvate dissimilation in mycobacteria
which requires isocitrate lyase and carbon dioxide fixation. PLoS Pathog. 7,
e1002091.
Buchmeier, N.A., Newton, G.L., Koledin, T., and Fahey, R.C. (2003).
Association of mycothiol with protection of Mycobacterium tuberculosis
from toxic oxidants and antibiotics. Mol. Microbiol. 47, 1723–1732.
Buchmeier, N.A., Newton, G.L., and Fahey, R.C. (2006). A mycothiol synthase
mutant of Mycobacterium tuberculosis has an altered thiol-disulfide content
and limited tolerance to stress. J. Bacteriol. 188, 6245–6252.
Bunkenborg, J., Garcıa, G.E., Paz, M.I.P., Andersen, J.S., and Molina, H.
(2010). The minotaur proteome: avoiding cross-species identifications
deriving from bovine serum in cell culture models. Proteomics 10, 3040–3044.
Carls, R.A., and Hanson, R.S. (1971). Isolation and characterization of tricar-
boxylic acid cycle mutants of Bacillus subtilis. J. Bacteriol. 106, 848–855.
Cole, S.T., Brosch, R., Parkhill, J., Garnier, T., Churcher, C., Harris, D., Gordon,
S., Eiglmeier, K., Gas, S., and Barry, C.E. (1998). Deciphering the biology of
Mycobacterium tuberculosis from the complete genome sequence. Nature
393, 537–544.
de Carvalho, L.P.S., Fischer, S.M., Marrero, J., Nathan, C., Ehrt, S., and Rhee,
K.Y. (2010). Metabolomics of Mycobacterium tuberculosis reveals compart-
mentalized co-catabolism of carbon substrates. Chem. Biol. 17, 1122–1131.
Engel, P., Kramer, R., andUnden, G. (1994). Transport of C4-dicarboxylates by
anaerobically grown Escherichia coli. Energetics andmechanism of exchange,
uptake and efflux. Eur. J. Biochem. 222, 605–614.
Eoh, H., and Rhee, K.Y. (2013). Multifunctional essentiality of succinate meta-
bolism in adaptation to hypoxia in Mycobacterium tuberculosis. Proc. Natl.
Acad. Sci. USA 110, 6554–6559.
Eoh, H., and Rhee, K.Y. (2014). Methylcitrate cycle defines the bactericidal es-
sentiality of isocitrate lyase for survival of Mycobacterium tuberculosison fatty
acids. Proc. Natl. Acad. Sci. USA 111, 4976–4981.
Gill, W.P., Harik, N.S., Whiddon, M.R., Liao, R.P., Mittler, J.E., and Sherman,
D.R. (2009). A replication clock for Mycobacterium tuberculosis. Nat. Med.
15, 211–214.
Griffin, J.E., Gawronski, J.D., Dejesus, M.A., Ioerger, T.R., Akerley, B.J., and
Sassetti, C.M. (2011). High-resolution phenotypic profiling defines genes
essential for mycobacterial growth and cholesterol catabolism. PLoS
Pathog. 7, e1002251.
Janausch, I., Kim, O., and Unden, G. (2001). DctA- and Dcu-independent
transport of succinate in Escherichia coli: contribution of diffusion and of alter-
native carriers. Arch. Microbiol. 176, 224–230.
Jurecka, A., Zikanova, M., Kmoch, S., and Tylki-Szyma�nska, A. (2014).
Adenylosuccinate lyase deficiency. J. Inherit. Metab. Dis. 38, 231–242.
Kasbekar, M., Fischer, G., Mott, B.T., Yasgar, A., Hyvonen, M., Boshoff,
H.I.M., Abell, C., Barry, C.E., and Thomas, C.J. (2016). Selective small mole-
cule inhibitor of the Mycobacterium tuberculosis fumarate hydratase reveals
an allosteric regulatory site. Proc. Natl. Acad. Sci. USA 113, 7503–7508.
Kim, J.-H., O’Brien, K.M., Sharma, R., Boshoff, H.I.M., Rehren, G.,
Chakraborty, S., Wallach, J.B., Monteleone, M., Wilson, D.J., Aldrich, C.C.,
et al. (2013). A genetic strategy to identify targets for the development of drugs
that prevent bacterial persistence. Proc. Natl. Acad. Sci. USA 110,
19095–19100.
MacLean, B., Tomazela, D.M., Shulman, N., Chambers, M., Finney, G.L.,
Frewen, B., Kern, R., Tabb, D.L., Liebler, D.C., and MacCoss, M.J. (2010).
Skyline: an open source document editor for creating and analyzing targeted
proteomics experiments. Bioinformatics 26, 966–968.
Maksymiuk, C., Balakrishnan, A., Bryk, R., Rhee, K.Y., and Nathan, C.F.
(2015). E1 of a-ketoglutarate dehydrogenase defends Mycobacterium tuber-
culosis against glutamate cataplerosis and nit oxidative stress. Proc. Natl.
Acad. Sci. USA 112, E5834–E5843.
Marrero, J., Rhee, K.Y., Schnappinger, D., Pether, K., and Ehrt, S. (2010).
Gluconeogenic carbon flow of tricarboxylic acid cycle intermediates is critical
forMycobacterium tuberculosis to establish andmaintain infection. Proc. Natl.
Acad. Sci. USA 107, 9819–9824.
Marrero, J., Trujillo, C., Rhee, K.Y., and Ehrt, S. (2013). Glucose phosphoryla-
tion is required for Mycobacterium tuberculosis persistence in mice. PLoS
Pathog. 9, e1003116.
McKinney, J., Homer zoo Betrump, K., Munoz-Elıas, E., Maczka, A., Chen, B.,
Chan, W., Swenson, D., Sacchetti Ni, J., Jacobs, W., and Russell, D. (2000).
Persistence of Mycobacterium tuberculosis in macrophages and mice re-
quires the glyoxylate shunt enzyme isocitrate lyase. Nature 406, 735–738.
Mechaly, A.E., Haus, A., Miras, I., Barilone, N., Weber, P., Shepard, W., Alzari,
P.M., and Bellinzoni, M. (2012). Conformational changes upon ligand binding
in the essential class II fumarase Rv1098c from Mycobacterium tuberculosis.
FEBS Lett. 586, 1606–1611.
Mercado-Lubo, R., Leatham, M.P., Conway, T., and Cohen, P.S. (2009).
Salmonella enterica serovar Typhimurium mutants unable to convert malate
to pyruvate and oxaloacetate are avirulent and immunogenic in BALB/c
mice. Infect. Immun. 77, 1397–1405.
Merkley, E.D., Metz, T.O., Smith, R.D., Baynes, J.W., and Frizzell, N. (2014).
The succinated proteome. Mass Spectrom. Rev. 33, 98–109.
Munoz-Elıas, E., and McKinney, J.D. (2005). Mycobacterium tuberculosis iso-
citrate lyases 1 and 2 are jointly required for in vivo growth and virulence. Nat.
Med. 11, 638–644.
Nagai, R., Brock, J.W., Blatnik, M., Baatz, J.E., Bethard, J., Walla, M.D.,
Thorpe, S.R., Baynes, J.W., and Frizzell, N. (2007). Succination of protein thiols
during adipocyte maturation: a biomarker of mitochondrial stress. J. Biol.
Chem. 282, 34219–34228.
Pandey, A.K., and Sassetti, C.M. (2008). Mycobacterial persistence requires
the utilization of host cholesterol. Proc. Natl. Acad. Sci. USA 105, 4376–4380.
Park, S.J., and Gunsalus, R.P. (1995). Oxygen, iron, carbon, and superoxide
control of the fumarase fumA and fumC genes of Escherichia coli: role of the
arcA, fnr, and soxR gene products. J. Bacteriol. 177, 6255–6262.
Peterson, A.C., Russell, J.D., Bailey, D.J., Westphall, M.S., and Coon, J.J.
(2012). Parallel reaction monitoring for high resolution and high mass accuracy
quantitative, targeted proteomics. Mol. Cell. Proteomics 11, 1475–1488.
Puckett, S., Trujillo, C., Eoh, H., Marrero, J., Spencer, J., Jackson, M.,
Schnappinger, D., Rhee, K., and Ehrt, S. (2014). Inactivation of fructose-1,6-bi-
sphosphate aldolase prevents optimal co-catabolism of glycolytic and
Cell Chemical Biology 24, 1–10, March 16, 2017 9

Please cite this article in press as: Ruecker et al., Fumarase Deficiency Causes Protein and Metabolite Succination and Intoxicates Mycobacteriumtuberculosis, Cell Chemical Biology (2017), http://dx.doi.org/10.1016/j.chembiol.2017.01.005
gluconeogenic carbon substrates in Mycobacterium tuberculosis. PLoS
Pathog. 10, e1004144.
Sassetti, C.M., Boyd, D.H., and Rubin, E.J. (2003). Genes required for myco-
bacterial growth defined by high density mutagenesis. Mol. Microbiol.
48, 77–84.
Schmidt, M.M., and Dringen, R. (2010). Fumaric acid diesters deprive cultured
primary astrocytes rapidly of glutathione. Neurochem. Int. 57, 460–467.
Shevchenko, A., Wilm,M., Vorm, O., andMann,M. (1996). Mass spectrometric
sequencing of proteins from silver-stained polyacrylamide gels. Anal. Chem.
68, 850–858.
Silva, J.C., Gorenstein, M.V., Li, G.-Z., Vissers, J.P.C., and Geromanos, S.J.
(2006). Absolute quantification of proteins by LCMSE: a virtue of parallel MS
acquisition. Mol. Cell. Proteomics 5, 144–156.
Song, C.W., Kim, D.I., Choi, S., Jang, J.W., and Lee, S.Y. (2013). Metabolic en-
gineering of Escherichia coli for the production of fumaric acid. Biotechnol.
Bioeng. 110, 2025–2034.
Spivak, M., Weston, J., Bottou, L., K€all, L., and Noble, W.S. (2009).
Improvements to the percolator algorithm for peptide identification from
shotgun proteomics data sets. J. Proteome Res. 8, 3737–3745.
Sullivan, L.B., Martinez-Garcia, E., Nguyen, H., Mullen, A.R., Dufour, E.,
Sudarshan, S., Licht, J.D., Deberardinis, R.J., and Chandel, N.S. (2013). The
proto-onco metabolite fumarate binds glutathione to amplify ROS-dependent
signaling. Mol. Cell 51, 236–248.
Trujillo, C., Blumenthal, A., Marrero, J., Rhee, K.Y., Schnappinger, D., and Ehrt,
S. (2014). Triosephosphate isomerase is dispensable in vitro yet essential for
Mycobacterium tuberculosis to establish infection. MBio 5, e00085.
10 Cell Chemical Biology 24, 1–10, March 16, 2017
Tseng, C.P., Yu, C.C., Lin, H.H., Chang, C.Y., and Kuo, J.T. (2001). Oxygen-
and growth rate-dependent regulation of Escherichia coli fumarase (FumA,
FumB, and FumC) activity. J. Bacteriol. 183, 461–467.
Venugopal, A., Bryk, R., Shi, S., Rhee, K., Rath, P., Schnappinger, D., Ehrt, S.,
and Nathan, C. (2011). Virulence of Mycobacterium tuberculosis depends on
lipoamide dehydrogenase, a member of three multienzyme complexes. Cell
Host Microbe 9, 21–31.
Wei, J.-R., Krishnamoorthy, V., Murphy, K., Kim, J.-H., Schnappinger, D.,
Alber, T., Sassetti, C.M., Rhee, K.Y., and Rubin, E.J. (2011). Depletion of anti-
biotic targets has widely varying effects on growth. Proc. Natl. Acad. Sci. USA
108, 4176–4181.
Woods, S.A., Schwartzbach, S.D., and Guest, J.R. (1988). Two biochemically
distinct classes of fumarase in Escherichia coli. Biochim. Biophys. Acta
954, 14–26.
Xu, G., Chen, X., Liu, L., and Jiang, L. (2013). Fumaric acid production in
Saccharomyces cerevisiae by simultaneous use of oxidative and reductive
routes. Bioresour. Technol. 148, 91–96.
Zhang, Z., Tan, M., Xie, Z., Dai, L., Chen, Y., and Zhao, Y. (2011). Identification
of lysine succinylation as a new post-translational modification. Nat. Chem.
Biol. 7, 58–63.
Zhang, Y.J., Ioerger, T.R., Huttenhower, C., Long, J.E., Sassetti, C.M.,
Sacchettini, J.C., and Rubin, E.J. (2012). Global assessment of genomic re-
gions required for growth in Mycobacterium tuberculosis. PLoS Pathog. 8,
e1002946.
Zheng, L., Cardaci, S., Jerby, L., MacKenzie, E.D., Sciacovelli, M., Johnson,
T.I., Gaude, E., King, A., Leach, J.D.G., Edrada-Ebel, R., et al. (2015).
Fumarate induces redox-dependent senescence by modifying glutathione
metabolism. Nat. Commun. 6, 6001.

Cell Chemical Biology, Volume 24
Supplemental Information
Fumarase Deficiency Causes Protein and Metabolite
Succination and Intoxicates
Mycobacterium tuberculosis
Nadine Ruecker, Robert Jansen, Carolina Trujillo, Susan Puckett, PradeepaJayachandran, Gerardo G. Piroli, Norma Frizzell, Henrik Molina, Kyu Y.Rhee, and Sabine Ehrt

Supplemental Figures
Figure S1, related to Figure 1. Confirmation of the generation of Fum-DUC. (A) Southern blot design showing the genomic locus of fumC and the same locus after successful replacement of fumC with a hygromycin resistance cassette via homologous recombination. The strain carries an additional copy of fumC with a C-terminal DAS-tag under the control of reverse Tet repressor (T38S38) and expresses SspB under the control of wt TetR (TSC10). (B) Southern blot showing successful replacement of the native fumC gene. Chromosomal DNAs were restricted with PmlI and SacI and hybridized with the probe indicated in A.
WT Fum locus
Fum-DUC Fum locus attL5 and tweety integration sites
glpx fumC Rv1097c Rv1096 phoH2
SacI PmlI probe
4733bp
glpx fumC Rv1097c Rv1096 phoH2
SacI PmlI probe
1209bp
SacI
HygR
fumC-DAS T38S38 sspB ZeoR KanR TSC10
A B
Fum-DUC WT
- 5 kb - 4 kb
- 3 kb
- 2 kb
- 1 kb
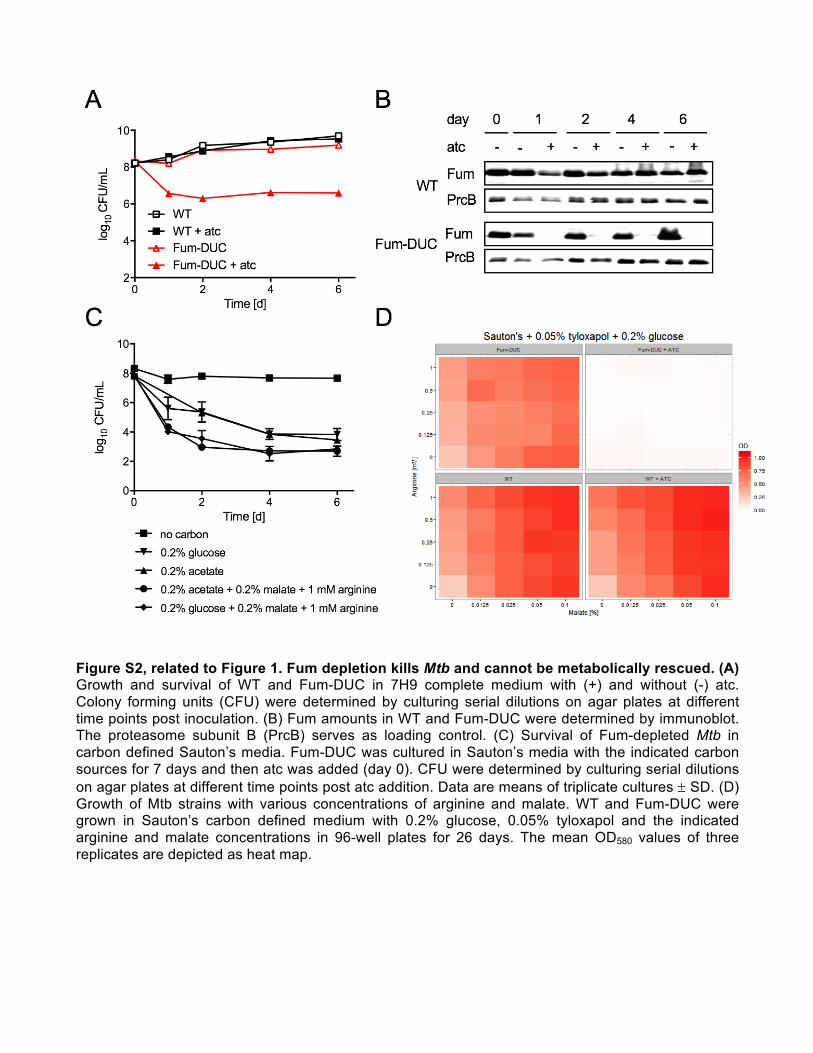
Figure S2, related to Figure 1. Fum depletion kills Mtb and cannot be metabolically rescued. (A) Growth and survival of WT and Fum-DUC in 7H9 complete medium with (+) and without (-) atc. Colony forming units (CFU) were determined by culturing serial dilutions on agar plates at different time points post inoculation. (B) Fum amounts in WT and Fum-DUC were determined by immunoblot. The proteasome subunit B (PrcB) serves as loading control. (C) Survival of Fum-depleted Mtb in carbon defined Sauton’s media. Fum-DUC was cultured in Sauton’s media with the indicated carbon sources for 7 days and then atc was added (day 0). CFU were determined by culturing serial dilutions on agar plates at different time points post atc addition. Data are means of triplicate cultures ± SD. (D) Growth of Mtb strains with various concentrations of arginine and malate. WT and Fum-DUC were grown in Sauton’s carbon defined medium with 0.2% glucose, 0.05% tyloxapol and the indicated arginine and malate concentrations in 96-well plates for 26 days. The mean OD580 values of three replicates are depicted as heat map.

Figure S3, related to Figure 2. Mtb requires Fum to establish and maintain infection in mice. Hematoxylin and eosin stained lung sections were isolated on day 56 and day 112 from mice infected with WT or Fum-DUC. Scale bar, 1 mm. Cellular infiltration was evident on day 56 and increased in magnitude in mouse lungs infected with WT Mtb and in lungs infected with Fum-DUC but not treated with doxy. In contrast, mice infected with Fum-DUC and treated with doxy starting day 10 or day 35 did not show similar cellular infiltrates. These data are consistent with the growth of WT Mtb and Fum-DUC in doxy-free mice and killing of Fum-DUC in doxy-treated mice.

Figure S4, related to Figure 3. (A) Viability of bacteria after growth on filters was determined by CFU. Depicted are mean values of biological triplicates ± SD. (B) Fum levels in bacteria grown on filters were determined by immunoblot. PrcB serves as a loading control.
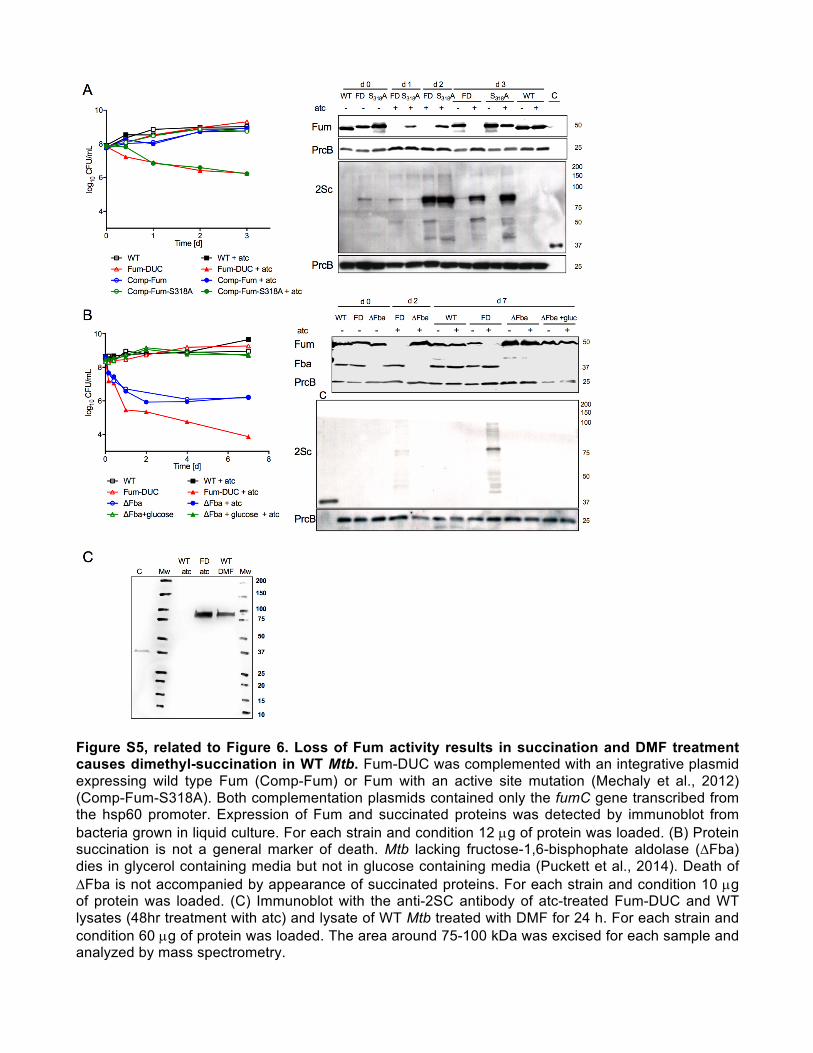
Figure S5, related to Figure 6. Loss of Fum activity results in succination and DMF treatment causes dimethyl-succination in WT Mtb. Fum-DUC was complemented with an integrative plasmid expressing wild type Fum (Comp-Fum) or Fum with an active site mutation (Mechaly et al., 2012) (Comp-Fum-S318A). Both complementation plasmids contained only the fumC gene transcribed from the hsp60 promoter. Expression of Fum and succinated proteins was detected by immunoblot from bacteria grown in liquid culture. For each strain and condition 12 µg of protein was loaded. (B) Protein succination is not a general marker of death. Mtb lacking fructose-1,6-bisphophate aldolase (∆Fba) dies in glycerol containing media but not in glucose containing media (Puckett et al., 2014). Death of ∆Fba is not accompanied by appearance of succinated proteins. For each strain and condition 10 µg of protein was loaded. (C) Immunoblot with the anti-2SC antibody of atc-treated Fum-DUC and WT lysates (48hr treatment with atc) and lysate of WT Mtb treated with DMF for 24 h. For each strain and condition 60 µg of protein was loaded. The area around 75-100 kDa was excised for each sample and analyzed by mass spectrometry.

Supplemental References Mechaly, A.E., Haouz, A., Miras, I., Barilone, N., Weber, P., Shepard, W., Alzari, P.M., and Bellinzoni, M. (2012). Conformational changes upon ligand binding in the essential class II fumarase Rv1098c from Mycobacterium tuberculosis. FEBS Lett 586, 1606–1611.
Puckett, S., Trujillo, C., Eoh, H., Marrero, J., Spencer, J., Jackson, M., Schnappinger, D., Rhee, K., and Ehrt, S. (2014). Inactivation of Fructose-1,6-Bisphosphate Aldolase Prevents Optimal Co-catabolism of Glycolytic and Gluconeogenic Carbon Substrates in Mycobacterium tuberculosis. PLoS Pathog 10, e1004144.