Download - Taxonomy of six Antarctic anascan Bryozoa

http://journals.cambridge.org Downloaded: 11 Dec 2013 IP address: 41.215.33.66
Antarctic Science 5 (2): 129-136 (1993)
Taxonomy of six Antarctic anascan Bryozoa P.J. HAYWARD and J.S. RYLAND
Marine, Environmental and Evolutionary Research Group, School of Biological Sciences, University College of Swansea, Singleton Park, Swansea SA2 8PP, UK
Abstract: Six species of anascan cheilostome Bryozoa are described from Antarctica. Apiophragma gen.nov. is introduced for Megapora hyalina Waters, 1904. Cellaria dennanti Waters, 1904 non MacGillivray, 1895 is redescribed as Swanomia belgica sp.nov. Paramawsonia Androsova, 1972 is restricted to its type species, Cellaria dennanti MacGillivray, 1895. Micropora notialis sp.nov. is distinguished from M. brevissima Waters, 1904. Two new species are described in the genus Cellaria.
Received 5 February 1992, accepted 29 May 1992
Key words: Bryozoa, Cheilostomata, Antarctica
Introduction
The cheilostome bryozoan fauna of Antarctica is rich in species and displays a high degree of endemicity. In recent years the list of endemic species compiled by Rogick (1965) has been augmented by numerous new, or newly recognized species (e.g. Hayward & Ryland 1990, Hayward 1992). Her total of 321 species reported from Antarctica has been considerably modified as the taxonomic identities of many species reported by earlier authors have been reassessed (e.g. Hayward & Thorpe 1989, Hayward & Ryland 1992). The taxonomic descriptions of early authors (e.g. Waters 1904) are frequently brief, and inadequate for the purposes of modem systematic research. Further, while some especially distinctive species are readily recognized from published illustrations, in many other cases morphological details now known to be important in cheilostome taxonomy cannot be distinguished in original drawings, and it is necessary to refer to type and other original specimens when conducting taxonomic research. The need for taxonomic precision in marine ecology does not have to be emphasized, and for many bryozoan faunas will only be achieved following a careful systematic reappraisal of all previously described species. Faunal records unsubstantiated by descriptions, illustrations orpreservedspecimens must often beviewed with scepticism.
The anascan cheilostomes of Antarctica are reasonably well known. The abundant and diverse assemblages of erect cellularines, so conspicuous in benthic samples, are well described by Kluge (1914) and Hastings (1943), and membraniporine and flustrine species describedby the former author may be recognized from his illustrations. However, a number of species describedby Waters (1904) have continued to pose taxonomic problems, and new species continue to be discovered in cryptic communities. Six species of anascan cheilostomes are described in this paper, from various Antarctic localities. Micropora brevissima Waters, 1904 is shown to comprise two very similar species, and the Cellaria dennanti described by Waters (1904) from the Bellingshausen Sea
proves to be a different species from the Australian fossil species C. dennanti MacGillivray, 1895. Megapora hyalina Waters, 1904 is redescribed as type species of the new genus Apiophragrna, and two new specieso f Cellaria are introduced.
Materials
Materials usedin thisstudy originated fromthevery substantial accumulations of Antarctic Bryozoa formerly held by the Institute of Oceanographic Sciences, Deacon Laboratory, Wormley, augmented by newly collected samples from Signy Island, supplied by the British Antarctic Survey. Additional collections of Bryozoa from the Ross Sea weremadeavailable for study by the New Zealand Oceanographic Institute, Wellington, through the courtesy of Dr. D.P. Gordon. A preliminary examination of the collections made by the 1989 EPOS 3 cruise in the Weddell Sea yielded a further sample of Cellaria incula, sp.nov. The type specimen of Micropora brevissiina Waters was kindly lent by the Institute Royale des Sciences Naturelles, Bruxelles. All 10s and BAS specimens listed have been deposited in the Natural History Museum, London, and registration numbers are indicated by the prefix BMNH. Details of all collecting stations are given in Appendix I.
Apiophragma gen.nov
Diagnosis: Colony encrusting; autozooids with short gymnocyst, continuous with incurved lateral walls; cryptocyst extensive, with reduced opesia comprising less than one- third total autozooid length, pierced by a single pair of opesiules; bordered by a raised, crenellated, mural rim; opesia reduced, but larger than operculum, with apair of stout condyles for articulation of the operculum; oral spines present; no avicularia; basal pore chambers present; ovicell hyperstomial, imperforate.
Type species: Megapora hyalina Waters, 1904.
Remarks: The boreal/Arctic species Megapora ringens (Busk)
129

http://journals.cambridge.org Downloaded: 11 Dec 2013 IP address: 41.215.33.66
130 P.J. HAYWARD and J.S. RYLAND
resembles M. lzyalina in its depressed cryptocyst, thickly calcified mural rim, reduced opesia with lateral condyles for the articulation of the operculum, and distal arc of spines (see Ryland & Hayward 1977: 1, p.107). However, the strongly trifoliate opesia of M. ringens, with the proximo-lateral notches accommodating the depressor muscles of the frontal membrane, suggests an affinity with Amplaiblestrum Gray and Ramphonotus Norman. The opesia ofM. hyalina is much smaller, narrowly bell-shaped, and the frontal wall is pierced by paired opesiules, recalling the morphology of the microporidan genus Manzonella Jullien (Gordon 1984). In Manzonella, the opesia is exactly coincident with the operculum, whereas in Megapora hyalina a short area of frontal membrane persists proximal to the pronounced condyles marking the proximal corners of the operculum. The new genus Apiophragma (from apios (Gr.), a pear, andplzragma (Gr.), a wall, referring to the shape of the mural rim)is accordingly introduced for M. lzyalina Waters, within the family Microporidae Gray. The ovicell of M. lzyalina was figuredby Waters(l904)but not described, anditsmorphology is still incompletely known.
Apiophragma hyalina (Waters) Fig. 1.
Megapora hyalina Waters, 1904: 39, p1.2, figs.l3a,b; text -fig. 1.
Material: Discovery Stn. 1948.
Descr@tion: Autozooids oval, 0.5 x 0.4 mm, separated by deep grooves. Gymnocyst and lateral walls distinct, smooth; frontal membrane bounded by a thickly crenellate mural rim;
cryptocyst flat, granular. Opesia comprising less than one- third total frontal length of autozooid, more or less bell- shaped, with straight proximal edge, and stout, conspicuous condyles at the point of constriction on each side. Opesiules elongate oval, situated about halfway along length of cryptocyst. Six short, stout spines disposedin an arc at distal end of autozooid. Ovicell (figured by Waters (1904)) prominent, elongate oval, perhaps with a narrow, elongate frontal fenestra.
Remarks: Waters (1904) recorded this species from two stations in the Bellingshausen Sea. A single small colony was recovered from Discovery Stn. 1948, off the South Shetland Islands, and there have been no other published reports of it. Ovicells were not present in the 'Discovery' specimen and remain to be properly described.
Micropora brevissima Waters Fig. 2d,e
Micropora brevissima Waters, 1904: 40 (in part), p1.2, figs.7a-c
? not Micropora brevissima: Waters, 1905: 234
Micropora brevissima: Hayward & Thorpe, 1989: 937 (in part), not figs.6a,b
Material: COTYPE: Institute Royale, Bruxelles no.596, 70°23'S, 82"47'W, Expedition Antarctique Belge.
Other material: Terra Nova stns. 194,316. Discovery Stns. 39,159,170,190,363,474,1948,1958, WS482. BANZARE Stn. 39/40.
Fig. 1. Apiophragma hyalina. a., View of the entire colony, x 50. b. detail to show a single autozooid, x190.

http://journals.cambridge.org Downloaded: 11 Dec 2013 IP address: 41.215.33.66
ANTARCTIC BRYOZOA 131
Remarks: Waters' (1904) original account of this species is unsatisfactory, and has been a persistent source of confusion when trying to identify it again. It is apparent that he founded the diagnosis of M. brevissirna partly upon material collected by the Expedition Antarctique Belge in the Bellingshausen Sea, and partly upon specimens from Cape Horn collected by the 'Romanche', which Waters examined during his preparation of the 'Belgica' report. He notes (Waters, 1904, p.40) that certain features, such as the pronounced knobs adjacent to theopesia are present only inCapeHornspecimens, but refers to the presence of oral spines without stating whether they occur in all of the specimens examined by him. His fig. 7a & 7b are drawn from 'Belgica' material while fig. 7c is of an avicularian mandible from a Cape Horn specimen. Hayward & Thorpe (1989) accepted the wide range in morphological characteristics implied in Waters' account, and the 'Discovery' specimens assigned to this species revealed a geographical distribution even broader than that established by Waters, from Cape Horn to the Falkland Islands, and from the Palmer Archipelago to the Ross Sea. However, further material from the 'Discovery' collections, together with fresh specimens from Signy Island suggested that this morphological variation might be discontinuous, and SEM micrographs of a larger number of specimens, over the whole of the supposed geographical range of M. brevissima have established conclusively that two rather similar but nonetheless distinct species may be recognized.
The only remaining type specimen of M. brevissima, that figured by Waters (1904), is unfortunately in apoor condition. It consists of just seven autozooids, several of which are incomplete. The ovicell figured by Waters is now missing as is the single avicularium; both of these structures, it should be noted, were depicted on the periphery of the colony. However, the dimensions of the autozooids and their opesiae areclosely similartospecimens from theSouthShetland Isles and from Enderby Land here assigned toM. brevissima. The opesia in all cases is 2.5 - 3 times as wide as long; the almost flat frontal wall has about 40 small pores; the avicularium measures 0.12-0.15 mm in length, scarcely longer than the width of the opesia; the ovicell has a smooth band of calcification bounding the aperture, and is often surmounted by an avicularium. Finally, as noted by Waters, the lateral walls are not noticeably thickened adjacent to the opesia. Contrary to the impression given by Waters, there are no oral spines. Specimens listed above as conformingto this diagnosis establish a purely Antarctic distribution for M. brevissima Waters, ranging from the Palmer Archipelago to the Ross Sea. A suite of specimens from Cape Horn, the Falkland Islands, Signy Island and the Palmer Archipelago, some of which were assigned toM. brevissima by Hayward & Thorpe (1989) are now recognized as differing from M. brevissima in the proportions of the opesia, the presence of paired distal oral spines in early ontogeny, by a tendency to thickening of the lateral walls adjacent to the opesia, by a proportionately
larger avicularium, and in the ovicell, which rarely has an avicularium distal to it. These specimens are here assigned to Micropora notialis sp.nov.
Measurements (means and standard deviations), mm:
M. brevissima M. brevissima &type Disc. Stn. 1948
n n Autozooid length 7 0.65k0.05 20 0.6320.07 Autozooid width 7 0.4520.03 20 0.44k0.05 Opesia length 6 0.06+0.008 20 0.05k0.004 Opesia width 6 0.16+0.005 20 0.1520.009 Avicularium length 20 0.1420.01
Micropora notialis sp.nov. Fig. 2a-c
Micropora brevissima Waters, 1904: 40 (in part)
?Micropora brevissima: Waters, 1905: 234
Micropora brevissima: Hayward & Thorpe, 1989: 937 (in part), figs.6a & b.
Holozype: BMNH reg.no.1992.1.17.3, Operation Tabarin, Stn. 1541.
Paratype: BMNH reg.no.1992.1.17.4, Powell Rock, Signy Island, 3.3.1986, British Antarctic Survey.
Other Material: Discovery Stns. 388,1948, WS85, WS221. BANZARE. Stn. 42. Tabarin Stn. 1541.
Description: Colony forming broad, thin, spreading sheets on hard substrata. Autozooids oval to hexagonal; rounded distally, tapered proximally, separated by indistinct sutures; rather flat, with just the central area of the frontal wall gently convex. Opesia semicircular, its width slightly more than twice its length, proximal edge straight, disto-lateral border smoothly rounded. Two basally-jointed, cylindrical spines developed on distal edge of opesia, widely spaced; present only in first few generations of autozooids from growing edge, lost in later ontogeny, and the bases obscured by calcification. Lateral walls of autozooid raised adjacent to proximal ccrners of opesia, forming a pair of prominent knobs. Cryptocyst finely granular, depressed peripherally, convex medially, with 20-40 small, simple pores; opesiules large, oval, each with a single stellate pore medio-proximal to it, obscured in later ontogeny. Avicularia sparsely distributed, often very infrequent, each at distal end of an autozooid; rostrum acute to frontal plane, disto-laterally directed, triangular, with hooked tip; no palate below mandible; proximal opesia wide, crescentic, with broad, cryptocystal rim below a complete, stout crossbar. Ovicell longer than broad, convex, prominent; immersed in distally succeeding autozooid, the cryptocyst of which is continuous with the distal portion of the ovicell; aperture bounded by a rim of smooth calcification, peaked medially and sometimes developing a low umbo.

http://journals.cambridge.org Downloaded: 11 Dec 2013 IP address: 41.215.33.66
132 P.J. HAYWARD and J.S. RYLAND
Fig. 2. a-c, Micropora notialis: a. Autozooids at the growing edge, showing distal oral spines, x 115 (specimen from Antarctic Peninsula); b. Group of ovicelled zooids, x 80 (specimen from off Lively Island, East Falkland); c. An avicularium, x 90 (specimen from Signy Island). d-e Micropora brevissima: d. Group of ovicelled autozooids, the ovicells capped by small avicularia, x 90 (specimen from South Shetland Islands); e. Autozooids and avicularia from close to the colony edge, x 90 (specimen from Enderby Land).

http://journals.cambridge.org Downloaded: 11 Dec 2013 IP address: 41.215.33.66
ANTARCTIC BRYOZOA 133
Measurements (means and standard deviations of 20 values), mm:
Autozooid length 0.82+0.06 Autozooid width 0.4320.04 Opesia length 0.1020.006
Avicularium length 0.3420.03
Remarks: Micropora notialis is confusingly similar to M. brevissima Waters. It is distinguished by its relatively larger, longer opesia, which does not have the quadrangular appearance of that of M. brevissima, and by its much larger avicularium. Actively growing colonies of M. notiatis may also be recognized by the presence of paired spines on the autozooids close to the growing edge. However, the most constant distinction lies in the ovicell, which in M. notiulis is immersed in the cryptocystal calcification of the distally succeeding autozooid, but in M. brevissima is usually intimately associated with an avicularium.
The geographical ranges of the two species overlap. M. notialis occurs from Cape Horn to the Falkland Islands, and off the southern Patagonian Shelf, and ranges south to South Georgia and the Palmer Archipelago. M. brevissima is widespreadin Antarcticshelfwaters, including those of the Antarctic Peninsula, but perhaps reaches its northern limit at South Georgia.
Micropora stenostoma (Busk), from Tasmania, resembles M. notialis in possessing spines. However, in this species a pair of stout, inflated, lateral oral spines is found on all autozooids, including those in late ontogeny. M. steizostonza (syntypes: BMNH 1854.11.15.217, 1892.7.18.1) also has smaller autozooids than M. notialis (0.45 x 0.25 mm) and proportionately smaller avicularia; further, its relatively larger ovicells are borne by dimorphic autozooids with orifices twice as wide as non-brooding autozooids. Busk (1854)listedaspecimen ofM. stenostorna from East FalMand (BMNH 1899.7.1.1479); this consists of a decalcifiedbalsam mount of fewer than 30 autozooids. Autozooids measure 0.5 x 0.35 mm, several bear spines, and orifice dimensions are 0.10 x 0.15 mm. This specimen may be a fragment of M. notialis, butshows insufficient detail for absolutecertainty.
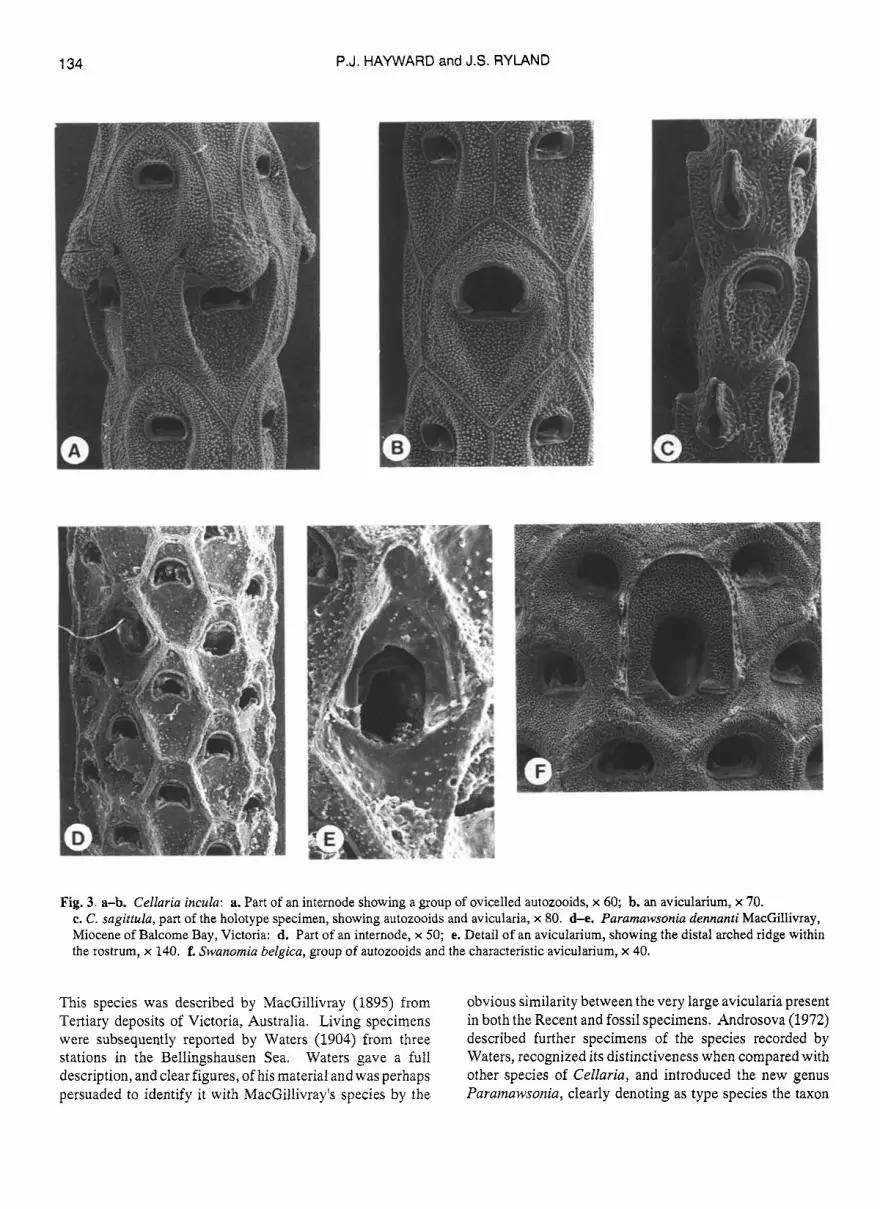
Cellaria incula s p a . Fig. 3a,b
Opesia width 0.2120.01
Holotype: BMNH reg. no.1992.1.17.1, BANZARE Stn. 39/40
Other material: EPOS 3 Stn. 271.
Colony developing a delicate, diffuse, rather spindly form; exceeding 5 cm high, often richly branched. Internodes commonly 10 mm long, 0.8-1.0 rnm wide, the nodes formed from bundles of intertwined rhizoids. Autozooids in alternating whorls of four, 0.7-0.8 x c. 0.4 mm; hexagonal, separated by distinct, thin, mural rims; cryptocyst granular, the central area concave and flanked by curving cryptocystal ridges, which do not meet either proximally or distally.
Opesia in distal half of autozooid, but distant from distal edge; semicircular, with a straight proximal edge within which is visible astout articular plate, its corners rounded and projecting. Avicularia infrequent; vicarious, as large as an autozooid, with a hood-like, semicircular rostrum, markedly larger than an autozooid opesia, projecting from the frontal plane of the internode. Ovicell with a rounded, cowl-like distal hood overhanging the aperture.
Remarks: The projecting, distal hood overhanging the ovicell aperture in this species recalls those seen in species of theunjointed cellariid genus Stomhypselosaria. In C. incula, however, the colony consists of slender internodes linked by conspicuous cuticular nodes, a feature characteristic of all species of Cellaria. The opesia, cryptocystal ridges and avicularia of C. incula are all also typical for the genus Cellaria. This species is immediately distinguished from all other southern hemisphere species of Cellaria by its rounded avicularium rostrum, and by the prominent, anvil-shaped articular plate within the opesia, as well as by its unusual ovicell hood. Generic distinctions within the Cellariidae are in need of systematic revision.
Cellaria sagittula sp.nov. Fig. 3c
Holotype: New Zealand Oceanographic Institute reg. no. H-591, NZOI Stn. A534
Description: Internode slender, curved, with autozooids disposed in triple whorls. Autozooids 0.4-0.5 x 0.2mm, with coarsely granular calcification; a complete, oval cryptocystal ridge projecting conspicuously from the frontal plane enclosing an area 0.3 x 0.15 mm. Opesia in distal third of autozooid, wider thanlong, the disto-lateral rim more or less semicircular, the proximal rim developed as a projecting, convex lip; an inconspicuous denticle present in each proximo-lateral corner. Avicularia interzooidal, each intercalated within an autozooid whorl; rostrum prominent, distally directed, shaped like a slender arrow head. Ovicells not present.
Remarks: This distinctive species is represented by the holotype specimen only, a single basal internode 5 mm long, with the ancestrula and anchoring rhizoids still intact. Its autozooids are the smallest of any Antarctic or subantarctic species of Cellaria, and it is immediately recognized by its projecting cryptocystal ridge and its characteristic avicularia. Colony form remains unknown, as does the morphology of the ovicell, and more material is required before a complete descriptive account can be given. C. sagittulu is probably inconspicuous among mixed samples of erect, bushy anascans, yet differs so markedly from other describedspecies that there should be no difficulty in recognizing it again.
Parainawsonia denizanti (MacGillivray) Fig. 3d & e.
Cellaria denizanti MacGillivray, 1895: 31, p1.22, figs.l4,14a
http://journals.cambridge.org Downloaded: 11 Dec 2013 IP address: 41.215.33.66
134 P.J. HAYWARD and J.S. RYLAND
Fig. 3. a-b. Cellaria incula: a. Part of an internode showing a group of ovicelled autozooids, x 60; b. an avicularium, x 70. c. C. sagittula, part of the holotype specimen, showing autozooids and aviculai-ia, x 80. d-e. Paramawsonia dennanti MacGillivray, Miocene o f Balcome Bay, Victoria: d. Part of an internode, x 50; e. Detail of an avicularium, showing the distal arched ridge within the rostrum, x 140. f. Swanomia belgica, group of autozooids and the characteristic avicularium, x 40.
This species was described by MacGillivray (1895) from Tertiary deposits of Victoria, Australia. Living specimens were subsequently reported by Waters (1904) from three stations in the Bellingshausen Sea. Waters gave a full description, and clear figures, of his material and was perhaps persuaded to identify it with MacGillivray's species by the
obvious similarity between the very large avicularia present in both the Recent and fossil specimens. Androsova (1972) described further specimens of the species recorded by Waters, recognized its distinctiveness when compared with other species of Cellaria, and introduced the new genus Paramawsonia, clearly denoting as type species the taxon

http://journals.cambridge.org Downloaded: 11 Dec 2013 IP address: 41.215.33.66
ANTARCTIC BRYOZOA 135
Cellaria dennanti MacGillivray 1895. An apparent link between early Tertiary faunas of southern Australiaand those of present day Antarctica is of great interest. Unfortunately, re-examination of MacGillivray's species, and comparison with 'Discovery' specimens of Antarctic 'Paramawsonia dennanti' shows that two separate species have been confused. MacGillivray's illustration (1895, p1.22, fig. 14a) shows three autozooids together with a single avicularium. However, the avicularium was obviously badly worn, to the extent that the triangular rostral rim was largely removed, leaving an arched, semielliptical shape which Waters (1904, p1.2, fig.9a; p1.8, fig.4) identified with thesemielliptical rostrum of his material. SEM micrographs of Miocenespecimens of Cellaria denriartti (Fig. 3d & e) from Balcome Bay, Victoria, show the actual shape of a relatively unworn avicularium; the rostral rim is broadly triangular, but basal to it is an archedridge, apparently continuous with the condyles on which the mandible of the avicularium pivoted. Abrasion of the rostrum would result in this ridge becoming prominent, and approximating to the rostral rim of the much larger avicularium seen in Waters' species (Fig. 39.
Paramawsonia Androsova, 1972 is defined by its type species, Cellaria dennunti MacGillivray, 1895. At present, generic distinctions within the Cellariidae are rather indistinct, and the taxonomic status of Pararnawsoitia should only be decided following a revision of the genera presently included within the family. However, the species described by Waters (1904) and Androsova (1972) belongs to neither genus, and is specifically distinct from Cellaria dennanti. In its deeply concave, thickly calcified cryptocyst, relatively large opesia, with especially pronounced condyles, andits grossly enlarged avicularium, this species accords with the diagnosis of Swanomia Hayward and Thorpe, 1989 (= Mawsortia Livingstone, 1928) to which it is here assigned.
Swanomia belgica spnov. Fig. 3f
Cellaria dennanti Waters, 1904: 36, p1.2, figs.9a-f; p1.8, fig.4
Paramawsonia dennanti: Androsova, 1972: 340, fig.14.
Holotype: BMNH reg.no.1992.1.17.2, Discovery Stn. 187.
Other material: Discovery Stns. 172, 177, 1948.
Description: Colony with stout, cylindrical or nodulated stems 3 mm wide, branching dichotomously at infrequent intervals, height exceeding 40 mm; attached by chitinous rhizoids. Autozooids in alternating whorls of 10-12, hexagona1,separatedby thin,raisedsutures, 0.8-1.OxO.6-0.8 mm. Cryptocyst depressed, with thick, finely nodular calcification. Opesia one-third total autozooid length, slightly wider than long, with finely crenellate rim; proximal border
straight, with a widely-spaced pair of knob-like condyles, linked by a low ridge. Avicularia frequent, larger than autozooids, with an evenly concavecryptocyst, andan elongate oval, central opesia; condyles short, thick and blunt, rostrum semielliptical, equivalent to two-thirds of length of avicularium. Ovicells undescribed.
Remarks: Waters' (1904) material originated from three stations in the Bellingshausen Sea, Androsova's (1972) from nine stations in western Antarctica, from Grahamland to the Ross Sea.
Acknowledgements
We are grateful to the Trustees of the Leverhulme Trust for generous financial support of this work, and to the Institute of Oceanographic Sciences, Deacon Laboratory, and the British Antarctic Survey, for providing the specimens. We thank Dr. J.J. Lopez Gappa for drawing our attention to Micropora stenostoma (Busk).
References
ANDROSOVA, E.I. 1972. Bryozoa Cheilostomata Anasca of the Antarctic and Subantarctic. Issledovaniia Fauni Morei, 11, 315-344.
BUSK, G. 1854. Catalogue of marine Polyzoa in the collection of the British Museum. 11. Cheilostomata @art). London: Trustees of the British Museum,
GORDON, D.P. 1984. The marine fauna of New Zealand: Bryozoa: Gymnolaemata from the Kermadec Ridge. New Zealand Oceanographic Institute, Memoir, 91, 1-144.
HASTINGS, A.B. 1943. Polyzoa (Bryozoa) I. Discovery Reports, 22,301-510. HAYWARD, P.J. 1992. Some Antarctic andsub-Antarctic speciesof Celleporidae
(Bryozoa, Cheilostomata). Journal of Zoology, London, 226,283-310. HAYWARD, P.J. & RYLAND, J.S. 1990. Some Antarctic and Subantarctic species
of Microporellidae (Bryozoa: Cheilostomata).Journal ofNatura1 History,
HAYWARD, P.J. & RYIAND, J.S. 1991. New and little-known species of Cheilostomata (Bryozoa) from Antarctica and the Subantarctic southwest Atlantic. Bulletin du Museum National d'Histoire Naturelle, Paris, 13,
HAYWARD, P.J. & THORPE, J.P. 1989. Membraniporoidea, Microporoidea and Cellarioidea (Bryozoa, Cheilostomata)collected by Discovery Investigations. Journal of Natural History, 23,913-959.
KLUGE, G.A. 1914. Die Bryozoen der deutschen Siidpolar-Expedition, 1901-3. I. Deutsche Siidpolar-Expedition, 14, 601-678.
MACGILLIVRAY, P.H. 1895. A monograph of the Tertiary Polyzoa of Victoria. Transactions of the Royal Society of Victoria (ns.), 4, 1-166.
RoGIcK,M.D. 1965. BryozoaoftheAntarctic. In P.VwOm& J.VANMW~HEM eds., Biogeography and Ecology in Antarctica. Monographiae Biologicae,
RYLAND, J.S. & HAYWARD, P. 1977. British Anascan Bryozoans. Synopses of the British Fauna (n.s.), 10, 1-188.
WATERS, A.W. 1904. Bryozoa. Resultats du Voyage du S.V. Belgica: Zoologie, 4, 1-114.
WATERS, A.W. 1905. Bryozoa from near Cape Hom. Journal of the Linnean Society, Zoology, 29,230-251.
55-120.
24,1263-1287.
24 1-261,
IS, 401-413.
http://journals.cambridge.org Downloaded: 11 Dec 2013 IP address: 41.215.33.66
136 P.J. HAYWARD and J.S. RYLAND
Appendix I. Details of collecting stations
a. "Discovery" Stations
39 159 170 172 177 187 190 363 388 474 1948 1958 WS85 ws221 WS482
25.3.26 21.1.27 23.2.27 26.2.27 5.3.27 18.3.27 24.3.27 26.2.30 16.4.30 12.11.30 4.1.37 .2.37
25.3.27 4.6.28 16.11.28
b. "Terra Nova" Stations
194 22.02.11 316 09.02.11
c. B.A.N.Z.A.R.E. Stations
39/40 17.01.30 42 26.01.30
East Cumberland Bay, South Georgia. Off South Georgia. Off Clarence Island. South Shetlands. Off South Shetlands. Neumayer Channel, Palmer Archipelago. Bismarck Strait, Palmer Archipelago. Off Zavodovski Island, South Sandwich Islands Off Cape Hom, Off Shag Rocks, South Georgia. Off Clarence & Elephant Islands. South Orkney Islands. Off Lively Island, East Falkland. Patagonian Shelf.
Off Oates Land. Off Glacier Tongue, McMurdo Sound.
d. Operation Tabarin, Stn.1541: Boat Harbour, Goudier Island
e. New Zealand Oceanographic Institute
Stn.A534
f. EPOS3 Stn.271 12.02.89
53"52.5'S, 36'08'W. 6I025.5'S, 53'46'W. 62"59'S, 60'28'W. 63"17.5'S, 61'17'W.
56°19.5'S, 67'09.75'W.
60°49.4'S, 52"40'W.
48"23'S, 65"lO'W. 63"10'S, 57"16.5'W.
69"43'S, 163"24'E.
Depth
179-235 m. 160 m. 342 m. 525 m. 1080 m. 259-354 m. 315 m. 329-278 m. 121 m. 199 m. 490-610 m.
79 m. 76-91 m. 125 m.
329-366 m. 348-457 m.
66"10'S, 49'41'E, to 66"12'S, 49"37'E. 300 m. 65'50'S, 54'23'E. 220 m.
77a36.7'S, 166"08'E, to 77"36'S, 166'12'E. 380-366 m.
73"17'S, 20°59.4'W, to 73'16.4'S, 20°54.6'W. 399-352 m.







