foraging leafcutter ants: olfactory memory underlies delayed avoidance of plants unsuitable for the...
TRANSCRIPT

lable at ScienceDirect
Animal Behaviour 82 (2011) 453e458
Contents lists avai
Animal Behaviour
journal homepage: www.elsevier .com/locate/anbehav
Foraging leafcutter ants: olfactory memory underlies delayed avoidanceof plants unsuitable for the symbiotic fungus
N. Saverschek*, F. RocesBehavioural Physiology and Sociobiology, Biocenter, University of Würzburg, Germany
a r t i c l e i n f o
Article history:Received 28 September 2010Initial acceptance 1 November 2010Final acceptance 9 May 2011Available online 16 July 2011MS. number: 10-00651
Keywords:Acromyrmex ambiguusaversive learningleafcutter antmemoryodourolfactionsubstrate selection
* Correspondence: N. Saverschek, Behavioural PBiocenter, University of Würzburg, Am Hubland, D-97
E-mail address: [email protected]
0003-3472/$38.00 � 2011 The Association for the Studoi:10.1016/j.anbehav.2011.05.015
Leafcutter ants are known to reject previously accepted plants if they prove to be unsuitable for theirsymbiotic fungus: a phenomenon that involves avoidance learning. Workers need to associate thedetrimental effects of the incorporated plant with its characteristics, chemical and/or physical features,thus allowing plant recognition at the foraging site on subsequent days, and its avoidance. We addressedthe question of to what extent olfactory memory underlies delayed avoidance of plants unsuitable for thesymbiotic fungus. Odour is an important plant characteristic used as a recognition cue outside the nestduring foraging. Acromyrmex ambiguus foragers were able to learn about a plant’s unsuitability for theirsymbiotic fungus, rejecting this substrate in a binary choice experiment in the laboratory. Presented withleaf discs of two plant species simultaneously, one known to be unsuitable and one suitable, individualforagers significantly preferred the suitable plant species. When presented only with the plant odour,foragers steered away from the odour of the known unsuitable plant species, moving towards thesimultaneously presented odour of a suitable plant species. Foragers were therefore able to identify plantspecies and recall information about substrate suitability for the fungus through the plant’s odour alone.None the less, foragers showed a significantly stronger avoidance of known unsuitable substrate whenthey could make contact with the leaf discs, suggesting the use of additional cues for plant identification.It is argued that not only appetitive, but also aversive learned responses are involved in the process ofplant recognition by foraging leafcutter ants.� 2011 The Association for the Study of Animal Behaviour. Published by Elsevier Ltd. All rights reserved.
Herbivorous insects are often confronted with a highly diverseenvironment, in which both plant availability and quality stronglyvary interindividually, and also on a seasonal basis. The selection ofsuitable food plants may therefore be complex, involving recogni-tion, assessment of the plant’s nutritional value and detection ofrepellent compounds, the level of which may be increased asa response to feeding (Karban & Myers 1989). Particularly ina changing and unpredictable environment, the occurrence oflearned foraging responses should be of particular adaptive value(Krebs & Inman 1994): insects may show both learned preferencefor high-quality resources, and postingestive learned avoidance ofnutritionally deficient or toxic foods (Bernays 1993).
Leafcutter ants are polyphagous herbivores that, despite theircatholicity, show distinct preferences in their plant choice. Foragingdecisions about the suitability of substrates are very complex, asleafcutter ants do not harvest for themselves, but for their symbioticfungus, which represents the sole food source of the developingbrood. The ants’ plant preferences are therefore expected to be
hysiology and Sociobiology,074 Würzburg, Germany.zburg.de (N. Saverschek).
dy of Animal Behaviour. Published
partially driven by the requirements of the fungus. Leafcutter antcolonies occur not only in habitats with highly diverse plantcommunities, harvesting 50e80% of the occurring plant species;they also usually harvest several species simultaneously (Cherrett1989; Wirth et al. 2003). Out in the field, foragers decide on plantmaterial for their symbiotic fungus based on both innate preferencesand theirownprevious experience. A further level of ‘qualitycontrol’occurs inside the nest, in the fungus gardens. Physical plant char-acteristics such as hardness can lead to postselection of plantmaterial by inside nest workers (Camargo et al. 2003). Moreimportantly, if a substrate proves to be unsuitable for the fungusowing to its chemical components, foragers stop harvesting thatsubstrate, even if it is harmless for them (Ridley et al. 1996; Northet al. 1999). With a fungicide undetectable by the ants, but harmfulto the fungus, Ridley et al. (1996), North et al. (1999), Herz et al.(2008) and recently Saverschek et al. (2010) demonstrated theinvolvementof the symbiotic fungus.Workers initially incorporatingbaits or leaves containing the fungicide into the fungus garden, andeven naïveworkers, stopped harvesting these substrates in responseto their deleterious effects on the fungus, a phenomenon thatinvolves avoidance learning by foragers, even though the substrateswere harmless for them (Saverschek et al. 2010).
by Elsevier Ltd. All rights reserved.

N.Saverschek, F.Roces / Animal Behaviour 82 (2011) 453e458454
Delayed avoidance of substrate has already been shown inseveral leafcutter ant species (Acromyrmex lundi, Acromyrmexoctospinosus, Acromyrmex subterraneus, Atta cephalotes, Atta laevi-gata, Atta sexdens, Atta colombica), both in the laboratory (Knappet al. 1990; Ridley et al. 1996; North et al. 1999; Camargo et al.2003; Herz et al. 2008) and in the field (Ridley et al. 1996;Saverschek et al. 2010). It occurs very fast, within 24 h, is speciesspecific (Herz et al. 2008; Saverschek et al. 2010), and is based ona postforaging mechanism via effects of the incorporated plantmaterial on the fungus garden. This was demonstrated by removingexperienced workers (those that incorporated the fungicide-containing plant fragments or bait) from the ‘treated’ garden, andintroducing naïve workers. After exposure to the garden, naïveworkers were shown to reject the previously unsuitable plantfragments or bait at the foraging arena on subsequent days,although they had never come into contact with them previously(North et al. 1999; Herz et al. 2008).
For delayed avoidance of previously accepted substrate tooccur, workers have to associate the detrimental effects of theincorporated plant with its characteristics (chemical and/orphysical features), thus allowing its recognition at the foraging siteon subsequent days, and its avoidance. Plant identification istherefore of utmost importance out in the field, as well as insidethe nest. Foraging leafcutter ants show marked preferences forleaves of certain plant species, individual plants, and even leaveswithin individual plants. Leaf features such as secondarycompounds (Hubbell & Wiemer 1983; Howard 1987, 1988),toughness (Nichols-Orians & Schultz 1990) and nutrient content(Berish 1986; Meyer et al. 2006) have been identified as influ-encing the choice of plant species. However, the relative impor-tance of these leaf traits for plant identification and choice, and inparticular the question of which physical and/or chemical char-acteristics are still available after the incorporation of the plantinto the fungus and could be used for plant recognition, remainsopen.
It is already known that the smell of harvested substrate canserve either as a mere orientation cue, with foragers walkingupwind towards the odour until they find the substrate, in leaf-cutter ants (Littledyke & Cherrett 1978; Therrien & McNeil 1990)and in desert ants, Cataglyphis (Wolf & Wehner 2000), or asa learned cue used as decision criterion during food collection inleafcutter ants (Roces 1990, 1994), carpenter ants (Provecho &Josens 2009) and stingless bees (Reichle et al. 2010). Odour istherefore expected to be an important cue for plant recognition, notonly outside the nest, but also inside the nest.
We addressed the question of to what extent olfactory memoryunderlies delayed avoidance of plants unsuitable for the symbioticfungus. Would the odour of an unsuitable plant species be suffi-cient to recall the negative association foragers had made with thisplant in the fungus garden, and subsequently cause foragers toavoid this plant during foraging? To answer this question, we tookadvantage of the known phenomenon of delayed avoidance ofplants unsuitable for the fungus in the leafcutter ant Acromyrmexambiguus, by testing individual foragers with a binary choicebefore and after the incorporation of unsuitable substrate into thefungus garden. In the initial control series, individuals wereoffered leaf discs of two different plant species simultaneously,and their spontaneous preferences were tested. Afterwards, one ofthe tested plant species was made unsuitable for the fungus, butnot for the ants, by infiltrating fragments with a fungicide.Twenty-four hours after its incorporation into the colony, indi-vidual preferences for the two plant species were evaluated again,testing the foragers’ ability to identify and therefore avoid theplant that had proven to be unsuitable for the fungus based on itsodour alone.
METHODS
Experiments were conducted with ‘subcolonies’ obtained fromfive large laboratory colonies (AeE) of A. ambiguus, collected inUruguay in 2002, and maintained in the laboratory at 25 �C andunder a 12:12 h light:dark cycle. Each subcolony was placed in anartificial nest consisting of three transparent Plexiglas boxes con-nected by short PVC tubes. The central box (19 � 8.5 and 8.5 cmhigh) contained the fungus on moistened expanded clay pebbles tokeep nest humidity high. The two other boxes (each 19 � 19 cm and8.5 cm high) served as a feeding box and a refuse box, respectively.Holes with a fine metallic mesh on the nest cover allowed aircirculation, thus creating a different, dryer microclimate from theone inside the fungus box: an ‘inside’ and ‘outside’ for thesubcolony. About 1000 ml of fungus garden (fungus and gardeningworkers) were taken from the mother colony and placed in theartificial nest, together with approximately 1000 foragers. Subcol-onies were established at least 4 days before the start of theexperiment to ensure well-established fungus gardens and activeforaging behaviour. The ants received fresh rose leaves (Rosacanina) and water every day, and honey water every other day. Allplant species used during the experiments are readily acceptedstandard feeding plants used year round for the maintenance of thelaboratory colonies.
Individual foragers’ plant preferences were tested in binarychoice experiments with two different set-ups. In the first, named‘leaf set-up’, single foragers could come into direct contact with theleaf discs of the two tested plant species, and the collection of onewas recorded as a choice. In the second experimental set-up, the‘odour set-up’, foragers had to choose one of the two offered plantspecies based solely on the plant odours. Two experimental serieswere conducted with each set-up. First, in the control series, twoplant species known as suitable to the subcolonies, blackberry andplum, were offered, and individual preferences recorded. After-wards, subcolonies received leaf discs of blackberry treated withfungicide, to create an unsuitable substrate known to the workers.As a consequence, one plant species was now known as unsuitablein the treatment series conducted 24 h later, whereas the otherplant species was known as suitable. The whole procedure wasrepeated in independent assays and with different subcoloniesusing a second pair of plants, privet and blackberry, with privetbeing the plant treated with the fungicide in the treatment series.
To change the suitability of the tested substrate, an aqueoussolution of cycloheximide (CHX; SigmaeAldrich, Deisenhofen,Germany) was used. It is undetectable by the ants but has beenproven to be fungicidal to the attine fungus in the laboratory(Ridley et al. 1996; North et al. 1999; Herz et al. 2008). To maintainleaf-specific properties, such as odour, surface characteristics andtoughness, the leaf internal airspace was infiltrated with anaqueous solution (0.03%, w/w) of cycloheximide (see Herz et al.2008). Each subcolony received 110 leaf discs after the initialcontrol series, which were readily accepted and incorporated intothe fungus garden.
Experimental Set-ups and Procedure
For the choice experiments, the feeding box of the subcolonywas disconnected and replaced with a PVC tube leading directlyfrom the fungus chamber to an entry platform. A wooden bridgeconnected the entry platform to the experimental feeding arena,1.5 m away, and served as a ‘main foraging trail’. A transfer bridge tosingle out individual foragers to the experimental set-up wasplaced 1 m away from the entry platform, and 50 cm away from theexperimental feeding arena. To guarantee a well-establishedforaging column, subcolonies foraged along the main trail for at
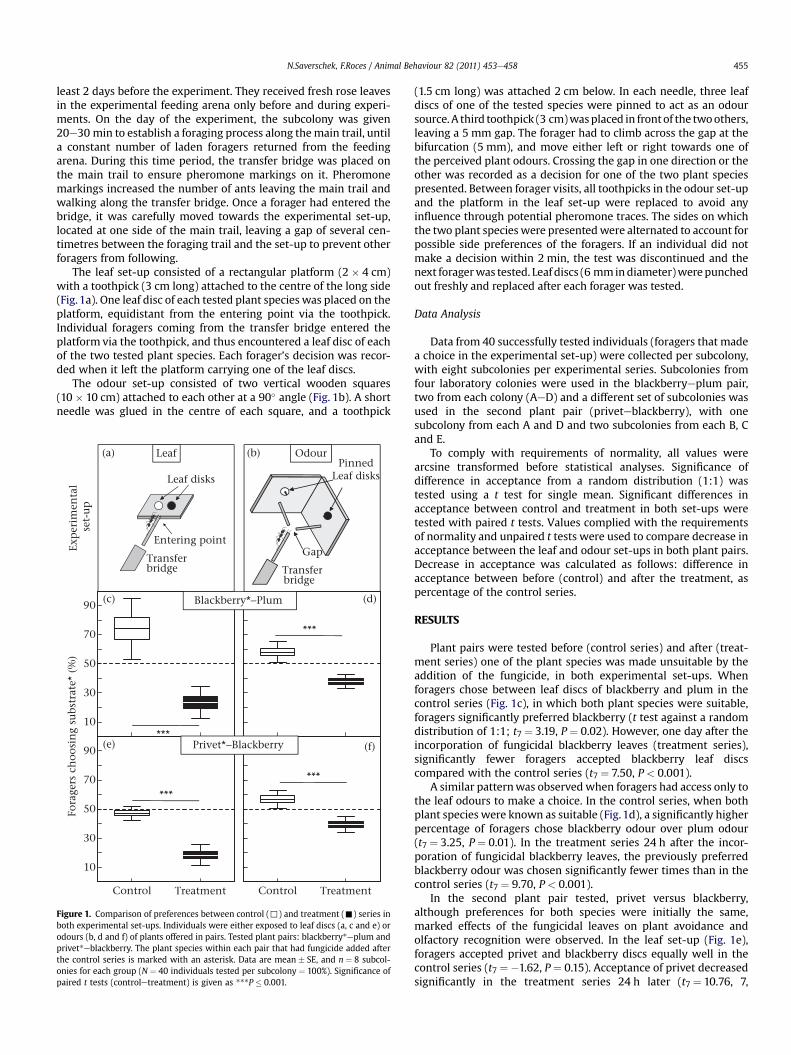
N.Saverschek, F.Roces / Animal Behaviour 82 (2011) 453e458 455
least 2 days before the experiment. They received fresh rose leavesin the experimental feeding arena only before and during experi-ments. On the day of the experiment, the subcolony was given20e30 min to establish a foraging process along themain trail, untila constant number of laden foragers returned from the feedingarena. During this time period, the transfer bridge was placed onthe main trail to ensure pheromone markings on it. Pheromonemarkings increased the number of ants leaving the main trail andwalking along the transfer bridge. Once a forager had entered thebridge, it was carefully moved towards the experimental set-up,located at one side of the main trail, leaving a gap of several cen-timetres between the foraging trail and the set-up to prevent otherforagers from following.
The leaf set-up consisted of a rectangular platform (2 � 4 cm)with a toothpick (3 cm long) attached to the centre of the long side(Fig.1a). One leaf disc of each tested plant species was placed on theplatform, equidistant from the entering point via the toothpick.Individual foragers coming from the transfer bridge entered theplatform via the toothpick, and thus encountered a leaf disc of eachof the two tested plant species. Each forager’s decision was recor-ded when it left the platform carrying one of the leaf discs.
The odour set-up consisted of two vertical wooden squares(10 � 10 cm) attached to each other at a 90� angle (Fig. 1b). A shortneedle was glued in the centre of each square, and a toothpick
Transfer bridge
PinnedLeaf disks
Gap
***
***
Control Treatment
Transfer bridge
Entering point
Leaf disks
10
30
50
70
90
***
10
30
50
70
90
***
Control Treatment
Blackberry*−Plum
Privet*−Blackberry
Leaf Odour
Fora
gers
ch
oosi
ng
subs
trat
e* (
%)
Exp
erim
enta
l s
et-u
p
(a) (b)
(c)
(e)
(d)
(f)
Figure 1. Comparison of preferences between control (,) and treatment (-) series inboth experimental set-ups. Individuals were either exposed to leaf discs (a, c and e) orodours (b, d and f) of plants offered in pairs. Tested plant pairs: blackberry*eplum andprivet*eblackberry. The plant species within each pair that had fungicide added afterthe control series is marked with an asterisk. Data are mean � SE, and n ¼ 8 subcol-onies for each group (N ¼ 40 individuals tested per subcolony ¼ 100%). Significance ofpaired t tests (controletreatment) is given as ***P � 0.001.
(1.5 cm long) was attached 2 cm below. In each needle, three leafdiscs of one of the tested species were pinned to act as an odoursource. A third toothpick (3 cm)wasplaced in frontof the twoothers,leaving a 5 mm gap. The forager had to climb across the gap at thebifurcation (5 mm), and move either left or right towards one ofthe perceived plant odours. Crossing the gap in one direction or theother was recorded as a decision for one of the two plant speciespresented. Between forager visits, all toothpicks in the odour set-upand the platform in the leaf set-up were replaced to avoid anyinfluence through potential pheromone traces. The sides on whichthe two plant species were presentedwere alternated to account forpossible side preferences of the foragers. If an individual did notmake a decision within 2 min, the test was discontinued and thenext foragerwas tested. Leaf discs (6 mmindiameter)werepunchedout freshly and replaced after each forager was tested.
Data Analysis
Data from 40 successfully tested individuals (foragers that madea choice in the experimental set-up) were collected per subcolony,with eight subcolonies per experimental series. Subcolonies fromfour laboratory colonies were used in the blackberryeplum pair,two from each colony (AeD) and a different set of subcolonies wasused in the second plant pair (priveteblackberry), with onesubcolony from each A and D and two subcolonies from each B, Cand E.
To comply with requirements of normality, all values werearcsine transformed before statistical analyses. Significance ofdifference in acceptance from a random distribution (1:1) wastested using a t test for single mean. Significant differences inacceptance between control and treatment in both set-ups weretested with paired t tests. Values complied with the requirementsof normality and unpaired t tests were used to compare decrease inacceptance between the leaf and odour set-ups in both plant pairs.Decrease in acceptance was calculated as follows: difference inacceptance between before (control) and after the treatment, aspercentage of the control series.
RESULTS
Plant pairs were tested before (control series) and after (treat-ment series) one of the plant species was made unsuitable by theaddition of the fungicide, in both experimental set-ups. Whenforagers chose between leaf discs of blackberry and plum in thecontrol series (Fig. 1c), in which both plant species were suitable,foragers significantly preferred blackberry (t test against a randomdistribution of 1:1; t7 ¼ 3.19, P ¼ 0.02). However, one day after theincorporation of fungicidal blackberry leaves (treatment series),significantly fewer foragers accepted blackberry leaf discscompared with the control series (t7 ¼ 7.50, P < 0.001).
A similar patternwas observed when foragers had access only tothe leaf odours to make a choice. In the control series, when bothplant species were known as suitable (Fig.1d), a significantly higherpercentage of foragers chose blackberry odour over plum odour(t7 ¼ 3.25, P ¼ 0.01). In the treatment series 24 h after the incor-poration of fungicidal blackberry leaves, the previously preferredblackberry odour was chosen significantly fewer times than in thecontrol series (t7 ¼ 9.70, P < 0.001).
In the second plant pair tested, privet versus blackberry,although preferences for both species were initially the same,marked effects of the fungicidal leaves on plant avoidance andolfactory recognition were observed. In the leaf set-up (Fig. 1e),foragers accepted privet and blackberry discs equally well in thecontrol series (t7 ¼ �1.62, P ¼ 0.15). Acceptance of privet decreasedsignificantly in the treatment series 24 h later (t7 ¼ 10.76, 7,
N.Saverschek, F.Roces / Animal Behaviour 82 (2011) 453e458456
P < 0.001), as observed with the previous plant pair tested. Whenforagers chose between odours of the two offered plant species(Fig. 1f), privet was significantly preferred in the control series(t7 ¼ 3.07, P ¼ 0.02), and acceptance decreased significantly in thetreatment series 24 h later (t8 ¼ 8.14, P < 0.001).
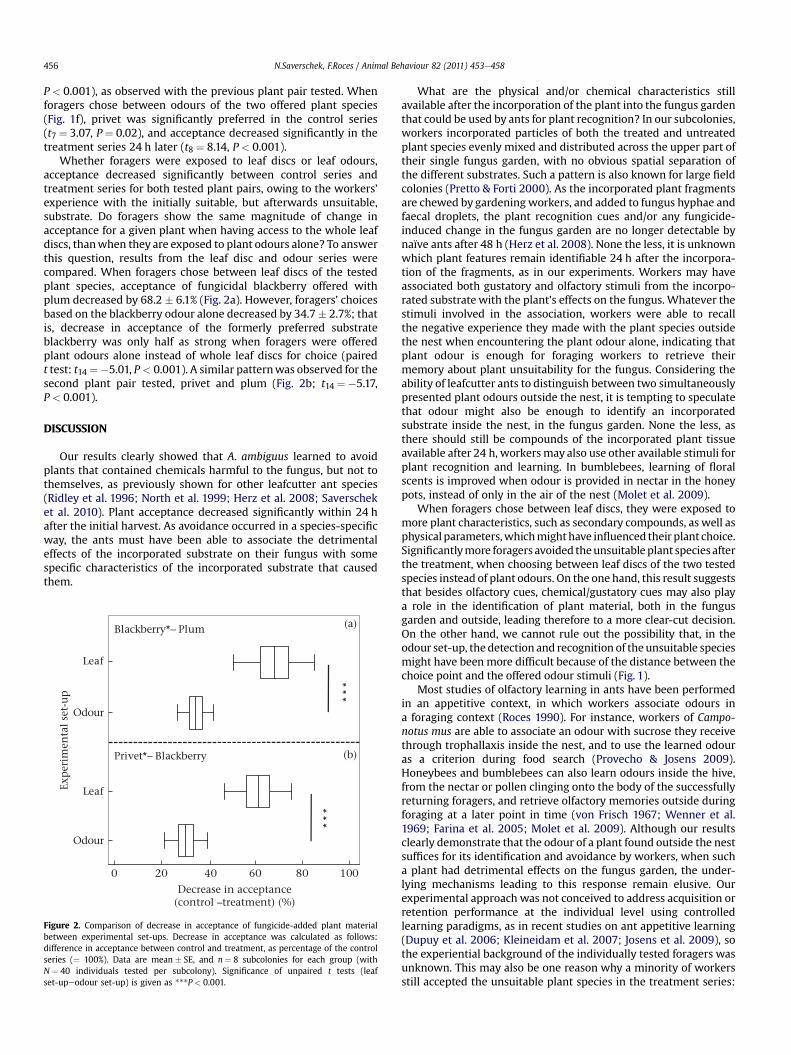
Whether foragers were exposed to leaf discs or leaf odours,acceptance decreased significantly between control series andtreatment series for both tested plant pairs, owing to the workers’experience with the initially suitable, but afterwards unsuitable,substrate. Do foragers show the same magnitude of change inacceptance for a given plant when having access to the whole leafdiscs, thanwhen they are exposed to plant odours alone? To answerthis question, results from the leaf disc and odour series werecompared. When foragers chose between leaf discs of the testedplant species, acceptance of fungicidal blackberry offered withplum decreased by 68.2 � 6.1% (Fig. 2a). However, foragers’ choicesbased on the blackberry odour alone decreased by 34.7 � 2.7%; thatis, decrease in acceptance of the formerly preferred substrateblackberry was only half as strong when foragers were offeredplant odours alone instead of whole leaf discs for choice (pairedt test: t14 ¼ �5.01, P < 0.001). A similar patternwas observed for thesecond plant pair tested, privet and plum (Fig. 2b; t14 ¼ �5.17,P < 0.001).
DISCUSSION
Our results clearly showed that A. ambiguus learned to avoidplants that contained chemicals harmful to the fungus, but not tothemselves, as previously shown for other leafcutter ant species(Ridley et al. 1996; North et al. 1999; Herz et al. 2008; Saverscheket al. 2010). Plant acceptance decreased significantly within 24 hafter the initial harvest. As avoidance occurred in a species-specificway, the ants must have been able to associate the detrimentaleffects of the incorporated substrate on their fungus with somespecific characteristics of the incorporated substrate that causedthem.
Exp
erim
enta
l se
t-u
p
Privet*− Blackberry
Blackberry*− Plum
**
*
**
*
0 20 40 60 80 100
Decrease in acceptance(control –treatment) (%)
Leaf
(a)
(b)
Odour
Leaf
Odour
Figure 2. Comparison of decrease in acceptance of fungicide-added plant materialbetween experimental set-ups. Decrease in acceptance was calculated as follows:difference in acceptance between control and treatment, as percentage of the controlseries (¼ 100%). Data are mean � SE, and n ¼ 8 subcolonies for each group (withN ¼ 40 individuals tested per subcolony). Significance of unpaired t tests (leafset-upeodour set-up) is given as ***P < 0.001.
What are the physical and/or chemical characteristics stillavailable after the incorporation of the plant into the fungus gardenthat could be used by ants for plant recognition? In our subcolonies,workers incorporated particles of both the treated and untreatedplant species evenly mixed and distributed across the upper part oftheir single fungus garden, with no obvious spatial separation ofthe different substrates. Such a pattern is also known for large fieldcolonies (Pretto & Forti 2000). As the incorporated plant fragmentsare chewed by gardening workers, and added to fungus hyphae andfaecal droplets, the plant recognition cues and/or any fungicide-induced change in the fungus garden are no longer detectable bynaïve ants after 48 h (Herz et al. 2008). None the less, it is unknownwhich plant features remain identifiable 24 h after the incorpora-tion of the fragments, as in our experiments. Workers may haveassociated both gustatory and olfactory stimuli from the incorpo-rated substrate with the plant’s effects on the fungus. Whatever thestimuli involved in the association, workers were able to recallthe negative experience they made with the plant species outsidethe nest when encountering the plant odour alone, indicating thatplant odour is enough for foraging workers to retrieve theirmemory about plant unsuitability for the fungus. Considering theability of leafcutter ants to distinguish between two simultaneouslypresented plant odours outside the nest, it is tempting to speculatethat odour might also be enough to identify an incorporatedsubstrate inside the nest, in the fungus garden. None the less, asthere should still be compounds of the incorporated plant tissueavailable after 24 h, workersmay also use other available stimuli forplant recognition and learning. In bumblebees, learning of floralscents is improved when odour is provided in nectar in the honeypots, instead of only in the air of the nest (Molet et al. 2009).
When foragers chose between leaf discs, they were exposed tomore plant characteristics, such as secondary compounds, as well asphysical parameters,whichmighthave influenced their plant choice.Significantlymore foragers avoided theunsuitableplant species afterthe treatment, when choosing between leaf discs of the two testedspecies instead of plant odours. On the one hand, this result suggeststhat besides olfactory cues, chemical/gustatory cues may also playa role in the identification of plant material, both in the fungusgarden and outside, leading therefore to a more clear-cut decision.On the other hand, we cannot rule out the possibility that, in theodour set-up, the detection and recognition of the unsuitable speciesmight have been more difficult because of the distance between thechoice point and the offered odour stimuli (Fig. 1).
Most studies of olfactory learning in ants have been performedin an appetitive context, in which workers associate odours ina foraging context (Roces 1990). For instance, workers of Campo-notus mus are able to associate an odour with sucrose they receivethrough trophallaxis inside the nest, and to use the learned odouras a criterion during food search (Provecho & Josens 2009).Honeybees and bumblebees can also learn odours inside the hive,from the nectar or pollen clinging onto the body of the successfullyreturning foragers, and retrieve olfactory memories outside duringforaging at a later point in time (von Frisch 1967; Wenner et al.1969; Farina et al. 2005; Molet et al. 2009). Although our resultsclearly demonstrate that the odour of a plant found outside the nestsuffices for its identification and avoidance by workers, when sucha plant had detrimental effects on the fungus garden, the under-lying mechanisms leading to this response remain elusive. Ourexperimental approach was not conceived to address acquisition orretention performance at the individual level using controlledlearning paradigms, as in recent studies on ant appetitive learning(Dupuy et al. 2006; Kleineidam et al. 2007; Josens et al. 2009), sothe experiential background of the individually tested foragers wasunknown. This may also be one reason why a minority of workersstill accepted the unsuitable plant species in the treatment series:

N.Saverschek, F.Roces / Animal Behaviour 82 (2011) 453e458 457
they may have not experienced its detrimental effects on thefungus, thus continuing to evaluate the encountered plant disc (orperceived odour) as suitable.
One possible explanation for the delayed plant avoidanceobserved in our study is aversive learning. Workers may haveassociated the plant odour with the negative effects of the plant onthe fungus, mediated by the fungicide added to the plant tissue.Upon detection of the plant odour, workers recalled the informa-tion about plant suitability for the fungus, steering away from it.Odour aversion after olfactory learning has recently been reportedin a detailed investigation on honeybees (Carcaud et al. 2009).Individuals bees trained to associate an odour with an electricshock, using the sting extension reflex (Núñez et al. 1983) asa conditioning protocol, avoided such odour in a choice situation. Inour study, leafcutter ants recalled information about the unsuit-ability of a substrate gained inside the nest, resulting in theavoidance of the odour of that substrate during foraging thefollowing day. Unlike honeybees in the study mentioned above,leafcutter ants do not experience the negative effects themselves,and the aversive responses to a specific plant odour are broughtabout through the effects of the plant substrate on their symbioticfungus. Because of lack of direct evidence for the occurrence ofaversive learning at the individual level, however, these argumentsremain a working hypothesis worth pursuing.
Another mechanism that could potentially account for theobserved delayed avoidance of previously suitable plants is latentinhibition, as known in honeybees (Fernández et al. 2009; Chandraet al. 2010). Latent inhibition allows animals to ignore stimuli thatdo not predict relevant events. In our study, foragers might havelearned only that the particular odour does not mean food. Thisexplanation, however, appears very unlikely. First, colonies harvestseveral plant species simultaneously, so workers inside a nestchamber are exposed to a mixture of odours arising from bothsuitable and unsuitable plant species, making latent inhibitiontowards a specific olfactory stimulus improbable. And second,experiments with the leafcutter ant A. sexdens rubropilosa haveshown that in the context of avoidance of previously acceptedplants, highly volatile compounds within the nest do not affectforaging decisions if naïve workers are prevented from interactingwith the symbiotic fungus (North et al. 1999).
It is known that foraging leafcutter ants orient upwind to odourstimuli (Littledyke & Cherrett 1978; Therrien & McNeil 1990) anduse learned odours in an appetitive context as decision criteriaduring plant selection: during recruitment in A. lundi, recruits learnthe odour of the leaf fragment carried by the scout worker, and usethe learned cue to orient to and decide which material should becollected at the food source (Roces 1990, 1994). The power ofolfactory learning in Acromyrmex leafcutter ants is suggested by thefact that only one learning event suffices to direct the response ofworkers to a fragment with the associated odour (Roces 1990;1994). The present results go beyond these findings by showingthat in the context of foraging, individuals may also avoid thedirection from which the odour of an already-known unsuitablesubstrate comes, moving away from it, underlining the involve-ment of not only appetitive, but also aversive learned responsesduring plant recognition by foragers. The ability of foraging ants tolearn about changes in the palatability of the available plantsthrough their effects on the fungus, and recognize the specificunsuitable plant species, can be regarded as main adaptations toprotect the mutualistic fungus against loading with harmfulcompounds, thus assuring fungal growth and ultimately colonyfitness. In highly diverse, spatially and temporarily variable envi-ronments, with many fungicidal plant compounds potentiallyunknown to the ants, it is advantageous for foragers to be able toavoid unsuitable plants through a mechanism that involves learned
responses to the effects of the plant on the fungus, and does not relyonly on the harmful effects that the plant compounds have on theindividuals directly.
Acknowledgments
We deeply thank H. Herz for inspiring initial work onAcromyrmex lundi. This research was supported by the GermanResearch Foundation (DFG), Sonderforschungsbereich 567/TPC4‘Mechanisms of Interspecific Interactions of Organisms’.
References
Berish, C. W. 1986. Leaf-cutting ants (Atta cephalotes) select nitrogen-rich forage.American Midland Naturalist, 115, 268e276.
Bernays, E. A. 1993. Aversion learning and feeding. In: Insect Learning (Ed. byD. R. Papaj & A. C. Lewis), pp. 1e17. New York: Chapman & Hall.
Camargo, R. S., Forti, L. C., de Matos, C. A., Lopes, J. F., de Andrade, A. P. &Ramos, V. M. 2003. Post-selection and return of foragedmaterial by Acromyrmexsubterraneus brunneus (Hymenoptera: Formicidae). Sociobiology, 42, 93e102.
Carcaud, J., Roussel, E., Giurfa, M. & Sandoz, J. 2009. Odour aversion after olfactoryconditioning of the sting extension reflex in honeybees. Journal of ExperimentalBiology, 212, 620e626.
Chandra, S. B. C., Wright, G. A. & Smith, B. H. 2010. Latent inhibition in the honeybee, Apis mellifera: is it a unitary phenomenon? Animal Cognition, 13, 805e815.
Cherrett, J. M. 1989. Leaf-cutting ants. In: Ecosystems of the World 14B: Tropical RainForest Ecosystems (Ed. by H. Lieth & M. J. Werger), pp. 473e488. Amsterdam:Elsevier.
Dupuy, F., Sandoz, J.-C., Giurfa, M. & Josens, R. 2006. Individual olfactory learningin Camponotus ants. Animal Behaviour, 72, 1081e1091.
Farina, W. M., Grüter, C. & Diaz, P. C. 2005. Social learning of floral odours insidethe honeybee hive. Proceedings of the Royal Society B, 272, 1923e1928.
Fernández, V. M., Arenas, A. & Farina, W. M. 2009. Volatile exposure within thehoneybee hive and its effect on olfactory discrimination. Journal of ComparativePhysiology A, 195, 759e768.
von Frisch, K. 1967. The Dance Language and Orientation of Bees. Cambridge,Massachussetts: Harvard University Press.
Herz, H., Hölldobler, B. & Roces, F. 2008. Delayed rejection in a leaf-cutting antafter foraging on plants unsuitable for the symbiotic fungus. Behavioral Ecology,19, 575e582.
Howard, J. J. 1987. Leafcutting ant diet selection: the role of nutrients, water andsecondary chemistry. Ecology, 68, 503e515.
Howard, J. J.1988. Leafcutting ant diet selection: relative influence of leaf chemistryand physical features. Ecology, 69, 250e260.
Hubbell, S. P. & Wiemer, D. F. 1983. Host plant selection by an attine ant. In: SocialInsects in the Tropics (Ed. by P. Jaisson), pp. 133e154. Paris: Université Paris-Nord.
Josens, R., Eschbach, C. & Giurfa, M. 2009. Differential conditioning and long-termolfactory memory in individual Camponotus fellah ants. Journal of ExperimentalBiology, 212, 1904e1911.
Karban, R. & Myers, J. H.1989. Induced plant responses to herbivory. Annual Reviewof Ecology, Evolution and Systematics, 20, 331e348.
Kleineidam, C., Ruchty, M., Casero-Montes, Z. & Roces, F. 2007. Thermal radiationas a learned orientation cue in leaf-cutting ants (Atta vollenweideri). Journal ofInsect Physiology, 53, 478e487.
Knapp, J., Howse, P. & Kermarrec, A. 1990. Factors controlling foraging patterns inthe leaf-cutting ant Acromyrmex octospinosus (Reich). In: Applied Myrmecology:a World Perspective (Ed. by R. Vander Meer & K. Jaffe), pp. 382e409. Boulder,Colorado: Westview Press.
Krebs, J. R. & Inman, A. J. 1994. Learning and foraging: individuals, groups, andpopulations. In: Behavioral Mechanisms in Evolutionary Ecology (Ed. byL. A. Real), pp. 46e65. Chicago: The University of Chicago Press.
Littledyke, M. & Cherrett, J. 1978. Olfactory responses of leaf-cutting ants Attacephalotes (L) and Acromyrmex octospinosus (Reich) (Hymenoptera, Formicidae)in the laboratory. Bulletin of Entomological Research, 68, 273e282.
Meyer, S., Roces, F. & Wirth, R. 2006. Selecting the drought stressed: effects ofplant stress on intraspecific and within-plant herbivory patterns of the leaf-cutting ant Atta colombica. Functional Ecology, 20, 973e981.
Molet, M., Chittka, L. & Raine, N. E. 2009. How floral odours are learned inside thebumblebee (Bombus terrestris) nest. Naturwissenschaften, 96, 213e219.
Nichols-Orians, C. & Schultz, J. 1990. Interactions among leaf toughness, chemistryand harvesting by attine ants. Ecological Entomology, 15, 311e320.
North, R. D., Jackson, C.W. &Howse, P. E.1999. Communication between the fungusgarden and workers of the leaf-cutting ant Atta sexdens rubropilosa regardingchoice of substrate for the fungus. Physiological Entomology, 24, 127e133.
Núñez, J. A., Maldonado, H., Miralto, A. & Balderrama, N. 1983. The stingingresponse of the honeybee: effects of morphine, naloxone and some opioidpeptides. Pharmacology Biochemistry and Behavior, 19, 921e924.
Pretto, D. R. & Forti, L. C. 2000. Dyed baits distribution in nests of Atta sexdens L.(Hym., Formicidae). Journal of Applied Entomology, 124, 37e39.
Provecho, Y.& Josens, R.2009.Olfactorymemoryestablishedduring trophallaxis affectsfood search behaviour in ants. Journal of Experimental Biology, 212, 3221e3227.

N.Saverschek, F.Roces / Animal Behaviour 82 (2011) 453e458458
Reichle, C., Jarau, S., Aguilar, I. & Ayasse, M. 2010. Recruits of the stingless beeScaptotrigona pectoralis learn food odors from the nest atmosphere. Natur-wissenschaften, 97, 519e524.
Ridley, P., Howse, P. E. & Jackson, C. W. 1996. Control of the behaviour ofleaf-cutting ants by their ‘symbiotic’ fungus. Experientia, 52, 631e635.
Roces, F. 1990. Olfactory conditioning during the recruitment process in a leaf-cutting ant. Oecologia, 83, 261e262.
Roces, F. 1994. Odour learning and decision-making during food collection in theleaf-cutting ant Acromyrmex lundi. Insectes Sociaux, 41, 235e239.
Saverschek, N., Herz, H., Wagner, M. & Roces, F. 2010. Avoiding plants unsuitablefor the symbiotic fungus: learning and long-term memory in leaf-cutting ants.Animal Behaviour, 79, 689e698.
Therrien, P. & McNeil, J. N. 1990. The discovery of new recources and subsequenttrail formation by Acromyrmex octospinosus in Guadeloupe. In: Applied Myrme-cology: a World Perspective (Ed. by R. K. Vander Meer & K. Jaffé), pp. 373e381.Boulder, Colorado: Westview Press.
Wenner, A., Wells, P. & Johnson, D. 1969. Honey bee recruitment to food sources:olfaction or language? Science, 164, 84e86.
Wirth, R., Herz, H., Ryel, R., Beyschlag, W. & Hölldobler, B. 2003. Herbivory of Leaf-cutting Ants: a Case Study on Atta colombica in the Tropical Rainforest of Panama.Berlin: Springer.
Wolf, H. & Wehner, R. 2000. Pinpointing food sources: olfactory and anemotacticorientation in desert ants, Cataglyphis fortis. Journal of Experimental Biology, 203,857e868.