fossil cercopithecidae from the hadar formation and...
TRANSCRIPT

Stephen R. FrostDepartment of Anatomy,New York College ofOsteopathic Medicine, NYITOld Westbury, New York11568, U.S.A. E-mail:[email protected]
Eric DelsonDivision of Paleontology,American Museum ofNatural History New York,NY 10024 and Department ofAnthropology, Lehmen Collegeand the Graduate School,City University of New York,U.S.A. E-mail:[email protected]
Received 10 May 2002Revision received6 September 2002 andaccepted 9 September 2002
Key Words:: Hadar, Ahmado,Geraru, Leadu,Cercopithecidae, Parapapio,Theropithecus,Cercopithecoides,Rhinocolobus, Pliocene,Pleistocene, paleontology.
Fossil Cercopithecidae from the HadarFormation and surrounding areas of theAfar Depression, Ethiopia
Hadar is well known as one of the most productive early hominin sitesin the world. Between 1972 and 1994 a large sample of fossilcercopithecid specimens was collected from Hadar and the nearbysites of Geraru, Ahmado, and Leadu. At least five, and possibly six,species are present in the sample, including two chronological sub-species of Theropithecus oswaldi. T. o. cf. darti is known from theMiddle Pliocene deposits in the Hadar area, along with Parapapiocf. jonesi, cf. Rhinocolobus turkanaensis, and a new species ofCercopithecoides, C. meaveae. There are also isolated molars from theMiddle Pliocene of a large colobine which most likely represent cf.R. turkanaensis, but may also represent another large colobine knownfrom the nearby site of Maka in the Middle Awash. T. o. oswaldiis represented from younger deposits of Late Pliocene and EarlyPleistocene age, along with the large colobine Cercopithecoides kimeui.
Throughout the sequence Theropithecus oswaldi is by far the mostabundant cercopithecid, with the other taxa being comparatively rare.The Parapapio material from Hadar is important as the only securelyidentifiable material of the genus in the East African Pliocene.Furthermore, the Hadar material includes the only associated post-cranial remains for the genus. If the tentative identification ofRhinocolobus is correct, then the Hadar sample is the only knownoccurrence outside of the Turkana Basin. Cercopithecoides meaveae is anew species, currently only known from the Hadar region, mostimportantly by the associated partial skeleton from Leadu. It appearsto show adaptations for terrestrial locomotion. Finally, Cercopithe-coides kimeui, a very large colobine previously known from OlduvaiGorge, Koobi Fora, and Rawi is recorded from the uppermost part ofthe Formation.
� 2002 Elsevier Science Ltd. All rights reserved.
Journal of Human Evolution (2002) 43, 687–748doi:10.1053/jhev.2002.0603Available online at http://www.idealibrary.com on
Introduction
The Hadar Formation of Ethiopia is amongthe most famous of hominin fossil sites,known for the many important finds ofAustralopithecus afarensis as well as a palateof early Homo associated with Oldowanstone tools (Johanson et al., 1982; Kimbelet al., 1994, 1996). A large number ofcercopithecids has been collected from theHadar Formation, as well as from severallocalities in the surrounding area, includ-ing Leadu, Ahmado, and Geraru (Taieb
0047–2484/02/110687+62 $35·00/0
et al., 1976; Szalay & Delson, 1979; Kalbet al., 1982b; Delson, 1984; Kalb, 1993)(Figure 1).
The sample of fossil cercopithecidsincludes 690 specimens spanning a timerange from older than 3·4 Ma through�1·8–1·6 Ma. There are at least fivespecies present, including three colobinesand two cercopithecines. Furthermore,there are two successive chrono-subspeciesof Theropithecus oswaldi. Initial collectionsfrom Hadar, Leadu, Ahmado, and Geraruwere made by the International Afar
� 2002 Elsevier Science Ltd. All rights reserved.
688 .
Figure 1. Location of Hadar, Leadu, Geraru, and Ahmado relative to other paleoanthropologicalcollecting regions in the Afar Depression.
Research Expedition between 1973 and1977; further collections were made atHadar between 1990 and 1994 by theInstitute of Human Origins. Specimenscollected after July, 1999 were not studied.
Taieb et al. (1976) tentatively listedthree cercopithecid species present in theHadar area: Papio sp., Theropithecus sp., andColobus sp. Johanson et al. (1982) includedfour cercopithecid species in their faunal listfor Hadar: Parapapio sp., Theropithecus sp.,Colobinae gen. et sp. indet., and Colobus sp.The only specimens from the Hadar regionthat have been described in detail are thoseof Theropithecus oswaldi cf. darti from theHadar Formation (Eck, 1993). The otherHadar monkeys have been discussed in anumber of reviews by Delson (Szalay &Delson, 1979; Delson, 1984; 1994; Delson
et al., 2000). In addition to Theropithecusdarti (here T. o. cf. darti), he listed Parapapiocf. jonesi, cf. Rhinocolobus turkanaensis,and Colobinae sp. ‘‘A’’. Finally, from theLate Pliocene upper part of the KadaHadar Member, Kimbel et al. (1996)included Theropithecus oswaldi (T. o. oswaldihere) among the species present fromMakaamitalu. The material other thanTheropithecus from Ahmado and Geraru hasnot been previously discussed. A partialskeleton of a medium-sized colobine fromLeadu has been mentioned by severalauthors, but never described in detail (Eck,1976; Szalay & Delson, 1979; Delson,1984; 1994). This paper describes all ofthe material that was available as of 1999.However, because most of the Theropithecusmaterial has been described previously,

689
and/or is currently under analysis by G. G.Eck, it will only be briefly discussed. Inaddition to the partial skeleton from Leadu,there is a sizeable amount of postcranialmaterial, most of which is likely to representTheropithecus oswaldi. Thorough functionalmorphological and ecomorphological analy-sis of this material is beyond the scope of thispaper, but some indices of functional impor-tance are included for descriptive purposes.
Figure 2. Chronostratigraphy of Hadar Formation, Ahmado, Geraru, and Leadu. Dated tuffs are shownby bold lines, and their chronological position is marked on the scale to the right, where the name is given,along with their ages. References for the dates of the tuffs are: BKT-3 (Kimbel et al., 1996); BKT-2(Kimbel et al., 1994); KHT, TT-4 (Walter, 1994); SHT (Walter & Aronson, 1993). The chronologicalposition of Ahmado and Geraru is not well known, and only their approximate age is shown following thestratigraphic relationships given in Kalb (1993). Their length is only meant to indicate that these sites havesome thickness of stratigraphic section.
Stratigraphic context and geologic ageof material
The geology of the Hadar Formation hasbeen described in several publications (e.g.,Taieb, et al. 1976; Aronson & Taieb, 1981;Tiercelin, 1986; Yemane, 1997; Kimbel &Walter, 2000). The Hadar Formation iswell controlled chronologically, with Ar/Ardated tuffs, paleomagnetic correlation, and
biochronology (Walter & Aronson, 1993;Walter, 1994; Kimbel & Walter, 2000). Thechronostratigraphy of the Hadar Formationand surrounding areas is shown in Figure 2.The Hadar Formation ranges in age fromover 3·4 Ma to less than 2·33 Ma, but witha large disconformity between 2·92 and2·33 Ma. The site of Pinnacle, at Hadar, islatest Pliocene to Early Pleistocene in age(G. G. Eck, personal communication 2000).
The Hadar Formation is divided intofour members, from lowest to highest: theBasal, Sidi Hakoma, Denen Dora, and KadaHadar. The upper three members aremarked at their bases by tuffs of the samename. These tephras (along with severalothers and a basalt) have been radio-metrically dated (Figure 2). The KadaHadar Member is divided into two parts bythe large disconformity mentioned above. Inthis paper, reference will be made to the

690 .
upper and lower Kada Hadar Member forthe sediments above and below the discon-formity, in a manner similar to the BurgiMember of the Koobi Fora Formation(Brown & Feibel, 1991).
The sites of Leadu, Geraru, and Ahmado,however, are less well known. Descriptionsof the stratigraphic positions of these threesites, along with some biostratigraphic cor-relations to the Hadar Formation, are givenby Kalb (1993). Leadu is thought to beroughly contemporary with the main part ofthe Hadar Formation (i.e., approximately3·4–2·9 Ma), and Ahmado is roughlyequivalent stratigraphically to the SidiHakoma Member. Geraru on the otherhand is less well placed, but possibly equiva-lent to the Denen Dora or Kada HadarMembers.
Each local collecting area at Hadar orother nearby site is assigned an A.L. (AfarLocality) number, with the initial digit oftenindicating the year of collection; specimensare then assigned sequential numbers withinthat locality, and individual parts of associ-ated specimens usually given letters. Thus,the mandible of ‘‘Lucy’’ is A.L.288–1i andwas collected in the third season.
In addition to the colobine partial skel-eton mentioned above, the site of Leadu(A.L.2) has produced an isolated lower rightmolar of Theropithecus and a few cranialfragments. The lower molar is similar in sizeto lower molars of T. o. cf. darti from themain part of the Hadar Formation.
Ahmado (A.L.100) has produced a largebut fragmentary collection of cercopithecidfossils, 104 individual specimens in total, ofwhich 68 are isolated teeth. There are twosmall mandibular fragments, and a frontalfragment, but most of the remaining 33fossils from Ahmado are highly fragmentarypostcranial elements, generally the denseportions of long bones. Thus, the site mayrepresent a higher energy depositionalenvironment than that of the main HadarFormation. Of those specimens diagnos-
able below the family level, 50 representTheropithecus, five are papionins other thanTheropithecus (most likely the Parapapiospecies that is present in the HadarFormation), and a single specimen is from alarge colobine similar in size to Rhinocolobus.The Theropithecus teeth from Ahmado aresimilar in size to the T. o. cf. darti materialfrom Hadar, but slightly smaller. They aresignificantly smaller than teeth of T. brumpti,T. o. oswaldi or T. o. leakeyi. Their size,along with the absence of any specimensunambiguously assignable to T. brumptifrom the Afar region, make the AhmadoTheropithecus likely to be T. o. cf. darti.
There are only three cercopithecid fossilsfrom Geraru (locs. A.L.18, A.L.74, andA.L.99). These are a left mandibular frag-ment with M3 assignable to Theropithecus, afemale mandibular fragment with damagedteeth, and an isolated upper male canine.The latter two specimens are also probablyTheropithecus, but they cannot be securelyallocated. The M3 of the first specimen isrelatively large in comparison with thesample from the main Hadar Formation.This specimen, therefore, may representeither an unusually large individual of T. o.cf. darti, or possibly T. o. oswaldi, and thusat least some areas of Geraru would mostlikely be younger than the main part of theHadar Formation, given that tooth sizein Theropithecus oswaldi appears stronglycorrelated with age (Figure 16; Eck, 1993).
Measurements
Standard caliper measurements wererecorded on all dental specimens that suf-ficiently preserved the relevant morphology.For incisors and canines these measure-ments are the maximum labio-lingual crownbreadth (‘‘W’’ in Tables 1–6); maximummesio-distal length (‘‘L’’ in Tables 1–6) andcrown height as measured labially from thecervix to the occlusal tip (‘‘O’’ in Tables1–6). For all premolars except the P3, only
691
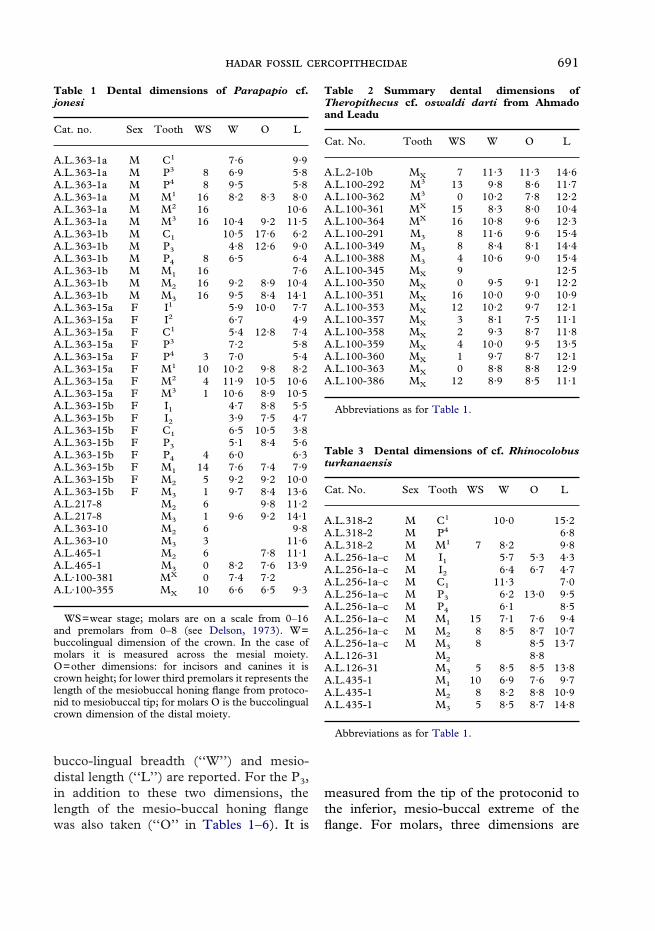
Table 1 Dental dimensions of Parapapio cf.jonesi
Cat. no. Sex Tooth WS W O L
A.L.363-1a M C1 7·6 9·9A.L.363-1a M P3 8 6·9 5·8A.L.363-1a M P4 8 9·5 5·8A.L.363-1a M M1 16 8·2 8·3 8·0A.L.363-1a M M2 16 10·6A.L.363-1a M M3 16 10·4 9·2 11·5A.L.363-1b M C1 10·5 17·6 6·2A.L.363-1b M P3 4·8 12·6 9·0A.L.363-1b M P4 8 6·5 6·4A.L.363-1b M M1 16 7·6A.L.363-1b M M2 16 9·2 8·9 10·4A.L.363-1b M M3 16 9·5 8·4 14·1A.L.363-15a F I1 5·9 10·0 7·7A.L.363-15a F I2 6·7 4·9A.L.363-15a F C1 5·4 12·8 7·4A.L.363-15a F P3 7·2 5·8A.L.363-15a F P4 3 7·0 5·4A.L.363-15a F M1 10 10·2 9·8 8·2A.L.363-15a F M2 4 11·9 10·5 10·6A.L.363-15a F M3 1 10·6 8·9 10·5A.L.363-15b F I1 4·7 8·8 5·5A.L.363-15b F I2 3·9 7·5 4·7A.L.363-15b F C1 6·5 10·5 3·8A.L.363-15b F P3 5·1 8·4 5·6A.L.363-15b F P4 4 6·0 6·3A.L.363-15b F M1 14 7·6 7·4 7·9A.L.363-15b F M2 5 9·2 9·2 10·0A.L.363-15b F M3 1 9·7 8·4 13·6A.L.217-8 M2 6 9·8 11·2A.L.217-8 M3 1 9·6 9·2 14·1A.L.363-10 M2 6 9·8A.L.363-10 M3 3 11·6A.L.465-1 M2 6 7·8 11·1A.L.465-1 M3 0 8·2 7·6 13·9A.L·100-381 MX 0 7·4 7·2A.L·100-355 MX 10 6·6 6·5 9·3
WS=wear stage; molars are on a scale from 0–16and premolars from 0–8 (see Delson, 1973). W=buccolingual dimension of the crown. In the case ofmolars it is measured across the mesial moiety.O=other dimensions: for incisors and canines it iscrown height; for lower third premolars it represents thelength of the mesiobuccal honing flange from protoco-nid to mesiobuccal tip; for molars O is the buccolingualcrown dimension of the distal moiety.
Table 2 Summary dental dimensions ofTheropithecus cf. oswaldi darti from Ahmadoand Leadu
Cat. No. Tooth WS W O L
A.L.2-10b MX 7 11·3 11·3 14·6A.L.100-292 M3 13 9·8 8·6 11·7A.L.100-362 M3 0 10·2 7·8 12·2A.L.100-361 MX 15 8·3 8·0 10·4A.L.100-364 MX 16 10·8 9·6 12·3A.L.100-291 M3 8 11·6 9·6 15·4A.L.100-349 M3 8 8·4 8·1 14·4A.L.100-388 M3 4 10·6 9·0 15·4A.L.100-345 MX 9 12·5A.L.100-350 MX 0 9·5 9·1 12·2A.L.100-351 MX 16 10·0 9·0 10·9A.L.100-353 MX 12 10·2 9·7 12·1A.L.100-357 MX 3 8·1 7·5 11·1A.L.100-358 MX 2 9·3 8·7 11·8A.L.100-359 MX 4 10·0 9·5 13·5A.L.100-360 MX 1 9·7 8·7 12·1A.L.100-363 MX 0 8·8 8·8 12·9A.L.100-386 MX 12 8·9 8·5 11·1
Abbreviations as for Table 1.
Table 3 Dental dimensions of cf. Rhinocolobusturkanaensis
Cat. No. Sex Tooth WS W O L
A.L.318-2 M C1 10·0 15·2A.L.318-2 M P4 6·8A.L.318-2 M M1 7 8·2 9·8A.L.256-1a–c M I1 5·7 5·3 4·3A.L.256-1a–c M I2 6·4 6·7 4·7A.L.256-1a–c M C1 11·3 7·0A.L.256-1a–c M P3 6·2 13·0 9·5A.L.256-1a–c M P4 6·1 8·5A.L.256-1a–c M M1 15 7·1 7·6 9·4A.L.256-1a–c M M2 8 8·5 8·7 10·7A.L.256-1a–c M M3 8 8·5 13·7A.L.126-31 M2 8·8A.L.126-31 M3 5 8·5 8·5 13·8A.L.435-1 M1 10 6·9 7·6 9·7A.L.435-1 M2 8 8·2 8·8 10·9A.L.435-1 M3 5 8·5 8·7 14·8
Abbreviations as for Table 1.
bucco-lingual breadth (‘‘W’’) and mesio-distal length (‘‘L’’) are reported. For the P3,in addition to these two dimensions, thelength of the mesio-buccal honing flangewas also taken (‘‘O’’ in Tables 1–6). It is
measured from the tip of the protoconid tothe inferior, mesio-buccal extreme of theflange. For molars, three dimensions are

692 .
reported: the bucco-lingual crown breadthtaken across the mesial loph(id) (‘‘W’’ inTables 1–6); the bucco-lingual crownbreadth taken across the distal loph(id)(‘‘O’’ in Tables 1–6); and mesiodistal crownlength (‘‘L’’ in Tables 1–6).
Dimensions of some postcranial elementsare also reported in Table 8. For humeri,maximum mediolateral and anteroposteriordiameters are given for the proximalend. Distally, the biepicondylar breadth isreported, and is equivalent to the maxi-mum mediolateral dimension of the distalhumerus (see Figure 10, dimension B). Asecond mediolateral dimension records thetotal width of the distal humerus from thelateral extreme of the lateral epicondyle tothe medial limit of the trochlea (see Figure10, dimension A; following Harrison, 1989).This dimension effectively records the totalhumeral width sans the medial epicondyle.Distal articular breadth is the mediolateraldimension across both the capitulum andtrochlea, and is taken anteriorly (see Figure12, dimension B). The length of the medialtrochlear flange is taken from the mostproximal point on the trochlea to the distaltip of the flange (see Figure 12, dimensionA). Femoral measurements reported are theanteroposterior diameter of the femoralhead (see Figure 15, dimension B), themaximum mediolateral breadth of the femurfrom the trochanteric tuberosity to themedial end of the head, and finally theprojection of the greater trochanter proximalto the femoral neck (see Figure 15, dimen-sion A; following Ting, 2001). On the distalend, both the condylar anteroposterior
depth and maximum mediolateral breadthare given. The ulna, radius, and tibia arerepresented by a single specimen for the taxaincluded here (all from the Leadu colobine),and the measurements included in the tableare only those available on these specimens.On the ulna, the height of the olecranonprocess is measured from the proximal tip ofthe trochlear notch to the proximal tip of theolecranon process. Also recorded are theproximodistal length of the trochlear notch,and the total ulnar breadth measured acrossthe radial and trochlear articular facets. Onthe radius, the maximum diameter of thehead and its diameter perpendicular to themaximum are recorded. The neck length ismeasured in two ways: first from the distallimit of the radial head to the proximal limitof the radial tuberosity (see Harrison, 1989);second is the mechanical length from thecapitular articular facet to the midpoint ofthe radial tuberosity (see Jolly, 1972). Onthe tibia, only the anteroposterior depth ofthe tibial condyles and the mediolateralbreadth across both condyles are recorded.
Table 4 Dental dimensions of Cercopithecoideskimeui
Cat. No. Sex Tooth WS W O L
A.L.603-1a F M2 8 11·3 10·4 11·2A.L.603-1a F M3 5 10·4 8·8 11·7
Abbreviations as for Table 1.
Systematic paleontology
The specimens discussed in this paperinclude only those from the HadarFormation, Leadu, Ahmado, and Geraruthat were available as of 1999. All of thesespecimens are housed in the NationalMuseum of Ethiopia, but the NME acro-nym is not used in the text. Three of thesamples described below are only tentativelyreferred to named taxa. Emended diagnosesare provided for these named taxa to facili-tate comparison and to make explicit thereasons for the tentative allocation. Thereferred material from Hadar is thendescribed in detail.
Several types of data are provided for eachspecies or subspecies described. In theby-taxon lists of ‘‘Specimens included’’,specimen numbers preceded by a questionmark are only tentatively allocated due to an

693
Catalog of elements of the adult male partial skeleton of Cercopithecoidesmeaveae from Leadu
Cat. no. Side Element Notes
A.L.2-27b Right Mandible Ramus fragmentA.L.2-28 Right Femur Distal fragment 1/3 shaftA.L.2-30 Bone FragmentA.L.2-34 Face Complete dentitionA.L.2-34 Mandible Complete dentitionA.L.2-35 Endocranial castA.L.2-36 Phalanx Distal fragmentA.L.2-37 Right Ulna Distal fragmentA.L.2-38 Right Fibula Distal fragmentA.L.2-39 Left Fibula Proximal fragmentA.L.2-40 Metacarpal V Proximal fragmentA.L.2-41 Metatarsal Distal fragmentA.L.2-42 Metatarsal Distal fragmentA.L.2-44 Left NavicularA.L.2-44 Left CuboidA.L.2-45 Thoracic vertebra BodyA.L.2-45 Caudal vertebraA.L.2-46 Lumbar vertebraA.L.2-47 Caudal vertebraA.L.2-48 Caudal vertebraA.L.2-51 Lumbar vertebra BodyA.L.2-52 Caudal vertebraA.L.2-53 Caudal vertebraA.L.2-54 Caudal vertebra Proximal fragmentA.L.2-55 Caudal vertebra Distal fragmentA.L.2-56 Caudal vertebra Proximal fragmentA.L.2-57 Caudal vertebra Distal fragmentA.L.2-58 Caudal vertebra Proximal fragmentA.L.2-60 Lumbar vertebra Spinous processA.L.2-61 Bone FragmentA.L.2-62 Left Scapula GlenoidA.L.2-63 Left Humerus Proximal fragmentA.L.2-64 Left Humerus Distal fragmentA.L.2-65 Left Ulna Proximal fragmentA.L.2-66 Left Radius Proximal fragmentA.L.2-67 Left Radius Distal fragmentA.L.2-68 Right Ischium TuberosityA.L.2-69 Left Ischium TuberosityA.L.2-70 Right Femur Head, neck, and lesser trochanterA.L.2-71 Left Innominate AcetabulumA.L.2-72 Left Femur Proximal fragmentA.L.2-73 Left Femur Shaft fragmentA.L.2-74 Left Femur Distal fragmentA.L.2-75 Left PatellaA.L.2-76 Right PatellaA.L.2-77 Left Tibia Proximal fragmentA.L.2-78 Left Tibia Distal fragmentA.L.2-79 Postcranial Shaft fragmentA.L.2-80 Right IliumA.L.2-80 Bone FragmentsA.L.2-81 Postcranial MidshaftA.L.2-83 Caudal vertebraA.L.2-84 Postcranial Shaft fragmentA.L.2-86 Frontal Glabellar regionA.L.2-87 Left Zygomatic (jugal)A.L.2-88 Right Frontal Orbitotemporal rimA.L.2-117 Right Tibia Proximal fragmentA.L.2-119 Mesial phalanxA.L.2-122 Metatarsal II Proximal fragmentA.L.2-? Caudal vertebra Distal fragment
Table 5
694 .
1 Parapapio is best diagnosed in the face, as no facialmaterial of Parapapio ado is complete enough to observethe diagnostic features of the genus, it is only tentativelyincluded in Parapapio.
Table 6 Dental dimensions of Cercopithecoidesmeaveae
Cat. No. Sex Tooth WS W O L
A.L.249-23 P3 5·5A.L.660-2 M1 8 7·5 7·3 8·0A.L.660-2 M2 6 7·5 9·2A.L.660-2 M3 2 7·4 7·5 9·5A.L.2-34 M I1 4·9 5·9 4·9A.L.2-34 M I2 5·5 6·3 4·7A.L.2-34 M C1 7·0 25·0 12·0A.L.2-34 M P3 2 6·8 4·4 4·9A.L.2-34 M P4 2 7·6 5·2 5·6A.L.2-34 M M1 8 7·9 7·4 8·8A.L.2-34 M M2 5 8·6 7·8 8·6A.L.2-34 M M3 1 8·8 7·6 9·2A.L.2-34 M I1 4·7 3·9 3·5A.L.2-34 M I2 5·1 5·0 3·5A.L.2-34 M C1 8·5 5·0A.L.2-34 M P3 4·6 11·6 8·5A.L.2-34 M P4 7 4·9 8·0A.L.2-34 M M1 8 6·5 6·8 8·3A.L.2-34 M M2 6 7·4 7·5 8·6A.L.2-34 M M3 2 7·8 7·9 12·2A.L.231-1a M I2 4·2 5·8 3·2A.L.231-1a M C1 8·2 4·6A.L.231-1a M P3 4·3 12·9 7·2A.L.231-1a M P4 4·9 6·6A.L.231-1a M M1 12 6·6 6·6 8·8A.L.231-1a M M2 8 7·0 7·4 9·2A.L.231-1a M M3 4 7·0 7·3 10·5
Abbreviations as for Table 1.
absence of diagnostic morphology, e.g., forisolated teeth and unassociated postcranialmaterial; those preceded by (cf.) representmaterial which is tentatively assignedbecause the morphology is ambiguous,being different from the typical material, butnot necessarily distinct enough to warranttaxonomic separation. The geographic dis-tribution of each species or subspecies isalso given. Underlined localities indicatethose included in this paper. Localities pre-ceded by a question mark yield specimensthat are too incomplete to provide a defini-tive diagnosis; those preceded by (cf.) yieldrelatively complete specimens which differslightly in their morphology from or areotherwise difficult to allocate to the namedtaxon.
Family Cercopithecidae Gray, 1821Subfamily Cercopithecinae Gray, 1821Tribe Papionini Burnett, 1828Genus Parapapio Jones, 1937(=or including: Papio Erxleben, 1777:Houghton, 1925; Gear, 1926, in part;Leakey & Leakey, 1976, in part. CercocebusGeoffroy, 1812: Hopwood, 1936, inpart. Papio (Simopithecus) (Andrews,1916): Dietrich, 1942. BrachygnathopithecusKitching, 1952, in part. Papio (Parapapio):Delson, 1975. Papionini gen. et. sp. indet.B. Leakey & Leakey, 1976.)Type species: Parapapio broomi Jones, 1937Other included species: Pp. antiquus
(Haughton, 1925); Pp. ado (Hopwood,1936)1; Pp. jonesi Broom, 1940; Pp. whiteiBroom, 1940; Pp. species nova Leakeyet al., in press.
Generic diagnosisThis diagnosis largely follows those ofFreedman (1957), Eisenhart (1974), andSzalay & Delson (1979). Parapapio is anextinct genus of medium sized Africanpapionins, thought to be conservativerelative to other known African papioningenera. When viewed in lateral profile,the cranium of Parapapio is distinguishedfrom those of Papio, Pliopapio, Theropithecus,Mandrillus, Gorgopithecus, Paradolichopithe-cus and some Macaca by the lack of ananteorbital drop, a relatively thin browridge, and lack of an ophryonic groove.It is also different from Papio (Papio),Gorgopithecus, Lophocebus, Cercocebus andMandrillus in that it generally lacks post-canine and suborbital fossae, though shallowfossae are present in some individuals,especially in Pp. jonesi and Pp. antiquus.Maxillary ridges in the males are also absentor only poorly developed, unlike Papio(Papio), Theropithecus (Omopithecus), and

695
Mandrillus. The mandible is distinctfrom those of Papio (Papio), Mandrillus,Lophocebus, Theropithecus (Omopithecus),Gorgopithecus and most Macaca in its lack ofcorpus fossae. The dentition is indistin-guishable from that of Papio. The limitedpostcranial evidence from Hadar appearsto suggest more adaptations for arborealpositional behaviors than in Theropithecus orextant Papio.
Parapapio jonesi Broom, 1940Holotype: TMP STS 565, fromSterkfontein Type Site Member 4.Distribution: Sterkfontein Mbr. 4 (type
locality); Makapansgat Mbs. 2–4. Nospecimens from the Hadar sample aredefinitively included in this taxon (seebelow).
Specific diagnosisThe different species of Parapapio havenot been well diagnosed, particularly thethree that are generally recognized atMakapansgat and Sterkfontein: Pp. broomi,Pp. jonesi and Pp. whitei. Freedman (1957)essentially divided them into dental sizecategories with Pp. jonesi the smallest, andPp. whitei the largest. This diagnosis followsthose of Maier (1970), Eisenhart (1974) andSzalay & Delson (1979).
Pp. jonesi is a medium sized papioninsimilar to Macaca thibetana in size and levelof sexual dimorphism (Delson et al., 2000).In molar and cranial size it is significantlysmaller than Pp. whitei, most subspecies ofPapio hamadryas, and Theropithecus oswaldidarti. Maxillary fossae are generally betterdeveloped than in other species of the genus,as are the maxillary ridges. These two fea-tures yield a muzzle dorsum that is moresquared in cross-section than that of Pp.broomi, but similar to Pp. whitei. The ros-trum is relatively tall and deep, and shorterin comparison to the neurocranium thanthat of Pp. whitei. The premaxillae projectfurther anteriorly beyond the canine than
do those of Pp. broomi. This produces anincisive arc that is more rounded in Pp. jonesithan in Pp. broomi, which tends to havea more flattened incisive arc. The M3 isnot reduced distally, distinguishing it fromPp. antiquus.
Parapapio cf. jonesi Broom, 1940(=or including Parapapio cf. jonesi: Szalay &Delson, 1979)Specimens included: NME ?A.L.217–8,
A.L.363–1a-l, 10, ?12, 15a-b, ?A.L.465–1, ?A.L.100–348, 354, 355, 365, 381.
Distribution: Hadar Fm. Sidi Hakoma andKada Hadar Mbs.; ?Ahmado; ?Maka, Fm.‘‘W’’ sub-Sidi Hakoma Tuff.
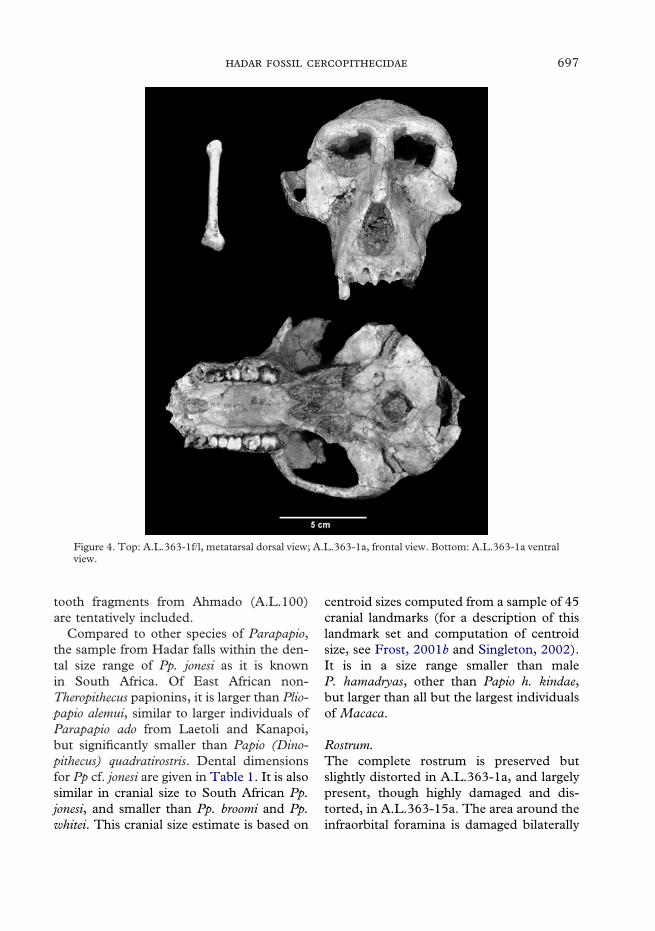
DescriptionThe majority of specimens and all of thediagnostic material of this taxon comes froma single locality, A.L.363, in Unit 3u-s in theuppermost part of the Kada Hadar Memberof the Hadar Formation below the discon-formity. The best specimen is a nearly com-plete male skull with a possibly associatedpartial skeleton, A.L.363–1a-l (Figures 3–6;9; 13). The cranium is nearly complete:most of the right side, including the zygo-matic arch is preserved, but the left zygo-matic arch, temporal squama, occipital, andinferior parietal, as well as the region aroundinion are absent. All of the cheek teeth arepresent, but they are so highly worn as topreserve almost no crown morphology, indi-cating that this was an older adult individ-ual. The right canine is also preserved, but isheavily damaged; the alveoli for the otherteeth are present. The mandible is wellpreserved on the left side (lacking onlythe coronoid process), but the right side isbroken behind the M2; the incisor alveoli areempty, and the right P3 and the left M1 areseverely damaged.
There is also a partly distorted femaleface and mandibular corpus, A.L.363–15(Figures 6 and 7), with nearly completedentition, lacking only the upper right lateral

696 .
Figure 3. Male Parapapio cf. jonesi skull A.L.363-1a. Dorsal view above. Right lateral view below.
incisor, upper left canine and lower rightcentral incisor. The surface bone of thewhole specimen is expanded and heavilycracked. The right side is better preserved,however, with the orbital rim, and pieces ofthe zygomatic arch. The right temporal hasalso been approximately joined although itdoes not fully contact. The orbit andzygoma are totally lacking on the left side.Other than the right temporal, the neuro-cranium is lacking. The palate is completelypreserved.
From the same locality there is also a leftmandibular corpus fragment with M2–3
(A.L.363-10), and the distal half of a
right humerus (A.L.363-12) is tentativelyassigned to this taxon (Delson, 1984;Delson et al., 2000). Two additional speci-mens are included from other Hadar locali-ties, but they are less securely placed inthis taxon. They are diagnosed by beingpapionins other than Theropithecus of thesame dental size as the A.L.363 crania, anda lack of any contradictory morphologicalevidence. A.L.217-8 (from SH 1-2-3s) is aright mandibular corpus with M2–3, andA.L.465-1 (from SH 1-s) is a left corpus,also with M2–3. Both of these specimens lackthe inferior margin and preserve little of thecorpus depth. Finally, five isolated teeth and
697
tooth fragments from Ahmado (A.L.100)are tentatively included.
Compared to other species of Parapapio,the sample from Hadar falls within the den-tal size range of Pp. jonesi as it is knownin South Africa. Of East African non-Theropithecus papionins, it is larger than Plio-papio alemui, similar to larger individuals ofParapapio ado from Laetoli and Kanapoi,but significantly smaller than Papio (Dino-pithecus) quadratirostris. Dental dimensionsfor Pp cf. jonesi are given in Table 1. It is alsosimilar in cranial size to South African Pp.jonesi, and smaller than Pp. broomi and Pp.whitei. This cranial size estimate is based on
centroid sizes computed from a sample of 45cranial landmarks (for a description of thislandmark set and computation of centroidsize, see Frost, 2001b and Singleton, 2002).It is in a size range smaller than maleP. hamadryas, other than Papio h. kindae,but larger than all but the largest individualsof Macaca.
Figure 4. Top: A.L.363-1f/l, metatarsal dorsal view; A.L.363-1a, frontal view. Bottom: A.L.363-1a ventralview.
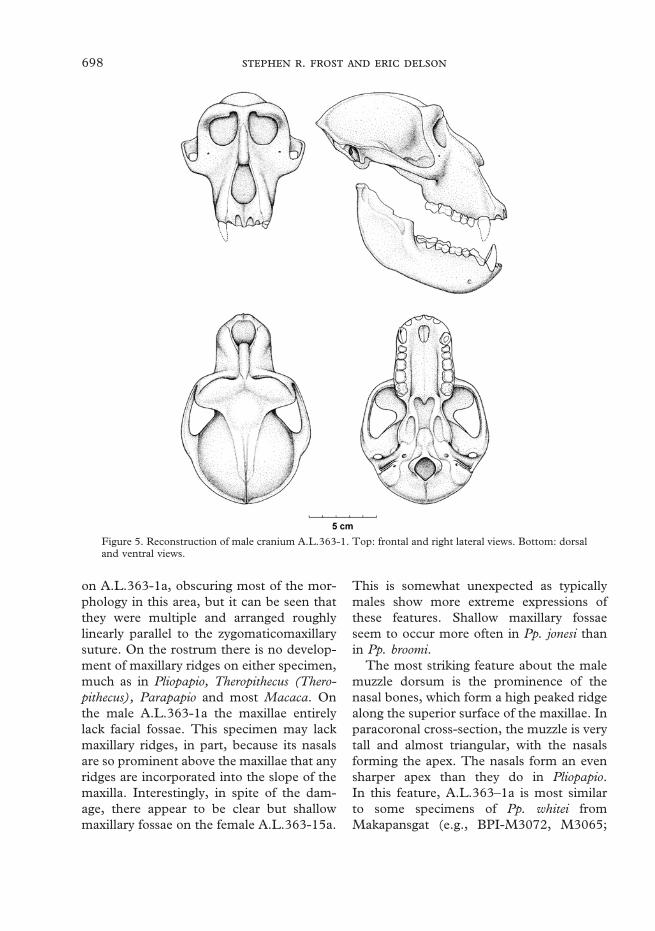
Rostrum.The complete rostrum is preserved butslightly distorted in A.L.363-1a, and largelypresent, though highly damaged and dis-torted, in A.L.363-15a. The area around theinfraorbital foramina is damaged bilaterally
698 .
Figure 5. Reconstruction of male cranium A.L.363-1. Top: frontal and right lateral views. Bottom: dorsaland ventral views.
on A.L.363-1a, obscuring most of the mor-phology in this area, but it can be seen thatthey were multiple and arranged roughlylinearly parallel to the zygomaticomaxillarysuture. On the rostrum there is no develop-ment of maxillary ridges on either specimen,much as in Pliopapio, Theropithecus (Thero-pithecus), Parapapio and most Macaca. Onthe male A.L.363-1a the maxillae entirelylack facial fossae. This specimen may lackmaxillary ridges, in part, because its nasalsare so prominent above the maxillae that anyridges are incorporated into the slope of themaxilla. Interestingly, in spite of the dam-age, there appear to be clear but shallowmaxillary fossae on the female A.L.363-15a.
This is somewhat unexpected as typicallymales show more extreme expressions ofthese features. Shallow maxillary fossaeseem to occur more often in Pp. jonesi thanin Pp. broomi.
The most striking feature about the malemuzzle dorsum is the prominence of thenasal bones, which form a high peaked ridgealong the superior surface of the maxillae. Inparacoronal cross-section, the muzzle is verytall and almost triangular, with the nasalsforming the apex. The nasals form an evensharper apex than they do in Pliopapio.In this feature, A.L.363–1a is most similarto some specimens of Pp. whitei fromMakapansgat (e.g., BPI-M3072, M3065;
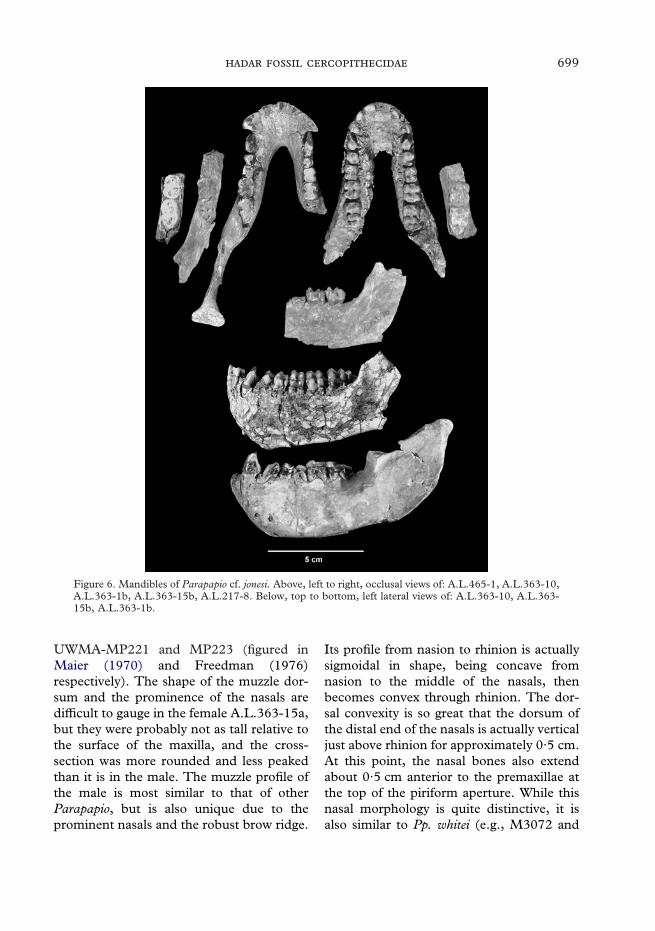
699
Figure 6. Mandibles of Parapapio cf. jonesi. Above, left to right, occlusal views of: A.L.465-1, A.L.363-10,A.L.363-1b, A.L.363-15b, A.L.217-8. Below, top to bottom, left lateral views of: A.L.363-10, A.L.363-15b, A.L.363-1b.
UWMA-MP221 and MP223 (figured inMaier (1970) and Freedman (1976)respectively). The shape of the muzzle dor-sum and the prominence of the nasals aredifficult to gauge in the female A.L.363-15a,but they were probably not as tall relative tothe surface of the maxilla, and the cross-section was more rounded and less peakedthan it is in the male. The muzzle profile ofthe male is most similar to that of otherParapapio, but is also unique due to theprominent nasals and the robust brow ridge.
Its profile from nasion to rhinion is actuallysigmoidal in shape, being concave fromnasion to the middle of the nasals, thenbecomes convex through rhinion. The dor-sal convexity is so great that the dorsum ofthe distal end of the nasals is actually verticaljust above rhinion for approximately 0·5 cm.At this point, the nasal bones also extendabout 0·5 cm anterior to the premaxillae atthe top of the piriform aperture. While thisnasal morphology is quite distinctive, it isalso similar to Pp. whitei (e.g., M3072 and

700 .
Figure 7. Female Parapapio cf. jonesi face A.L.363-15a. Frontal and right lateral above dorsal and ventralviews.
MP221) which possesses a less extreme ver-sion of the sigmoidal profile. Although thereis extensive damage to the nasals, the femalespecimen clearly lacks anteorbital drop andhas a profile that is typical of Parapapio,being relatively linear from nasion toprosthion.
In the male specimen, the premaxillo-maxillary suture follows the lateral rim of thepiriform aperture at a margin of less than2 mm before curving laterally anterior to thecanine. Unlike that of T. gelada it neverenters the piriform aperture. The nasal pro-cess of the premaxilla projects posteriorly toapproximately the midpoint of the nasalsbefore it is covered by the maxilla. Thepremaxillomaxillary suture is complexlycurved in lateral view. Initially it arcsinferiorly following the curvature of the
nasals, but then becomes concave-up alongthe lateral margin of the piriform aperturebefore curving inferiorly again anterior tothe canine root. Once again, for the femalemost of the morphology is obscured, but thepremaxillomaxillary suture is somewhatpreserved on the left side. It appears that itwas considerably straighter in its course thanthat of the male. The premaxillae projectrelatively far anteriorly beyond the canine,and there is a modest diastema separatingthe canine from the incisors. In thesefeatures, A.L.363-1a is similar to Pp.whitei (BPI-M3065, M3072, UWMA-MP221, and MP223) and Pp jonesi (TMPSTS 565, holotype). Known specimens ofPp. broomi seem to lack this area, except fora large male from Bolt’s Farm, UWMA BF43, which is only tentatively assigned to this

701
species [and was designated Pp. whitei byFreedman (1965)]. This specimen has pre-maxillae that do not project nearly as farbeyond the canine, forming a straighter linebetween the canines.
The piriform aperture of the maleA.L.363-1a is basically oval in outline, as arethose of most papionins, but its inferior limitis not ‘‘V’’-shaped, instead being morerounded. The unique nasal morphology ofthis specimen gives the superior part of theaperture a somewhat distinctive shape. Thenasals form a semicircular arch that projectsperpendicularly over the superior limit of thepiriform aperture. While the breadth acrossboth nasals is only about 1 cm, rhinionprojects approximately 0·5 cm above andanterior to the lateral edge of the nasals. Thewidest portion of the piriform aperture isinferior to midheight. The piriform apertureof the female is heavily damaged and dis-torted, but what is preserved appears to besimilar to the male, except for the areaaround the nasals. Viewed laterally, theplane defined by the rim of the piriformaperture is concave-up, and inclined at anangle of approximately 40� to the occlusalplane, in both the male A.L.363-1a and thefemale A.L.363-15a.
The maxillary dental arcade is typical ofmost papionins. The male arcade is ‘‘U’’-shaped in outline, whereas that of thefemale, as far as can be determined, is moreparabolic. The postcanine tooth rows of themale are fairly straight and parallel. The M2
is the most laterally positioned tooth, so thatthere is a slight arc to the tooth row. Thecanines are positioned laterally relative tothe other teeth, particularly the P3. Theincisors form a smoothly bowing archbetween the canines, projecting moreanteriorly than they do in Theropithecus.There is also a short diastema between thecanine and I2. The female maxillary dentalarcade is not as well preserved, but allowingfor this it appears to be similar to that of themale, except that the canines do not project
laterally beyond the P3 and the cheek toothrows converge more anteriorly.
In lateral view, the occlusal surface of thedentition is rather straight. The palate issimilar to those of other papionins, beingrectangular in outline in both the male andfemale. The palate is deep and deepensposteriorly in the male, reaching a maximumdepth of approximately 13 mm. It is deep,but of more constant depth in the female.The absolute depth of the female specimenis difficult to judge due to distortion, but issimilar to that of the male. The alveolarprocesses of the male are nearly perpendicu-lar to the main floor of the palate. Those ofthe female are far more sloping, but theiroriginal shape is difficult to determine.Overall, the palate shape is rather unlike thatof most Parapapio, which tends to bebroader and comparatively shallow. Forinstance, even in the large male Pp. whiteiUWMA MP223 from Makapansgat, thepalate reaches a maximum depth of only10 mm, even though it is a substantiallylarger specimen than A.L.363-1a.
Zygomatic arch.The anterior surface of the zygomatic pro-cess of the maxilla arises superior to themesial part of the M3 in the male. Itsposition in the female is difficult to assessdue to damage and distortion, but it isprobably somewhere above the distal M2 ormesial M3. The anterior surface of thezygoma curve smoothly posteriorly inboth the male and female specimens. Theanterior surface in the male A.L.363-1a is smooth, lacking suborbital fossaeentirely, like Pp. whitei and Pp. broomi fromMakapansgat and Sterkfontein, but unlikePp jonesi from Makapansgat (e.g., M3051,M3054). This pattern is also similar tothe anterior surface in T. oswaldi, Papio(Dinopithecus), Parapapio and Mandrillus,and may be primitive for African papionins.The region of the infraorbital foramina isdamaged bilaterally. The female A.L.363-15a

702 .
preserves slight suborbital fossae (there is achance this is due to distortion, however).The inferior edge of the zygoma archessharply laterally away from the maxilla. Thealveolar process extends fairly far below thezygomatic root, i.e. the maxilla is compara-tively deep in the male, but less so in thefemale.
In superior view, the zygomatic arches ofboth specimens curve sharply posteriorly,and are not widely flaring. The anteriorportion does not jut out laterally as it does inPapio (Papio), Cercocebus, Theropithecus, andLophocebus, but slopes more posteriorly as inT. oswaldi, P. (Dinopithecus), and Mandrillus.In this morphology, it is the same as otherknown specimens of Parapapio. In bothA.L.363-1a and A.L.363-15a, the mostlateral portion of the zygomatic arch is in itsposterior portion near to where it attaches tothe neurocranium. In overall morphologythe zygomatic arch is thin and lightly built.
Orbital region.Both orbits are well preserved in A.L.363-1a, and the right orbit is present but dis-torted in A.L.363-15a. The supraorbitaltorus of the male is one of the most robustknown for such a small cercopithecid. It isconsiderably thicker than in all known SouthAfrican Parapapio. In superoinferior thick-ness it approaches 12 mm, whereas thethickest brow ridge for any South AfricanParapapio is that of UWMA BF 43 which isapproximately 7 mm. Most other specimenshave brow ridges that are considerably moregracile than this. In frontal view, thesupraorbital torus of A.L.363-1a forms sep-arate arches over each orbit, so that it islowest in the sagittal plane, then reachesmaximum height over the midpoints of theorbits. This is in part due to bulging over themidpoints of the orbits as in robust speci-mens of Papio, T. oswaldi, and Mandrillus.The supraorbital notches are distinct andcut deeply into the torus. The supraorbitalrim of the female is considerably more
gracile, but shows a similar morphology.The supraorbital rim is approximately 7 mmin thickness and has distinct supraorbitalnotches. This is thicker than other knownfemale Parapapio specimens, which are gen-erally less than 5 mm and lack the deepsupraorbital notches.
The interorbital region is narrow in bothspecimens. Glabella is not prominent ineither cranium, so that nasion is the mostanterior point on the midline of the frontal.Like other known Parapapio, the profile inthis region clearly lacks anteorbital drop.The lacrimal fossa lies within the orbit onA.L.363-1a, and its anterior border seems tobe at the lacrimal–maxilla suture. Due todamage, its position on A.L.363-15a is notclear. The lateral orbital rim of the male isrobust. The frontozygomatic suture is themost posterior point of the orbital rim; itcurves anteriorly both superior and inferiorto this point. The frontal process of thezygomatic slopes anteriorly and increases inbreadth inferiorly as it approaches the zygo-matic arch. The orbits themselves are circu-lar in outline in A.L.363-1a. The orbit of thefemale is too distorted to be sure of itsoriginal shape. The internal morphologyis largely obscured by matrix in bothspecimens.
Calvaria.The calvaria is preserved only in A.L.363-1a. It is slightly crushed in the vicinity ofbregma, and the vault is missing on the leftside superior to the auditory meatus. It isoval in shape, being widest above theauditory meatus. Postorbital constrictionis modest, being more mild than that ofTheropithecus and P. (Dinopithecus). Thesupraorbital torus is separated from the cal-varia by an ophryonic groove. The frontalrises superiorly over 1 cm above the supra-orbital rim, reaching its maximum heightanterior to bregma, before flattening to apoint midway between bregma and lambda.This morphology is similar to that of other

703
known specimens of Parapapio (Freedman,1957). The temporal lines are stronglymarked anteriorly, curving sharply mediallyposterior to the orbital rim. At approxi-mately the midpoints of the orbits the tem-poral lines curve sharply posteriorly and donot meet in the midline until about 1 cmanterior to lambda. At this point they form ashort and low sagittal crest. The regionaround inion is absent, but just lateral to thisthere is a well-developed nuchal crest thatreaches its maximum height of about 5 mmjust posterior to the auditory meatus.
Basicranium.The basicranium is well preserved inA.L.363-1a, except for the portions nearinion and around to the left mastoid. Theoccipital plane is relatively flat and inclinedat an angle of 45� relative to the FrankfurtHorizontal. The mastoid processes are lowand the digastric groove nearly imper-ceptible. The auditory meatus is angledposteriorly at an angle approximately 30� tothe coronal plane. The inferior surface of themeatus is distinctive: it is pinched up into asharp crest that follows the length of thetube. In this morphology it is similar to themale BPI M3051 from Makapansgat, alsoassigned to Pp. jonesi (Maier, 1970). Thetips of the postglenoid processes are broken,but their bases are preserved. They are rela-tively small and gracile in comparison tothose of Theropithecus. The articular sur-face for the mandibular condyle is sellar inshape, being convex anteroposteriorly andconcave mediolaterally. The eminence is notas prominent as that of Theropithecus. Thechoanae are clearly narrow, but they arelargely obscured by matrix making itimpossible to determine their height. Thebasioccipital has a sharp break in slope:immediately anterior to the foramen mag-num it is nearly parallel with the FrankfurtHorizontal, but approximately 1 cm anteriorto this, the slope of the clivus increases byabout 60�.
Facial hafting.The only specimen in which the relationshipbetween the face and neurocranium can bestudied is A.L.363-1a. The glenoid fossa isonly slightly elevated above the level of theocclusal plane. The frontal is significantlyelevated above the orbits. The angulation ofthe face on the neurocranium is similarto that of most papionins, but is lessklinorhynch than Papio hamadryas ursinusand Paradolichopithecus. It is less airorhynchthan Theropithecus gelada.
Mandible.Overall, the mandible is very similar toother well-preserved specimens of Parapapiobroomi (BPI M3067) and Pp. jonesi (BPIM3061) from South Africa. The symphysisof A.L.363-1b slopes at an angle of approxi-mately 50� to the occlusal plane. InA.L.363-15b the symphysis appears morevertical, but this may be due to the damage.This angle is similar to that of Papio, Man-drillus and most species of Theropithecus. Thesymphysis is pierced by a median mentalforamen. There appear to have been faint,triangular mental ridges. The superior trans-verse tori of A.L.363-1b and A.L.363-15bextend posteriorly to the middle of P3 insuperior view. The inferior torus extendsonly a small amount further to the mesialend of P4.
The lateral surface of the corpus of themale shows only a shallow fossa. The femalespecimen also seems to lack corpus fossae,although it is too damaged to be certain. InA.L.363-1b the deepest part of the corpus ispositioned relatively far anteriorly, approxi-mately under the M1/M2 contact. InA.L.363-15b it is difficult to be certain, butit was probably in a similar position. Theoblique line emerges near the level of themesial lophid of M3 or the distal lophid ofM2, and is weakly developed. The extra-molar sulcus is smooth and weakly devel-oped. The gonial area is not expanded, andcurves smoothly to the ramus. If present at

704 .
all, the mylohyoid line is poorly developed.In superior view, the cheek teeth rows areparallel. The canines project laterally in themale, whereas in the female they are in linewith the incisors, which form a short arcanterior to the canines. In lateral view thetooth rows are slightly concave-up.
The ramus is nearly complete in A.L.363-1b, and its anterior portion is preserved inA.L.363-15a and A.L.363-10. The ramus islow, as would be expected given the shallowelevation of the glenoid fossa. It is relativelylong anteroposteriorly and back-tilted, simi-lar to that of Papio, T. (Omopithecus) andMandrillus but significantly more than in T.(Theropithecus). On A.L.363-1b the inferiorlimit of a deep triangular fossa is preserved,and the lateral surface of the ramus is other-wise relatively smooth.
Dentition.Every element of the adult dentition is rep-resented in this sample. There is also anisolated right lower dP4 of a small papionin,from the nearby site of Ahmado, which mayrepresent this species as well. The incisorsare preserved only in the female A.L.363-15, and they are typically papionin inmorphology. The upper incisors lack lingualcingula. The upper central incisors arebroad and spatulate with a vertical lingualgroove. In anterior view, the crown flaresconsiderably from cervix to apex, althoughmore medially than laterally. The lateralincisor is generally similar but has anarrower crown. Its lingual surface is moretightly curved than in the central incisor.The crown is also less flaring in anteriorview, more asymmetrical, and angledmedially. The lower incisors clearly lacklingual enamel. They are ‘‘squared’’ anteri-orly in occlusal view. In anterior view, thecrowns are less flaring than those of theuppers. The distal margin of the lateralincisor is tightly curved and angles mesially.
The canines are typical cercopithecidteeth, being highly sexually dimorphic. The
upper canines of A.L.363-1a are heavilybroken and damaged. What is preservedshows a tooth that was much larger incaliber than the female’s. It is triangular incross-section, with a sharp distal border.Mesially, there is a deep sulcus. The upperfemale canines are relatively compressedlabiolingually. The crown is low and roughlytriangular in labial view. The distal edge isslightly sigmoidal. There is a slight mesialgroove on the root. The female C1s are lowand otherwise substantially smaller thanthose of the male. The C1s of A.L.363-1bare heavily worn, but can be seen to berelatively tall and large in caliber, and thereis a distinct sulcus on the mesial surface ofthe root. The C1s of A.L.363-15b are com-plete but damaged. They are small, beingsimilar in size to the lower incisors, and arerelatively short in the mesiodistal direction.
The upper premolars are typical bicuspidteeth. The P3 is smaller than the P4, butneither has well developed mesial or distalfoveae. The P3 crown is also more triangularin outline in occlusal view. The P3 has a tallprotoconid, and on A.L.363-15b there is alarge metaconid. Both specimens preserve aparaconid that is better developed than thatof Pliopapio, T. (Theropithecus), and Papio.The mesiobuccal flange is relatively short.The male’s is significantly longer than thatof the female, but still shorter than thoseof most papionins. The P4 is more molari-form, with a clear lingual notch, and com-paratively large talonid. That of A.L.363-15b has a small hypoconid. The P4 ofA.L.363-1b has a slight mesiobuccalextension.
The molars are similar to those of mostpapionins. The crowns are low, with a largeamount of basal flare (Figure 8), althoughless than is present in Mandrillus, Cercocebusor Lophocebus. The cusps are low and buno-dont, and the notches between them areshallow. The cusp tips are closely approxi-mated due to the flaring crown. The uppermolars sometimes develop small cuspules

705
in the lingual clefts. The cross-loph(id)suniting the cusps are poorly developed. Ofthe upper molars the M2 is the largest,although the M3 is the longest and similar insize. The lower molars increase in size fromanterior to posterior.
Postcrania.There are several postcranial elementsassociated with the male skull A.L.363-1a-b.These are discussed below under the differ-ent anatomical regions. A.L.363-12 is a dis-tal fragment of a right humerus that mayalso represent Parapapio. In spite of the largecollections of Parapapio from South Africathere are no postcranial fossils directlyassociated with diagnostic cranial material(Elton, 2001). The Hadar sample is the onlypostcranial material assigned to Parapapioknown that is directly associated withdiagnostic cranial remains.
Axial skeleton.A.L.363-1g preserves all three sacral ver-tebrae. It is slightly larger than the sacrumassociated with the female T. o. cf. dartipartial skeleton A.L.193-6. In morphologyit appears to be typical for the family,with a well-developed distal body and zyga-pophyses for articulation with the firstcaudal vertebra. The neural canal is alsolarge and patent distally. A.L.363-1k is athoracic vertebra, which appears typical forcercopithecids. A.L.363-1h/8 is a fragmentof a right rib.
Figure 8. Box and whisker plot of M3 basal flare: the ratio of M3 mesial breadth/M3 mesial breadth at theheight of the notch. The central bar of each box represents the median, or 50th percentile. The left andright of each box represent the value of the 25th and 75th percentiles respectively. The whiskers extend tothe farthest observation that is less than 1·5 times the length of the box. Any individuals outside of thewhisker range are marked separately. Some data from M. Singleton.
Forelimb.A.L.363-12 is a partial right humerus, pre-serving the distal half of the shaft, althoughthere is considerable damage, so that onlythe distal 3 cm are well preserved (Figure 9).This specimen is apparently not associatedwith the cranial material, and it almost

706 .
Figure 9. Hadar distal humeri, anterior above distal views (distal of casts except A.L.2-64). Upper rowsleft to right: A.L.259-1 (T. o. cf. darti, probable male), A.L.196-3c (T. o. cf. darti, associated with femalemandible), A.L.363-12 (cf. Pp. jonesi, probable male; reversed). Lower rows left to right: A.L.300-1 (cf.Rhinocolobus turkanaensis); A.L.2-64 (Cercopithecoides meaveae); A.L.222-14 (?Cercopithecoides meaveae);A.L.577-1 (?Cercopithecoides kimeui, distal view not available).
certainly represents a different individualfrom A.L.363-1. It is morphologically dis-tinct from and slightly larger than thehumeri associated with Hadar T. o. cf. dartiand those identified as T. o. cf. darti fromHadar by Krentz (1992, 1993; see list inDelson et al., 1993). The medial epicondyleis long, large, and projects medially. Therelative projection of the medial epicondyleis closer to the means for more arboreallyadapted cercopithecid species, but is stillwithin the range for humeri allocated to T. o.cf. darti (see Figure 10). Its angle of retro-flection is relatively small, being outside ofthe T. o. cf. darti range, but slightly less thanthat of A.L.300-1 (here allocated to cf. Rhi-nocolobus turkanaensis), while similar to thoseof Cercopithecus mitis, Lophocebus albigena,
and the extant African colobines. It isslightly more retroflexed than the Asianforms in Figure 11. The capitulum is roundand projecting, and the zona conoidea is flat.The medial trochlear flange is short, anddoes not come to a sharp angle, unlike thoseof the T. o. cf. darti. The length of thetrochlear flange relative to the articularwidth is shorter than for any other humerusfrom Hadar. It falls within the lower rangefor L. albigena and Macaca fascicularis,as well as near the means for the morearboreally adapted colobines (see Figure12). The coronoid fossa is very deep androunded. The radial fossa is quite shallowand low. This specimen has a m. brachio-radialis flange that is less prominent thanin T. o. cf. darti and significantly shorter

707
proximodistally. Posteriorly, the olecranonfossa is broad and deep, with a smallforamen on its superior surface. Ciochon(1993) identified this specimen as Rhinocolo-bus turkanaensis, but Delson (1984; Delsonet al., 2000) considered it more likely tobe Parapapio. The latter view is followedhere as the coronoid fossa is deeper thanthe radial (Olivier & Caix, 1959; Szalay& Delson, 1979), and the specimen notflattened anteroposteriorly, a feature whichcharacterizes the only known humerus allo-cated to Rhinocolobus in the Turkana Basin(see discussion of that genus below). Fur-thermore it is from the same locality asseveral other specimens of Pp. cf. jonesi.However, one feature that may argue for itsbeing Rhinocolobus is its comparatively large
size, and Ciochon’s identification remains apossibility. From the locomotor viewpoint,this specimen appears to be less adapted to aterrestrial substrate than are Theropithecus,Papio, Paradolichopithecus or Mandrillus, andit may have been more similar to taxa suchas Lophocebus or the more arboreal membersof Macaca.
Figure 10. Box and whisker plot of medial epicondyle projection: the ratio of biepicondylar breadth(B)–medial distal articular limit to lateral epicondyle (A)�100/biepicondylar breadth (B). Some datafrom T. Harrison.
Hindlimb.Proximal and distal ends of the rightfemur are preserved. A.L.363-1c is theentire proximal end including the greatertrochanter, head and shaft to approximately1 cm distal to the lesser trochanter (seeFigure 13). It has a relatively long neckin comparison to the other proximal femorafrom Hadar. The head is not cranially

708 .
Figure 11. Box and whisker plot of angle of medial epicondyle relative to axis of distal articular surface.Some data from T. Harrison.
oriented, with a neck-shaft angle of 114�,which overlaps that of many species, but isless than M. fascicularis and most extantcolobines (see Figure 14). The greatertrochanter is approximately 9 mm taller thanthe head and hooks sharply medially. Whenthe height of the greater trochanter abovethe neck is compared to femoral head diam-eter, A.L.363-1c is within the range of mostextant cercopithecines, though greater thanthat of L. albigena and most extant colobines(see Figure 15). It does overlap with thefemora assigned to T. o. cf. darti by Krentz(1992; Delson et al., 1993). The lessertrochanter is long and medially oriented.The gluteal fossa extends inferior to the m.quadratus femoris insertion. The fovea capitisis short and oval. A.L.363-1d is the distalend of the same femur. It is very similar to
the other distal femora from the same hori-zons (most of these presumably representT. o. cf. darti) except that the patellar groovemay be deeper and narrower than the others.It also has higher medial and lateral margins.
A.L.363-1f/l is a right fifth metatarsal (seeFigure 4), 64 mm in length. This is similarto fifth metatarsals of male extant Papio, butsmaller than those of male Mandrillus. It isslightly longer than would be expected for afifth metatarsal of a cercopithecid with abody mass of 16 kg (as estimated from thedentition by Delson et al., 2000). It is con-siderably smaller than the fifth metatarsalof Paracolobus chemeroni (KNM-BC 3aa). Itis similar in overall morphology to the fifthmetatarsals of other cercopithecids. Theproximal articulation for the cuboid is trian-gular in outline and is continuous with the

709
articular surface for the fourth metatarsalmedially. Inferiorly and laterally, there is asmall sulcus between the cuboidal articularsurface and the edge of the basal tubercle.This sulcus is larger than in modern Papio,but not as deep or strongly rimmed as it isin Mandrillus. There is also a small articularfacet on the inferior surface of the tuberclefor a sesamoid bone.
Figure 12. Box and whisker plot of medial trochlear flange length (A)/distal humeral articular breadth (B).Some data from T. Harrison.
RemarksWhile the most completely preserved indi-vidual of this sample shows several uniquefeatures, many of these may be explained byindividual variation. It is also not possible tobe certain if the unique features of thisindividual are related to its advanced age, sothat younger members of the taxon might be
different. However, there are a number offeatures that may well warrant specific dis-tinction. The most striking are the shape ofthe nasals, the thickness of the supraorbitaltorus, and the narrow and deep palate.Before it can be determined more confi-dently whether the Afar taxon is conspecificwith any of the South African forms a com-prehensive review of the large South Africansample of Parapapio is required with morethorough diagnoses of those taxa and abetter understanding of their variability.Pending such a revision, the Afar materialis recognized as most similar to Pp. jonesi,particularly the material from Makapansgat,and is here termed Pp. cf. jonesi. It resemblesPp. jonesi and Pp. whitei, but is distinct fromPp. broomi, in that the nasals are more

710 .
Figure 13. Hadar proximal femora, posterior view. Top row left to right: A.L.363-1d (Pp. cf. jonesi, male),A.L.175-21 (likely T. o. cf. darti, probable male), A.L.341-5a (likely T. o. cf. darti, probable female).Lower row left to right: KNM-ER 4420 (C. williamsi, male from Koobi Fora), A.L.2-72+A.L.2-71 andA.L.2-70 (C. meaveae).
prominent, facial fossae are variably present(one of two Hadar specimens has them), thepremaxillae are more anteriorly projecting,and the temporal lines are more stronglymarked. The Afar sample resembles Pp.jonesi more than it does Pp. whitei due toits relatively shorter rostrum, considerablysmaller size, and the crest on the ventralsurface of the tympanic. Lastly, it showsnone of the distal molar reduction andanterior molar lengthening of Pp. antiquus(Maier, 1970), nor the distinctive facialarchitecture of that species.
At present, Parapapio can be diagnosedonly on facial evidence, and thus the Hadar
material is the only definitive evidence of thegenus in East Africa in the Pliocene andPleistocene. All other samples or individualspecimens that have been assigned toParapapio from the Pliocene or Pleistoceneof East Africa (e.g., Laetoli, Kanapoi,and Omo) lack diagnostic facial material.These assignments, including the genericallocation of Pp. ado, therefore must beconsidered tentative. From the LateMiocene Nawata Formation at Lothagam,there is a small species of Parapapio whichdoes preserve facial material (Leakey et al.,in press). Its relationship to later material isunknown.

711
Figure 14. Box and whisker plot of femoral neck-shaft angle. Some data from Ting (2001).
As discussed by Elton (2001), amongothers, there are no associations yet knownbetween cranial and postcranial elements ofParapapio from southern Africa. The Hadarfemur A.L.363-1 c/d and metatarsal -1f/l aremost likely from the same individual asthe male skull, while the distal humerusA.L.363-12 probably represents a differentindividual of the same population. Theinformation about locomotor adaptationfrom these specimens is the first evidenceknown for the genus. The overall pattern ofmorphology of the humerus of Parapapiomaterial from Hadar suggests a taxon witha more arboreal habitus than Mandrillus,Papio or Theropithecus, being closer to thearboreal macaques or mangabeys. The fem-oral morphology is more ambiguous, over-
lapping the ranges of both more terrestrialand more arboreal taxa.
Genus Theropithecus Geoffroy, 1843(=or including Macacus Ruppell, 1835 (inpart). Gelada Gray, 1843. SimopithecusAndrews, 1916. Theropythecus Vram, 1922(lapsus?). Papio Erxleben, 1777: Broom &Jensen, 1946 (in part); Buettner-Janusch,1966 (in part). Dinopithecus Broom, 1937:Arambourg, 1947 (in part); Broom &Hughes, 1949 (in part). Brachygnatho-pithecus Kitching, 1952 (in part). Gorgo-pithecus Broom & Robinson, 1949: Kitching,1953 (in part). T. (Omopithecus) Delson,1993.)Type species Theropithecus gelada (Ruppell,1835)

712 .
Other included species: T. oswaldi(Andrews, 1916), T. brumpti (Arambourg,1947), ?T. baringensis (R. E. F. Leakey,1969).
Figure 15. Box and whisker plot of the height of the greater trochanter above the neck�100/antero-posterior diameter of the femoral head. Some data from Ting (2001).
Generic diagnosisEck & Jablonski (1987), Jablonski (1993)and Delson (1993) have all providedrecent diagnoses for the genus Theropithecus,and these are followed here. Theropithecusis a medium to very large-sized papionin.The neurocranium can be distinguishedfrom those of Papio (Papio), Parapapio,Mandrillus and Pliopapio by the presence of awell developed anteriorly positioned sagittalcrest. The postorbital region is greatlyconstricted and the zygomatic arches are
widely flaring, yielding a large infratem-poral fossa. The lower portion of the faceis tall, due to the posterior portion ofthe maxilla being deep, resulting in thetemporomandibular joint being elevatedrelative to the occlusal plane. A steep ante-orbital drop characterizes the facial profile.This is unlike the anteorbital region ofParapapio, Cercocebus, Lophocebus andsome Macaca. Furthermore, it is generallysteeper and the vertical segment is longerthan in Papio and Pliopapio. The pre-maxillae are short in comparison to themaxillae, unlike Pliopapio, Papio, Lophoce-bus, Mandrillus, Cercocebus, Gorgopithecusand Macaca but similar to those ofParadolichopithecus.

713
Theropithecus is most clearly distinguishedfrom all other papionins by its dentition.The incisors are small relative to the molarteeth, particularly in comparison to Papio,Mandrillus, Cercocebus, and Lophocebus. Thecheek teeth are highly derived, being highcrowned and straight sided with a lowamount of basal flare (see Figure 8) and alarge amount of cuspal relief, deeply exca-vated notches and foveae. The cusps them-selves are columnar in form, being separatedby deep basins. On the lower molars, themedian cleft is flattened at its base into a‘‘pocket’’, the lophids are angled mesio-lingually, and there is sometimes a largedistal accessory cuspule present on M1–2.
While T. gelada has several known autapo-morphies in the postcranium relative toother papionins, only a few are known forthe fossil species. One of the most importantis the presence of elongate first and shortsecond metacarpals. This feature givesTheropithecus the highest opposability indexof any cercopithecid and is related to‘‘manual grazing’’ behavior. It is known inT. brumpti from the Omo and West Turkana(Jablonski, 1986; Jablonski et al., 2002).Both manual and pedal phalanges are shortrelative to their breadth and to overall footlength (Jolly, 1972; Strasser, 1992). Thefemur shows a reverse carrying angle, poss-ibly related to ‘‘bottom shuffling’’ locomotorbehavior (Krentz, 1993).
RemarksTheropithecus is a widely distributed genus inthe fossil record, and often easily diagnoseddue to its highly distinctive molar dentition.Delson (1993) recognized two subgenera,which we accept here. The subgenus(Omopithecus) includes the enigmaticspecies T. (O.) brumpti and possibly ?T.baringensis. There is some question as towhether ?T. baringensis truly represents aspecies of Theropithecus, Papio (as originallydescribed) or some other primitive papioningenus. Delson & Dean (1993) provide a
thorough review of these arguments, andtheir position is followed here with it tenta-tively recognized as a primitive member ofthe genus, possibly in the T. (Omopithecus)subgenus. It should be noted, however, thisacceptance is mainly based on contextualevidence as described in Leakey (1993).
There are specimens from a number offossil localities that are diagnosable to thegenus, but which cannot be allocated toeither of the recognized subgenera, basedsolely on the preserved morphology. Thesesites include: Ain Jourdel, Algeria; Fejej,Ethiopia; Kanam East, and Nyeri, Kenya;Kaiso, Uganda; Senga, Zaire; Malemba,Malawi; Cueva Victoria, Spain; andMirzapur, India. Given the wide geographicdistribution and chronological range of T.oswaldi, and the relatively restricted rangeand distribution of T. brumpti and T. gelada,these fossils most likely represent T. oswaldi.However, because they cannot be diagnosedto any species based on their preservedmorphology they are best regarded asTheropithecus sp. indeterminate. Some ofthese have been tentatively included in thehypodigms given for the subspecies of T.oswaldi to indicate where they would best fitif they were to represent T. oswaldi, but theyare preceded by a question mark to indicatethe tentative nature of this allocation.
Two of these specimens have been desig-nated as holotypes. The single tooth fromAin Jourdel is the type of Cynocephalusatlanticus (Thomas, 1884) and that fromMirzapur of Theropithecus delsoni (Gupta &Sahni, 1981). However, neither of these canbe said, with any confidence, to be con-specific with the more complete materialallocated to T. oswaldi, nor can they beclearly diagnosed as separate species (seealso Delson, 1993). This presents a nomen-clatural problem, as T. atlanticus (Thomas,1884) would have priority over T. oswaldi(Andrews, 1916) if the populations involvedwere judged conspecific. On the other hand,Alemseged & Geraads (1998) argued that

714 .
fossils from Ahl al Oughlam in Morocco areconspecific with the holotype of T. atlanticusfrom Ain Jourdel, and that this taxon is inturn specifically distinct from T. oswaldi. Werespond that the Ahl al Oughlam materialis consistent with the variation seen withinT. oswaldi and does not warrant specificdistinction; furthermore, its position relativeto the Ain Jourdel molar is no more clearcut than that of any other population ofT. oswaldi. Given the ambiguity surroundingT. atlanticus, the name has not been appliedto what we here term T. oswaldi for the sakeof clarity, but this is also not intended toimply the specific distinction of T. atlanticus.
Theropithecus (Theropithecus) Geoffroy, 1843(=or including Macacus Ruppell, 1835 (inpart). Gelada Gray, 1843. SimopithecusAndrews, 1916. Theropythecus Vram, 1922(lapsus?). Papio Erxleben, 1777: Broom &Jensen, 1946 (in part); Buettner-Janusch,1966 (in part). Dinopithecus Broom, 1937:Broom & Hughes, 1949 (in part). Brachy-gnathopithecus Kitching, 1952 (in part).Gorgopithecus Broom & Robinson, 1949:Kitching, 1953 (in part).)Type species Theropithecus gelada (Ruppell,1835)Other included species: T. oswaldi(Andrews, 1916)
Subgeneric diagnosisDelson (1993) divided Theropithecus intotwo subgenera, which are recognized here.His diagnoses are followed, along withdescriptions by Eck & Jablonski (1987) andLeakey (1993). T. (Theropithecus) aremedium to very large members of Thero-pithecus distinguished from T. (Omopithecus)in all of the following features. The muzzle isshorter and the face more airorhynch. Themaxillary ridges are either weakly developedor absent. When they are present, they arerounded in cross-section, unlike those of T.brumpti that are more triangular. The dorsal
surface of the muzzle is sellar. It is roundedand convex in parasagittal cross-section andconcave in profile. The zygomatic arch isrobust, but not greatly expanded and flared.The greater tuberosity of the humerusprojects proximally above the head (Krentz,1993).
Theropithecus oswaldi (Andrews, 1916)(See subspecies for synonymy.)Holotype: BMNH-M11539 (lectotype)from Kanjera, KenyaSubspecies included: T. o. oswaldi(Andrews, 1916), T. o. leakeyi (Hopwood,1934), T. o. darti (Broom & Jensen,1946).Specimens included: see subspecificdescriptions below.Distribution: see subspecific descriptionsbelow, plus ?Mirzapur, India; ?CuevaVictoria, Spain.
Specific diagnosisThe concept of Theropithecus oswaldi usedhere follows that of Leakey (1993). Threechronologically sequential subspecies arerecognized within Africa, each of whichspans a large geographic area. This species isdistinguished from the other undisputedmembers of the genus Theropithecus, T.gelada and T. brumpti, largely on the basisof characters in the cranium and anteriordentition.
There are several morphological trendsdisplayed by the subspecies of T. oswaldi,which show their origins in T. o. darti andtheir most extreme expressions in T. o.leakeyi. These trends are also features thatdistinguish this species from T. brumpti andto a lesser extent T. gelada.
Through time there is a general increasein body size from early T. o. darti (similar insize to T. gelada) to the largest T. o. leakeyi(similar in size to Gorilla females) (e.g.,Jolly, 1972; Eck, 1987; Krentz, 1993;Delson et al., 2000). Thus, early members ofthis species can be separated from T. brumpti
partly because they are smaller, and later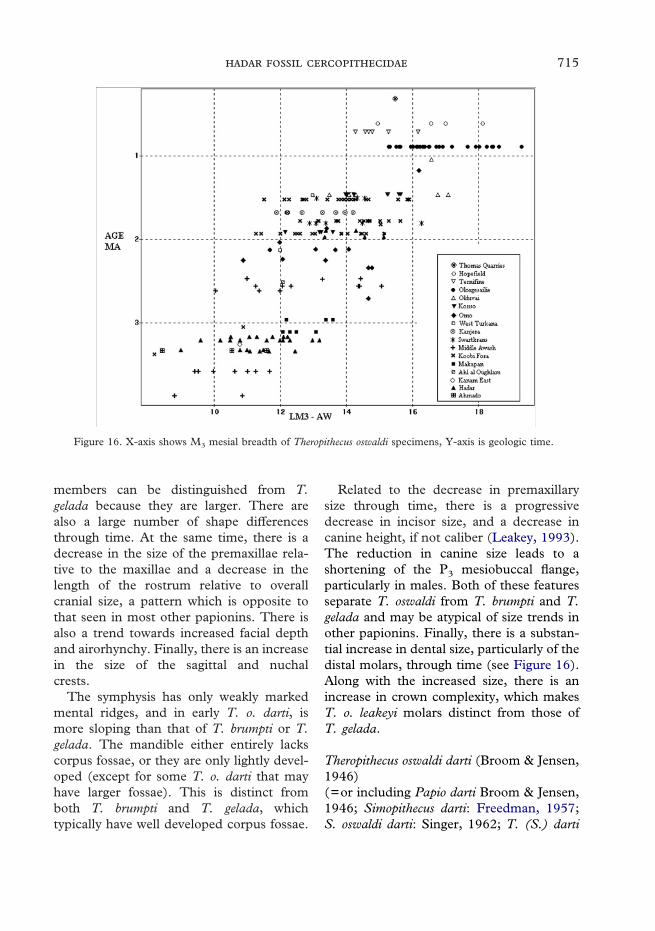
715
Figure 16. X-axis shows M3 mesial breadth of Theropithecus oswaldi specimens, Y-axis is geologic time.
members can be distinguished from T.gelada because they are larger. There arealso a large number of shape differencesthrough time. At the same time, there is adecrease in the size of the premaxillae rela-tive to the maxillae and a decrease in thelength of the rostrum relative to overallcranial size, a pattern which is opposite tothat seen in most other papionins. There isalso a trend towards increased facial depthand airorhynchy. Finally, there is an increasein the size of the sagittal and nuchalcrests.
The symphysis has only weakly markedmental ridges, and in early T. o. darti, ismore sloping than that of T. brumpti or T.gelada. The mandible either entirely lackscorpus fossae, or they are only lightly devel-oped (except for some T. o. darti that mayhave larger fossae). This is distinct fromboth T. brumpti and T. gelada, whichtypically have well developed corpus fossae.
Related to the decrease in premaxillarysize through time, there is a progressivedecrease in incisor size, and a decrease incanine height, if not caliber (Leakey, 1993).The reduction in canine size leads to ashortening of the P3 mesiobuccal flange,particularly in males. Both of these featuresseparate T. oswaldi from T. brumpti and T.gelada and may be atypical of size trends inother papionins. Finally, there is a substan-tial increase in dental size, particularly of thedistal molars, through time (see Figure 16).Along with the increased size, there is anincrease in crown complexity, which makesT. o. leakeyi molars distinct from those ofT. gelada.
Theropithecus oswaldi darti (Broom & Jensen,1946)(=or including Papio darti Broom & Jensen,1946; Simopithecus darti: Freedman, 1957;S. oswaldi darti: Singer, 1962; T. (S.) darti

716 .
darti: Jolly, 1972; T. (S.) darti: Szalay &Delson, 1979, in part; T. darti: Eck &Jablonski, 1984, 1987 Eck, 1993, Delson,1993, in part; T. o. darti: Leakey 1993, inpart)Holotype: UWMA MP1 (=M201, 1326/1)Distribution: Makapansgat (type locality).
No specimens from the Hadar sample aredefinitively included in this taxon (seebelow).
Subspecific diagnosisAn early subspecies of T. oswaldi smaller incranial, dental, and postcranial size than T.o. oswaldi and T. o. leakeyi. The rostrum islong relative to overall cranial size. Maxillaryand mandibular corpus fossae are variable,but often more pronounced than in latersubspecies of T. oswaldi. The incisors arerelatively large, although smaller comparedto molar width than those of Papio. Themale canines are tall, being similar in sizeto those of T. gelada and Papio. The P3
mesiobuccal honing flange is also relativelylong. The molar teeth show the specializa-tions of the genus, but weakly developed incomparison to later subspecies.
Theropithecus oswaldi cf. darti (Broom &Jensen, 1946)(=or including T. (S.) darti: Szalay &Delson, 1979, in part; T. darti: Eck &Jablonski, 1984, 1987, Eck, 1993, Delson,1993, in part; T. o. darti: Leakey 1993, inpart)Specimens included: see Eck, 1993; Delsonet al., 1993; and Frost, 2001b.Distribution: Hadar Fm. Sidi Hakoma–Lower Kada Hadar; Ahmado, ?Leadu;Maka; ?Bunketo; Matabaietu; Wee-ee;?Shungura Fm., C-6; Koobi Fora Fm., TuluBor Mbr.; ?Kanam East.
DescriptionEck (1993) has thoroughly described all ofthe T. o. cf. darti cranial material from the
Hadar Formation that was available at thetime. Krentz (1993) has described the longbones of the forelimb and hindlimb cata-logued in Delson et al. (1993). Someadditional material has been collected atHadar since these studies were published,and these are currently under analysis byG. G. Eck.
In overall cranial size, this material issimilar to T. gelada and medium-sized Papiohamadryas subspecies, such as P. h. cyno-cephalus. Both sexes are smaller than in T.brumpti, T. o. oswaldi and T. o. leakeyi, aswell as Papio (Dinopithecus) and Paradolicho-pithecus. The dentition of T. o. cf. darti fromthe Afar region is slightly smaller than thatfrom Makapan, and similar to that of mostsubspecies of P. hamadryas in size. It issmaller in dental size than younger sub-species of T. oswaldi. Dental dimensions ofT. o. cf. darti are given in Eck (1993).Selected postcranial specimens of HadarT. o. cf. darti are included in Figures 9and 13.
RemarksEck (1987, 1993), and others (e.g., Delson,1993), recognized T. o. darti at the specificlevel distinct from T. oswaldi. While agreeingthat T. darti is the sister taxon to T. oswaldiand likely its ancestor, Eck has argued forspecific distinction based upon severalcranial features. Most important of these arethe ‘‘concavo-convexo-concave’’ dorsal sur-face of the rostrum, the relatively largeincisors and male canines, and the smalloverall body (or at least cranial) and molarsize of T. o. darti. However, the only popu-lation of T. o. leakeyi to preserve the muzzleis that from the Middle Awash (Kalbet al., 1982b; Delson et al., 2000; Frost,2001b), which also presents the distinctive‘‘concavo-convexo-concave’’ dorsal rostralsurface. Furthermore, as pointed out by Eck(1987; 1993), the differences in the anteriordentition (Leakey, 1993; Frost, 2001b) andbody size (Delson et al., 2000) can be

717
interpreted as anagenetic trends which beginin T. o. darti, continue through T. o. oswaldiand find their most extreme expression inT. o. leakeyi. This situation requires thatthe three chronological forms be recognizedeither as three separate species, or assubspecies of T. oswaldi as is done here.
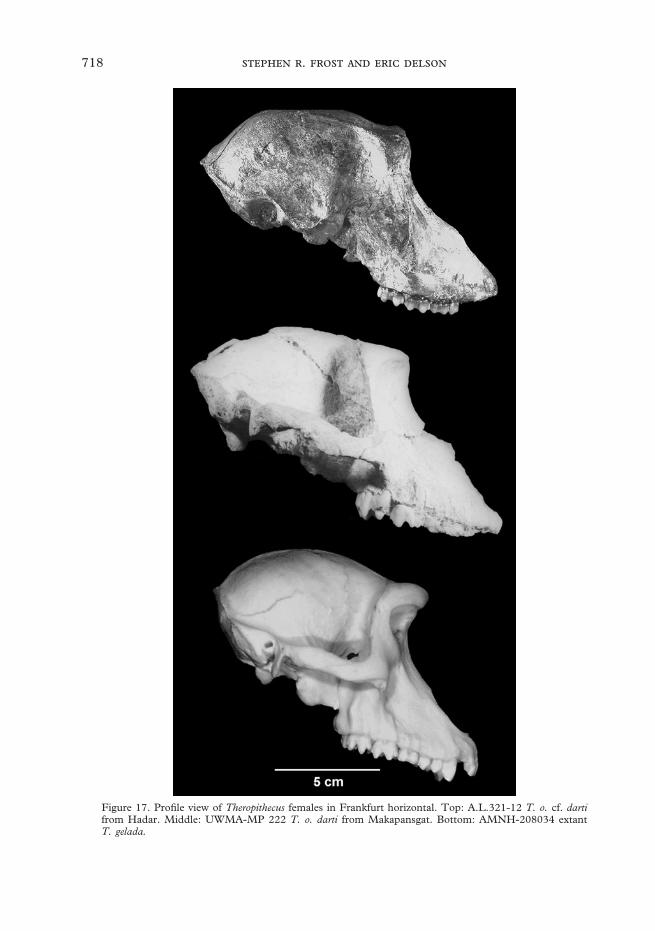
There is, however, considerable differencebetween the type sample of T. o. darti fromMakapansgat and the Hadar sample. In fact,the differences between these two assem-blages are larger than the variation amongdifferent samples of T. o. oswaldi or T. o.leakeyi. There are no male crania fromMakapansgat to compare with those fromHadar, but there are several fairly goodfemale crania. The nearly complete femalecranium UWMA-MP222 and the sub-adultfemale BPI-M3073 both possess browridges that project more anteriorly than dothose of the Hadar female A.L.321-12, sothat glabella is either directly superior to oreven slightly anterior to nasion. In this fea-ture the Makapansgat specimens are morelike T. gelada and some females of T. o.oswaldi (e.g., TMP SK 561 and BMNH-M14936 from Swartkrans and Kanjerarespectively). However, in A.L.185-5a thebrows are slightly more projecting than inA.L.321-12. The rostral profile of theMakapansgat specimens is deeper, moreconvex, and somewhat more similar to thatof T. gelada than A.L.321-12 (Figure 17).Both fossils appear to share longer nasalsthat are more prominent at rhinion, produc-ing a larger difference in slope between thenasals and premaxillae than is the case inT. gelada. However, there is damage to themuzzle dorsum and distal part of the nasalsand proximal part of the piriform aperture,rendering accurate assessment of the profiledifficult in MP222. Both of the femalecrania from Makapansgat have maxillaryridges, that while small, are more sharplymarked than are those of any of the Hadarspecimens. This feature, however, is variableamong later assemblages of T. oswaldi and
also among extant subspecies of Papio; it isthus probably consistent with geographicvariation within a widely distributed species.The variation in mandibular form atMakapansgat is greater than at Hadar (Eck& Jablonski, 1987; Eck, 1993): the femaleUWMA MP56 (M633) possesses only avery shallow corpus fossa, but the maleUWMA MP44 (M626) has a corpus fossathat is deeper than known in any otherspecimen assigned to the T. oswaldi lineage,and in fact overlaps in size with somespecimens of T. brumpti and T. gelada (seeEck, 1993: Table 2.12). Finally, theMakapansgat sample has slightly largermolar teeth than at Hadar, but this is con-sistent with the South African material beingslightly younger in age than most of theHadar material.
In all of the features discussed above, itmay be that the Hadar sample representsthe more conservative condition, with theMakapansgat material being derived inthe direction of T. gelada in the case of therostral profile, and browridges, or inthe direction of T. oswaldi in the case of thedentition. Given that the polarity of theabove features is not completely understood,and that there is damage to specimens inimportant areas (the brow of A.L.321-12and the distal nasals of MP222) it is prema-ture to argue for taxonomic distinction. Themandibular corpus fossa present in theMakapansgat male seems to distinguish thissample from all other T. oswaldi, especiallyconsidering the presence of only shallowor no fossae in both the earlier Hadarassemblage and later samples (e.g., fromSwartkrans, upper Kada Hadar, KoobiFora, Matabaietu, Kanjera, Olduvai,Olorgesailie, Ternifine). These features andothers may argue for the distinction of theMakapansgat sample at the subspecific level.If so, a new name will be required forthe Hadar and other early representatives ofT. oswaldi which lack the deep mandibularcorpus fossa.
718 .
Figure 17. Profile view of Theropithecus females in Frankfurt horizontal. Top: A.L.321-12 T. o. cf. dartifrom Hadar. Middle: UWMA-MP 222 T. o. darti from Makapansgat. Bottom: AMNH-208034 extantT. gelada.

719
Figure 17. Continued. Top: BM(NH) M14936 T. o. oswaldi from Kanjera. Middle: TMP-SK 561T. o. oswaldi from Swartkrans. Bottom: NME-L32–155 T. brumpti from the Shungura Formation.

720 .
Subspecific diagnosisA subspecies of T. oswaldi distinguishedfrom T. o. darti by its larger overall cranial,molar and postcranial size, and from T. o.leakeyi by its smaller size. The incisors andcanines are relatively smaller than those ofT. o. darti and are relatively larger than thoseof T. o. leakeyi. It is further distinguishedfrom T. o. leakeyi by having a relativelylonger muzzle. Mandibular corpus fossaeare generally shallower than those of T. o.darti. The symphysis is more vertical inprofile with less rugose mental ridges thanthat of T. o. darti. Relative to T. o. leakeyi,T. o. oswaldi develops mandibular corpusfossae more often, and has a more slopingsymphysis. Additionally, the anterior surfaceof the symphysis rarely develops a triangularanterior fossa as it does in T. o. leakeyi. The
molars are generally more complex, and aremore readily identifiable to the genus, thanthose of T. o. darti but they have less enamelcomplexity and fewer folds than do those ofT. o. leakeyi.
DescriptionThis material is currently under analysis byG. G. Eck, and will be described elsewhere.Kimbel et al. (1996) noted the presence ofT. o. oswaldi in the Hadar Formation abovethe large disconformity in the Kada HadarMember. Theropithecus o. oswaldi is the mostcommon and abundant African fossil cerco-pithecid and has been thoroughly describedfrom several localities, including Kanjera,Olduvai Gorge, Swartkrans, the ShunguraFormation, and Koobi Fora (Freedman,1957; Leakey & Whitworth, 1958; Jolly,1972; Eck, 1987; Delson, 1993; Leakey,1993). The Hadar material is generally simi-lar to that known from other Pliocene andearly Pleistocene sites.
Theropithecus oswaldi leakeyi (Hopwood,1934)(=or including S. leakeyi Hopwood, 1934;S. oswaldi olduvaiensis Leakey & Whitworth,1958; S. jonathani Leakey & Whitworth,1958; S. oswaldi mariae Leakey & Whitworth(nomen nudum), 1958; S. oswaldi hopefield-ensis Singer, 1962; S. oswaldi leakeyi:Leakey, 1965; T. (S.) oswaldi leakeyi: Jolly,1972; T. (S.) oswaldi mariae Jolly, 1972; T.(S.) oswaldi cf. oswaldi: Jolly, 1972; T. (S.)aff. oswaldi ssp. indet A, Szalay & Delson,1979; T. (S.) oswaldi hopefieldensis: Szalay &Delson, 1979; T. o. leakeyi: M. G. Leakey,1993)Holotype: BM(NH) M14680 from OlduvaiBed IV, TanzaniaRange: 1·65–�0·4 MaDistribution: Ternifine; Thomas Quarries;Asbole, Andalee, Bodo, Dawaitoli,Hargufia; Konso (upper); Shungura Fm.,Mbr. L; ?Nachukui Fm., NariokotomeMbr.; Olorgesailie; Kapthurin; Olduvai
Theropithecus oswaldi oswaldi (Andrews,1916)(=or including Cynocephalus atlanticusThomas, 1884: Alemseged & Geraads,1998; Simopithecus oswaldi Andrews, 1916;S. oswaldi oswaldi: Leakey & Whitworth,1958; S. danieli Freedman, 1957; T. (S.)darti danieli: Jolly, 1972; T. (S.) oswaldioswaldi: Szalay & Delson, 1979; T. oswaldi:Eck, 1987 (in part) T. oswaldi oswaldi:Leakey, 1993)Holotype: BM(NH) M11539 (lectotype)from Kanjera, KenyaSpecimens included: see Frost, 2001b.Distribution: ?Ain Jourdel, Ahl al Oughlam,Hadar Fm., Upper Kada Hadar Mbr.;?Geraru; Gamedah, Wilti Dora, Matabaietu,Halsaiya; Bouri Fm., Hatayae Mbr.; Konso(lower); Shungura Fm. Mbs. E3–G14 (H3–K); ?Fejej; Nachukui Fm., Kalochoro–Nariokotome Mbs.; Koobi Fora Fm. UpperBurgi–Okote Mbs.; Kanjera (type locality);?Marsabit; ?Kaiso; Peninj; Olduvai Bed I,Lower Bed II; ?Chiwondo Beds Unit3A; Swartkrans Mbs. 1 (including hangingremnant)-3; Gladysvale.

721
Beds Upper II–IV (type locality), Masek;?Nyeri; Hopefield; Gladysvale.
Subspecific diagnosisThis subspecies is distinguished from bothT. o. darti and T. o. oswaldi by its largercranial, molar, and postcranial size. Therostrum is shorter relative to cranial size andthe zygomatic is positioned more anteriorly(typically near M2). Cranial superstructuresare larger than in any other known cerco-pithecid. The posterior maxilla is deeper.The mandibular symphysis lacks mentalridges, and often has a triangular fossa onthe anterior surface between the roots ofthe canines. The area for the incisors isvery small, and the canine roots convergeinferiorly. The mandible completely lackscorpus fossae, the ramus is tall and vertical.The molar teeth are larger than those ofother subspecies. The M3 is larger relative tothe M2, which is in turn larger relative to theM1 than in other subspecies. Molar enamelcomplexity is also greater in this subspeciesthan in the older subspecies. The incisorsare relatively smaller. The canines are rela-tively shorter, though are often still verybroad. The P3 mesiobuccal flange is likewiseshort.
RemarksThere are no specimens of T. o. leakeyi fromthe sample discussed, but it is included herefor completeness. The taxon is known in theAfar Depression from the sites of Busidima,Andalee, Bodo, Dawaitoli and Hargufia(Kalb et al., 1980; 1982a,b; Clark et al.,1994; Gilbert et al., 2000; Alemseged &Geraads, 2001; Frost, 2001b).
Subfamily Colobinae Jerdon, 1867Genus Rhinocolobus M. G. Leakey, 1982(=or including Colobinae gen. et sp. nov.M. G. Leakey & R. E. F. Leakey, 1973;M.G. Leakey, 1976; Eck, 1976, 1977; cf.Genus et sp. nov. Omo M. G. Leakey &
R. E. F. Leakey, 1973. Colobinae gen. etsp. nov. 1. Szalay & Delson, 1979, in part)Type species Rhinocolobus turkanaensisM. G. Leakey, 1982
Generic diagnosisThe generic diagnosis for Rhinocolobus hasnot changed since its original descriptionby Leakey (1982), which will largely befollowed here. The most diagnostic featuresof the genus are concentrated in the face,which is airorhynchous overall, compared toother large colobines. The interorbital pillaris relatively narrow, distinguishing it frommost colobines other than Nasalis, Dolicho-pithecus and Libypithecus. The rostrum isrelatively long, at least sub-nasally, anddifferent from that of Cercopithecoides,Kuseracolobus, Colobus and Procolobus. Thenasals are extremely short, even relative toother colobines. The piriform aperture isunique among cercopithecids. It is very longanteroposteriorly and relatively wide for acolobine. It is oriented so that its margin liesin a plane that is at a low angle relative to thealveolar plane. In profile, the piriform aper-ture, and face in general, is concave inoutline, not unlike Pygathrix or Rhinopithecusin this one aspect, but is substantially longer.Additionally, it can be observed on somespecimens that there is what appears to be amuscle scar around the superior half of therim (personal observation). The supraorbitaltori are prominent, projecting, and separ-ated from the neurocranium by a deepophryonic groove. The neurocranium isrelatively long and narrow, with only a smallposterior sagittal crest in the male. Nuchalcrests are present in both males and females.Unfortunately, there is little facial materialpreserved in the sample from Hadar.
The mandibular symphysis is pierced by amedian mental foramen (in the Turkanabasin material, but see below) which distin-guishes it from Colobus, Paracolobus, Kusera-colobus, Paracolobus, but not from Procolobus

722 .
(Procolobus). The symphysis also lacks men-tal ridges as do those of most colobines, butis unlike the symphysis of P. (Procolobus).It is relatively sloping in profile, whencompared to other large colobines such asParacolobus mutiwa, or Cercopithecoides. Thesymphysis and corpus are relatively deep,narrow, and deepen posteriorly, similar toParacolobus, but different from Cercopithe-coides. The corpus lacks facial fossae orprominentia laterales, further separating itfrom Cercopithecoides. The gonial region islarge, and in the males it appears to havebeen inferiorly expanded.
Dentally, the incisors are small relative tothe molar teeth, the P3 has a reduced proto-cone as appears to typify the Colobina, butthe P4 has a prominent metaconid. Thedistal lophid of the M3 is typically equal toor narrower than the mesial, being widerthan the mesial in only 42% of the Turkanasample.
There is a partial skeleton associated witha mandible from Koobi Fora. Based on thisspecimen, Rhinocolobus is distinguished fromthe other large colobines by possessing post-cranial adaptations associated with arboreallocomotion (Birchette, 1982; Ciochon,1993). The humeral head is broad, sphericaland higher than the greater tuberosity. Dis-tally, the humerus is characterized by beinganteroposteriorly flat and mediolaterallybroad. The medial trochlear keel is short,the capitulum is relatively spherical, and thezona conoidea is prominent in comparisonto other cercopithecids. The medial epi-condyle is long and projects medially. Onthe proximal ulna, the olecranon is shorterthan that of Paracolobus; it is less retroflexedthan in Paracolobus, and significantly less sothan in Cercopithecoides williamsi from KoobiFora.
Rhinocolobus turkanaensis M. G. Leakey,1982, Type Species(Synonymy as for genus.)
Holotype: NME Omo 75 1969–1012 fromShungura Fm. Lower Mb. G.Distribution: Shungura Fm. A–G (typelocality); Usno Fm; Koobi Fora Fm.
Specific diagnosisAs for genus
cf. Rhinocolobus turkanaensis M. G. Leakey,1982, Type Species(=or including Colobinae gen. et sp. nov. 1.Szalay & Delson, 1979, in part)Specimens included: NME A.L.318–2,A.L.256–1a-c, A.L.248–5, A.L.435–1,A.L.126–31; ?A.L.300–1Distribution: Hadar Fm. Sidi Hakoma–Denen Dora Mbs.
DescriptionRhinocolobus was previously described onlyfrom the Koobi Fora and Omo ShunguraFormations of the Turkana Basin. TheHadar material was initially identified asRhinocolobus by Delson (Szalay & Delson,1979, as Colobinae gen. et. sp. nov.) andlater in several subsequent review articles(Delson, 1984; 1994; Delson et al., 2000).In cranial and dental size, Rhinocolobusis larger than all other known Colobinaeexcept for the two named species of Para-colobus and Cercopithecoides kimeui. Dentaldimensions of cf. R. turkanaensis are given inTable 3.
Maxilla.The maxilla is well preserved in specimensfrom the Omo Shungura and Koobi ForaFormations of the Turkana Basin, but fromthe Afar region there is only a single maxil-lary fragment, A.L.318-2, from unit 2-3s ofthe Denen Dora Member of the Hadar Fm.It is a fragment of a left maxilla from a malewith the first incisor, canine root, and P3 toM1. There is some distortion of this speci-men so that the premolars, canine, andincisors are spread apart more than theywould have been in life. Little of the bony

723
morphology is preserved. The zygoma ispotentially at the distal M1/anterior M2. Thecurvature of the premaxilla, in its mesio-distal arc, is similar to some of theKoobi Fora material, e.g. KNM-ER 4448.This impression may be suspect given thedistortion of this specimen.
Figure 18. Specimens of cf. Rhinocolobus turkanaensis. Anticlockwise from top left: A.L.318-2 lateral view;A.L.318-2 ventral view; A.L.435-1 lateral view; A.L.435-1 medial view; A.L.256-1a–c right lateral view;A.L.256-1a–c dorsal view.
Mandible.The mandible is more completely preservedand represented by several specimens. All ofthose identifiable to sex are male. The mostcomplete, A.L.256-1a-c (Figure 18), is thatof a male reconstructed from three frag-ments, with left I1–P3, M2–3, right I1–M3,but the right M2 lacks its distal portion,from unit SH-u1–s. It is very similar to themandibles Omo 75s �70 C68 and L412-1from the Shungura Formation, but lacks theinferior portion of the corpus. A.L.248-5,also SH-1s-u is an edentulous symphyseal
fragment of a male, which preserves theentire depth of the symphysis. A.L.435-1,from DD 2s-u, (Figure 18) is a right corpusfragment with well preserved M1–3 andapproximately 2·5 cm of the corpus belowthe alveolar process. A.L.126-31 (SH 1s-u)is a right corpus fragment with M2–3 butdoes not preserve much of the corpus belowthe teeth.
The symphysis is robust, and deeper thanthat of Cercopithecoides williamsi, but shal-lower than male specimens of Rhinocolobusfrom the Omo, and Paracolobus. The incisivealveolar process is slightly proclined, inferiorto which the symphysis is quite steep inprofile, then 1·5–2 cm below the alveoli,there is a break in the slope, and the sym-physis extends posteriorly at a shallow angle.This symphyseal shape is typical of Rhino-colobus and Paracolobus. Unlike Rhinocolobusfrom the Turkana basin, there is no median

724 .
mental foramen. Both transverse tori arewell developed, but the inferior one extendsfurther posteriorly than the superior. Thereare distinct but slight corpus fossae presenton A.L.248-5, just anterior to the mentalforamen, which lies inferior to the P4.A.L.256-1, A.L.126-31, and A.L.435-1 allshow that the corpus was not broad and hada narrow extramolar sulcus and weaklydeveloped oblique line. While the margin isnot preserved, A.L.435-1 preserves enoughof the corpus to show that the fossae did notextend far posteriorly, and that the corpuswas deeper than is the case for Cercopithe-coides. All of this is consistent with Rhino-colobus from the Turkana basin, except forthe lack of a median mental foramen.
Dentition.The upper dentition is preserved only onA.L.318-2. The upper central incisor israther small, given the size of the molars, butits morphology is normal for colobines, andsimilar to others of this species from KoobiFora. Its crown does not flare towardsthe apex, but is approximately the samemesiodistal length at the alveolus as at thetip. There appears to have been a lingualcingulum present. Of the upper canine,most of the crown is missing, but its cervicalarea and root are preserved. Its morphologyis typical of male cercopithecids, being largerelative to the other teeth, with a deep mesialsulcus. The P3 is present, but its crown isdamaged so that little morphology can beobserved. The P4 is damaged so that theprotocone is missing, but the paracone ispreserved. The paracone is tall as would beexpected of a colobine, and there is whatappears to be a well-developed transverseloph. The M1 is the only upper molar pre-served. The hypocone is damaged, and thedistolingual corner is missing, but the toothis otherwise intact. It is typically colobine,with a large amount of cusp relief, a lownotch, and sharp transverse lophs. Thecrown also has only a low amount of flare.
The lower incisors, canines, and pre-molars are preserved only on A.L.256-1a,where they are heavily worn. The incisorsare typical of colobines, being relativelysmall peg-like teeth. Enamel is clearlypresent on the lingual surface of the teeth.The lower central incisors are small, but aretoo worn to preserve any details of theircrown morphology. The lower lateralincisors are also quite worn, but some detailscan be seen. There is a small distal cuspuleor ‘‘lateral prong’’ present. The crownwould have been narrow and tilted mesially.The lower canines are typical for malecercopithecids, being very large relative tothe other teeth. Though both are missing theapical parts of their crowns they would havebeen tall fairly conical teeth, but with amesial groove, and distal tubercle.
The P3 is a highly sexually dimorphictooth in cercopithecids. The protoconid istall and conical. The mesiobuccal honingflange is long compared to female speci-mens, but shorter than that of a cercopithe-cine male. It is also relatively anteriorlyoriented. There is more of a paraconiddeveloped than would be the case on acercopithecine. The talonid is also large andwell developed. The only preserved P4 isrelatively complete but is missing the enamelon the lingual side. The cusps are highrelative to the well-developed talonid. Themetaconid is similar in size to the protoco-nid. There is even slight development of aparaconid. The mesiodistal axis of the toothis in line with the molar row.
The lower molars are represented on afew mandibles that are assignable to thisspecies. A.L.256-1 preserves the right M1–3
and left M2–3, A.L.126-31 preserves theright M2–3, and A.L.435-1 preserves the leftM1–3. In general the lower molars are typicalof the colobines with high cuspal relief,sharp cross-lophids, and deep lingualnotches. The crowns show very little basalflare, and the cusp tips are widely spaced.The teeth are very ‘‘clean’’ lacking extra

725
cuspules. The M1–2 have the typicalcolobine pattern of the distal lophid beingwider than the mesial. In the two measur-able M3‘s the distal loph is equal to themesial on A.L.126-31 and wider onA.L.435-1. This may hint at colobinanaffinity for this species. The buccal cusps arerelatively columnar or ‘‘pinched’’, which isfairly common among the colobines. Thetuberculum sextum is very small or absenton A.L.126-31 and A.L.256-1, but larger onA.L.435-1.
Postcrania.No postcrania are directly associated withany of the dental material, but there is asingle distal fragment of a right humerus,A.L.300-1 which is likely to represent thistaxon (Figure 9). It is larger than and mor-phologically distinct from the T. o. cf. dartidistal humeri, which predominate in theHadar sample. Morphologically it is verysimilar to the distal humerus from KoobiFora assigned to this taxon, and discussedby Birchette (1982) in his description of P.chemeroni. If the Koobi Fora allocations arecorrect, Rhinocolobus has a humerus that isflattened anteroposteriorly, an extended andmedially oriented medial epicondyle, and anarticular area that is narrow relative tobiepicondylar breadth. A.L.300-1 is similarto the Koobi Fora material in all of theserespects. The medial epicondyle is retro-flexed at an angle approximately 38�, whichis similar to extant African colobines,KNM-ER 1542 and A.L.363-12, and muchless than KNM-ER 4420, Cercopithecoidesmeaveae and the T. o. cf. darti sample(Figure 11). Relative to overall biepicondy-lar breadth the medial epicondyle projectionis large, being within the range of most ofthe arboreal colobines, as well as Lophocebusalbigena and Cercopithecus mitis (see Figure10). Overall, the distal humerus A.L.300–1is quite broad mediolaterally and flat antero-posteriorly. Also similar to the Koobi Forahumerus, the zona conoidea is prominent
and the medial trochlear flange relativelyshort, and not sharp. The length of theflange, relative to distal articular breadth isshorter than in Semnopithecus entellus, but isgenerally in the upper part of the range forarboreal colobines, and in the lower part ofthe range of more terrestrial cercopithecines(see Figure 12). Ciochon (1993), in amultivariate morphometric analysis of cer-copithecid forelimbs, allocated this speci-men to T. brumpti. A.L.300-1, however,lacks several features typical of T. brumptisuch as an extended medial trochlear flangeand retroflexed medial epicondyle (Krentz,1992, 1993). Furthermore, this assignmentseems unlikely given the absence of anycranial remains of this species outside of theTurkana Basin.
Genus Cercopithecoides Mollet, 1947(=or including Parapapio Jones, 1937:Broom, 1940; Broom & Robinson, 1950;Freedman, 1957, in part. Brachygnatho-pithecus Kitching, 1952, in part. Colobinaecf. Cercopithecoides Leakey & Leakey, 1973.cf. Colobinae gen. et sp. nov. A: Eck, 1976;1977; Colobinae gen. et sp. indet. smallerSzalay & Delson, 1979)Type species: Cercopithecoides williamsiMollet, 1947Other included species: C. kimeui Leakey,
M. G. 1982; C. meaveae sp. nov.; C. sp.nov. Leakey et al., 2002.
Generic diagnosisThe diagnosis for this genus is little changedfrom Szalay & Delson (1979) and theemended diagnoses of Leakey (1982) andFreedman (1957). Medium to very largecolobines with globular, rounded calvaria.The rostrum is short in comparison toneurocranial length, very different fromParacolobus, Rhinocolobus, Dolichopithecus,and Nasalis. It is not as short as that ofRhinopithecus, however. The frontal processof the zygomatic bone is narrow, unlike

726 .
Paracolobus and Rhinocolobus. The inter-orbital region is broad, which is distinctfrom Libypithecus, Rhinocolobus, and Nasalis.The supraorbital tori are thick and separatedfrom the calvaria by a deep ophryonicgroove, which is distinct from Colobus, andmost species of Presbytis and Trachypithecus.The calvaria itself lacks a sagittal crest, atleast anteriorly, which separates it fromParacolobus and Procolobus. The P3 lacksa protocone, which is similar to modernAfrican colobines, but different fromLibypithecus, Kuseracolobus, Paracolobus andRhinocolobus.
The mandibular symphysis is steep, butshallow and pierced by a median mentalforamen (except in C. meaveae), which isdifferent from all colobines other thanProcolobus (Procolobus) and Rhinocolobusfrom the Turkana Basin. The mandibularcorpus is shallow and thick, which contrastsgreatly with that of Kuseracolobus, Rhinocolo-bus, Paracolobus and to some degree Colobus,but is not unlike that of Procolobus (Piliocolo-bus). The gonial region is unexpandedor expanded only a small amount. Thiscontrasts with mandibles of Kuseracolobus,Rhinocolobus, Paracolobus mutiwa, andColobus.
Associated postcrania are known for C.williamsi from an associated skeleton fromKoobi Fora (the morphology of which maynot be typical for the South African material,see remarks below). It is distinguishedfrom all other known colobines, exceptDolichopithecus, in the degree of its adapta-tions for terrestrial habitus (Birchette, 1981;1982; Ting, 2001). The associated post-crania of C. meaveae also show adaptationsfor terrestrial habitus, as are describedbelow.
RemarksIn addition to Cercopithecoides meaveae,which is described below, Cercopithecoidescurrently has two named species, with a
third in press. The best known is the typespecies, C. williamsi, which has been col-lected from many Pliocene and Pleistocenesites in South Africa and Angola, and alsohas been recognized by Leakey (1982) atKoobi Fora. A second larger species, C.kimeui is known from Olduvai Gorge, KoobiFora (Leakey & Leakey, 1973b; Leakey,1982), Rawi (Ditchfield et al., 1999; Frostet al., in review), and Hadar. Finallya species significantly smaller than theprevious two has been recognized fromLothagam. It is of uncertain stratigraphicprovenance, but may be from the ApakMember of the Nachukui Formation(Leakey et al., in press).
There is some question as to whether C.williamsi from Koobi Fora is conspecificwith the material from Southern Africa. AtSterkfontein, Kromdraai, Swartkrans, andBolt’s Farm C. williamsi is the only colobineknown in the cranio-dental sample. Thereare also postcranial remains from these sites,identified by Ciochon (1993) as those ofa large colobine, which lack the extremeterrestrial adaptations of East African C.williamsi. If this material indeed representsC. williamsi, and its morphology is typical ofthe South African species, then the EastAfrican material is unlikely to represent thesame species. If so, a new name will berequired for at least KNM-ER 4420.
There is also considerable variation withinand among the populations represented atMakapansgat, Sterkfontein, Kromdraai, andHasgaat, both in size and cranial mor-phology. The Makapansgat sample aloneshows a large amount of cranial variability[compare the male crania UWMA MP113(=M2999) and BPI M3055, figured byFreedman (1961) and Maier (1970)respectively].
Freedman (1957) had originally recog-nized the Cercopithecoides material fromSwartkrans as a separate species, C. molletti,based on its larger dental size. However,based on increased variation shown by

727
additional specimens from Makapansgathe later synonymized this material with C.williamsi (Freedman, 1961). Delson (1984)has also recognized the material fromKromdraai along with that from Swartkrans,as a ‘‘large variant’’ of C. williamsi. If thesepopulations are distinct, then the nameC. molletti (Freedman, 1957) is available. IfKromdraai alone is distinct it would requirea new name. The purpose of this paper isnot to revise the systematics of Cercopithe-coides, and therefore C. meaveae is, for now,included in Cercopithecoides as currentlyrecognized.
Cercopithecoides kimeui, Leakey M. G. 1982(=or including Cercopithecoides sp. nov.:Leakey & Leakey, 1973. cf. ColobinaeLeakey & Leakey, 1973. Cercopithecoides sp.2: Szalay & Delson, 1979. ?Cercopithecoidessp(p): Szalay & Delson, 1979, in part. Papio/Parapapio sp. Ditchfield et al., 1999)Holotype: NMT 068/6514 from MLKOlduvai Gorge (Middle Bed II)Specimens included: NME A.L.603–1;?A.L.577–1.Distribution: Hadar, Pinnacle locality;
Koobi Fora Formation KBS and OkoteMembers; Rawi Formation; OlduvaiGorge, Middle Bed II (type locality), ?BedIII; (and possibly the Lokochot and TuluBor Mbs of the Koobi Fora Fm.).
Specific diagnosisThis diagnosis follows that of Leakey(1982). A species of Cercopithecoides largerthan C. williamsi (and far larger than C.meaveae and C. sp. nov. from Lothagam).The mandibular corpus is broader and morerobust than that of C. williamsi, and haslarge prominentia laterales. The upper molarsare unique among the Colobinae, in beinglow-crowned. They are also quite broad andquadrate in outline, and show more flarethan do other colobine molars.
DescriptionThe best Afar specimen, and the only oneassignable to this species with any confi-dence, is A.L.603–1a (Figure 19), a partialskull of a female from the Pinnacle locality.It is very similar to the partial craniumKNM-ER 398 from Koobi Fora. Althoughbadly weathered, most of the cranium andmandibular corpus are preserved, and thedescription below is based entirely on thisspecimen. The neurocranium is essentiallycomplete, but the brow ridges are damagedlateral to the interorbital pillar. The left sidepreserves most of the orbit, except for someof the rim on the superior half. The rightzygomatic bone is largely missing, but on theleft it is preserved anterior to the frontalprocess. The face is otherwise largely com-plete. The basicranium is obscured bymatrix and a little damaged as well, butsome details can be discerned. The com-plete, but damaged and weathered, man-dibular corpus is present, but both rami arelacking. Three isolated teeth associated withthis specimen fit back onto their roots:the right M2–3 and the left M3. Also fromPinnacle is a distal fragment of a largehumerus, A.L.577-1. This specimen is froma very large cercopithecid, appears to showsome colobine affinities, and is distinct in itsmorphology from Theropithecus humeri. Infact, it is slightly larger than most of thehumeri assigned to this latter taxon. It ispotentially from C. kimeui, but no humeriare known for this species with which tocompare this specimen.
This is a very large cercopithecid by anymeasure. The female cranium from Hadar isin most measurements similar in size to themale holotype of Rhinocolobus turkanaensis,except that the face is smaller. The face isalso smaller than that of the holotype ofParacolobus chemeroni. Overall, it is similar tothe female crania of C. kimeui (ER 398 andER 991) from Koobi Fora. The calvaria is alittle smaller than the male from Olduvai.The face is substantially smaller than the
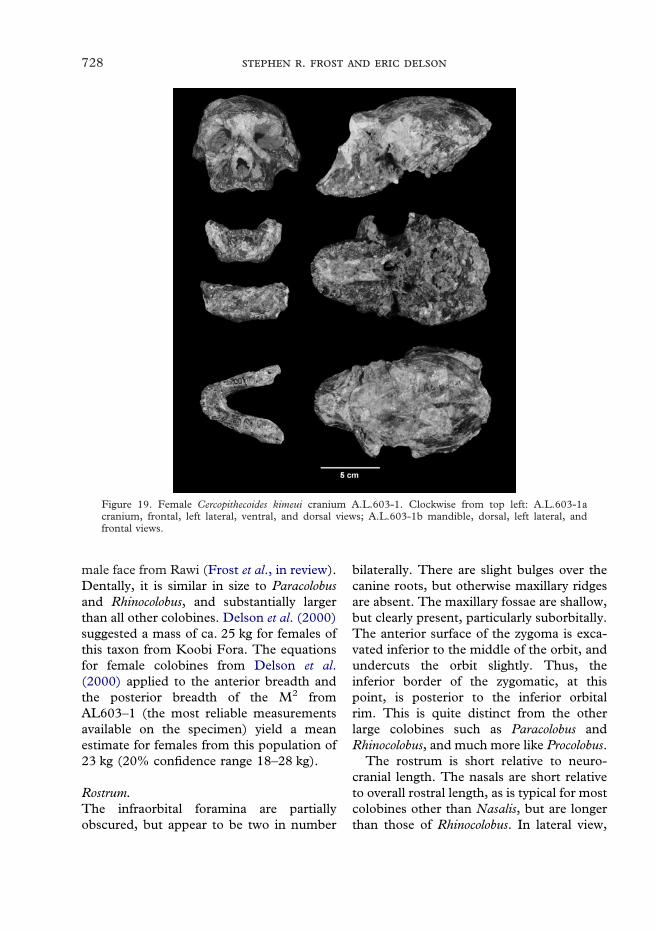
728 .
male face from Rawi (Frost et al., in review).Dentally, it is similar in size to Paracolobusand Rhinocolobus, and substantially largerthan all other colobines. Delson et al. (2000)suggested a mass of ca. 25 kg for females ofthis taxon from Koobi Fora. The equationsfor female colobines from Delson et al.(2000) applied to the anterior breadth andthe posterior breadth of the M2 fromAL603–1 (the most reliable measurementsavailable on the specimen) yield a meanestimate for females from this population of23 kg (20% confidence range 18–28 kg).
Figure 19. Female Cercopithecoides kimeui cranium A.L.603-1. Clockwise from top left: A.L.603-1acranium, frontal, left lateral, ventral, and dorsal views; A.L.603-1b mandible, dorsal, left lateral, andfrontal views.
Rostrum.The infraorbital foramina are partiallyobscured, but appear to be two in number
bilaterally. There are slight bulges over thecanine roots, but otherwise maxillary ridgesare absent. The maxillary fossae are shallow,but clearly present, particularly suborbitally.The anterior surface of the zygoma is exca-vated inferior to the middle of the orbit, andundercuts the orbit slightly. Thus, theinferior border of the zygomatic, at thispoint, is posterior to the inferior orbitalrim. This is quite distinct from the otherlarge colobines such as Paracolobus andRhinocolobus, and much more like Procolobus.
The rostrum is short relative to neuro-cranial length. The nasals are short relativeto overall rostral length, as is typical for mostcolobines other than Nasalis, but are longerthan those of Rhinocolobus. In lateral view,

729
the profile is relatively vertical and straightfrom glabella to prosthion, with most of therostral length being distal to rhinion, similarto the profile of Colobus. As in the C.meaveae, but different from most Colobus,rhinion is comparatively prominent. Thealveolar process shows a strong normalcurve of Spee, as seems to be commonamong colobines.
The premaxillae are relatively small(partly because the incisor roots are quitesmall) being short, and rounded in superior-view. In general, females have more roundedpremaxillae than do males. However, thepremaxillae of A.L.603-1a are different fromthose of females of Kuseracolobus, mostColobus, and some C. williamsi, which aremore squared in superior view. The nasalprocesses of the premaxillae form the lateralborders of the piriform aperture and extendsuperior to its superior border, and thenterminate lateral to the nasals, considerablyinferior to nasion. They do not contactthe frontal as in Presbytis, Trachypithecus,and some Colobus and Procolobus. Thepremaxillo–maxillary sutures follow a fairlystraight course from their termination at thenasals to their inferior limit mesial tothe canine. They bulge slightly laterallyaround the piriform aperture, maintaining adistance of about 2–3 mm from its rim.
Relative to its height, the piriform aper-ture is broader than those of most colobines,but narrower than those of most papionins.It is oval in outline, but its inferior polecomes to a sharp angle at nasospinale. Inlateral view, the piriform aperture is inclinedat an angle of approximately 45� to theocclusal plane.
While most of the teeth are absent, thealveolar process is preserved and the shapeof the dental arcade can be observed. Thecheek teeth form straight rows from M3 toC1, with lateral borders that bulge slightly,being widest at M1–2. The alveolar processesare wide overall, to accommodate the broadmolars. The incisors form a flat arc from
side to side. This yields a dental arcade thatis ‘‘U’’-shaped, being somewhat less para-bolic than in many smaller colobines. Thismay in part be due to the relatively broadsquare molars. The palate is fairly long andsquare in outline. It is relatively deep com-pared to most colobines, and deepensposteriorly. The exact depth is difficult todetermine as much of the palate is coveredin matrix, but at least 1 cm of depth can beobserved near the M3.
Zygomatic arch.The zygomatic process of the maxilla liessuperior to the M2. This position is similarto that of the females of C. kimeui fromKoobi Fora and is relatively posterior com-pared to other colobines. It is further pos-terior than in any specimens of C. williamsi,and similar to the female of Rhinocolobusturkanaensis KNM-ER 1485. Only Paracolo-bus chemeroni, P. mutiwa and the male holo-type of Rhinocolobus have a zygoma that ismore posteriorly positioned. On the otherhand, it is more anteriorly positioned inA.L.603–1a than is the case in most ofthe larger papionins, such as Papio andMandrillus.
When viewed anteriorly, the zygomataare shallow. The inferior margin curvessmoothly upward from its origin and reachesa maximum height under the lateral part ofthe orbit, then on the same curve it con-tinues downward. This point of maximumheight of the inferior border also corre-sponds with the portion of the zygoma thatmost undercuts the orbit. While the zygo-matic arches are absent posterior to thefrontal process, what can be seen of them onthe left side shows them to be swept back insuperior view.
Orbital region.Both brow ridges and the lateral portions ofthe orbits are missing. Only the medial 2 cmof the supraorbital torus is present. This

730 .
portion of the torus is thick, projecting androbust, and is separated from the neurocra-nium by a wide ophryonic groove. Althoughmuch of the orbits are damaged, they prob-ably were broader than tall, and were likelyto have been slightly laterally oriented. As istypical of the Colobinae, the interorbitalregion is broad, even taking the overall sizeof the specimen into account. Glabella isquite prominent and projects anterior tonasion.
Calvaria.The neurocranium is globular in overallappearance, and is relatively long and nar-row in superior view. In Frankfurt Horizon-tal, the frontal bone rises from the ophryonicgroove, in a smooth arc, and reaches itsmaximum height at approximately bregma.The temporal lines do not form a sagittalcrest, but remain separated by at least 2 cm.They are well marked anteriorly, but fadeposteriorly. The nuchal crests are damagedbilaterally, but it can be seen that they werewell developed and reached nearly 1 cm inheight on the right side. In superior view, thewidest part of the neurocranium is at thelevel of the auditory meatus, but it is notsignificantly wider than the rest of the vault,in part because postorbital constriction isminimal.
Basicranium.The basicranium is reasonably well pre-served, but is partially obscured by adheringmatrix. The occipital plane is inclined atapproximately 45� relative to FrankfurtHorizontal. The mastoid processes are smalland pyramidal in shape, and the digastricgroove is shallow and wide. The postglenoidprocess is tall and broad, and separated fromthe glenoid fossa by a wide groove. Theglenoid fossa is shallow and flat.
Facial hafting.Facial hafting is similar to that in Colobusand other Cercopithecoides. The glenoid fossa
is only slightly above the alveolar plane,thus the face is not deep as in Theropithecus.The face is less airorhynchous than that ofParacolobus chemeroni, and much less so thanin Rhinocolobus.
Mandible.The mandibular corpora are largely pre-served but heavily weathered. The rami andgonial areas are totally lacking. While themandible is edentulous, it does possess theroots for the entire dentition. The symphysisis relatively shallow, but has a nearly verticalprofile. Its anterior surface is pierced by amedian mental foramen. This trait is rareamong colobines, but occurs in Cercopithe-coides (except for C. meaveae), Rhinocolobusfrom the Turkana Basin, and P. (Procolo-bus). On the genial surface, both transversetori are present, but the inferior is partiallyobscured by some adhering matrix. Thesuperior surface of the superior transversetorus extends posteriorly to the distal part ofthe P3, and the inferior back to the mesialpart of the P4.
As with other mandibles allocated toCercopithecoides, the corpus is thick androbust, but shallow, being deepest under M1
then shallowing posteriorly. The lateral sur-face of the corpus has a very shallow fossa,largely due to the large prominentia lateralesthat are present. These actually form thedeepest portion of the corpus in lateral view,which is below the M1. The mental foramenis obscured by matrix.
Dentition.The only teeth preserved with A.L.603–1are the right M2–3 and left M3. The uppermolars are distinctive from those of othercolobines. They are quadrate in occlusalview, being short, broad, and low crowned.Even though the teeth are low crowned, arelatively large amount of crown height ismade up by the cusps above the buccalnotch, whereas in papionins most of thecrown height is below the level of the buccal

731
notch. Also, unlike papionin molars, thecusps are relatively widely spaced, and thecrowns are less flaring buccally. The M2 islarger than the M3. Dental measurementsare given in Table 4.
In addition to the three molars preserved,all of the roots are present, and a few com-ments can be made about the relative sizesof the other teeth. The incisors would havebeen small relative to the cheek teeth. Thecanine roots are small, which is the basis forthe diagnosis of this specimen as female.The premolars are large, and the P4 is largerthan the P3. The P3 mesiobuccal flange isshort, which is also consistent with thisspecimen being a female. The P4 is broaderthan the P3. The molars are large, and theM2 is the largest of the upper molars. Forthe lower molars, the first is smallest, andthe third is the largest.
Postcranium.There are no postcrania that are definitelyassociated with the cranium. However, anisolated left distal humerus, A.L.577-1,which is also from the Pinnacle site, may beassignable to this taxon (Figure 9). It is avery large humerus, similar to the largestspecimens of Theropithecus oswaldi in size. Italso has a medial trochlear flange that issharp and well developed, longer than thoseof most colobines, but within the ranges ofS. entellus, D. ruscinensis, and M. pentelicus. Itis shorter than the most Theropithecus orPapio, but is in the lower end of their vari-ation. Unlike most distal humeri allocated toT. o. oswaldi it has an articular area thatis narrow relative to total biepicondylarbreadth, a very prominent zona conoidea,and a tall radial fossa, all of which arefeatures that may be associated with colob-ines (Olivier & Caix, 1959; Delson, 1973).The medial epicondyle is stout, large andretroflexed. These features are generallyassociated with more use of terrestrial sub-strates during locomotion. In overall appear-ance it is similar to A.L.2-64, the distal
humerus associated with the C. meaveae, butmuch larger.
Cercopithecoides meaveae sp. nov.(=or including Colobinae gen. et sp. indet,smaller, Szalay & Delson, 1979, in part.Colobinae gen. et sp. nov. A. Eck, 1976,1977, Delson, 1984, 1994, in part.)Etymology: Named in honor of Dr MeaveG. Leakey, National Museums of Kenya, forall of her contributions to the study of fossilcercopithecoids, especially colobines, and toAfrican paleontology in general.Holotype: NME A.L.2–34 (and associatedelements listed in Table 5) a partial skeletonfrom Leadu, which is of uncertain chrono-logical position, but possibly dates between2·5 and 3 Ma.Specimens included: NME A.L.231-1a,A.L.249-23, ?A.L.222-14, A.L.660-2Distribution: Leadu (type locality); HadarFm., Sidi Hakoma Mbr.
Specific diagnosisA medium-sized species of colobine, mostlikely assignable to Cercopithecoides, similarto C. sp. nov. from Lothagam, Kuseracolobusaramisi and larger individuals of Semno-pithecus entellus shistacea in overall size. It issmaller than C. williamsi and substantiallysmaller than C. kimeui. The glabellar regionand supraorbital torus are both prominent,but less so than is the case in C. williamsi.The supraorbital rim is separated from thecalvaria by a sulcus, which is not as deep asthat of C. williamsi. The mandibular sym-physis lacks a median mental foramen,unlike C. williamsi and C. kimeui, but similarto the new species from Lothagam. Thesymphysis is distinct from that of theLothagam species in being considerablymore shallow and less sloping, as well as lessflattened across the face of the symphysis.The inferior margin of the mandibularcorpus is not thickened, and does not havestrongly developed prominentia laterales.

732 .
Dentally, the upper molars are high crownedwith well developed lophs, unlike the molarsof C. kimeui. The molars show none of theunusual wear pattern of C. kimeui and someC. williamsi (e.g., UWMA BF 43 andDGUNL LEBA01). The postcrania, in par-ticular the shoulder, elbow, and hip, appearto show similar, though less extreme, adap-tations for terrestrial locomotion to those ofKNM-ER 4420.
Figure 20. Cercopithecoides meaveae. A.L.2-34+A.L.2-86 (holotype male) and A.L.660-2 maxilla(unknown sex). Top: A.L.2-34 dorsal vew. Middle Row: A.L.2-34/86 frontal view, A.L.660-2 ventralview, and A.L.2-34 ventral view. Bottom: A.L.2-34/86 right lateral view.
DescriptionThis description is based upon the holotypeassociated partial skeleton from Leadu(A.L.2-34 and associated elements listed inTable 5) near Hadar (Figures 20–24), whichrepresents a single adult male individual,and upon four specimens from the SidiHakoma Member of the Hadar Formation.These are a nearly complete mandible(A.L.231-1a, from SH 2s-u) (Figure 21), an

733
isolated P3 (A.L.249-23, from SH 1), a rightmaxillary fragment with M1–3 (A.L.660-2,from SH 1-2) (Figure 20), and a distalfragment of a left humerus (A.L.222-14,from SH 1-3s) (Figure 9). C. meaveaeis larger in size than Colobus, Procolobus,Libypithecus and Mesopithecus, but smallerthan Paracolobus, Rhinocolobus and Dolicho-pithecus. It is similar in size to the largestSemnopithecus, and to Kuseracolobus. Kusera-colobus is similar to A.L.2-34 in those fewaspects of the face that are preserved, but itis considerably different in the morphologyof its mandible and postcranium (Frost,2001a). Other colobines of similar size areknown from below the Sidi Hakoma Tuff atWee-ee and Matabaietu in the MiddleAwash (Frost, 2001b), from the OmoShungura Formation in Members B,C,D
and G (Eck, 1976; Leakey, 1987), from theTulu Bor Member at Koobi Fora (Leakey,1976; Delson, 1984), and from the UpperLaetolil beds (Leakey & Delson, 1987).These fossils are only isolated teeth, andtwo postcranial elements, so their specificstatus is indeterminate at this time. Dentaldimensions for Cercopithecoides meaveae aregiven in Table 6.
Figure 21. Mandibles of Cercopithecoides meaveae. Left column, top to bottom: A.L.231-1a dorsal, lateral,and medial views. Right column, top to bottom: A.L.2-34 dorsal, lateral and frontal views.
Rostrum.Overall, the muzzle is shorter relative tocranial size than that of Libypithecus, Rhino-colobus, Paracolobus, and Dolichopithecus butlonger than that of Colobus, being similar inproportion to that of Cercopithecoides. As istypical for the subfamily, it lacks maxillaryfossae and ridges. The muzzle is highbut rounded in cross-section, describing a

734 .
Figure 22. Partial skeleton of Cercopithecoides meaveae from Leadu. Elements are listed in Table 5.
generally smooth parabola. In lateral view,the profile is steep from glabella to nasion,with glabella being only slightly prominent,and curving in a smooth, concave-up arc torhinion followed by a slightly concave slopefrom rhinion to prosthion. When viewedsuperiorly, the muzzle is fairly short andsquared in outline, not unlike that of otherspecies of Cercopithecoides.
The nasal bones are short and consider-ably broader inferiorly than superiorly,where they are quite narrow. Relative to the
overall length of the muzzle, the nasals areshorter than those of Nasalis (including N.(Simias)), and longer than those of Rhino-colobus, Paracolobus and Rhinopithecus, butotherwise similar to those of most colobines.The nasal process of the premaxilla extends1·5 cm beyond the superior limit of thepiriform aperture as a thin sliver to the baseof the interorbital pillar. The premaxillo–maxillary suture runs as a fairly smooth arcfrom this point to the alveolar process, beingnearly straight in both anterior and lateral
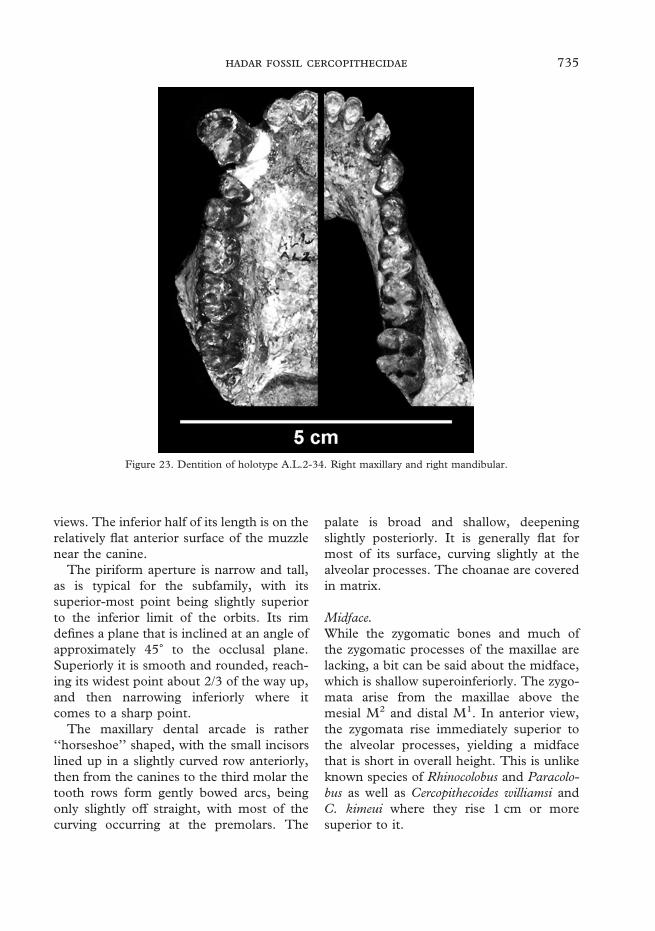
735
views. The inferior half of its length is on therelatively flat anterior surface of the muzzlenear the canine.
The piriform aperture is narrow and tall,as is typical for the subfamily, with itssuperior-most point being slightly superiorto the inferior limit of the orbits. Its rimdefines a plane that is inclined at an angle ofapproximately 45� to the occlusal plane.Superiorly it is smooth and rounded, reach-ing its widest point about 2/3 of the way up,and then narrowing inferiorly where itcomes to a sharp point.
The maxillary dental arcade is rather‘‘horseshoe’’ shaped, with the small incisorslined up in a slightly curved row anteriorly,then from the canines to the third molar thetooth rows form gently bowed arcs, beingonly slightly off straight, with most of thecurving occurring at the premolars. The
palate is broad and shallow, deepeningslightly posteriorly. It is generally flat formost of its surface, curving slightly at thealveolar processes. The choanae are coveredin matrix.
Figure 23. Dentition of holotype A.L.2-34. Right maxillary and right mandibular.
Midface.While the zygomatic bones and much ofthe zygomatic processes of the maxillae arelacking, a bit can be said about the midface,which is shallow superoinferiorly. The zygo-mata arise from the maxillae above themesial M2 and distal M1. In anterior view,the zygomata rise immediately superior tothe alveolar processes, yielding a midfacethat is short in overall height. This is unlikeknown species of Rhinocolobus and Paracolo-bus as well as Cercopithecoides williamsi andC. kimeui where they rise 1 cm or moresuperior to it.

736 .
Orbital region.Relatively little of the orbital region is pre-served. The orbital rim is separated from theneurocranium by a broad ophryonic groove.What is preserved of the supraorbital rim isfairly thick superoinferiorly. The interorbitalpillar is broad, as is expected for a colobine,but it is broader in comparison to overallcranial size than that of Nasalis, Rhinocolobusand Libypithecus. The lacrimal bone makesup the posterior half of the lacrimal fossawith the maxilla forming its anterior rim,which is also the orbital rim, so that the fossalies entirely within the orbit. Overall, theorbits themselves are rather large and tallrelative to the size of the face, but theirbreadth is not preserved.
Endocast and facial hafting.The natural partial endocast, A.L.2-35,preserves much of the front of the brain. It
fits tightly with the face, and allows anestimate of the relative sizes of the neuro-cranium and face. The braincase would havebeen relatively large compared to the face, asin Colobus, Procolobus, and other species ofCercopithecoides. It is comparatively smallerthan that of Libypithecus, Paracolobus andRhinocolobus. There is enough preserved totell that the neurocranium was not long andnarrow as in Cercopithecoides kimeui, butmore rounded in superior view. The occlu-sal plane was likely only slightly inferiorto the glenoid fossa and basicranium, due tothe relatively low facial height.
Figure 24. Forelimb elements of Cercopithecoides meaveae. Clockwise from upper left: distal humerusA.L.2-64, proximal humerus A.L.2-63, proximal radius A.L.2-66, proximal ulna A.L.2-65.
Mandible.There are two well-preserved mandibles.A.L.2-34 is a corpus and complete denti-tion. The inferior margin is intact from theleft M3 to the right M2. The right ramus islacking, but the left is partially preserved.

737
None of the gonial region remains, however.A.L.231-1a is a nearly complete male man-dible, in left and right halves. It preservesmost of the margin on the left, except for inthe gonial area, and on the right it preservesfrom below M1 back to gonion, except for asmall piece just anterior to gonion. Bothrami are largely intact. Of the dentition, theleft C1–M3 and right I2, P3–M3 are preserved.
The symphysis is squared and vertical inprofile, but shallow. It is considerably shal-lower than that of Kuseracolobus. The sym-physeal profile is distinct from the Lothagamspecies of Cercopithecoides, which is consid-erably deeper, more sloping, and extendsmore posteriorly than does that of C.meaveae. The incisive alveolar process formsa very slightly curving arc. As is typical of thesubfamily, there is no median mentalforamen. This is unlike the case in Rhino-colobus from the Turkana Basin (but notHadar) and Cercopithecoides from SouthAfrica and Kenya. Both transverse tori arerepresented, and the plenum alveolare isshort and steep, extending back only todistal P3.
The corpus is shallow and broad,especially when compared to Rhinocolobus,Paracolobus, and Kuseracolobus. Corpus fos-sae are absent, as are prominentia laterales.The corpus bulges laterally in the middle ofits height, like that of other species of Cerco-pithecoides, and unlike Paracolobus chemeroniand Kuseracolobus, which are widest near theinferior margin due to large prominentia lat-erales. In lateral view, the corpus is generallyshallow, and relatively even in overall depththroughout its length, but with a bulgeapproximately under the M1. The mentalforamen is double bilaterally in A.L.2-34and single bilaterally in A.L.231-1a, and liesinferior to the P3/P4 contact and P4 respec-tively. The gonial region is only minimallyexpanded.
In superior view, the oblique line mergesin with the corpus at about M1–2. The wholecorpus is quite broad, with a wide extra-
molar sulcus present. On the medial side ofthe corpus, inferior to the M3 there is noridge. The ramus is vertical, but dorso-ventrally low and anteroposteriorly long.This is similar to mandibles of other speciesof Cercopithecoides, but unlike those ofRhinocolobus and Paracolobus and is whatwould be expected from the short midfaceand low glenoid fossae relative to thealveolar plane. There is only a shallow trian-gular fossa, and the coronoid process ishigher than the condyle. Laterally on theramus, the anterior edge of the masseterictuberosity is not marked by a ridge, but doesbulge laterally.
Dentition.The incisors are small and peg-like. Theupper incisors are only preserved on A.L.2-34, and their crowns preserve well-developed lingual cingula. The I1 crown isspatulate and not significantly larger thanthe I2. Its crown is widest near the cervix,then remains nearly even in width through-out its remaining height. The I2 is a morecaniniform tooth that is substantially widerat the cervix than it is at its apex. Its crownis also angled mesially, when viewedanteriorly. The lower incisors are small andhave nonflaring crowns with enamel on theirlingual aspects. The I2 is narrower than theI1, its crown is slightly mesially angled, andit has a well developed ‘‘lateral prong’’. Theupper canine crowns are large, which is thebasis for the identification of this specimenas male. The lower canine is best preservedon A.L.231-1a, but also fragmentarily onA.L.2-34. They are typical of male individ-uals, being large in caliber. The root has adistinct mesial sulcus in A.L.231-1a, butthis feature is unobservable in A.L.2-34 dueto damage. The canine seems to lack thedisto-buccal tubercle of the Kuseracolobus,having only a distal cingulum.
The upper premolars are bicuspid teeth,as is typical for the family. The P3 has aslight mesiobuccal flange, and the protocone

738 .
is greatly reduced, but not completelyabsent. The P4 has a well-developed proto-cone, no flange, and a more strongly devel-oped talon, giving a more quadrate outlinein occlusal view. As is expected in a male,the P3 has a fairly long mesiobuccal flange.The paraconid projects slightly from thesuperio-anterior surface of the mesiobuccalflange producing a distinct change in slopebetween the protoconid and the end of themesiobuccal flange, much as in Kusera-colobus. The protoconid is tall and projectsabove the crowns of the other teeth. Distally,the talonid bulges medially. The P4 has ashort mesiobuccal flange as well, which islonger in A.L.2-34 than in A.L.231-1a. Itscrown is set in line with the molar tooth row.On both individuals, the metaconid is welldeveloped. Laterally, there is a bit of ashelf to the buccal cleft as in the molars.The talonid is also well developed in bothindividuals, but there are no accessorycusps.
The upper molars are preserved in bothA.L.2-34 and A.L.660-2. As is typical ofcolobines, the molars have crowns withminimal basal flare, widely spaced cusps, alarge amount of cuspal relief, and sharpcross-lophs. The upper molars are all similarin size, but they do increase in size distally.The lower molars have tall cusps, and lowlingual notches. They are basically the sameon both mandibles, except that the M3 ofA.L.231-1a lacks a tuberculum sextum andhas a more reduced hypoconulid than is thecase on A.L.2-34, which has a small tuber-culum sextum. The molars increase in sizefrom front to back, and the distal lophid ofthe M2 is wider than the mesial lophid. Themesial and distal lophids of the M3 aresub-equal, but the distal may be a bit wider.The molar rows are fairly straight, butdiverge slightly distally.
Postcrania.Several elements of the axial skeleton, fore-limb, and hindlimb are associated with the
cranial material from Leadu. This partialskeleton is one of the better specimens in allof the African Pliocene, and it is certainlythe most complete single cercopithecid indi-vidual from Ethiopia. The elements fromthis partial skeleton are described belowunder the individual regions. The only post-cranial element that may represent thistaxon, other than the partial skeleton fromLeadu, is a distal fragment of a humerus:A.L.222-14, from the Sidi Hakoma Memberin the Hadar Formation. It is not directlyassociated with any cranial material, but istentatively assigned to this taxon basedon its similarity to the distal humeral frag-ment A.L.2-63, associated with the face andmandible from Leadu.
Vertebrae.One thoracic, five lumbar, and twelve caudalvertebrae are preserved (see Table 5). Theyare typical of cercopithecids in their mor-phology. The lumbar region was clearlylong, but it is impossible to gauge its lengthrelative to the other vertebral segments. It isclear that this individual possessed a longtail, each of the twelve caudal vertebrae arequite long, and they still do not represent thecomplete length of the tail.
Forelimb.A left scapular glenoid fragment with a smallbit of the spine and coracoid process, A.L.2-64, is preserved. The glenoid cavity is some-what concave, being much more curvedthan is generally true for most colobines.More striking than the curvature is themedial bulging of the distal part of thecavity, whose border curves sharply mediallymaking a very circular posterior portion,very much like that of KNM-ER 4420.
The proximal and distal ends of the lefthumerus are preserved (A.L.2-63 and -64respectively), and there is also a distalhumerus from Hadar, A.L.222-14, whichhas been partially reconstructed to reversecrushing distortion, that is quite similar to

739
Figure 25. Projection of the greater tuberosity relative to the humeral head. Categories follow those fromHarrison (1989): (A), greater tuberosity well above head; (B), slightly above; (C), equal; (D), slightlybelow; (E), well below. Some data from T. Harrison: AMNH specimens scored by Harrison were rescoredhere, and found to have equivalent results.
A.L.2-64. When the proximal end is viewedlaterally, the greater tuberosity is even withor slightly higher than the head, althoughthis depends to some degree on how theshaft is oriented (this error is indicated byhatches in Figure 25). Thus, there is someerror in this estimate, and some fossilsscored as class B or D may be class C or viceversa. Regardless of orientation, there wouldbe no errors between classes A and B orclasses D and E. In A.L.2-63, the greatertuberosity is higher than in most extantcolobines, but is within the range of all
except for Nasalis (Figure 25). Compared toother Pliocene fossil colobines, the projec-tion of the greater tuberosity is similar tothat of P. chemeroni, lower than that ofKNM-ER 4420 (assigned to C. williamsi),and higher than that of KNM-ER 1542(assigned to Rhinocolobus turkanaensis). It isimpossible to tell how retroflexed the shaftwould have been, as only the proximal 3 cmare preserved. The lateral surface of thegreater tuberosity is marked by a modestm. infraspinatus fossa. The head is relativelybroad and spherical and comes to a slight

740 .
point posteriorly. The bicipital groove iswide relative to the size of the humerus andshallow, and its lateral rim curls over slightlymedially. The little that is visible of thedeltoid crest appears fairly strongly devel-oped, although not to the degree seen inKNM-ER 4420.
Distally, the humerus shows a number ofadaptations to a terrestrial habitus. As seemsto be typical of colobines, the articular sur-face is narrow relative to the total biepi-condylar width. The medial trochlear flangeis long and sharp, particularly in comparisonto modern colobines. Relative to distalarticular width, the trochlear flange is longerthan in any extant colobine other thanSemnopithecus entellus, and is generallywithin the range of the more terrestrialcercopithecines (see Figure 12). The zonaconoidea is more prominent than in mostcercopithecines, but not as large as in the cf.Rhinocolobus humeri KNM-ER 1542o andA.L. 300–1. The capitulum is more cylindri-cal than that of Colobus, Procolobus, andRhinocolobus, but more spherical than that ofKNM-ER 4420 or Papio. It is also fairlylong. The radial fossa is deeper and tallerthan the coronoid fossa on A.L.2–64 as isthe normal condition in the Colobinae(Olivier & Caix, 1959; Szalay & Delson,1979). On A.L.222–14, the radial is deeperthan but similar in height to the coronoid.The medial epicondyle is long, and retro-flexed at 64� relative to the distal articularsurface in A.L.2–64 and 58� in A.L.222–14(see Figure 11). This is within the range ofmodern Papio hamadryas and Theropithecusgelada, as well as T. o. cf. darti. This isoutside of the range of all extant colobines,except for some individuals of Semnopithecusentellus. It is also more retroflexed thanParacolobus chemeroni and Rhinocolobusturkanaensis, including A.L.300-1, but simi-lar to the 65� of KNM-ER 4420. In spite ofthe high angle of retroflection, the medialepicondyle projects relatively far mediallydue to its large overall size. Thus, the ratio
of the medial epicondyle relative to overallhumeral breadth is modest overlapping alarge number of taxa, but still smaller thanthe more arboreal colobines (see Figure 10).The olecranon fossa is proximodistally shortand mediolaterally broad. In overall mor-phology, the humerus appears similar to thatof KNM-ER 4420.
A.L.2–65 is the proximal end of the leftulna preserving the olecranon, trochlear andradial fossae, and 1·5 cm of the shaft distalto the radial articulation. The olecranonprocess is tall, modestly retroflexed, and tiltsmedially. The trochlear notch is deep, andthe medial end of its superior border is onlyslightly more proximal than the lateral end.This feature is different than in most terres-trial papionins, where the medial end issignificantly more proximal than the lateral.The radial articulation is singular. Theradial notch is deep and cuts considerablyinto the area of the trochlear articular sur-face. Thus, when it is articulated with thehead of the radius A.L.2-66 in the pronatedposition it matches the oblong radial headtightly. The shaft distal to the radialnotch is mediolaterally thin, but deepanteroposteriorly. The area of the inter-osseus border is damaged, but part is pre-served just distal to the radial notch, whichis strongly marked. The distal end of theright ulna, A.L.2-37, is also present, pre-serving the head and styloid process. As istypical for the family, the styloid has a largearticulation for the pisiform and triquetrum,which is separated from the head by a well-marked sulcus.
A.L.2-66 and A.L.2-67 are the proximaland distal ends of the left radius respectively.A.L.2-66 preserves the head, neck, tuberos-ity and 2 cm of the shaft. In proximal view,the head is broad and oval in outline, beingconsiderably broader in the transversedimension than its width in the anteropos-terior direction. The neck is short, robustand square. The shaft is damaged at theoblique line, but it does not appear to have

741
a well-marked interosseus border. Notenough of the shaft is preserved to discernits degree of curvature.
Hindlimb.Parts of both os coxae are preserved.Of the left side there is only a smallfragment, A.L.2-71, consisting of theacetabulum and a bit of the surroundingbone, with the head of the proximal femurA.L.2-72 still fixed in the acetabulum.A.L.2-80 is the right innominate, whichpreserves the acetabulum, and some of theright ilium, including the sacral articularfacet. There are also many small fragmentsof this ilium present, but unattached. Theilium is narrow and tall, as is typical for thefamily. Both ischial tuberosities are preservedand they are large. It is not possible to becertain whether or not the attached callositieswould have been continuous in the midline.
Large parts of both femora are preserved.Left proximal femur A.L.2-72, which is stillarticulated in the acetabulum of A.L.2-71,preserves the head, neck, greater trochanterand part of the shaft (see Figure 13).A.L.2-73 is a segment of the midshaft, andA.L.2-74 is the distal end with approxi-mately one-third of the shaft. Of the rightfemur, A.L.2-70/80 preserves the head, neckand small portion of the shaft with the lessertrochanter, and provides the antimere of thearea obscured by the acetabulum on the left(Figure 13). A.L.2-81 is the middle half ofthe femoral shaft. A.L.2-28 is the distalarticular surface and one-third of the shaft.Thus, a fair amount of femoral morphologyis preserved. While it is impossible to unam-biguously determine the femoral length as asmall segment is missing just below thelesser trochanter and there is considerabledamage to the shaft, but it would have beenapproximately 24 to 25 cm in length. This isslightly larger than male Nasalis larvatus(23 cm) and shorter than male Paracolobuschemeroni (27 cm).
The head is spherical, with a large andround fovea capitis. The articular surfacedoes not spread onto the posterior surface ofthe neck, implying less abduction of the hip.Relative to the size of the head, the neck islong compared to other colobines. The neckis oriented at an angle of 114� to the shaft.This is in the lower range for most arborealcercopithecid taxa, and the upper part of therange for the more terrestrial species (seeFigure 14). One of the most striking featuresof the proximal femur is the large andprominent greater trochanter. It is extremelytall and straight. It does not curve mediallyas in Theropithecus (Krentz, 1993). Relativeto the diameter of the head, it is shorter thanthe sampled Theropithecus gelada, in thelower range for Papio hamadryas, and withinthe middle of the ranges for Mandrillussphinx and T. o. cf. darti. It is generally tallerthan that of other colobines, but slightlyshorter than that of KNM-ER 4420 (seeFigure 15). The gluteal fossa is deep, butdoes not extend below the level of them. quadratus femoris insertion. The lessertrochanter is also long and is orientedposteriorly.
Distal to the lesser trochanter, the shaft isrobust, and thickens distally. In anteriorview the shaft is straight without either anormal or reverse carrying angle. In lateralview, the midshaft is quite bowed, especiallyin comparison to other colobines. The patel-lar groove is fairly tall, but is also broad anddeep. The lateral margin is stronger than themedial, and is elevated considerably to forma prominent ridge. The medial and lateralcondyles are symmetrical, and deep in lat-eral view. A.L.2-75 and 76 are the left andright patellae respectively. They are largesesamoids that articulate well with theirrespective grooves on the femora.
A.L.2-77 and 117 are the left and rightproximal tibiae, respectively. A.L.2-78 is thedistal end of the left tibia. Proximally, thecondyles are transversely narrow and deep inthe anteroposterior plane. The tuberosity is

742 .
large and prominent, and the shaft is narrowand deep in cross-section. Unfortunately,not enough is present to estimate overalllength. The medial malleolus is broken. Theastragalar facet is highly asymmetrical, indi-cating that the lateral border of the astra-galus would have been much higher than themedial.
Of the fibulae, distal right and proximalleft fragments are preserved, numberedA.L.2-38 and 39 respectively. They are typi-cal of cercopithecids in morphology. Theastragalar facet of the lateral malleoluswould have been relatively vertically aligned.
Only two elements of the tarsus have beenpreserved, both catalogued as A.L.2-44.One is the left navicular, the other the leftcuboid. In overall proportions, the latter isproximodistally longer than it is wide.Although it is slightly damaged distally, bothproximal and distal facets for the ectocunei-form can be seen. This condition is typicalof Asian colobines, and may be primi-tive for the subfamily (Strasser, 1988).Unfortunately a population is required todetermine the significance of this feature, asit varies to some degree within species.Additionally, a fragment of the proximal endof the left second metatarsal (A.L.2-122)and distal fragments of two metatarsals(A.L.2-41 and -42) are preserved.
RemarksEck (1976; 1977) noted the affinity of theLeadu specimen with Cercopithecoides wil-liamsi from South Africa in his discussion ofsimilarly sized colobine molar teeth fromthe Omo, and his opinion is borne out byour analysis. This material is here referredto Cercopithecoides for several reasons, pend-ing a thorough review of the genus. Thebroad interorbital pillar is unlike Nasalis,Libypithecus, and Rhinocolobus. The nasalsare longer than those of Rhinopithecus andRhinocolobus, and significantly shorter thanthose of Nasalis. The face is less projectingthan that of Paracolobus and Rhinocolobus.
The symphysis is shallow, similar to Cerco-pithecoides williamsi, C. kimeui and Procolobusbut different from Kuseracolobus, Paracolo-bus, Rhinocolobus and Colobus. The corpus isshallow and broad, similar to other speciesof Cercopithecoides and P.(Procolobus), butdistinct from Kuseracolobus, Rhinocolobus,Paracolobus and to some extent Colobus. Thecorpus is deepest inferior to the M1–2, againsimilar to other species of Cercopithecoides.The gonial region is only very slightly ex-panded, similar to other Cercopithecoides andParacolobus, but distinct from Kuseracolobus,Rhinocolobus, Paracolobus mutiwa and mostColobus. The ramus is short and deep, as inCercopithecoides, but unlike Rhinocolobus andParacolobus. One feature of the mandiblethat is unlike other species of Cercopithecoides(except for the new species from Lothagam)is the absence of a median mental canal.
Finally, in the postcranium, there aremany features related to terrestriality. Thisis unlike most extant colobines or Rhino-colobus turkanaensis, but similar to Cerco-pithecoides williamsi from Koobi ForaDolichopithecus ruscinensis, and to a lesserextent Paracolobus chemeroni or P. mutiwa.The humerus has a long medial trochlearkeel, and a retroflexed medial epicondyle.The ulnar olecranon is moderately retro-flexed. On the femur, the greater trochanteris long and the lesser trochanter is pos-teriorly oriented, which is unlike Paracolobuschemeroni, but similar to P. mutiwa (Ting,2001).
It seems clear, for the reasons given in thediagnosis, that this taxon is specifically dis-tinct from C. kimeui and C. williamsi, as wellas from the new species from Lothagam.
Colobinae genus et species indet., large size(cf. size B of Eck, 1976)(=or including Colobinae gen. et sp. nov. 1.Szalay & Delson, 1979, in part. cf.Rhinocolobus, Delson, 1984, 1994, in part)Specimens included: NME A.L.100–356,A.L.109–14, A.L.403–44.

743
DescriptionThere are three isolated molar teeth of largecolobines in the sample. These are all withinthe size range of cf. Rhinocolobus turkanaensis(dental dimensions given in Table 7) andderive from similar stratigraphic levels, withA.L.403-44 being from below the SidiHakoma tuff, and A.L.109-14 from the SidiHakoma Member above the tuff. A.L.100-356 is from Ahmado, which may be roughlyequivalent in age to the Sidi HakomaMember (Kalb, 1993). Thus, they all dateto approximately 3·4 Ma. Given the pres-ence of two colobines of similar dental sizein the Afar region: cf. Rhinocolobus turka-naensis from Hadar and cf. Cercopithecoidessp. indet. from Maka in the Middle Awash(Frost, 2001b), and the fact that these twotaxa are as yet virtually indistinguishabledentally, it is best to leave these specimensunidentified to genus. All of these molars arenormal for colobines with deep notches,high cuspal relief, and sharp cross-loph(id)s.A.L.100-356 is an M3, and shows a distalloph that is considerably narrower than themesial. The crown is virtually unworn, andthere is no root. The cusps are quite tall, andthe buccal notch fairly short. The other twoare lower molars. A.L.403-44 could beeither an M1 or M2. A.L.109-14 is an M3. Ithas a distal lophid that is as wide as themesial lophid.
Table 7 Dental dimensions of Colobinae indet.size B
Cat. No. Tooth WS W O L
A.L.100-356 M3 0 9·8 7·6 10·5A.L.109-14 M3 0 9·2 9·2 15·7A.L.403-44 MX 2 7·9 8·5 10·2
Abbreviations as for Table 1.
Summary and conclusions
The cercopithecid sample from the Hadarregion is important for understanding sev-
eral aspects of the evolution of the family.The distribution of the various speciesthrough the Hadar Formation and in thesurrounding sites is given in Table 9. Inthe Hadar sample, Parapapio. cf. jonesi is theonly cercopithecine other than Theropithecus.It is quite rare, being fairly well representedonly in the Kada Hadar Member, where itrepresents just over 20 % of the cercopithe-cids recovered. This is largely due to a singlelocality, A.L.363, where at least three indi-viduals of Pp. cf. jonesi were recovered.Given the relatively small number of indi-viduals from this member, and the low over-all abundance of this taxon in the HadarFormation, this relatively high local abun-dance is probably the result of samplingbias. No specimens of this taxon havebeen collected so far from the Kada HadarMember above the disconformity or fromthe Pinnacle site.
While Parapapio is a rare taxon at Hadar,its presence is important. In South Africa,Parapapio is diverse (four commonly recog-nized species) and known from severalPliocene sites (though absent fromPleistocene ones). In spite of several tenta-tive assignments in the literature, the Hadarmaterial is the only definitive evidence forthe genus in East Africa in the Pliocene,because all of the known diagnostic featuresare located in the face. While there is aMiocene sample from Lothagam withfacial material that represents this genus(Leakey et al., in press), none of the otherPliocene fossils from East Africa assigned toParapapio preserve enough of the face todiagnose them unequivocally as Parapapio.The sample from Laetoli, including the typespecimen of Cercocebus ado, was transferredto Parapapio by Leakey & Delson (1987)based on the lack of mandibular corpusfossae and presence of typical papionin(rather than Cercocebus-like) dentition. Amandible from Kanapoi, initially assignedby Patterson (1968) to Parapapio jonesibased on dental size, was referred to the

744 .
Table 8 Dimensions taken on various postcranial elements
A.L.363-12 A.L.2-63/64 A.L.222-14 A.L.300-1 A.L.577-1
HumeriProximal mediolateral dimension 29Proximal anteroposterior dimension 32Biepicondylar breadth 42 35 32 46 44Lateral epicondyle to medial edge oftrochlea
35 31 27 40
Distal articular breadth 30 24 20 31 29Medial trochlear flange length 17 19 15 20 22
Ulnae A.L.2-65
Height of olecranon above trochlearfossa
10
Proximodistal trochlear fossa length 19Mediolateral breadth across radial facetand trochlear notch
22
Radii A.L.2-66
Maximum head diameter 18Perpendicular diameter 15Proximodistal neck length 9Proximodistal length from head tomid-tuberosity
25
Femora A.L.363-1c/d A.L.2-70/72
Anteroposterior head diameter 21 23Maximum mediolateral breadth 47 10Greater trochanter projection 12 12Bicondylar breadth 38 38Anteroposterior condyle length 31 31
Tibia A.L.2-77
Proximal anteroposterior depth 23Proximal mediolateral breadth 39
For more specific information on measurements taken, see text.
same species as is present at Laetoli byLeakey & Delson (1987). More recent col-lections from Kanapoi have recovered alarger sample of this species, which seems toconfirm this diagnosis (Leakey et al., 1995).This sample also lacks facial material,however and the allocation of Pp. ado toParapapio must therefore continue to beonly tentative.
The Hadar sample of Pp. cf. jonesi is alsoimportant because of the associated post-crania. In spite of being quite common inSouth Africa, no postcrania are directly
associated with any cranial remains ofParapapio. From Laetoli, there are severalhand and foot bones that have beenallocated to Pp. ado (Leakey & Delson,1987), but most of the elements from Hadarare previously unknown for the taxon. Whilethese elements will require a more thoroughanalysis, they do seem to indicate that Pp.cf. jonesi was adapted for more arboreallocomotion than modern Papio hamadryas,Theropithecus gelada, or T. o. cf. darti.
The genus Theropithecus is one of themost widespread and common mammalian

745
genera during the Pliocene and Pleistocenein Africa. It is by far the most abundantcercopithecid taxon at Hadar, where itmakes up 75–95% of the cercopithecidspecimens identifiable to the generic level, indifferent strata. Two time-successive sub-species of T. oswaldi occur at Hadar: T. o. cf.darti from the Basal Member through thelower Kada Hadar Member in the HadarFormation, and from Ahmado and possiblyLeadu; and T. o. oswaldi from the upper partof the Kada Hadar Member, the Pinnaclesite, and possibly Geraru. In addition to thelarge increase in overall size and morpho-logical evolution in this species, comparisonof the Hadar and Makapansgat populationsof T. o. darti shows that there was a largeamount of geographic variability in this sub-species, perhaps more than was present inthe later subspecies T. o. oswaldi (see Figure17) and T. o. leakeyi. Alternatively, morethan one subspecies may need to be recog-nized. The postcranial material from Hadarassigned to this taxon generally show lessextreme adaptations for terrestriality than dothose of modern Theropithecus or Papio. Thisis similar to the description of the locomotorbehavior reconstructed for T. o. darti byKrentz (1993).
Colobines are rare in the Hadar forma-tion, but they are more diverse than thecercopithecines, with three or perhaps even
four species represented. There is a largecolobine, tentatively identified as cf. Rhino-colobus turkanaensis, present in the SidiHakoma and Denen Dora Members of theHadar Formation. While more certain diag-nosis of this taxon must await the discoveryof facial material, this would represent thefirst occurrence of Rhinocolobus outside ofthe Turkana Basin. Three isolated teethfrom the Sidi Hakoma Member and itsprobable equivalent at Ahmado documentthe presence of a large colobine which mostlikely represents cf. Rhinocolobus turkanaen-sis, but the possibility that they are betterconsidered cf. Cercopithecoides sp. indet., asknown from Maka in the Middle Awash,cannot be excluded. Another large colobine,Cercopithecoides kimeui, has been identified atHadar from the Pinnacle locality, probablyof early Pleistocene age. This species is alsoknown from Bed II (and probably III) atOlduvai Gorge, from the upper Burgi andKBS Members at Koobi Fora, and fromRawi on the Homa Peninsula (Ditchfieldet al., 1999; Frost et al., in review). AtOlduvai and Koobi Fora it is associated withTheropithecus oswaldi oswaldi as it is atHadar. Cercopithecoides kimeui may representa colobine which is typically associated withfaunas representing relatively open habitats.
The new taxon Cercopithecoides meaveae isa smaller colobine known from Leadu and
Table 9 Distribution of cercopithecid species in the Hadar Formation and surrounding areas
Member/site Geologic agePp. cf.jonesi
T. o. cf.darti
T. o.oswaldi
cf. R.turkanaensis C. meaveae C. kimeui Col. B.
Pinnacle �1·7 X XU. Kada Hadar �2·33 XGeraru �Kada Hadar? ?Leadu ? X XL. Kada Hadar 2·92–3·18 X XDenen Dora 3·18–3·22 X X ?Sidi Hakoma 3·22–3·18 Ma X X XBasal >3·4 Ma X XAhmado �Sidi Hakoma ? X X
‘‘X’’ indicates present in the specified horizon, ‘‘?’’ designates tentative allocations. Rows represent the differentmembers of the Hadar Formation and/or sites outside the Hadar Formation proper.

746 .
the Sidi Hakoma Member at Hadar. It issimilar in size to the largest extant colobines.This taxon adds to the known diversitywithin the genus Cercopithecoides. It appearsto be closely related to C. williamsi, at leastas represented at Koobi Fora by KNM-ER4420. Both C. meaveae and ER 4420 showadaptations of the elbow and hip that implymore frequent use of terrestrial behaviorsthan in extant African colobines, Paracolo-bus, and Rhinocolobus. Whether or not thisterrestrial adaptation is shared with some orall of the South African Cercopithecoidesmaterial is unknown as there are no post-crania directly associated with the latter(Elton, 2001). If Ciochon’s identification ofcolobine postcranial elements which showmore arboreal adaptations from sites wherethe only known colobine is C. williamsi iscorrect, then this may imply that the EastAfrican populations assigned to C. williamsiare not conspecific with the southernsamples, which are themselves quite variablemorphologically.
Acknowledgments
We thank the Ethiopian Centre for Researchand Conservation of Cultural Heritage, theEthiopian Ministry of Information andCulture, and the National Museum ofEthiopia for permission to study the materialin their care. We are grateful to Bill Kimbel,Don Johanson, and the Institute for HumanOrigins for their support of our study of theHadar material (and patience during themany years ED procrastinated), and alongwith the I.A.R.E., all of their effort in col-lecting and preparing the fossils describedhere. We also thank M. G. Leakey for apre-publication copy of her manuscript onthe fossil monkeys from Lothagam and dis-cussion of the differences between those andHadar taxa. Our thanks also go to M. G.Leakey, B. R. Benefit, C. J. Jolly, and N. J.Jablonski for comments on an earlier draft of
this manuscript. G. G. Eck kindly providedthe photograph of the lateral view of A.L.2-34. T. Harrison, N. Ting, and M. Singletondeserve very special thanks for sharingtheir data on humeri, femora, and M3srespectively, which greatly improved thispaper.
This research was supported by theWenner-Gren Foundation (Gr.6436), theL. S. B. Leakey Foundation, a NYCEPfellowship (NSF BIR 9602234), predoctoralsupport from the 3D Visualization project atAMNH (NSF ACI 9982351), a CUNYpredissertation travel award, and a pre-doctoral fellowship from the SmithsonianInstitution, Evolution of Terrestrial Eco-systems Consortium to SF. ED receivedsupport from the L.S.B. Leakey Founda-tion, the PSC-CUNY Faculty ResearchAward Program (grants 11483E and6-68479), the John Simon GuggenheimMemorial Foundation and NSF (BNS7915091, 8113628 and 8419939). We alsothank Anita Cleary for the drawings of theParapapio skull.
References
Alemseged, Z. & Geraads, D. (1998). Theropithe-cus ‘‘atlanticus’’ Thomas 1884 (Cercopithecidae,Primates) fossils from Ahl al Oughlam, Casa Blanca,Morocco. J. hum. Evol. 34, 609–621.
Alemseged, Z. & Geraads, D. (2001). A new MiddlePleistocene fauna from the Busidima-Telalak regionof the Afar, Ethiopia. C. r. Acad. Sci. Paris, Sciences dela Terre et des planetes 331, 549–556.
Aronson, J. A. & Taieb, M. (1981). Geology andpaleogeography of the Hadar hominid site, Ethiopia.In (G. Rapp, C. F. Vondra, Eds) Hominid Sites:Their geological settings, pp. 165–195. Boulder, CO:Westview Press.
Birchette, M. (1981). Postcranial remains of Cercopithe-coides. Am. J. phys. Anthrop. 54 (Suppl.), 201.
Birchette, M. (1982). The postcranial skeleton ofParacolobus chemeroni. Ph.D. dissertation, HarvardUniversity.
Brown, F. H. & Feibel, C. S. (1991). Stratigraphy,Depositional Environments and Palaeogeography ofthe Koobi Fora Formation. In (J. M. Harris, Ed.)Koobi Fora Research Project. Vol. 3, Stratigraphy,artiodactyls and paleoenvironments, pp. 321–346.Oxford: Clarendon Press.

747
Ciochon, R. L. (1993). Evolution of the cercopithecoidforelimb, phylogenetic and functional implicationsfrom morphometric analyses. Univ. Calif. Publ. Geol.Sci. 138, 1–251.
Delson, E. (1973). Fossil colobine monkeysof the circum-Mediterranean region and theevolutionary history of the Cercopithecidae(Primates, Mammalia). Ph.D. dissertation,Columbia University.
Delson, E. (1984). Cercopithecid biochronology of theAfrican Plio-Pleistocene: Correlation among easternand southern hominid-bearing localities. Cour.Forsch.-Inst. Senckenberg 69, 199–218.
Delson, E. (1988). Chronology of South Africanaustralopith site units. In (F. E. Grine, Ed.)Evolutionary History of the ‘‘Robust’’ Australopithecines,pp. 317–324. New York: Aldine.
Delson, E. (1993). Theropithecus fossils from Africa andIndia and the taxonomy of the genus. In (N. G.Jablonski, Ed.) Theropithecus: The Rise and Fall of aPrimate Genus, pp. 157–189. Cambridge: CambridgeUniversity Press.
Delson, E. (1994). Evolutionary history of thecolobine monkeys in paleoenvironmental perspec-tive. In (A. G. Davies & J. F. Oates, Eds) ColobineMonkeys: Their Ecology, Behaviour and Evolution,pp. 11–43. Cambridge: Cambridge University Press.
Delson, E. & Dean, D. (1993). Are Papio baringensisR. Leakey, 1969, and P. quadratirostris Iwamoto,1982, species of Papio or Theropithecus? In (N. G.Jablonski, Ed.) Theropithecus: The Rise and Fall of aPrimate Genus, pp. 125–156. Cambridge: CambridgeUniversity Press.
Delson, E., Eck, G. G., Leakey, M. G. & Jablonski,N. G. (1993). A partial catalogue of fossil remains ofTheropithecus. In (N. G. Jablonski, Ed.) Theropithecus:The Rise and Fall of a Primate Genus, pp. 499–525.Cambridge: Cambridge University Press.
Delson, E., Terranova, C. J., Jungers, W. L., Sargis,E. J., Jablonski, N. G. & Dechow, P. C. (2000). Bodymass in Cercopithecidae (Primates, Mammalia):estimation and scaling in extinct and extant taxa.Anthrop. Pap. Am. Mus. nat. Hist. 83, 1–159.
Ditchfield, P., Hicks, J., Plummer, T., Bishop, L. C. &Potts, R. (1999). Current research on the LatePliocene and Pleistocene deposits north of Homamountain, southwestern Kenya. J. hum. Evol. 36,123–150.
Eck, G. G. (1976). Cercopithecoidea from Omo groupdeposits. In (Y. Coppens, F. C. Howell, G. L.Isaac & R. E. F. Leakey, Eds) Earliest Man andEnvironments in the Lake Rudolf Basin, pp. 332–344.Chicago: University of Chicago Press.
Eck, G. G. (1977). Diversity and frequency distributionof Omo group Cercopithecoidea. J. hum. Evol. 6, 55–63.
Eck, G. G. (1987). Theropithecus oswaldi from theShungura Formation, Lower Omo Basin, South-western Ethiopia. In: Les faunes Plio-Pleistocenesde la Basse Vallee de l’Omo (Euthiopie) Tome 3, Cerco-pithecidae de la Formation de Shungura, pp. 123–139.Cahiers de Paleontologie, Travaux de PaleontologieEst-Africaine. Paris: CNRS.
Eck, G. G. (1993). Theropithecus darti from the HadarFormation, Ethiopia. In (N. G. Jablonski, Ed.)Theropithecus: The Rise and Fall of a Primate Genus,pp. 15–83. Cambridge: Cambridge University Press.
Eck, G. G. & Jablonski, N. G. (1987). The skull ofTheropithecus brumpti compared with those of otherspecies of the genus Theropithecus. In Les faunesPlio-Pleistocenes de la Basse Vallee de l’Omo (Euthiopie).Tome 3, Cercopithecidae de la Formation de Shungura,pp. 11–122. Cahiers de Paleontologie, Travaux dePaleontologie Est-Africaine. Paris: CNRS.
Eisenhart, W. L. (1974). The fossil cercopithecoids ofMakapansgat and Sterkfontein. B.A. Thesis, HarvardCollege.
Elton, S. (2001). Locomotor and habitat classifi-cations of cercopithecoid postcranial material fromSterkfontein Member 4, Bolt’s Farm and SwartkransMembers 1 and 2, South Africa. Paleontol. afr. 37,115–126.
Freedman, L. (1957). Fossil Cercopithecoidea ofSouth Africa. Annls. Transv. Mus. 23, 121–262.
Freedman, L. (1961). Some new fossil cercopithe-coid specimens from Makapansgat, South Africa.Paleontol. afr. 7, 7–45.
Freedman, L. (1965). Fossil and subfossil primatesfrom the limestone deposits at Taung, Bolt’s Farm,and Witkrans South Africa. Paleontol. afr. 9, 19–48.
Freedman, L. (1976). South African fossil cercopithe-coidea: A reassessment including a description ofnew material from Makapansgat, Sterkfontein andTuang. J. hum. Evol. 5, 297–315.
Frost, S. R. (2001a). New Early Pliocene Cercopithe-cidae (Mammalia: Primates) from Aramis, MiddleAwash Valley, Ethiopia. Am. Mus. Novit. 3350,1–36.
Frost, S. R. (2001b). Fossil Cercopithecidae fromthe Afar Depression, Ethiopia: Species systematicsand comparison to the Turkana Basin. Ph.D.Dissertation, City University of New York.
Frost, S. R., Plummer, T., Bishop, L. C., Ditchfield,P. D., Ferraro, J. & Hicks, J. (In review). Partialcranium of Cercopithecoides kimeui Leakey, 1982 fromRawi Gully, Southwestern Kenya. Am. J. phys.Anthrop.
Gilbert, W. H., Asfaw, B. & White, T. D. (2000).Paleontology. In (J. D. Clark, K. D. Schick &W. H. Gilbert, Eds) The Acheulean and thePlio-Pleistocene deposits of the Middle AwashValley, Ethiopia. Royal Museum of Central Africa,(Belgium) Annales–Sciences Geologiques 104, 183–192.
Harrison, T. (1989). New postcranial remains ofVictoriapithecus from the middle Miocene of Kenya.J. hum. Evol. 18, 3–54.
Jablonski, N. G. (1986). The hand of Theropithecusbrumpti. In (P. Lee & J. Else, Eds) PrimateEvolution, Proceedings of the 10th Congress ofthe International Primatological Society, Vol. 1.Cambridge: Cambridge University Press.
Jablonski, N. J., Leakey, M. G., Kiarie, C. & Anton, M.(2002). A new skeleton of Theropithecus brumpti(Primates: Cercopithecidae) from Lomekwi, WestTurkana. J. hum. Evol. (in press).

748 .
Johanson, D. C., Taieb, M. & Coppens, Y. (1982).Pliocene hominids from the Hadar Formation,Ethiopia (1973–1977): stratigraphic, chronologic,and paleoenvironmental contexts, with notes onhominid morphology and systematics. Am. J. phys.Anthrop. 57, 373–402.
Jolly, C. J. (1972). The classification and natural his-tory of Theropithecus (Simopithecus) (Andrews, 1916),baboons of the African Plio-Pleistocene. Bull. Br.Mus. Nat. Hist., Geology, 22, 1–122.
Kalb, J. E. (1993). Refined stratigraphy of the hominid-bearing Awash Group, Middle Awash Valley, AfarDepression, Ethiopia. Newsletters Stratigraphy 29,21–62.
Kalb, J. E., Jaegar, M., Jolly, C. J. & Kana, B. (1982a).Preliminary geology, palaeontology and paleoecologyof a Sangoan site at Andalee, Middle Awash Valley,Ethiopia. J. Archaeol. Sci. 9, 349–363.
Kalb, J. E., Jolly, C. J., Tebedge, S., Mebrate, A.,Smart, C., Oswald, E. B., Whitehead, P. F., Wood,C. B., Tsrha, A. & Rawn-Schatzinger, V. (1982b).Vertebrate faunas from the Awash group, MiddleAwash Valley, Afar, Ethiopia. J. Vert. Paleontol. 2,237–258.
Kalb, J. E., Wood, C. B., Smart, C., Oswald, E. B.,Mebrate, A., Tebedge, S. & Whitehead, P. (1980).Preliminary geology and palaeontology of the BodoD’ar hominid site, Afar, Ethiopia. Palaeogeogr.,Palaeoclimatol., Palaeoecol. 30, 107–120.
Kimbel, W. H. & Walter, R. C. (2000). Hadar. In(E. Delson, I. Tattersall, J. A. Van Couvering &A. S. Brooks, Eds) Encyclopedia of Human Evolutionand Prehistory, 2nd edn. pp. 299–302. New York:Garland.
Kimbel, W. H., Johanson, D. C. & Rak, Y. (1994).The first skull and other new discoveries ofAustralopithecus afarensis at Hadar, Ethiopia. Nature368, 449–451.
Kimbel, W. H., Walter, R. C., Johanson, D. C., Reed,K. E., Aronson, J. L., Assefa, Z., Marean, C. W.,Eck, G. G., Bobe, R., Hovers, E., Rak, Y., Vondra,C., Yemane, T., York, D., Chen, Y., Evensen, N. M.& Smith, P. E. (1996). Late Pliocene Homo andOldowan tools from the Hadar Formation (KadaHadar Member), Ethiopia. J. hum. Evol. 31, 549–561.
Krentz, H. B. (1992). The forelimb anatomy of Thero-pithecus brumpti and Theropithecus oswaldi from theShungura Formation, Ethiopia. Ph.D. Dissertation,University of Washington.
Krentz, H. B. (1993). Postcranial anatomy of extantand extinct species of Theropithecus. In (N. G.Jablonski, ed.) Theropithecus: The Rise and Fall of aPrimate Genus, pp. 383–422. Cambridge: CambridgeUniversity Press.
Leakey, M. G. (1982). Extinct large colobines from thePlio-Pleistocene of Africa. Am. J. phys. Anthrop. 58,153–172.
Leakey, M. G. (1993). Evolution of Theropithecusin the Turkana Basin. In (N. G. Jablonski, Ed.)Theropithecus: The Rise and Fall of a Primate Genus,pp. 85–123. Cambridge: Cambridge UniversityPress.
Leakey, M. G. & Delson, E. (1987). Fossil Cercopithe-cidae from the Laetolil Beds, Tanzania. In (M. D.Leakey & J. M. Harris, Eds) The Pliocene Site ofLaetoli, northern Tanzania, pp. 91–107. Oxford:Oxford University Press.
Leakey, M. G., Teaford, M. & Ward, C. V. (In press).Cercopithecidae from Lothagam. In (M. G. Leakey& J. M. Harris, Eds) Lothagam: Dawn of humanity ineastern Africa. New York: Columbia University Press(in press).
Maier, W. (1970). New fossil Cercopithecoideafrom the Lower Pleistocene cave deposits of theMakapansgat Limeworks, South Africa. Palaeont. afr.3, 69–107.
Olivier, G. & Caix, M. (1959). L’humerus duSemnopitheque. Mammalia 23, 78–99.
Singleton, A. M. (2002). Patterns of cranialshape variation in the Papionini (Primates: Cerco-pithecinae). J. hum. Evol. 42, 547–578.
Strasser, E. (1992). Hindlimb proportions, allometry,and biomechanics in Old World monkeys (Primates,Cercopithecidae). Am. J. phys. Anthrop. 87, 187–213.
Szalay, F. S. & Delson, E. (1979). Evolutionary Historyof the Primates. San Diego: Academic Press.
Taieb, M., Johanson, D. C., Coppens, Y. & Aronson,J. L. (1976). Geological and palaeontological back-ground of Hadar hominid site, Afar, Ethiopia. Nature260, 289–293.
Tiercelin, J.-J. (1986). The Pliocene Hadar Formation,Afar Depression, Ethiopia. In (L. E. Frostic, R. W.Reneaut, I. Read & J-J. Tiercelin, Eds) Sedimentationin the African Rifts, pp. 221–240. Oxford: BlackwellScientific Publications.
Ting, N. (2001). The hip and thigh of Paracolobusmutiwa and Paracolobus chemeroni. M.A. Thesis,University of Missouri, Columbia.
Walter, R. C. (1994). Age of Lucy and the First Family:single-crystal 40Ar/39Ar dating of the Denen Doraand lower Kada Hadar members of the HadarFormation, Ethiopia. Geology 22, 6–10.
Walter, R. C. & Aronson, J. L. (1993). Age andsource of the Sidi Hakoma Tuff, Hadar Formation,Ethiopia. J. hum. Evol. 25, 229–240.
Yemane, T. (1997). Stratigraphy and sedimentologyof the Hadar formation, Afar, Ethiopia. Ph.D.Dissertation. Iowa State University.