
Expression of novel lipocalin-like milk protein gene is developmentally-regulated during lactation in the tammar wallaby, Macropus eugenii
Josephine F. Trotta,*, Michael J. Wilson a,1,2, Russell C. Hoveyb,1,Denis C. Shawc, Kevin R. Nicholasa
aDivision of Molecular Biology and Genetics, Victorian Institute of Animal Science, 475 Mickleham Road, Attwood, VIC 3049, AustraliabMolecular and Cellular Endocrinology Section, Center for Cancer Research, NCI, NIH, Bethesda, MD 20892-1402, USA
cDivision of Biochemistry and Molecular Biology, John Curtin School of Medical Research, The Australian National University,
Canberra, ACT 2600, Australia
Received 24 August 2001; received in revised form 9 November 2001; accepted 11 December 2001
Received by A.J. van Wijnen
Abstract
We have identified a novel whey protein (late lactation protein B; LLPB) that is first secreted in the milk of the tammar wallaby around day
200 of lactation. The LLPB cDNA clone of 843 base pairs encodes a mature protein of 156 amino acids. LLPB shares 65 and 48% nucleotide
and deduced amino acid identity, respectively, with a previously identified late lactation protein A (LLPA). Both these proteins share
significant amino acid sequence homology with the lipocalin protein family. Expression of the LLPB gene is induced between days 200 and
240 of lactation, in contrast to expression of the LLPA gene, which is induced at around 145 days of lactation. Maximal expression of both
genes in mammary explants from tammars at 213 days of lactation required a combination of prolactin, insulin and hydrocortisone.
Transcripts of LLPA, LLPB and b -lactoglobulin (TBLG) were localized to the same cells by in situ hybridization. A substantial level of
alveolar maturation is required for expression of the LLP genes, unlike TBLG, which is expressed in immature alveoli. We hypothesize that
the temporal expression of the LLPB and LLPA genes may be regulated both by endocrine stimuli and factors intrinsic to the mammary gland.
q 2002 Elsevier Science B.V. All rights reserved.
Keywords: Late lactation protein; Mammary gland; Autocrine regulation; Milk composition; Marsupial
1. Introduction
Macropodid marsupials such as the tammar wallaby
(Macropus eugenii) have a unique lactation compared to
eutherian species. After a short 26.5-day gestation they
give birth to an altricial neonate and during the subsequent
lactation of 300–350 days all the major components of the
milk change significantly, presumably to provide appropri-
ate nutrition for the extensive physiological development of
the pouch young (Tyndale-Biscoe and Janssens, 1988). In
contrast, eutherians give birth to a more developed young
(Renfree, 1993), lactate for less time and do not change the
composition of their milk during lactation (Green, 1984).
Lactation in the tammar wallaby can be divided into four
phases: phases 1, 2A, 2B and 3 (Tyndale-Biscoe and Renfree,
1987; Tyndale-Biscoe and Janssens, 1988). Phase 1 encom-
passes pregnancy and lactogenesis at parturition. Phase 2
commences at lactogenesis and is divided into phase 2A
(days 0–100) when the young is permanently attached to
the teat, and phase 2B (days 100–200) when the young is
intermittently attached to the teat but permanently in the
pouch. The young is entirely dependent on milk during
phase 2 of lactation. Phase 3 (days 200–340) commences
when the young first exits the pouch and consumes herbage
in addition to milk. During the last 100–150 days of lactation
the young develops the metabolic capacity to become nutri-
tionally independent of milk (Tyndale-Biscoe and Renfree,
1987).
Gene 283 (2002) 287–297
0378-1119/02/$ - see front matter q 2002 Elsevier Science B.V. All rights reserved.
PII: S0378-1119(01)00883-6
www.elsevier.com/locate/gene
Abbreviations: aa, amino acid(s);a-lac,a-lactalbumin; BLG, b-lactoglo-
bulin; bp, base pair(s); cDNA, DNA complementary to RNA; cpm, counts
per minute; DIG, digoxigenin; ELP, early lactation protein; F, hydrocorti-
sone; HPLC, high-performance liquid chromatography; I, insulin; LLP, late
lactation protein; PRL, prolactin; SA, serum albumin; SDS, sodium dodecyl
sulfate; SDS–PAGE, SDS–polyacrylamide gel electrophoresis; SEM, stan-
dard error of the mean; SSC, 0.15 M NaCl/0.015 M Na3citrate, pH 7.6;
VEGP, von Ebner’s gland protein; WAP, whey acidic protein
* Corresponding author. Present address: Room 204, Terrill Hall, Depart-
ment of Animal Science, University of Vermont, Burlington, VT 05405,
USA. Tel.: 11-802-656-5894; fax: 11-802-656-8196.
E-mail address: [email protected] (J.F. Trott).1 These authors contributed equally to this work.2 Present address: Department of Pathology, University of Cambridge,
Tennis Court Road, Cambridge CB2 1QP, UK.

The concentration and composition of milk carbohydrates,
lipids and proteins changes significantly during the course of
the lactation cycle of the tammar wallaby (Messer and Green,
1979; Green, 1984; Green and Merchant, 1988), concurrent
with increases in both the size (Findlay, 1982) and weight
(Stewart, 1984) of the mammary gland. Specific changes in
the expression of individual milk protein genes (Bird et al.,
1994) lead to a large increase in milk protein content at the
onset of phase 3 of lactation (Green et al., 1980; Nicholas,
1988a). A novel whey protein, late lactation protein A
(LLPA), is first detected in the milk during the later stages
of lactation in a large group of marsupials including the
tammar wallaby (Nicholas et al., 1987; Collet et al., 1989),
the red kangaroo (Macropus rufus; McKenzie et al., 1983),
grey kangaroo (Macropus giganteus; McKenzie et al., 1983),
the quokka (Setonix brachyurus; Beg and Shaw, 1994) and
the brushtail possum (Trichosurus vulpecula; Piotte et al.,
1998). LLPA is first secreted in the milk of the tammar
wallaby around day 160 of lactation and increases in concen-
tration to constitute approximately 25% of the total milk
protein at around day 250 of lactation (Nicholas et al.,
1987). The function of this protein remains to be determined,
but amino acid (aa) sequence analysis suggests that, like
tammar b-lactoglobulin (TBLG), it is a member of the lipo-
calin group of proteins (Collet and Joseph, 1993). The
mechanisms regulating the induction of LLPA gene expres-
sion at the onset of phase 3 of lactation are unknown.
However, the tammar wallaby is capable of concurrent asyn-
chronous lactation, whereby two young of different ages
suckle from adjacent mammary glands secreting milk of
different composition (Lincoln and Renfree, 1981; Nicholas,
1988a). This suggests that in addition to an endocrine control
of milk composition ( Nicholas, 1988b), intramammary or
autocrine factors regulate the synthesis of specific milk
constituents in the tammar wallaby.
Analysis of whey proteins by SDS–PAGE reveals the
secretion of a second milk protein specifically during late
lactation in the tammar wallaby (Nicholas et al., 1987). This
protein was named late lactation protein B (LLPB) but its
molecular properties have remained unidentified. We report
here the cloning and characterization of the cDNA for LLPB
and the developmentally- and hormonally-regulated expres-
sion of the gene during lactation. In addition, the spatial
distribution of LLP gene expression has been examined.
Our results suggest that the temporal expression of LLP
genes may be regulated by factors intrinsic to the mammary
gland, responding either to changes in the sucking pattern of
the young as it relinquishes the teat, or maternally-regulated
mammary gland development.
2. Materials and methods
2.1. Materials
Ovine prolactin (NIDDK-oPRL-20 AFP10677C) was a
gift from the National Hormone and Pituitary Program,
NIDDK, NIH. Bovine insulin and hydrocortisone were
purchased from Sigma-Aldrich (St Louis, MO). Sodium
methohexitone was purchased from Eli Lilly Australia
Pty. Ltd. (West Ryde, Australia), oxytocin from Heriot
Agvet Pty. Ltd. (Rowville, Australia) and [a-32P]dCTP
from Bresatec (Adelaide, Australia).
2.2. Animals
A colony of tammar wallabies was kept in open grassy
yards at the Victorian Institute of Animal Science, Attwood,
and provided feed and water ad libitum. All animal experi-
mentation was performed according to the Victorian Insti-
tute of Animal Science Animal Experimentation Ethics.
2.3. SDS–PAGE analysis of milk proteins
Milk was collected as described previously (Green et al.,
1980) with a few modifications. Pouch young were gently
removed from the teat 2–3 h prior to milking to allow milk
accumulation. Ectothermic young (less than approximately
180 days-old) were kept warm (32 8C) and moist. Home-
othermic young (greater than 180 days-old) were kept in
calico sacks. Mothers were anaesthetized with an i.v. injec-
tion of sodium methohexitone (8 mg/kg body weight) into
the lateral tail vein, followed by an intramuscular injection
of 1 IU oxytocin. Milk was removed by gentle massage of
the gland and teat, collected in 1.5-ml tubes and stored at
220 8C. After the mothers recovered, the pouch young were
transferred back into the pouch. Whole milk samples were
centrifuged at 3000 £ g for 15 min at 4 8C, and the fat layer
removed. The skim milk was centrifuged at 50; 000 £ g for
30 min at 12 8C and the casein pellet discarded. The super-
natant was diluted and whey proteins analyzed by 20%
SDS–PAGE.
2.4. Protein isolation and amino acid sequencing
Whey proteins were fractionated by high performance
liquid chromatography as previously described (Simpson
et al., 2000) and fractions analyzed by SDS–PAGE. If a
positive identification was necessary, up to 20 N-terminal
aa were sequenced using an ABI 477A pulsed-liquid phase
sequencer (Applied Biosystems). If the N-terminus of a
protein was blocked and could not be sequenced, the protein
was subjected to tryptic digestion followed by peptide
sequencing (Shaw et al., 1993).
2.5. Oligonucleotide design and synthesis
LLPB specific oligonucleotides with sequences deduced
from aa sequence data were designed to regions of lowest
ambiguity and inosine was substituted in positions of high
ambiguity to reduce the number of possible oligonucleo-
tides. Two pools of oligonucleotides (5 0-RAAYTCNCT-
RTCNGTIAGDATIGC-3 0, 5 0-RAAYTCNGARTCNGTI-
AGDATIGC-3 0) in the sense orientation were designed to
J.F. Trott et al. / Gene 283 (2002) 287–297288

screen a cDNA library. PCR primers to amplify phage lgt11
cDNA inserts were designed from the sequence of the phage
lgt11 arms (gtllF 5 0-GCGACGACTCCTGGAGCCCG-3 0,
gtllR 5 0-TGGTAGCGACCGGCGCTCAG-3 0).
2.6. Screening of the cDNA library
The cDNA library was constructed in lgt11 using total
RNA using Trizol (Life Technologies Inc.) isolated from
tammar wallaby mammary glands at day 270 of lactation.
Degenerate LLPB oligonucleotide probes were labeled with
[a-32P]dCTP using terminal deoxy transferase and hybri-
dized overnight to plaque lifts in Church buffer (Sambrook
and Russell, 2001) at 40 8C. Membranes were washed to a
stringency of 2£ SSC, 0.1% SDS at 45 8C. LLPB cDNA was
isolated, cloned into pGEM-7fz 2 (Promega) and
sequenced. The library was re-screened with a partial
LLPB cDNA clone. Forward and reverse primers to the
phage lgt11 were used for PCR to select clones containing
at least 900 bp of cDNA. The PCR reaction mix included 5
ml of phage, 2.5 units Taq DNA Polymerase, 40 pmol gt11R
and gtllF, 2.5 mM MgCl2, 80 mM dNTP, in 1£ PCR buffer
(10 mM Tris–HCl (pH 9), 50 mM KCl, 0.01% gelatin, 0.1%
Triton X-100). Reactions were amplified using a hot start of
92 8C for 5 min, then cycled 30 times at 92 8C for 30 s, 54.7
8C for 1 min, 72 8C for 1 min, followed by 72 8C for 10 min.
The PCR products of two reactions were cloned into pGEM-
T (Promega), and sequenced using an ABI 373A automated
DNA sequencer (Applied Biosystems; Griffith University,
Qld, Australia). Phage DNA for these clones was isolated by
liquid lysis (Sambrook and Russell, 2001), digested with
EcoRI and the cDNA inserts were subsequently cloned
into pGEM-7f-. Full-length sequence in both directions,
obtained using internal sequencing primers, was subjected
to sequence analysis using DNASIS and SeqEd 1.0, and
percentage identities were calculated from the alignment
created using the ClustalW website (http://www2.ebi.a-
c.uk/clustalw).
2.7. Northern and slot blot analysis
Total RNA was extracted using Trizol (Life Technologies
Inc.) from either the mammary glands of tammar wallabies
at various stages of lactation or from cultured mammary
explants. Total RNA (10 mg) was fractionated through an
18% formaldehyde, 1.4% agarose gel and transferred to
Zeta-Probe GT membrane (Bio-Rad) using 50 mM NaOH.
For slot blot analysis total RNA (1 mg) was diluted in 10
mM NaOH, 1 mM EDTA and transferred to Zeta-Probe GT
membrane.
All membranes were hybridized in Church buffer
(Sambrook and Russell, 2001) containing 50 mg/ml herring
sperm DNA at 65 8C with LLPB cDNA that had been
labeled by random priming with [a-32P]dCTP. Membranes
were then washed to high stringency (65 8C; 0:1£ SSC,
0.1% SDS) and exposed to X-ray film at 80 8C.
Following autoradiography of the slot blots, the
membrane containing each slot of RNA was isolated, placed
in a scintillation vial with 3 ml organic counting scintillant
(Amersham), and counted in a Wallac 1410 liquid scintilla-
tion counter (Pharmacia). The counts per minute (cpm) for
each slot were corrected for background radioactivity on the
filter.
2.8. Explant culture
Mammary gland from tammars at day 213 of lactation
were cut into approximately 1–2-mg explants, planted onto
lens paper siliconized with Aquasil (Pierce) and floated on
M199 media (with Earle’s salts). Media was changed daily.
Explants were cultured in insulin (1 mg/ml) for 4 days,
followed by 4 days in media supplemented with either insu-
lin, insulin and hydrocortisone (50 ng/ml) or insulin, hydro-
cortisone and prolactin (200 ng/ml). Explants were collected
and pooled prior to culture (T0) and after 4 or 8 days of
culture.
2.9. In situ hybridization
Mammary tissue from a tammar wallaby euthanized at
day 218 of lactation was fixed in 4% paraformaldehyde,
dehydrated and embedded in paraffin. Riboprobes were
transcribed from LLPB, LLPA and TBLG cDNA cloned
into pGEM-7f1, pGEM-3f and pPCRScript, respectively.
Antisense cRNA probes were synthesized from plasmid
linearized with HindIII, HindIII and EcoRV, respectively.
Sense cRNA probes for LLPB, LLPA and TBLG were
synthesized from plasmid linearized with XhoI, PvuII and
NotI, respectively. Riboprobes were generated using T7
(antisense for LLPB and LLPA, sense for TBLG), SP6
(sense for LLPB and LLPA) or T3 (antisense for TBLG)
polymerase (Maxiscript kit, Ambion, Austin, TX) in the
presence of DIG-11-dUTP (RNA labeling kit, Boehringer
Mannheim). Probes were subsequently reduced to approxi-
mately 100–300 nucleotides in length by limited alkaline
hydrolysis (Angerer and Angerer, 1992). In situ hybridiza-
tion with LLPB, LLPA and TBLG cRNA was performed as
previously described (Seagroves et al., 2000). Hybridized
cRNA was localized by incubating sections with an anti-
DIG polyclonal antibody (1:125, Boehringer Mannheim)
followed by chromogenic detection with NBT/BCIP (Boeh-
ringer Mannheim). Sections were counterstained with
Nuclear Fast red.
3. Results
3.1. Identification and cloning of LLPB cDNA
Separation of whey proteins in milk from phases 2B and 3
of lactation by SDS–PAGE showed that LLPA was first
secreted in abundance after 168 days of lactation and a
second protein, named LLPB, was secreted in abundance
only after 198 days of lactation (Fig. 1). In contrast, a-
J.F. Trott et al. / Gene 283 (2002) 287–297 289
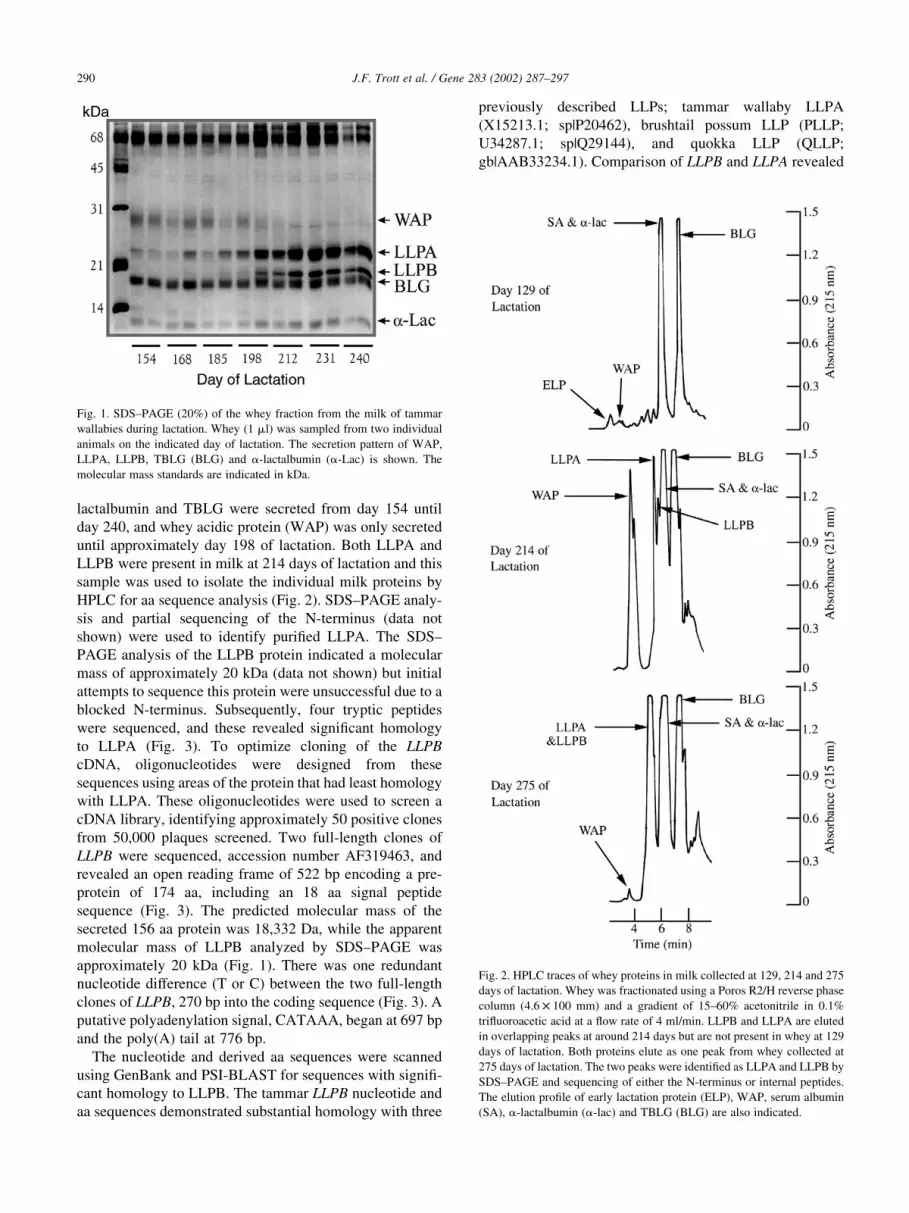
lactalbumin and TBLG were secreted from day 154 until
day 240, and whey acidic protein (WAP) was only secreted
until approximately day 198 of lactation. Both LLPA and
LLPB were present in milk at 214 days of lactation and this
sample was used to isolate the individual milk proteins by
HPLC for aa sequence analysis (Fig. 2). SDS–PAGE analy-
sis and partial sequencing of the N-terminus (data not
shown) were used to identify purified LLPA. The SDS–
PAGE analysis of the LLPB protein indicated a molecular
mass of approximately 20 kDa (data not shown) but initial
attempts to sequence this protein were unsuccessful due to a
blocked N-terminus. Subsequently, four tryptic peptides
were sequenced, and these revealed significant homology
to LLPA (Fig. 3). To optimize cloning of the LLPB
cDNA, oligonucleotides were designed from these
sequences using areas of the protein that had least homology
with LLPA. These oligonucleotides were used to screen a
cDNA library, identifying approximately 50 positive clones
from 50,000 plaques screened. Two full-length clones of
LLPB were sequenced, accession number AF319463, and
revealed an open reading frame of 522 bp encoding a pre-
protein of 174 aa, including an 18 aa signal peptide
sequence (Fig. 3). The predicted molecular mass of the
secreted 156 aa protein was 18,332 Da, while the apparent
molecular mass of LLPB analyzed by SDS–PAGE was
approximately 20 kDa (Fig. 1). There was one redundant
nucleotide difference (T or C) between the two full-length
clones of LLPB, 270 bp into the coding sequence (Fig. 3). A
putative polyadenylation signal, CATAAA, began at 697 bp
and the poly(A) tail at 776 bp.
The nucleotide and derived aa sequences were scanned
using GenBank and PSI-BLAST for sequences with signifi-
cant homology to LLPB. The tammar LLPB nucleotide and
aa sequences demonstrated substantial homology with three
previously described LLPs; tammar wallaby LLPA
(X15213.1; sp|P20462), brushtail possum LLP (PLLP;
U34287.1; sp|Q29144), and quokka LLP (QLLP;
gb|AAB33234.1). Comparison of LLPB and LLPA revealed
J.F. Trott et al. / Gene 283 (2002) 287–297290
Fig. 1. SDS–PAGE (20%) of the whey fraction from the milk of tammar
wallabies during lactation. Whey (1 ml) was sampled from two individual
animals on the indicated day of lactation. The secretion pattern of WAP,
LLPA, LLPB, TBLG (BLG) and a-lactalbumin (a-Lac) is shown. The
molecular mass standards are indicated in kDa.
Fig. 2. HPLC traces of whey proteins in milk collected at 129, 214 and 275
days of lactation. Whey was fractionated using a Poros R2/H reverse phase
column (4:6 £ 100 mm) and a gradient of 15–60% acetonitrile in 0.1%
trifluoroacetic acid at a flow rate of 4 ml/min. LLPB and LLPA are eluted
in overlapping peaks at around 214 days but are not present in whey at 129
days of lactation. Both proteins elute as one peak from whey collected at
275 days of lactation. The two peaks were identified as LLPA and LLPB by
SDS–PAGE and sequencing of either the N-terminus or internal peptides.
The elution profile of early lactation protein (ELP), WAP, serum albumin
(SA), a-lactalbumin (a-lac) and TBLG (BLG) are also indicated.

65 and 48% homology at the nucleotide and derived aa (Fig.
4) levels, respectively. LLPB shares aa sequence similarity
of 47 and 45% with that for PLLP and QLLP, respectively
(Fig. 4), while LLPA has sequence similarity of 74 and 87%
with PLLP and QLLP, respectively (Fig. 4). The aa
sequence of LLPB has significant sequence identity with
human (41%; sp|P31025) and porcine (39%; sp|P53715)
von Ebner’s gland proteins (VEGP; Fig. 4) in addition to
the dog major allergen F1 protein (41%; sp|O18873) and
human oderant-binding protein 2B (39%; ref|NP_055396).
J.F. Trott et al. / Gene 283 (2002) 287–297 291
Fig. 3. Nucleic acid sequence of LLPB cDNA and the derived aa sequence. The 18 aa signal peptide is shaded, and the sequenced tryptic peptides are
underlined. The LLPB nucleotide sequence is numbered in normal text, with the inferred aa sequence numbered in bold. Amino acid residues 55 (F), 95 (Y) and
128 (M) were determined by peptide sequence to be different from the derived aa residues of M, W and F, respectively. The putative polyadenylation signal is
boxed. A redundant base pair substitution of T for C was found at 399 bp.
Fig. 4. Alignment of aa sequences of LLPB (TLLPB), LLPA (TLLPA), quokka LLPA (QLLP), brushtail possum LLP (PLLP), pig tear VEGP (PVEGP),
human tear VEGP (HVEGP) and TBLG. Amino acid residues that are identical in the six proteins are shaded. Amino acid residues that are identical in at least
five of the six proteins are in bold. Asterisks indicate the aa residues common to all LLPs. The lipocalin motifs 1, 2 and 3 are numbered and boxed, and the
critical residues are in italics and bold. Motif 1 is found in all lipocalins. Motif 2 is found only in the kernel lipocalin TBLG (TDYxxY). Motif 3 is found both in
PVEGP and HVEGP as well as TBLG, but a P substitutes for the R in the LLPs.

3.2. Developmental regulation of LLPB and LLPA gene
expression
Northern blot analysis of RNA from mammary tissue
using LLPB and LLPA cDNA probes revealed specific
hybridization to transcripts of approximately 950 and 900
nucleotides respectively (Fig. 5A). The LLPB transcript was
detectable in total RNA from mammary glands after day
200 of lactation by both Northern and slot blot (Fig. 5).
Extended exposure of Northern blots failed to detect any
LLPB mRNA in tissue collected at, or before 200 days of
lactation (data not shown). LLPA mRNA was easily
detected by Northern blot at about 170 days of lactation
(Fig. 5A), although a transcript was evident at approxi-
mately 145 days post-partum following extended autoradio-
graphic exposure (data not shown). Maximum accumulation
of mRNA for LLPA was at approximately 245 days of lacta-
tion (Fig. 5B). The time when LLPB showed maximal accu-
mulation could not be established because samples were
collected only until 270 days of lactation.
Northern analysis of RNA from tammar mammary tissue
at day 213 of lactation revealed high levels of expression of
the LLPA, LLPB and TBLG genes (Fig. 5C). When explants
of mammary tissue were cultured for 4 days in media
supplemented with insulin, the expression of all genes was
significantly decreased. After 4 additional days incubation
in the presence of either insulin or insulin and hydrocorti-
sone, the expression levels declined even further. In
contrast, when medium in the latter 4-day incubation period
was supplemented with prolactin in addition to insulin and
J.F. Trott et al. / Gene 283 (2002) 287–297292
Fig. 5. Developmental and hormonal regulation of the LLPB and LLPA genes during lactation. (A) Northern blot of total mammary gland RNA (10 mg) from
lactating tammars divided into six groups of three animals; group 1 (days 95, 96 and 106); group 2 (days 140, 144 and 144); group 3 (days 169, 172 and 174);
group 4 (days 198, 202 and 202); group 5 (days 231, 242 and 244); group 6 (days 275, 276 and 276). The average day of lactation for each of the six groups is
notated above the data. The Northern blots were probed sequentially with cDNA for LLPB and LLPA. LLPB mRNA was not detected in RNA collected from
mammary glands at or prior to 200 days of lactation, even after extended exposure of blots (data not shown). LLPA mRNA could be detected in RNA collected
from mammary glands at 145 days of lactation following extended exposure of Northern blots (data not shown). TBLG mRNA is expressed throughout
lactation, increasing in concentration in phase 3 (Simpson et al., 2000) and confirms the quality of the RNA (republished with permission of the Journal of
Biological Chemistry from Simpson et al., 2000; permission conveyed through Copyright Clearance Center, Inc.). (B) Quantification of both LLPB and LLPA
mRNA by slot blot analysis of total RNA (1 mg) from the mammary gland at the six grouped time points of lactation (using the same RNA as analyzed by the
Northern blot). Each value represents the mean ^ SEM for three animals. X, LLPB; B, LLPA. (C) Northern blot of mammary gland explant total RNA (10 mg)
from tammars at day 213 of lactation. Explants were cultured for 0, 4 or 8 days. Total RNA is shown for a loading comparison. T0, Time zero; I4, insulin (I)
treatment for 4 days; I4-IFPRL4, I for 4 days followed by 4 days treatment with I, hydrocortisone (F) and prolactin (PRL).

hydrocortisone, expression of all three genes was stimulated
or maintained compared to the expression after 4 days in
insulin alone.
3.3. Spatial expression of LLPB, LLPA and TBLG mRNA
Hybridization of LLPB and LLPA cRNA to serial sections
of day 218 mammary gland demonstrated identical distribu-
tion of mRNA expression for both genes within the alveolar
epithelium (Fig. 6A,B). Since all cells within an alveolus
expressed both genes (Fig. 6A,B), we conclude that the
same cells express both LLP genes. We also observed differ-
ences in the level of LLPA and LLPB mRNA expression
between alveoli at day 218 of lactation. Specifically,
J.F. Trott et al. / Gene 283 (2002) 287–297 293
Fig. 6. In situ hybridization of LLPB and LLPA cRNA to serial sections of tammar wallaby mammary gland at day 218 of lactation. (A–D) Sections were
hybridized with LLPB and LLPA antisense DIG-labeled riboprobes. The dark precipitate localizes probe hybridized within mammary epithelial cells. (A,B)
Distended fully-functional alveoli in sections of mammary gland. (A) Section hybridized with LLPB antisense riboprobe. (B) Serial section to (A) hybridized
with LLPA antisense riboprobe. Comparison with (A) shows that all cells within a given alveolus expressed both LLPB and LLPA. The same alveolus is
indicated by an asterisk. (C,D) A mixture of distended fully-functional alveoli and functionally-immature alveoli in sections of mammary gland. (C) Section
hybridized with LLPB antisense riboprobe. (D) Serial section to (C) hybridized with LLPA antisense riboprobe. Comparison with (C) shows that there is
heterogeneous expression of both genes and that only fully differentiated secretory alveoli (white arrows) express both LLPB and LLPA. Functionally-
immature alveoli (black arrowheads) are not expressing the two LLP genes. (E,F) Sections hybridized with (E) LLPB and (F) LLPA sense DIG-labeled
riboprobes, showing no non-specific hybridization. Scale bar in F: 25 mm for panels A,B; 100 mm for panels C–F.

LLPA and LLPB mRNA was detected in fully-functional
alveoli with a distended lumen lined by cells that were
actively secreting lipid, whereas neither LLPA or LLPB
was expressed in functionally-immature alveoli. These
immature alveoli were frequently interspersed among the
distended, fully-functional alveoli. Hybridization of LLPB
and TBLG cRNA to serial sections of day 218 mammary
gland showed that TBLG was expressed by the same cells as
LLPB (Fig. 7A,B). Transcripts of LLPB were undetectable
in the same immature alveoli that were expressing TBLG
mRNA (Fig. 7C,D). It is noteworthy that the cellular expres-
sion of LLPB was sometimes heterogeneous within alveoli
J.F. Trott et al. / Gene 283 (2002) 287–297294
Fig. 7. In situ hybridization of LLPB and TBLG cRNA to serial sections of tammar wallaby mammary gland at day 218 of lactation. (A–E) Sections of
mammary gland were hybridized with LLPB and TBLG antisense DIG-labeled riboprobes. The dark precipitate localizes probe hybridized within mammary
epithelial cells. (A) Section of mammary gland hybridized with LLPB antisense riboprobe. (B) Serial section to (A) hybridized with TBLG antisense riboprobe.
Comparison with (A) shows that all cells within a given alveolus expressed both LLPB and TBLG. The same alveolus is indicated by an asterisk. (C) Section of
mammary gland hybridized with LLPB antisense riboprobe. (D) Serial section to (C) hybridized with TBLG antisense riboprobe. Comparison with (C) shows
that functionally-immature alveoli (black arrowheads) only express TBLG, whereas fully-differentiated secretory alveoli (white arrows) are expressing both
LLPB and TBLG. (E) Section of mammary gland hybridized with LLPB antisense riboprobe showing adjacent cells within an alveolus expressing (arrow) or not
expressing (arrowhead) LLPB mRNA. (F) Section of mammary gland hybridized with TBLG sense DIG-labeled riboprobe, showing no non-specific hybri-
dization. Scale bar in F: 50 mm for panels A,B,F; 100 mm for panels C,D; 25 mm for panel E.

(Fig. 7E), a phenomenon that was not observed for TBLG or
LLPA. The sense strand riboprobes did not hybridize to
comparable sections.
4. Discussion
We have identified LLPB as a novel whey protein that has
not yet been characterized in any other species. This protein
shares significant identity with tammar LLPA (Nicholas et
al., 1987; Collet et al., 1989), while the LLP in two other
marsupial species, the quokka (Beg and Shaw, 1994) and the
brushtail possum (Piotte et al., 1998), display greater aa
identity to LLPA. This suggests that these LLPs are direct
orthologues of LLPA and these marsupials may also posses
an additional LLP locus. It is worth noting that a report by
Woodlee et al. (1993) shows that LLPA may exist as a two-
allele polymorphism in the tammar wallaby. These authors
identified the polymorphisms by polyacrylamide gel elec-
trophoresis of whey on native gels and N-terminal sequence
analysis. They labeled the polymorphic forms of this protein
as LLPa and LLPb, but the N-terminal sequence only corre-
sponds to that for LLPA. In addition, the N-terminus of
native LLPB is blocked and cannot be sequenced. Thus,
the notation used by these authors to describe these poly-
morphisms of LLPA should not be confused with our use of
the abbreviations LLPA and LLPB.
The LLPs share significant aa sequence homology with
the lipocalins (Collet and Joseph, 1993), a family of proteins
which function broadly as extracellular carriers of small
hydrophobic molecules (Flower, 1996). The lipocalins
have been assigned into two sets; a core set of closely
related ‘kernel’ lipocalins and the more divergent ‘outlier’
lipocalins (Flower, 1996). The kernel lipocalins are charac-
terized by the presence of three conserved motifs, but the
outlier lipocalins share only the first motif, which is diag-
nostic of family membership. Motif 1 is GXWY/W/F and is
not strictly shared by the LLPs since they contain a Y
instead of a W in the third aa position. In addition, neither
LLPB nor LLPA possesses motifs 2 or 3, therefore classify-
ing both these proteins as putative outlier lipocalins.
Interestingly, the LLPs have the highest level of homology
with human and tear VEGP, dog major allergen F1 protein
and human oderant binding protein 2B. Human oderant bind-
ing protein 2B is a putative oderant carrier and strongly
expressed by the prostate and the mammary gland (Lacazette
et al., 2000). Dog allergen F1 is a salivary protein, produced
by tongue epithelial cells (Konieczny et al., 1997). VEGP is
secreted into saliva by the Von Ebner’s gland, a small acinar
salivary gland of the tongue (Flower, 1996), and also into tear
fluid by the lachrymal gland (Redl et al., 1992). This is of
interest in terms of the evolution of the mammary glands
from sebaceous and apocrine glands (Renfree, 1993).
Homology to these members of the lipocalin superfamily
may give some insight into the function of LLPs. For exam-
ple, tear VEGP binds palmitic and oleic acids (Flower, 1996)
and another member of the lipocalin family, bovine BLG
(BBLG; Flower, 1996), also binds palmitic acid (Puyol et
al., 1991) as well as retinol in the intestine of the young
(Said et al., 1989). Fatty acid ligands, including palmitic
acid, bind BBLG inside the central conical calyx that is a
structural characteristic of the lipocalin family (Wu et al.,
1999). In bovine milk, fatty acids are physiologically
bound to BBLG (Diaz de Villegas et al., 1987; Perez et al.,
1991), and hence BBLG could enhance the uptake of palmitic
acid from milk (Burczynski et al., 1990). The BBLG could
also enhance the activity of pregastric lipase (Perez et al.,
1991), suggesting a role for this protein during the digestion
of milk fat by the young (Perez and Calvo, 1995). It is note-
worthy that there is a large increase in the lipid content of
milk at around day 200 of lactation in the tammar wallaby
(Nicholas, 1988a) with oleic and palmitic acid making up
70% of the total lipids (Green and Merchant, 1988). This
increase in lipids corresponds with the induction of LLPB
and the increased synthesis of LLPA and TBLG. Although
there is no sequence homology between the LLPs and fatty
acid binding proteins (data not shown) a role for LLPs in lipid
transport and absorption in the tammar wallaby deserves
further investigation. In addition, it has also been proposed
that tear VEGP may have an antibacterial function (Flower,
1996). Milk also has anti-microbial properties (Blackburn et
al., 1989), and structural and functional links between milk
components and anti-microbial secretion of other integu-
ment-derived glands have been noted (Hayssen and Black-
burn, 1985).
The unique pattern of induction for the LLPB and LLPA
genes has raised the question of whether a new population of
cells arises to express each of these genes (Tyndale-Biscoe
and Renfree, 1987), or whether the same cells express both
genes. The mammary gland expresses LLPA in the same
cells that are expressing TBLG and a -lactalbumin after
200 days of lactation (Joseph and Collet, 1994). We have
extended this observation and demonstrated the expression
of the LLPB, LLPA and TBLG genes in the same cells at day
218 of lactation using in situ hybridization with gene-speci-
fic cRNA probes. Taken collectively, these data indicate
that the same epithelial cells most likely express the full
complement of milk protein genes in phase 3 of lactation,
and the induction of the LLPA and LLPB genes cannot be
explained by the growth of a new population of mammary
epithelial cells.
The spatial distribution of LLPB and LLPA mRNA was
heterogeneous within the day 218 lactating mammary gland.
In particular, morphologically-underdeveloped and func-
tionally-immature alveoli did not express either LLPB or
LLPA, whereas distended fully-functional alveoli expressed
both genes. The functionally-immature alveoli were capable
of expressing milk protein mRNAs since they expressed
TBLG mRNA. These alveoli were also morphologically
similar to those found in phase 2 mammary glands (Findlay,
1982) which cannot be hormonally-induced to express
LLPB or LLPA (Nicholas, 1988b). It is noteworthy that
J.F. Trott et al. / Gene 283 (2002) 287–297 295

the LLPA, LLPB and TBLG genes all have similar hormonal
requirements for expression in mammary explant culture
using tissue from this stage of lactation, suggesting it is
unlikely that endocrine influences regulate their differential
expression during early phase 3. Thus, local regulation of
gene expression probably regulates the temporal induction
of LLPA and LLPB gene expression. The local regulation of
LLPB gene expression is indicated by our finding that only
certain cells within some alveoli expressed LLPB mRNA.
This phenomenon likely contributes to the temporal induc-
tion of LLPB synthesis that occurs at around 200 days of
lactation. Furthermore, the LLPB and LLPA genes are only
transcribed in distended, fully-functional alveoli, suggesting
that a certain level of structural development of the gland is
required for their expression. One of the factors that controls
alveolar structure and cell morphology is the extracellular
matrix (Barcellos-Hoff et al., 1989), which is directly
involved in determining whether particular milk protein
genes are expressed by mammary epithelial cells (Schmid-
hauser et al., 1995). It is possible that the extracellular
matrix regulates the extent of alveolar development,
which in turn regulates expression of the LLP genes.
The large increase in size (Findlay, 1982) and weight
(Stewart, 1984) of the gland during the transition from
phase 2B to phase 3 of lactation is indicative of the changing
development of the gland, as is the presence of both func-
tionally-mature and -immature alveoli. The increase in size
is partly due to increases in the cytoplasm:nucleus ratio and
alveolar diameter (Findlay, 1982; Joseph and Collet, 1994).
The other contributing factor is an increase in the number of
cells per gland until 210–240 days of lactation (Stewart,
1984). Although mitotic figures in sections of the tammar
mammary gland are minimal after day 40 of lactation
(Findlay, 1982), the level of binucleated epithelial cells
increases after day 200 of lactation, indicating DNA repli-
cation (Joseph and Collet, 1994). The level of expression of
the LLPB and LLPA genes increases dramatically from day
200 to day 240 of lactation and may be partially mediated by
an increase in the number of cells per gland around this time
(Stewart, 1984). However, our present results indicate that
this increased synthesis of LLPA and -B mRNA and protein
is due to increasing numbers of alveoli assuming full func-
tionality (Trott and Nicholas, personal observation), thereby
increasing the total output of these proteins by the mammary
gland between days 200 and 240 of lactation.
The endocrine and putative local mammary factors that
regulate the induction of LLPB and LLPA gene expression at
the onset of phase 3 of lactation are not yet clear. In the
current study we found the LLPB, LLPA and TBLG genes all
responded to the same hormonal stimulation in cultured
explants from phase 3 mammary gland. It is well known
that the tammar wallaby is capable of concurrent asynchro-
nous lactation, whereby two young of different ages suck
adjacent mammary glands that are at different stages of
development, and secrete milk of different composition
(Lincoln and Renfree, 1981; Nicholas, 1988a). This finding
suggests that in addition to endocrine control, intramam-
mary or autocrine factors regulate the synthesis of the
LLP genes. Around day 200 of lactation the young begins
leaving the pouch, supplementing a milk diet with herbage
(Tyndale-Biscoe and Janssens, 1988). The sucking pattern
of the young is presumed to reduce in frequency, coincident
with changes in gene expression and the most rapid growth
of the mammary gland (Stewart, 1984). The change in suck-
ing pattern may cue these events, or they may be the result
of maternally-regulated mammary gland development.
Previously we have reported the isolation and characteriza-
tion of a factor in tammar milk that regulates the sucking-
dependent secretion of milk (Hendry et al., 1998). However,
it remains to be determined whether the sucking frequency
of the pouch young influences the control of milk composi-
tion.
Acknowledgements
J.F.T. was supported by a Nancy Millis Research Award
from the Department of Natural Resources and the Environ-
ment, Victoria, Australia. We thank Dr. Tracie Webster for
assistance with graphics.
References
Angerer, L.M., Angerer, R.C., 1992. In situ hybridization to cellular RNA
with radiolabelled RNA probes. In Situ Hybridization: A Practical
Approach, Oxford University Press, Oxford, UK, pp. 15–32.
Barcellos-Hoff, M.H., Aggeler, J., Ram, T.G., Bissell, M.J., 1989. Func-
tional differentiation and alveolar morphogenesis of primary mammary
cultures on reconstituted basement membrane. Development 105, 223–
235.
Beg, O.U., Shaw, D.C., 1994. The complete primary structure of late lacta-
tion protein from quokka (Setonix brachyurus). J. Protein Chem. 13,
513–516.
Bird, P., Hendry, K.A.K., Shaw, D., Wilde, C.J., Nicholas, K.R., 1994.
Progressive changes in milk protein gene expression and prolactin bind-
ing during lactation in the tammar wallaby (Macropus eugenii). J. Mol.
Endocrinol. 13, 117–125.
Blackburn, D.G., Hayssen, V., Murphy, C.J., 1989. The origins of lactation
and the evolution of milk: A review with new hypotheses. Mammal.
Rev. 19, 1–26.
Burczynski, F.J., Moran, J.B., Cai, Z.S., Forker, E.L., 1990. b-Lactoglobu-
lin enhances the uptake of free palmitate by hepatocyte monolayers: the
relative importance of diffusion and facilitated dissociation. Can. J.
Physiol. Pharmacol. 68, 201.
Collet, C., Joseph, R., 1993. A novel member of the lipocalin superfamily:
tammar wallaby late-lactation protein. Biochim. Biophys. Acta 1167,
219–222.
Collet, C., Joseph, R., Nicholas, K.R., 1989. Molecular cloning and char-
acterization of a novel marsupial milk protein gene. Biochem. Biophys.
Res. Commun. 164, 1380–1383.
Diaz de Villegas, M.C., Oria, R., Sala, F.J., Calvo, M., 1987. Lipid binding
by b-lactoglobulin of cow milk. Milchwissenschaft 42, 357.
Findlay, L., 1982. The mammary glands of the tammar wallaby (Macropus
eugenii) during pregnancy and lactation. J. Reprod. Fertil. 65, 59–66.
Flower, D.R., 1996. The lipocalin protein family: structure and function.
Biochem. J. 318, 1–14.
J.F. Trott et al. / Gene 283 (2002) 287–297296

Green, B., 1984. Composition of milk and energetics of growth in marsu-
pials. Symp. Zool. Soc. Lond. 51, 369–387.
Green, B., Merchant, J.C., 1988. The composition of marsupial milk. In:
Tyndale-Biscoe, C.H., Janssens, P.A. (Eds.). The Developing Marsu-
pial: Models for Biomedical Research, Springer, Heidelberg, pp. 41–54.
Green, B., Newgrain, K., Merchant, J., 1980. Changes in milk composition
during lactation in the tammar wallaby (Macropus eugenii). Aust. J.
Biol. Sci. 33, 35–42.
Hayssen, V., Blackburn, D.G., 1985. a-lactalbumin and the origins of lacta-
tion. Evolution 39, 1147–1149.
Hendry, K.A.K., Simpson, K.J., Nicholas, K.R., Wilde, C.J., 1998. Auto-
crine inhibition of milk secretion in the lactating tammar wallaby
(Macropus eugenii). J. Mol. Endocrinol. 21, 169–177.
Joseph, R., Collet, C., 1994. Double staining in situ study of mRNAs
encoding milk proteins in the mammary gland of the tammar wallaby
(Macropus eugenii). J. Reprod. Fertil. 101, 241–246.
Konieczny, A., Morgenstern, J.P., Bizinkauskas, C.B., Lilley, C.H., Brauer,
A.W., Bond, J.F., Aalberse, R.C., Wallner, B.P., Kasaian, M.T., 1997.
The major dog allergens, Can f 1 and Can f 2, are salivary lipocalin
proteins: cloning and immunological characterization of the recombi-
nant forms. Immunology 92, 577–586.
Lacazette, E., Gachon, A.M., Pitiot, G., 2000. A novel human odorant-
binding protein gene family resulting from genomic duplicons at
9q34: differential expression in the oral and genital spheres. Hum.
Mol. Genet. 9, 289–301.
Lincoln, D.W., Renfree, M.B., 1981. Mammary gland growth and milk
ejection in the agile wallaby, Macropus agilis, displaying concurrent
asynchronous lactation. J. Reprod. Fertil. 63, 193–203.
McKenzie, H.A., Muller, V.J., Treacy, G.B., 1983. Whey proteins of milk
of the red (Macropus rufus) and eastern grey (Macropus giganteus)
kangaroo. Comp. Biochem. Physiol. 74B, 259–271.
Messer, M., Green, B., 1979. Milk carbohydrates of marsupials. II. Quali-
tative and quantitative changes in milk carbohydrates during lactation
in the tammar wallaby (Macropus eugenii). Aust. J. Biol. Sci. 32, 519–
531.
Nicholas, K.R., 1988. Control of milk protein synthesis in the marsupial
Macropus eugenii: a model system to study prolactin-dependent devel-
opment. In: Tyndale-Biscoe, C.H., Janssens, P.A. (Eds.). The Develop-
ing Marsupial: Models for Biomedical Research, Springer, Heidelberg,
pp. 68–85.
Nicholas, K.R., 1988b. Asynchronous dual lactation in a marsupial, the
tammar wallaby (Macropus eugenii). Biochem. Biophys. Res.
Commun. 154, 529–536.
Nicholas, K.R., Messer, M., Elliott, C., Maher, F., Shaw, D.C., 1987. A
novel whey protein synthesised only in late lactation by the mammary
gland from the tammar (Macropus eugenii). Biochem. J. 241, 899–904.
Perez, M.D., Calvo, M., 1995. Interaction of b-lactoglobulin with retinol
and fatty acids and its role as a possible biological function for this
protein: A review. J. Dairy Sci. 78, 978–988.
Perez, M.D., Sanchez, L., Aranda, P., Ena, J.M., Oria, R., Calvo, M., 1991.
Effect of b-lactoglobulin on the activity of pregastric lipase. A possible
role for this protein in ruminant milk. Biochim. Biophys. Acta 1123,
151.
Piotte, C.P., Hunter, A.K., Marshall, C.J., Grigor, M.R., 1998. Phylogenetic
analysis of three lipocalins present in the milk of Trichosurus vulpecula
(Phalangridae, Marsupialia). J. Mol. Evol. 46, 361–369.
Puyol, M., Perez, J.M.E., Calvo, M., 1991. Interaction of bovine b-lacto-
globulin and other bovine and human whey proteins with retinol and
fatty acids. Agric. Biol. Chem. 55, 2515–2520.
Redl, B., Holzfeind, P., Lottspeich, F., 1992. cDNA cloning and sequencing
reveals human tear prealbumin to be a member of the lipophilic-ligand
carrier protein superfamily. J. Biol. Chem. 267, 20282–20287.
Renfree, M.B., 1993. Ontogeny, genetic control, and phylogeny of female
reproduction in monotreme and therian mammals. In: Szalay, F.S.,
Novacek, M.J., McKenna, M.C. (Eds.). Mammal Phylogeny; Mesozoic
Differentiation, Multituberculates, Monotremes, Early Therians, and
Marsupials, Springer, New York, pp. 4–20.
Said, H.M., Ong, D.E., Shingleton, J.L., 1989. Intestinal uptake of retinol:
enhancement by bovine milk b-lactoglobulin. Am. J. Clin. Nutr. 49,
690.
Sambrook, J., Russell, D.W., 2001. Molecular Cloning: A Laboratory
Manual, 3rd Edition. Cold Spring Harbor Laboratory Press, Cold Spring
Harbor, NY.
Schmidhauser, C., Myers, C.A., Mossi, R., Casperson, G.F., Bissell, M.J.,
1995. Extracellular matrix dependent gene regulation in mammary
epithelial cells. In: Wilde, C.J., Peaker, M., Knight, C.H. (Eds.). Inter-
cellular Signalling in the Mammary Gland, Plenum, New York, pp.
107–119.
Seagroves, T.N., Lydon, J.P., Hovey, R.C., Vonderhaar, B.K., Rosen, J.M.,
2000. C/EBPbeta (CCAAT/enhancer binding protein) controls cell fate
determination during mammary gland development. Mol. Endocrinol.
14, 359–368.
Shaw, D., Messer, M., Scrivener, A.M., Nicholas, K.R., Griffiths, M., 1993.
Isolation, partial characterization and amino acid sequence of a-lactal-
bumin from platypus (Ornithorynchus anatinus) milk. Biochim.
Biophys. Acta 1161, 177–186.
Simpson, K.J., Ranganathan, S., Fisher, J.A., Janssens, P.A., Shaw, D.C.,
Nicholas, K.R., 2000. The gene for a novel member of the whey acidic
protein family encodes three four-disulfide core domains and is asyn-
chronously expressed during lactation. J. Biol. Chem. 275, 23074–
23081.
Stewart, F., 1984. Mammogenesis and changing prolactin receptor concen-
trations in the mammary glands of the tammar wallaby (Macropus
eugenii). J. Reprod. Fertil. 71, 141–148.
Tyndale-Biscoe, C.H., Janssens, P.A., 1988. In: Tyndale-Biscoe, C.H.,
Janssens, P.A. (Eds.). The Developing Marsupial: Models for Biome-
dical Research, Springer, Heidelberg, pp. 1–8.
Tyndale-Biscoe, C.H., Renfree, M.B., 1987. Reproductive Physiology of
Marsupials, Cambridge University Press, Cambridge, UK.
Woodlee, G.L., Gooley, A.A., Collet, C., Cooper, D.W., 1993. Origin of
late lactation protein from b-lactoglobulin in the tammar wallaby. J.
Hered. 84, 460–465.
Wu, S.-Y., Perez, D., Puyol, P., Sawyer, L., 1999. b-Lactoglobulin binds
palmitate within its central cavity. J. Biol. Chem. 274, 170–174.
J.F. Trott et al. / Gene 283 (2002) 287–297 297







