
p(1a((1efic
d
Molecular and Cellular Neuroscience 15, 343–354 (2000)
doi:10.1006/mcne.1999.0829, available online at http://www.idealibrary.com on MCN
A
Myelin Basic Protein Gene DosageEffects in the PNS
Candra Smith-Slatas* and Elisa Barbarese* ,1
*Combined Degree Program and Neuroscience Graduate Program and Department ofNeurology, University of Connecticut Health Center, Farmington, Connecticut 06030
in(otwwcgtp
itstnImodCbetfitb(vmmeta
Myelin basic protein (MBP) plays an essential adhesiverole in the formation of compact myelin in the centralnervous system (CNS), but not in the peripheral nervoussystem (PNS). Morphologic data suggest that MBP con-trols the number of cytoplasmic channels or Schmidt–Lanterman incisures (SLI) present in PNS myelin. The lev-els of connexin-32 (Cx32) and myelin-associatedglycoprotein (MAG), two components of the incisures, areinversely proportional to the levels of MBP in sciaticnerves of mice affected by the shiverer (shi) mutation,while protein zero (P0) and peripheral membrane protein22 (PMP22), two structural components of compact mye-lin, remain constant. The levels of P0, PMP22, Cx32, andMAG mRNA do not vary in relationship to the levels ofMBP. This indicates that MBP exerts its effect on Cx32and MAG at a posttranscriptional level and suggests anew function for MBP in regulating gene expression in thePNS.
INTRODUCTION
Myelin from the peripheral nervous system (PNS) ofthe homozygous mutant mouse shiverer (shi/shi) is com-
act despite the total absence of myelin basic proteinMBP) (Kirschner and Ganser, 1980; Mikoshiba et al.,980; Rosenbluth, 1980). It has been postulated that thedhesive role of MBP in central nervous system myelinCNS) is performed by protein zero (P0) in PNS myelinD’Urso et al., 1990; Filbin et al., 1990; Martini et al.,995). If this were the case, the level of P0 may belevated in PNS myelin from shi/shi mice to compensateor the absence of MBP. Further morphologic studiesndicated that the number of cytoplasmic channelsalled Schmidt–Lanterman incisures (SLI) was elevated
nc
1 To whom correspondence and reprint requests should be ad-ressed. Fax: (860) 679-4446. E-mail: [email protected].
1044-7431/00 $35.00Copyright © 2000 by Academic Press
ll rights of reproduction in any form reserved.
n shi/shi PNS myelin, suggesting that several compo-ents may be up-regulated in the absence of MBPGould et al., 1995). We sought to determine if the levelsf connexin 32 (Cx32) and myelin-associated glycopro-ein (MAG), two components of the incisures, and P0
ere increased in shi/shi PNS myelin in comparison toild type (wt) animals, and if the levels of these proteins
orrelated with the levels of MBP. In order to distin-uish whether gene expression was regulated throughranscription or translation, the levels of several myelinroteins cognate mRNA were measured.A comparison of morphologic and biochemical stud-
es suggests that there is an inverse correlation betweenhe levels of MBP and the number of SLI. In the ratciatic nerve, the 18.5-kDa isoform of MBP is 2% of theotal myelin proteins (Greenfield et al., 1973), and theumber of incisures is 25 per internode (Hiscoe, 1947).n the guinea pig sciatic nerve, MBP is 16% of the total
yelin proteins (Greenfield et al., 1973), and the numberf incisures is 12 per internode for a fibre of equivalentiameter and internodal distance (Webster, 1964). In theNS of anyone species, there is a positive correlationetween the levels of MBP and myelin thickness (Shinet al., 1992), which may not be significant in the PNS; inhe latter, there is a positive correlation between thebre diameter and the number of SLI (Hiscoe, 1947). In
he absence of MBP, in homozygous shi mice, the num-er of incisures is double that found in wt sciatic nervesGould et al., 1995), while myelin thickness is reportedlyery midly reduced (Peterson and Bray, 1984) or nor-al (Rosenbluth, 1980). The present study uses ho-ozygous (shi/shi) and heterozygous shi (1/shi) mice to
xamine the correlation between the level of MBP andhe number of SLI in the sciatic nerves at a biochemicalnd molecular level because these animals differ ge-
etically only by the number of functional MBP geneopies.343

ttC
aprfibobb1Absbps
Ri
tatslst
t
344 Smith-Slatas and Barbarese
The shi mutation results from a large deletion in theMBP gene with no MBP protein or messenger RNAbeing produced (Roach et al., 1983). It is functionally anull allele for MBP and introduction of the wt MBP genein the genome of homozygous shi animals rescues themutant phenotype (Readhead et al., 1987). MBP is amyelin-specific component localized to the compact re-gions of CNS and PNS myelin (Omlin et al., 1982). Inboth the CNS and the PNS, the amount of MBP isdirectly proportional to the number of gene copiespresent, and 1/shi myelin contains half the amount ofMBP found in wt (Barbarese et al., 1983; Popko et al.,1987).
SLI are cytoplasmic channels within the compact my-elin sheath. They contain Schwann cell cytoplasm, a feworganelles, and gap junctions composed of Cx32 pro-teins that link successive wrappings of myelin (Peters etal., 1991). The following proteins have been assigned toSLI: 29,39-cyclic nucleotide phosphohydrolase, MAG,S100, Cx32, E-cadherin, DM-20, and actin (Puckett et al.,1987; Trapp et al., 1989; Mata et al., 1990; Bergoffen et al.,1993; Fannon et al., 1995; Griffiths et al., 1995; Andersonet al., 1997; Garbern et al., 1997). P0, MBP, P2, andperipheral myelin protein-22 (PMP22) are present in thecompact regions of the myelin sheath (Trapp et al., 1981;Omlin et al., 1982; Welcher et al., 1991; Snipes et al., 1992;Anderson et al., 1997; Garbern et al., 1997).
There is concordance between the levels of myelinprotein mRNAs and their corresponding proteins dur-ing the active period of myelination as indicated by theanalysis of the steady state levels of MAG, MBP, Cx32,PMP22, and P0 mRNA in the rat sciatic nerve (Stahl etal., 1990; Gupta et al., 1990; Scherer et al., 1995; Griffithset al., 1989; Gow et al., 1994; Kuhn et al., 1993; Snipes etal., 1992). In this paper we have used a ribonucleaseprotection assay (RPA) to examine the steady statemRNA levels of PMP22, Cx32, and MAG in 90-day-oldshi/shi and wt animals to determine if gene expressionwas similarly regulated.
RESULTS
Localization of Cx32 and MAG to SLI in shi/shiSciatic Nerves
In a previous study the increased density of SLI inshi/shi sciatic nerves was estimated from morphologicdata (Gould et al., 1995). In order to further characterizehese structures, immunofluorescence was performed
o assess the localization of two of their components,x32 and MAG. Whole sciatic nerves were stained withnti-Cx32, anti-MAG, and with fluorescein-conjugatedhalloidin to delineate SLI and nodes of Ranvier. Theesults showed that Cx32 and MAG were found inlamentous actin-positive incisures (arrowheads) inoth wt and shi/shi sciatic nerves. Other compartmentsf the nerve such as the outer mesaxons, Schwann cellodies (arrows), and the nodes of Ranvier, which haveeen reported to contain MAG and Cx32 (Trapp et al.,989; Scherer et al., 1995), were also positively stained.ll incisures-like structures identified by phalloidin la-eling contained Cx32 and MAG, indicating that theupernumerary incisures in shi/shi sciatic nerves areona fide SLI and suggesting that the levels of the tworoteins should be commensurably increased in theciatic nerves of shi/shi mice.
elationship between MBP Levels and Incisuresn 1/shi Mice Sciatic Nerves
In order to further establish the relationship betweenhe levels of MBP and the density of SLI, the latter wasssessed in 90-day-old 1/shi mice sciatic nerves (n 5 2)hat contain half the amount of MBP present in wtciatic nerves (see below). Fluorescein-conjugated phal-oidin was used to identify the incisures. Examples oftained nerves are shown in Fig. 1. The analysis showedhat 1/shi nerves contained 6.0 incisures per 100 mm,
which is intermediate between wt and shi/shi nerves ofhe same age (3.9 and 7.5 SLI/100 mm for wt and shi/shi
sciatic nerves, respectively) (Gould et al., 1995), indicat-ing that there is an inverse correlation between theamount of MBP present in the nerve and the number ofincisures.
Correlation between MBP Levels and Other MyelinProteins
The relationship between the number of functionalMBP genes and the level of SLI components was quan-tified by immunodot blot analysis of wt, 1/shi, andshi/shi sciatic nerve components. In a first step, theprotein profile of PNS myelin from adult wt and shi/shimice was obtained to insure the identity of the proteinsrecognized by the battery of antisera used.
Membrane and soluble fractions of 90-day-old wt andshi/shi sciatic nerves were analyzed for the presence ofmyelin proteins by Western blot. Compact myelinmembrane proteins P0, PMP22, and MBP, and SLI pro-teins MAG and Cx32 were recovered exclusively in themembrane fraction of both genotypes. Cx32 was also
present in the membrane fraction of liver obtained fromthe same animals.
345Gene Expression in PNS Myelin
FIG. 1. Localization of filamentous actin (fluorescein-conjugated phalloidin), Cx32, and MAG proteins in sciatic nerves. Wt, shi/shi, and 1/shisciatic nerves were incubated with fluorescein-conjugated phalloidin, Cx32 antibodies (1:75) (wt and shi/shi), and MAG antibodies (1:50) (wt and
shi/shi), followed by a Texas red-conjugated secondary antibody (1:50). Schwann cell body (arrow); Schmidt-Lanterman incisures (arrowhead).The scale bar in each panel represents 25 mm.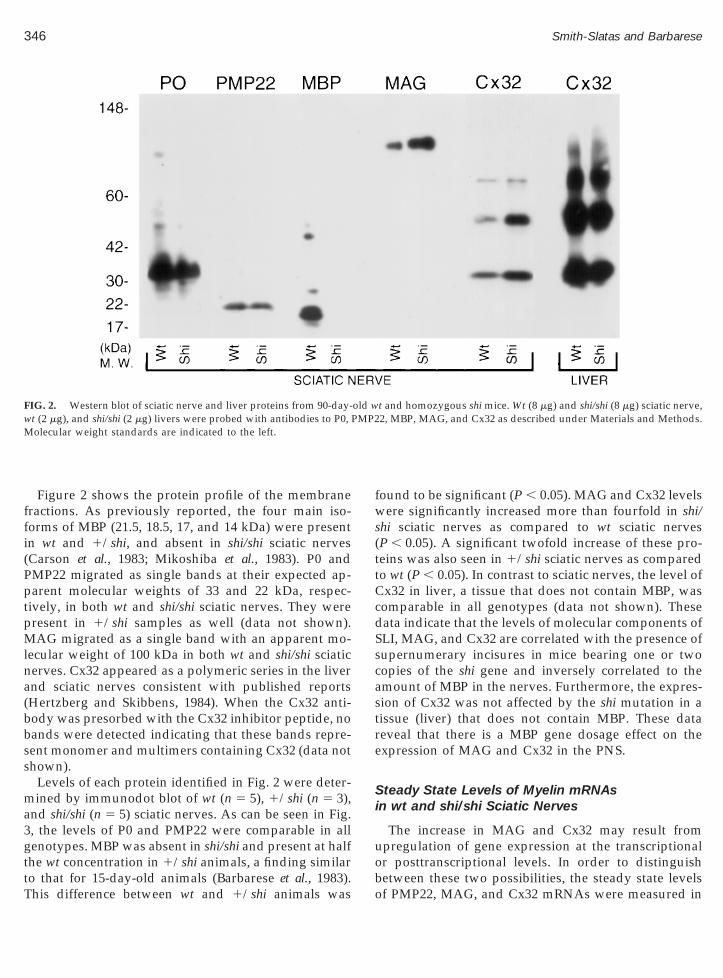
pMlna(bbss
ma3g
T
fws
w PMP2
346 Smith-Slatas and Barbarese
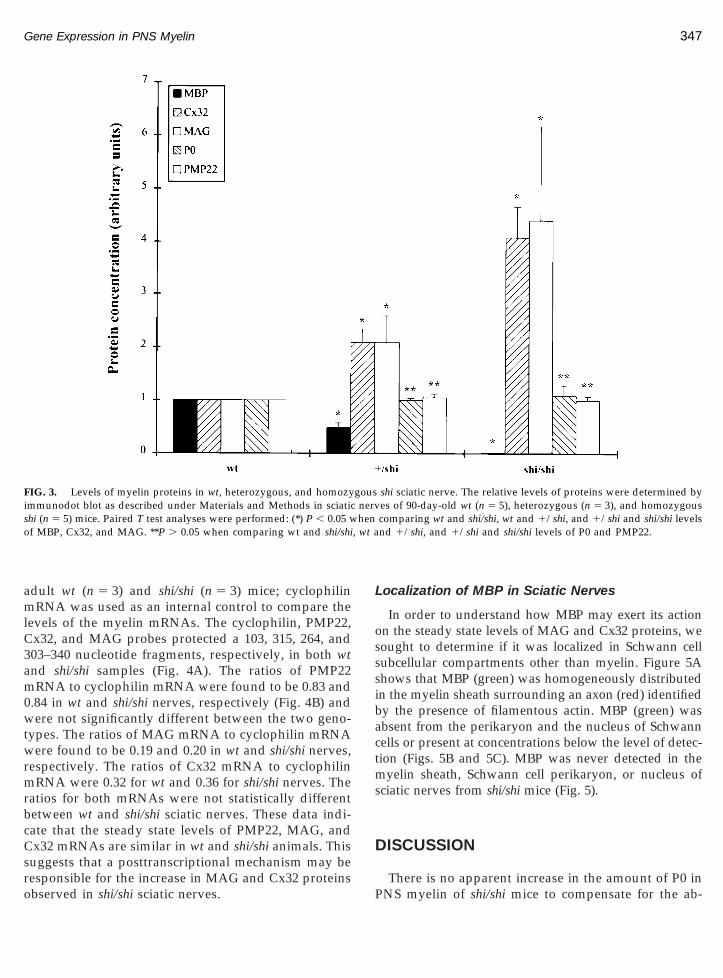
Figure 2 shows the protein profile of the membranefractions. As previously reported, the four main iso-forms of MBP (21.5, 18.5, 17, and 14 kDa) were presentin wt and 1/shi, and absent in shi/shi sciatic nerves(Carson et al., 1983; Mikoshiba et al., 1983). P0 andPMP22 migrated as single bands at their expected ap-parent molecular weights of 33 and 22 kDa, respec-tively, in both wt and shi/shi sciatic nerves. They were
resent in 1/shi samples as well (data not shown).AG migrated as a single band with an apparent mo-
ecular weight of 100 kDa in both wt and shi/shi sciaticerves. Cx32 appeared as a polymeric series in the livernd sciatic nerves consistent with published reportsHertzberg and Skibbens, 1984). When the Cx32 anti-ody was presorbed with the Cx32 inhibitor peptide, noands were detected indicating that these bands repre-ent monomer and multimers containing Cx32 (data nothown).
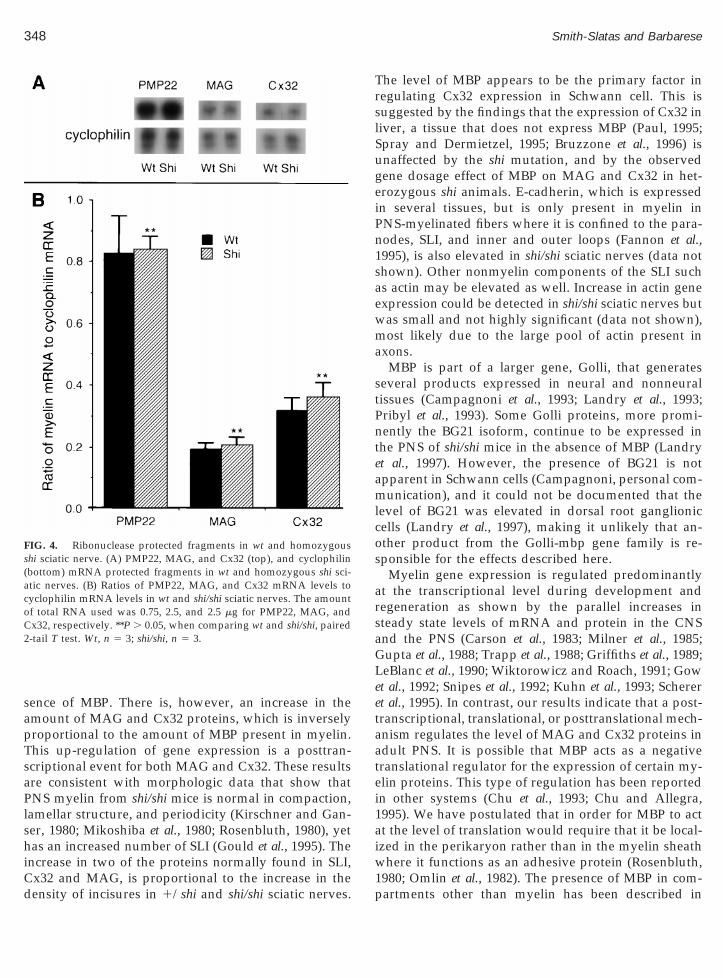
Levels of each protein identified in Fig. 2 were deter-ined by immunodot blot of wt (n 5 5), 1/shi (n 5 3),
nd shi/shi (n 5 5) sciatic nerves. As can be seen in Fig., the levels of P0 and PMP22 were comparable in allenotypes. MBP was absent in shi/shi and present at half
the wt concentration in 1/shi animals, a finding similar
FIG. 2. Western blot of sciatic nerve and liver proteins from 90-day-t (2 mg), and shi/shi (2 mg) livers were probed with antibodies to P0,
Molecular weight standards are indicated to the left.
to that for 15-day-old animals (Barbarese et al., 1983).his difference between wt and 1/shi animals was
ound to be significant (P , 0.05). MAG and Cx32 levelsere significantly increased more than fourfold in shi/
hi sciatic nerves as compared to wt sciatic nerves(P , 0.05). A significant twofold increase of these pro-teins was also seen in 1/shi sciatic nerves as comparedto wt (P , 0.05). In contrast to sciatic nerves, the level ofCx32 in liver, a tissue that does not contain MBP, wascomparable in all genotypes (data not shown). Thesedata indicate that the levels of molecular components ofSLI, MAG, and Cx32 are correlated with the presence ofsupernumerary incisures in mice bearing one or twocopies of the shi gene and inversely correlated to theamount of MBP in the nerves. Furthermore, the expres-sion of Cx32 was not affected by the shi mutation in atissue (liver) that does not contain MBP. These datareveal that there is a MBP gene dosage effect on theexpression of MAG and Cx32 in the PNS.
Steady State Levels of Myelin mRNAsin wt and shi/shi Sciatic Nerves
The increase in MAG and Cx32 may result fromupregulation of gene expression at the transcriptionalor posttranscriptional levels. In order to distinguish
t and homozygous shi mice. Wt (8 mg) and shi/shi (8 mg) sciatic nerve,2, MBP, MAG, and Cx32 as described under Materials and Methods.
old w
between these two possibilities, the steady state levelsof PMP22, MAG, and Cx32 mRNAs were measured in
mlC3am0wtw
rbcC
hen, wt a
347Gene Expression in PNS Myelin
adult wt (n 5 3) and shi/shi (n 5 3) mice; cyclophilinRNA was used as an internal control to compare the
evels of the myelin mRNAs. The cyclophilin, PMP22,x32, and MAG probes protected a 103, 315, 264, and03–340 nucleotide fragments, respectively, in both wtnd shi/shi samples (Fig. 4A). The ratios of PMP22RNA to cyclophilin mRNA were found to be 0.83 and
.84 in wt and shi/shi nerves, respectively (Fig. 4B) andere not significantly different between the two geno-
ypes. The ratios of MAG mRNA to cyclophilin mRNAere found to be 0.19 and 0.20 in wt and shi/shi nerves,
respectively. The ratios of Cx32 mRNA to cyclophilinmRNA were 0.32 for wt and 0.36 for shi/shi nerves. Theatios for both mRNAs were not statistically differentetween wt and shi/shi sciatic nerves. These data indi-ate that the steady state levels of PMP22, MAG, andx32 mRNAs are similar in wt and shi/shi animals. This
suggests that a posttranscriptional mechanism may be
FIG. 3. Levels of myelin proteins in wt, heterozygous, and homozyimmunodot blot as described under Materials and Methods in sciaticshi (n 5 5) mice. Paired T test analyses were performed: (*) P , 0.05 wof MBP, Cx32, and MAG. **P . 0.05 when comparing wt and shi/shi
responsible for the increase in MAG and Cx32 proteinsobserved in shi/shi sciatic nerves.
Localization of MBP in Sciatic Nerves
In order to understand how MBP may exert its actionon the steady state levels of MAG and Cx32 proteins, wesought to determine if it was localized in Schwann cellsubcellular compartments other than myelin. Figure 5Ashows that MBP (green) was homogeneously distributedin the myelin sheath surrounding an axon (red) identifiedby the presence of filamentous actin. MBP (green) wasabsent from the perikaryon and the nucleus of Schwanncells or present at concentrations below the level of detec-tion (Figs. 5B and 5C). MBP was never detected in themyelin sheath, Schwann cell perikaryon, or nucleus ofsciatic nerves from shi/shi mice (Fig. 5).
DISCUSSION
shi sciatic nerve. The relative levels of proteins were determined byes of 90-day-old wt (n 5 5), heterozygous (n 5 3), and homozygouscomparing wt and shi/shi, wt and 1/shi, and 1/shi and shi/shi levelsnd 1/shi, and 1/shi and shi/shi levels of P0 and PMP22.
gousnerv
There is no apparent increase in the amount of P0 inPNS myelin of shi/shi mice to compensate for the ab-
ls
TrslSugeiPn1saewma
stPnteamlcos
arsaG
etaatei
C2
348 Smith-Slatas and Barbarese
sence of MBP. There is, however, an increase in theamount of MAG and Cx32 proteins, which is inverselyproportional to the amount of MBP present in myelin.This up-regulation of gene expression is a posttran-scriptional event for both MAG and Cx32. These resultsare consistent with morphologic data that show thatPNS myelin from shi/shi mice is normal in compaction,amellar structure, and periodicity (Kirschner and Gan-er, 1980; Mikoshiba et al., 1980; Rosenbluth, 1980), yet
has an increased number of SLI (Gould et al., 1995). Theincrease in two of the proteins normally found in SLI,
FIG. 4. Ribonuclease protected fragments in wt and homozygousshi sciatic nerve. (A) PMP22, MAG, and Cx32 (top), and cyclophilin(bottom) mRNA protected fragments in wt and homozygous shi sci-atic nerves. (B) Ratios of PMP22, MAG, and Cx32 mRNA levels tocyclophilin mRNA levels in wt and shi/shi sciatic nerves. The amountof total RNA used was 0.75, 2.5, and 2.5 mg for PMP22, MAG, and
x32, respectively. **P . 0.05, when comparing wt and shi/shi, paired-tail T test. Wt, n 5 3; shi/shi, n 5 3.
Cx32 and MAG, is proportional to the increase in thedensity of incisures in 1/shi and shi/shi sciatic nerves.
he level of MBP appears to be the primary factor inegulating Cx32 expression in Schwann cell. This isuggested by the findings that the expression of Cx32 iniver, a tissue that does not express MBP (Paul, 1995;pray and Dermietzel, 1995; Bruzzone et al., 1996) isnaffected by the shi mutation, and by the observedene dosage effect of MBP on MAG and Cx32 in het-rozygous shi animals. E-cadherin, which is expressedn several tissues, but is only present in myelin inNS-myelinated fibers where it is confined to the para-odes, SLI, and inner and outer loops (Fannon et al.,995), is also elevated in shi/shi sciatic nerves (data nothown). Other nonmyelin components of the SLI suchs actin may be elevated as well. Increase in actin genexpression could be detected in shi/shi sciatic nerves butas small and not highly significant (data not shown),ost likely due to the large pool of actin present in
xons.MBP is part of a larger gene, Golli, that generates
everal products expressed in neural and nonneuralissues (Campagnoni et al., 1993; Landry et al., 1993;ribyl et al., 1993). Some Golli proteins, more promi-ently the BG21 isoform, continue to be expressed in
he PNS of shi/shi mice in the absence of MBP (Landryt al., 1997). However, the presence of BG21 is notpparent in Schwann cells (Campagnoni, personal com-unication), and it could not be documented that the
evel of BG21 was elevated in dorsal root ganglionicells (Landry et al., 1997), making it unlikely that an-ther product from the Golli-mbp gene family is re-ponsible for the effects described here.
Myelin gene expression is regulated predominantlyt the transcriptional level during development andegeneration as shown by the parallel increases inteady state levels of mRNA and protein in the CNSnd the PNS (Carson et al., 1983; Milner et al., 1985;upta et al., 1988; Trapp et al., 1988; Griffiths et al., 1989;
LeBlanc et al., 1990; Wiktorowicz and Roach, 1991; Gowet al., 1992; Snipes et al., 1992; Kuhn et al., 1993; Scherert al., 1995). In contrast, our results indicate that a post-ranscriptional, translational, or posttranslational mech-nism regulates the level of MAG and Cx32 proteins indult PNS. It is possible that MBP acts as a negativeranslational regulator for the expression of certain my-lin proteins. This type of regulation has been reportedn other systems (Chu et al., 1993; Chu and Allegra,
1995). We have postulated that in order for MBP to actat the level of translation would require that it be local-ized in the perikaryon rather than in the myelin sheathwhere it functions as an adhesive protein (Rosenbluth,
1980; Omlin et al., 1982). The presence of MBP in com-partments other than myelin has been described in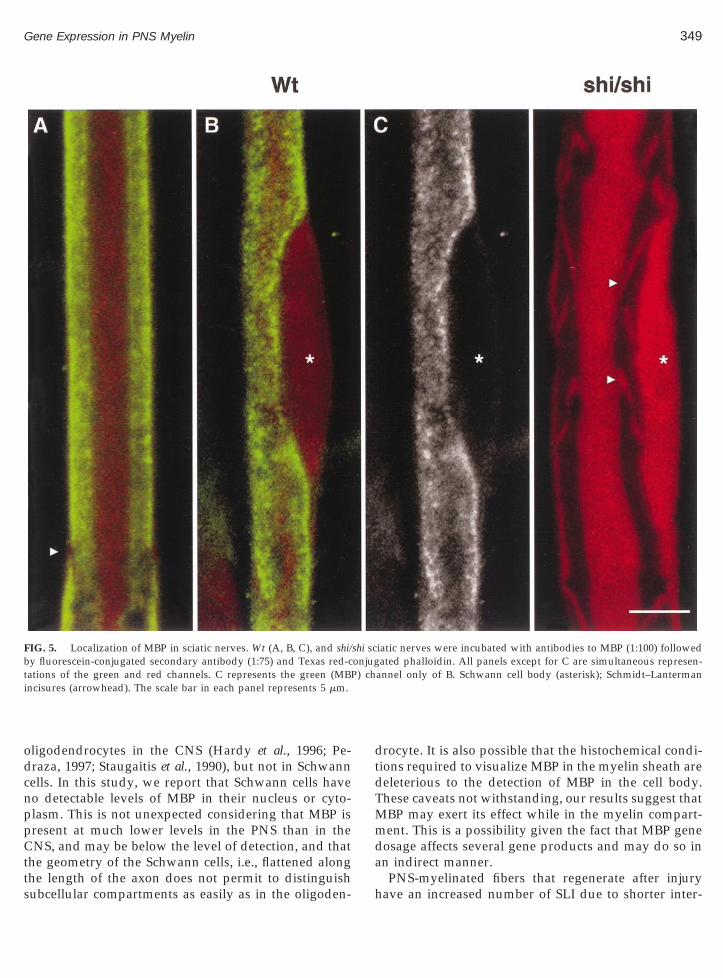
cnppCtts
) ch
349Gene Expression in PNS Myelin
oligodendrocytes in the CNS (Hardy et al., 1996; Pe-draza, 1997; Staugaitis et al., 1990), but not in Schwannells. In this study, we report that Schwann cells haveo detectable levels of MBP in their nucleus or cyto-lasm. This is not unexpected considering that MBP isresent at much lower levels in the PNS than in theNS, and may be below the level of detection, and that
he geometry of the Schwann cells, i.e., flattened along
FIG. 5. Localization of MBP in sciatic nerves. Wt (A, B, C), and shi/by fluorescein-conjugated secondary antibody (1:75) and Texas red-ctations of the green and red channels. C represents the green (MBPincisures (arrowhead). The scale bar in each panel represents 5 mm.
he length of the axon does not permit to distinguishubcellular compartments as easily as in the oligoden-
drocyte. It is also possible that the histochemical condi-tions required to visualize MBP in the myelin sheath aredeleterious to the detection of MBP in the cell body.These caveats not withstanding, our results suggest thatMBP may exert its effect while in the myelin compart-ment. This is a possibility given the fact that MBP genedosage affects several gene products and may do so inan indirect manner.
iatic nerves were incubated with antibodies to MBP (1:100) followedated phalloidin. All panels except for C are simultaneous represen-
annel only of B. Schwann cell body (asterisk); Schmidt–Lanterman
shi sconjug
PNS-myelinated fibers that regenerate after injuryhave an increased number of SLI due to shorter inter-

1attdvm(bsttnbs
caicb
350 Smith-Slatas and Barbarese
nodes (Ghabriel and Allt, 1980; Hiscoe, 1947). The levelsof MBP transcripts and protein that accumulate in theregenerated fibers, however, is comparable to that ofuninjured animals suggesting that they are not influ-enced by that morphologic peculiarity (Gupta et al.,988; LeBlanc and Poduslo, 1990). The increase in MAGnd Cx32 levels and in the number of incisures wherehey are found may represent an adaptive mechanismo low levels or absence of MBP. A recent study hasemonstrated that gap junctions in the incisures pro-ide a radial pathway for the rapid diffusion of smallolecules from the outer to the inner myelin layers
Balice-Gordon et al., 1998). Thus, the role of SLI haseen postulated to be in the delivery of nutrients to theheath and in removal of toxic metabolites. It is possiblehat the stability of the myelin sheath is compromised inhe absence of MBP, and that removal of componentseeds to be accelerated, hence the increase in the num-er of incisures. In that context, the increase in inci-ures, and therefore in Cx32 and MAG, in shi/shi sciatic
nerves would represent a part in a general responseaimed at maintaining the integrity of the MBP-deficientmyelin sheath by facilitating more active transport ofnutrients and removal of toxic metabolites (Paul, 1995;Scherer et al., 1995). However, our earlier study indi-ates that the increase in SLI in shi/shi sciatic nerves ispparent as soon as myelin starts being made, suggest-ng that MBP must affect the initial levels of incisuresomponents, rather than their levels after myelin hasecome compact (Gould et al., 1995). Furthermore, there
is no evidence that shi/shi myelin is degenerating, andthe number of Schwann cells which increases in regen-erating fibers and leads to shorther myelin internodes(Ghabriel and Allt, 1980) appears to be the same inshi/shi and wt mice (Gould et al., 1995). Thus, a morelikely explanation is that MBP directly or indirectlyregulates the expression of Cx32 and MAG and possiblyother myelin and nonmyelin components as well dur-ing normal development. This would represent a newrole for this protein traditionally considered to be solelya structural element of the myelin membrane.
MBP could act to affect the stability or the turnoverrate of proteins of the SLI, or could act as an inhibitor ofprotein translation. There is no evidence that MBP is anRNA binding protein that could directly regulate thetranslation of Cx32 mRNA or MAG mRNA. Nor is thereany data on possible regulatory sites in Cx32 or MAGmRNA. There are several mechanisms for the control oftranslation initiation in animals (Gray and Wickens,1998) and one or more can be at play for either Cx32 or
MAG mRNA. Analysis of Cx32 mRNA with an RNAfolding program reveals the presence of putative stem–loop structures with a single-stranded RNA bulge sim-ilar to those which have been implicated in negativetranslational regulation of mRNAs such as ferritin andtransferrin mRNAs (Chu et al., 1993; Burd and Drey-fuss, 1994; Chu and Allegra, 1995; McCarthy and Koll-mus, 1995; Henderson and Kuhn, 1997; Muckenthalerand Hentze, 1997). It seems unlikely, however, giventhe complexity of the SLI, that MBP regulates all theircomponents directly or by the same mechanism.
EXPERIMENTAL METHODS
Animals. Wt and 1/shi mice with the same geneticbackground (C3HeB/FeJ) were obtained from the Jack-son Laboratories (Bar Harbour, ME). The latter weremated to produce shi/shi animals. Mice were euthanizedaccording to a protocol approved by the committee forLaboratory Animal Care at the University of Connecti-cut Health Center. Sciatic nerves were removed andfixed in 4% buffered paraformaldehyde solution, orsnap frozen in liquid nitrogen, and stored at 275°C.Livers were removed and processed in the lattermanner.
Immunofluorescence. Sciatic nerves were desheathedand teased into fiber bundles. For staining with anti-MBP, the nerves were permeabilized with a solution of5% acetic acid in methanol for 10 min. All nerves werefurther incubated in Buffer A (0.1% Triton X-100, 5%normal goat serum, 0.1 M NaCl, 0.05 M Tris–HCl, pH7.6) for 30 min at room temperature, with gentle rock-ing. Incubations in primary and secondary antibodiesdiluted in Buffer A were done overnight at 4°C withrocking. Each incubation with antibodies was followedby four 10 min washes in Buffer B (0.05% Triton X-100,0.1 M NaCl, 0.05 M Tris–HCl, pH 7.6) at room temper-ature with rocking. Nerves were otherwise processedaccording to standard protocols and examined byconfocal fluorescence microscopy. Mouse anti-Cx32(R5.21C) was used at 1:75 dilution. Mouse anti-MAGwas used at 1:50 dilution. Mouse anti-MBP (SternbergerMonoclonals Inc., Baltimore, MD) was used at a 1:100dilution. Texas-red-conjugated goat anti-mouse second-ary antibody was used at 1:50 dilution. Fluorescein-conjugated goat anti-mouse secondary antibody wasused at 1:75 dilution.
Phalloidin staining and SLI counts. Two fixed sci-atic nerves from two 1/shi mice were incubated influorescein-conjugated phalloidin (Sigma Chemical Co.,St. Louis, MO) (1:70 dilution in PBS) overnight at 4°C
and examined by confocal fluorescence microscopy. Atotal of 209 myelinated axons were sampled, and the
iCo
lw(
mur
pafmtnad31gccCm
ba
(M
351Gene Expression in PNS Myelin
number of SLI was determined as described in Gould etal. (1995). Sciatic nerves from wt and shi/shi mice werencubated with Texas red-conjugated phalloidin (Sigmahemical Co., St. Louis, MO) (1:70 dilution in PBS)vernight at 4°C.Tissue preparation. The detergent-independent iso-
ation of sciatic nerves and liver membranous fractionas performed according to the method of Hertzberg
Hertzberg, 1984).Protein determination. Protein content was deter-ined by the method of Lowry (Lowry et al., 1951)
sing the BioRad DC Protein Assay kit (Bio-Rad Labo-atories, Hercules, CA).
Western blot. Protein samples were run on 8–16%olyacrylamide gradient gels (Novex, San Diego, CA)ccording to the method of Laemmli (1970) and trans-erred to supported nitrocellulose according to the
ethod of Towbin (Towbin et al., 1979). Immunodetec-ion was performed with antibodies to myelin compo-ents and horseradish peroxidase-conjugated second-ry antibodies. Antibodies were used at the followingilutions: mouse anti-MAG (1/1000), rabbit anti-P0 (1/000), rabbit anti-PMP22 (1/2000), mouse anti-MBP (1/000), mouse anti-Cx32 (1/1000), horseradish-conju-ated donkey anti-rabbit (1/1000), and horseradish-onjugated sheep anti-mouse (1/1000). Enhancedhemiluminescence reagents (Supersubstrate kit, Piercehemical Co., Rockford, IL) were used to detect im-une complexes.Immuno-dot blot and quantitative analysis. Solu-
ilized membrane fractions from 90-day-old wt, 1/shi,nd shi/shi sciatic nerves having the same protein con-
centration were serially diluted and blotted in duplicatedirectly onto supported nitrocellulose. Immunodetec-tion was performed as for Western blot. The immunecomplexes visualized on X-ray films were photo-graphed with a digital camera, and densitometry andquantification were done using the Molecular Dynam-ics ImageQuaNT analysis system (Molecular Dynamics,Sunnyvale, CA). The intensity levels of P0, PMP22,MBP, MAG, and Cx32 proteins were measured in wt,1/shi, and shi/shi samples, and those in wt sampleswere set arbitrarily to 1. The amounts of the proteinsfrom 1/shi and shi/shi were expressed relative to the wtvalues. Protein levels were determined in at least fiveindependent experiments for wt and shi/shi samples andin at least three independent experiments for 1/shisamples. Each sample consisted of two sciatic nervesfrom the same animal. The mean and standard devia-
tion were calculated for each protein and a paired T testwas performed using Microsoft Excel (Microsoft Corp.).RNA isolation. Total RNA was isolated from sciaticnerves with Tri Reagent (Sigma BioSciences, St. Louis,MO) according to the manufacturer’s instructions. Eachpreparation consisted of two sciatic nerves crushed inliquid nitrogen and suspended in 1 ml of Tri Reagent.The yield of RNA averaged around 1 mg per sciaticnerve. RNA samples were stored at 275°C until use.
Ribonuclease protection assay (RPA). RPA wasperformed using the Hybspeed RPA kit from Ambion(Ambion, Austin, TX) according to the manufacturer’sinstructions. The protected fragments were separatedby electrophoresis on 6% TBE-Urea gels (Novex, SanDiego, CA) and detected by autoradiography. Filmswere photographed with a digital camera, and quanti-fication by densitometry was done using MolecularDynamics ImageQuaNT, Version 4.2 (Molecular Dy-namics, Sunnyvale, CA). At least three independentRPA determinations were done for each mRNA in wtand shi/shi sciatic nerve samples. The mean and stan-dard deviation of the ratios of myelin mRNA to controlmRNA intensity values were calculated and a paired Ttest was performed using Microsoft Excel (MicrosoftCorp.).
cDNA templates. The following cloned DNAs wereused to prepare the antisense RNA probes: a 515-bp ratPMP22 cDNA inserted in the HindIII site of pRc/CMVD’Urso, personal communication); a 2,049-bp rat L-
AG cDNA inserted in the ApaI site of Bluescript KS1(Salzer et al., 1987); and a 1458-bp rat Cx32 cDNA in-serted in the KpnI site of pGEM3Zf1 (Werner et al.,1991).
Antisense RNA probes. Template for cyclophilinwas obtained from Ambion (Ambion Inc., Austin, TX).The cyclophilin antisense probe was 165 nucleotideslong and protected a 103-nucleotide fragment. ThePMP22 containing plasmid was cut with XmnI and SP6polymerase was used to make a 424 nucleotide probe,which protected a 315-nucleotide fragment. The Cx32containing plasmid cut with KpnI and T7 polymerasewas used to make a 281-nucleotide probe, which pro-tected a 264-nucleotide fragment. The L-MAG contain-ing plasmid cut with EcoRI and T3 polymerase wasused to make a 361-nucleotide probe, which protected a341-nucleotide fragment of L-MAG and a 303-nucleo-tide fragment of S-MAG. These two fragments couldnot be resolved in the gel system used.
In vitro transcription of [a-32P]UTP (800 Ci/mmol)(DuPont NEN, Boston, MA) labeled antisense RNAprobes and RNA markers was performed with theMaxiscript In Vitro Transcription kit from Ambion
(Ambion Inc., Austin, TX), according to the manufac-turer’s instructions. Labeled probes were gel purified
M(gbMpmbCSMZpfacjsPrwiorL(NWaw
B
B
B
B
C
C
C
C
D
F
F
G
G
G
G
G
352 Smith-Slatas and Barbarese
using 6% TBE-Urea gels (Novex, San Diego, CA). TheRNA Century Marker Plus Template set was from Am-bion.
Antibodies, plasmids, and reagents. Mouse anti-AG antibody was a generous gift from Dr. R. Quarles
NIH, Bethesda, MD). Rabbit anti-P0 was a generousift from Dr. M. Filbin (Hunter College, NY, NY). Rab-it anti-PMP22 was a generous gift from Dr. H. W.uller (Laboratory for Molecular Neurobiology, De-
artment of Neurology, University of Dusseldorf, Ger-any). Mouse anti-MBP was purchased from Stern-
erger Monoclonals Inc. (Baltimore, MD). Mouse anti-x32 (R5.21C) was obtained from The Developmentaltudies Hybridoma Bank (University of Iowa, IA).ouse and rabbit anti-Cx32 were purchased from
ymed Laboratories (San Francisco, CA). An inhibitoreptide for the Cx32 antibodies was also purchased
rom Zymed Laboratories. Texas red-conjugated goatnti-mouse IgG was purchased from Accurate Chemi-al & Scientific Corp. (Westbury, NY). Fluorescein-con-ugated goat anti-mouse IgG was purchased from Jack-on ImmunoResearch Laboratories, Inc. (West Grove,A). Horseradish peroxidase-conjugated donkey anti-abbit, sheep anti-rat, and sheep anti-mouse antibodiesere purchased from Amersham Life Sciences (Buck-
nghamshire, England). Rat PMP22 cDNA was a gener-us gift from Drs. H. W. Muller and D. D’Urso (Hein-ich-Heine-University, Dusseldorf, Germany). Rat-MAG cDNA was a generous gift from Dr. J. Salzer
New York University School of Medicine, New York,Y). Rat Cx32 cDNA was a generous gift from Dr. R.erner (University of Miami School of Medicine, Mi-
mi, FL). Except where otherwise noted all reagentsere from Sigma Chemical Co. (St. Louis, MO).
ACKNOWLEDGMENTS
This work was supported by NIH Grant NS19943 (E.B.), and by agraduate fellowship from the U. of CT. Health Ctr. (C.S.-S). We thankDrs. D’Urso, M. Filbin, H. W. Muller, R. Quarles, and J. Salzer for theirgenerous gifts of reagents, and Dr. D. Oliver (U. Conn. Health Ctr.Farmington, CT) for his comments on the manuscript. We also thankFrank Morgan and Maureen Pons for technical assistance. The Cx32antibody developed by Dr. Daniel Goodenough was obtained formthe Developmental Studies Hybridoma Bank maintained by The Uni-versity of Iowa, Department of Biological Sciences, Iowa City, IA52242, under contract NO1-HD-7-3263 from the NICHD.
REFERENCES
Anderson, T. J., Montague, P., Nadon, N., Nave, K.-A., and Griffiths,I. R. (1997). Modification of Schwann cell phenotype with Plp
transgenes: Evidence that the PLP and DM20 isoproteins are tar-geted to different cellular domains. J. Neurosci. Res. 50: 13–22.
Balice-Gordon, R. J., Bone, L. J., and Scherer, S. S. (1998). Functionalgap junctions in the Schwann cell myelin sheath. J. Cell Biol. 142:1095–1104.
arbarese, E., Nielson, M. L., and Carson, J. H. (1983). The effect of theshiverer mutation on myelin basic protein expression in homozy-gous and heterozygous mouse brain. J. Neurochem. 40: 1680–1686.
ergoffen, J., Scherer, S. S., Wang, S., Scott, M. O., Bone, L. J., Paul,D. L., Chen, K., Lensch, M. W., Chance, P. F., and Fischbeck, K. H.(1993). Connexin mutations in X-linked Charcot-Marie-Tooth dis-ease. Science 262: 2039–2042.
ruzzone, R., White, T. W., and Paul, D. (1996). Connections withconnexins: The molecular basis of direct intercellular signaling. Eur.J. Biochem. 238: 1–27.
urd, C. G., and Dreyfuss, G. (1994). Conserved structures and diver-sity of functions of RNA-binding proteins. Science 265: 615–621.
ampagnoni, A. T., Pribyl, T. M., Campagnoni, C. W., Kampf, K.,Amur-Umarjee, S., Landry, C. F., Handley, V. W., Newman, S. L.,Garbay, B., and Kitamura, K. (1993). Structure and developmentalregulation of Golli-mbp, a 105-kilobase gene that encompasses themyelin basic protein gene and is expressed in cells in the oligoden-drocyte lineage in the brain. J. Biol. Chem. 268: 4930–4938.
arson, J. H., Nielson, M. L., and Barbarese, E. (1983). Developmentalregulation of myelin basic protein expression in mouse brain. Dev.Biol. 96: 485–492.
hu, E., and Allegra, C. J. (1995). The role of thymidylate synthase asan RNA binding protein. BioEssays 18: 191–198.
hu, E., Voeller, D., Koeller, D. M., Drake, J. C., Takimoto, C. H.,Maley, G. F., and Allegra, C. J. (1993). Identification of an RNAbinding site for human thymidylate synthase. Proc. Natl. Acad. Sci.USA 90: 517–521.’Urso, D., Brophy, P. J., Staugaitis, S. M., Gillespie, C. S., Frey, A. B.,Stempak, J. G., and Colman, D. R. (1990). Protein zero of peripheralnerve myelin: Biosynthesis, membrane insertion, and evidence forhomotypic interaction. Neuron 2: 449–460.
annon, A. M., Sherman, D. L., Ilyina-Gragerova, G., and Brophy, P.J. (1995). Novel E-cadherin-mediated adhesion in peripheral nerve:Schwann cell architecture is stabilized by autotypic adherens junc-tions. J. Cell Biol. 129: 189–202.
ilbin, M. T., Walsh, F. S., Trapp, B. D., Pizzey, J. A., and Tennekoon,G. I. (1990). Role of myelin P0-protein as a homophilic adhesionmolecule. Nature 344: 871–872.arbern, J. Y., Cambi, F., Tang, X.-M., Sima, A. A. F., Vallat, J. M.,Bosch, E. P., Lewis, R., Shy, M., Sohi, J., Kraft, G., Chen, K. L., Joshi,I., Leonard, D. G. B., Johnson, W., Raskind, W., Dlouhy, S. R., Pratt,V., Hodes, E. M., Bird, T., and Kamholz, J. (1997). Proteolipidprotein is necessary in peripheral as well as central myelin. Neuron19: 205–218.habriel, M. M., and Allt, G. (1980). Schmidt-Lanterman incisures I. Aquantitative teased fibre study of remyelinating peripheral nervefibres. Acta Neuropathol. 52: 85–95.ould, R. M., Byrd, A. L., and Barbarese, E. (1995). The number ofSchmidt-Lanterman incisures is more than doubled in shiverer PNSmyelin sheaths. J. Neurocytol. 24: 85–98.ow, A., Friedrich, V. L., and Lazzarini, R. A. (1992). Myelin basicprotein gene contains separate enhancers for oligodendrocyte andSchwann cell expression. J. Cell Biol. 119: 605–616.ow, A., Friedrich, V. L., and Lazzarini, R. A. (1994). Many naturally
occurring mutations of myelin proteolipid protein impair its intra-cellular transport. J. Neurosci. Res. 37: 574–583.
G
H
H
H
H
H
K
K
L
L
L
L
L
M
M
M
M
M
M
M
O
P
P
P
P
P
P
P
R
R
R
S
353Gene Expression in PNS Myelin
Gray, N. K., and Wickens, M. (1998). Control of translation initiatioinin animals. Annu. Rev. Cell Dev. Biol. 14: 399–458.
Greenfield, S., Brostoff, S., Eylar, E. H., and Morell, P. (1973). Proteincomposition of myelin of the peripheral nervous system. J. Neuro-chem. 20: 1207–1216.riffiths, I. R., Dickinson, P., and Montague, P. (1995). Expression ofthe proteolipid protein gene in glial cells of the post-natal periph-eral nervous system of rodents. Neuropathol. Appl. Neurobiol. 21:97–110.
Griffiths, I. R., Mitchell, L. S., McPhilemy, K., Morrison, S., Kyriakides,E., and Barrie, J. A. (1989). Expression of myelin protein genes inSchwann cells. J. Neurocytol. 18: 345–352.
Gupta, S. K., Poduslo, J. F., Dunn, R., Roder, J., and Mezei, C. (1990).Myelin-associated glycoprotein gene expression in the presenceand absence of Schwann cell-axonal contact. Dev. Neurosci. 12:22–23.
Gupta, S. K., Poduslo, J. F., and Mezei, C. (1988). Temporal changes inPO and MBP gene expression after crush-injury of the adult pe-ripheral nerve. Mol. Brain Res. 4: 133–141.ardy, R. J., Lazzarini, R. A., Colman, D. R., and Friedrich, V. L.(1996). Cytoplasmic and nuclear localization of myelin basic proteinreveals heterogeneity among oligodendrocytes. J. Neurosci. Res. 46:246–257.enderson, B. R., and Kuhn, L. C. (1997). Interactions between iron-regulatory proteins and their RNA target sequences, iron-respon-sive elements. Prog. Mol. Subcell. Biol. 18: 117–139.ertzberg, E. L., and Skibbens, R. V. (1984). A protein homologous tothe 27,000 Dalton liver gap junction protein is present in a widevariety of species and tissues. Cell 39: 61–69.ertzberg, E. L. (1984). A detergent-independent procedure for theisolation of gap junctions from rat liver. J. Biol. Chem. 259: 9936–9943.iscoe, E. B. (1947). Distribution of nodes and incisures in normal andregenerated nerve fibres. Anat. Rec. 99: 447–475.
irschner, D. A., and Ganser, A. L. (1980). Compact myelin exists inthe absence of basic protein in the shiverer mutant mouse. Nature283: 207–210.
uhn, G., Lie, A., Wilms, S., and Muller, H. W. (1993). Coexpressionof PMP22 gene with MBP and P0 during de novo myelination andnerve repair. Glia 8: 256–264.
aemmli, U. K. (1970). Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature 227: 680–685.
andry, C. F., Ellison, J. A., Pribyl, T. M., Campagnoni, A. T., Kampf,K., and Campagnoni, A. T. (1993). Myelin basic protein gene ex-pression in neurons: Developmental and regional changes in pro-tein targeting within neuronal nuclei, cell bodies, and processes. J.Neurosci. 16: 2452–2462.
andry, C. F., Ellison, J., Skinner, E., and Campagnoni, A. T. (1997).Golli-MBP proteins mark the earliest stages of fibre extension andterminal arboration in the mouse peripheral nervous system.J. Neurosci. Res. 50: 265–271.
eBlanc, A. C., and Poduslo, J. F. (1990). Axonal modulation of myelingene expression in the peripheral nerve. J. Neurosci. Res. 26: 317–326.
owry, O. H., Rosebrough, N. J., Farr, A. L., and Randall, R. J. (1951).Protein measurement with the folin phenol reagent. J. Biol. Chem.193: 265–275.artini, R., Mohajeri, M. H., Kasper, S., Giese, K. P., and Schachner,M. (1995). Mice doubly deficient in the genes for P0 and myelinbasic protein show that both proteins contribute to the formation of
the major dense line in peripheral nerve myelin. J. Neurosci. 15:4488–4495.ata, M., Alessi, D., and Fink, D. (1990). S100 is preferentially dis-tributed in myelin-forming Schwann cells. J. Neurocytol. 19: 432–442.cCarthy, J. E. G., and Kollmus, K. (1995). Cytoplasmic mRNA–protein interactions in eukaryotic gene expression. Trends Biochem.Sci. 20: 191–197.ikoshiba, K., Kohsaka, S., Takamatsu, K., and Tsukada, Y. (1980).Neurochemical and morphological studies on the myelin of theperipheral nervous system from shiverer mutant mice: Absence ofbasic proteins common to central nervous system. Brain Res. 204:455–460.ikoshiba, K., Takamatsu, K., and Tsukada, Y. (1983). Peripheralnervous system of shiverer mutant mice: Developmental change ofmyelin components and immunohistochemical demonstration ofthe absence of MBP and presence of P2 protein. Dev. Brain Res. 7:71–79.ilner, R. J., Lai, C., Nave, K.-A., Lenoir, D., Ogata, J., and Sutcliffe,J. G. (1985). Nucleotide sequences of two mRNAs for rat brainmyelin proteolipid protein. Cell 42: 931–939.uckenthaler, M., and Hentze, M. W. (1997). Mechanisms for post-transcriptional regulation by iron-responsive elements and ironregulatory proteins. Prog. Mol. Subcell. Biol. 18: 93–115.mlin, F. X., Webster, H. D., Palkovits, C. G., and Cohen, S. R. (1982).Immunocytochemical localization of basic protein in major denseline regions of the central and peripheral myelin. J. Cell Biol. 95:242–248.
aul, D. L. (1995). New functions for gap junctions. Curr. Opin. CellBiol. 7: 665–672.
edraza, L. (1997). Nuclear transport of myelin basic protein.J. Neurosci. Res. 50: 258–264.
eters, A., Palay, S. L., and Webster, H. deF. (1991). The fine structureof the nervous system: Neurons and their supporting cells. OxfordUniv. Press, New York.
eterson, A. C., and Bray, G. M. (1984). Hypomyelination in theperipheral nervous system of the shiverer mice and shiverer 7normal chimaera. J. Comp. Neurol. 227: 348–356.
opko, B., Puckett, C., Lai, E., Shine, H. D., Readhead, C., Takahashi,N., Hunt, S. W., Sidman, R. L., and Hood, L. (1987). Myelin deficientmice: Expression of myelin basic protein and generation of micewith varying levels of myelin. Cell 48: 713–721.
ribyl, T. M., Campagnoni, C. W., Kampf, K., Kashima, T., Handley,V. W., McMahon, J., and Campagnoni, A. T. (1993). The 179 kilo-base human golli-mbp gene: Structure and expression in the im-mune and central nervous system. Proc. Natl. Acad. Sci. USA 90:10695–10699.
uckett, C., Hudson, L., Ono, K., Friedrich, V., Benecke, J., Dubois-Dalcq, M., and Lazzarini, R. A. (1987). Myelin-specific proteolipidprotein is expressed in myelinating Schwann cells but is not incor-porated into myelin sheaths. J. Neurosci. Res. 18: 511–518.
eadhead, C., Popko, B., Takahashi, N., Sidman, R. L., and Hood, L.(1987). Expression of myelin basic protein gene in transgenic shiv-erer mice: Correction of the dysmyelinating phenotype. Cell 48:703–712.
oach, A., Boylan, K., Horvath, S., Prusiner, S. B., and Hood, L. E.(1983). Characterization of cloned cDNA representing rat myelinbasic protein: Absence of expression in brain of shiverer mutantmice. Cell 34: 799–806.
osenbluth, J. (1980). Peripheral myelin in the mouse mutant shiverer.J. Comp. Neurol. 193: 729–739.
alzer, J. L., Holmes, W. P., and Colman, D. R. (1987). The amino acid
sequences of the myelin-associated glycoproteins: Homology to theimmunoglobulin gene superfamily. J. Cell Biol. 104: 957–965.
S
T
T
T
T
W
W
W
354 Smith-Slatas and Barbarese
Scherer, S. S., Deschenes, S. M., Xu, Y., Grinspan, J. B., Fischbeck, K.H., and Paul, D. L. (1995). Connexin32 is a myelin-related protein inthe PNS and CNS. J. Neurosci. 15: 8281–8294.
Shine, H. D., Readhead, C., Popko, B., Hood, L., and Sidman, R. L.(1992). Morphometric analysis of normal, mutant, and transgenicCNS: Correlation of myelin basic protein expression to myelino-genesis. J. Neurochem. 58: 342–349.
Snipes, G. J., Suter, U., Welcher, A. A., and Shooter, E. M. (1992).Characterization of a novel peripheral nervous system myelin pro-tein (PMP-22/SR13). J. Cell Biol. 117: 225–238.
Spray, D. C., and Dermietzel, R. (1995). X-linked dominant Charcot-Marie-Tooth disease and other potential gap-junction diseases ofthe nervous system. Trends Neurosci. 18: 256–262.
Stahl, N., Harry, J., and Popko, B. (1990). Quantitative analysis ofmyelin protein gene expression during development in the ratsciatic nerve. Mol. Brain Res. 8: 209–212.
taugaitis, S. M., Smith, P. R., and Colman, D. R. (1990). Expression ofmyelin basic protein isoforms in nonglial cells. J. Cell. Biol. 110:1719–1727.
owbin, H., Staehelin, T., and Gordon, J. (1979). Electrophoretic trans-fer of proteins from polyacrylamide gels to nitrocellulose sheets:
Procedures and some applications. Proc. Natl. Acad. Sci. USA 76:4350–4354.rapp, B. D., Andrews, S. B., Wong, A., O’Connell, M., and Griffin, J.W. (1989). Co-localization of the myelin-associated glycoproteinand the microfilament components, F-actin and spectrin, inSchwann cells of myelinated nerve fibres. J. Neurocytol. 18: 47–60.
rapp, B. D., Hauer, P., and Lemke, G. (1988). Axonal regulation ofmyelin protein mRNA levels in actively myelinating Schwann cells.J. Neurosci. 8: 3515–3521.
rapp, B. D., Itoyama, Y., Sternberger, N. H., Quarles, R. H., andWebster, H. D. (1981). Immunocytochemical localization of P0 pro-tein in Golgi complex membranes and myelin of developing ratSchwann cells. J. Cell Biol. 90: 1–6.ebster, H. deF. (1964). The relationship between Schmidt-Lanter-mann incisures and myelin segmentation during Wallerian degen-eration. Annals N.Y. Acad. Sci. 122: 29–38.elcher, A. A., Suter, U., DeLeon, M., Snipes, G. J., and Shooter, E. M.(1991). A myelin protein is encoded by the homologue of a growtharrest-specific gene. Proc. Natl. Acad. Sci. USA 88: 7195–7199.erner, R., Levine, E., Rabadan-Diehl, C., and Dahl, G. (1991). Gatingproperties of connexin32 cell–cell channels and their mutants ex-pressed in Xenopus oocytes. Proc. R. Soc. Lond. 243: 5–11.
Wiktorowicz, M., and Roach, A. (1991). Regulation of myelin basic
protein gene transcription in normal and shiverer mutant mice.Dev. Neurosci. 13: 143–150.Received August 9, 1999Revised November 1, 1999
Accepted November 24, 1999







