
Peripheral T-Cell Tolerance Associated with Prostate Cancer Is
Independent from CD4+CD25+ Regulatory T Cells
Elena Degl’Innocenti,1Matteo Grioni,
1Giusy Capuano,
1Elena Jachetti,
1Massimo Freschi,
2
Maria T.S. Bertilaccio,1Rodrigo Hess-Michelini,
1Claudio Doglioni,
2and Matteo Bellone
2
1Cancer Immunotherapy and Gene Therapy Program and 2Unita Operativa Anatomia Patologica, Istituto Scientifico San Raffaele,Milan, Italy
Abstract
CD4+CD25+Foxp3+ regulatory T cells (Treg) are thought tosuppress the natural and vaccine-induced immune responseagainst tumor-associated antigens (TAA). Here, we show thatTreg accumulate in tumors and tumor-draining lymph nodesof aging transgenic adenocarcinoma of the mouse prostate(TRAMP) male mice, which spontaneously develop prostatecancer. TAA overexpression and disease progression associatealso with induction of TAA-specific tolerance. TAA-specificT cells were found in the lymphoid organs of tumor-bearingmice. However, they had lost the ability to release IFN-; andkill relevant targets. Neither in vivo depletion of Treg by PC61monoclonal antibody followed by repeated vaccinations withantigen-pulsed dendritic cells nor the combined treatmentwith 1-methyl-L-tryptophan inhibitor of the enzyme indole-amine 2,3-dyoxigenase, PC61 antibody, and dendritic cellvaccination restored the TAA-specific immune response. Tregdid not seem to control the early phases of toleranceinduction, as well. Indeed, depletion of Treg, starting at week6, the age at which TRAMP mice are not yet tolerant, andprolonged up to week 12, did not avoid tolerance induction. Asimilar accumulation of Treg was found in the lymph nodesdraining the site of dendritic cell vaccination both in TRAMPand wild-type animals. Hence, we conclude that Treg accrualis a phenomenon common to the sites of an ongoing immuneresponse, and in TRAMP mice in particular, Treg aredispensable for induction of tumor-specific tolerance. [CancerRes 2008;68(1):292–300]
Introduction
With a population aging rapidly, prostate cancer is one of theleading causes of death in adult men (1). Indeed, prostatectomyand radiotherapy are potentially curative for organ-confineddiseases, and treatment of locally advanced and/or metastaticcancer remains palliative.
Because of the expression of unique tumor-associated antigens(TAA), prostate cancer is an ideal candidate for immunotherapy.Vaccination strategies were tested in phase I trials (2), and prostatecancer was the object of the first randomized phase III vaccine trial(3). Clinical results were encouraging but far from the predictionbased on animal models. Indeed, many of the preclinical models
were based on transplantable tumors (4), wherein in vitro passagesmay cause substantial genetic/epigenetic alterations of the tumorcells, and cell engraftment does not mimic the natural dynamicinteractions among neoplastic cells, stroma, and the immuneresponse (5). Also, those models may not correctly recapitulate themechanisms the tumor develops to escape natural and vaccine-induced T-cell responses (6).
Prostate cancer cells may lose expression of relevant antigens,acquire defects in antigen presentation, release immunosuppres-sive substances, block T-cell function favoring apoptosis, and,finally, promote development and recruitment of regulatory T cells(Treg) inside tumors and draining lymph nodes (7). The lattermechanism, especially the role of CD4+CD25+ Treg in cancer, hasbeen the focus of intense investigation in recent years.
Treg (5–10% of the peripheral CD4+ T cells) are mostly generatedin the thymus and represent an essential mechanism of peripheraltolerance to self antigens (8). Absence of Treg, which is causedeither genetically or by depletion, favors autoimmunity (9). Tregsuppress not only CD4+ T and natural killer cells but also CTL (10).Treg can be identified by several cell surface markers, among whichis the interleukin 2 (IL-2) receptor a-chain CD25. In addition, Tregselectively express Foxp3, a forkhead/winged helix transcriptionfactor that controls master genes in Treg development/function (8).
Treg can be also induced in the periphery from naiveCD4+CD25� T cells (11), although it is not yet known whetherthey substantially contribute to peripheral tolerance. Conversion ofCD4+CD25� T cells into Treg seems rather common in tumor-draining lymph nodes (TDLN; refs. 12–14).
Prostate cancer (15) and other neoplasms associate with Tregaccumulation in the blood and/or in tumors, and this mayinversely correlate with patients’ survival (16), therefore suggest-ing that Treg promote tumor-immune privilege. It remains to beelucidated whether their gathering directly affects the tumor-specific immune response. Data in animal models suggest a rolefor Treg in tolerance induction against TAA. Indeed, depletion ofTreg before tumor challenge favors induction of tumor-specificimmune responses and tumor eradication (17). Depletion of Tregalso increases the therapeutic index of several cancer treatments(18, 19). Those models, however, were based on transplantabletumors.
Transgenic adenocarcinoma of the mouse prostate (TRAMP)mice express the SV40 large T antigen (Tag) selectively on theprostate epithelium under the control of the rat probasin regulatoryelement, whose expression is influenced by sexual hormones (20).Hence, male mice remain healthy until puberty. In the followingweeks, TRAMP mice progressively overexpress Tag and invariablydevelop spontaneous mouse prostate intraepithelial neoplasia,adenocarcinoma, and seminal vesicles, lymph node, and visceralmetastases, therefore resembling human prostate cancer (21). Also,the immune response against Tag, a surrogate tissue-specific TAA
Requests for reprints: Matteo Bellone, Laboratory of Cellular Immunology, CancerImmunotherapy and Gene Therapy Program, 3P-A1, Dibit, Istituto Scientifico SanRaffaele, Via Olgettina 58, 20132, Milan, Italy. Phone: 39-02-2643-4789; Fax: 39-02-2643-4786; E-mail: [email protected].
I2008 American Association for Cancer Research.doi:10.1158/0008-5472.CAN-07-2429
Cancer Res 2008; 68: (1). January 1, 2008 292 www.aacrjournals.org
Research Article
Research. on February 10, 2016. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from
antigen in TRAMP mice, has been well characterized in othermodels and is dominated by CTL specific for the sequence 404 to411 (Tag-IV; ref. 22). In TRAMP mice, the immune response againstTag is characterized by thymic deletion of high-avidity CTL (23),which allows induction of low-avidity Tag-specific CTL in younghealthy mice, upon vaccination with Tag-IV–pulsed dendritic cells.Prostate cancer development and progression parallel induction ofa profound state of peripheral tolerance, which cannot be rescuedby dendritic cell vaccination (24).
We investigated whether Treg accumulate in the tumor andTDLN of TRAMP mice during disease development and whetherthey have a role in Tag tolerance.
Materials and Methods
Mice, tumor cell lines, and reagents. Heterozygous C57/BL6 TRAMPmice and wild-type (WT) mice were housed and bred in a specific
pathogen-free animal facility and treated in accordance with the European
Union guidelines and the approval of the Institutional Ethical Committee.Animals were typed for Tag expression by PCR-based screening assay.3
Isolation of mouse tail genomic DNA was performed as described (25).
RMA is a H-2b Rauscher virus–induced thymoma (26). B6/K-0 is a kidney
cell line expressing Tag (27). TRAMP-C1 is a prostate cancer, originatedfrom a TRAMP mouse (28). C26-GM (H-2d) is a colon carcinoma genetically
modified to produce granulocyte/macrophage colony-stimulating factor.
Unless specified, all chemical reagents were from Sigma-Aldrich, and
monoclonal antibodies (mAb) were from BD PharMingen.Dendritic cell preparation and immunization protocols. Dendritic
cells were prepared and characterized as previously described (29).
Dendritic cells were pulsed with 2 Amol/L Tag-IV (Research Genetics)
peptide for 1 h at 37jC, washed, and suspended at 1 � 106/mL in PBS.Dendritic cells (5 � 105) were injected i.d. into mice.
In vitro cytotoxicity assay. Splenocytes were restimulated in vitro in
the presence of 1 Amol/L Tag-IV peptide. Day 5 blasts were tested forcytolytic activity in a standard 4 h 51Cr release assay (29). 51Cr release of
target cells alone was always <25% of maximal 51Cr release (target cells
in 0.25 mol/L SDS).
Flow cytometry analyses. PE-labeled Kb/Tag-IV pentamers (ProIm-mune) were generated using a C411L-substituted synthetic peptide to
enhance binding to H2-Kb (30). Kb/OVA (SIINFEKL) pentamers were used as
negative control. Cells were incubated with the pentamer complex for
30 min at +4jC, and then, without washing, PerCP-Cy-5.5–conjugated anti-CD8 and the FITC-conjugated anti-CD44 mAbs were added for additional
15 min at +4jC. Dead cells were excluded by physical variables and/or
by the addition of ToPro-5 (Molecular Probes) immediately beforefluorescence-activated cell sorting analysis. For intracellular cytokine
measurement, day 5 blasts were stimulated in vitro with B6/K-0 or RMA
cells (1:1 ratio) or phorbol 12-myristate 13-acetate/ionomycin and stained
with FITC-labeled anti-CD44, PerCP-Cy 5.5–labeled anti-CD8, and antigen-presenting cells (APC)–labeled anti–IFN-g antibody as previously described
(29). For enumeration of CD4+CD25+Foxp3+ cells, TDLN and collagenase
D–digested prostates were processed on a cell strainer, stained with FITC-
labeled anti-CD4, PerCP-Cy 5.5–labeled anti-CD8 antibody, and APC-labeledanti-CD25 (clone PC61), permeabilized, and finally stained with PE-labeled
anti-Foxp3 (eBioscience) according to the manufacturer’s instructions. In all
experiments, cells were analyzed on a BD FacsCalibur.Isolation of CD4+CD25+ and CD4+CD25� cells and in vitro
proliferation assay. CD4+CD25+ T cells were isolated using a Treg isolation
kit (Miltenyi Biotec) following the manufacturer’s instructions. The three
cell populations (i.e., unfractionated CD4+ T cells, CD4+CD25+ cells, andCD4+CD25� cells) were cultured for 72 h in plates previously coated with
5 Ag/mL anti-CD3 mAb. The incorporation of [3H]thymidine by prolifer-
ating T cells (triplicate cultures) during the last 16 h of culture was
measured. Proliferation index was calculated assigning value of 1 to cpm
obtained from unstimulated cultures.
In vivo depletion of CD25+ cells and/or blocking of indoleamine 2,3-dyoxigenase enzyme. Five hundred micrograms of monoclonal anti-CD25
antibody (clone PC61; American Type Culture Collection) were injected i.p.
into mice at day �4. In selected experiments, treatment was repeated at day
+3 or every 2 weeks. Mice received 1-methyl-tryptophan dissolved indrinking water at 5 mg/mL as previously reported (31).
Histology and immunohistochemistry. Organs were fixed in 4%
formalin for 6 h, then embedded, and included in paraffin wax. H&Eand Tag staining of 5-Am-thick sections were performed as previously
described (24). For foxp3 staining, deparaffinized and rehydrated sections
were immersed in EDTA (pH 9.0) and followed the procedures described in3 As described in www.jax.org.
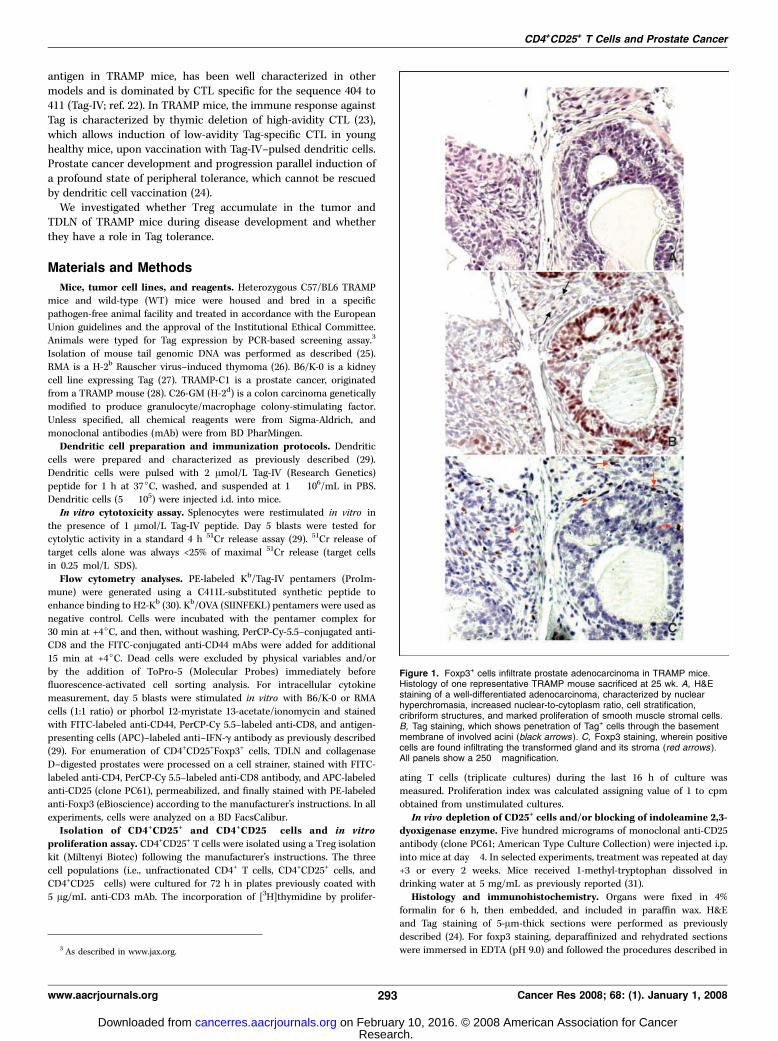
Figure 1. Foxp3+ cells infiltrate prostate adenocarcinoma in TRAMP mice.Histology of one representative TRAMP mouse sacrificed at 25 wk. A, H&Estaining of a well-differentiated adenocarcinoma, characterized by nuclearhyperchromasia, increased nuclear-to-cytoplasm ratio, cell stratification,cribriform structures, and marked proliferation of smooth muscle stromal cells.B, Tag staining, which shows penetration of Tag+ cells through the basementmembrane of involved acini (black arrows ). C, Foxp3 staining, wherein positivecells are found infiltrating the transformed gland and its stroma (red arrows ).All panels show a 250� magnification.
CD4+CD25+ T Cells and Prostate Cancer
www.aacrjournals.org 293 Cancer Res 2008; 68: (1). January 1, 2008
Research. on February 10, 2016. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from
ref. (24). Slides were incubated with streptavidin 1:1,000 for 30 min,followed by incubation with 3,3¶-diaminobenizidine tetrahydrochloride
for 5 min, and the counter stain was done with Mayer hematoxylin.
Macroscopic and microscopic specimens were evaluated by a pathologist
in a blind fashion. Histology sections were scored as previously described
(24) with partial modifications: the score of 5 was given to well-differentiated adenocarcinoma and 6 was given to metastases and/or
neuroendocrine tumors.
Statistical analysis. Statistical analyses were performed using Student’s
t test and log-rank test. Comparison of survival curves was consideredstatistically significant for P < 0.05.
Results
CD4+CD25+Foxp3+ Treg accumulate in prostates and TDLNof aged TRAMP mice. Firstly, we investigated whether Foxp3+ cellsaccumulate in prostates of TRAMP mice. The analysis was initiallyconducted in 25-week-old to 27-week-old mice. At this age, TRAMPmice present a significant enlargement of the urogenital apparatus(2.5 F 0.9 g; n = 6) when compared with age-matched and sex-matched WT littermates (0.9 F 0.2; n = 6; P < 0.008) and a well-differentiated adenocarcinoma (Fig. 1A) with penetration of Tag+
cells through the basement membrane of involved acini (Fig. 1B,black arrows). We also found several Foxp3+ cells infiltrating thetransformed gland and its stroma (Fig. 1C, red arrows), which wereabsent in WT prostates (not shown).
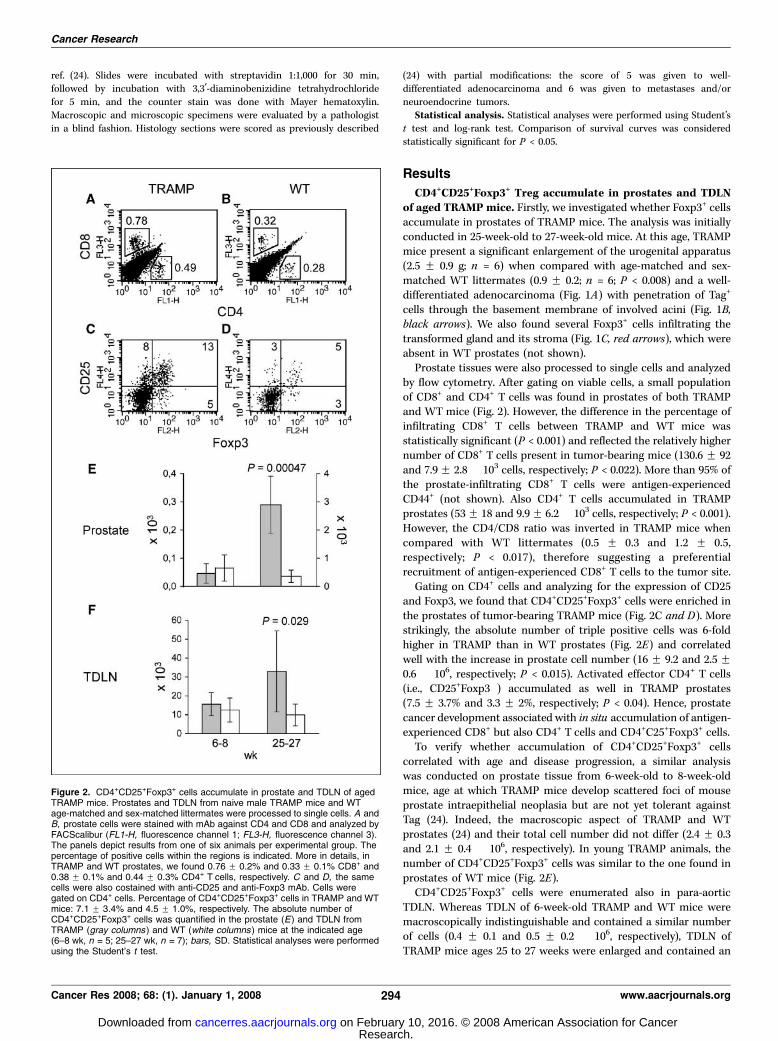
Prostate tissues were also processed to single cells and analyzedby flow cytometry. After gating on viable cells, a small populationof CD8+ and CD4+ T cells was found in prostates of both TRAMPand WT mice (Fig. 2). However, the difference in the percentage ofinfiltrating CD8+ T cells between TRAMP and WT mice wasstatistically significant (P < 0.001) and reflected the relatively highernumber of CD8+ T cells present in tumor-bearing mice (130.6 F 92and 7.9 F 2.8 � 103 cells, respectively; P < 0.022). More than 95% ofthe prostate-infiltrating CD8+ T cells were antigen-experiencedCD44+ (not shown). Also CD4+ T cells accumulated in TRAMPprostates (53 F 18 and 9.9 F 6.2 � 103 cells, respectively; P < 0.001).However, the CD4/CD8 ratio was inverted in TRAMP mice whencompared with WT littermates (0.5 F 0.3 and 1.2 F 0.5,respectively; P < 0.017), therefore suggesting a preferentialrecruitment of antigen-experienced CD8+ T cells to the tumor site.
Gating on CD4+ cells and analyzing for the expression of CD25and Foxp3, we found that CD4+CD25+Foxp3+ cells were enriched inthe prostates of tumor-bearing TRAMP mice (Fig. 2C and D). Morestrikingly, the absolute number of triple positive cells was 6-foldhigher in TRAMP than in WT prostates (Fig. 2E) and correlatedwell with the increase in prostate cell number (16 F 9.2 and 2.5 F0.6 � 106, respectively; P < 0.015). Activated effector CD4+ T cells(i.e., CD25+Foxp3�) accumulated as well in TRAMP prostates(7.5 F 3.7% and 3.3 F 2%, respectively; P < 0.04). Hence, prostatecancer development associated with in situ accumulation of antigen-experienced CD8+ but also CD4+ T cells and CD4+C25+Foxp3+ cells.
To verify whether accumulation of CD4+CD25+Foxp3+ cellscorrelated with age and disease progression, a similar analysiswas conducted on prostate tissue from 6-week-old to 8-week-oldmice, age at which TRAMP mice develop scattered foci of mouseprostate intraepithelial neoplasia but are not yet tolerant againstTag (24). Indeed, the macroscopic aspect of TRAMP and WTprostates (24) and their total cell number did not differ (2.4 F 0.3and 2.1 F 0.4 � 106, respectively). In young TRAMP animals, thenumber of CD4+CD25+Foxp3+ cells was similar to the one found inprostates of WT mice (Fig. 2E).
CD4+CD25+Foxp3+ cells were enumerated also in para-aorticTDLN. Whereas TDLN of 6-week-old TRAMP and WT mice weremacroscopically indistinguishable and contained a similar numberof cells (0.4 F 0.1 and 0.5 F 0.2 � 106, respectively), TDLN ofTRAMP mice ages 25 to 27 weeks were enlarged and contained an
Figure 2. CD4+CD25+Foxp3+ cells accumulate in prostate and TDLN of agedTRAMP mice. Prostates and TDLN from naive male TRAMP mice and WTage-matched and sex-matched littermates were processed to single cells. A andB, prostate cells were stained with mAb against CD4 and CD8 and analyzed byFACScalibur (FL1-H, fluorescence channel 1; FL3-H, fluorescence channel 3).The panels depict results from one of six animals per experimental group. Thepercentage of positive cells within the regions is indicated. More in details, inTRAMP and WT prostates, we found 0.76 F 0.2% and 0.33 F 0.1% CD8+ and0.38 F 0.1% and 0.44 F 0.3% CD4+ T cells, respectively. C and D, the samecells were also costained with anti-CD25 and anti-Foxp3 mAb. Cells weregated on CD4+ cells. Percentage of CD4+CD25+Foxp3+ cells in TRAMP and WTmice: 7.1 F 3.4% and 4.5 F 1.0%, respectively. The absolute number ofCD4+CD25+Foxp3+ cells was quantified in the prostate (E) and TDLN fromTRAMP (gray columns ) and WT (white columns ) mice at the indicated age(6–8 wk, n = 5; 25–27 wk, n = 7); bars, SD. Statistical analyses were performedusing the Student’s t test.
Cancer Research
Cancer Res 2008; 68: (1). January 1, 2008 294 www.aacrjournals.org
Research. on February 10, 2016. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from
increased number of cells when compared with age-matched WTcontrols (1.5 F 0.7 and 0.6 F 0.4 � 106, respectively; P < 0.036).Moreover, CD4+CD25+Foxp3+ cells were significantly increased onlyin TDLN of TRAMP mice ages 25 to 27 weeks (Fig. 2F).
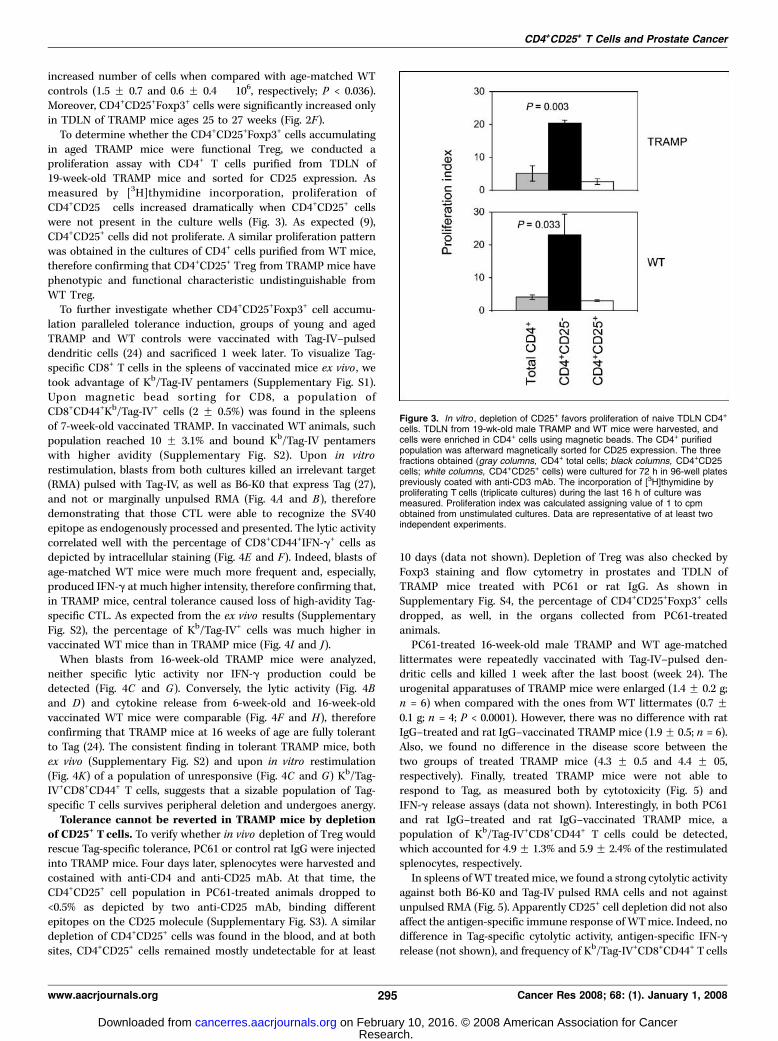
To determine whether the CD4+CD25+Foxp3+ cells accumulatingin aged TRAMP mice were functional Treg, we conducted aproliferation assay with CD4+ T cells purified from TDLN of19-week-old TRAMP mice and sorted for CD25 expression. Asmeasured by [3H]thymidine incorporation, proliferation ofCD4+CD25� cells increased dramatically when CD4+CD25+ cellswere not present in the culture wells (Fig. 3). As expected (9),CD4+CD25+ cells did not proliferate. A similar proliferation patternwas obtained in the cultures of CD4+ cells purified from WT mice,therefore confirming that CD4+CD25+ Treg from TRAMP mice havephenotypic and functional characteristic undistinguishable fromWT Treg.
To further investigate whether CD4+CD25+Foxp3+ cell accumu-lation paralleled tolerance induction, groups of young and agedTRAMP and WT controls were vaccinated with Tag-IV–pulseddendritic cells (24) and sacrificed 1 week later. To visualize Tag-specific CD8+ T cells in the spleens of vaccinated mice ex vivo , wetook advantage of Kb/Tag-IV pentamers (Supplementary Fig. S1).Upon magnetic bead sorting for CD8, a population ofCD8+CD44+Kb/Tag-IV+ cells (2 F 0.5%) was found in the spleensof 7-week-old vaccinated TRAMP. In vaccinated WT animals, suchpopulation reached 10 F 3.1% and bound Kb/Tag-IV pentamerswith higher avidity (Supplementary Fig. S2). Upon in vitrorestimulation, blasts from both cultures killed an irrelevant target(RMA) pulsed with Tag-IV, as well as B6-K0 that express Tag (27),and not or marginally unpulsed RMA (Fig. 4A and B), thereforedemonstrating that those CTL were able to recognize the SV40epitope as endogenously processed and presented. The lytic activitycorrelated well with the percentage of CD8+CD44+IFN-g+ cells asdepicted by intracellular staining (Fig. 4E and F). Indeed, blasts ofage-matched WT mice were much more frequent and, especially,produced IFN-g at much higher intensity, therefore confirming that,in TRAMP mice, central tolerance caused loss of high-avidity Tag-specific CTL. As expected from the ex vivo results (SupplementaryFig. S2), the percentage of Kb/Tag-IV+ cells was much higher invaccinated WT mice than in TRAMP mice (Fig. 4I and J).
When blasts from 16-week-old TRAMP mice were analyzed,neither specific lytic activity nor IFN-g production could bedetected (Fig. 4C and G). Conversely, the lytic activity (Fig. 4Band D) and cytokine release from 6-week-old and 16-week-oldvaccinated WT mice were comparable (Fig. 4F and H), thereforeconfirming that TRAMP mice at 16 weeks of age are fully tolerantto Tag (24). The consistent finding in tolerant TRAMP mice, bothex vivo (Supplementary Fig. S2) and upon in vitro restimulation(Fig. 4K) of a population of unresponsive (Fig. 4C and G) Kb/Tag-IV+CD8+CD44+ T cells, suggests that a sizable population of Tag-specific T cells survives peripheral deletion and undergoes anergy.Tolerance cannot be reverted in TRAMP mice by depletion
of CD25+ T cells. To verify whether in vivo depletion of Treg wouldrescue Tag-specific tolerance, PC61 or control rat IgG were injectedinto TRAMP mice. Four days later, splenocytes were harvested andcostained with anti-CD4 and anti-CD25 mAb. At that time, theCD4+CD25+ cell population in PC61-treated animals dropped to<0.5% as depicted by two anti-CD25 mAb, binding differentepitopes on the CD25 molecule (Supplementary Fig. S3). A similardepletion of CD4+CD25+ cells was found in the blood, and at bothsites, CD4+CD25+ cells remained mostly undetectable for at least
10 days (data not shown). Depletion of Treg was also checked byFoxp3 staining and flow cytometry in prostates and TDLN ofTRAMP mice treated with PC61 or rat IgG. As shown inSupplementary Fig. S4, the percentage of CD4+CD25+Foxp3+ cellsdropped, as well, in the organs collected from PC61-treatedanimals.
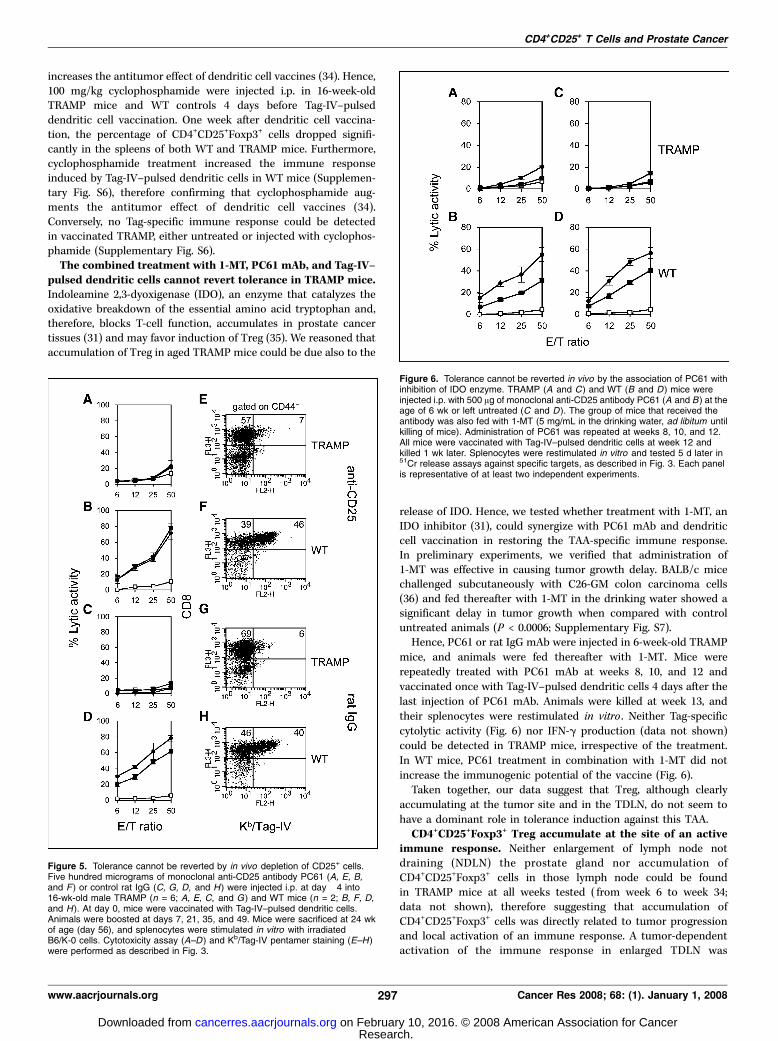
PC61-treated 16-week-old male TRAMP and WT age-matchedlittermates were repeatedly vaccinated with Tag-IV–pulsed den-dritic cells and killed 1 week after the last boost (week 24). Theurogenital apparatuses of TRAMP mice were enlarged (1.4 F 0.2 g;n = 6) when compared with the ones from WT littermates (0.7 F0.1 g; n = 4; P < 0.0001). However, there was no difference with ratIgG–treated and rat IgG–vaccinated TRAMP mice (1.9 F 0.5; n = 6).Also, we found no difference in the disease score between thetwo groups of treated TRAMP mice (4.3 F 0.5 and 4.4 F 05,respectively). Finally, treated TRAMP mice were not able torespond to Tag, as measured both by cytotoxicity (Fig. 5) andIFN-g release assays (data not shown). Interestingly, in both PC61and rat IgG–treated and rat IgG–vaccinated TRAMP mice, apopulation of Kb/Tag-IV+CD8+CD44+ T cells could be detected,which accounted for 4.9 F 1.3% and 5.9 F 2.4% of the restimulatedsplenocytes, respectively.
In spleens of WT treated mice, we found a strong cytolytic activityagainst both B6-K0 and Tag-IV pulsed RMA cells and not againstunpulsed RMA (Fig. 5). Apparently CD25+ cell depletion did not alsoaffect the antigen-specific immune response of WT mice. Indeed, nodifference in Tag-specific cytolytic activity, antigen-specific IFN-grelease (not shown), and frequency of Kb/Tag-IV+CD8+CD44+ T cells
Figure 3. In vitro , depletion of CD25+ favors proliferation of naive TDLN CD4+
cells. TDLN from 19-wk-old male TRAMP and WT mice were harvested, andcells were enriched in CD4+ cells using magnetic beads. The CD4+ purifiedpopulation was afterward magnetically sorted for CD25 expression. The threefractions obtained (gray columns, CD4+ total cells; black columns, CD4+CD25�
cells; white columns, CD4+CD25+ cells) were cultured for 72 h in 96-well platespreviously coated with anti-CD3 mAb. The incorporation of [3H]thymidine byproliferating T cells (triplicate cultures) during the last 16 h of culture wasmeasured. Proliferation index was calculated assigning value of 1 to cpmobtained from unstimulated cultures. Data are representative of at least twoindependent experiments.
CD4+CD25+ T Cells and Prostate Cancer
www.aacrjournals.org 295 Cancer Res 2008; 68: (1). January 1, 2008
Research. on February 10, 2016. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from

could be detected in PC61-vaccinated mice when compared withrat IgG–vaccinated ones (Fig. 5).
We previously reported that full tolerance against Tag could befound in TRAMP mice as early as by weeks 10 to 11 (24). Although wefound evidence of Treg accumulation only at later time points, wehypothesized that failure of PC61 treatment in reverting toleranceagainst Tag could be due to a deferred schedule. Hence, PC61antibodies were injected in TRAMP mice starting at week 6, time atwhich TRAMP mice are not tolerant (Fig. 2). To prolong CD25+ T-celldepletion, PC61 was injected every 2 weeks until week 12, when allmice were vaccinated with Tag-IV–pulsed dendritic cells and killed1 week later. Ex vivo analysis of lymphoid organs confirmed a10-fold reduction of CD4+CD25+Foxp3+ cells in PC61-injectedanimals when compared with their controls (e.g., TDLN, 9.8 F0.8% and 0.8 F 0.8%, respectively). However, such treatment did notbreak Tag-specific tolerance in TRAMP mice (data not shown).
Those unexpected findings prompted us to verify the antitumoreffects of PC61 mAb in a subcutaneous model of cancer. Hence,
PC61 or control rat IgG were injected i.p. at days �1 and +3 in WTC57BL/6 mice, and at day 0, animals were challenged subcutane-ously with TRAMP-C1 cells, a prostate adenocarcinoma cell line(28). As shown in Supplementary Fig. S5, PC61-treated miceshowed a significant delay (P < 0.026) in tumor growth, thereforeconfirming that PC61 treatment is effective against subcutaneousprostate cancer (32).
An alternative explanation to our findings is that antibodytreatment also depleted activated/effector CD25+ T cells. Using asimilar depletion protocol, concomitant depletion of effectors andconsequent reduction of the antigen-specific immune response hasnot been reported thus far (e.g., refs. 17–19). Also, in WT animals,we found that depletion of CD25+ cells by PC61 treatment did notimpair the immune response induced by Tag-IV–pulsed dendriticcells (Fig. 5), therefore suggesting that CD25+ effectors were notaltered by PC61 pretreatment. To further address this issue, weattempted to impair Treg function by cyclophosphamide, which,at low doses, decreases Treg number and functionality (33) and
Figure 4. Young TRAMP mice are able tomount a Tag-specific CTL response, whereasadult tumor-bearing mice are tolerant to Tag.Tag-IV–pulsed dendritic cells were injected oncei.d. into 6-wk-old (A, E, and I ) and 16-wk-old(C, G, and K ) male TRAMP mice and their WTage-matched and sex-matched littermates (B, F, Jand D, H, L, respectively). After 7 d, animals werekilled, and their splenocytes were stimulatedin vitro with irradiated B6/K-0 cells and tested5 d later for cytotoxic activity (measured as51Cr release); unpulsed (white squares ) orTag-IV–pulsed (black squares ) RMA and B6/K-0(black circles ) cells were used as targets (A–D ).E–H, blasts were tested for IFN-g production (uponstimulation with B6/K-0 or an irrelevant target)and analyzed by FACScalibur after costainingwith mAb against CD8, CD44, and IFN-g.Percentage of CD8+IFN-g+ T cells in TRAMP andWT mice: 6 wk, 9 F 2.5% and 55 F 11%; 16 wk,0.5 F 0.6% and 42 F 18%, respectively (n = 4).IFN-g–producing cells in the presence of theirrelevant target (RMA) were <2% (not shown) andwere subtracted. I–L, cells were also costainedwith mAb against CD8, CD44, and Kb/Tag-IVpentamer to identify a Tag-specific population(percentage of positive cells is indicated in eachquadrant). Percentage of CD8+Kb/Tag-IV+ T cells inTRAMP and WT mice: 6 wk, 14 F 2.5% and59 F 1.6%; 16 wk, 6 F 2.8% and 58 F 9.1%,respectively (n = 4). Staining with Kb/OVApentamers gave background values of positivecells <1% (Supplementary Fig. S1). The differentintensity of pentamer staining is due to differentbatches of custom grade pentamers used in thetwo sets of experiments. Data correspond to one ofat least three independent experiments, whichgave similar results.
Cancer Research
Cancer Res 2008; 68: (1). January 1, 2008 296 www.aacrjournals.org
Research. on February 10, 2016. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from
increases the antitumor effect of dendritic cell vaccines (34). Hence,100 mg/kg cyclophosphamide were injected i.p. in 16-week-oldTRAMP mice and WT controls 4 days before Tag-IV–pulseddendritic cell vaccination. One week after dendritic cell vaccina-tion, the percentage of CD4+CD25+Foxp3+ cells dropped signifi-cantly in the spleens of both WT and TRAMP mice. Furthermore,cyclophosphamide treatment increased the immune responseinduced by Tag-IV–pulsed dendritic cells in WT mice (Supplemen-tary Fig. S6), therefore confirming that cyclophosphamide aug-ments the antitumor effect of dendritic cell vaccines (34).Conversely, no Tag-specific immune response could be detectedin vaccinated TRAMP, either untreated or injected with cyclophos-phamide (Supplementary Fig. S6).The combined treatment with 1-MT, PC61 mAb, and Tag-IV–
pulsed dendritic cells cannot revert tolerance in TRAMP mice.Indoleamine 2,3-dyoxigenase (IDO), an enzyme that catalyzes theoxidative breakdown of the essential amino acid tryptophan and,therefore, blocks T-cell function, accumulates in prostate cancertissues (31) and may favor induction of Treg (35). We reasoned thataccumulation of Treg in aged TRAMP mice could be due also to the
release of IDO. Hence, we tested whether treatment with 1-MT, anIDO inhibitor (31), could synergize with PC61 mAb and dendriticcell vaccination in restoring the TAA-specific immune response.In preliminary experiments, we verified that administration of1-MT was effective in causing tumor growth delay. BALB/c micechallenged subcutaneously with C26-GM colon carcinoma cells(36) and fed thereafter with 1-MT in the drinking water showed asignificant delay in tumor growth when compared with controluntreated animals (P < 0.0006; Supplementary Fig. S7).
Hence, PC61 or rat IgG mAb were injected in 6-week-old TRAMPmice, and animals were fed thereafter with 1-MT. Mice wererepeatedly treated with PC61 mAb at weeks 8, 10, and 12 andvaccinated once with Tag-IV–pulsed dendritic cells 4 days after thelast injection of PC61 mAb. Animals were killed at week 13, andtheir splenocytes were restimulated in vitro . Neither Tag-specificcytolytic activity (Fig. 6) nor IFN-g production (data not shown)could be detected in TRAMP mice, irrespective of the treatment.In WT mice, PC61 treatment in combination with 1-MT did notincrease the immunogenic potential of the vaccine (Fig. 6).
Taken together, our data suggest that Treg, although clearlyaccumulating at the tumor site and in the TDLN, do not seem tohave a dominant role in tolerance induction against this TAA.CD4+CD25+Foxp3+ Treg accumulate at the site of an active
immune response. Neither enlargement of lymph node notdraining (NDLN) the prostate gland nor accumulation ofCD4+CD25+Foxp3+ cells in those lymph node could be foundin TRAMP mice at all weeks tested ( from week 6 to week 34;data not shown), therefore suggesting that accumulation ofCD4+CD25+Foxp3+ cells was directly related to tumor progressionand local activation of an immune response. A tumor-dependentactivation of the immune response in enlarged TDLN was
Figure 5. Tolerance cannot be reverted by in vivo depletion of CD25+ cells.Five hundred micrograms of monoclonal anti-CD25 antibody PC61 (A, E, B,and F ) or control rat IgG (C, G, D, and H ) were injected i.p. at day �4 into16-wk-old male TRAMP (n = 6; A, E, C, and G) and WT mice (n = 2; B, F, D,and H ). At day 0, mice were vaccinated with Tag-IV–pulsed dendritic cells.Animals were boosted at days 7, 21, 35, and 49. Mice were sacrificed at 24 wkof age (day 56), and splenocytes were stimulated in vitro with irradiatedB6/K-0 cells. Cytotoxicity assay (A–D ) and Kb/Tag-IV pentamer staining (E–H)were performed as described in Fig. 3.
Figure 6. Tolerance cannot be reverted in vivo by the association of PC61 withinhibition of IDO enzyme. TRAMP (A and C ) and WT (B and D ) mice wereinjected i.p. with 500 Ag of monoclonal anti-CD25 antibody PC61 (A and B ) at theage of 6 wk or left untreated (C and D ). The group of mice that received theantibody was also fed with 1-MT (5 mg/mL in the drinking water, ad libitum untilkilling of mice). Administration of PC61 was repeated at weeks 8, 10, and 12.All mice were vaccinated with Tag-IV–pulsed dendritic cells at week 12 andkilled 1 wk later. Splenocytes were restimulated in vitro and tested 5 d later in51Cr release assays against specific targets, as described in Fig. 3. Each panelis representative of at least two independent experiments.
CD4+CD25+ T Cells and Prostate Cancer
www.aacrjournals.org 297 Cancer Res 2008; 68: (1). January 1, 2008
Research. on February 10, 2016. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from

suggested by the increased population of activated CD4+CD25+
Foxp3� cells only in TDLN from aged and tumor-bearing TRAMPmice (12.3 F 7 � 103; n = 6) and not in age-matched WT animals(3 F 1.3 � 103; n = 6; P < 0.021) or young TRAMP mice (3.9 F3.3 � 103, n = 5).
Hence, we verified whether CD4+CD25+Foxp3+ cells accumulatedalso in lymph node draining the site of vaccination (VDLN). Asfound in TDLN, the absolute number of cells in VDLN, from WTmice vaccinated 1 week before with Tag-IV–pulsed dendritic cells,increased dramatically from an average of 0.95 F 0.4 � 106 for aNDLN to 12.9 F 3.6 � 106 cells in VDLN (n = 10; P < 0.000002). Theabsolute number of CD4+CD25+Foxp3+ cells increased as well,being only 30 F 16 � 103 in NDLN and reaching the 299 F 78 � 103
units in VDLN (P < 0.000006). Curiously enough, also in VDLN, wefound a significant increase in the absolute number and not in thepercentage of Treg, therefore confirming that, during an activeimmune response, increase in Treg number in VDLN parallelsincrease of other lymph node cell populations, such as total CD4(3.4 F 1 and 0.3 F 0.1 � 106 cells in VDLN and NDLN, respectively),CD8 (2.5 F 0.5 and 0.2 F 0.1 � 106), and especially Kb/Tag-IV+ cells(9.5 F 2.5%). Similar results were obtained in vaccinated TRAMPmice (data not shown).
All together, our data suggest that Treg accumulation at the siteof an active immune response is physiologic and, at least in theTRAMP model, is not essential for tolerance induction to a TAA.
Discussion
The mechanism by which peripheral tolerance is induced intumor-bearing subjects is ill defined (4), although findings inanimal models suggest that TAA drained or carried to the lymphnodes and presented by APC in a noninflammatory context driveclonal deletion or anergy (37). Tolerance in 10-week-old to 11-week-old TRAMP mice (24) is likely induced in the absence of localinflammation because the prostate gland at that time lacks aninflammatory infiltrate and TDLN are not enlarged (data notshown). Although several previous findings were consistent withperipheral deletion of TAA-specific T cells in tumor-bearingTRAMP mice (24, 38), in those mice, we consistently found apopulation of Kb/Tag-IV+CD8+CD44+ T cells, which neitherexpanded in culture nor killed or produced IFN-g upon specificstimulation, therefore implying that a sizable population of Tag-specific T cells survive peripheral deletion and undergo anergy intumor-bearing TRAMP mice.
As it occurs in human prostate cancer patients (15), we foundevidence of a progressive accumulation of functional Treg both atthe tumor site and in TDLN of TRAMP mice developingspontaneous prostate cancer. However, Treg depletion by PC61mAb followed by repeated vaccinations did not rescue T cells fromanergy. Also, repetitive PC61 injections followed by dendritic cellvaccination did not break tolerance. Most importantly, thecombined treatments did not modify both the early and advancedphases of disease progression. We excluded malfunction of themAb, because mice that received a single dose of PC61 antibodyshowed approximately a 10-fold reduction in the CD4+CD25+ T-cellpopulation in lymphoid organs. Furthermore, PC61 treatment waseffective against TRAMP-C1 tumors.
As for our TRAMP-C1 model, most of the reports on successfulPC61-mediated depletion of Treg were conducted in mice in whichtumor cells from in vitro established tumor cell lines were injecteds.c. The engraftment of even a small number of tumor cells does
not mimic the natural development of a tumor mass (37) and likelycauses inflammation at the site of injection (39). This maydramatically alter the dynamic interactions between tumor andimmune cells (5). Hiura et al. recently reported that both Treg andantitumor effector T cells were primed in the same TDLN of atumor growing subcutaneously. (12). PC61 mAb are usually injecteda few days before or at the time of tumor cell implantation. Suchtreatment schedule may give an unrealistic advantage to effectorT cells, likely depleting thymic-derived Treg and preventing theinduction of peripherally induced tumor-specific Treg. Thosemodels may more closely mimic the condition of a prophylacticvaccination, wherein PC61 pretreatment increases the immuno-genic potential of the vaccine (18), likely recruiting high-avidityT cells (40) and natural killer cells (17).
However, those tumor transplantation models did not addressthe peculiar condition of tumor-bearing subjects, wherein effectorT cells, negatively selected for avidity or exhausted by prolongedTAA presentation in the draining lymph nodes (41), are atcompetition with both thymus-derived and peripherally inducedTreg expanded by conversion of CD4+CD25� lymphocytes (13).Indeed, when high-avidity tumor-specific T cells were lackingbecause of deletion (42), as it occurs in TRAMP mice and mostlikely in humans, or when PC61 treatment was began at later timepoints since subcutaneous tumor challenge (43), depletion of Tregby PC61 treatment did not increase vaccine efficacy even against asubcutaneous tumor.
Tien et al. (32) found increased CD4+CD25+ T cells in thetransgenic model of spontaneous prostate dysplasia 12T-7s LPB-Tag. PC61 treatment in those mice reduced, but did not prevent,tumor growth. The different findings obtained in TRAMP and12T-7s LPB-Tag models may depend on 12T-7s LPB-Tag micedeveloping a far less aggressive prostate disease. Indeed, the lowamount of antigen released from prostate cells in 12T-7s LPB-Tagmice may likely spare prostate-specific T-cell clones from toleranceinduction and favor their activation in conditions of reducedcompetition (i.e., depletion of CD25+ cells).
Our experimental settings addressed the role of Treg in a modelof spontaneous prostate cancer development, wherein endogenouslow avidity CD8+ T cells are tolerized during cancer developmentand progression. Hence, our data substantially extend, in a modelthat more closely resemble the human disease, the observationmade in a more artificial context (44) by Mihalyo et al. (45). Indeed,PC61 pretreatment did not avoid tolerization also of high-avidityHA-specific transgenic CD4+ T cells upon adoptive transfer intoTRAMP mice expressing HA in the prostate.
The process of Treg generation and accrual in tumor-bearingsubjects may follow a kinetic common to autoantigens: thepersistence of the Ag; in this case, the TAA would lead tocontinuous generation of Treg in the periphery. As suggested (46),removal of Treg might have only limited or no effect in thatcondition, as Treg may be continuously replenished from thememory pool. In the TRAMP model in particular, tolerance is soprofound that even transient depletion of Treg is not sufficient torestore TAA-specific immune response. This might occur also inprostate cancer patients.
Depletion of Treg in vivo has been already attempted in humanswith variable results. Dannull et al. (47) reported that in renalcancer patients pretreated with the recombinant IL-2 diphteriatoxin conjugate denileukin, Treg were selectively eliminated and avaccine-mediated antitumor immunity was enhanced. However,another group reported no reduction in the number of Treg or their
Cancer Research
Cancer Res 2008; 68: (1). January 1, 2008 298 www.aacrjournals.org
Research. on February 10, 2016. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from

function in melanoma patients receiving denileukin (48). Further-more, depletion is only transient, and Treg rapidly repopulate thehuman body (49).
Other strategies may be implemented to break tolerance inprostate cancer patients and in TRAMP mice. Also, inhibition ofIDO by 1-MT did not revert tolerance in our transgenic model. Onthe other hand, high levels of nitrotyrosines in T cells infiltratingprostate cancer from humans and TRAMP mice suggested a localproduction of peroxynitrites (50). Indeed, in vitro inhibition ofarginase and nitric oxide synthase activity restored T-cellresponsiveness to tumor. It would be interesting to investigatewhether in vivo inhibition of those enzymes may breakTag-tolerance in TRAMP mice.
In addition to the mechanisms cited above, cytokines, such asIL-6, IL-10, and transforming growth factor-h, prostaglandins, andeven prostate-specific antigen are all factors that may directly and/or indirectly impair T-cell function while favoring tumor cell growth(7). Impairment in tumor antigen expression or its processing and
presentation by both tumor cells and APC, as well as alteredexpression of B7 family molecules by APC, may likely reduce effectiveimmunosurveillance of prostate cancer (7). Finally, FasL expressedon tumor cells, together with other proapoptotic mechanisms, mayfavor programmed T-cell death at the tumor site (7). Hence, furtherstudies are warranted to better define tumor immunosuppressivemechanisms and design more effective and vigorous combinatorialstrategies for generating productive immune responses.
Acknowledgments
Received 7/9/2007; revised 9/17/2007; accepted 10/22/2007.Grant support: Associazione Italiana per la Ricerca sul Cancro, Ministero della
Salute, CARIPLO, and Ministero dell’Istruzione dell’Universita e della Ricerca.The costs of publication of this article were defrayed in part by the payment of page
charges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.
We thank Dr. Tevethia (Pennsylvania State University College of Medicine) forproviding us B6/K-0 and several colleagues at Cancer Immunotherapy and GeneTherapy program for helpful discussions and critical comments on the manuscript.
CD4+CD25+ T Cells and Prostate Cancer
www.aacrjournals.org 299 Cancer Res 2008; 68: (1). January 1, 2008
References1. Jemal A, Siegel R, Ward E, et al. Cancer statistics, 2006.CA Cancer J Clin 2006;56:106–30.2. Marrari A, Iero M, Pilla L, et al. Vaccination therapy inprostate cancer. Cancer Immunol Immunother 2007;56:429–45.3. Small EJ, Schellhammer PF, Higano CS, et al. Placebo-controlled phase III trial of immunologic therapy withsipuleucel-T (APC8015) in patients with metastatic,asymptomatic hormone refractory prostate cancer.J Clin Oncol 2006;24:3089–94.4. Pardoll D. Does the immune system see tumors asforeign or self? Annu Rev Immunol 2003;21:807–39.5. Ochsenbein AF, Klenerman P, Karrer U, et al. Immunesurveillance against a solid tumor fails because ofimmunological ignorance. Proc Natl Acad Sci U S A1999;96:2233–8.6. Rabinovich GA, Gabrilovich D, Sotomayor EM. Immu-nosuppressive strategies that are mediated by tumorcells. Annu Rev Immunol 2006;25:267–96.7. Miller AM, Pisa P. Tumor escape mechanisms inprostate cancer. Cancer Immunol Immunother 2007;56:81–7.8. Rudensky AY, Gavin M, Zheng Y. FOXP3 and NFAT:partners in tolerance. Cell 2006;126:253–6.9. Sakaguchi S. Naturally arising Foxp3-expressingCD25+CD4+ regulatory T cells in immunological toler-ance to self and non-self. Nat Immunol 2005;6:345–52.10. Murakami M, Sakamoto A, Bender J, Kappler J,Marrack P. CD25+CD4+ T cells contribute to the controlof memory CD8+ T cells. Proc Natl Acad Sci U S A 2002;99:8832–7.11. Kretschmer K, Apostolou I, Jaeckel E, Khazaie K, vonBoehmer H. Making regulatory T cells with definedantigen specificity: role in autoimmunity and cancer.Immunol Rev 2006;212:163–9.12. Hiura T, Kagamu H, Miura S, et al. Both regulatoryT cells and antitumor effector T cells are primed in thesame draining lymph nodes during tumor progression.J Immunol 2005;175:5058–66.13. Valzasina B, Piconese S, Guiducci C, Colombo MP.Tumor-induced expansion of regulatory T cells byconversion of CD4+CD25- lymphocytes is thymus andproliferation independent. Cancer Res 2006;66:4488–95.14. Liu VC, Wong LY, Jang T, et al. Tumor evasion of theimmune system by converting CD4+CD25- T cells intoCD4+CD25+ T regulatory cells: role of tumor-derivedTGF-h. J Immunol 2007;178:2883–92.15. Miller AM, Lundberg K, Ozenci V, et al.CD4+CD25high T cells are enriched in the tumor andperipheral blood of prostate cancer patients. J Immunol2006;177:7398–405.16. Curiel TJ, Coukos G, Zou L, et al. Specific recruitment
of regulatory T cells in ovarian carcinoma fostersimmune privilege and predicts reduced survival. NatMed 2004;10:942–9.17. Shimizu J, Yamazaki S, Sakaguchi S. Induction oftumor immunity by removing CD25+CD4+ T cells: acommon basis between tumor immunity and autoim-munity. J Immunol 1999;163:5211–8.18. Sutmuller RP, van Duivenvoorde LM, van Elsas A,et al. Synergism of cytotoxic T lymphocyte-associatedantigen 4 blockade and depletion of CD25(+) regulatoryT cells in antitumor therapy reveals alternative path-ways for suppression of autoreactive cytotoxic Tlymphocyte responses. J Exp Med 2001;194:823–32.19. Turk MJ, Guevara-Patino JA, Rizzuto GA, EngelhornME, Sakaguchi S, Houghton AN. Concomitant tumorimmunity to a poorly immunogenic melanoma isprevented by regulatory T cells. J Exp Med 2004;200:771–82.20. Greenberg NM, DeMayo F, Finegold MJ, et al.Prostate cancer in a transgenic mouse. Proc Natl AcadSci U S A 1995;92:3439–43.21. Shappel SB, Thomas GV, Roberts RL, et al. Prostatepathology of genetically engineered mice: definitionsand classification. The consensus report from the BarHarbor meeting of the mouse models of Human CancerConsortium Prostate Pathology Commitee. Cancer Res2004;64:2270–305.22. Mylin LM, Schell TD, Roberts D, et al. Quantitation ofCD8(+) T-lymphocyte responses to multiple epitopesfrom simian virus 40 (SV40) large T antigen in C57BL/6mice immunized with SV40, SV40 T-antigen-trans-formed cells, or vaccinia virus recombinants expressingfull-length T antigen or epitope minigenes. J Virol 2000;74:6922–34.23. Zheng X, Gao JX, Zhang H, Geiger TL, Liu Y, Zheng P.Clonal deletion of simian virus 40 large T antigen-specific T cells in the transgenic adenocarcinoma ofmouse prostate mice: an important role for clonaldeletion in shaping the repertoire of T cells specific forantigens overexpressed in solid tumors. J Immunol 2002;169:4761–9.24. Degl’Innocenti E, Grioni M, Boni A, et al. PeripheralTcell tolerance occurs early during spontaneous prostatecancer development and can be rescued by dendriticcell immunization. Eur J Immunol 2005;35:66–75.25. Sambrook J, Russel DW. Preparation and analysis ofeukariotic genomic DNA. In: Sambrook J, Russel DW,editors. Molecular cloning. A laboratory manual Thirdedition. Cold Spring Harbor (NY): Cold Spring Labora-tory Press; 2001. p. 6.26–6.7.26. Ljunggren HG, Karre K. Host resistance directedselectively against H-2-deficient lymphoma variants.Analysis of the mechanism. J Exp Med 1985;162:1745–59.
27. Tanaka Y, Tevethia SS. In vitro selection of SV40 Tantigen epitope loss variants by site-specific cytotoxicT lymphocyte clones. J Immunol 1988;140:4348–54.28. Foster BA, Gingrich JR, Kwon ED, Madias C, Green-berg NM. Characterization of prostatic epithelial celllines derived from transgenic adenocarcinoma of themouse prostate (TRAMP) model. Cancer Res 1997;57:3325–30.29. Camporeale A, Boni A, Iezzi G, et al. Critical impactof the kinetics of dendritic cells activation on the in vivoinduction of tumor-specific T lymphocytes. Cancer Res2003;63:3688–94.30. Schell TD, Knowles BB, Tevethia SS. Sequential lossof cytotoxic T lymphocyte responses to simian virus40 large T antigen epitopes in T antigen transgenicmice developing osteosarcomas. Cancer Res 2000;60:3002–12.31. Uyttenhove C, Pilotte L, Theate I, et al. Evidence for atumoral immune resistance mechanism based ontryptophan degradation by indoleamine 2,3-dioxyge-nase. Nat Med 2003;9:1269–74.32. Tien AH, Xu L, Helgason CD. Altered immunityaccompanies disease progression in a mouse model ofprostate dysplasia. Cancer Res 2005;65:2947–55.33. Lutsiak ME, Semnani RT, De Pascalis R, Kashmiri SV,Schlom J, Sabzevari H. Inhibition of CD4(+)25+T regulatory cell function implicated in enhancedimmune response by low-dose cyclophosphamide.Blood 2005;105:2862–8.34. Liu JY, Wu Y, Zhang XS, et al. Single administration oflow dose cyclophosphamide augments the antitumoreffect of dendritic cell vaccine. Cancer ImmunolImmunother 2007;56:1597–604.35. Mellor AL, Munn DH. IDO expression by dendriticcells: tolerance and tryptophan catabolism. Nat RevImmunol 2004;4:762–74.36. Chiodoni C, Paglia P, Stoppacciaro A, Rodolfo M,Parenza M, Colombo MP. Dendritic cells infiltratingtumors cotransduced with granulocyte/macrophagecolony-stimulating factor (GM-CSF) and CD40 ligandgenes take up and present endogenous tumor-associat-ed antigens, and prime naive mice for a cytotoxicT lymphocyte response. J Exp Med 1999;190:125–33.37. Spiotto MT, Fu YX, Schreiber H. Tumor immunitymeets autoimmunity: antigen levels and dendritic cellmaturation. Curr Opin Immunol 2003;15:725–30.38. Drake CG, Doody AD, Mihalyo MA, et al. Androgenablation mitigates tolerance to a prostate/prostatecancer-restricted antigen. Cancer Cell 2005;7:239–49.39. Bellone M, Iezzi G, Martin-Fontecha A, et al.Rejection of a nonimmunogenic melanoma by vaccina-tion with natural melanoma peptides on engineeredantigen-presenting cells. J Immunol 1997;158:783–9.40. Ercolini AM, Ladle BH, Manning EA, et al.
Research. on February 10, 2016. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from

Cancer Research
Cancer Res 2008; 68: (1). January 1, 2008 300 www.aacrjournals.org
Recruitment of latent pools of high-avidity CD8(+)T cells to the antitumor immune response. J ExpMed 2005;201:1591–602.41. Dong H, Strome SE, Salomao DR, et al. Tumor-associated B7–1 promotes T-cell apoptosis: a potentialmechanism of immune evasion. Nat Med 2002;8:793–800.42. Souders NC, Sewell DA, Pan ZK, et al. Listeria-basedvaccines can overcome tolerance by expanding lowavidity CD8+ T cells capable of eradicating a solid tumorin a transgenic mouse model of cancer. Cancer Immun2007;7:1–12.43. Ko K, Yamazaki S, Nakamura K, et al. Treatment ofadvanced tumors with agonistic anti-GITR mAb and itseffects on tumor-infiltrating Foxp3+CD25+CD4+ regula-tory T cells. J Exp Med 2005;202:885–91.
44. Badovinac VP, Haring JS, Harty JT. Initial T cellreceptor transgenic cell precursor frequency dictatescritical aspects of the CD8(+) T cell response toinfection. Immunity 2007;26:827–41.45. Mihalyo MA, Hagymasi AT, Slaiby AM, Nevius EE,Adler AJ. Dendritic cells program non-immunogenicprostate-specific T cell responses beginning at earlystages of prostate tumorigenesis. Prostate 2007;67:536–46.46. Akbar AN, Vukmanovic-Stejic M, Taams LS, MacallanDC. The dynamic co-evolution of memory and regula-tory CD4+ T cells in the periphery. Nat Rev Immunol2007;7:231–7.47. Dannull J, Su Z, Rizzieri D, et al. Enhancement ofvaccine-mediated antitumor immunity in cancer
patients after depletion of regulatory T cells. J ClinInvest 2005;115:3623–33.48. Attia P, Maker AV, Haworth L, Rogers-Freezer L,Rosenberg SA. Inability of a fusion protein of IL-2and diphtheria toxin (Denileukin Diftitox, DAB389IL-2, ONTAK) to eliminate regulatory T lymphocytesin patients with melanoma. J Immunother 2005;28:582–92.49. O’Mahony D, Morris JC, Quinn C, et al. A pilot studyof CTLA-4 blockade after cancer vaccine failure inpatients with advanced malignancy. Clin Cancer Res2007;13:958–64.50. Bronte V, Kasic T, Gri G, et al. Boosting antitumorresponses of T lymphocytes infiltrating human prostatecancers. J Exp Med 2005;201:1257–68.
Research. on February 10, 2016. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from

2008;68:292-300. Cancer Res Elena Degl'Innocenti, Matteo Grioni, Giusy Capuano, et al.
Regulatory T Cells+CD25+Is Independent from CD4Peripheral T-Cell Tolerance Associated with Prostate Cancer
Updated version
http://cancerres.aacrjournals.org/content/68/1/292
Access the most recent version of this article at:
Material
Supplementary
http://cancerres.aacrjournals.org/content/suppl/2008/01/23/68.1.292.DC1.html
Access the most recent supplemental material at:
Cited articles
http://cancerres.aacrjournals.org/content/68/1/292.full.html#ref-list-1
This article cites 49 articles, 27 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/68/1/292.full.html#related-urls
This article has been cited by 15 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
Research. on February 10, 2016. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from







