
ANALISIS METABOLOMIK SEMASA PEMASAKAN
DARIPADA PELBAGAI TISU BUAH
Garcinia mangostana Linn.
UNIVERSITI KEBANGSAAN MALAYSIA
SITI FARAH BINTI MAMAT

PERAKUAN TEStS SARJANA I DOKTORFALSAFAH (CERTIFICATION OF MASTERS / DOCTORAL THESIS)
Nama Penuh Pengarang (Author's Full Name)
No. Pendaftaran Pelajar (Student's Registration No.)
sic IMIP14 kSiwi\ \cnrT
Sesi Akademik (Academic
p860k Session)
Tajuk Tesis ANA Usk S 1NaoLetciz se1t&g PcaMPw.Atv
(Thesis Title) DARIPADA PELPjCp 1\su P"JM'I Qc4rcjt1jq
(hcffl9csftlflq Utho.
Merujuk kepada Klausa 4.2 Dasar Harts Intelek Pelajar UKM (Tambahan), tesis adatah hak milik pelajar. Saya mengaku tesis mi sebagai: (With regard to Clause 4.2 of the UKM Student Intellectual Property Policy (Supplementary), the thesis is the student's property. I hereby declare this thesis as:)
RAHSIA Mengandungi maklumat rahsia di bawah AKTA RAHSIA RASMI (CONFIDENTIAL) 1972
(Consisting of classified information under the OFFICIAL SECRETS ACT 1972)
TERHAD Mengandungi maklumat TERHAD yang telah ditentukan oleh (RESTRICTED) organisasi/badan di mane penyelidikan dijalankan
(Consisting of RESTRICTED information which has been determined by the organisation/body where the research was conducted)
I ,,/f AKSES Saya membenarkan tesis mi diterbitkan secara akses terbuka,
TERBUKA teks penuh atau dibuat salirtan untuk tujuan pengajian, ITIDAK TERHAD pembelajaran, penyelidikan sahaja. (OPEN ACCESS! (I allow this thesis to be published through open access, full text or NON-RESTRICTED) copied for study, learning and research purposes only.)
Bagi kategori Akses Terbuka/Tidak Terhad, saya membenarkan tesis (Sarjana/Doktor Falsafah) ini di simpan di Perpustakaan Universiti Kebangsaan Malaysia (UKM)* dengan syarat-syarat kegunaan seperti berikut: (For the Open Access/Non-Restricted category, I allow this (Master's/Doctoral) Thesis to be kept in the Universiti Kebangsaan Malaysia (UKM) Library with the following usage conditions:)
1. Perpustakaan UKM mempunyai hak untuk membuat salman untuk tujuan pengajian, pembelajaran, penyelidikan sahaja. (11KM Library has the right to reproduce the thesis for study, learning and research purposes only.)
2. Perpustakaan (Jniversiti Kebangsaan Malaysia dibenarkan membuat saW (1) salman tesis ni untuk tujuan pertukaran antara institusi pengajian tinggi dan mana-mana badan/ agensi kerajaan, tertakluk kepada terms dan syarat. (UKM Library is allowed to make one (1) copy of this thesis for exchange purpose among higher education institutions and any government body/agency, subject to terms and conditions.)

DISAHKAN OLEH: (VERIFIED BY:)
TANDATANGhN PELAJAR TAND dPENYELIAI (STUDENT'S SIGNATURE)
PEN—ElUCI JK SJCWAZAH (SUPERVISOR'S / CIIAIRrZPCON
SUPERVISION COMMITTEE SIGNATURE)
30 8014 -
OrvJ AL(k'.144 k' won KAD PENGENALAN I NAMA PENYELIAI W'GwJip
NO. PASPORT PENOERUSWAZAH (IDENTITY CARD/PASSPORT
(SUPERVISOR'S /CHAIRPERSON NO.) SUPERVISION COMMITTEE NAME)
Tarikhf Tarikh/ Date: (0 / to /t q Date: tI.( tt(c

ANALISIS METABOLOMIK SEMASA PEMASAKAN DARIPADA PELBAGAI
TISU BUAH Garcinia mangostana Linn.
2019
SITI FARAH BINTI MAMAT
TESIS YANG DIKEMUKAKAN UNTUK MEMPEROLEHI
IJAZAH SARJANA BIOLOGI SISTEM (SAINS BIOMOLEKUL)
INSTITUT BIOLOGI SISTEM
UNIVERSITI KEBANGSAAN MALAYSIA
BANGI

ii
PENGAKUAN
Saya akui karya ini adalah hasil kerja saya sendiri kecuali nukilan dan ringkasan yang tiap-
tiap satunya telah saya jelaskan sumbernya.
12 Jun 2019 SITI FARAH BINTI MAMAT
P88609

iii
PENGHARGAAN
Dengan nama Allah S.W.T yang Maha Pemurah lagi Maha Penyayang.
Terlebih dahulu, bersyukur saya ke hadrat Ilahi kerana dengan izin-Nya saya telah berjaya
melengkapkan penulisan tesis sebagai syarat untuk menamatkan pengajian di Institut
Biologi Sistem (INBIOSIS), UKM dalam bidang Sarjana Biologi Sistem (Sains
Biomolekul) dalam jangka masa yang telah diperuntukkan.
Sekalung penghargaan ditujukan khas kepada penyelia utama saya, Dr. Wan Mohd
Aizat Wan Kamaruddin kerana telah membimbing serta mendidik saya sepanjang tempoh
pengajian saya di INBIOSIS. Beliau banyak membantu saya dengan memupuk pelbagai
kemahiran dalam diri saya seperti kebolehan merangka dan menjalankan eksperimen,
kemampuan untuk menyampaikan maklumat dengan berkesan melalui pemantapan teknik
penulisan dan kemahiran berkomunikasi semasa melakukan pembentangan.
Seterusnya, jutaan terima kasih juga ingin saya sampaikan kepada Prof. Madya Dr.
Syarul Nataqain Baharum, Dr. Kamalrul Azlan Azizan dan Prof. Emeritus Dr. Normah
Mohd Noor merangkap penyelia bersama dalam kajian ini. Perkongsian ilmu yang
bermanfaat serta bimbingan untuk menjalankan kajian menjadi antara pendorong utama
saya untuk menyiapkan kajian ini. Selain itu, tidak lupa juga kepada felo penyelidik yang
lain di INBIOSIS serta warga kerja INBIOSIS terutamanya Puan Sarah Ibrahim, Puan
Rafidah Ahmad dan En. Syahmi Afiq Mustaza yang telah membantu secara langsung dalam
aspek teknikal analisis metabolomik.
Tidak lengkap rasanya pengajian saya di INBIOSIS tanpa rakan-rakan
seperjuangan yang sentiasa mendorong saya. Terima kasih tidak terhingga kepada rakan-
rakan kajian manggis iaitu Azhani Abdul Rahman, Salahuddin Sanusi dan Othman Mazlan,
rakan-rakan makmal metabolomik iaitu Ku Nurul Aqmar Ku Bahaudin, Yosmetha A/P
Mayalvanan, Arief Izzairy Zamani, Aqil Fitri Rosli, Fahmeeda Mohamad Jazamuddin,
Rubashiny A/P Veeramohan dan Shuhaila Sharif. Tidak lupa juga kepada rakan-rakan
pengajian yang lain termasuklah Marhaini Mostapha, Khaidatul Akmar Kamaruzaman,
Muhammad Redha Abdullah Zawawi, Muhammad Iqmal Abdullah, Pang Sze Lei dan lain-
lain. Tidak ketinggalan, ingin saya kongsikan kejayaan saya bersama ibu bapa saya, Sdiah
binti Hussin dan Mamat bin Sulong serta keluarga saya yang sentiasa menjadi tulang
belakang saya dalam apa jua yang saya lakukan. Terakhir, ucapan terima kasih juga ingin
saya sampaikan kepada pihak yang terlibat secara langsung atau tidak langsung dalam
kajian ini termasuklah biasiswa Zamalah Pendidikan UKM dan geran penyelidikan
Sciencefund (02-01-02-SF1237) yang menyumbang dari segi kewangan.
Terima kasih.

iv
ABSTRAK
Garcinia mangostana Linn. (manggis) adalah buah tropika bersifat klimakterik yang
digelar sebagai “Permaisuri buah” ekoran daripada rasanya yang manis dan unik. Manggis
mempunyai pelbagai fungsi biologi dan farmaseutikal seperti anti-kanser, anti-radang, anti-
malaria dan anti-oksida berikutan kandungan sebatian bioaktif yang tinggi terutamanya
xanton. Meskipun permintaan pasaran manggis semakin meningkat, namun pengetahuan
mengenai mekanisme pemasakan manggis yang kompleks masih lagi kurang. Oleh itu,
pendekatan biologi sistem menggunakan platform metabolomik telah digunakan dalam
kajian ini bagi meningkatkan kefahaman tentang pengawalaturan metabolik semasa
pemasakan manggis. Metabolit manggis telah diekstrak daripada empat peringkat
pemasakan (peringkat 0, 2, 4, dan 6) dan dibezakan tisunya (perikarpa, isi dan biji
manggis). Pada awalnya, beberapa kaedah pengekstrakan menggunakan bahan pelarut
berbeza dengan nisbah yang berlainan telah diuji bagi mendapatkan jumlah metabolit yang
paling optimum. Dua kaedah terbaik telah dipilih dan digunakan untuk menganalisis
kesemua sampel manggis iaitu metanol/klorofom/air (3:1:1) disertai teknik sonikasi bagi
analisis kromatografi gas-spektrometri jisim (GC-MS) dan metanol/asid formik (599:1)
disertai teknik sonikasi bagi analisis kromatografi cecair-spektrometri jisim (LC-MS).
Corak perubahan metabolit bagi setiap kumpulan eksperimen seterusnya diproses
menggunakan analisis statistik univariat dan multivariat serta analisis varians dua-hala
(two-way ANOVA) dan kaedah bioinformatik. Sebanyak 155 metabolit telah dikenalpasti
secara putatif merangkumi pelbagai kumpulan metabolit primer dan sekunder termasuklah
gula, asid amino, asid organik, fenolik, terpenoid, alkaloid dan xanton. Hasil kajian
mencadangkan bahawa metabolit berpengaruh tinggi dengan nilai pembolehubah
berkepentingan dalam unjuran (VIP) melebihi 1.00 (VIP ≥1.00) dan signifikan secara
statistik (p ≤0.05) terdiri daripada metabolit primer (contohnya L-manopiranosa, myo-
inositol, arabinofuranosa, asid galakturonik, asid L-(+)-tartarik dan butanal) dan metabolit
sekunder (contohnya neoisotegana, epirobinatidiol-(4β,8)-katesin, α-mangostin dan
gartanin). Metabolit ini mungkin menyumbang kepada degradasi dinding sel,
perkembangan rasa, warna dan aroma serta sistem pertahanan di dalam pelbagai tisu
manggis. Analisis tapak jalan seterusnya mendapati bahawa pengawalaturan metabolisme
galaktosa, metabolisme askorbat dan aldarat, metabolisme glioksilat dan dikarboksilat,
penyalingtukaran pentosa dan glukuronat serta kitaran asid trikarboksilik (TCA) berlaku
pada tahap yang berbeza semasa pemasakan manggis. Ini adalah laporan pertama yang
telah berjaya merungkai metabolom dan tapak jalan metabolik semasa pemasakan manggis
menggunakan pendekatan GC-MS dan LC-MS serta analisis bioinformatik. Hasil kajian ini
diharap akan menjadi asas kepada penambahbaikan dalam penghasilan buah dan
pembangunan teknologi lepas tuai buah manggis serta peningkatan nilai komersial dan
ekonomi buah yang bermanfaat tinggi ini pada masa hadapan.

v
METABOLOMICS ANALYSIS OF Garcinia mangostana Linn. DURING RIPENING IN
DIFFERENT FRUIT TISSUES
ABSTRACT
Garcinia mangostana Linn. (mangosteen) is a climacteric tropical fruit that has been
dubbed as the “Queen of fruit” due to its unique taste and flavour. Mangosteen possesses
various biological and pharmaceutical functions such as anti-cancer, anti-inflammation,
anti-malaria and anti-oxidant due to the high content of bioactive compounds especially
xanthones. Despite the increasing market demand for mangosteen, less is known about its
complex ripening mechanism. Hence, systems biology approach using metabolomics
platform was applied in this study to enhance the understanding of the metabolic regulation
during mangosteen ripening. Metabolites were extracted from four ripening stages (stages
0, 2, 4 and 6) and separated according to tissue types (pericarp, aril and seed). Initially, a
few extraction methods using different solvents with different ratio were tested to obtain
the most optimal number of total metabolites. The two best methods were chosen and used
to analyse all mangosteen samples which were methanol/chloroform/water (3:1:1) assisted
with sonication for gas chromatography-mass spectrometry (GC-MS) and methanol/formic
acid (599:1) assisted with sonication for liquid chromatography-mass spectrometry (LC-
MS) analyses. The metabolite distribution pattern between experimental groups were then
analysed using univariate and multivariate statistical analyses as well as two-way ANOVA
and bioinformatics approach. A total of 155 metabolites were putatively identified,
covering a broad range of primary and secondary metabolites including sugars, amino
acids, organic acids, phenolics, terpenoids, alkaloids and xanthones. The results suggest
that highly influence metabolites with variable importance of projection (VIP) value
exceeding 1.00 (VIP ≥1.00) and statistically significant (p ≤0.05) consists of primary
metabolites (for examples L-mannopyranose, myo-inositol, arabinofuranose, galacturonic
acid, L-(+)-tartaric acid and butanal) and secondary metabolites (for examples
neoisostegane, epirobinetinidol-(4β,8)-catechin, α-mangostin and gartanin). Such
metabolites may contribute to the cell wall degradation, flavour, colour and aroma
developments as well as defense systems in various manggosteen tissues. Pathway analysis
revealed that galactose metabolism, ascorbate and aldarate metabolism, glyoxylate and
dicarboxylate metabolism, pentose and glucuronate interconversions as well as
tricarboxylic acid (TCA) were differentially regulated during mangosteen ripening. This is
the first report that has successfully profiled the metabolome and metabolic pathways of
mangosteen ripening using GC-MS and LC-MS as well as bioinformatics analysis
approaches. The findings of this project could be the basis for improving the production of
the fruit and the development of related post-harvest technologies as well as increasing the
commercial and economical value of this highly beneficial fruit in the future.

vi
KANDUNGAN
Halaman
PENGAKUAN ii
PENGHARGAAN iii
ABSTRAK iv
ABSTRACT v
KANDUNGAN vi
SENARAI JADUAL x
SENARAI RAJAH xii
SENARAI SINGKATAN xiv
BAB I PENDAHULUAN
1.1 Pengenalan 1
1.2 Permasalahan Kajian 5
1.3 Objektif Kajian 7
1.3.1 Objektif umum 7
1.3.2 Objektif khusus 7
BAB II KAJIAN KEPUSTAKAAN
2.1 Pemasakan Buah 8
2.1.1 Buah klimakterik dan bukan-klimakterik 8 2.1.2 Perubahan struktur dan degradasi dinding sel 10 2.1.3 Perubahan kandungan metabolit primer dan sekunder 10
2.2 Garcinia mangostana Linn. 12
2.2.1 Proses pemasakan manggis 15 2.2.2 Manfaat dan bioaktiviti manggis 17 2.2.3 Potensi manggis di pasaran 23
2.3 Pendekatan Biologi Sistem Berasaskan Metabolomik 25
2.3.1 Penyediaan dan pengekstrakan sampel 28 2.3.2 Analisis sampel menggunakan teknik analitikal 31 2.3.3 Analisis data 37

vii
BAB III BAHAN DAN KAEDAH
3.1 Bahan 41
3.1.1 Sumber sampel 41 3.1.2 Bahan kimia dan reagen 41
3.2 Kaedah 42
3.2.1 Pensampelan dan penyediaan sampel 42 3.2.2 Pengekstrakan metabolit 42 3.2.3 Penyediaan sampel bagi analisis GC-MS 44 3.2.4 Penyediaan sampel bagi analisis LC-MS dan LC-
MS/MS 45
3.2.5 Parameter analisis GC-MS 45
3.2.6 Parameter analisis LC-MS 46 3.2.7 Pemprosesan data GC-MS 46 3.2.8 Pemprosesan data LC-MS 47 3.2.9 Analisis multivariat 49
3.2.10 Analisis bioinformatik 49
BAB IV HASIL
4.1 Perbandingan Kaedah Pengekstrakan Metabolit 50
4.1.1 Perbandingan lima kaedah pengekstrakan metabolit
disertai proses derivatisasi MSTFA bagi analisis GC-
MS 50 4.1.2 Perbandingan dua kaedah pengekstrakan metabolit
disertai derivatisasi menggunakan BSTFA bagi
analisis GC-MS 59
4.1.3 Perbandingan dua kaedah pengekstrakan serta mod
pengionan berbeza bagi analisis LC-MS 67
4.2 Pemprofilan Metabolit Tisu Perikarpa, Isi dan Biji Manggis
daripada Empat Peringkat Pemasakan Berbeza 68
4.2.1 Pemprofilan metabolit putatif menggunakan analisis
GC-MS 68 4.2.2 Pemprofilan metabolit putatif menggunakan analisis
LC-MS 76 4.2.3 Analisis tapak jalan metabolomik 90
BAB V PERBINCANGAN
5.1 Perbandingan Kaedah Pengekstrakan Metabolit 94
5.1.1 Perbandingan keberkesanan derivatisasi menggunakan
MSTFA dan BSTFA terhadap pengesanan dan
pengenalpastian metabolit 94

viii
5.1.2 Perbandingan jenis dan nisbah pelarut serta teknik lisis
terhadap pengekstrakan metabolit menggunakan lima
kaedah pengekstrakan berbeza dan derivatisasi
MSTFA 96
5.1.3 Kesan nisbah pelarut terhadap pengekstrakan
metabolit menggunakan dua kaedah pengekstrakan
berbeza dan derivatisasi BSTFA 99 5.1.4 Perbandingan kaedah pengekstrakan dan mod
pengionan LC-MS terhadap pengenalpastian metabolit 101
5.2 Pengawalaturan Metabolisme Metabolit Primer dan Sekunder
Semasa Pemasakan Manggis 102
5.2.1 Analisis multivariat mendedahkan pemisahan
kelompok metabolit mengikut peringkat pemasakan
berdasarkan jenis tisu yang berbeza 102 5.2.2 Pengawalaturan metabolik gula, asid amino, asid
organik serta fitohormon menyumbang kepada ciri-ciri
pemasakan manggis 103
5.2.3 Pengawalaturan dinamik metabolisme sekunder
manggis melibatkan metabolit fenolik, terpenoid,
alkaloid dan xanton 107
BAB VI KESIMPULAN DAN CADANGAN
6.1 Kesimpulan 114
6.2 Cadangan Kajian Lanjutan 116
RUJUKAN 118
Lampiran A Perbandingan Kromatogram Ion Jumlah (TIC) bagi Lima
Kaedah Pengekstrakan Berbeza (Kaedah 1, 2, 3, 4 dan 5) 140
Lampiran B Senarai Metabolit yang Dikenalpasti daripada Lima Kaedah
Pengekstrakan Berbeza (Kaedah 1, 2, 3, 4 dan 5) Menggunakan
Analisis GC-MS 141
Lampiran C Plot VIP bagi Analisis Perbandingan Lima Kaedah
Pengekstrakan 145
Lampiran D Perbandingan Kromatogram Ion Jumlah (TIC) bagi Sampel
Perikarpa, Isi dan Biji Manggis Menggunakan Kaedah
Pengekstrakan 3 146

ix
Lampiran E Perbandingan Kromatogram Ion Jumlah (TIC) bagi Sampel
Perikarpa, Isi dan Biji Manggis Menggunakan Kaedah
Pengekstrakan 5 147
Lampiran F Senarai Metabolit yang Dikenalpasti daripada Dua Kaedah
Pengekstrakan Berbeza (Kaedah 3 dan 5) Menggunakan
Analisis GC-MS 148
Lampiran G Plot VIP bagi Analisis Perbandingan Dua Kaedah
Pengekstrakan (Kaedah 3 dan 5) 151
Lampiran H Senarai Puncak (Pasangan RT: m/z) dalam Kromatogram bagi
Analisis Perbandingan Kaedah Pengekstrakan 1 dan 5 pada
Dua Mod Pengionan LC-MS 152
Lampiran I Senarai Metabolit daripada Analisis Pemprofilan Menggunakan
GC-MS 158
Lampiran J Senarai Metabolit VIP ≥1.00 yang Dikenalpasti daripada
Analisis GC-MS 162
Lampiran K Senarai Metabolit daripada Analisis Pemprofilan Menggunakan
LC-MS 163
Lampiran L Senarai RT: m/z yang Tidak Dapat Dikenalpasti daripada
Analisis LC-MS 171
Lampiran M Senarai Metabolit VIP ≥1.00 yang Dikenalpasti daripada
Analisis LC-MS 177
Lampiran N Senarai Metabolit Bukan Berpengaruh Tinggi yang
Dikenalpasti Melalui Kedua-dua Analisis LC-MS dan LC-
MS/MS 179
Lampiran O Struktur Molekul α-Mangostin Berserta Kedudukan Tapak
Fragmentasi yang Dicadangkan 180

x
SENARAI JADUAL
No. Jadual Halaman
Jadual 2.1 Perbezaan buah manggis dengan mesta 14
Jadual 2.2 Indeks kematangan manggis di Malaysia 15
Jadual 2.3 Klasifikasi terpenoid 22
Jadual 2.4 Kajian metabolomik terhadap pemasakan buah menggunakan
teknik nuklear magnetik resonans (NMR) dan spektrometri jisim
(MS) 33
Jadual 3.1 Senarai pangkalan data umum yang digunakan untuk
pengenalpastian metabolit bagi data daripada analisis LC-MS 48
Jadual 4.1 Ringkasan bagi lima kaedah pengekstrakan yang berbeza daripada
segi campuran pelarut, nisbah pelarut dan kaedah lisis 51
Jadual 4.2 Senarai metabolit yang diperoleh daripada analisis gambar rajah
Venn untuk pengekstrakan metabolit daripada perikarpa manggis
peringkat 6 menggunakan lima kaedah pengekstrakan berbeza 57
Jadual 4.3 Ringkasan bagi dua kaedah pengekstrakan (kaedah 3 dan 5) yang
berbeza dari segi nisbah pelarut 60
Jadual 4.4 Hasil analisis gambar rajah Venn bagi pengekstrakan metabolit
daripada tisu perikarpa, isi dan biji manggis peringkat 6
menggunakan dua kaedah pengekstrakan berbeza (kaedah 3 dan 5) 65
Jadual 4.5 Perbandingan dua kaedah pengekstrakan dan mod pengionan bagi
analisis LC-MS untuk sampel perikarpa manggis peringkat 6 68
Jadual 4.6 Analisis sisa ramalan pengesahan silang-analisis varians (CV-
ANOVA) model PLS-DA bagi analisis pemprofilan metabolit
perikarpa, isi dan biji manggis pada peringkat pemasakan berbeza
menggunakan GC-MS 73
Jadual 4.7 Analisis sisa ramalan pengesahan silang-analisis varians (CV-
ANOVA) model PLS-DA bagi analisis pemprofilan metabolit
perikarpa, isi dan biji manggis pada peringkat pemasakan berbeza
menggunakan LC-MS 80
Jadual 4.8 Senarai metabolit berpengaruh tinggi (VIP ≥1.00 dan p ≤0.05)
yang dikenalpasti melalui analisis LC-MS/MS 87

xi
Jadual 4.9 Hasil analisis tapak jalan metabolomik (MetPA) 90

xii
SENARAI RAJAH
No. Rajah Halaman
Rajah 2.1 Corak pemasakan buah klimakterik dan bukan klimakterik 9
Rajah 2.2 Keratan rentas buah manggis 13
Rajah 2.3 Struktur rangka asas xanton 18
Rajah 2.4 Xanton utama di dalam manggis 20
Rajah 2.5 Sistem kromatografi gas-spektrometri jisim (GC-MS) 35
Rajah 2.6 Sistem kromatografi cecair-spektrometri jisim (LC-MS) 36
Rajah 4.1 Bilangan dan jenis metabolit yang diekstrak daripada perikarpa
manggis peringkat 6 menggunakan lima kaedah pengekstrakan
berbeza 53
Rajah 4.2 Plot skor PCA (a) dan PLS-DA (b) yang membandingkan taburan
metabolit di dalam perikarpa manggis peringkat 6 menggunakan
lima kaedah pengekstrakan berbeza 54
Rajah 4.3 Plot pemuatan PCA (a) dan PLS-DA (b) yang membandingkan
taburan metabolit di dalam perikarpa manggis peringkat 6
menggunakan lima kaedah pengekstrakan berbeza 55
Rajah 4.4 Analisis gambar rajah Venn untuk analisis perbandingan lima
kaedah pengekstrakan bagi perikarpa manggis peringkat 6 57
Rajah 4.5 Bilangan dan jenis metabolit yang diekstrak daripada perikarpa, isi
dan biji manggis peringkat 6 menggunakan dua kaedah
pengekstrakan berbeza (kaedah 3 dan 5) 60
Rajah 4.6 Plot skor PCA (a) dan PLS-DA (b) yang membandingkan taburan
metabolit di dalam perikarpa, isi dan biji manggis peringkat 6
menggunakan dua kaedah pengekstrakan berbeza (kaedah 3 dan 5) 62
Rajah 4.7 Plot pemuatan PCA (a) dan PLS-DA (b) yang membandingkan
taburan metabolit di dalam tisu perikarpa, isi dan biji manggis
peringkat 6 menggunakan dua kaedah pengekstrakan berbeza
(kaedah 3 dan 5) 63

xiii
Rajah 4.8 Analisis gambar rajah Venn yang membandingkan metabolit yang
diekstrak daripada tisu perikarpa, isi dan biji manggis peringkat 6
menggunakan dua kaedah pengekstrakan berbeza (kaedah 3 dan 5) 64
Rajah 4.9 Analisis PCA dan PLS-DA bagi analisis pemprofilan metabolit
perikarpa, isi dan biji manggis pada peringkat pemasakan berbeza
menggunakan GC-MS 70
Rajah 4.10 Ujian permutasi validasi model PLS-DA bagi analisis pemprofilan
metabolit perikarpa, isi dan biji manggis pada peringkat
pemasakan berbeza menggunakan GC-MS 72
Rajah 4.11 Graf metabolit berpengaruh tinggi (VIP ≥1.00 and p ≤0.05) bagi
kumpulan metabolit gula (a), glikosida gula (b), asid gula (c),
alkohol gula (d), alkohol (e), asid organik (f), aldehid (g) dan lain-
lain (h) 74
Rajah 4.12 Analisis PCA dan PLS-DA bagi analisis pemprofilan metabolit
perikarpa, isi dan biji manggis pada peringkat pemasakan berbeza
menggunakan LC-MS 78
Rajah 4.13 Ujian permutasi validasi model PLS-DA bagi analisis pemprofilan
metabolit perikarpa, isi dan biji manggis pada peringkat
pemasakan berbeza menggunakan LC-MS 79
Rajah 4.14 Graf metabolit berpengaruh tinggi (VIP ≥1.00 and p ≤0.05) bagi
kumpulan metabolit gula (a), asid gula (b), fosfat gula (c),
glikosida (d), asid organik (e), asid lemak (f), asid amino dan
terbitan (g), kuinon (h), alkaloid (i), terpenoid (j) dan xanton (k) 82
Rajah 4.15 Graf metabolit berpengaruh tinggi (VIP ≥1.00 and p ≤0.05) bagi
kumpulan metabolit fenolik (a) dan lain-lain (b) 83
Rajah 4.16 Pemetaan metabolit dalam tapak jalan metabolik pemasakan
manggis 93

xiv
SENARAI SINGKATAN
Δppm Ralat jisim molekul
1-MCP 1-Metilsiklopropena
ACC Asid 1-aminosiklopropana-1-karboksilik
AMDIS Automated Mass Spectral Deconvolution and Identification
System
ANOVA Analisis varians
APCI Pengionan kimia tekanan atmosfera
AutoMS Spektrometri jisim automatik
BSTFA N,O-Bis(trimetilsilil)trifluoroasetamida
CE-MS Elektroforesis kapilari-spektrometri jisim
ChEBI Chemical Entities of Biological Interest
CV-ANOVA Pengesahan silang-analisis varians
DF Darjah kebebasan
DMMBP 1,2-Dimetoksi-13-metil-[1,3]benzodioksolo[5,6-c]fenantridina
ES-242-4 (3S,3'S,4S,4'S)-7,7',9,9'-Tetrametoksi-3,3'-dimetil-3,3',4,4'-
tetrahidro-1H,1'H-5,5'-bibenzo[g]isokromena-4,4',10,10'-tetrol
ESI Pengionan semburan elektro
F Nilai ujian F
FiehnLib Fiehn Metabolomics Library
GC-FID Kromatografi gas-pengesan pengionan api
GC-MS Kromatografi gas-spektrometri jisim
GMD Golm Metabolic Databases
HMDB Human Metabolome Database
HPLC Kromatografi cecair berprestasi tinggi

xv
KEGG Kyoto Encyclopedia of Genes and Genomes
LC-MS Kromatografi cecair-spektrometri jisim
LC-MS/MS Kromatografi cecair-spektrometri jisim seiring
LC-QTOF-MS Kromatografi cecair-masa penerbangan tetrakutub-spektrometri
jisim
MetPA Analisis tapak jalan metabolit
MRM Pemantauan tindak balas berganda
MS Purata persegi
MS Spektrometri jisim
MSEA Analisis pengkayaan set metabolit
MSTFA N-Metil-N-(trimetilsilil)trifluoroasetamida
m/z Nisbah jisim kepada caj
NIST National Institute of Standards and Technology
NMR Resonans magnetik nuklear satu dimensi
p Nilai kebarangkalian
PC Komponen utama
PCA Analisis komponen utama
PLS-DA Analisis diskriminan-separa kuasa dua terkecil
q Tetrakutub
RI Indeks penahanan
RT Masa penahanan
SAM S-Adenosil-metionina
SD Sisihan piawai
SE Ralat piawai
SIR Rakaman ion terpilih

xvi
SRM Pemantauan tindak balas terpilih
SS Jumlah persegi
TCA Asid trikarboksilik
TIC Kromatogram ion jumlah
TMCS Trimetilklorosilana
TOF Masa penerbangan
UPLC Kromatografi cecair berprestasi ultra
UV Ultraungu
VIP Pembolehubah berkepentingan dalam unjuran

BAB I
PENDAHULUAN
1.1 PENGENALAN
Pemasakan buah adalah satu proses yang diprogram secara genetik dan melibatkan
pelbagai siri pengubahsuaian daripada aspek fisiologi, biokimia dan struktur buah
seperti perubahan struktur dinding sel, kadar metabolisme, pengumpulan pigmen serta
biosintesis metabolit sekunder (Mworia et al. 2012; Prasanna et al. 2007). Bagi buah
klimakterik, perubahan ini dirangsang oleh peningkatan kadar kandungan hormon
etilena semasa pemasakan terjadi (Osorio et al. 2013). Daripada aspek biokimia, proses
pemasakan melibatkan perubahan pada kadar kandungan metabolit primer seperti gula,
asid organik, alkohol serta metabolit sekunder seperti fenolik, flavonoid, terpenoid dan
alkaloid (Baldi et al. 2018; Osorio & Fernie 2014). Perubahan pada kadar kandungan
metabolit ini turut melibatkan penghasilan pelbagai nutrisi semasa pemasakan seperti
serat, asid askorbik dan pelbagai vitamin serta anti-oksida (Osorio et al. 2013; Prasanna
et al. 2007). Hal ini seterusnya akan menyebabkan perubahan daripada segi fisiologi
buah di mana rupa, tekstur, rasa dan aroma buah akan berubah sehinggalah ianya
mencapai tahap akhir pemasakan. Perubahan yang berlaku ini akan meningkatkan daya
tarikan buah kepada hidupan lain termasuklah manusia dan haiwan, yang seterusnya
membawa kepada penyebaran biji benih secara meluas.
Garcinia mangostana Linn. atau lebih dikenali sebagai manggis yang turut
digelar sebagai “Permaisuri buah” adalah buah tropika yang berasal daripada famili
Clusiaceae. Manggis ditanam secara meluas di selatan Vietnam, Myanmar, Thailand
dan juga Malaysia. Di Malaysia, manggis ditanam secara meluas termasuklah di
beberapa kawasan utama iaitu negeri Perak, Johor, Kelantan, Kedah dan Pahang.
Manggis dikelaskan sebagai buah klimakterik berdasarkan pada peningkatan kadar

2
respirasi dan kadar penghasilan hormon etilena sepanjang proses pemasakan buah ini
terjadi (Lerslerwong et al. 2013; Palapol et al. 2009b). Manggis, terutamanya perikarpa
atau kulit luar manggis digunakan dalam perubatan tradisional untuk mengubati
pelbagai jenis penyakit seperti jangkitan luka, cirit-birit, sakit perut, leukorea, gonorea
dan ulser yang kronik (Chaijaroenkul et al. 2014; Mohamed et al. 2014; Shibata et al.
2011). Pada masa kini, manggis juga telah digunakan sebagai ramuan dalam makanan
tambahan bernutrisi dan produk kosmetik (Mohamed et al. 2014).
Sehingga kini, para penyelidik telah melaporkan bahawa manggis mempunyai
pelbagai manfaat seperti anti-radang, anti-malaria, anti-alergi, anti-oksida dan anti-
kanser (Ayman et al. 2019; Chaijaroenkul et al. 2014; Ibrahim et al. 2016; Mohamed et
al. 2014; Shibata et al. 2011; Thong et al. 2015). Kehadiran pelbagai sebatian bioaktif
terutamanya xanton (Mohamed et al. 2014; Nguyen et al. 2005) telah dilaporkan
menyumbang kepada fungsi biologikal dan farmaseutikal di dalam manggis. Selain itu,
kehadiran metabolit sekunder seperti fenolik, alkaloid dan terpenoid turut menyumbang
kepada bioaktiviti manggis termasuklah sebagai anti-mutagenik, anti-proliperatif dan
anti-mikrob (Kampa et al. 2004; Qin et al. 2017; Zarena & Sankar 2012). Kajian lepas
lebih tertumpu kepada pemencilan dan penggunaan sebatian bioaktif daripada manggis,
namun, mekanisme molekular yang berlaku semasa pemasakan manggis masih belum
difahami serta diperincikan dengan baik dan mendalam terutamanya melibatkan
pengawalaturan penghasilan metabolit primer dan sekunder. Antara faktor utama yang
mempengaruhi perubahan dalam komposisi metabolit buah adalah proses pemasakan.
Justeru itu, pendekatan biologi sistem seperti metabolomik diketengahkan dalam kajian
ini untuk mengkaji corak perubahan metabolit manggis semasa proses pemasakan
terjadi seterusnya merungkai pemasakan manggis yang kompleks.
Metabolomik digunakan untuk mengkaji pelbagai metabolit sasaran atau bukan
sasaran yang mempunyai berat molekular yang rendah (50-1500 Dalton) secara global
(Aretz & Meierhofer 2016; Patti et al. 2012; Zhang et al. 2016). Sehingga kini,
metabolomik telah diaplikasikan secara meluas dalam pelbagai cabang penyelidikan
termasuklah kajian toksikologi tumbuhan, nutrisi, sains tumbuhan dan biologi sistem
(Robertson et al. 2011; Sumner et al. 2003). Metabolomik didefinisikan sebagai analisis
pemprofilan metabolit secara komprehensif di dalam cecair biologi, sel, tisu, organ atau

3
organisma (Johnson et al. 2016). Analisis metabolomik menawarkan pengukuran
metabolit secara kualitatif dan kuantitatif untuk mengenalpasti metabolit yang
memainkan peranan sama ada sebagai substrat atau produk dalam sesuatu tapak jalan
biokimia (Aizat et al. 2014; Fiehn 2002) dan merupakan kaedah yang berpotensi tinggi
untuk mengkaji dan meneroka pengawalaturan fungsi dan selular daripada mana-mana
sampel hidupan (Patti et al. 2012). Metabolit bertindak sebagai rekod visual bagi
keadaan selular dan merupakan gambaran langsung kepada fenotip dan aktiviti
biokimia, berbeza dengan gen atau protein (Baharum & Azizan 2018; Patti et al. 2012).
Ini disebabkan fungsi gen dipengaruhi oleh pengawalaturan epigenetik dan protein pula
dipengaruhi oleh modifikasi pasca-translasi (Patti et al. 2012).
Analisis metabolomik umumnya dikategorikan kepada dua iaitu analisis sasaran
dan analisis bukan sasaran. Analisis sasaran memfokuskan kepada pengukuran
metabolit tertentu yang telah disasarkan. Analisis ini diaplikasikan untuk menilai aras
sesuatu sebatian di dalam sampel yang wujud dalam keadaan atau rawatan tertentu.
Kaedah ini kebiasaannya memerlukan tahap penulenan sebatian yang lebih tinggi dan
kaedah pemencilan metabolit yang spesifik terhadap metabolit yang ingin dipencilkan.
Berbeza dengan kaedah analisis bukan sasaran (turut dikenali sebagai analisis
komprehensif), analisis ini memfokuskan kepada pengenalpastian metabolit secara
maksimum dan melibatkan lebih daripada satu tapak jalan biokimia (Cevallos-Cevallos
et al. 2009; Patti et al. 2012; Soga et al. 2003).
Terdapat dua jenis teknik utama yang sering digunakan dalam kajian
metabolomik iaitu resonans magnetik nuklear (NMR) dan spektrometri jisim (MS). MS
sering digabungkan dengan sistem pemisah sebatian seperti kromatografi gas (GC),
kromatografi cecair (LC) atau elektroforesis kapilari (CE) (Aizat et al. 2014; Goodacre
et al. 2004; Patti et al. 2012). Pengasingan sebatian bagi analisis GC-MS dilakukan
berdasarkan kadar kemeruapan sebatian dengan cara mengalirkan gas lengai seperti
helium, nitrogen, argon atau hidrogen yang akan bertindak untuk membawa sampel
melalui fasa pegun di dalam kolum (Hussain & Maqbool 2014). Sebatian yang
mempunyai kadar kemeruapan yang lebih tinggi akan berimigrasi dengan lebih pantas
di dalam kolum berbanding sebatian yang mempunyai kadar kemeruapan yang lebih
redah. Bagi sebatian berkutub yang tidak meruap, proses derivatisasi kimia diperlukan

4
untuk menukarkan sebatian ini kepada molekul terbitan yang meruap dan stabil haba
bagi membolehkan sebatian ini sesuai untuk dianalisis menggunakan sistem GC-MS.
Sistem LC mampu memisahkan pelbagai jenis sebatian organik dengan efektif
termasuklah molekul kecil, peptida, protein dan juga metabolit gula (Kang 2012).
Aplikasi kaedah LC-MS dalam kajian metabolomik telah berkembang secara meluas
setelah beberapa teknik pengionan seperti sumber ion tekanan atmosfera (API),
pengionan semburan elektro (ESI) dan pengionan kimia tekanan atmosfera (APCI)
diperkenalkan. Teknik-teknik ini mempunyai tahap sensitiviti yang tinggi sekali gus
membolehkan sistem ini mengenalpasti dan memisahkan sebatian dalam julat yang
lebih luas berbanding kaedah GC-MS (Kang 2012). Teknologi LC-MS menggabungkan
teknik pemisahan kromatografi cecair berprestasi tinggi (HPLC) yang mempunyai
resolusi tinggi dan fungsi analisis yang fleksibel dengan pengenalpastian metabolit
menggunakan komponen MS (Shulaev 2006). Komponen sebatian akan dipisahkan di
dalam kolum HPLC dan seterusnya memasuki bahagian MS di mana ion yang terhasil
daripada sebatian akan dikenal pasti. Analisis LC-MS seringkali diikuti dengan analisis
spektrometri jisim seiring (MS/MS) menggunakan teknik seperti spektrometri jisim
automatik (AutoMS) dan pemantauan tindak balas berganda (MRM) untuk kajian yang
lebih teliti dan mendalam mengenai sifat molekular sesuatu metabolit.
Umumnya, analisis GC-MS digunakan untuk mengenalpasti dan
mengkuantifikasi sebatian meruap dan stabil haba seperti alkohol dan minyak pati
tumbuhan (Baharum et al. 2010) serta kumpulan metabolit berkutub dan semi-kutub
yang kebanyakannya terdiri daripada metabolit primer seperti gula, asid organik dan
asid amino (Gullberg et al. 2004; Lee et al. 2013; Okazaki et al. 2016; t’Kindt et al.
2009). Sementara itu, analisis LC-MS pula memfokuskan kepada jenis metabolit yang
lebih pelbagai serta mempunyai struktur yang lebih kompleks, merangkumi metabolit
primer (seperti gula disakarida dan polisakarida) serta metabolit sekunder (seperti
fenolik, flavonoid, alkaloid dan xanton), bergantung kepada jenis fasa pegun yang
digunakan di dalam sistem LC-MS (De Vos et al. 2007; Lee et al. 2013; t’Kindt et al.
2009).

5
Bagi kajian ini, kedua-dua analisis GC-MS dan LC-MS telah digunakan untuk
mengenalpasti dan mengukur aras metabolit jumlah bagi perikarpa, isi dan biji manggis
daripada peringkat pemasakan yang berbeza iaitu peringkat 0 (hijau), 2 (hijau
kekuningan disertai tompokan merah jambu), 4 (merah keperangan) dan 6 (ungu gelap)
yang ditentukan berdasarkan Indeks Kematangan Manggis Malaysia (Osman & Milan
2006). Data daripada kedua-dua sistem analitikal GC-MS dan LC-MS diproses dan
dikaji menggunakan beberapa analisis seperti analisis statistik univarit dan multivariat
serta analisis tapak jalan metabolik. Akhir sekali, mekanisme molekular semasa proses
pemasakan manggis telah dirungkaikan dan dibincangkan berdasarkan corak perubahan
metabolit manggis semasa proses ini terjadi. Kajian ini membekalkan maklumat asas
yang boleh digunakan bagi penghasilan teknologi untuk meningkatkan jangka hayat
lepas tuai buah manggis dan penambahbaikan proses penyimpanan buah manggis,
seterusnya meningkatkan nilai komersial dan ekonomi buah tropika ini. Secara tidak
langsung, kadar pengeksportan buah manggis akan turut meningkat.
1.2 PERMASALAHAN KAJIAN
Penggunaan manggis, terutamanya kulit buah manggis sebagai bahan perubatan
alternatif untuk mengubati pelbagai jenis penyakit terutamanya luka dan cirit-birit telah
bermula sejak beberapa abad yang lepas (Osman & Milan 2006). Potensi manggis
sebagai ejen penyembuhan ini mendapat perhatian para penyelidik serta industri yang
seterusnya membawa kepada penggunaan buah ini dalam penghasilan pelbagai jenis
ubat-ubatan, produk makanan tambahan dan juga produk kosmetik (Mohamed et al.
2014). Di Malaysia, manggis pernah dipilih sebagai buah kemegahan negara dalam
Dasar Pertanian Negara Ketiga (1998-2010) (Abu Dardak et al. 2011). Kualiti manggis
dari negara ini yang tinggi dan bebas daripada bahan kimia diterima baik serta mendapat
permintaan tinggi daripada negara Eropah dan China (Abu Dardak et al. 2011). Pasaran
manggis turut dijangka terus berkembang dan permintaan manggis segar dunia dijangka
meningkat sebanyak lebih 3% setahun kerana wujudnya pasaran baru di negara Arab
seperti Arab Saudi, Emeriah Arab Bersatu dan Qatar (Abu Dardak et al. 2011).
Namun, pada tahun 2017, laporan statistik yang dikeluarkan oleh Lembaga
Pemasaran Pertanian Persekutuan (FAMA), Malaysia menyatakan kadar pengeksportan

6
manggis ke negara luar telah berkurang sebanyak 63.6% pada tahun 2015 berbanding
tahun sebelumnya iaitu tahun 2014 (Lembaga Pemasaran Pertanian Persekutuan 2017).
Antara salah satu faktor yang menyebabkan kemerosotan dari segi kadar eksport
manggis adalah persaingan daripada negara jiran seperti Thailand dan Indonesia.
Justeru itu, kualiti manggis yang dieksport haruslah dititikberatkan kerana faktor ini
mempengaruhi permintaan manggis di pasaran. Teknologi lepas tuai memainkan
peranan penting dalam memastikan manggis dapat dieksport dalam keadaan yang
terbaik. Kunci kepada penambahbaikan dan peningkatan kualiti buah adalah dengan
memahami mekanisme pemasakan sesuatu buah itu.
Walau bagaimanapun, mekanisme pemasakan manggis yang kompleks masih
tidak difahami dengan jelas dan terperinci sehingga kini. Manggis yang dituai pada
peringkat pemasakan yang berbeza akan menghasilkan kualiti buah masak yang berbeza
kerana kadar metabolit termasuklah sebatian bioaktif seperti fenolik dan xanton di
dalam buah manggis dipengaruhi oleh beberapa faktor termasuklah peringkat
pemasakan dan pengurusan lepas tuai buah (Palapol et al. 2009b; Zadernowski et al.
2009). Selain itu, identiti serta kadar sebatian bioaktif ini serta metabolit sekunder yang
lain seperti terpenoid dan alkaloid di dalam buah manggis pada peringkat pemasakan
yang berbeza masih belum dilaporkan dengan terperinci. Perbezaan kandungan
metabolit antara tisu manggis yang berbeza juga masih tidak jelas dan memerlukan
kajian lanjutan bagi meneroka metabolit jumlah daripada tisu ini.
Pendekatan metabolomik digunakan dalam kajian ini untuk mengkaji corak
perubahan metabolit semasa proses pemasakan manggis sekaligus merungkai
mekanisme pemasakan manggis dan seterusnya mendalami proses molekular yang
terlibat. Justeru itu, pemahaman ini akan menjadi asas serta membantu kepada kajian
seterusnya pada masa hadapan. Pemahaman ini diharap turut dapat menyumbang
kepada industri pertanian di negara ini melalui penghasilan bahan atau teknik bagi
kawalan tanaman di ladang atau melalui penambahbaikan teknologi lepas tuai buah.
Hal ini seterusnya akan meningkatkan nilai komersial buah-buahan tempatan
terutamanya manggis, serta industri pembuatan seperti penghasilan makanan dan
minuman, produk kosmetik serta pil makanan tambahan berasaskan manggis.

7
1.3 OBJEKTIF KAJIAN
1.3.1 Objektif umum
Matlamat kajian ini adalah untuk mengkaji corak perubahan metabolit manggis semasa
proses pemasakan dengan cara memprofil metabolit manggis pada peringkat
pemasakan yang berbeza iaitu peringkat 0, 2, 4 dan 6. Tisu manggis yang berbeza iaitu
perikarpa, isi dan biji digunakan untuk perbandingan yang lebih terperinci.
1.3.2 Objektif khusus
Objektif bagi kajian ini adalah seperti berikut:
1) Mendapatkan kaedah pengekstrakan metabolit daripada manggis yang sesuai
dengan membandingkan faktor jenis dan nisbah pelarut serta teknik
pengekstrakan
2) Memprofil metabolit daripada perikarpa, isi dan biji manggis pada empat
peringkat pemasakan yang berbeza iaitu peringkat 0, 2, 4 dan 6 menggunakan
kaedah GC-MS dan LC-MS
3) Pemetaan tapak jalan biokimia pemasakan manggis menggunakan analisis
bioinformatik

8
BAB II
KAJIAN KEPUSTAKAAN
2.1 PEMASAKAN BUAH
Buah-buahan merupakan makanan yang mempunyai kepentingan dari segi ekonomi
serta memainkan peranan penting dalam diet pemakanan manusia dan haiwan herbivor
dengan membekalkan pelbagai nutrien seperti vitamin, serat dan mineral yang
diperlukan untuk mengekalkan hidup yang sihat (Giovannoni et al. 2017; Osorio &
Fernie 2014). Buah-buahan yang masak menjadi tarikan kepada hidupan yang turut
bertindak sebagai agen penting dalam penyebaran dan evolusi perkembangan biji benih
(Giovannoni et al. 2017). Proses pemasakan buah bermula apabila biji benih buah telah
mengalami proses pertumbuhan dan perkembangan yang lengkap.
Pemasakan buah merupakan satu proses yang kompleks, tersusun dan telah
dijadualkan secara genetik yang membawa kepada penghasilan rasa buah yang enak
disertai dengan aroma yang harum (Giovannoni 2001; Giovannoni et al. 2017; Osorio
& Fernie 2014). Proses pemasakan buah melibatkan satu siri perubahan dan
pengubahsuaian dari aspek fisiologi, biokimia serta struktur buah yang menyebabkan
perubahan pada rupa, tekstur, rasa dan aroma buah. Perubahan dan pengubahsuaian ini
termasuklah pada kadar metabolisme, peleraian dan penyusunan semula polisakarida
dinding sel, penghasilan dan pengumpulan pigmen serta penghasilan metabolit
sekunder (Giovannoni et al. 2017; Mworia et al. 2012; Prasanna et al. 2007).
2.1.1 Buah klimakterik dan bukan-klimakterik
Terdapat dua pengkelasan utama bagi pemasakan buah iaitu buah klimakterik dan
bukan-klimakterik yang ditentukan berdasarkan perbezaan pada kadar respirasi dan

9
kadar penghasilan hormon etilena (Aizat et al. 2013; Giovannoni 2001; Giovannoni et
al. 2017; Saladié et al. 2015) seperti yang dipamerkan di dalam Rajah 2.1 di bawah.
Buah klimakterik seperti tomato, pisang, avokado, epal dan manggis mengalami
peningkatan pada kadar respirasi dan kadar penghasilan hormon etilena sepanjang
proses pemasakan berlaku (Giovannoni et al. 2017; Osorio & Fernie 2014; Osorio et al.
2013). Kadar hormon etilena adalah minimum pada peringkat awal pemasakan dan
meningkat secara mendadak sehingga mencapai kadar optimum semasa kemuncak
proses pemasakan, seterusnya menurun pada peringkat akhir pemasakan sebelum buah
memasuki fasa senesens (Kou & Wu 2018; Paul et al. 2012; Prasanna et al. 2007) (Rajah
2.1).
Hormon etilena diperlukan untuk pengawalaturan serta untuk melengkapkan
proses pemasakan dalam buah klimakterik (Osorio et al. 2013). Oleh itu, buah
klimakterik boleh mengalami pemasakan yang sempurna meskipun telah berpisah
daripada pokok induk. Berbeza dengan buah bukan-klimakterik seperti strawberi dan
buah-buahan sitrus yang mana ianya tidak bergantung kepada kehadiran hormon etilena
untuk merangsang proses pemasakan. Proses pemasakan buah bukan klimakterik akan
terhenti sekiranya buah terpisah daripada pokok induk sebelum proses pemasakan
lengkap (Kou & Wu 2018; Paul et al. 2012).
Rajah 2.1 Corak pemasakan buah klimakterik dan bukan klimakterik
Sumber: Paul dan Pellny (2003)

10
2.1.2 Perubahan struktur dan degradasi dinding sel
Pelembutan buah yang berlaku disebabkan perubahan atau degradasi pada struktur
polisakarida dinding sel adalah antara fenomena utama yang terjadi semasa proses
pemasakan (Brummell 2006; Osorio & Fernie 2014). Kelas-kelas utama polisakarida
dinding sel yang terlibat semasa pemasakan buah adalah pektin, selulosa dan
hemiselulosa (Brummell 2006). Pelembutan buah semasa proses pemasakan tidak
semestinya melibatkan perubahan pada keseluruhan komposisi dinding sel tetapi ianya
berlaku mungkin hanya disebabkan perubahan pada sebahagian daripada komponen
polisakarida (Robinson & Davies 2000). Pelembutan buah semasa proses pemasakan
terjadi disebabkan berlakunya pengubahsuaian pada polisakarida dinding sel primer
serta lamela tengah yang menyebabkan struktur sel menjadi lemah. Lamela tengah
adalah lapisan yang kaya dengan pektin yang berada di antara sel dan bertindak sebagai
penyambung antara sel (Jarvis et al. 2003; Saladié et al. 2007).
Kajian lepas menggunakan mikroskop elektron mendapati bahawa pembubaran
pada lapisan lamela tengah adalah merupakan perubahan awal yang belaku pada
dinding sel semasa proses pemasakan terjadi (Crookes & Grierson 1983; Hallett et al.
1992). Proses ini diikuti dengan gangguan pada lapisan dinding sel primer, menandakan
telah berlakunya proses degradasi yang signifikan pada dinding sel (Brummell 2006).
Seterusnya, perubahan pada ikatan antara polimer akan menyebabkan pemisahan sel
meningkat, dan apabila disertai dengan perubahan turgor akan menyebabkan
pelembutan dan perubahan pada struktur buah (Brummell 2006). Tekanan turgor
dikatakan menurun semasa pemasakan buah (Harker & Sutherland 1993; Shackel et al.
1991) disebabkan pengumpulan larutan osmotik di dalam apoplas serta kehilangan
kandungan air di dalam buah. Pengurangan pada tekanan turgor ini seterusnya akan
mengurangkan tekanan pengembangan terhadap dinding sel buah (Brummell 2006).
2.1.3 Perubahan kandungan metabolit primer dan sekunder
Kebanyakan buah mengalami perubahan dari segi warna, rasa, tekstur, aroma dan juga
rintangan terhadap patogen semasa proses pemasakan berlaku (Giovannoni 2001;
Giovannoni et al. 2017). Perkembangan pada rasa, warna dan aroma buah terjadi
disebabkan penghasilan dan perubahan kandungan metabolit primer dan sekunder

11
(Baldi et al. 2018; Osorio & Fernie 2014). Metabolit primer adalah merupakan sebatian
perantara bagi tapak jalan katabolik dan anabolik yang terjadi dalam semua tumbuhan
seperti gula dan terbitan (metabolit terbitan gula seperti asid gula, alkohol gula dan
fosfat gula), asid organik, asid amino dan asid lemak.
Metabolit primer berperanan penting dalam pertumbuhan dan perkembangan
tumbuhan serta diperlukan oleh tumbuhan untuk terus hidup. Kumpulan metabolit ini
terlibat secara langsung dalam proses fotosintesis, kitaran Krebs (kitaran asid
trikarboksilik) serta rantaian pengangkutan elektron (Klaser Cheng 2017). Gula atau
gula alkohol ditukarkan ke dalam bentuk kanji di dalam buah (contoh: mangga, pisang,
buah kiwi) atau lipid (contoh: zaitun) atau disimpan oleh buah dalam bentuk gula
penurun (contoh: tomato) dan sukrosa (contoh: tembikai, anggur) (Osorio & Fernie
2014). Sukrosa, glukosa dan fruktosa adalah metabolit gula yang paling banyak terdapat
di dalam tumbuhan serta merupakan komponen makanan yang terbanyak diperoleh
daripada tumbuhan. Rasa buah yang manis ditentukan oleh jumlah kandungan gula serta
nisbah antara gula yang berlainan yang hadir di dalam buah. Kemanisan buah adalah
ciri-ciri utama buah yang menentukan kualiti buah tersebut.
Selain daripada metabolit gula, asid organik juga adalah metabolit penting yang
menyumbang kepada rasa dan kualiti buah (Osorio & Fernie 2014). Asid organik turut
berperanan sebagai pelopor kepada biosintesis pelbagai metabolit sekunder (Klaser
Cheng 2017). Kandungan asid organik bergantung kepada beberapa aktiviti tapak jalan
utama seperti glikolisis dan kitaran asid trikarboksilik (TCA). Asid organik utama
terdapat dalam buah adalah metabolit perantaraan sitrat dan malat. Manakala, beberapa
metabolit perantaraan TCA yang lain iaitu oksalat, suksinat, isositrat dan fumarat hadir
dalam jumlah yang lebih rendah berbanding sitrat dan malat (Klaser Cheng 2017).
Metabolit sekunder pula tidak terlibat secara langsung dalam pertumbuhan dan
perkembangan tumbuhan (Klaser Cheng 2017; Lattanzio et al. 2008). Namun,
kumpulan metabolit ini diperlukan oleh tumbuhan untuk adaptasi dengan alam sekitar
termasuklah untuk ketahanan tumbuhan terhadap serangan herbivor, patogen, penyakit,
haiwan penyebar biji benih serta tekanan abiotik (hujan, kemarau dan sebagainya)
(Lattanzio et al. 2008; Wink & Schimmer 2018).

12
Selain itu, metabolit sekunder juga berperanan sebagai isyarat untuk penyebaran
biji benih dan pendebungaan (Wink & Schimmer 2018). Berbeza dengan metabolit
primer yang hadir dalam kebanyakan tumbuhan, metabolit sekunder lebih bersifat
spesifik terhadap sesetengah spesies tumbuhan atau hanya dihasilkan dan terkumpul
dalam sesetengah tisu bagi tumbuhan tersebut (Klaser Cheng 2017; Lattanzio et al.
2008). Biosintesis metabolit sekunder adalah melibatkan beberapa tapak jalan utama
iaitu tapak jalan sikimat, mevalonat dan asetat ( Klaser Cheng 2017; Lattanzio et al.
2008). Tapak jalan sikimat menghasilkan metabolit fenilpropanoid manakala tapak
jalan mevalonat pula menghasilkan terpenoid. Asid lemak dan poliketida pula terhasil
melalui tapak jalan asetat (Klaser Cheng 2017; Lattanzio et al. 2008). Kumpulan
metabolit ini menyumbang kepada rasa, warna, dan kadar rintangan buah terhadap
tekanan biotik dan abiotik semasa proses pemasakan terjadi (Baldi et al. 2018; Tohge
et al. 2014).
2.2 Garcinia mangostana Linn.
Garcinia mangostana Linn. merupakan salah satu buah-buahan tropika yang ditanam
secara meluas di negara-negara Asia Tenggara termasuklah Selatan Vietnam,
Myanmar, Thailand dan Malaysia. Umumnya, Garcinia mangostana Linn. tergolong
dalam famili Clusiaceae (Guttiferae) dan dikenali sebagai manggis di Malaysia. Selain
itu, buah ini juga turut dikenali dengan beberapa nama lain iaitu masta, mestor,
sementoh atau semetah di Malaysia dan Indonesia, mangkhut di Thailand, cay mang cut
di Vietnam, mangostan di Filipina dan mangostin di India (Osman & Milan 2006). Di
Malaysia, penanaman manggis bertumpu di beberapa negeri utama iaitu Johor,
Kelantan, Perak, Kedah dan Pahang (Abu Dardak et al. 2011; Jabatan Pertanian Negeri
Pulau Pinang 2018).
Pokok manggis merupakan sejenis tumbuhan malar hijau tropika yang
mempunyai batang berkayu dengan ketinggian antara 6-15 meter (Osman & Milan
2006) dan kanopi pokok yang membentuk piramid bersaiz sederhana iaitu 9-12 meter
lebar. Tumbuhan berbunga ini mempunyai daun berwarna hijau yang berbentuk bujur
dan eliptik, berukuran antara 15-25 cm panjang dan 7-13 cm lebar serta mempunyai
struktur yang tebal, keras dan licin (Brunner & Morales-Payan 2010; Failla &

13
Gutiérrez-Orozco 2017; Jabatan Pertanian Negeri Pulau Pinang 2018; Osman & Milan
2006). Setiap organ pokok manggis termasuk daun, batang dan buah manggis
menghasilkan getah berwarna kuning (Lan 1989; Osman & Milan 2006).
Buah manggis pula bersaiz purata antara 6-8 cm daripada garis pusat. Kulit buah
manggis mempunyai permukaan yang licin dan bulat serta mempunyai warna yang
berubah mengikut peringkat pemasakan (daripada warna hijau kepada ungu gelap).
Kulit manggis mempunyai ketebalan antara 0.6-1.0 cm (Jabatan Pertanian Negeri Pulau
Pinang 2018). Selain itu, terdapat empat hingga lapan bekas kelopak stigma yang akan
kelihatan dengan jelas apabila manggis menghampiri peringkat akhir pemasakan.
Jumlah kelopak stigma ini menunjukkan jumlah isi di dalam setiap buah manggis
(Jabatan Pertanian Negeri Pulau Pinang 2018; Osman & Milan 2006). Isi buah manggis
adalah berwarna putih, bertekstur lembut dan berair serta mempunyai gabungan rasa
masam dan manis disertai aroma yang menyenangkan (Lan 1989; Thong et al. 2015).
Sesetengah isi manggis mempunyai biji yang ringan berwarna perang dan melekat
dengan isi buah manggis. Biji buah manggis yang matang tidak mempunyai
endosperma, rata dan hadir dalam pelbagai saiz yang kebanyakannya adalah
berketebalan 0.3-0.5 cm dan berukuran 1.0-1.5 cm (Lan 1989; Osman & Milan 2006).
Rajah 2.2 di bawah menunjukkan keratan rentas bagi buah manggis:
Rajah 2.2 Keratan rentas buah manggis
Selain daripada manggis, terdapat satu lagi klon Garcinia mangostana L. yang
didaftarkan oleh Jabatan Pertanian Malaysia iaitu mesta. Perbezaan ketara antara

14
manggis dengan mesta adalah mesta mempunyai bentuk yang bujur serta isi yang halus
dan kering tanpa kehadiran biji (Abu Dardak et al. 2011; Sandrang et al. 2015). Batang
pokok mesta pula lebih licin serta mempunyai cabang yang lebih rapat dan rendah
berbanding pokok manggis sementara daun mesta lebih panjang dan tirus. Skop
penanaman mesta di Malaysia adalah lebih kecil berbanding manggis dan
kebanyakannya boleh didapati di Temerloh, Maran dan Kuantan di negeri Pahang
(Sandrang et al. 2015). Selain itu, prospek pasaran manggis di peringkat domestik dan
antarabangsa juga lebih besar berbanding mesta. Justeru itu, manggis digunakan
sebagai organisma pilihan dalam kajian ini. Perbezaan antara buah manggis dengan
mesta dapat dilihat seperti dalam Jadual 2.1.
Jadual 2.1 Perbezaan buah manggis dengan mesta. Buah dituai dari plot tumbuhan
Universiti Kebangsaan Malaysia (UKM), Bangi, Selangor
Manggis Mesta
Pokok manggis disifatkan sebagai tumbuhan diesius, di mana pokok manggis
terbahagi kepada dua organ pembiakan iaitu “jantan” dan “betina”. Pokok manggis
yang ditanam secara penanaman biji benih mengambil masa selama 6-12 tahun untuk
berbuah bergantung kepada kaedah pengurusan, jenis tanah dan baka. Di Malaysia,
pokok manggis kebiasaannya menghasilkan buah sebanyak dua kali setahun, iaitu
antara bulan Jun-Ogos dan November-Januari (Osman & Milan 2006). Walau
bagaimanapun, tumbuhan bermusim seperti manggis ini boleh dipengaruhi oleh
perubahan pada cuaca persekitaran yang lembap dan kering, menyebabkan sesetengah
pokok hanya berbuah pada salah satu musim sahaja. Cuaca dan persekitaran yang sesuai
akan menggalakkan pendebungaan pokok manggis, seterusnya meningkatkan
penghasilan buah manggis (Apiratikorn et al. 2012; Lerslerwong et al. 2013). Musim
kering yang singkat juga diperlukan untuk merangsang dan memulakan proses antesis
pokok manggis (Osman & Milan 2006). Osman dan Milan (2006) turut melaporkan

15
bahawa julat suhu 25-35°C dan taburan hujan tahunan sebanyak 1200 mm atau lebih
disertai kelembapan yang tinggi tanpa musim kering yang berpanjangan adalah paling
optimum untuk proses antesis manggis.
2.2.1 Proses pemasakan manggis
Proses kematangan manggis umumnya adalah tidak seragam, kesan daripada proses
antesis yang berterusan pada pokok induk (Osman & Milan 2006). Proses kematangan
bermula apabila terdapat perubahan warna pada kulit manggis. Manggis memasuki fasa
kematangan bermula seawal minggu ke-14 selepas antesis dan mengambil masa kira-
kira selama dua minggu iaitu pada minggu ke-14 hingga minggu ke-15. Menurut Indeks
Kematangan Manggis Malaysia seperti yang dilaporkan oleh Osman dan Milan (2006),
proses pemasakan atau kematangan manggis terdiri daripada tujuh peringkat bermula
daripada muda semasa kulit luar manggis (perikarpa) berwarna hijau sehingga masak
di mana perikarpa manggis bertukar kepada ungu gelap. Perincian bagi indeks
kematangan manggis adalah seperti yang dinyatakan dalam Jadual 2.2.
Jadual 2.2 Indeks kematangan manggis di Malaysia
Indeks
kematangan
Tempoh selepas
antesis
Perincian
Peringkat 0
Minggu 12
Buah tidak matang dan berwarna hijau kekuningan.
Perikarpa, isi dan biji manggis belum dapat dipisahkan
sepenuhnya pada peringkat ini. Lapisan getah sangat tebal
pada kulit dalam buah.
Peringkat 1
Minggu 14
(purata 17 hari
selepas peringkat 0)
Buah sedang matang dan berwarna kuning atau hijau
pucat disertai titik merah jambu. Perikarpa, isi dan biji
mulai dapat dipisahkan dengan sempurna. Lapisan getah
tebal pada kulit dalam buah.
Peringkat 3
Minggu 14
(purata 3 hari
selepas peringkat 2)
Buah menghampiri kematangan dan berwarna merah
jambu yang sekata di keseluruhan perikarpa. Lapisan
getah nipis pada kulit dalam buah. Buah pada peringkat
ini sesuai untuk dituai bagi tujuan eksport.
bersambung...

16
...sambungan
Peringkat 4
Minggu 15
(purata 1-2 hari
selepas peringkat 3)
Buah matang dan berwarna merah atau merah
keperangan. Perikarpa, isi dan biji dapat dipisahkan
dengan mudah dan sempurna. Getah sangat nipis atau
tiada pada lapisan dalam kulit buah.
Peringkat 5
Minggu 15
(purata 1-2 hari
selepas peringkat 4)
Buah masak dan berwarna merah-ungu. Tiada kehadiran
getah pada kulit dalam buah. Pada peringkat ini, buah
sesuai dimakan sebagai buah segar.
Peringkat 6
Minggu 15
(purata 1 hari
selepas peringkat 5)
Buah masak dengan sempurna dan berwarna ungu, ungu
gelap atau hitam. Buah sesuai dimakan sebagai buah
segar tetapi tidak sesuai dieksport kerana mudah rosak.
Sumber: Diadaptasi daripada Osman dan Milan (2006)
Kondo et al. (2002) adalah sebahagian kumpulan penyelidik yang memulakan
kajian awal dalam merungkai mekanisme pemasakan dalam manggis. Kajian ini
memfokuskan kepada metabolisme asid absisik di dalam perikarpa, isi dan biji manggis
dan melaporkan kadar metabolisme yang berbeza pada peringkat perkembangan dan
pematangan manggis di dalam ketiga-tiga tisu ini. Kajian oleh Palapol et al. (2009a)
pula melaporkan faktor transkripsi MYB iaitu GmMYB10 memainkan peranan untuk
mengatur proses biosintesis antosianin dalam manggis semasa pemasakan berlaku.
Sementara itu, perkembangan warna dan kualiti manggis serta kadar penghasilan
hormon etilena dan kandungan antosianin pula dikaji oleh Palapol et al. (2009b) dengan
membandingkan enam peringkat pemasakan manggis yang berbeza. Kajian ini turut
melaporkan antosianin utama yang ditemui pada bahagian luar dan dalam perikarpa
manggis seperti sianidin-soforosida, sianidin-glukosida dan sianidin-
glukosidapentisida melalui analisis kromatografi cecair berprestasi tinggi/ spektrometri
jisim (HPLC/MS).

17
Terbaharu, Abdul-Rahman et al. (2017) mengkaji pemasakan manggis pada
peringkat RNA dengan membandingkan tiga peringkat pemasakan iaitu peringkat 0, 2
dan 6. Kajian ini membekalkan data yang penting untuk kajian-kajian pemprofilan
transkriptom pada masa akan datang. Selain itu, penglibatan metabolit utama seperti
asid 2-aminoisobutirik, pikosa dan beberapa asid amino (fenilanalina, valina dan
isoleusina) telah dilaporkan menunjukkan perubahan yang seiring dengan proses
pemasakan apabila dianalisis menggunakan kromatografi gas-spektrometri jisim (GC-
MS) (Parijadi et al. 2017). Namun, kajian secara komprehensif terhadap profil metabolit
di dalam manggis semasa pemasakan berlaku masih lagi terhad dan memerlukan kajian
yang lebih mendalam.
2.2.2 Manfaat dan bioaktiviti manggis
Manggis digelar sebagai “Permaisuri buah” ekoran daripada rasanya yang unik dan
enak (Fairchild 1915). Selain itu, manggis juga didapati mengandungi jumlah kalori
yang rendah dan sesuai dimakan begitu sahaja atau ditinkan, dibekukan atau diproses
menjadi jus minuman, kordial, jem atau gula-gula. Perikarpa manggis kebiasaannya
dicampurkan dengan bahan atau buah-buahan lain sebelum dijadikan produk seperti
jus, kapsul makanan tambahan, pencuci muka, atau sabun mandian. Penggunaan
manggis sebagai bahan semula jadi dalam pembuatan produk-produk kosmetik dan
kesihatan semakin meningkat dari semasa ke semasa.
Selain digelar sebagai “Permaisuri Buah”, manggis turut mendapat gelaran
“Buah Masa Hadapan” kerana potensinya untuk dikomersialkan pada masa hadapan
serta “Superfruit” kerana ciri-ciri perubatan yang terdapat pada buah ini (Failla &
Gutiérrez-Orozco 2017; Gutierrez-Orozco & Failla 2013; Saleem 2009). Buah manggis
terutamanya bahagian perikarpa telah digunakan sebagai ramuan dalam perubatan
tradisional oleh masyarakat setempat sejak zaman dahulu lagi kerana keberkesanannya
untuk mengubati pelbagai jenis penyakit termasuklah jangkitan luka, cirit-birit, disentri,
sakit perut, leukorea, gonorea dan ulser kronik (Osman & Milan 2006; Pedraza-
Chaverri et al. 2008; Yaacob & Tindall 1995). Dalam perubatan tradisional, perikarpa
manggis dikeringkan dan disapu pada luka. Bagi tujuan mengubati sakit perut dan

18
disentri pula, perikarpa manggis direbus dan dicampurkan dengan air, air limau atau air
beras sebelum diminum (Osman & Milan 2006; Yaacob & Tindall 1995).
Manggis terkenal dengan kehadiran metabolit bioaktif terutamanya xanton
(Mohamed et al. 2014; Nguyen et al. 2005) yang mempunyai ciri-ciri biologikal dan
farmaseutikal seperti anti-radang (Chen et al. 2008; Ibrahim et al. 2016), anti-malaria
(Chaijaroenkul et al. 2014), anti-alergi (Ibrahim et al. 2016; Nakatani et al. 2002), anti-
oksida (Mohamed et al. 2014; Thong et al. 2015), anti-kanser (Ibrahim et al. 2016;
Matsumoto et al. 2003; Shibata et al. 2011), anti-diabetes (Ibrahim et al. 2016) dan anti-
obesiti (Ibrahim et al. 2016; Liu et al. 2015). Pelbagai kajian telah dilakukan untuk
merungkai potensi manggis sebagai ramuan dalam penjagaan kesihatan melalui
penghasilan ubat-ubatan yang memfokuskan kepada xanton (Failla & Gutiérrez-Orozco
2017; Shan et al. 2011; Watanapokasin et al. 2010).
Kebanyakan xanton adalah terdiri daripada metabolit tidak-berkutub dan semi-
kutub (El-Seedi et al. 2009). Xanton mempunyai struktur kimia yang unik iaitu struktur
sistem aromatik trisiklik (C6-C3-C6). Kehadiran kumpulan fungsi isoprin, metoksil dan
hidroksil di pelbagai lokasi pada cincin A dan B (Rajah 2.3) menghasilkan struktur
xanton yang pelbagai. Xanton yang dipencilkan daripada sumber semula jadi boleh
dikategorikan kepada enam kumpulan iaitu xanton mudah, glikosida xanton, xanton
terprenil, xantonolignoid, bisxanton dan xanton pelbagai (El-Seedi et al. 2010; Vieira
& Kijjoa 2005).
Rajah 2.3 Struktur rangka asas xanton
Sumber: Pangkalan data ChemSpider

19
Sehingga tahun 2012, sebanyak 278 xanton dilaporkan telah ditemui daripada
sumber semula jadi iaitu di dalam 20 famili tumbuhan tinggi (Gentianaceae, Guttiferae,
Moraceae, Clusiaceae dan Polygalaceae) yang merangkumi 122 spesies dan 44 genera.
Selain itu, xanton juga turut dikenalpasti dan dipencilkan daripada 19 spesies kulat dan
tiga spesies liken (Negi et al. 2013; Vieira & Kijjoa 2005). Xanton daripada tumbuhan
tinggi paling banyak di dapati di dalam famili tumbuhan Clusiaceae (55 spesies dalam
12 genera termasuk manggis) dan Gentianaceae (28 spesies dalam 8 genera).
Terdapat sekurang-kurangnya sebanyak 68 xanton telah ditemui dan
dikenalpasti di beberapa bahagian pokok manggis yang berbeza termasuklah perikarpa
(Mohamed et al. 2014; Suksamrarn et al. 2003; Thong et al. 2015), isi (Suksamrarn et
al. 2003; Thong et al. 2015; Wittenauer et al. 2012), biji (Suksamrarn et al. 2003), daun
(Hamid et al. 2012; Parveen & Khan 1988), getah manggis yang berwarna kuning
(Sukatta et al. 2013) serta teras kayu atau batang pokok manggis (Harrison & Nilar
2002). Perikarpa manggis dilaporkan mempunyai kandungan xanton yang paling tinggi
dan terdapat sekurang-kurangnya sebanyak 50 xanton telah dikenalpasti di dalam
perikarpa manggis (El-Seedi et al. 2009; Pedraza-Chaverri et al. 2008). Antara xanton
utama yang ditemui di dalam manggis adalah mangostin (nama mangostin
kemudiannya ditukarkan kepada α-mangostin) yang mula ditemui pada tahun 1855
(Pedraza-Chaverri et al. 2008; Schmid 1855). Selain α-mangostin, β-mangostin, γ-
mangostin, gartanin dan garsinon E juga telah dilaporkan sebagai xanton utama di
dalam manggis (El-Seedi et al. 2010; El-Seedi et al. 2009; Shan et al. 2011; Sukatta et
al. 2013; Wittenauer et al. 2012) (Rajah 2.4).

20
Rajah 2.4 Xanton utama di dalam manggis
Sumber: Pangkalan data METLIN
Selain daripada xanton, manggis juga mengandungi sebatian bioaktif lain yang
terdiri daripada metabolit sekunder seperti tanin, antosianin, prosianidin dan fenolik
termasuklah asid fenolik, fenol dan flavonoid yang semakin menjadi tumpuan para
penyelidik pada masa kini. Pelbagai kajian yang memfokuskan kepada bioaktiviti bagi
metabolit sekunder telah dijalankan, antaranya, Pothitirat et al. (2009) telah
membandingkan aktiviti anti-oksida dan aktiviti bakteria pendorong anti-jerawat bagi
sebatian fenolik, flavonoid dan tanin daripada manggis muda dan matang. Selain itu,
terdapat juga kajian yang telah dijalankan untuk menentukan aktiviti anti-oksida bagi
sebatian prosianidin (Qin et al. 2017) dan sebatian fenolik serta flavonoid (Zarena &
Sankar 2012) daripada perikarpa manggis.
Kumpulan fenolik merupakan kumpulan metabolit sekunder yang paling
banyak didapati di dalam tumbuhan termasuklah buah-buahan (Dai & Mumper 2010;
Lattanzio et al. 2008). Metabolit di dalam kumpulan ini mempunyai struktur satu cincin
aromatik yang mengandungi satu atau lebih bahan ganti hidroksi, dan terhasil melalui
tapak jalan sikimat/fenilpropanoid atau tapak jalan “poliketida” asetat/malonat, atau
kedua-duanya (Lattanzio et al. 2008). Pengkelasan fenolik adalah rumit, namun fenolik
boleh diklasifikasikan kepada beberapa kumpulan berdasarkan tapak jalan biosintetik

21
dan ciri strukturnya. Antaranya adalah polifenol (flavonoid, flavonol, xanton), asid
fenolik, koumarin, fenolik glikosida, kuersetin, tanin dan lignin.
Fenolik kebiasaannya hadir dalam bentuk ester atau konjugat glikosida kepada
sebatian semula jadi yang lain seperti flavonoid, alkohol, sterol atau glukosida (Dai &
Mumper 2010; Lattanzio et al. 2008). Zarena dan Sankar (2012) melaporkan beberapa
fenolik utama di dalam ekstrak perikarpa manggis yang berbeza. Asid kafeik, asid t-
sinamik, asid vanilik, asid sinapik dan asid siringik dikenalpasti sebagai asid fenolik
utama dalam ekstrak asid fenolik berasid terhidrolisis manakala asid galik pula adalah
asid fenolik utama di dalam ekstrak asid fenolik beralkali terhidrolisis. Selain itu,
ekstrak asid fenolik bebas dan ekstrak asid fenolik berasid terhidrolisis turut
menunjukkan aktiviti antioksida yang memberangsangkan berdasarkan analisis asai
DPPH yang dijalankan (Zarena & Sankar 2012).
Aktiviti anti-oksida fenolik adalah penting untuk melindungi tumbuhan
daripada sinaran ultra ungu (UV) yang merbahaya. Radiasi sinaran UV boleh
merangsang penghasilan spesies oksigen reaktif (ROS) yang boleh mempengaruhi
DNA, protein dan membran, seterusnya menyebabkan berlakunya perubahan pada
metabolisme tumbuhan (Lattanzio et al. 2008). Selain daripada berfungsi sebagai anti-
oksida, fenolik turut dilaporkan mempunyai aktiviti anti-mutagenik, anti-proliperatif
dan anti-mikrob (Kampa et al. 2004; Zarena & Sankar 2012). Selain itu, sesetengah
kumpulan fenolik seperti flavonoid, terutamanya klorofil dan antosianin berperanan
dalam perkembangan warna dan rasa tumbuhan, termasuklah buah-buahan (Zarena &
Sankar 2012).
Flavonoid dikategorikan sebagai polifenol dan dikelaskan berdasarkan
kehadiran rangka struktur difenilpropana (C6C3C6). Kelas utama bagi flavonoid adalah
flavon, flavanon, flavonol, isoflavonoid, antosianin dan kalkon (Panche et al. 2016).
Flavonoid merangkumi lebih daripada 4000 metabolit sekunder yang turut hadir
sebagai konjugat kepada gula (Miean & Mohamed 2001; Rhodes & Price 1996) di
dalam tumbuhan. Katesin yang dijumpai di dalam ekstrak asid fenolik bebas dan
kuersetin di dalam ekstrak asid fenolik berasid terhidrolisis telah dilaporkan sebagai
flavonoid utama di dalam perikarpa manggis (Zarena & Sankar 2012).

22
Terpenoid (atau juga dipanggil sebagai isoprenoid) adalah kumpulan metabolit
yang mempunyai struktur isoprena dan dikategorikan mengikut rangka karbon seperti
yang diringkaskan di dalam jadual di bawah (Jadual 2.3). Kumpulan terpenoid
menjalankan pelbagai fungsi penting kepada tumbuhan (Cheng et al. 2007; Grassmann
2005). Selain daripada fenolik, terpenoid juga merupakan kumpulan utama yang
bertindak sebagai anti-oksida tumbuhan (Grassmann 2005).
Jadual 2.3 Klasifikasi terpenoid
Kelas terpenoid Bilangan atom
karbon
Bilangan subunit
isoprena
Formula
molekul
Contoh terpenoid
Monoterpenoid 10 2 C10H16 Mentol
Timol
Seskuiterpenoid 15 3 C15H24 Asid absisik
Farnesol
Diterpenoid 20 4 C20H32 Karnosol
Asid giberelik
Triterpenoid 30 6 C30H48 Asid oleanolik
Asid ursolik
Tetraterpenoid 40 8 C40H64 Karotin
Kriptoxantin
Politerpenoid > 40 > 8 (C5H8)n Getah
Sumber: Grassmann (2005)
Monoterpenoid dan diterpenoid dikenalpasti sebagai komponen utama di dalam
pati minyak tumbuhan dan memainkan peranan sebagai agen alelopati. Kumpulan
metabolit ini terlibat dalam interaksi antara satu tumbuhan dengan tumbuhan yang lain
serta tumbuhan-patogen/herbivor (Cheng et al. 2007; Grassmann 2005; Zwenger &
Basu 2008). Karetenoid di bawah kumpulan tetraterpenoid pula berfungsi sebagai
aksesori pigmen untuk menyerap cahaya matahari dan dalam masa yang sama
membekalkan perlindungan kepada tumbuhan daripada kerosakan molekular yang
disebabkan oleh cahaya matahari. Karetenoid juga membekalkan pigmentasi kepada
tumbuhan dan mempunyai fungsi yang sama seperti flavonoid (klorofil dan antosianin)
dalam perkembangan warna dan rasa tumbuhan serta buah-buahan.
Terpenoid digunakan secara meluas sebagai bahan perasa tambahan, wangian,
farmaseutikal dan racun serangga (Tholl 2015). Meskipun skop kajian terhadap
terpenoid di dalam manggis masih lagi kecil, namun terdapat beberapa kajian kualitatif
dan kuantitatif yang telah melaporkan kehadiran kumpulan metabolit ini di dalam
beberapa bahagian manggis. Kehadiran skualina dan vitamin E telah dilaporkan oleh

23
Hapsari et al. (2018) di dalam biji manggis manakala terpenoid lain iaitu si-kar-otenal
telah dikenalpasti di dalam tisu yang sama semasa biji manggis mengalami fasa
percambahan (Mazlan et al. 2019). Selain itu, terpenoid juga dilaporkan hadir di dalam
perikarpa (Adnyana et al. 2016; Tjitrosemito & Poerwanto 2008), batang pokok, getah
dan isi manggis (Tjitrosemito & Poerwanto 2008).
Alkaloid pula adalah kumpulan bagi pelbagai metabolit yang mempunyai berat
molekul yang rendah serta dikelaskan berdasarkan kehadiran atom nitrogen asas
(Kabera et al. 2014; Zulak et al. 2006). Alkaloid dijumpai di dalam 20% daripada
spesies tumbuhan termasuklah tumbuhan daripada famili Apocyanaceae,
Ranunculaceae, Papaveraceae, Solanaceae dan Rutaceae. Alkaloid boleh didapati
daripada pelbagai bahagian pokok yang berbeza daripada famili tumbuhan ini,
antaranya seperti nikotin pada daun, kinkonina dan kuinina pada batang, nibidina pada
biji dan rawelfinina pada akar pokok (Wansi et al. 2013). Alkaloid kebanyakannya
terhasil daripada asid amino triptofan, fenilalanina, tirosina dan asid antranilik melalui
tapak jalan sikimat (Klaser Cheng 2017).
Seperti fenolik dan terpenoid, alkaloid juga mempunyai pelbagai aktiviti biologi
dan farmakologi dan digunakan secara meluas di dalam bidang perubatan (Croteau et
al. 2000; Kabera et al. 2014) serta sebagai perangsang, narkotik dan racun seperti
kafein, morfin, nikotin, dan kokain (Zulak et al. 2006). Sementara itu, lain-lain alkaloid
seperti anopterina, alpinina dan 1,2-dimetoksi-13-metil-[1,3]benzodioksolo[5,6-
c]fenantridina (DMMBP) dipercayai terlibat dalam proses percambahan biji benih
manggis dengan memainkan peranan dalam pertahanan daripada serangan herbivor dan
serangga (Mazlan et al. 2019). Selain itu, alkaloid turut dilaporkan menyumbang kepada
aktiviti anti-oksida dalam perikarpa manggis (Sinaga & Siregar 2016).
2.2.3 Potensi manggis di pasaran
Manggis menjadi tumpuan di beberapa negara termasuklah China-Taiwan, Jepun,
Australia dan Eropah ekoran daripada rasanya yang unik. Manggis mula terkenal di
Barat Eropah dan Amerika Syarikat sejak abad ke-20 ekoran daripada kesan positifnya
terhadap kesihatan manusia (Brunner & Morales-Payan 2010). Negara-negara Asia
menjadi pengeluar utama kepada pasaran manggis di Eropah. Abu Dardak et al. (2011)

24
turut melaporkan permintaan daripada negara-negara Timur Tengah juga semakin
meningkat. Pasaran antarabangsa berkembang dari semasa ke semasa, terutamanya
negara China yang menganggap manggis sebagai makanan “sejuk” di dalam konsep
budaya yin-yang yang diamalkan penduduk di negara tersebut. Manggis juga dikatakan
pelengkap kepada buah durian, di mana manggis bertindak sebagai “penyejuk” kepada
buah durian yang bersifat “panas” (Osman & Milan 2006). Peningkatan kadar
permintaan manggis segera dan produk-produk berasaskan manggis dianggarkan
bernilai lebih RM50 juta setahun di pasaran domestik dan RM580 juta setahun di
pasaran antarabangsa (Abu Dardak et al. 2011). Namun, kadar pengeksportan manggis
ke negara luar telah dilaporkan berkurang sebanyak 63.6% pada tahun 2015 berbanding
tahun sebelumnya iaitu tahun 2014 (Lembaga Pemasaran Pertanian Persekutuan 2017)
ekoran daripada persaingan yang tinggi daripada negara jiran seperti Thailand dan
Indonesia.
Bagi tujuan eksport ke pasaran domestik dan antarabangsa, manggis biasanya
dieksport sebagai buah segar atau telah diproses di dalam bentuk sejuk beku. Buah
manggis mempunyai jangka hayat lepas tuai yang pendek iaitu selama dua minggu
(Osman & Milan 2006). Bagi buah segar, manggis disimpan di dalam suhu sejuk di
bawah 18 °C untuk meningkatkan daya tahan (sehingga 21 hari) dari kerosakan semasa
proses pengangkutan (Abu Dardak et al. 2011). Bagi tujuan eksport buah manggis
segar, pelbagai kajian telah dijalankan untuk memanjangkan tempoh penuaian,
meningkatkan jangka hayat lepas tuai manggis serta meningkatkan hasil penanaman
manggis. Antaranya, kajian oleh Lerslerwong et al. (2013) melaporkan bahawa
penggunaan asid 2-kloroetanafosfonik (etefon) boleh mempercepatkan proses
pemasakan manggis selama satu minggu jika manggis yang masih berada pada pokok
induk dirawat dengan etefon apabila menghampiri tempoh penuaian (Lerslerwong et al.
2013). Etefon adalah sejenis sebatian yang bertindak menggalakkan perembesan etilena
oleh buah, terutamanya buah klimakterik seperti manggis (Lerslerwong et al. 2013; Li
et al. 2017). Hormon ini akan bertindak mengawal pelbagai proses pertumbuhan,
perkembangan dan senesens tumbuhan termasuklah pemasakan buah selain tindak balas
tumbuhan terhadap alam sekitar (Abeles et al. 2012; Lieberman 1979; Yang & Hoffman
1984).

25
Lerslerwong et al. (2013) juga melaporkan bahawa 1-metilsiklopropena (1-
MCP) boleh melambatkan proses pemasakan manggis jika kaedah yang sama
dikenakan iaitu dengan cara merawat buah yang masih berada pada pokok induk dengan
1-MCP apabila buah menghampiri tempoh penuaian. Sebatian 1-MCP bertindak
sebagai perencat kepada hormon etilena dan telah terbukti boleh melambatkan proses
pemasakan dan dalam masa yang sama mengekalkan kualiti pelbagai jenis tanaman
(Blankenship & Dole 2003; Watkins 2008). Selain itu, satu kajian oleh penyelidik di
Indonesia iaitu Poerwanto et al. (2006) melaporkan bahawa jangka hayat selepas tuai
manggis boleh dipanjangkan kepada 40 hari jika manggis disalut dengan kitosan dan
dibungkus dengan plastik filem polietilena serta disimpan pada suhu 15 °C. Kitosan
adalah metabolit terbitan kepada kitin dan merupakan sebatian anti-mikrob semulajadi
yang banyak digunakan untuk tujuan pengawetan buah-buahan dan sayur-sayuran
(Devlieghere et al. 2004; Jianglian & Shaoying 2013).
Sementara itu, di Malaysia, Institut Penyelidikan dan Kemajuan Pertanian
Malaysia (MARDI) telah memperkenalkan anak benih “Advance Planting Material”
yang mampu mempercepatkan pengeluaran hasil tanaman manggis sekaligus
memendekkan tempoh pulangan modal pelaburan bagi pengusaha tanaman (Abu
Dardak et al. 2011). Namun begitu, kajian terhadap mekanisme semasa pemasakan
buah manggis yang kompleks masih lagi kurang. Pemasakan buah adalah antara faktor
utama yang menyumbang kepada perubahan dalam komposisi buah. Pemahaman
terhadap pemasakan manggis boleh menyumbang kepada eksploitasi potensi manggis
yang lebih luas sama ada sebagai buah segar atau penghasilan produk-produk berasakan
manggis, sekaligus meningkatkan nilai ekonomi buah tropika ini setanding dengan
negara pengeluar utama seperti Thailand dan Indonesia.
2.3 PENDEKATAN BIOLOGI SISTEM BERASASKAN METABOLOMIK
Pendekatan biologi sistem telah digunakan sebagai platform untuk merungkai
pemasakan manggis dalam kajian ini. Biologi sistem didefinisikan sebagai kajian
saintifik mengenai ciri sistematik dan interaksi dinamik sesuatu komponen biologi,
sama ada molekul, sel, organisma atau keseluruhan spesies secara kualitatif mahupun
kuantitatif (Klipp et al. 2016). Definisi “sistem” dalam aspek “biologi sistem” pula

26
adalah satu set bahagian yang berhubung dan membentuk satu keseluruhan yang
kompleks. Sistem hidupan adalah dinamik dan kompleks serta tingkah laku mereka
sukar untuk diramal jika hanya merujuk kepada beberapa bahagian tertentu sahaja. Bagi
memahami biologi pada peringkat sistem, struktur dan dinamik bagi fungsi selular dan
fungsi organisma haruslah dikaji dan bukan hanya tertumpu kepada komponen terpencil
sesuatu sel atau organisma (Kitano 2002).
Antara pendekatan yang seringkali digunakan dalam biologi sistem adalah
genomik, preoteomik, transkriptomik, metabolomik dan glikomik serta disokong
dengan kajian bioinformatik (Aizat et al. 2018; Zierer et al. 2015). Genomik adalah
kajian terhadap gen, proteomik dijalankan untuk mengkaji aras dan identiti protein dan
transkriptomik pula adalah kajian pada peringkat RNA manakala metabolomik
memfokuskan kepada pengenalpastian metabolit di dalam sesuatu sampel biologi
(Aizat et al. 2018; Zierer et al. 2015). Pendekatan metabolomik telah digunakan dalam
kajian ini bagi merungkai mekanisme pemasakan buah manggis (Garcinia mangostana
Linn.) pada pelbagai peringkat pemasakan dengan cara melihat kepada komposisi
metabolit di dalam jenis tisu manggis yang berbeza. Oleh itu, kaedah metabolomik akan
dibincangkan dengan lebih terperinci dalam penerangan seterusnya.
Metabolomik adalah antara kaedah “omik” yang terbaharu di dalam bidang
kajian sistem biologi (Ryan & Robards 2006). Metabolomik adalah satu kajian saintifik
mengenai profil metabolit yang hadir di dalam sel, tisu atau organisma (Wei et al. 2018).
Metabolit merupakan molekul kecil (50-1500 Dalton) yang mengalami perubahan
kimia semasa proses metabolisme, manakala metabolom pula didefinisikan sebagai
metabolit jumlah yang hadir di dalam sel, tisu atau organisma (Aretz & Meierhofer
2016; Patti et al. 2012; Zhang et al. 2016). Metabolit dianggap sebagai gambaran
langsung kepada keadaan selular di dalam sesuatu organisma (Patti et al. 2012). Berbeza
dengan gen dan protein, metabolit lebih menggambarkan fenotip serta aktiviti biokimia
sesuatu organisma (Baharum & Azizan 2018; Patti et al. 2012). Hal ini kerana fungsi
gen boleh berubah kerana dipengaruhi oleh pengawalaturan epigenetik, manakala
protein pula akan berubah apabila mengalami modifikasi semasa proses pasca-translasi
(Aretz & Meierhofer 2016; Patti et al. 2012).

27
Analisis metabolomik terbahagi kepada dua iaitu analisis sasaran dan bukan
sasaran (Roberts et al. 2012; Shulaev 2006). Analisis sasaran sesuai digunakan untuk
mengukur kepekatan metabolit yang disasarkan dengan tepat atau mengkaji struktur
metabolit tersebut dengan lebih terperinci (Bingol 2018; Roberts et al. 2012). Maklumat
asas mengenai metabolit yang ingin disasarkan seperti struktur, berat molekul ataupun
kumpulan fungsi diperlukan sebagai panduan untuk menjalankan analisis metabolomik
sasaran termasuklah bagi menentukan teknik dan parameter sistem analitikal yang akan
digunakan serta pengenalpastian metabolit melalui pencarian dalam pangkalan data
metabolomik. Penggunaan piawai dalaman pula membolehkan analisis dijalankan
secara kuantitatif atau semi-kuantitatif (Roberts et al. 2012). Namun, limitasi bagi
analisis sasaran ini adalah metabolit yang ingin dikaji haruslah diekstrak dan hadir
dalam bentuk asli serta berkepekatan tinggi. Oleh itu, kaedah ini tidak sesuai digunakan
untuk mengkaji penanda metabolit yang unik atau perubahan metabolit secara global
(Shulaev 2006).
Sementara itu, analisis bukan sasaran atau turut dikenali sebagai analisis cap jari
metabolik tidak memfokuskan kepada identifikasi atau pengukuran metabolit secara
spesifik (Bingol 2018; De Vos et al. 2007). Sebaliknya, analisis bukan sasaran adalah
satu analisis komprehensif yang memfokuskan kepada kesemua metabolit di dalam
sesuatu sampel biologi termasuklah metabolit yang tidak diketahui atau belum pernah
ditemui (Roberts et al. 2012). Analisis ini digunakan untuk mengenalpasti jumlah profil
atau corak unik perubahan metabolit di dalam sesuatu sampel biologi yang mencirikan
sesuatu sel atau tisu (De Vos et al. 2007; Shulaev 2006) serta untuk pengesanan dan
diagnostik penanda biologi. Pelbagai teknik analitikal boleh digunakan untuk
menjalankan analisis metabolomik seperti nuklear magnetik resonans (NMR),
kromatografi gas-spektrometri jisim (GC-MS), kromatografi cecair (LC-MS) dan
elektroforesis kapilari (CE-MS) (Bingol 2018). Secara amnya, analisis metabolomik
terbahagi kepada beberapa bahagian atau langkah utama iaitu penyediaan dan
pengekstrakan sampel, analisis sampel menggunakan kaedah analitikal (pemisahan dan
pengesanan metabolit) dan analisis data (Cevallos-Cevallos et al. 2009; Kim &
Verpoorte 2010).

28
Kajian lepas menggunakan pendekatan metabolomik terhadap manggis banyak
memfokuskan kepada pengenalpastian dan pengukuran tahap sebatian bioaktif
terutamanya xanton (Sugita et al. 2017; Sukatta et al. 2013; Zarena & Sankar 2009;
Zhou et al. 2008). Terdapat juga kajian metabolomik telah dilakukan terhadap
organisma tertentu yang telah didedahkan terhadap ekstrak manggis bagi mengkaji
aktiviti anti-malaria manggis seperti yang telah dilakukan terhadap parasit manusia,
Plasmodium falciparum (Chaijaroenkul et al. 2014). Selain itu, pemprofilan asid fenolik
manggis turut dijalankan oleh Zadernowski et al. (2009) dengan membandingkan tisu
manggis yang berbeza iaitu kulit luar, kulit dalam dan isi manggis. Beberapa tahun
selepas itu, Zarena & Sankar (2012) pula memprofil metabolit asid fenolik dan
flavonoid di dalam perikarpa manggis menggunakan kaedah pengekstrakan yang
berbeza.
Terbaharu, kajian terhadap komposisi metabolit semasa perkembangan biji
manggis turut dijalankan oleh Mazlan et al. (2018a) dan Mazlan et al. (2018b)
manakala beberapa kajian untuk merungkai pemasakan manggis telah dijalankan oleh
Parijadi et al. (2017) dan Gondokesumo et al. (2019) dengan membandingkan peringkat
pemasakan manggis yang berbeza. Walaubagaimanapun, kajian oleh Parijadi et al.
(2017) yang membandingkan tisu manggis yang berbeza pada tujuh peringkat
pemasakan hanya melibatkan penggunaan satu teknik analitikal iaitu GC-MS manakala
kajian oleh Gondokesumo et al. (2019) pula memfokuskan kepada metabolit xanton di
dalam tisu perikarpa dan bukan keseluruhan metabolom manggis.
2.3.1 Penyediaan dan pengekstrakan sampel
Penyediaan sampel adalah satu langkah yang kritikal di dalam setiap analisis
metabolomik dan perlu dijalankan menggunakan teknik yang betul (Kim & Verpoorte
2010). Proses ini akan mempengaruhi sebatian atau metabolit yang akan dipisahkan dan
dikenalpasti melalui analisis analitikal serta menentukan ketepatan hasil dan tahap
sensitiviti bagi keseluruhan analisis metabolomik yang akan dijalankan (Kim &
Verpoorte 2010; Mushtaq et al. 2014). Penyediaan sampel merangkumi beberapa tahap
iaitu pengumpulan sampel (atau penuaian bagi sampel buah), pengeringan dan diikuti
dengan pengekstrakan sampel. Sampel pepejal seperti tisu tumbuhan atau buah (daun,

29
kulit, biji dan sebagainya) biasanya dikisar menjadi serbuk halus bagi meningkatkan
kadar penyerapan pelarut, seterusnya menggalakkan pembebasan metabolit oleh sel
semasa proses pengekstrakan (Cevallos-Cevallos et al. 2009). Cecair nitrogen (-190°C)
sering digunakan semasa proses pengisaran sampel bagi menghentikan semua aktiviti
enzim serta metabolisme yang berlaku dalam sampel biologi kerana aktiviti ini akan
mengubah profil metabolit asal sampel biologi tersebut (Abdel-Farid et al. 2007;
Mushtaq et al. 2014).
Seterusnya, sampel yang telah dikisar akan dikeringkan menggunakan beberapa
teknik seperti pengeringan menggunakan ketuhar, ketuhar gelombang mikro, udara
panas (Harbourne et al. 2009) atau pengeringan sejuk-beku (Mushtaq et al. 2014) bagi
membuang kandungan air di dalam sampel. Proses pengeringan sampel sangat penting
bagi memudahkan penyimpanan sampel untuk jangka masa panjang kerana proses ini
akan menghalang atau mengurangkan sebarang aktiviti enzim serta pertumbuhan
mikrob pada sampel. Selain itu, perbezaan kandungan kelembapan (kandungan air) di
dalam sampel juga boleh mengganggu proses analisis menggunakan instrumen
analitikal seperti GC-MS, LC-MS dan NMR. Kandungan air akan menyebabkan
resolusi spektrum NMR yang terhasil akan berkurang serta kadar kebolehulangan bagi
analisis GC-MS menurun (Mushtaq et al. 2014; Rangarajan & Ghosh 2011).
Pengeringan sejuk-beku dilaporkan mampu memberi hasil pengeringan yang
paling bagus berbanding teknik-teknik pengeringan yang lain (Mushtaq et al. 2014).
Bagi teknik pengeringan sejuk-beku, sampel harus disejukkan menggunakan cecair
nitrogen terlebih dahulu sebelum dimasukkan ke dalam ruang khas yang mempunyai
suhu dan tekanan yang rendah supaya proses pengeringan boleh berlaku. Teknik ini
sangat sesuai digunakan bagi sampel yang sensitif terhadap suhu yang tinggi. Sampel
yang telah dikeringkan haruslah disimpan pada suhu -80 °C atau di dalam cecair
nitrogen bagi mengelakkan berlakunya penyerapan semula air daripada persekitaran
(Mushtaq et al. 2014).
Selain daripada proses penyediaan sampel, proses pengekstrakan juga adalah
langkah penting dalam analisis metabolomik kerana langkah ini akan menentukan
jumlah metabolit dan jenis metabolit yang akan diekstrak daripada sampel (Cevallos-

30
Cevallos et al. 2009; Drouin et al. 2018). Terdapat dua faktor yang mempengaruhi
proses pengekstrakan iaitu faktor kimia dan faktor fizikal (Kim & Verpoorte 2010;
Mushtaq et al. 2014). Faktor kimia seperti jenis pelarut, tahap kekutuban serta nisbah
pelarut ditentukan berdasarkan tujuan analisis metabolomik yang dijalankan.
Bagi analisis metabolomik sasaran, pemilihan pelarut adalah bergantung kepada
jenis metabolit yang ingin diekstrak (Cevallos-Cevallos et al. 2009). Disebabkan
kehadiran metabolit yang pelbagai di dalam sampel biologi, penggunaan hanya satu
jenis pelarut adalah mustahil untuk mengekstrak semua jenis metabolit. Oleh itu, bagi
analisis bukan sasaran atau analisis metabolomik global, gabungan pelarut seringkali
digunakan untuk memaksimumkan hasil yang bakal diperoleh (Kim & Verpoorte 2010;
Mushtaq et al. 2014). Pelarut umum seperti air sesuai digunakan untuk mengekstrak
pelbagai jenis metabolit berkutub termasuklah gula. Pelarut tidak berkutub seperti
kloroform pula sesuai digunakan untuk mengekstrak metabolit tidak berkutub.
Gabungan pelbagai pelarut seperti air-metanol-kloroform dengan nisbah yang berbeza
sering digunakan dalam kajian metabolomik pada masa kini untuk meningkatkan
kepelbagaian jenis metabolit yang merangkumi metabolit tidak berkutub, metabolit
yang berkutub tinggi serta metabolit hidrofilik dan hidrofobik (Mushtaq et al. 2014).
Faktor fizikal seperti teknik sonikasi atau gelombang ketuhar turut digunakan
untuk meningkatkan hasil pengekstrakan metabolit (Cevallos-Cevallos et al. 2009; Das
& Eun 2018; Mushtaq et al. 2014). Teknik ini dijalankan bertujuan untuk meningkatkan
kadar pembebasan metabolit daripada sel (Cevallos-Cevallos et al. 2009; Das & Eun
2018). Teknik sonikasi melibatkan pendedahan sampel terhadap nadi atau gelombang
bunyi berfrekuensi tinggi untuk memecahkan sel manakala teknik gelombang ketuhar
pula digunakan untuk memanaskan pelarut melebihi tekanan takat didih tekanan
atmosfera pelarut tersebut bagi meningkatkan kadar penyerapan pelarut ke dalam
sampel (Kaufmann & Christen 2002; Vinatoru 2001).

31
2.3.2 Analisis sampel menggunakan teknik analitikal
a. Teknik analitikal dalam kajian pemasakan buah
Teknik analitikal bagi kajian metabolomik terbahagi kepada dua pengkelasan utama
iaitu resonans magnetik nuklear (NMR) dan spektrometri jisim (MS) (Cambiaghi et al.
2017; De Vos et al. 2007; Theodoridis et al. 2012a). Kedua-dua teknik ini telah
dibuktikan keberkesanannya melalui kajian-kajian lepas untuk mengkaji tentang
pemasakan buah seperti yang disenaraikan di dalam Jadual 2.4. Sistem NMR
menawarkan pengenalpastian dan kuantifikasi metabolit secara langsung, terutamanya
metabolit yang hadir dalam kepekatan yang tinggi di dalam sesuatu sampel biologi
(Theodoridis et al. 2012b). Berbanding sistem MS, sistem NMR adalah lebih kukuh dan
mempunyai kadar kebolehulangan yang lebih tinggi. Sistem NMR membenarkan
pengenalpastian struktur sesuatu molekul dengan tepat dan berkesan (Theodoridis et al.
2012b). Namun, seperti yang dinyatakan sebelum ini, NMR hanya boleh mengenalpasti
metabolit yang berkepekatan tinggi di dalam sesuatu sampel (Cambiaghi et al. 2017;
Theodoridis et al. 2012b) serta melibatkan kos permulaan pemasangan alat yang tinggi
(Theodoridis et al. 2012b).
Sistem MS pula beroperasi dengan cara menukarkan molekul analit kepada
keadaan bercaj (ionised). Ion dan fragmen ion yang terhasil semasa proses pegionan
akan dianalisis berdasarkan nisbah jisim terhadap caj (mass to charge ratio) atau nilai
m/z (Metz et al. 2017; Pitt 2009; Theodoridis et al. 2012b). MS yang digabungkan
dengan sistem pemisahan seperti GC, LC dan CE berperanan penting dan seringkali
digunakan bagi analisis pemprofilan metabolit secara holistik ekoran daripada tahap
sensitiviti sistem ini yang tinggi serta boleh didapati secara meluas (Metz et al. 2017).
Sistem MS mempunyai tahap sensitiviti yang lebih tinggi berbanding sistem NMR
(Cambiaghi et al. 2017; Theodoridis et al. 2012b) kerana sistem ini berkeupayaan untuk
mengenalpasti metabolit yang hadir dalam kepekatan yang rendah di dalam sesuatu
sampel biologi yang tidak dapat dikesan melalui sistem NMR (De Vos et al. 2007;
Theodoridis et al. 2012a).
Bilangan sistem MS yang digunakan di seluruh dunia juga lebih tinggi
berbanding sistem NMR berikutan daripada kos pemasangan yang lebih rendah. Oleh

32
itu, bilangan pengguna serta pakar dalam penggunaan MS yang berkemampuan untuk
membekalkan serta mengendalikan data metabolomik juga adalah lebih tinggi. Justeru
itu, sistem MS semakin banyak digunakan bagi tujuan pemprofilan metabolik serta
pengenalpastian metabolit sasaran terutamanya sejak beberapa tahun kebelakangan ini
(Griffiths et al. 2010; Theodoridis et al. 2012b). Dalam kajian ini, sistem GC dan LC
yang dilengkapi dengan sistem MS telah digunakan untuk memprofil metabolit
daripada manggis.
.
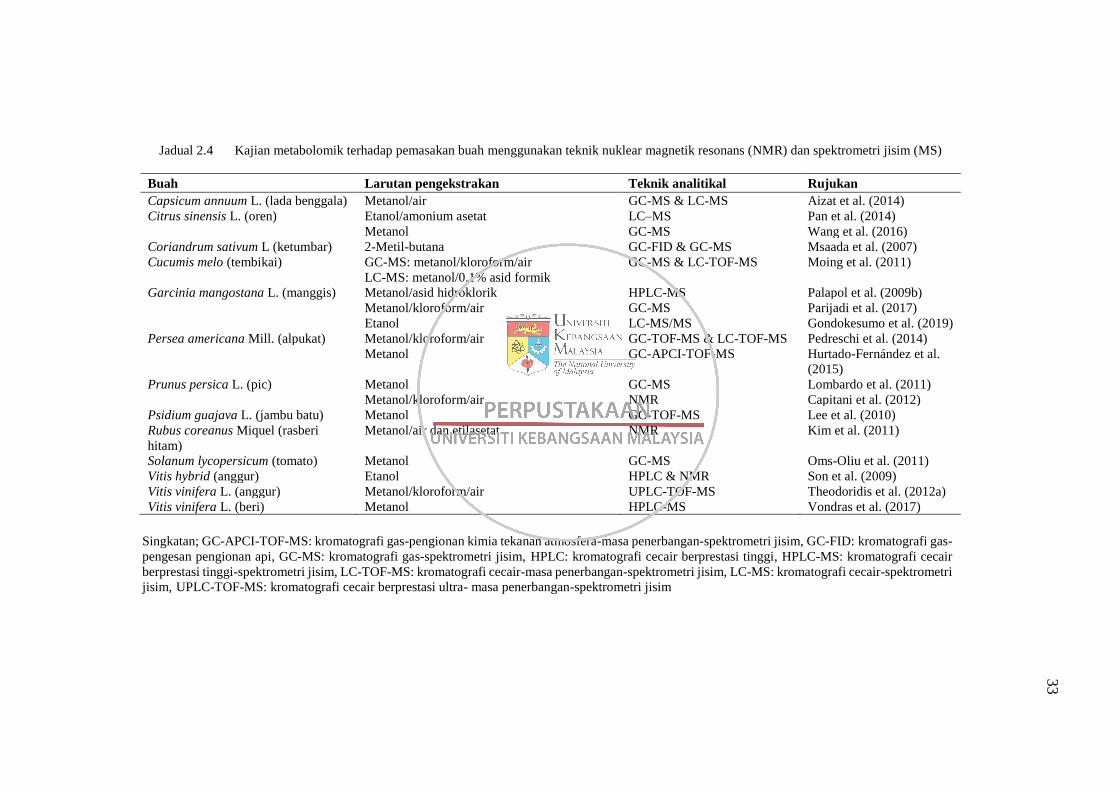
33
Jadual 2.4 Kajian metabolomik terhadap pemasakan buah menggunakan teknik nuklear magnetik resonans (NMR) dan spektrometri jisim (MS)
Buah Larutan pengekstrakan Teknik analitikal Rujukan
Capsicum annuum L. (lada benggala) Metanol/air GC-MS & LC-MS Aizat et al. (2014)
Citrus sinensis L. (oren) Etanol/amonium asetat LC–MS Pan et al. (2014)
Metanol GC-MS Wang et al. (2016)
Coriandrum sativum L (ketumbar) 2-Metil-butana GC-FID & GC-MS Msaada et al. (2007)
Cucumis melo (tembikai) GC-MS: metanol/kloroform/air
LC-MS: metanol/0.1% asid formik
GC-MS & LC-TOF-MS Moing et al. (2011)
Garcinia mangostana L. (manggis) Metanol/asid hidroklorik HPLC-MS Palapol et al. (2009b)
Metanol/kloroform/air GC-MS Parijadi et al. (2017)
Etanol LC-MS/MS Gondokesumo et al. (2019)
Persea americana Mill. (alpukat) Metanol/kloroform/air GC-TOF-MS & LC-TOF-MS Pedreschi et al. (2014)
Metanol GC-APCI-TOF-MS Hurtado-Fernández et al.
(2015)
Prunus persica L. (pic) Metanol GC-MS Lombardo et al. (2011)
Metanol/kloroform/air NMR Capitani et al. (2012)
Psidium guajava L. (jambu batu) Metanol GC-TOF-MS Lee et al. (2010)
Rubus coreanus Miquel (rasberi
hitam)
Metanol/air dan etilasetat NMR Kim et al. (2011)
Solanum lycopersicum (tomato) Metanol GC-MS Oms-Oliu et al. (2011)
Vitis hybrid (anggur) Etanol HPLC & NMR Son et al. (2009)
Vitis vinifera L. (anggur) Metanol/kloroform/air UPLC-TOF-MS Theodoridis et al. (2012a)
Vitis vinifera L. (beri) Metanol HPLC-MS Vondras et al. (2017)
Singkatan; GC-APCI-TOF-MS: kromatografi gas-pengionan kimia tekanan atmosfera-masa penerbangan-spektrometri jisim, GC-FID: kromatografi gas-
pengesan pengionan api, GC-MS: kromatografi gas-spektrometri jisim, HPLC: kromatografi cecair berprestasi tinggi, HPLC-MS: kromatografi cecair
berprestasi tinggi-spektrometri jisim, LC-TOF-MS: kromatografi cecair-masa penerbangan-spektrometri jisim, LC-MS: kromatografi cecair-spektrometri
jisim, UPLC-TOF-MS: kromatografi cecair berprestasi ultra- masa penerbangan-spektrometri jisim

34
b. Kromatografi gas-spektrometri jisim (GC-MS) dan kromatografi cecair-
spektrometri jisim (LC-MS)
Sehingga kini, kajian metabolomik yang dilakukan terhadap pemasakan manggis adalah
tidak bersifat komprehensif dan hanya memfokuskan kepada salah satu platform analitikal
sahaja (GC-MS atau LC-MS) atau kumpulan metabolit tertentu terutamanya xanton dan
fenolik sahaja (Gondokesumo et al. 2019; Ji et al. 2007; Parijadi et al. 2017; Yang et al.
2017; Zarena et al. 2012; Zarena & Sankar 2009). Dalam kajian ini, kedua-dua analisis GC-
MS dan LC-MS telah digunakan bagi memaksimumkan pengenalpastian jumlah metabolit
yang hadir dalam sampel manggis (perikarpa, isi dan biji) pada peringkat pemasakan yang
berbeza (peringkat 0, 2, 4 dan 6) merangkumi pelbagai jenis metabolit termasuklah
metabolit tidak berkutub sehingga berkutub, hidrofilik dan hidrofobik.
GC-MS menawarkan kaedah pemisahan sebatian yang sangat berkesan disertai
tahap sensitiviti, kebolehulangan dan resolusi yang tinggi, menjadikan sistem ini sangat
sesuai digunakan untuk menganalisis sampel biologi yang kompleks (Niessen 1999;
Theodoridis et al. 2012b). GC-MS mampu memisahkan pelbagai jenis sebatian dengan
berkesan terutamanya metabolit (dan metabolit terbitan) yang meruap, metabolit berkutub
dan semi-kutub yang kebanyakannya adalah metabolit primer seperti metabolit gula, asid
amino, asid organik, alkohol, sterol, toksin, hormon dan asid lemak (Fiehn 2016).
Kumpulan metabolit ini mempengaruhi serta menentukan ciri-ciri serta kualiti sesuatu
tumbuhan atau buah (Osorio & Fernie 2014), sekaligus menyebabkan analisis GC-MS ini
banyak digunakan dalam kajian metabolomik terhadap tumbuhan dan buah. Analisis GC-
MS adalah satu proses yang melibatkan pemisahan sampel di dalam turus komponen GC
diikuti dengan proses pengionan dan pemecahan sampel apabila sampel keluar dari hujung
turus (Rajah 2.5). Kemudian, sampel akan dipecahkan kepada serpihan berdasarkan berat
jisim dan membentuk corak pemecahan yang seterusnya akan dikesan oleh pengesan MS.
Corak pemecahan serta masa penahanan (RT) adalah spesifik dan unik kepada setiap
sebatian di dalam sampel, justeru dianggap sebagai ciri pengenalpastian atau turut
dikatakan sebagai cap jari molekul bagi sebatian tersebut.

35
Rajah 2.5 Sistem kromatografi gas-spektrometri jisim (GC-MS)
Sumber: Hussain and Maqbool (2014)
Analisis sampel menggunakan GC-MS memerlukan proses tambahan selepas
pengekstrakan sampel dijalankan iaitu derivatisasi sampel. Proses derivatisasi akan
menukarkan sebatian berkutub yang tidak meruap atau kurang meruap kepada molekul
terbitan yang lebih meruap dan stabil haba bagi membolehkan sebatian dapat dikenalpasti
di dalam sistem GC-MS pada suhu yang sesuai tanpa berlakunya penguraian terma (Knapp
1979; Orata 2012) atau pengubahsuaian pada struktur molekul (Kühnel et al. 2007; Orata
2012). Selain mengubah suai kumpulan fungsi metabolit agar dapat dianalisis di dalam
sistem GC-MS, proses derivatisasi juga dijalankan bertujuan untuk mengurangkan
penjerapan analit di dalam sistem GC-MS serta meningkatkan tindak balas pengesanan MS
(Orata 2012). Proses derivatisasi yang berkesan akan menghasilkan puncak metabolit yang
tajam, bersimetri, jelas dan mempunyai resolusi yang tinggi pada kromatogram (Orata
2012), seterusnya memudahkan proses pengenalpastian bagi setiap individu metabolit.
Kromatografi cecair-spektrometri jisim (LC-MS) pula adalah satu teknik analitikal
kimia yang menggabungkan keupayaan pemisahan fizikal kromatografi cecair dengan
kebolehan analisis jisim oleh spektrometri jisim untuk mengenalpasti kehadiran metabolit
di dalam sesuatu sampel biologi (Rajah 2.6). LC-MS telah menjadi salah satu teknik pilihan
dan telah digunakan secara meluas dalam pelbagai bidang kajian termasuklah bidang
farmaseutikal, alam sekitar, biokimia, bioteknologi dan sebagainya sejak beberapa dekad

36
terakhir pada abad ke-20 (Niessen 1999) kerana kebolehan sistem ini untuk memisahkan
dan mengenalpasti julat molekul yang besar (Theodoridis et al. 2012b).
Rajah 2.6 Sistem kromatografi cecair-spektrometri jisim (LC-MS)
Sumber: Anon (2017)
Antara kelas metabolit yang boleh dikenalpasti melalui sistem LC-MS adalah
metabolit sekunder tumbuhan seperti alkaloid, saponin, asid fenolik, fenilpropanoid,
flavonoid dan glukosinolat (De Vos et al. 2007; Huhman & Sumner 2002; Moco et al.
2006) yang mempunyai pelbagai kepentingan dari sudut ekonomi. Kajian mengenai
metabolit sekunder tumbuhan banyak difokuskan kepada kesan rintangan tumbuhan
terhadap tekanan persekitaran, bioaktiviti serta perkembangan warna dan rasa dalam
tumbuhan atau pemasakan buah (De Vos et al. 2007). Analisis LC-MS seringkali diikuti
dengan analisis spektrometri jisim seiring (MS/MS) menggunakan teknik seperti
spektrometri jisim automatik (AutoMS) dan pemantauan tindak balas berganda (MRM)
untuk kajian yang lebih teliti dan mendalam mengenai sifat molekular sesuatu metabolit.

37
2.3.3 Analisis data
a. Pemprosesan data mentah
Pengendalian dan pemprosesan data mentah adalah proses yang paling mencabar dan
mengambil masa yang paling lama dalam suatu kajian metabolomik (Shulaev 2006;
Theodoridis et al. 2012b). Data yang terhasil daripada analisis MS adalah besar dan
kompleks serta terdiri daripada tiga dimensi yang mewakili masa penahanan (retention
time) (RT), nilai m/z dan intensiti signal (Theodoridis et al. 2012b). Perlombongan data
seterusnya diperlukan untuk menjalankan proses penapisan bunyi, pembetulan garis dasar,
pemusatan, pengenyahkonvolusi spektrum, normalisasi, pemilihan puncak, integrasi
puncak dan penyelarasan masa penahanan (Cambiaghi et al. 2017; Theodoridis et al.
2012b). Proses ini akan menukarkan data matriks tiga dimensi kepada dua dimensi yang
terhasil di dalam bentuk jadual tersusun yang mengandungi nilai m/z, RT dan intensiti yang
sepadan bagi setiap puncak yang dikenalpasti.
Proses perlombongan data ini biasanya dijalankan menggunakan perisian yang
dikhususkan kepada sesuatu instrumen analisis seperti Bruker® DataAnalysis dan Bruker®
ProfileAnalysis untuk instrumen daripada Bruker, perisian MassHunter untuk instrumen
Agilent, perisian MarkerLynx untuk instrumen Waters dan perisian Turbo Mass daripada
Perkin Elmer (Theodoridis et al. 2012b). Terdapat juga perisian yang boleh dicapai secara
percuma dan terbuka kepada umum seperti MZmine (Pluskal et al. 2010; Theodoridis et al.
2012b), XCMS (Smith et al. 2006; Theodoridis et al. 2012b) dan MET-IDEA (Broeckling
et al. 2006; Lei et al. 2012) yang boleh memproses data daripada pelbagai instrumen
analisis. Perisian-perisian ini beroperasi dengan menggunakan fail data berformat NetCDF
(Fiehn 2008).
Selain itu, perisian seperti Leco Chromatof dan AMDIS (Automated Mass Spectral
Deconvolution and Identification System) menawarkan analisis tambahan iaitu
pengenyahkonvolusi data khususnya untuk pemprosesan data daripada analisis GC-MS
(Behrends et al. 2011; Shulaev 2006). Perisian ini jarang digunakan untuk analisis data

38
daripada instrumen yang lain seperti LC-MS kerana proses pengenyahkonvolusi data
daripada instrumen ini boleh dilakukan menggunakan perisian khusus seperti Bruker®
DataAnalysis. Data daripada analisis metabolomik seringkali menghasilkan kromatogram
yang mempunyai puncak sebatian yang bertindih. Proses pegenyahkonvolusi dijalankan
bertujuan untuk mengekstrak spektrum jisim tunggal daripada puncak yang bertindih
seterusnya menghasilkan spektrum jisim yang spesifik kepada hanya satu sebatian sahaja
(Behrends et al. 2011).
Analisis metabolomik turut menawarkan beberapa pangkalan data yang kukuh bagi
proses pengenalpastian metabolit. Antaranya adalah National Institute of Standards and
Technology (NIST), METLIN (Tautenhahn et al. 2012), Human Metabolome Database
(HMDB) (Wishart et al. 2012), Chemical Entities of Biological Interest (ChEBI) (Marco-
Ramell et al. 2018), MassBank (Horai et al. 2010) dan Kyoto Encyclopedia of Genes and
Genomes (KEGG) (Kanehisa & Goto 2000; Marco-Ramell et al. 2018). Pangkalan data ini
menawarkan proses anotasi yang pantas dan tepat menggunakan maklumat seperti masa
penahanan (RT), nilai m/z serta fragmentasi ion yang terdapat pada metabolit atau sebatian
yang berjaya dikesan melalui instrumen analisis metabolomik (Garg et al. 2015).
b. Analisis statistik univariat dan multivariat
Analisis data metabolomik biasanya disertai dengan analisis univariat dan analisis
multivariat. Analisis statistik seperti t-test atau analisis varians (ANOVA) diperlukan untuk
mengenalpasti komponen-komponen atau metabolit yang signifikan bagi setiap kumpulan
sampel yang dibandingkan (Vinaixa et al. 2012). Analisis ini boleh dijalankan
menggunakan beberapa perisian seperti SPSS dan GraphPad Prism serta pelayan
MetaboAnalyst (https://www.metaboanalyst.ca/). MetaboAnalyst adalah satu pelayan web
yang direka untuk menjalankan analisis komprehensif, visualisasi dan interpretasi bagi data
metabolomik yang kompleks. Pelayan ini boleh mengendalikan data metabolomik daripada
pelbagai platform MS dan NMR yang melibatkan pelbagai jenis analisis (analisis sasaran,
bukan sasaran, kuantitatif) (Xia et al. 2015b).

39
MetaboAnalyst boleh menjalankan pelbagai analisis serta visualisasi data
menggunakan pelbagai kaedah univariat dan multivariat seperti t-test, ANOVA, analisis
komponen utama (PCA), analisis diskriminan-separa kuasa dua terkecil (PLS-DA) dan
pengelompokan peta haba. MetaboAnalyst turut menawarkan pelbagai analisis interpretasi
data metabolomik seperti analisis pengkayaan set metabolit (MSEA) dan analisis tapak
jalan metabolomik (MetPA). Selain daripada pelayan Metaboanalyst, perisian SIMCA-P
juga sering diaplikasikan dalam kajian metabolomik untuk menjalankan analisis
multivariat seperti PCA dan PLS-DA. Seperti MetaboAnalyst, SIMCA-P juga boleh
mengendalikan data metabolomik dari pelbagai sumber. MetaboAnalyst dan SIMCA-P
turut menawarkan analisis pra-pemprosesan data seperti normalisasi, penskalaan, dan
transformasi sebelum analisis multivariat dijalankan.
PCA dan PLS-DA adalah dua analisis multivariat utama yang dijalankan untuk
melihat taburan metabolit di dalam setiap kumpulan sampel yang dibandingkan. PCA
adalah analisis data tanpa seliaan yang dijalankan dengan cara melakukan transformasi data
terhadap set pembolehubah baru atau dikenali sebagai komponen utama (PCs) yang tidak
berkait antara satu sama lain (Jolliffe 2011; Yi et al. 2016). PCA dijalankan bertujuan untuk
mengurangkan dimensi data metabolomik yang mengandungi sejumlah besar
pembolehubah yang saling berkaitan dengan mengekalkan sebanyak mungkin variasi yang
wujud di dalam setiap pembolehubah asal (Jolliffe 2011). Dalam analisis PCA, data boleh
dilihat dalam bentuk plot 2-dimensi atau 3-dimensi yang dikenali sebagai plot skor dan plot
pemuatan. Plot skor lazimnya digunakan untuk menentukan corak pemisahan metabolit dan
melihat taburan metabolit antara kelompok manakala plot pemuatan pula digunakan untuk
mengenalpasti pembolehubah atau metabolit yang mempengaruhi pemisahan antara
kumpulan metabolit (Ren et al. 2015; Xia et al. 2015b).
Analisis PLS-DA pula adalah satu teknik analisis seliaan yang memerlukan
pengguna untuk menentukan kumpulan metabolit secara manual. Seperti analisis PCA,
hasil analisis PLS-DA juga boleh dilihat dalam bentuk plot skor dan plot pemuatan.
Analisis ini digunakan untuk menjalankan analisis diskriminasi serta pengenalpastian
pembolehubah atau metabolit berkaitan yang berpotensi menyumbang kepada pemisahan

40
antara metabolit melalui analisis pembolehubah berkepentingan dalam unjuran (VIP).
Secara ringkasnya, semakin tinggi nilai VIP sesuatu metabolit, semakin tinggi pengaruh
metabolit tersebut terhadap pemisahan antara kelompok metabolit (Azizan et al. 2016; Kind
et al. 2007). Analisis PLS-DA banyak digunakan bagi tujuan pengenalpastian penanda
biologi serta pengelasan penyakit (Kind et al. 2007; Xia et al. 2015b).

BAB III
BAHAN DAN KAEDAH
3.1 BAHAN
3.1.1 Sumber sampel
Buah manggis dituai dari plot tumbuhan Universiti Kebangsaan Malaysia (UKM) yang
terletak di Bangi, Selangor (2°55′09.0″N 101°47′04.8″E) semasa musim pemasakan
manggis pada bulan Jun hingga Ogos 2014 dan Jun hingga Ogos 2016. Buah yang dituai
pada tahun 2014 telah digunakan untuk mencapai objektif kajian yang pertama manakala
buah yang dituai pada tahun 2016 pula telah digunakan untuk mencapai objektif ke-2. Buah
manggis dipetik pada empat peringkat pemasakan yang berbeza iaitu semasa peringkat 0
(hijau), 2 (hijau kekuningan disertai tompokan merah jambu), 4 (merah keperangan) dan
6 (ungu gelap) berdasarkan Indeks Kematangan Manggis Malaysia (Osman & Milan 2006).
3.1.2 Bahan kimia dan reagen
Pelarut gred analisis terdiri daripada metanol dan kloroform diperoleh dari Syarikat Merck
(Jerman) dan asid formik diperoleh daripada APS Ajax Finechem (Auburn, Australia).
Piridin, metoksiamina hidroklorida (MeOX), N-metil-N-(trimetilsilil) trifluoroasetamina
(MSTFA), N,O-bistrifluoroasetamina (BSTFA), (±)-naringenin, asid isopimarik, asid sitrik
dan asid malik diperoleh daripada Sigma Aldrich, St. Loius, MO, USA. α-Mangostin dan
γ-mangostin diperoleh daripada ChamFaces (Wuhan Economic and Technological
Development Zone, Wuhan, Hubei).

42
3.2 KAEDAH
3.2.1 Pensampelan dan penyediaan sampel
Buah manggis yang telah dipetik dibawa ke makmal dan diasingkan kepada jenis tisu yang
berbeza iaitu perikarpa, isi dan biji kecuali manggis yang dipetik pada peringkat 0
pemasakan kerana biji dan isi sukar dipisahkan daripada kulit pada peringkat ini (Osman
& Milan 2006). Kemudian, setiap tisu manggis dikisar secara berasingan ke dalam bentuk
serbuk halus menggunakan alu dan lesung atau pengisar gred makmal yang telah
disejukkan terlebih dahulu dengan cecair nitrogen (-196°C). Semua peralatan yang
digunakan semasa penyediaan sampel iaitu alu, lesung, spatula, pisau pemotong dan papan
pemotong disterilkan terlebih dahulu menggunakan cecair etanol (75%). Sampel buah
manggis yang telah dikisar kemudian dikeringkan menggunakan pengering sejuk-beku
(Labconco, model: 74200) untuk mengeluarkan kandungan air sebelum disimpan di dalam
-80°C sebelum pengekstrakan dijalankan.
3.2.2 Pengekstrakan metabolit
Bagi pemilihan kaedah pengekstrakan yang paling sesuai untuk pemprofilan metabolit
daripada buah manggis menggunakan analisis kromatografi gas-spektrometri jisim (GC-
MS) dan kromatografi cecair-spektrometri jisim (LC-MS), pengoptimuman kaedah
pengekstrakan metabolit telah dijalankan terlebih dahulu. Sebanyak lima kaedah
pengekstrakan metabolit telah digunakan dan dibandingkan. Kaedah-kaedah ini
mempunyai perbezaan dari aspek kimia (jenis dan nisbah pelarut) serta aspek fizikal
(keberkesanan teknik sonikasi). Lima kaedah ini dipilih berdasarkan kemampuan untuk
mengekstrak pelbagai jenis metabolit merangkumi metabolit berkutub, tidak berkutub,
hidrofilik serta hidrofobik dan telah dibuktikan keberkesanannya melalui kajian-kajian
lepas dalam analisis metabolomik bagi sampel tumbuhan (Cadahía et al. 2015; De Vos et
al. 2007; Lisec et al. 2006; Okazaki et al. 2016). Kaedah yang berjaya mengekstrak
metabolit paling banyak serta merangkumi pelbagai kelas atau kumpulan metabolit

43
seterusnya dipilih untuk analisis keseluruhan peringkat pemasakan buah manggis
(peringkat 0, 2, 4, 6) dalam tisu manggis yang berbeza (perikarpa, isi dan biji).
a. Kaedah pengekstrakan 1
Kaedah 1 diubah suai daripada De Vos et al. (2007). Sebanyak 200 mg sampel yang telah
dikering-beku dilarutkan ke dalam larutan pengekstrakan sejuk yang disediakan segar (599
µL bagi 75% metanol dan 1.0 µL bagi 0.1% asid formik) berkadar isi padu per berat dengan
nisbah 3:1. Sampel divorteks selama 10 saat. Kemudian, setiap sampel disonikasi selama
20 minit pada frekuensi maksimum (40 kHz) secara berterusan di dalam air rendaman pada
suhu bilik. Selepas itu, sampel diempar selama sepuluh minit dengan kelajuan maksimum
(16,100 x g) pada suhu bilik. Supernatan yang berada pada lapisan atas (fasa berkutub)
diasingkan daripada pelet dan dikumpulkan ke dalam tiub baru.
b. Kaedah pengekstrakan 2
Kaedah 2 diperoleh daripada Lisec et al. (2006) dan diubah suai. Sebanyak kira-kira 200
mg sampel kering-beku dilarutkan ke dalam 1.5 mL larutan sejuk metanol (4°C) dan
divorteks selama 10 minit. Campuran ini kemudiannya dibiarkan selama 10 minit di dalam
air rendaman bersuhu 70°C. Kemudian, campuran diempar selama 10 minit pada kelajuan
11,000 x g dan supernatan yang diperoleh dipindahkan ke dalam tiub baru. Selepas itu,
sebanyak 0.75 mL kloroform sejuk ditambah, diikuti dengan 1.5 mL air suling. Campuran
divorteks selama 10 saat dan diempar selama 15 minit pada kelajuan 2,200 x g. Akhirnya,
supernatan diambil daripada lapisan atas (fasa berkutub) dan dimasukkan ke dalam tiub
yang baru.
c. Kaedah pengekstrakan 3
Kaedah 3 diubah suai daripada kaedah 1 dan kaedah 2 untuk membandingkan keberkesanan
teknik sonikasi yang diaplikasikan dalam kaedah 1 dengan menggunakan campuran pelarut
yang sama dengan kaedah 2. Sebanyak 200 mg serbuk sampel sejuk-beku dicampurkan
dengan larutan pengekstrakan yang terdiri daripada 1.5 mL metanol, 0.75 mL kloroform

44
dan 1.5 mL air suling. Campuran ini divorteks selama 10 saat dan disonikasi selama 15
minit pada kelajuan maksimum (40 kHz) secara berterusan di dalam air rendaman bersuhu
bilik. Kemudian, campuran ini diempar selama 10 minit pada kelajuan (16,100 x g) pada
suhu bilik. Supernatan yang berada pada lapisan atas (fasa berkutub) dikumpulkan dan
dipindahkan ke dalam tiub baru.
d. Kaedah pengekstrakan 4
Kaedah 4 diubah suai daripada Okazaki et al. (2016). Sebanyak 1.5 mL campuran larutan
pengekstrakan terdiri daripada metanol/kloroform/air suling (3:1:1 isipadu/isipadu)
dicampurkan dengan 200 mg sampel sejuk beku. Sampel kemudiannya divorteks selama
30 saat dan dibiarkan di dalam ais selama 30 minit sebelum dicampurkan dengan 0.4 mL
air suling (untuk mengekstrak metabolit akueus). Selepas itu, sampel divorteks kembali
selama 30 saat dan diempar pada kelajuan 1,400 rpm pada suhu 4°C selama dua minit untuk
memisahkan lapisan yang terbentuk. Supernatan yang diperoleh daripada lapisan atas (fasa
berkutub) dipindahkan ke dalam tiub baru.
e. Kaedah pengekstrakan 5
Kaedah 5 diubah suai daripada kaedah Cadahia et al. (2015). Sebanyak 200 mg serbuk
sampel dicampurkan dengan 0.4 mL kloroform sejuk dan disonikasi dalam air rendaman
selama 20 minit pada suhu bilik. Kemudian, 1.6 mL metanol sejuk dan air suling dengan
nisbah (3:1 isipadu/isipadu) ditambahkan ke dalam campuran dan disonikasi selama 20
minit. Campuran diempar pada kelajuan maksimum selama 10 minit dan akhir sekali,
supernatan pada lapisan atas (fasa berkutub) dikumpulkan dan dipindahkan ke dalam tiub
baru.
3.2.3 Penyediaan sampel bagi analisis GC-MS
Bagi analisis GC-MS, asid isopimarik digunakan sebagai piawai dalaman. Asid
isopimarik berkepekatan 100 ppm dicampurkan ke dalam setiap sampel yang telah
diekstrak (supernatan yang telah diasingkan pada langkah akhir setiap kaedah

45
pengekstrakan metabolit) sebelum sampel diderivatisasi menggunakan agen derivatisasi
MSTFA atau BSTFA. Sebanyak 50 µL supernatan yang mengandungi metabolit yang telah
diekstrak dan piawai dalaman dikeringkan dan dipekatkan terlebih dahulu di dalam vakum
pemekat (CHRIST, model: RVC2-18/MZ2C). Untuk derivatisasi MSTFA, kaedah diubah
suai daripada Okazaki et al. (2016). Sampel yang telah kering dilarutkan ke dalam 20
mg/mL metoksiamina hidroklorida di dalam piridin sebanyak 40 µL dan dibiarkan selama
90 minit pada suhu 40°C. Kemudian, sebanyak 40 µL MSTFA ditambah dan campuran
dibiarkan selama 30 minit pada suhu 40°C. Akhir sekali, sampel dipindahkan ke dalam
botol kaca untuk analisis GC-MS. Proses derivatisasi menggunakan BSTFA dijalankan
berdasarkan kaedah yang diubah suai daripada Cadahía et al. (2015). Sampel yang telah
kering dilarutkan ke dalam 40 µL 20 mg/mL metoksiamina hidroklorida dalam piridin dan
divorteks selama 10 saat. Kemudian, 40 µL BSTFA ditambah dan campuran divorteks
selama 10 saat. Seterusnya, sampel dibiarkan selama satu jam pada suhu 60°C sebelum
dipindahkan ke dalam botol kaca untuk analisis GC-MS.
3.2.4 Penyediaan sampel bagi analisis LC-MS dan LC-MS/MS
Bagi analisis LC-MS, supernatan yang mengandungi metabolit dicampurkan dengan
naringenin (100 ppm) yang berperanan sebagai piawai dalaman. Kemudian, sebanyak 50
µL sampel dilarutkan ke dalam 950 µL metanol untuk mencapai isi padu akhir sebanyak 1
mL. Kemudian, sampel dipindahkan ke dalam botol kaca untuk analisis LC-MS. Bagi
analisis LC-MS/MS pula, sebanyak 50 µL dari setiap kumpulan sampel yang telah
dicampurkan terlebih dahulu dengan naringenin disatukan menjadi satu sampel sebelum
dihantar untuk analisis.
3.2.5 Parameter analisis GC-MS
Sampel telah dianalisis menggunakan Perkin Elmer Clarus 600 Turbo Mass GC (Perkin
Elmer, USA) yang digabungkan dengan MS jenis tetrakutub yang beroperasi pada 70 eV.
Sampel sebanyak 1.0 μL telah disuntik ke dalam turus Elite 5MS (5% difenil 95%
dimetilpolisiloxan, 30.0 m x 0.25 mm ID x 250 µm). Parameter sistem GC-MS diubah suai

46
daripada Azizan et al. (2015) berdasarkan kesesuaian terhadap sampel yang digunakan. Gas
helium digunakan sebagai gas pembawa dan julat imbasan ditetapkan kepada 50-600 Da.
Suhu awal ditetapkan kepada 70°C untuk minit pertama, kemudian dinaikkan kepada 76°C
dengan kadar kenaikan sebanyak 1°C/minit sebelum dikekalkan selama satu minit.
Seterusnya, suhu dinaikkan kepada 300°C dengan kadar kenaikan sebanyak 6°C/minit dan
akhir sekali ditahan selama lima minit. Suhu injektor dan suhu pindahan ditetapkan kepada
250°C sementara suhu punca pula diubah kepada 300°C. Julat imbasan penuh berjaya
diperoleh selepas tempoh kelewatan pelarut selama lapan minit dengan nisbah pecahan
50:1.
3.2.6 Parameter analisis LC-MS
Pemisahan sebatian dilakukan menggunakan turus Thermo Scientific C18 (AcclaimTM
Polar Advantage II, 3 m x 150 mm, 3 µm) dalam sistem Thermo Scientific Dionex UltiMate
3000 UHPLC. Parameter sistem LC-MS diubah suai daripada Rosli et al. (2017) untuk
mendapatkan parameter yang paling sesuai dengan sampel yang digunakan. Isipadu sampel
yang disuntik adalah 1.0 μL. Elusi kecerunan dijalankan selama 25 minit (0.4 mL/min,
40°C) menggunakan H2O + 0.1% asid formik (A) dan 100% asetonitril (B). Kecerunan
bermula dengan 60% pelarut B (0-3 minit), 95% pelarut B (3-5 minit), 95% pelarut B (5-
10 minit), 60% pelarut B (10-20 minit) dan seterusnya 60% pelarut B (20-25 minit).
Spektrometri jisim beresolusi tinggi dijalankan menggunakan sistem MicroTOF QIII
Bruker Daltonic (Bruker, Germany). Pengionan ESI pada mod positif dan negatif
dijalankan menggunakan parameter berikut; voltan kapilari: 4500 V, tekanan pengabut: 1.2
bar, gas pengeringan: 8 L/min pada suhu 200 °C dan julat jisim: 50-1000 m/z. Bagi analisis
kromatografi cecair-spektrometri jisim seiring (LC-MS/MS), tetapan bagi sistem
kromatografi dan spektrometri jisim yang sama telah digunakan.
3.2.7 Pemprosesan data GC-MS
Data mentah GC-MS yang mengandungi maklumat nama metabolit, masa penahanan (RT),
nilai padanan, nilai padanan relatif dan kawasan puncak telah dijana daripada perisian

47
Turbo Mass (Perkin Elmer, USA). Pengenalpastian metabolit dijalankan menggunakan
perpustakaan spektrum jisim National Institute of Standards and Technology (NIST, 2008)
dengan nilai padanan minimum 700. Format data mentah GC-MS ini kemudian ditukarkan
kepada format .NetCDF sebelum dimuatnaik ke dalam perisian Automated Mass Spectra
Deconvolution and Identification System (AMDIS) versi 2.71 untuk pengenalpastian
puncak yang bertindih (proses pengenyahkonvolutan) dan seterusnya mendapatkan
spektrum jisim individu. Nilai padanan minimum bagi pengenalpastian metabolit
menggunakan AMDIS juga ditetapkan kepada 700.
Nilai padanan minimum ditetapkan kepada 700 bagi memastikan tiada hasil positif
palsu dan negatif palsu terjadi semasa proses pengenalpastian metabolit (Grapp et al. 2016;
Meyer et al. 2010) dan telah banyak digunakan dalam kajian lepas (Baharum et al. 2010;
Halket et al. 2005; Jonsson et al. 2005; Lee et al. 2013; Simon-Manso et al. 2013). Metabolit
yang mempunyai nilai padanan 700 dan ke atas serta mempunyai signal yang tinggi pada
kromatogram ion jumlah (TIC) telah dilaporkan dalam kajian ini manakala metabolit yang
mempunyai nama yang disertai dengan tanda soal (?) tidak diambil kira kerana metabolit
ini adalah berkemungkinan merupakan padanan positif palsu. Nilai indeks penahanan (RI)
bagi setiap metabolit yang dikenalpasti juga telah ditentukan menggunakan perisian
AMDIS. Kemudian, proses penjajaran secara manual menggunakan perisian Microsoft
Excel (2016) telah dijalankan terhadap data output yang diperoleh daripada AMDIS bagi
analisis seterusnya iaitu analisis statistik univariat dan multivariat.
3.2.8 Pemprosesan data LC-MS
Bagi analisis LC-MS, data mentah dijana daripada sistem MicroTOF QIII Bruker Daltonic
melalui perisian Bruker Compass DataAnalysisViewer versi 2.0 (Bruker Daltonics,
Germany) dan diperoleh dalam format .d. Seterusnya, data ini dimuatnaik ke perisian
ProfileAnalysis versi 2.1 (Bruker Daltonic, Germany) untuk pengelasan data (data
bucketing). Penjajaran puncak metabolit dijalankan dengan pengenalan data mentah
terhadap pencirian ciri-ciri molekul atau Find Molecular Features (FMF) menggunakan
parameter yang diubah suai daripada Krug et al. (2008) seperti berikut: ambang nisbah

48
signal kepada hingar (S/N): 5, pekali korelasi: 0.7, panjang minimum sebatian: 10 dan lebar
pelicinan: 3. Jadual pengkelasan (bucket table) sebatian dikira menggunakan ciri
pengkelasan lanjutan (advanced bucketing) menggunakan penetapan daripada penjajaran
masa (time alignment). Julat masa ditetapkan kepada 1 minit sehingga 25.02 minit dan julat
jisim ditetapkan kepada 50 hingga 1000 m/z. Senarai keseluruhan bagi nilai m/z berserta
RT kemudian dijana untuk seterusnya iaitu analisis multivariat dan pengenalpastian
metabolit.
Pengenalpastian metabolit bagi data yang diperoleh daripada analisis LC-MS
dijalankan berdasarkan struktur molekul yang dijana menggunakan Smart Formula
daripada perisian ProfileAnalysis versi 2.1 serta menggunakan pangkalan data
metabolomik METLIN (http://metlin.scripps.edu/) terhadap pangkalan data umum seperti
di dalam Jadual 3.1 di bawah.
Jadual 3.1 Senarai pangkalan data umum yang digunakan untuk pengenalpastian metabolit
bagi data daripada analisis LC-MS
Pangkalan data Pautan
Kyoto Encyclopedia of Genes and Genomes (KEGG) http://www.kegg.jp/kegg/
Chemical Entities of Biological Interest (ChEBI) https://www.ebi.ac.uk/chebi/
Human Metabolome Database (HMDB) http://www.hmdb.ca/
MetaCyc Metabolic Pathway Database (MetaCyc) https://metacyc.org/
Struktur molekul yang dicadangkan berdasarkan Smart Formula bagi setiap nilai
m/z dijadikan panduan bagi mengenalpasti metabolit menggunakan pangkalan data
METLIN. Pencarian dilakukan menggunakan carian mudah (simple search) menggunakan
parameter berikut; caj: negatif, toleransi: 30 ppm dan aduk: [M-H]-. Bagi data LC-MS/MS,
pencarian melalui pangkalan data METLIN dilakukan menggunakan padanan persamaan
pecahan (fragment similarity match) dan carian padanan spektrum MS/MS (MS/MS
spectrum match search) menggunakan parameter yang sama seperti carian mudah (simple
search). Pengenalpastian metabolit serta tapak fragmentasi pada struktur molekul metabolit
bagi data LC-MS/MS turut dilakukan melalui pangkalan data Metfrag (https://msbi.ipb-
halle.de/MetFragBeta/) menggunakan parameter pencarian yang ditetapkan seperti berikut;
ion aduk: [M-H]-, ppm carian: 30, pangkalan data: KEGG, ChEBI, HMDB dan MetaCyc.

49
Pencarian dilakukan dengan cara memasukkan nilai ion prakursor dan diikuti dengan nilai
m/z serta intensiti bagi setiap fragmen ion yang terhasil.
3.2.9 Analisis multivariat
Data daripada kedua-dua analisis GC-MS dan LC-MS yang telah diproses kemudian
dimuatnaik ke pelayan MetaboAnalyst 3.0 (www.metaboanalyst.ca) untuk normalisasi
integral. Parameter pelayan MetaboAnalyst ditetapkan seperti berikut; ciri normalisasi:
jumlah, transformasi data: transformasi log, penskalaan data: penskalaan pareto. Kumpulan
sampel yang mengandungi nilai hilang ditapis dan digantikan dengan nilai kecil (separuh
daripada nilai positif minimum data) (Xia et al. 2015a). Analisis peta haba dan analisis
statistik iaitu analisis varians (ANOVA) dua-hala turut dijalankan menggunakan data yang
telah diproses. Akhir sekali, data yang telah diproses dimuatnaik ke perisian SIMCA-P+
versi 14 (Umetrics AB, Umea, Sweden) untuk analisis data multivariat iaitu analisis
komponen utama (PCA) dan analisis diskriminan-separa kuasa dua terkecil (PLS-DA).
3.2.10 Analisis bioinformatik
Analisis gambar rajah Venn dilakukan menggunakan pelayan bioinformatik & genetik
evolusi yang boleh dicapai di pautan http://bioinformatics.psb.ugent.be/webtools/Venn/.
Analisis tapak jalan biokimia pemasakan manggis pula dijalankan melalui analisis tapak
jalan metabolomik (MetPA) (http://metpa.metabolomics.ca.) berdasarkan pencarian
terhadap pangkalan data KEGG. Pemetaan tapak jalan dilakukan berdasarkan hasil yang
diperoleh dari analisis MetPA serta rujukan terhadap beberapa pangkalan data seperti
KEGG dan MetaCyc serta kajian kepustakaan daripada kajian-kajian lepas.

BAB IV
HASIL
4.1 PERBANDINGAN KAEDAH PENGEKSTRAKAN METABOLIT
4.1.1 Perbandingan lima kaedah pengekstrakan metabolit disertai proses derivatisasi
MSTFA bagi analisis GC-MS
a. Pengenalpastian jumlah keseluruhan metabolit di dalam perikarpa manggis
Bagi tujuan pengoptimuman kaedah pengekstrakan metabolit, lima kaedah
pengekstrakan yang berbeza dibandingkan dan dinilai seperti yang diringkaskan di
dalam Jadual 4.1. Ini bagi mengenalpasti kaedah yang sesuai dan mampu memberikan
jumlah metabolit terbanyak daripada sampel manggis. Untuk tujuan pengoptimuman
ini juga, hanya sampel perikarpa manggis daripada peringkat akhir pemasakan
(peringkat 6) telah digunakan kerana manggis pada peringkat ini lebih mudah didapati.
Faktor kimia iaitu jenis pelarut dan nisbah pelarut serta faktor fizikal iaitu teknik
sonikasi yang memainkan peranan penting terhadap hasil pengekstrakan metabolit telah
dibandingkan dan dinilai keberkesanannya berdasarkan jumlah dan jenis metabolit yang
berjaya diekstrak (Jadual 4.1).
Kromatogram ion jumlah (TIC) yang diperoleh dari analisis GC-MS bagi
kelima-lima kaedah pengekstrakan (kaedah 1, 2, 3, 4 dan 5) dilampirkan di dalam
Lampiran A. Secara kualitatifnya, kaedah 3 dan kaedah 5 menghasilkan pemisahan
puncak yang lebih jelas, tajam dan merangkumi bilangan puncak yang lebih banyak
berbanding kaedah yang lain. Daripada analisis pengenalpastian metabolit yang
dijalankan terhadap pangkalan data National Institute of Standards and Technology
(NIST) pula, terdapat sebanyak 73 metabolit telah berjaya dikenalpasti secara putatif
daripada kelima-lima kaedah pengekstrakan (Lampiran B).

51
Jadual 4.1 Ringkasan bagi lima kaedah pengekstrakan yang berbeza daripada segi
campuran pelarut, nisbah pelarut dan kaedah lisis. Keputusan jumlah
metabolit dan metabolit primer yang dikenalpasti daripada analisis GC-MS
bagi tisu perikarpa manggis peringkat 6 ini juga dibandingkan
Kaedah Rujukan Pelarut Kaedah
lisis
Jumlah
metabolit
Jumlah
metabolit
primer
1 De Vos et al. (2007) Metanol (75%)/asid
formik (0.1%) (599:1)
Lisis
kimia &
sonikasi
28 17
2 Lisec et al. (2006) Metanol/kloroform/air
(2:1:2)
Lisis
kimia
22 20
3 Lisec et al. (2006) &
De Vos et al. (2007)
Metanol/kloroform/air
(2:1:2)
Lisis
kimia &
sonikasi
31 27
4 Okazaki et al. (2016) Metanol/kloroform/air
(3:1:1)
Lisis
kimia
14 13
5 Cadahia et al. (2015) Metanol/kloroform/air
(3:1:1)
Lisis
kimia &
sonikasi
34 28
Merujuk Jadual 4.1, kaedah 5 berjaya memprofilkan jumlah metabolit yang
paling tinggi iaitu sebanyak 34 metabolit dan diikuti dengan kaedah 3, 1, 2 dan 4 dengan
jumlah masing-masing sebanyak 31, 28, 22 dan 14 metabolit. Selain itu, kaedah 5 juga
menghasilkan jumlah metabolit primer yang paling tinggi iaitu sebanyak 28 metabolit
diikuti dengan kaedah 3, 2, 1 dan terakhir adalah kaedah 4 dengan jumlah masing-
masing sebanyak 27, 20, 17 dan 13 metabolit. Kaedah yang menggunakan campuran
pelarut metanol, kloroform dan air disertai dengan teknik sonikasi iaitu kaedah 3 dan
kaedah 5 berjaya mengekstrak jumlah metabolit serta jumlah metabolit primer yang
lebih tinggi jika dibandingkan dengan kaedah 1 yang menggunakan campuran larutan
metanol dengan asid formik serta kaedah 2 dan 4 yang menggunakan campuran larutan
metanol, kloroform dengan air tetapi tidak disertai dengan teknik sonikasi. Bagi
pengekstrakan metabolit sekunder pula, kaedah 1 berjaya mengekstrak jumlah
metabolit sekunder yang paling tinggi berbanding kaedah pengekstrakan yang lain
(Rajah 4.1).
Secara keseluruhannya, kelima-lima kaedah yang telah dikaji menunjukkan
pengekstrakan metabolit daripada kelas yang berbeza bagi sampel perikarpa manggis
pada peringkat akhir pemasakan ini (Rajah 4.1). Metabolit daripada kelas atau
kumpulan gula didapati merangkumi hampir separuh daripada jumlah keseluruhan
metabolit yang berjaya dikesan (49.32%). Kumpulan metabolit kedua terbesar pula

52
adalah kumpulan alkohol (termasuk alkohol gula) iaitu sebanyak 9.59%, diikuti dengan
kumpulan asid gula (8.22%), asid organik (6.85%) dan aldehid (1.37%). Kumpulan
metabolit sekunder turut dikenalpasti dalam peratusan kecil iaitu asid fenolik (5.48%)
dan sebatian aromatik (2.74%). Sebatian lain yang tidak tergolong di dalam mana-mana
kumpulan sebatian sebelum ini merangkumi 16.44% daripada keseluruhan metabolit
(Rajah 4.1).
Berdasarkan Rajah 4.1, kaedah 5 berjaya mengekstrak jumlah gula yang paling
banyak dan diikuti dengan kaedah 3. Kaedah 1, 2 dan 4 pula berjaya mengekstrak
jumlah gula yang sama. Alkohol berjaya diekstrak dalam jumlah yang sama kecuali
kaedah 1 yang menghasilkan jumlah metabolit yang lebih tinggi berbanding empat
kaedah pengekstrakan yang lain. Sementara itu, asid gula dan asid organik diekstrak
pada kadar yang berbeza bagi kebanyakan kaedah pengekstrakan yang digunakan.
Manakala, aldehid berjaya dikenalpasti melalui tiga kaedah pengekstrakan sahaja iaitu
kaedah 1, 2 dan 3. Metabolit sekunder yang merangkumi sebatian fenolik dan sebatian
aromatik hanya berjaya diekstrak melalui kaedah 1, 3 dan 5 dengan jumlah metabolit
sekunder paling banyak diperoleh daripada kaedah 1 (Rajah 4.1).
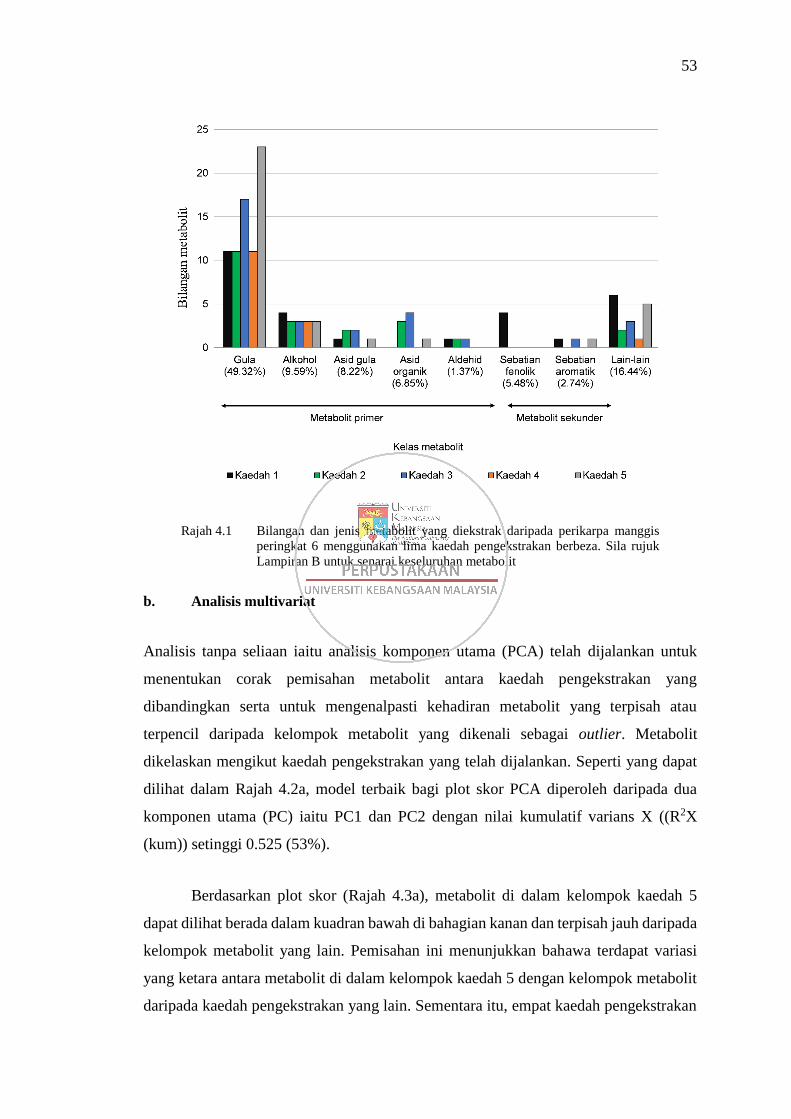
53
Rajah 4.1 Bilangan dan jenis metabolit yang diekstrak daripada perikarpa manggis
peringkat 6 menggunakan lima kaedah pengekstrakan berbeza. Sila rujuk
Lampiran B untuk senarai keseluruhan metabolit
b. Analisis multivariat
Analisis tanpa seliaan iaitu analisis komponen utama (PCA) telah dijalankan untuk
menentukan corak pemisahan metabolit antara kaedah pengekstrakan yang
dibandingkan serta untuk mengenalpasti kehadiran metabolit yang terpisah atau
terpencil daripada kelompok metabolit yang dikenali sebagai outlier. Metabolit
dikelaskan mengikut kaedah pengekstrakan yang telah dijalankan. Seperti yang dapat
dilihat dalam Rajah 4.2a, model terbaik bagi plot skor PCA diperoleh daripada dua
komponen utama (PC) iaitu PC1 dan PC2 dengan nilai kumulatif varians X ((R2X
(kum)) setinggi 0.525 (53%).
Berdasarkan plot skor (Rajah 4.3a), metabolit di dalam kelompok kaedah 5
dapat dilihat berada dalam kuadran bawah di bahagian kanan dan terpisah jauh daripada
kelompok metabolit yang lain. Pemisahan ini menunjukkan bahawa terdapat variasi
yang ketara antara metabolit di dalam kelompok kaedah 5 dengan kelompok metabolit
daripada kaedah pengekstrakan yang lain. Sementara itu, empat kaedah pengekstrakan
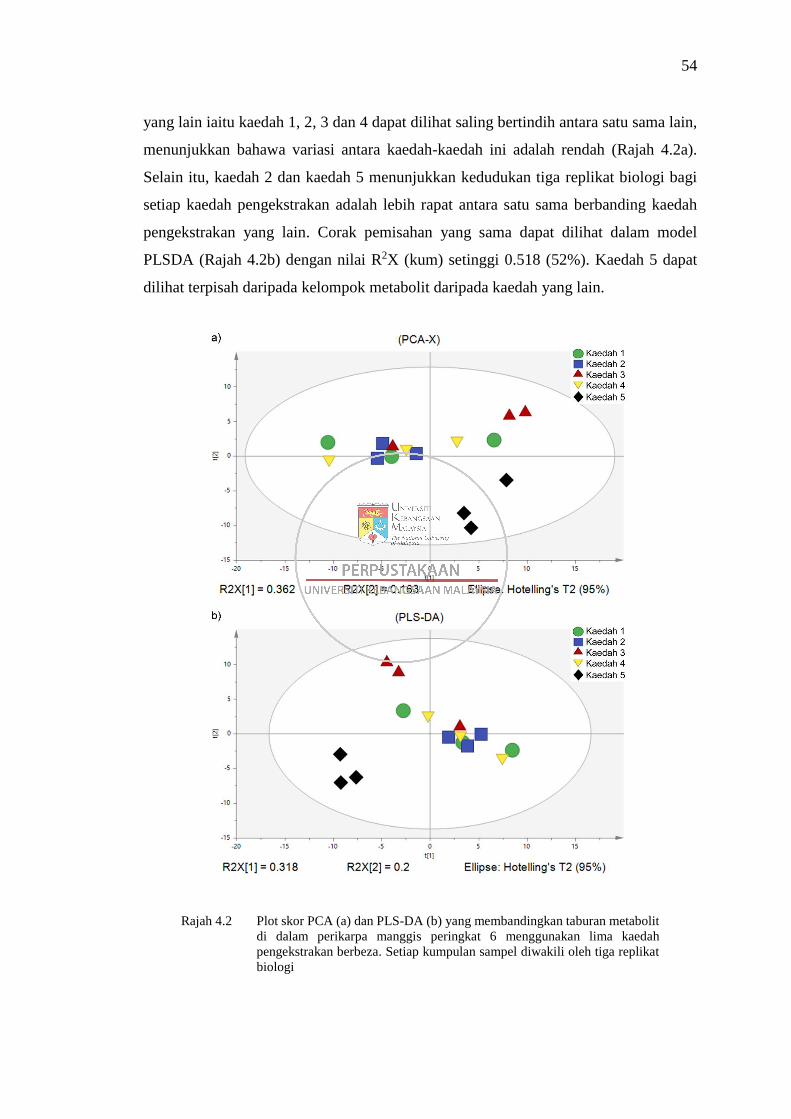
54
yang lain iaitu kaedah 1, 2, 3 dan 4 dapat dilihat saling bertindih antara satu sama lain,
menunjukkan bahawa variasi antara kaedah-kaedah ini adalah rendah (Rajah 4.2a).
Selain itu, kaedah 2 dan kaedah 5 menunjukkan kedudukan tiga replikat biologi bagi
setiap kaedah pengekstrakan adalah lebih rapat antara satu sama berbanding kaedah
pengekstrakan yang lain. Corak pemisahan yang sama dapat dilihat dalam model
PLSDA (Rajah 4.2b) dengan nilai R2X (kum) setinggi 0.518 (52%). Kaedah 5 dapat
dilihat terpisah daripada kelompok metabolit daripada kaedah yang lain.
Rajah 4.2 Plot skor PCA (a) dan PLS-DA (b) yang membandingkan taburan metabolit
di dalam perikarpa manggis peringkat 6 menggunakan lima kaedah
pengekstrakan berbeza. Setiap kumpulan sampel diwakili oleh tiga replikat
biologi
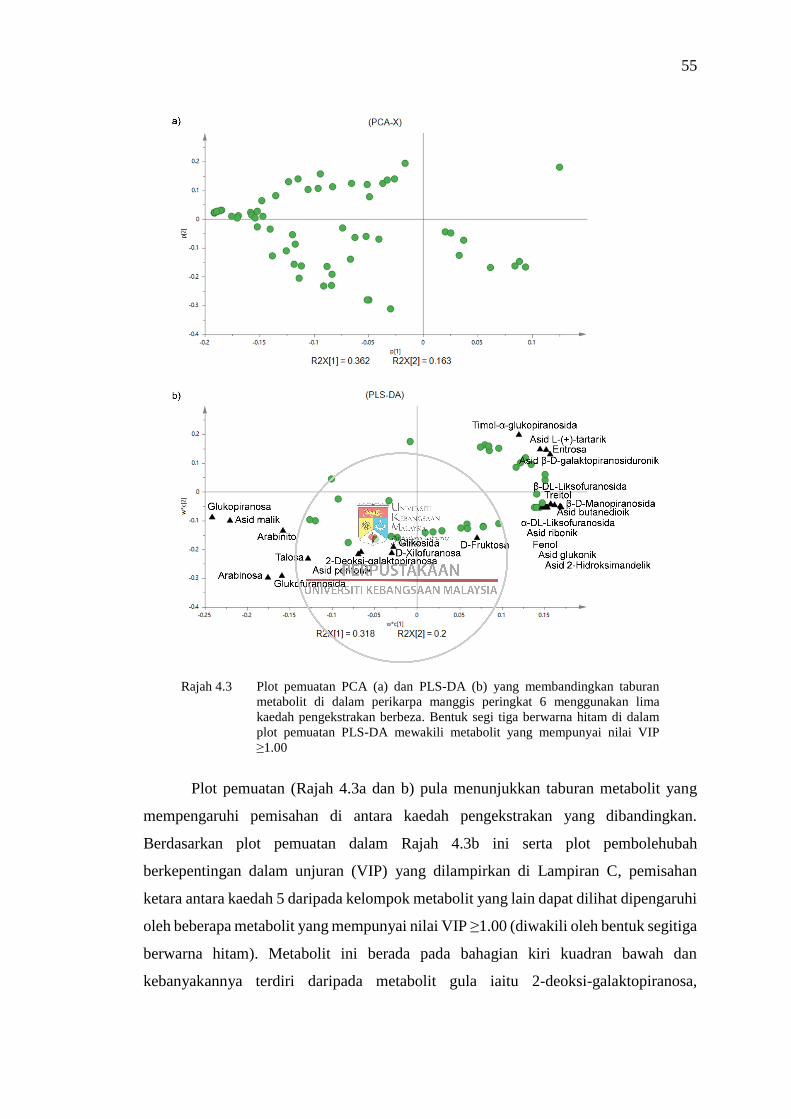
55
Rajah 4.3 Plot pemuatan PCA (a) dan PLS-DA (b) yang membandingkan taburan
metabolit di dalam perikarpa manggis peringkat 6 menggunakan lima
kaedah pengekstrakan berbeza. Bentuk segi tiga berwarna hitam di dalam
plot pemuatan PLS-DA mewakili metabolit yang mempunyai nilai VIP
≥1.00
Plot pemuatan (Rajah 4.3a dan b) pula menunjukkan taburan metabolit yang
mempengaruhi pemisahan di antara kaedah pengekstrakan yang dibandingkan.
Berdasarkan plot pemuatan dalam Rajah 4.3b ini serta plot pembolehubah
berkepentingan dalam unjuran (VIP) yang dilampirkan di Lampiran C, pemisahan
ketara antara kaedah 5 daripada kelompok metabolit yang lain dapat dilihat dipengaruhi
oleh beberapa metabolit yang mempunyai nilai VIP ≥1.00 (diwakili oleh bentuk segitiga
berwarna hitam). Metabolit ini berada pada bahagian kiri kuadran bawah dan
kebanyakannya terdiri daripada metabolit gula iaitu 2-deoksi-galaktopiranosa,

56
arabinosa, D-xilofuranosa, arabinitol, glikosida, glukopiranosa, glukofuranosida, talosa
dan asid pentonik serta asid organik iaitu asid malik (VIP ≥1.00).
c. Analisis perbandingan antara lima kaedah pengekstrakan berbeza
Analisis perbandingan menggunakan gambar rajah Venn (Rajah 4.4) menunjukkan
persamaan dan perbezaan bagi kelima-lima kaedah pengekstrakan metabolit yang telah
digunakan. Berdasarkah Rajah 4.4, terdapat lapan metabolit telah berjaya diekstrak
menggunakan kelima-lima kaedah pengekstrakan. iaitu tujuh Lapan metabolit ini terdiri
daripada tujuh metabolit primer iaitu arabinitol, D-glukosa, D-ribosa, D-manosa, D-
fruktosa, D-xilosa dan D-galaktosa dan satu metabolit daripada kategori “lain-lain” iaitu
timol-α-d-glukopiranosida (Jadual 4.2). Kaedah 1 dan 5 berjaya mengekstrak jumlah
metabolit unik yang paling banyak berbanding kaedah-kaedah yang lain iaitu sebanyak
16 metabolit unik bagi kaedah 1 dan 14 metabolit unik bagi kaedah 5. Metabolit unik
adalah merujuk kepada metabolit yang hanya berjaya diekstrak menggunakan kaedah
tertentu sahaja. Metabolit unik daripada kaedah 1 turut merangkumi lima metabolit
sekunder iaitu empat asid fenolik (asid salisilik, asid benzoik, asid 3,4-
dihidroksibenzoik dan 4-hidroksifeniletanol) dan satu sebatian aromatik (benzildehid)
(Jadual 4.2). Majoriti metabolit unik yang berjaya diekstrak menggunakan kaedah 5
pula adalah daripada kumpulan gula termasuklah talosa, manopiranosa dan D-
xilofuranosa. Satu asid gula iaitu asid pentonik turut dikenalpasti melalui kaedah 5.
Asid gula yang lain telah dikenalpasti sebagai metabolit unik bagi kaedah
pengekstrakan yang berbeza. Sebagai contoh, asid ribonik dan asid manonik hanya
hadir di dalam ekstrak perikarpa manggis daripada kaedah 2 manakala asid β-D-
galaktopiranosiduronik dan asid 2-keto-d-glukonik pula hanya dikenalpasti
menggunakan kaedah 3. Asid gula yang lain seperti asid glukonik pula adalah metabolit
unik bagi kaedah 1. Dua asid organik iaitu asid metilmaleik dan asid propanadioik
dikenalpasti sebagai metabolit unik bagi kaedah 3 manakala asid butanadioik pula
dikenalpasti sebagai metabolit unik bagi kaedah 2. Selain itu, asid organik seperti asid
malik dan asid L-(+)-tartarik pula berjaya diekstrak menggunakan lebih daripada satu
kaedah pengekstrakan. Asid malik telah dikenalpasti di dalam ekstrak daripada kaedah

57
2, 3 dan 5 manakala asid L-(+)-tartarik berjaya diekstrak melalui dua kaedah iaitu
kaedah 2 dan 3 (Jadual 4.2).
Rajah 4.4 Analisis gambar rajah Venn untuk analisis perbandingan lima kaedah
pengekstrakan bagi perikarpa manggis peringkat 6
Jadual 4.2 Senarai metabolit yang diperoleh daripada analisis gambar rajah Venn (rujuk
Rajah 4.4) untuk pengekstrakan metabolit daripada perikarpa manggis
peringkat 6 menggunakan lima kaedah pengekstrakan berbeza
Kaedah pengekstrakan Bilangan
metabolit
Metabolit
Kaedah 1, Kaedah 2, Kaedah 3, Kaedah 4 dan
Kaedah 5
8 Timol-α-d-glukopiranosida
Arabinitol
D-Glukosa
D-Ribosa
Manosa
D-Fruktosa
D-Xilosa
D-Galaktosa
Kaedah 1 , Kaedah 2, Kaedah 3 dan Kaedah 5 1 myo-Inositol
Kaedah 1, Kaedah 3, Kaedah 4 dan Kaedah 5 2 Arabinofuranosa
α-D-Glukopiranosida
Kaedah 1, Kaedah 2 dan Kaedah 3 1 Butanal
Kaedah 2, Kaedah 3 dan Kaedah 4 1 D-Xilopiranosa
Kaedah 2, Kaedah 3 dan Kaedah 5 2 Adonitol
Asid malik
bersambung...

58
sambungan...
Kaedah 3, Kaedah 4 dan Kaedah 5 1 D-Ribofuranosa
Kaedah 2 dan Kaedah 3 2 Liksosa
Asid L-(+)-tartarik
Kaedah 2 dan Kaedah 5 1 Arabinosa
Kaedah 3 dan Kaedah 4 1 D-Turanosa
Kaedah 3 dan Kaedah 5 3 2H-1-Benzopiran
Siklopenta-1,3-diena
Glukopiranosa
Kaedah 4 dan Kaedah 5 2 Glukofuranosida
α-DL-Arabinopiranosa
Kaedah 1
16 Asid glukonik
1,4-sikloheksadiena
Pirokatekol
Asid benzoik
Benzena
1,4-Pentadiena
Asid 3,4-dihidroksibenzoik
Benzaldehid
Asid salisilik
β-DL-Liksofuranosida
Asid 2-hidroksimandelik
α-DL-Liksofuranosida
α-l-Galaktofuranosida
Asid 3-merkaptobenzoik
Fenol
4-Hidroksifeniletanol
Kaedah 2 6 Asid ribonik
Asid butanadioik
Asid manonik
β-D-Manopiranosida
Asid 2-piridinakarboksilik
α-D-Galaktofuranosida
Kaedah 3 9 Asid β-D-
galaktopiranosiduronik Sorbopiranosa
Asid propanadioik
Inososa
Asid 2-keto-d-glukonik
Gulosa
Eritrosa
Asid α,β-L-idopiranuronik
Asid metilmaleik
Kaedah 4 3 Treitol
Pentitol
bersambung...

59
...sambungan
α-DL-Liksopiranosa
Kaedah 5 14 Liksopiranosida
Talosa
Glikosida
Manopiranosa
Secara keseluruhannya, kaedah pengekstrakan 3 (metanol: kloroform: air
dengan nisbah 2:1:2 disertai teknik sonikasi) dan kaedah pengekstrakan 5 (metanol;
kloroform: air dengan nisbah 3:1:1 disertai teknik sonikasi) menghasilkan jumlah
keseluruhan metabolit yang paling tinggi iaitu masing-masing sebanyak 31 dan 34
metabolit. Kaedah ini kemudian dipilih untuk proses pengoptimuman seterusnya iaitu
pengekstrakan metabolit daripada sampel tisu manggis yang berbeza iaitu perikarpa, isi
dan biji disertai derivatisasi menggunakan N,O-bis(trimetilsilil)trifluoroasetamida)
(BSTFA).
4.1.2 Perbandingan dua kaedah pengekstrakan metabolit disertai derivatisasi
menggunakan BSTFA bagi analisis GC-MS
a. Pengenalpastian jumlah keseluruhan metabolit di dalam perikarpa, isi dan biji
manggis
Selanjutnya, kajian diteruskan untuk mengenalpasti samada teknik pengekstrakan yang
digunakan adalah sesuai untuk jenis tisu manggis yang berbeza. Sampel tisu perikarpa,
isi dan biji manggis daripada peringkat 6 diekstrak menggunakan dua pengekstrakan
berbeza iaitu kaedah 3 (metanol: kloroform: air dengan nisbah 2:1:2 disertai teknik
sonikasi) dan kaedah 5 (metanol: kloroform: air dengan nisbah 3:1:1 disertai teknik
sonikasi). Dua kaedah ini dipilih berdasarkan jumlah metabolit terbanyak daripada
keputusan kajian pengoptimuman sebelum ini (Bab 4.1.1). Selain itu, ekstrak tisu
manggis juga diderivatisasi menggunakan BSTFA untuk menguji keberkesanan reagen
ini berbanding MSTFA yang telah digunakan sebelum ini.
Hasil daripada analisis GC-MS yang telah dijalankan, pemisahan puncak yang
jelas, tajam dan menyeluruh dapat dilihat bagi kedua-dua kaedah pengekstrakan
berdasarkan gambar rajah TIC pada Lampiran D dan Lampiran E. Secara
keseluruhannya, sebanyak 43 metabolit telah dikenalpasti hadir di dalam ketiga-tiga
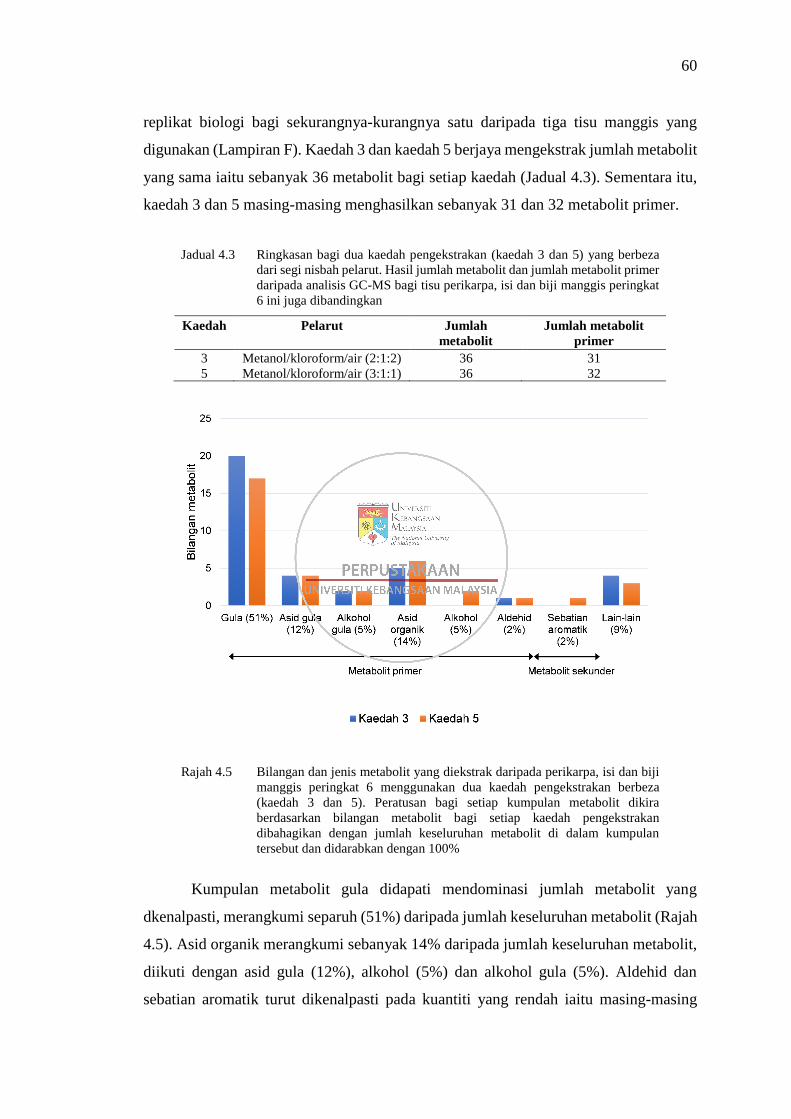
60
replikat biologi bagi sekurangnya-kurangnya satu daripada tiga tisu manggis yang
digunakan (Lampiran F). Kaedah 3 dan kaedah 5 berjaya mengekstrak jumlah metabolit
yang sama iaitu sebanyak 36 metabolit bagi setiap kaedah (Jadual 4.3). Sementara itu,
kaedah 3 dan 5 masing-masing menghasilkan sebanyak 31 dan 32 metabolit primer.
Jadual 4.3 Ringkasan bagi dua kaedah pengekstrakan (kaedah 3 dan 5) yang berbeza
dari segi nisbah pelarut. Hasil jumlah metabolit dan jumlah metabolit primer
daripada analisis GC-MS bagi tisu perikarpa, isi dan biji manggis peringkat
6 ini juga dibandingkan
Kaedah Pelarut Jumlah
metabolit
Jumlah metabolit
primer
3 Metanol/kloroform/air (2:1:2) 36 31
5 Metanol/kloroform/air (3:1:1) 36 32
Rajah 4.5 Bilangan dan jenis metabolit yang diekstrak daripada perikarpa, isi dan biji
manggis peringkat 6 menggunakan dua kaedah pengekstrakan berbeza
(kaedah 3 dan 5). Peratusan bagi setiap kumpulan metabolit dikira
berdasarkan bilangan metabolit bagi setiap kaedah pengekstrakan
dibahagikan dengan jumlah keseluruhan metabolit di dalam kumpulan
tersebut dan didarabkan dengan 100%
Kumpulan metabolit gula didapati mendominasi jumlah metabolit yang
dkenalpasti, merangkumi separuh (51%) daripada jumlah keseluruhan metabolit (Rajah
4.5). Asid organik merangkumi sebanyak 14% daripada jumlah keseluruhan metabolit,
diikuti dengan asid gula (12%), alkohol (5%) dan alkohol gula (5%). Aldehid dan
sebatian aromatik turut dikenalpasti pada kuantiti yang rendah iaitu masing-masing

61
sebanyak 2%. Sebanyak 9% daripada jumlah keseluruhan metabolit tidak dikategorikan
ke dalam mana-mana kelas metabolit dan dilabel sebagai “lain-lain”(Rajah 4.5).
Berdasarkan Rajah 4.5 di atas, kaedah 3 berjaya mengekstrak jumlah gula yang
lebih banyak berbanding kaedah 5. Selain itu, kedua-dua kaedah menghasilkan bilangan
metabolit asid gula, alkohol gula dan aldehid yang sama. Bagi asid organik, alkohol dan
sebatian aromatik pula, kaedah 5 menunjukkan hasil pengekstrakan yang lebih baik
berbanding kaedah 3. Selebihnya, kaedah 3 dan kaedah 5 turut mengekstrak metabolit
yang tidak dikategorikan kepada mana-mana kumpulan metabolit dengan bilangan yang
lebih tinggi ditunjukkan oleh kaedah 3 berbanding kaedah 5.
b. Analisis multivariat
Plot skor dan plot pemuatan PCA dihasilkan untuk menilai persamaan dan perbezaan
profil metabolit yang diperoleh daripada kaedah pengekstrakan 3 dan 5. Rajah 4.6a
menunjukkan plot PCA dihasilkan daripada dua komponen utama dengan jumlah
varians (R2) (campuran nilai R2X[1] dan R2X[2]) setinggi 0.607 (61%). Berdasarkan
plot skor PCA di dalam Rajah 4.6, dapat dilihat kebanyakan kelompok metabolit
daripada setiap kaedah pengekstrakan (kaedah 3 dan kaedah 5) serta tisu manggis yang
berbeza (perikarpa, isi dan biji) berjaya dipisahkan daripada satu sama lain. Pertindihan
hanya dapat dilihat berlaku pada kelompok metabolit daripada sampel isi antara kaedah
3 dan kaedah 5.
Plot skor PLS-DA (Rajah 4.6b) juga menunjukkan corak pemisahan yang
hampir sama di mana pertindihan antara kelompok metabolit hanya berlaku pada
kelompok metabolit daripada sampel isi bagi kaedah 3 dan 5 pada bahagian atas
kuadran tengah. Nilai R2 juga tidak berubah iaitu setinggi 0.607 (61%). Plot pemuatan
bagi plot PCA (Rajah 4.7a) dan PLS-DA (Rajah 4.7b) pula menunjukkan taburan
metabolit dalam setiap kelompok yang menyumbang kepada pemisahan yang berlaku
di dalam plot skor. Analisis VIP turut dijalankan untuk mengenalpasti metabolit yang
berpengaruh tinggi bagi pemisahan kelompok metabolit daripada kaedah pengekstrakan
dan jenis tisu yang berbeza. Metabolit yang mempunyai nilai VIP ≥1.00 diwakili oleh
simbol segitiga berwarna hitam (Rajah 4.7b) dan plot metabolit VIP dilampirkan di
dalam Lampiran G.
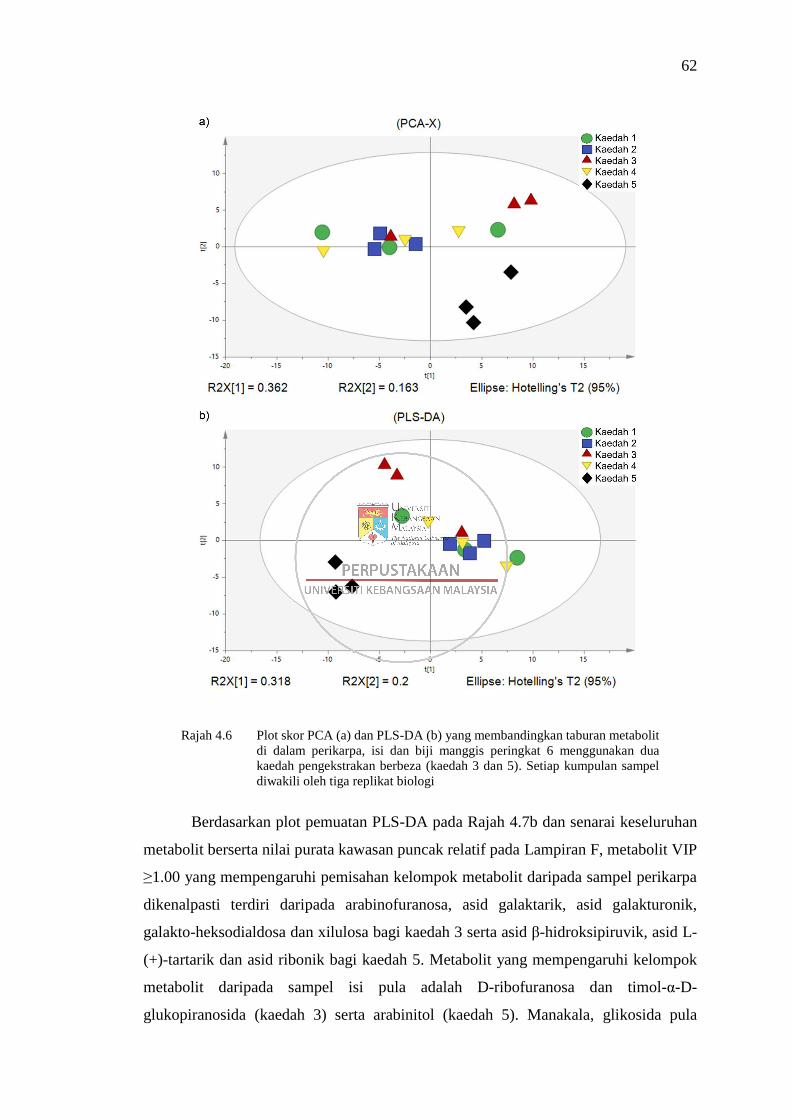
62
Rajah 4.6 Plot skor PCA (a) dan PLS-DA (b) yang membandingkan taburan metabolit
di dalam perikarpa, isi dan biji manggis peringkat 6 menggunakan dua
kaedah pengekstrakan berbeza (kaedah 3 dan 5). Setiap kumpulan sampel
diwakili oleh tiga replikat biologi
Berdasarkan plot pemuatan PLS-DA pada Rajah 4.7b dan senarai keseluruhan
metabolit berserta nilai purata kawasan puncak relatif pada Lampiran F, metabolit VIP
≥1.00 yang mempengaruhi pemisahan kelompok metabolit daripada sampel perikarpa
dikenalpasti terdiri daripada arabinofuranosa, asid galaktarik, asid galakturonik,
galakto-heksodialdosa dan xilulosa bagi kaedah 3 serta asid β-hidroksipiruvik, asid L-
(+)-tartarik dan asid ribonik bagi kaedah 5. Metabolit yang mempengaruhi kelompok
metabolit daripada sampel isi pula adalah D-ribofuranosa dan timol-α-D-
glukopiranosida (kaedah 3) serta arabinitol (kaedah 5). Manakala, glikosida pula
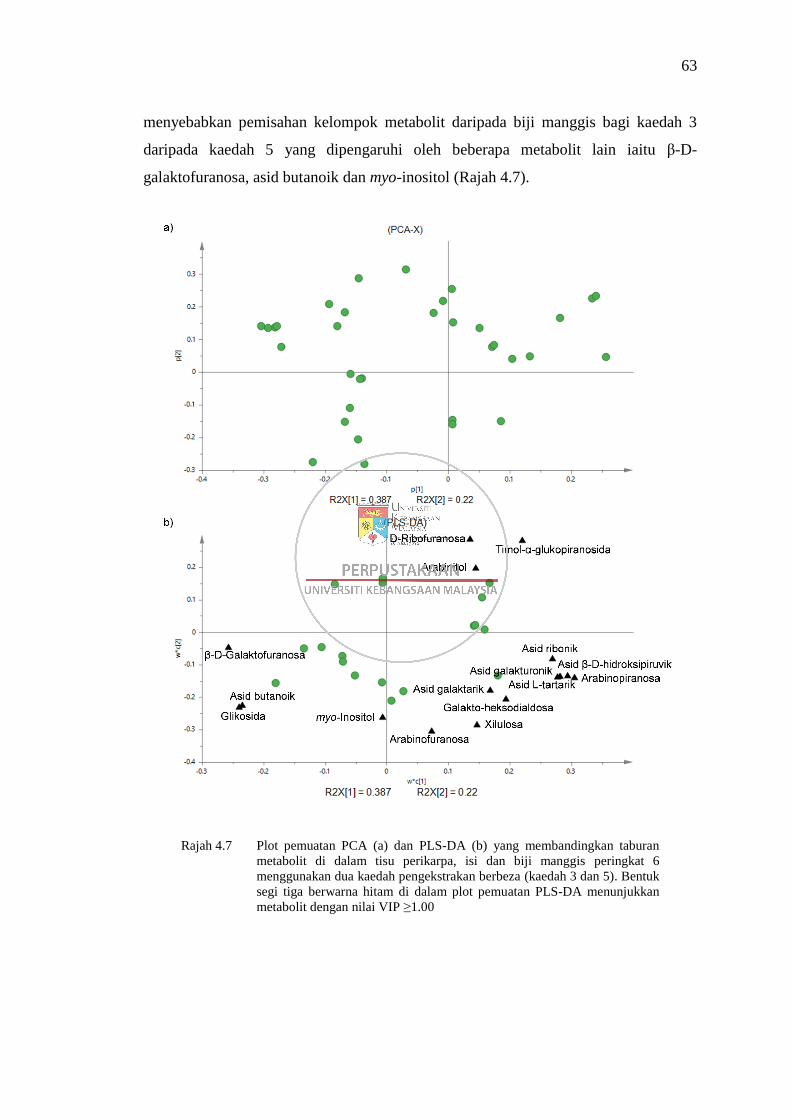
63
menyebabkan pemisahan kelompok metabolit daripada biji manggis bagi kaedah 3
daripada kaedah 5 yang dipengaruhi oleh beberapa metabolit lain iaitu β-D-
galaktofuranosa, asid butanoik dan myo-inositol (Rajah 4.7).
Rajah 4.7 Plot pemuatan PCA (a) dan PLS-DA (b) yang membandingkan taburan
metabolit di dalam tisu perikarpa, isi dan biji manggis peringkat 6
menggunakan dua kaedah pengekstrakan berbeza (kaedah 3 dan 5). Bentuk
segi tiga berwarna hitam di dalam plot pemuatan PLS-DA menunjukkan
metabolit dengan nilai VIP ≥1.00

64
c. Analisis perbandingan bagi dua kaedah pengekstrakan berbeza
Analisis perbandingan menggunakan gambar rajah Venn (Rajah 4.8 dan Jadual 4.4)
menunjukkan persamaan dan perbezaan antara dua kaedah pengekstrakan metabolit
berbeza tersebut mengikut jenis tisu manggis iaitu perikarpa, isi dan biji. Berdasarkan
gambar rajah Venn di bawah, kaedah 3 berjaya mengekstrak 10 metabolit yang sama
daripada ketiga-tiga jenis tisu manggis manakala kaedah 5 pula berjaya mengekstrak 13
metabolit daripada tisu yang sama.
Rajah 4.8 Analisis gambar rajah Venn yang membandingkan metabolit yang diekstrak
daripada tisu perikarpa, isi dan biji manggis peringkat 6 menggunakan dua
kaedah pengekstrakan berbeza (kaedah 3 dan 5)
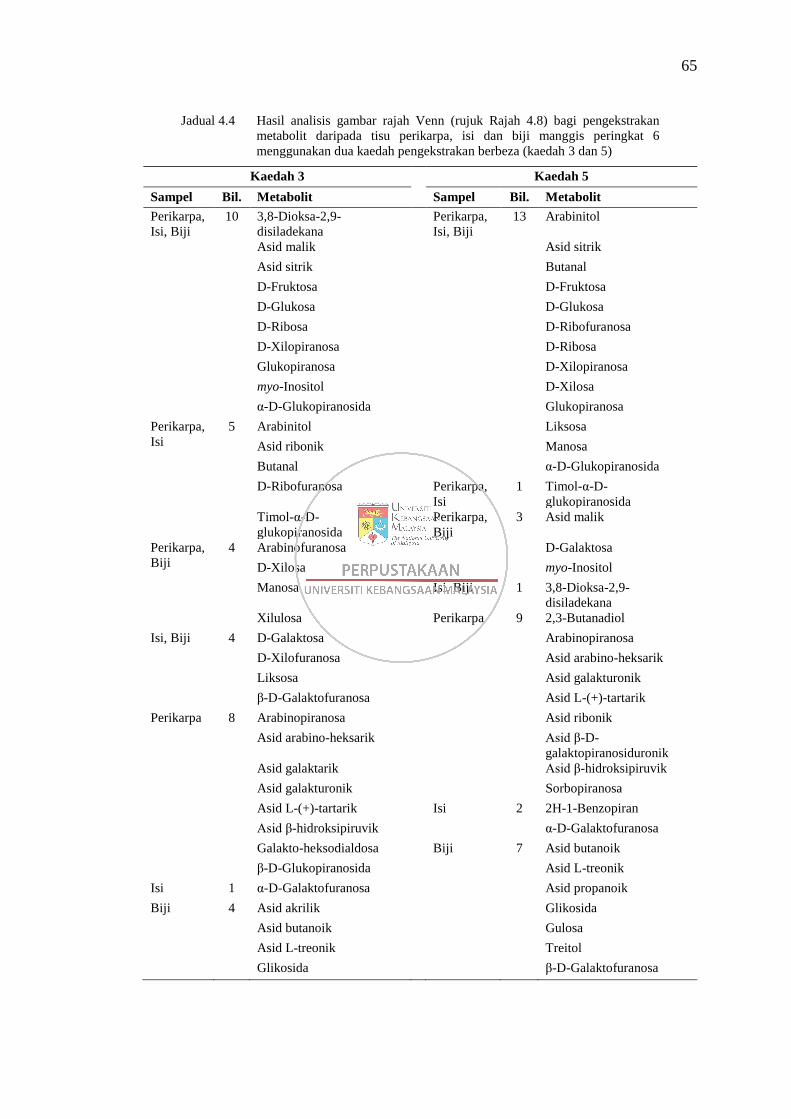
65
Jadual 4.4 Hasil analisis gambar rajah Venn (rujuk Rajah 4.8) bagi pengekstrakan
metabolit daripada tisu perikarpa, isi dan biji manggis peringkat 6
menggunakan dua kaedah pengekstrakan berbeza (kaedah 3 dan 5)
Kaedah 3 Kaedah 5
Sampel Bil. Metabolit Sampel Bil. Metabolit
Perikarpa,
Isi, Biji
10 3,8-Dioksa-2,9-
disiladekana
Perikarpa,
Isi, Biji
13 Arabinitol
Asid malik
Asid sitrik
Asid sitrik
Butanal
D-Fruktosa
D-Fruktosa
D-Glukosa
D-Glukosa
D-Ribosa
D-Ribofuranosa
D-Xilopiranosa
D-Ribosa
Glukopiranosa
D-Xilopiranosa
myo-Inositol
D-Xilosa
α-D-Glukopiranosida
Glukopiranosa
Perikarpa,
Isi
5 Arabinitol
Liksosa
Asid ribonik
Manosa
Butanal
α-D-Glukopiranosida
D-Ribofuranosa Perikarpa,
Isi
1 Timol-α-D-
glukopiranosida Timol-α-D-
glukopiranosida
Perikarpa,
Biji
3 Asid malik
Perikarpa,
Biji
4 Arabinofuranosa
D-Galaktosa
D-Xilosa
myo-Inositol
Manosa Isi, Biji 1 3,8-Dioksa-2,9-
disiladekana Xilulosa Perikarpa 9 2,3-Butanadiol
Isi, Biji 4 D-Galaktosa
Arabinopiranosa
D-Xilofuranosa
Asid arabino-heksarik
Liksosa
Asid galakturonik
β-D-Galaktofuranosa
Asid L-(+)-tartarik
Perikarpa 8 Arabinopiranosa
Asid ribonik
Asid arabino-heksarik
Asid β-D-
galaktopiranosiduronik Asid galaktarik
Asid β-hidroksipiruvik
Asid galakturonik
Sorbopiranosa
Asid L-(+)-tartarik Isi 2 2H-1-Benzopiran
Asid β-hidroksipiruvik
α-D-Galaktofuranosa
Galakto-heksodialdosa Biji 7 Asid butanoik
β-D-Glukopiranosida
Asid L-treonik
Isi 1 α-D-Galaktofuranosa
Asid propanoik
Biji 4 Asid akrilik
Glikosida
Asid butanoik
Gulosa
Asid L-treonik
Treitol
Glikosida
β-D-Galaktofuranosa

66
Berdasarkan Jadual 4.4, enam gula iaitu D-fruktosa, D-glukosa, D-ribosa, D-
xilopiranosa, glukopiranosa dan α-D-glukopiranosida serta satu asid organik iaitu asid
sitrik telah berjaya diekstrak menggunakan kedua-dua kaedah pengekstrakan (kaedah
3: dan kaedah 5) daripada semua tisu manggis (perikarpa, isi dan biji) peringkat 6 ini.
Gula yang lain pula hanya didapati di dalam tisu manggis yang tertentu, contohnya,
arabinopiranosa hanya didapati di dalam perikarpa manggis sementara α-D-
galaktofuranosa pula hanya ditemui di dalam isi manggis apabila diekstrak
menggunakan kedua-dua kaedah pengekstrakan. Arabinofuranosa, galakto-
heksodialdosa, D-xilofuranosa, β-D-glukopiranosida dan xilulosa hanya berjaya
diekstrak menggunakan kaedah 3 daripada sekurang-kurangnya satu tisu manggis
manakala gulosa dan sorbopiranosa pula didapati merupakan metabolit unik kepada
kaedah 5 (Jadual 4.4).
Kedua-dua kaedah pengekstrakan berjaya mengekstrak dua asid gula iaitu asid
galakturonik dan asid ribonik daripada perikarpa manggis. Asid ribonik juga ditemui di
dalam isi manggis daripada kaedah 3. Asid L-treonik pula merupakan metabolit unik
untuk biji manggis bagi kedua-dua kaedah pengekstrakan. Asid galaktarik hanya
dikenalpasti di dalam ekstrak perikarpa daripada kaedah 3 manakala asid β-D-
galaktopiranosiduronik dikenalpasti di dalam tisu yang sama daripada kaedah 5. Bagi
asid organik pula, asid sitrik dikenalpasti di dalam ketiga-tiga tisu manggis daripada
kaedah 3 dan 5 manakala asid malik ditemui di dalam ketiga-tiga tisu daripada kaedah
3 tetapi hanya ditemui di dalam perikarpa dan isi manggis daripada kaedah 5. Asid
arabino-heksarik dan asid L-(+)-tartarik dikenalpasti sebagai metabolit unik kepada tisu
perikarpa manakala asid butanoik merupakan metabolit unik kepada tisu biji bagi
kedua-dua kaedah pengekstrakan. Kaedah 5 turut berjaya mengekstrak satu asid organik
lain iaitu asid propanoik daripada biji manggis (Jadual 4.4).
Kaedah 3 dan 5 masing-masing berjaya mengekstrak dua metabolit gula yang
sama iaitu arabinitol dan myo-inositol daripada tisu yang berbeza. Arabinitol hadir di
dalam ketiga-tiga tisu daripada kaedah 5 tetapi hanya hadir di dalam perikarpa dan isi
bagi daripada kaedah 3. myo-Inositol pula dikenalpasti di dalam ketiga-tiga tisu bagi
daripada kaedah 3 tetapi hanya ditemui di dalam tisu perikarpa dan biji daripada kaedah
5. Selain itu, dua alkohol iaitu 2,3-butanadiol dan treitol masing-masing dikenalpasti

67
sebagai metabolit unik kepada perikarpa dan biji bagi kaedah 5. Butanal berjaya
diekstrak daripada kedua-dua kedah pengekstrakan (perikarpa dan isi bagi kaedah 3 dan
ketiga-tiga tisu bagi kaedah 5). Timol-α-D-glukopiranosida pula diekstrak daripada tisu
perikarpa dan isi manggis menggunakan kedua-dua kaedah pengekstrakan sementara
asid β-hidroksipiruvik pula hanya dikesan di dalam perikarpa. Asid akrilik hanya
dikenalpasti di dalam biji manggis apabila diekstrak menggunakan kaedah 3 manakala
3,8-dioksa-2,9-disiladekana dikenalpasti di dalam ketiga-tiga tisu daripada kaedah 3
dan tisu isi serta biji daripada kaedah 5 (Jadual 4.4).
Secara kesimpulannya, kaedah 3 dan 5 masing-masing menunjukkan hasil
pengekstrakan yang memberangsangkan dengan penghasilan jumlah metabolit yang
sama iaitu 36 metabolit. Namun, kaedah 5 memberikan jumlah metabolit primer yang
lebih tinggi (32 metabolit) berbanding kaedah 3 (31 metabolit). Kaedah 5 kemudiannya
dipilih untuk analisis pemprofilan metabolit daripada perikarpa, isi dan biji manggis
pada peringkat pemasakan yang berbeza (peringkat 0, 2, 4 dan 6) meskipun perbezaan
antara dua kaedah ini adalah tidak ketara.
4.1.3 Perbandingan dua kaedah pengekstrakan serta mod pengionan berbeza bagi
analisis LC-MS
Dua kaedah pengekstrakan iaitu kaedah 1 (metanol/asid formik dengan nisbah 599:1)
dan 5 (metanol/kloroform/air dengan nisbah 3:1:1) telah dibandingkan sekali lagi
menggunakan analisis LC-MS bagi mengkaji kesesuaian kaedah ini dengan sistem
analisis yang berbeza. Kaedah 1 dan kaedah 5 dipilih berdasarkan hasil analisis
perbandingan kaedah pengekstrakan yang telah dijalankan sebelum ini. Kaedah 1
dipilih kerana kemampuan kaedah ini untuk mengekstrak metabolit sekunder (Rajah
4.1) manakala kaedah 5 pula dipilih kerana kaedah ini mampu mengekstrak jumlah
metabolit yang lebih tinggi berbanding kaedah lain (Rajah 4.1 dan Rajah 4.5)
Sampel ekstrak daripada perikarpa manggis peringkat 6 diuji pada dua mod
pengionan semburan elektro (ESI) bagi analisis LC-MS iaitu mod positif dan mod
negatif. Hasil analisis diperoleh daripada perisian DataAnalysisViewer dan dinilai
berdasarkan bilangan puncak pada kromatogram yang diwakili dengan pasangan RT:
m/z yang diperoleh daripada setiap kaedah pengekstrakan pada mod pengionan yang

68
berbeza tanpa analisis pengenalpastian metabolit. Hasil analisis telah diringkaskan
seperti dalam Jadual 4.5 di bawah.
Jadual 4.5 Perbandingan dua kaedah pengekstrakan dan mod pengionan bagi analisis
LC-MS untuk sampel perikarpa manggis peringkat 6. Senarai penuh
bilangan puncak (pasangan RT: m/z) boleh dirujuk pada Lampiran H
Kaedah
pengekstrakan
Bilangan puncak (pasangan RT: m/z)
Mod pengionan
Positif Negatif
Kaedah 1 46 58
Kaedah 5 55 41
Kaedah 1 menghasilkan lebih banyak bilangan puncak pada mod negatif iaitu
sebanyak 58 pasangan RT: m/z manakala hanya 46 pasangan RT: m/z dihasilkan pada
mod positif. Berbeza dengan kaedah 1, kaedah 5 didapati lebih sesuai digunakan pada
mod positif kerana penghasilan bilangan pasangan RT: m/z adalah lebih tinggi pada
mod ini iaitu sebanyak 55 pasangan RT: m/z berbanding 41 pasangan RT: m/z pada mod
negatif. Secara keseluruhannya, kaedah 1 yang diuji pada mod pengionan negatif telah
memberikan hasil yang paling memberangsangkan. Oleh itu, kaedah ini kemudiannya
dipilih untuk pemprofilan metabolit daripada perikarpa, isi dan biji manggis yang dituai
pada empat peringkat pemasakan berbeza (peringkat 0, 2, 4 dan 6).
4.2 PEMPROFILAN METABOLIT TISU PERIKARPA, ISI DAN BIJI MANGGIS
DARIPADA EMPAT PERINGKAT PEMASAKAN BERBEZA
4.2.1 Pemprofilan metabolit putatif menggunakan analisis GC-MS
Analisis GC-MS telah dijalankan untuk menganalisis kandungan metabolit secara
global daripada perikarpa, isi dan biji manggis yang dituai pada empat peringkat
pemasakan yang berbeza iaitu peringkat 0, 2, 4 dan 6. Empat peringkat ini dipilih kerana
ianya merangkumi peringkat awal pemasakan (peringkat 0), peringkat pertengahan
(peringkat 2 dan 4) dan peringkat akhir pemasakan (peringkat 6). Terlebih dahulu,
manggis yang dituai daripada empat peringkat pemasakan ini diproses dan dipisahkan
kepada tiga jenis tisu yang berbeza iaitu perikarpa, isi dan biji kecuali manggis
peringkat 0 kerana ketiga-tiga tisu ini tidak dapat dipisahkan dengan sebaiknya pada
peringkat ini (Osman dan Milan 2006). Kemudian, setiap sampel diekstrak

69
menggunakan kaedah pengekstrakan yang telah dioptimumkan sebelum ini iaitu
menggunakan metanol/kloroform/air dengan nisbah 3:1:1 disertai teknik sonikasi
(kaedah pengekstrakan 5) dan derivatisasi menggunakan BSTFA memandangkan
kaedah ini mampu mengekstrak jumlah metabolit yang tinggi serta merangkumi
pelbagai kumpulan metabolit.
Daripada analisis GC-MS yang telah dijalankan, sebanyak 57 metabolit berjaya
dikenalpasti secara putatif seperti yang disenaraikan di dalam Lampiran I. Metabolit ini
merangkumi pelbagai kelas metabolit terutamanya metabolit primer iaitu gula yang
didapati dengan jumlah yang paling tinggi sebanyak 23 metabolit dan diikuti dengan
kumpulan glikosida gula (enam metabolit), alkohol gula (enam metabolit) dan asid
organik (enam metabolit). Metabolit alkohol, sebatian aromatik dan aldehid turut
dikenalpasti dengan jumlah yang kecil iaitu hanya satu metabolit bagi setiap kelas.
Selain itu, turut dikenalpasti adalah metabolit yang tidak dikategorikan kepada mana-
mana kelas metabolit atau “lain-lain” sebanyak 8 metabolit.
a. Analisis statistik data univariat dan multivariat
Analisis komponen utama (PCA) telah dijalankan untuk melihat taburan metabolit
antara peringkat pemasakan yang berbeza berdasarkan kepada jenis tisu manggis.
Analisis PCA dijalankan secara berasingan bagi setiap tisu manggis seperti yang dapat
dilihat dalam Rajah 4.9a bagi tisu perikarpa, Rajah 4.9b bagi tisu isi dan Rajah 4.9c
bagi tisu biji manggis. Ketiga-tiga plot skor PCA mencapai jumlah keseluruhan variasi,
R2 (jumlah bagi R2X[1] dan R2X [2]) yang tinggi dan melebihi 50% (0.5) iaitu 0.679
bagi sampel perikarpa, 0.628 bagi sampel isi dan 0.678 bagi sampel biji manggis.

70
Rajah 4.9 Analisis PCA dan PLS-DA bagi analisis pemprofilan metabolit perikarpa,
isi dan biji manggis pada peringkat pemasakan berbeza menggunakan GC-
MS. a) Plot skor PCA sampel perikarpa, b) plot skor PCA sampel isi, c) plot
skor PCA sampel biji, d) plot skor PLS-DA sampel perikarpa, e) plot skor
PLS-DA sampel isi dan f) plot skor PLS-DA sampel biji. Setiap kumpulan
sampel diwakili oleh tiga replikat biologi
Berdasarkan plot skor PCA yang telah dijana daripada perisian SIMCA-P+ versi
14 ini, perbezaan yang jelas disertai kedudukan yang rapat antara replikat biologi dapat
dilihat antara setiap peringkat pemasakan 0, 2, 4 dan 6 bagi ketiga-tiga jenis tisu
manggis. Sedikit pertindihan berlaku antara sampel daripada peringkat 0 dengan sampel
perikarpa daripada peringkat 2 (Rajah 4.9a). Kemudian, analisis PLS-DA (Rajah 4.9d,
e dan f) turut dijalankan dengan memfokuskan kepada analisis pengenalpastian
metabolit berkepentingan dalam unjuran (VIP) bagi mengesan metabolit yang
berkemungkinan sebagai penanda biologi dalam pemasakan buah manggis.
Seperti analisis PCA, analisis PLS-DA juga berjaya memisahkan kelompok
metabolit bagi peringkat pemasakan yang berbeza mengikut jenis tisu. Model PLS-DA
mencapai jumlah R2 yang tinggi dengan nilai yang sama atau hampir sama dengan
model PCA sepadan iaitu 0.679 bagi model PLS-DA sampel perikarpa (Rajah 4.9d),
0.628 bagi sampel isi (Rajah 4.9e) dan 0.680 (Rajah 4.9f) bagi sampel biji manggis.
Senarai metabolit yang mempunyai nilai melebihi 1.00 (VIP ≥1.00) telah disenaraikan
di dalam Lampiran J.

71
Analisis validasi bagi model PLS-DA yang merangkumi analisis permutasi
(Rajah 4.10) dan analisis sisa ramalan pengesahan silang-analisis varians (CV-ANOVA)
(Jadual 4.6) turut dijalankan untuk mengesahsahihkan model PLS-DA bagi setiap
sampel tisu. Berdasarkan hasil analisis validasi yang diperoleh, didapati semua model
PLS-DA adalah sah apabila dinilai berdasarkan analisis permutasi atau analisis CV-
ANOVA atau kedua-duanya. Berdasarkan analisis permutasi (Rajah 4.10), ketiga-tiga
model PLS-DA memberikan nilai R2Y asal (simbol bulat berwarna hijau pada
kedudukan paling kanan) dan nilai Q2Y asal (simbol kotak berwarna biru pada
kedudukan paling kanan) yang lebih tinggi berbanding nilai permutasi R2Y dan Q2Y di
sebelah kiri.
Selain itu, garisan regresi berwarna biru bagi titik Q2 bersilang dengan paksi
menegak (Y) pada sebelah kiri pada kedudukan (0.0, -0.415) bagi validasi model PLS-
DA perikarpa (Rajah 4.10a), (0.0, -0.503) bagi validasi model PLS-DA sampel isi
(Rajah 4.10b) dan (0.0, -0.416) bagi validasi model PLS-DA sampel biji manggis
(Rajah 4.10c). Persilangan garisan regresi Q2 pada titik negatif bagi paksi Y
menandakan bahawa model PLS-DA ini adalah sah dan tidak berlakunya overfitting (Li
et al. 2009). Manakala, analisis CV-ANOVA yang menunjukkan nilai p regresi 0.0007
bagi validasi model PLS-DA perikarpa dan 0.0104 bagi validasi model PLS-DA biji
juga membuktikan bahawa model PLS-DA adalah signifikan (p ≤0.05). Hanya nilai p
bagi validasi model sampel isi memberikan nilai melebihi 0.05 iaitu 0.5601 (Jadual 4.6).
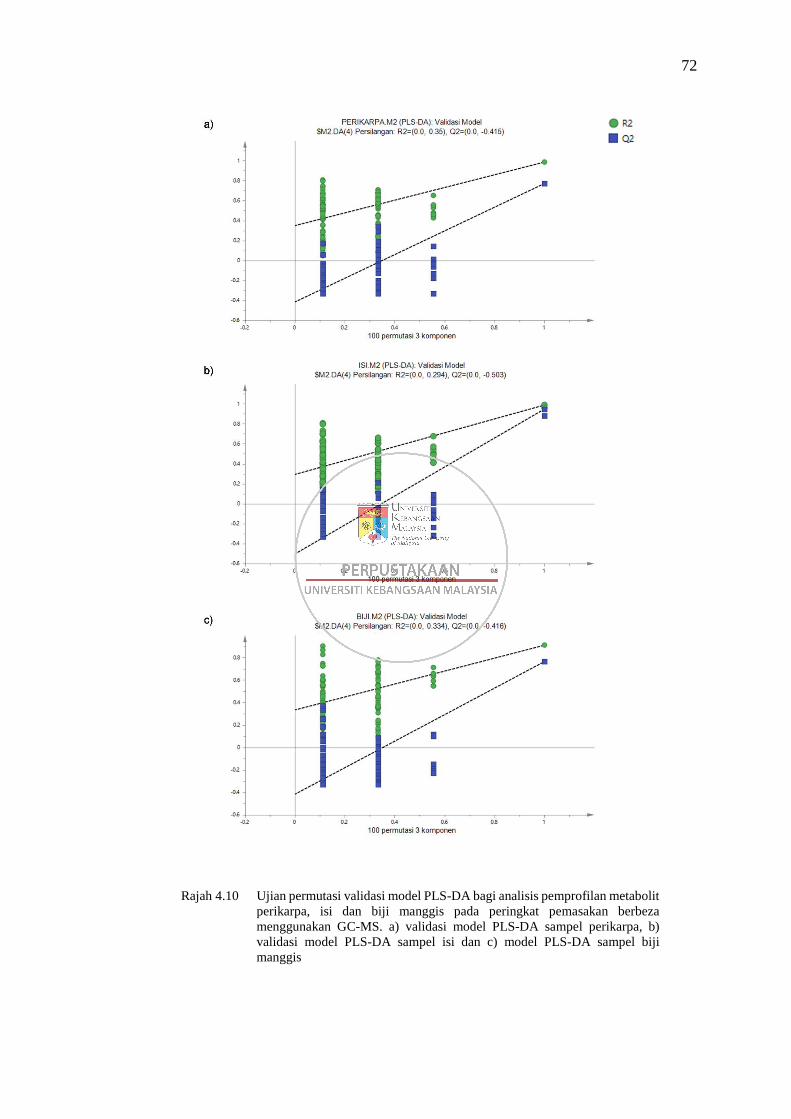
72
Rajah 4.10 Ujian permutasi validasi model PLS-DA bagi analisis pemprofilan metabolit
perikarpa, isi dan biji manggis pada peringkat pemasakan berbeza
menggunakan GC-MS. a) validasi model PLS-DA sampel perikarpa, b)
validasi model PLS-DA sampel isi dan c) model PLS-DA sampel biji
manggis
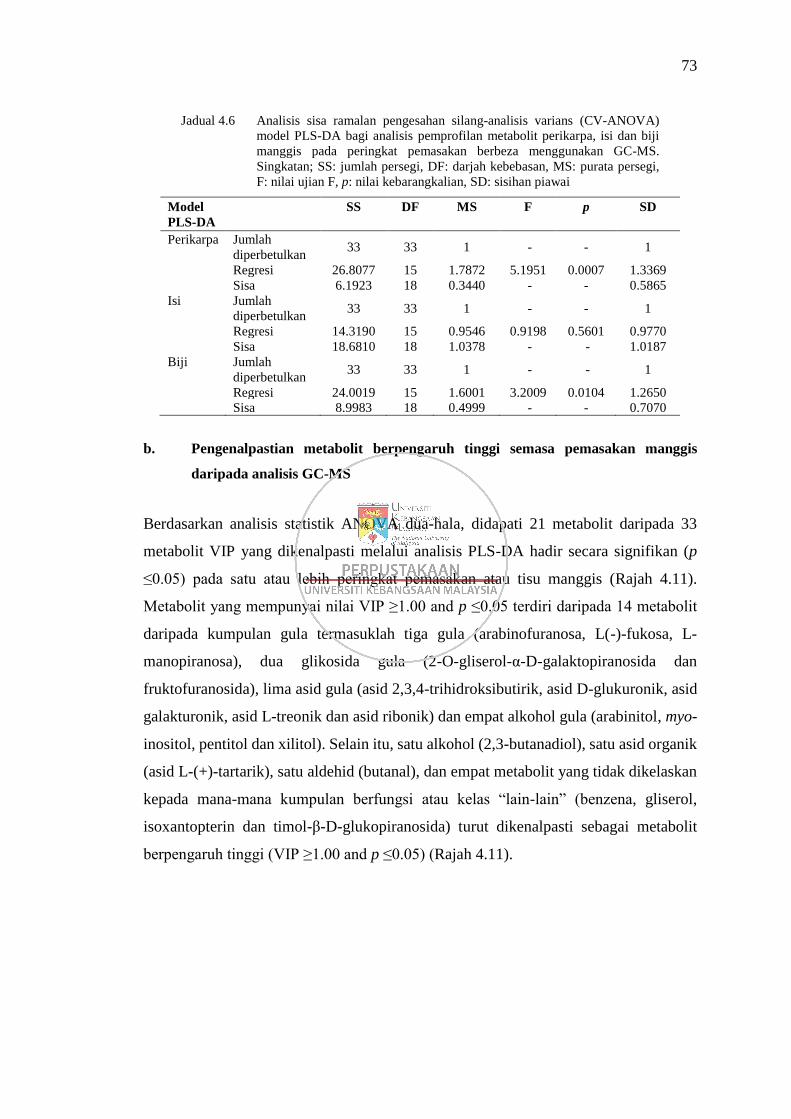
73
Jadual 4.6 Analisis sisa ramalan pengesahan silang-analisis varians (CV-ANOVA)
model PLS-DA bagi analisis pemprofilan metabolit perikarpa, isi dan biji
manggis pada peringkat pemasakan berbeza menggunakan GC-MS.
Singkatan; SS: jumlah persegi, DF: darjah kebebasan, MS: purata persegi,
F: nilai ujian F, p: nilai kebarangkalian, SD: sisihan piawai
Model
PLS-DA
SS DF MS F p SD
Perikarpa Jumlah
diperbetulkan 33 33 1 - - 1
Regresi 26.8077 15 1.7872 5.1951 0.0007 1.3369
Sisa 6.1923 18 0.3440 - - 0.5865
Isi Jumlah
diperbetulkan 33 33 1 - - 1
Regresi 14.3190 15 0.9546 0.9198 0.5601 0.9770
Sisa 18.6810 18 1.0378 - - 1.0187
Biji Jumlah
diperbetulkan 33 33 1 - - 1
Regresi 24.0019 15 1.6001 3.2009 0.0104 1.2650
Sisa 8.9983 18 0.4999 - - 0.7070
b. Pengenalpastian metabolit berpengaruh tinggi semasa pemasakan manggis
daripada analisis GC-MS
Berdasarkan analisis statistik ANOVA dua-hala, didapati 21 metabolit daripada 33
metabolit VIP yang dikenalpasti melalui analisis PLS-DA hadir secara signifikan (p
≤0.05) pada satu atau lebih peringkat pemasakan atau tisu manggis (Rajah 4.11).
Metabolit yang mempunyai nilai VIP ≥1.00 and p ≤0.05 terdiri daripada 14 metabolit
daripada kumpulan gula termasuklah tiga gula (arabinofuranosa, L(-)-fukosa, L-
manopiranosa), dua glikosida gula (2-O-gliserol-α-D-galaktopiranosida dan
fruktofuranosida), lima asid gula (asid 2,3,4-trihidroksibutirik, asid D-glukuronik, asid
galakturonik, asid L-treonik dan asid ribonik) dan empat alkohol gula (arabinitol, myo-
inositol, pentitol dan xilitol). Selain itu, satu alkohol (2,3-butanadiol), satu asid organik
(asid L-(+)-tartarik), satu aldehid (butanal), dan empat metabolit yang tidak dikelaskan
kepada mana-mana kumpulan berfungsi atau kelas “lain-lain” (benzena, gliserol,
isoxantopterin dan timol-β-D-glukopiranosida) turut dikenalpasti sebagai metabolit
berpengaruh tinggi (VIP ≥1.00 and p ≤0.05) (Rajah 4.11).
74
Rajah 4.11 Graf metabolit berpengaruh tinggi (VIP ≥1.00 and p ≤0.05) bagi kumpulan metabolit gula (a), glikosida gula (b), asid gula (c), alkohol gula (d),
alkohol (e), asid organik (f), aldehid (g) dan lain-lain (h). Data diperoleh daripada analisis pemprofilan metabolit perikarpa, isi dan biji manggis
pada peringkat pemasakan berbeza menggunakan analisis GC-MS. Paksi Y menunjukkan nilai kawasan puncak relatif (kpr) manakala nilai
purata ralat piawai daripada tiga replikat biologi digunakan sebagai bar ralat. Singkatan; PO: peringkat 0, P2: peringkat 2, P4: peringkat 4, P6:
peringkat 6

75
Kadar metabolit berpengaruh tinggi yang dikenalpasti adalah berbeza antara
peringkat pemasakan atau jenis tisu manggis atau kedua-duanya. Contohnya, hanya
arabinofuranosa dikenalpasti seawal peringkat pertama pemasakan iaitu peringkat 0
manakala metabolit gula yang lain hanya mula dikesan pada peringkat 2 di dalam tisu
yang berlainan iaitu di dalam biji bagi L(-)-fukosa dan perikarpa bagi L-manopiranosa
(Rajah 4.11a). Kadar arabinofuranosa kemudian menurun terutamanya di dalam tisu
perikarpa dan biji apabila menghampiri peringkat akhir pemasakan manakala L-
manopiranosa tinggi di dalam perikarpa pada peringkat 2 dan 6.
Selain itu, aras metabolit 2-O-gliserol-α-D-galaktopiranosida tinggi di dalam
tisu isi pada peringkat pertengahan hingga akhir pemasakan manakala fruktofuranosida
hanya hadir pada peringkat 4 (tisu isi) (Rajah 4.11b). Bagi metabolit berpengaruh tinggi
dari kelas asid gula pula (Rajah 4.11c), hanya asid galakturonik dan asid ribonik hadir
pada peringkat 0. Asid ribonik juga didapati hadir dalam ketiga-tiga tisu manggis pada
peringkat pemasakan yang tertentu. Manakala, asid gula yang lain (asid 2,3,4-
trihidroksibutirik, asid D-glukuronik dan asid L-treonik) hanya dikesan seawal
peringkat 2 dan ke atas pada kadar yang berbeza. Asid 2,3,4-trihidroksibutirik
(peringkat 4), asid D-glukuronik (peringkat 2) dan asid L-treonik (peringkat 2 hingga
6) didapati tinggi di dalam tisu biji manggis berbanding tisu yang lain (Rajah 4.11c).
Selain itu, untuk kelas alkohol gula, terdapat metabolit seperti arabinitol dan
myo-inositol telah hadir pada seawal peringkat 0 (Rajah 4.11d). Arabinitol
kemudiannya meningkat sehingga akhir pemasakan pada isi manggis manakala myo-
inositol pula kekal tinggi di dalam tisu biji berbanding tisu yang lain. Selain itu, pentitol
pula hanya ditemui pada peringkat 2 (biji) dan xilitol didapati unik kepada peringkat 4
pemasakan (perikarpa dan isi) (Rajah 4.11d). Metabolit lain seperti 2,3-butanadiol, L-
(+)-tartarik dan butanal juga didapati hadir seawal peringkat 0. Aras metabolit 2,3-
butanadiol kemudian meningkat dalam perikarpa peringkat 2 dan didapati tinggi pada
peringkat 4 dan 6 di dalam tisu isi (Rajah 4.11e). Manakala, asid L-(+)-tartarik
menunjukkan kadar paling tinggi dalam sampel perikarpa (peringkat 2 dan 4)
berbanding sampel isi (peringkat 4 dan 6) (Rajah 4.11f). Butanal pula didapati
meningkat dalam tisu isi dan biji pada peringkat 2 kepada peringkat 4 namun menurun
semasa peringkat 6 (Rajah 4.11g).

76
Manakala, metabolit daripada kumpulan “lain-lain” dilihat bersifat lebih
spesifik terhadap jenis tisu manggis. Contohnya, benzena hanya dikesan di dalam biji
(peringkat 2 dan 6) manakala isoxantopterin hanya hadir di dalam isi (peringkat 2) dan
timol-β-D-glukopiranosida unik kepada perikarpa manggis (peringkat 2). Gliserol pula
dikenalpasti di dalam ketiga-tiga tisu manggis pada peringkat 2 atau 4 (Rajah 4.11h).
4.2.2 Pemprofilan metabolit putatif menggunakan analisis LC-MS
Selain daripada analisis GC-MS, analisis LC-MS bukan sasaran juga telah dijalankan
dalam kajian ini bagi memprofilkan metabolit dalam perikarpa, isi dan biji manggis
pada peringkat 0, 2, 4 dan 6. Sampel manggis diekstrak menggunakan kaedah yang
telah dioptimumkan iaitu kaedah pengekstrakan 1 (metanol/asid formik dengan nisbah
599:1 disertai teknik sonikasi) kerana kaedah ini telah menunjukkan hasil
pengekstrakan metabolit sekunder yang paling berkesan berbanding kaedah-kaedah
yang lain (kaedah 2, 3, 4 dan 5) (Rajah 4.1). Mod pengionan LC-MS ditetapkan kepada
mod negatif kerana ekstrak manggis daripada kaedah pengekstrakan 1 didapati lebih
sesuai dianalisis pada mod negatif berdasarkan penghasilan puncak yang lebih banyak
iaitu 58 puncak berbanding 41 puncak pada mod positif (Jadual 4.5).
Hasilnya, sebanyak 1919 nilai RT: m/z (masa pengekalan: nisbah jisim kepada
caj) berjaya diekstrak daripada perisian ProfileAnalysis. Kemudian, data ditapis dengan
hanya mengambil kira metabolit yang hadir dalam ketiga-tiga replikat biologi dalam
sekurang-kurangnya satu kumpulan sampel yang dikaji. Dengan itu, sebanyak 416 RT:
m/z telah dipilih untuk proses seterusnya iaitu analisis statistik dan pengenalpastian
metabolit. Analisis varians dua-hala (two-way ANOVA) telah dijalankan dan hasil
analisis mendapati sebanyak 354 RT: m/z adalah signifikan secara statistik (p ≤0.05)
sama ada antara peringkat pemasakan, jenis tisu atau interaksi antara pembolehubah
peringkat pemasakan dan jenis tisu yang dibandingkan. Manakala, RT: m/z yang
selebihnya (62) adalah tidak signifikan secara statistik.
Daripada 416 RT: m/z yang diperoleh ini, sebanyak 98 metabolit telah berjaya
dikenalpasti (Lampiran K) menggunakan pangkalan data METLIN dan MetFrag
menerusi pencarian terhadap pangkalan data umum seperti KEGG, ChEBI, HMDB dan
MetaCyc. Walau bagaimanapun, terdapat juga sebanyak 102 RT: m/z yang tidak dapat

77
dikenalpasti dan telah dilaporkan di dalam Lampiran L. Kemudian, analisis multivariat
(analisis PCA dan PLS-DA) telah dijalankan terhadap gabungan data metabolit yang
telah dikenalpasti (98 metabolit) dan data RT: m/z yang tidak dapat dikenalpasti (102
RT: m/z).
a. Analisis statistik data multivariat
Analisis data multivariat (analisis PCA dan PLS-DA) telah dijalankan untuk mengkaji
taburan metabolit antara peringkat pemasakan yang berbeza dengan melihat kepada
setiap tisu manggis yang telah dibandingkan. Analisis PCA dan PLS-DA berjaya
memisahkan kelompok metabolit mengikut setiap peringkat pemasakan (peringkat 0, 2,
4 dan 6) dengan nilai R2 melebihi 50% (0.5) seperti yang dapat dilihat dalam Rajah 4.12
di bawah. Plot skor PCA dan PLS-DA bagi sampel perikarpa memberikan nilai varians
keseluruhan (R2) yang sama iaitu 0.717 (Rajah 4.12a dan d) manakala plot skor PCA
dan PLS-DA bagi sampel isi dan biji manggis masing-masing memberikan nilai R2
setinggi 0.690 (Rajah 4.12b dan e) dan 0.723 (Rajah 4.12c dan f).
Kesemua model PCA dan PLS-DA menunjukkan pemisahan yang jelas antara
kumpulan sampel, membuktikan bahawa jenis atau kadar metabolit yang berjaya
diekstrak daripada analisis LC-MS mempunyai perbezaan yang tinggi antara peringkat
pemasakan yang berbeza di dalam manggis. Kedudukan replikat biologi bagi setiap
kelompok adalah rapat antara satu sama lain, menandakan tahap kebolehulangan
replikat yang tinggi. Analisis PLS-DA dijalankan untuk mengenalpasti metabolit VIP
yang mempengaruhi pemisahan antara kumpulan sampel yang dibandingkan. Daripada
analisis yang dijalankan, 58 metabolit dikenalpasti mempunyai nilai VIP yang tinggi
(VIP ≥1.00) (Lampiran M), sekaligus mempengaruhi pemisahan antara kumpulan
sampel dan akan dibincangkan dengan lebih terperinci pada bahagian seterusnya.
Model PLS-DA juga telah disahsahihkan melalui ujian permutasi (Rajah 4.13) dan ujian
CV-ANOVA (Jadual 4.7).
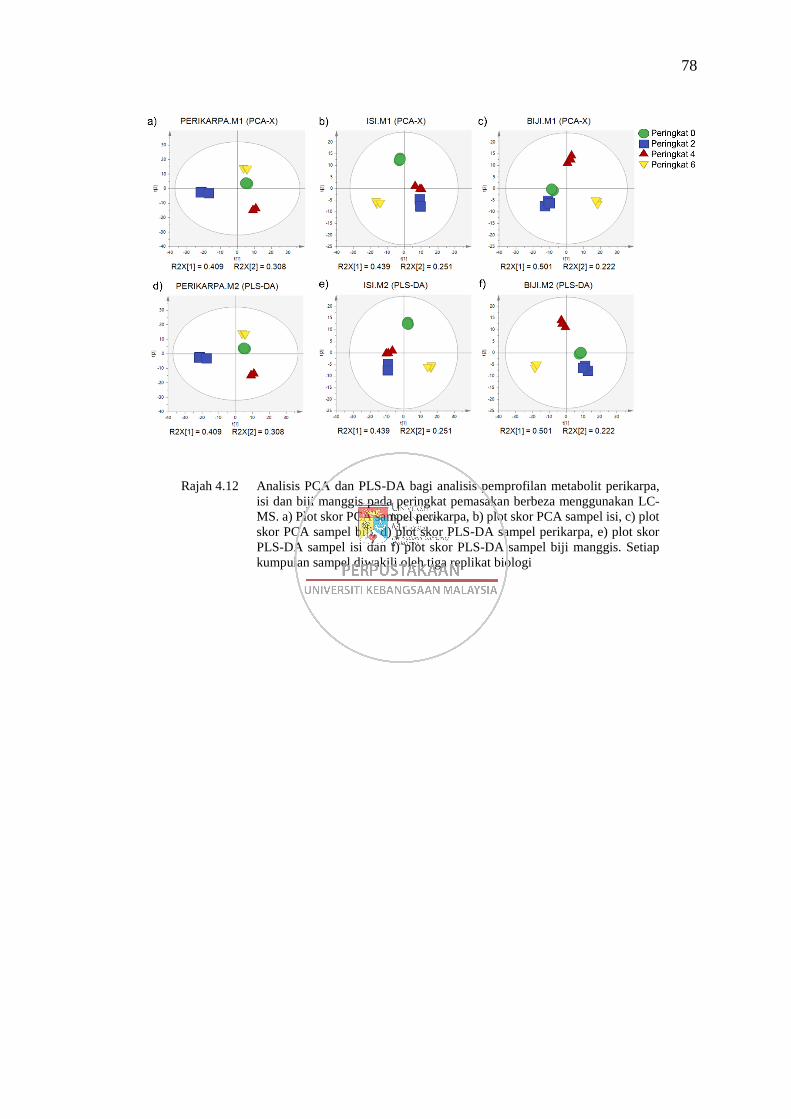
78
Rajah 4.12 Analisis PCA dan PLS-DA bagi analisis pemprofilan metabolit perikarpa,
isi dan biji manggis pada peringkat pemasakan berbeza menggunakan LC-
MS. a) Plot skor PCA sampel perikarpa, b) plot skor PCA sampel isi, c) plot
skor PCA sampel biji, d) plot skor PLS-DA sampel perikarpa, e) plot skor
PLS-DA sampel isi dan f) plot skor PLS-DA sampel biji manggis. Setiap
kumpulan sampel diwakili oleh tiga replikat biologi
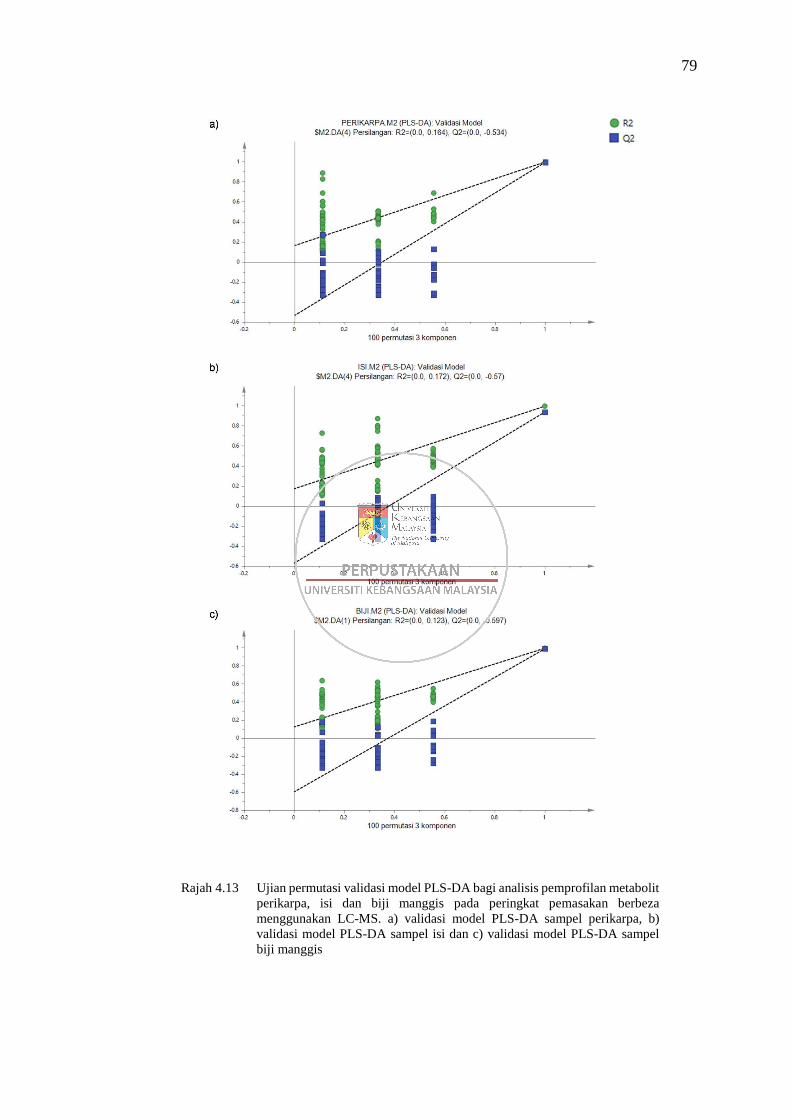
79
Rajah 4.13 Ujian permutasi validasi model PLS-DA bagi analisis pemprofilan metabolit
perikarpa, isi dan biji manggis pada peringkat pemasakan berbeza
menggunakan LC-MS. a) validasi model PLS-DA sampel perikarpa, b)
validasi model PLS-DA sampel isi dan c) validasi model PLS-DA sampel
biji manggis
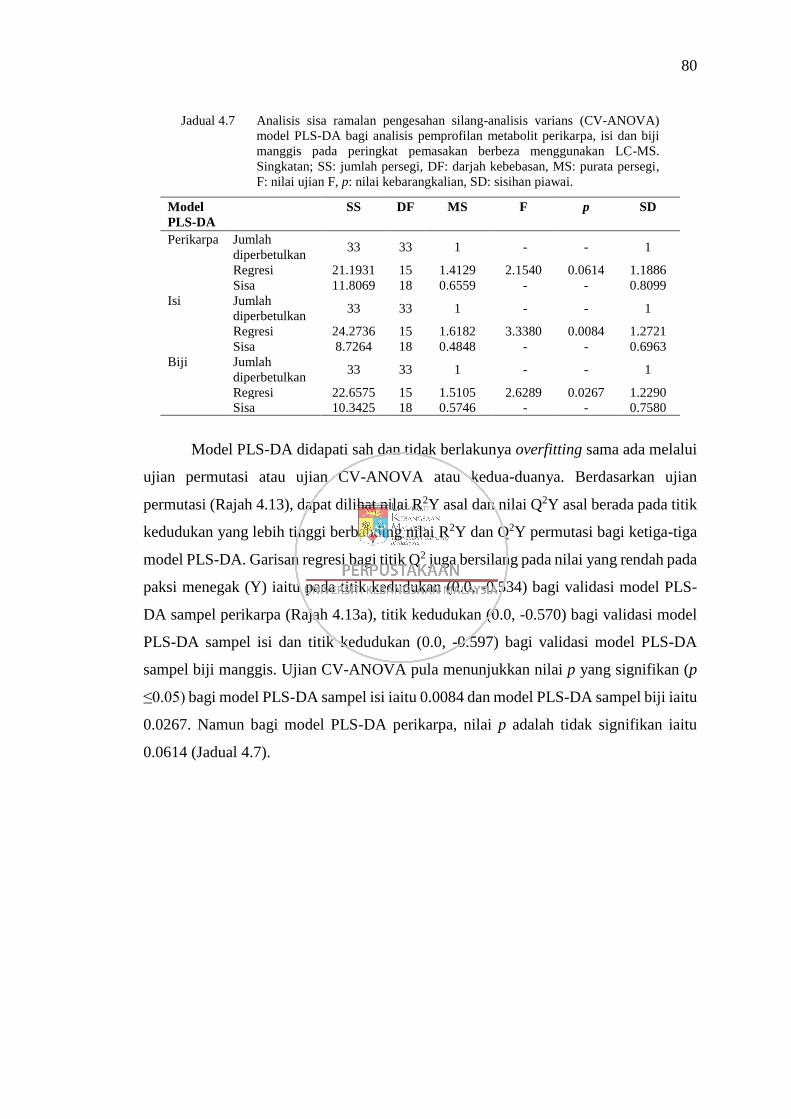
80
Jadual 4.7 Analisis sisa ramalan pengesahan silang-analisis varians (CV-ANOVA)
model PLS-DA bagi analisis pemprofilan metabolit perikarpa, isi dan biji
manggis pada peringkat pemasakan berbeza menggunakan LC-MS.
Singkatan; SS: jumlah persegi, DF: darjah kebebasan, MS: purata persegi,
F: nilai ujian F, p: nilai kebarangkalian, SD: sisihan piawai.
Model
PLS-DA
SS DF MS F p SD
Perikarpa Jumlah
diperbetulkan 33 33 1 - - 1
Regresi 21.1931 15 1.4129 2.1540 0.0614 1.1886
Sisa 11.8069 18 0.6559 - - 0.8099
Isi Jumlah
diperbetulkan 33 33 1 - - 1
Regresi 24.2736 15 1.6182 3.3380 0.0084 1.2721
Sisa 8.7264 18 0.4848 - - 0.6963
Biji Jumlah
diperbetulkan 33 33 1 - - 1
Regresi 22.6575 15 1.5105 2.6289 0.0267 1.2290
Sisa 10.3425 18 0.5746 - - 0.7580
Model PLS-DA didapati sah dan tidak berlakunya overfitting sama ada melalui
ujian permutasi atau ujian CV-ANOVA atau kedua-duanya. Berdasarkan ujian
permutasi (Rajah 4.13), dapat dilihat nilai R2Y asal dan nilai Q2Y asal berada pada titik
kedudukan yang lebih tinggi berbanding nilai R2Y dan Q2Y permutasi bagi ketiga-tiga
model PLS-DA. Garisan regresi bagi titik Q2 juga bersilang pada nilai yang rendah pada
paksi menegak (Y) iaitu pada titik kedudukan (0.0, -0.534) bagi validasi model PLS-
DA sampel perikarpa (Rajah 4.13a), titik kedudukan (0.0, -0.570) bagi validasi model
PLS-DA sampel isi dan titik kedudukan (0.0, -0.597) bagi validasi model PLS-DA
sampel biji manggis. Ujian CV-ANOVA pula menunjukkan nilai p yang signifikan (p
≤0.05) bagi model PLS-DA sampel isi iaitu 0.0084 dan model PLS-DA sampel biji iaitu
0.0267. Namun bagi model PLS-DA perikarpa, nilai p adalah tidak signifikan iaitu
0.0614 (Jadual 4.7).

81
b. Pengenalpastian metabolit berpengaruh tinggi dalam pemasakan manggis
daripada analisis LC-MS
Daripada 58 metabolit VIP (VIP ≥1.00) yang dikenalpasti melalui analisis multivariat
PLS-DA, didapati 55 metabolit hadir secara signifikan (p ≤0.05) di dalam sampel
manggis sama ada antara peringkat pemasakan berbeza, tisu yang berbeza atau interaksi
antara pembolehubah peringkat pemasakan dan jenis tisu (Rajah 4.14 dan Rajah 4.15).
Berdasarkan Rajah 4.14, metabolit berpengaruh tinggi ini merangkumi beberapa kelas
metabolit primer dan sekunder termasuklah satu metabolit gula (sukrosa), satu asid gula
(asid L-askorbik-2-glukosida), satu fosfat gula (D-glukosa 6-fosfat), satu glikosida (asid
L-askorbik-2-glukosida), dua asid organik (asid 2-butinaioik dan asid kuinik) dan satu
asid lemak iaitu 1-(3-metilbutanoil)-6-apiosilglukosa. Selain itu, turut dikenalpasti
adalah lima asid amino dan terbitan iaitu S-adenosilhomosisteina, asid aspartik, (S)-N-
[3-(3,4-metilenadioksifenil)-2-(asetiltio)metil-1-oksooprolil]-(S)-alanina benzil ester,
Asn-Trp-OH dan temokaprilat serta satu kuinon iaitu knifolona.
Sebanyak empat alkaloid (6-hirdoksiprotopina, normorfin 3-glukuronida,
oksinarkotina dan xantina), enam terpenoid ((asid (+)-absisik, giberelin A3, ginggolida
C, katalposida, montanol dan salvinorin A)) dan empat xanton (α-mangostin,
astikolorin B, gartanin dan kratoksiarborenon C) (Rajah 4.14) turut didapati mempunyai
nilai VIP ≥1.00 dan p ≤0.05). Selain itu, metabolit yang turut dikenalpasti dengan
jumlah tertinggi (19 metabolit) adalah metabolit daripada kumpulan fenolik seperti
epirobinatinidol-(4β,8)-katesin, kuersetin 3,3'-dimetil eter 4'-glukosida, neoisostegana
dan sebagainya (Rajah 4.15). Metabolit yang tidak dikelaskan ke dalam mana-mana
kumpulan turut dikenalpasti sebagai metabolit berpengaruh tinggi sebanyak lapan
metabolit seperti asid meso-tartarik monohidrat, fisalin K, melampodinin dan
sebagainya (Rajah 4.15).
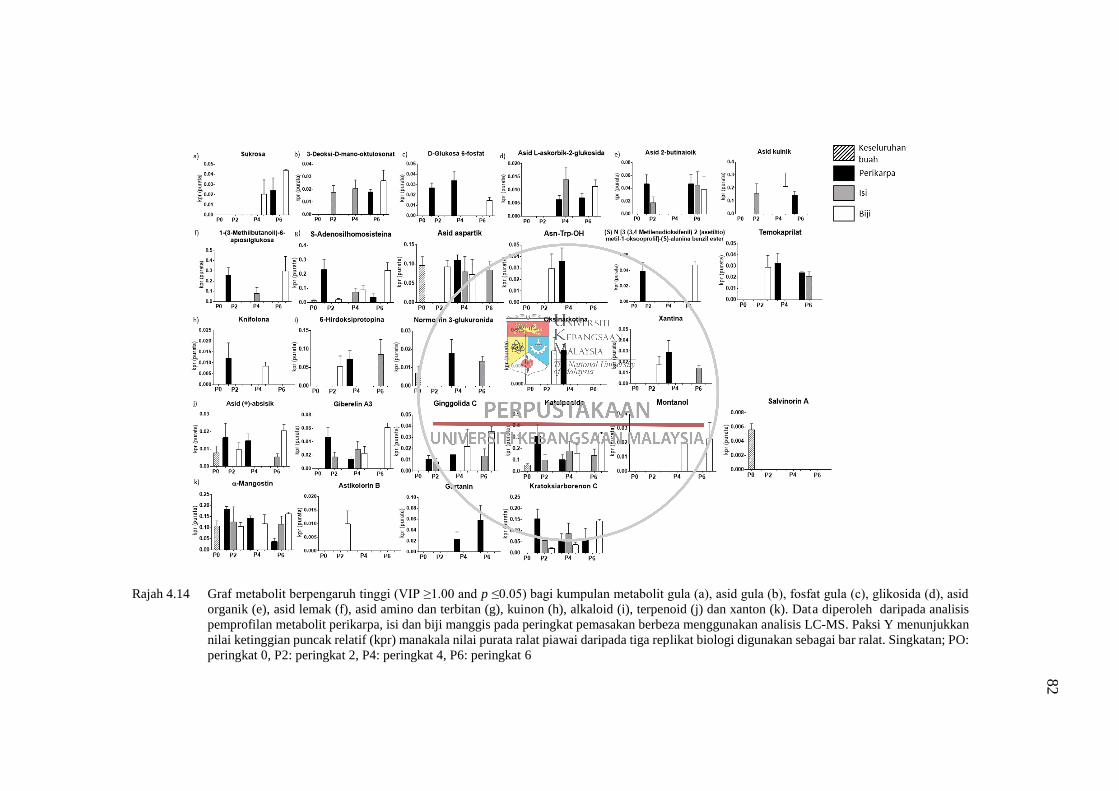
82
Rajah 4.14 Graf metabolit berpengaruh tinggi (VIP ≥1.00 and p ≤0.05) bagi kumpulan metabolit gula (a), asid gula (b), fosfat gula (c), glikosida (d), asid
organik (e), asid lemak (f), asid amino dan terbitan (g), kuinon (h), alkaloid (i), terpenoid (j) dan xanton (k). Data diperoleh daripada analisis
pemprofilan metabolit perikarpa, isi dan biji manggis pada peringkat pemasakan berbeza menggunakan analisis LC-MS. Paksi Y menunjukkan
nilai ketinggian puncak relatif (kpr) manakala nilai purata ralat piawai daripada tiga replikat biologi digunakan sebagai bar ralat. Singkatan; PO:
peringkat 0, P2: peringkat 2, P4: peringkat 4, P6: peringkat 6

83
Rajah 4.15 Graf metabolit berpengaruh tinggi (VIP ≥1.00 and p ≤0.05) bagi kumpulan metabolit fenolik (a) dan lain-lain (b). Data diperoleh daripada
analisis pemprofilan metabolit perikarpa, isi dan biji manggis pada peringkat pemasakan berbeza menggunakan analisis LC-MS. Paksi Y
menunjukkan nilai ketinggian puncak relatif (kpr) manakala nilai purata ralat piawai daripada tiga replikat biologi digunakan sebagai bar ralat.
Singkatan; PO: peringkat 0, P2: peringkat 2, P4: peringkat 4, P6: peringkat 6 dan ES-242-4: (3S,3'S,4S,4'S)-7,7',9,9'-Tetrametoksi-3,3'-dimetil-
3,3',4,4'-tetrahidro-1H,1'H-5,5'-bibenzo[g]isokromena-4,4',10,10'-tetrol

84
Kadar metabolit berpengaruh tinggi (VIP ≥1.00 and p ≤0.05) adalah berbeza
mengikut peringkat pemasakan atau tisu manggis yang berbeza. Sukrosa tidak dapat
dikesan pada peringkat awal pemasakan (peringkat 0 dan 2) namun didapati meningkat di
dalam tisu biji dan perikarpa pada peringkat akhir pemasakan iaitu peringkat 6 (Rajah
4.14a). Beberapa metabolit lain seperti 3-deoksi-D-mano-oktulosonat, D-glukosa 6-fosfat,
asid L-askorbik-2-glukosida, 2-butinaioik dan asid kuinik juga tidak ditemui pada
peringkat 0 namun ditemui pada peringkat 2 atau seterusnya. Contohnya, 3-deoksi-D-
mano-oktulosonat dikesan di dalam tisu isi pada peringkat 2 dan 4 dan di dalam perikarpa
dan biji pada peringkat 6) (Rajah 4.14b). D-glukosa 6-fosfat pula hadir di dalam perikarpa
(peringkat 2 dan 4) dan biji (peringkat 6) (Rajah 4.14c). Asid L-askorbik-2-glukosida hadir
pada kadar yang hampir konsisten di dalam perikarpa pada peringkat 4 dan 6 dan turut
menunjukkan kadar yang tinggi dalam isi (peringkat 4) dan biji (peringkat 6) (Rajah 4.14d).
Asid 2-butinaioik pula tinggi di dalam tisu perikarpa dan isi pada peringkat 2 dan 6
manakala asid kuinik pula hadir pada peringkat 2 (isi), 4 (biji) dan 6 (perikarpa) (Rajah
4.14e).
Daripada enam asid amino dan terbitan yang dikenalpasti sebagai metabolit
berpengaruh tinggi, hanya S-adenosilhomosisteina dan asid aspartik dikesan seawal
peringkat 0 pemasakan (Rajah 4.14f). S-adenosilhomosisteina kemudian meningkat dalam
perikarpa dan menurun kembali pada peringkat 6. Sementara dalam tisu biji, metabolit ini
meningkat sehingga peringkat 6. Asid aspartik pula tinggi pada peringkat 0 dan kekal tinggi
pada peringkat 2 (biji), peringkat 4 (perikarpa, isi dan biji) dan peringkat 6 (isi). Sementara
itu, Asn-Trp-OH, (S)-N-[3-(3,4-metilenadioksifenil)-2-(asetiltio)metil-1-oksooprolil]-(S)-
alanina benzil ester dan temokaprilat pula hanya dikenalpasti bermula peringkat 2 sama ada
dalam perikarpa atau biji manggis (Rajah 4.14g).
Metabolit alkaloid iaitu 6-hirdoksiprotopina, normorfin 3-glukuronida,
oksinarkotina dan xantina didapati bersifat spesifik terhadap jenis tisu pada peringkat
pemasakan yang tertentu (Rajah 4.14i). Contohnya, semua alkaloid hanya dikenalpasti di
dalam perikarpa semasa peringkat 4 sahaja manakala tiga daripadanya iaitu 6-

85
hirdoksiprotopina, normorfin 3-glukuronida dan xantina hanya ditemui dalam isi manggis
pada peringkat 6. Sementara itu, kehadiran 6-hirdoksiprotopina, oksinarkotina dan xantina
dalam biji manggis hanya dikenalpasti pada peringkat 2 (Rajah 4.14i).
Seterusnya, tiga daripada enam terpenoid yang dikenalpasti sebagai metabolit
berpengaruh tinggi iaitu asid (+)-absisik, katalposida dan salvinorin A dikenalpasti pada
peringkat 0 (Rajah 4.14j). Salvinorin A tidak ditemui dalam mana-mana tisu pada peringkat
2 dan seterusnya tetapi asid (+)-absisik dan katalposida serta dua lagi terpenoid iaitu
giberelin A3 dan ginggolida C didapati meningkat dalam tisu biji menuju peringkat akhir
pemasakan. Penurunan pada kadar giberelin A3 dan katalposida dapat dilihat berlaku dalam
perikarpa daripada peringkat 2 kepada peringkat 4. Montanol pula unik kepada tisu biji
(peringkat 4 dan 6) dan hadir pada kadar yang tinggi (Rajah 4.14j).
Kumpulan metabolit bioaktif utama dalam buah manggis iaitu xanton turut
dikenalpasti sebagai metabolit berpengaruh tinggi dalam pemasakan manggis (Rajah
4.14k). Berdasarkan kumpulan fungsi, xanton telah dikategorikan dalam kelas polifenol
(fenolik), namun dalam kajian ini, xanton akan dibincangkan sebagai satu kumpulan
metabolit yang berasingan daripada kumpulan fenolik. Dua daripada empat xanton yang
dikenalpasti sebagai metabolit berpengaruh tinggi iaitu α-mangostin dan kratoksiarborenon
C menunjukkan penurunan dalam perikarpa manggis sepanjang proses pemasakan.
Manakala dalam biji manggis pula, kadar kedua-dua metabolit ini meningkat sewaktu
proses pemasakan. α-Mangostin dan kratoksiarborenon C juga hadir di dalam tisu isi pada
peringkat 2, tetapi hanya hadir pada peringkat 6 untuk α-mangostin dan peringkat 4 untuk
kratoksiarborenon C bagi tisu yang sama. Manakala astikolorin B hanya dikenalpasti dalam
sampel biji (peringkat 2) dan gartanin hanya hadir dan didapati meningkat daripada
peringkat 4 ke 6 bagi sampel perikarpa (Rajah 4.14k).
Berdasarkan Rajah 4.15a, hanya enam daripada 19 fenolik iaitu 4'-metil-(-)-
epigalokatesin 3'-glukuronida, ginketin, kerkosporin, kuersetin 3,3'-dimetil eter 4'-
glukosida, leufolin A dan metilofiopogonona B didapati hadir pada peringkat 0 pemasakan.
Kemudian, 4'-metil-(-)-epigalokatesin 3'-glukuronida didapati meningkat dalam tisu biji

86
pada peringkat 6 bersama beberapa metabolit lain seperti 3-glukosil-2,3',4,4',6-
pentahidroksibenzofenon, 5,7,3'-trihidroksi-6,4',5'-trimetoksiflavanon dan bis-N-butil
ftalat. Bis-N-butil ftalat dan beberapa fenolik lain seperti epirobinatinidol-(4β,8)-katesin,
kuersetin 3,3'-dimetil eter 4'-glukosida, leufolin A, neoisostegana, okrokarpin E dan
soiuflavon A hadir dalam tisu perikarpa pada kadar yang tinggi pada peringkat 2. Kesemua
metabolit ini kemudiannya menurun dalam perikarpa apabila memasuki peringkat 4 tetapi
dua daripadanya iaitu bis-N-butil ftalat dan epirobinatinidol-(4β,8)-katesin kembali
meningkat dalam tisu yang sama pada peringkat akhir pemasakan manakala metabolit yang
selebihnya (kuersetin 3,3'-dimetil eter 4'-glukosida, leufolin A, neoisostegana, okrokarpin
E dan soiuflavon A) tidak ditemui pada peringkat 6 ini. Sebaliknya, leufolin A,
neoisostegana, okrokarpin E dan soiuflavon A didapati semakin meningkat dalam tisu biji
pada peringkat 6 (Rajah 4.15a).
Antara metabolit lain yang turut dikenalpasti mempunyai nilai VIP ≥1.00 dan p
≤0.05 adalah fisalin K dan melampodinin yang dikesan seawal peringkat 0 (Rajah 4.15b).
Fisalin K tidak ditemui dalam mana-mana tisu pada peringkat seterusnya manakala
melampodinin hadir pada kadar yang tinggi dalam tisu biji pada peringkat 2 dan mengalami
penurunan pada peringkat 4 dan 6. Sebaliknya, rodomirtoksin pula meningkat dalam tisu
yang sama sepanjang proses pemasakan. Rubriflordilakton B pula unik kepada tisu
perikarpa (peringkat 6).
c. Analisis LC-MS/MS
Analisis LC-MS/MS yang merangkumi analisis Auto-MS dan analisis LC-MS/MS sasaran
menggunakan teknik pemantauan tindak balas berganda (MRM) telah dijalankan untuk
mengenalpasti identiti metabolit yang diperoleh dari analisis LC-MS sebelum ini dengan
lebih tepat berdasarkan maklumat fragmentasi ion yang diperoleh. Dua metabolit piawai
iaitu α-mangostin dan γ-mangostin turut dianalisis bagi membandingkan fragmentasi ion
yang terhasil.
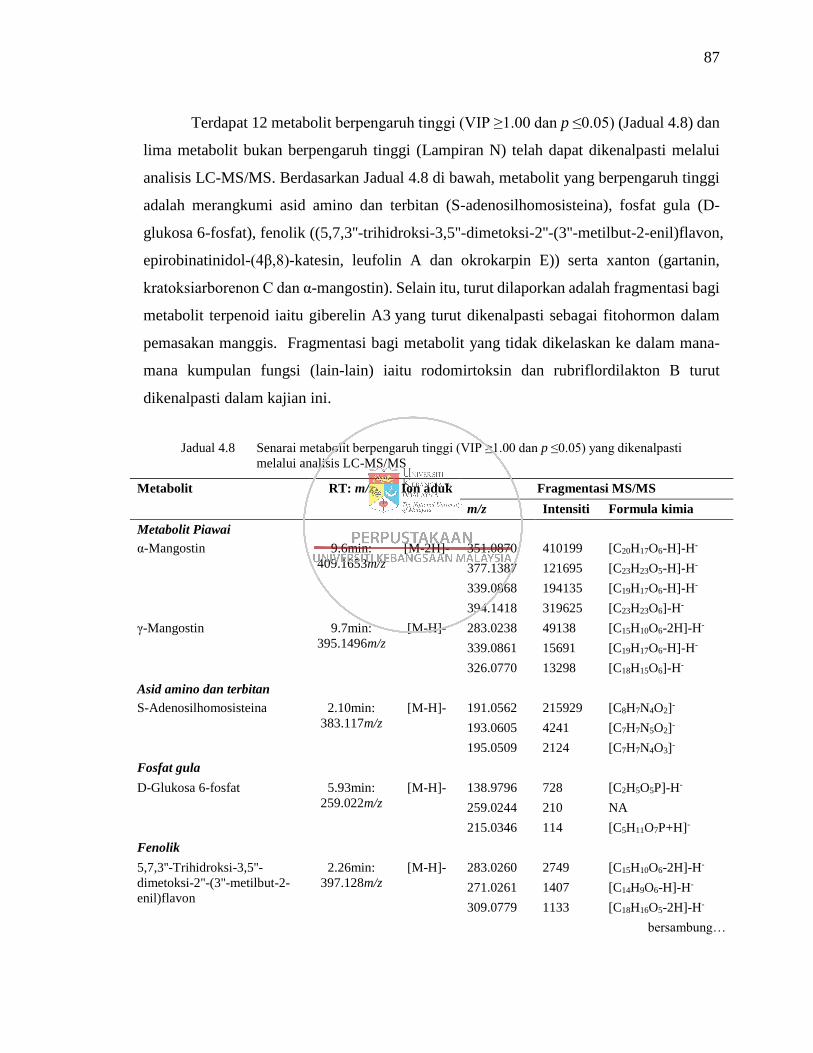
87
Terdapat 12 metabolit berpengaruh tinggi (VIP ≥1.00 dan p ≤0.05) (Jadual 4.8) dan
lima metabolit bukan berpengaruh tinggi (Lampiran N) telah dapat dikenalpasti melalui
analisis LC-MS/MS. Berdasarkan Jadual 4.8 di bawah, metabolit yang berpengaruh tinggi
adalah merangkumi asid amino dan terbitan (S-adenosilhomosisteina), fosfat gula (D-
glukosa 6-fosfat), fenolik ((5,7,3''-trihidroksi-3,5''-dimetoksi-2''-(3''-metilbut-2-enil)flavon,
epirobinatinidol-(4β,8)-katesin, leufolin A dan okrokarpin E)) serta xanton (gartanin,
kratoksiarborenon C dan α-mangostin). Selain itu, turut dilaporkan adalah fragmentasi bagi
metabolit terpenoid iaitu giberelin A3 yang turut dikenalpasti sebagai fitohormon dalam
pemasakan manggis. Fragmentasi bagi metabolit yang tidak dikelaskan ke dalam mana-
mana kumpulan fungsi (lain-lain) iaitu rodomirtoksin dan rubriflordilakton B turut
dikenalpasti dalam kajian ini.
Jadual 4.8 Senarai metabolit berpengaruh tinggi (VIP ≥1.00 dan p ≤0.05) yang dikenalpasti
melalui analisis LC-MS/MS
Metabolit RT: m/z Ion aduk Fragmentasi MS/MS
m/z Intensiti Formula kimia
Metabolit Piawai
α-Mangostin 9.6min:
409.1653m/z
[M-2H]- 351.0870 410199 [C20H17O6-H]-H-
377.1387 121695 [C23H23O5-H]-H-
339.0868 194135 [C19H17O6-H]-H-
394.1418 319625 [C23H23O6]-H-
γ-Mangostin 9.7min:
395.1496m/z
[M-H]- 283.0238 49138 [C15H10O6-2H]-H-
339.0861 15691 [C19H17O6-H]-H-
326.0770 13298 [C18H15O6]-H-
Asid amino dan terbitan
S-Adenosilhomosisteina 2.10min:
383.117m/z
[M-H]- 191.0562 215929 [C8H7N4O2]-
193.0605 4241 [C7H7N5O2]-
195.0509 2124 [C7H7N4O3]-
Fosfat gula
D-Glukosa 6-fosfat
5.93min:
259.022m/z
[M-H]- 138.9796 728 [C2H5O5P]-H-
259.0244 210 NA
215.0346 114 [C5H11O7P+H]-
Fenolik
5,7,3''-Trihidroksi-3,5''-
dimetoksi-2''-(3''-metilbut-2-
enil)flavon
2.26min:
397.128m/z
[M-H]- 283.0260 2749 [C15H10O6-2H]-H-
271.0261 1407 [C14H9O6-H]-H-
309.0779 1133 [C18H16O5-2H]-H-
bersambung…
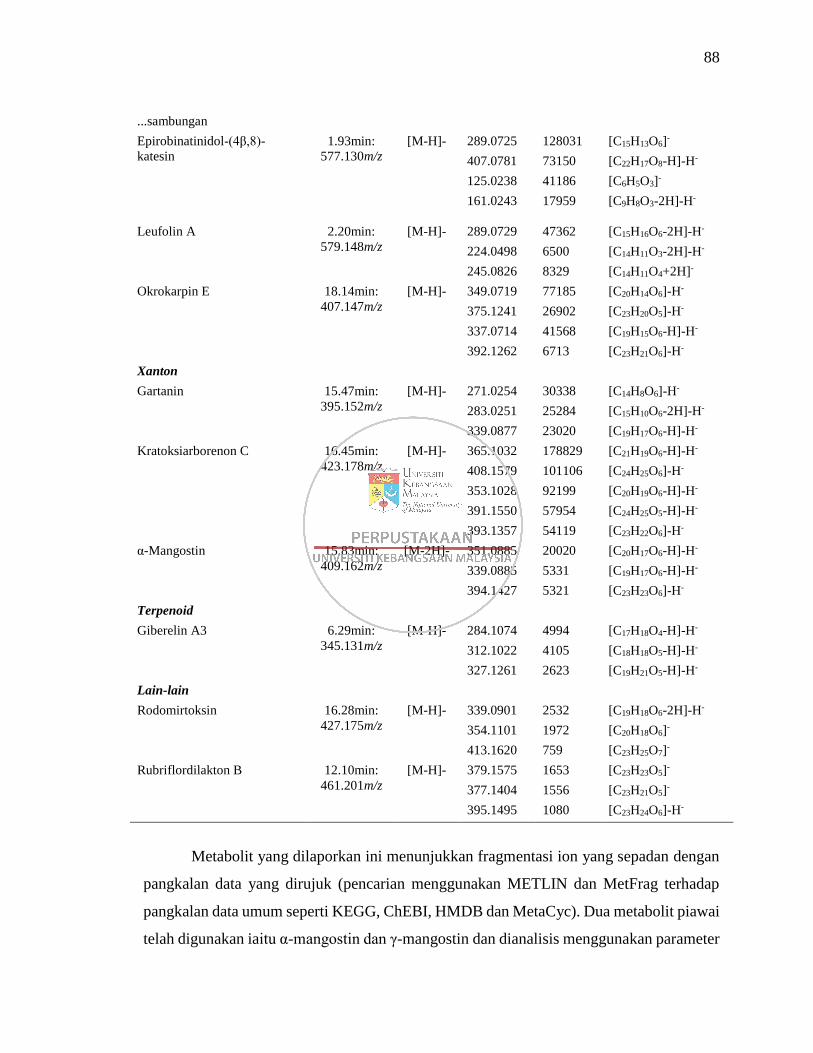
88
...sambungan
Epirobinatinidol-(4β,8)-
katesin
1.93min:
577.130m/z
[M-H]- 289.0725 128031 [C15H13O6]-
407.0781 73150 [C22H17O8-H]-H-
125.0238 41186 [C6H5O3]-
161.0243 17959 [C9H8O3-2H]-H-
Leufolin A 2.20min:
579.148m/z
[M-H]- 289.0729 47362 [C15H16O6-2H]-H-
224.0498 6500 [C14H11O3-2H]-H-
245.0826 8329 [C14H11O4+2H]-
Okrokarpin E 18.14min:
407.147m/z
[M-H]- 349.0719 77185 [C20H14O6]-H-
375.1241 26902 [C23H20O5]-H-
337.0714 41568 [C19H15O6-H]-H-
392.1262 6713 [C23H21O6]-H-
Xanton
Gartanin 15.47min:
395.152m/z
[M-H]- 271.0254 30338 [C14H8O6]-H-
283.0251 25284 [C15H10O6-2H]-H-
339.0877 23020 [C19H17O6-H]-H-
Kratoksiarborenon C 16.45min:
423.178m/z
[M-H]- 365.1032 178829 [C21H19O6-H]-H-
408.1579 101106 [C24H25O6]-H-
353.1028 92199 [C20H19O6-H]-H-
391.1550 57954 [C24H25O5-H]-H-
393.1357 54119 [C23H22O6]-H-
α-Mangostin 15.83min:
409.162m/z
[M-2H]- 351.0885 20020 [C20H17O6-H]-H-
339.0885 5331 [C19H17O6-H]-H-
394.1427 5321 [C23H23O6]-H-
Terpenoid
Giberelin A3 6.29min:
345.131m/z
[M-H]- 284.1074 4994 [C17H18O4-H]-H-
312.1022 4105 [C18H18O5-H]-H-
327.1261 2623 [C19H21O5-H]-H-
Lain-lain
Rodomirtoksin 16.28min:
427.175m/z
[M-H]- 339.0901 2532 [C19H18O6-2H]-H-
354.1101 1972 [C20H18O6]-
413.1620 759 [C23H25O7]-
Rubriflordilakton B 12.10min:
461.201m/z
[M-H]- 379.1575 1653 [C23H23O5]-
377.1404 1556 [C23H21O5]-
395.1495 1080 [C23H24O6]-H-
Metabolit yang dilaporkan ini menunjukkan fragmentasi ion yang sepadan dengan
pangkalan data yang dirujuk (pencarian menggunakan METLIN dan MetFrag terhadap
pangkalan data umum seperti KEGG, ChEBI, HMDB dan MetaCyc). Dua metabolit piawai
telah digunakan iaitu α-mangostin dan γ-mangostin dan dianalisis menggunakan parameter

89
dan sistem LC-MS/MS yang sama. Corak fragmentasi bagi α-mangostin di dalam sampel
manggis didapati sepadan dengan metabolit piawai yang telah digunakan manakala γ-
mangostin pula tidak dapat dikenalpasti di dalam mana-mana tisu manggis (Jadual 4.8).
Tapak fragmentasi yang dicadangkan pada struktur molekul α-mangostin yang
menghasilkan fragmen ion yang berlainan boleh dirujuk pada Lampiran O manakala tapak
fragmentasi pada struktur molekul lain-lain metabolit yang telah dikenalpasti daripada
analisis LC-MS telah dilampirkan di dalam cakera padat (compact disc) sebagai data
tambahan bagi kajian ini.

90
4.2.3 Analisis tapak jalan metabolomik
Berdasarkan analisis tapak jalan yang dijalankan melalui Analisis Tapak Jalan
Metabolomik (MetPA) terhadap model organisma Arabidopsis thaliana menggunakan
pelayan MetaboAnalyst, 10 tapak jalan yang mempunyai impak paling tinggi
disenaraikan seperti dalam Jadual 4.9 di bawah. Impak dikira dengan membahagikan
bilangan padanan dengan jumlah metabolit dalam tapak jalan yang sepadan. Impak
yang tinggi menggambarkan penglibatan metabolit yang lebih tinggi dalam sesuatu
tapak jalan (Chen et al. 2015), oleh itu ianya sangat berguna bagi membolehkan data
metabolit yang diperoleh daripada analisis GC-MS dan LC-MS dapat dikaji secara
global pada peringkat tapak jalan biosintesis metabolit.
Jadual 4.9 Hasil analisis tapak jalan metabolomik (MetPA). Analisis dijalankan
menggunakan data yang diperoleh daripada analisis pemprofilan metabolit
perikarpa, isi dan biji manggis pada peringkat pemasakan berbeza
menggunakan GC-MS dan LC-MS
Tapak jalan Jumlah
metabolit
Padanan Impak Metabolit
Metabolisme galaktosa 26 8 0.3077 D-fruktosa, D-galaktosa, D-glukosa,
D-manosa, gliserol, glukosa 6-fosfat,
myo-inositol, sukrosa
Metabolisme askorbat
dan aldarat
15 4 0.2667 asid D-glukuronik, D-galaktosa,
D-glukarat, myo-inositol
Metabolisme glioksilat
dan dikarboksilat
17 3 0.1765 asid isositrik, asid malik, asid sitrik
Penyalingtukaran pentosa
dan glukuronat
12 2 0.1667 D-xilosa, D-xilulosa
Kitaran asid
trikarboksilik (TCA)
20 3 0.1500 asid isositrik, asid malik, asid sitrik
Metabolisme kanji dan
sukrosa
30 4 0.1333 D-fruktosa, D-glukosa, glukosa 6-
fosfat, sukrosa
Metabolisme gula amino
dan gula nukleotida
41 5 0.1220 arabinosa, D-fruktosa, D-galaktosa,
D-manosa, glukosa 6-fosfat,
Tapak jalan pentosa
fosfat
18 2 0.1111 D-ribosa, glukosa 6-fosfat
Biosintesis Lisin 10 1 0.1000 asid L-aspartik
Metabolisme inositol
fosfat
24 2 0.0833 asid D-glukuronik, myo-inositol
Tapak jalan metabolisme galaktosa didapati menunjukkan padanan bilangan
metabolit yang paling tinggi dengan bilangan asal metabolit di dalam tapak jalan
tersebut iaitu sebanyak lapan metabolit. D-fruktosa, D-galaktosa, D-glukosa, D-
manosa, gliserol, glukosa 6-fosfat, myo-inositol dan sukrosa didapati terlibat dalam
tapak jalan ini. Kebanyakan metabolit yang terlibat dalam tapak jalan ini turut terlibat

91
di dalam beberapa tapak jalan yang lain. Contohnya, D-galaktosa turut terlibat dalam
metabolisme askorbat dan aldarat serta metabolisme gula amino dan gula nukleotida.
Glukosa 6-fosfat pula turut terlibat dalam metabolisme kanji dan sukrosa dan
metabolisme gula amino dan gula nukleotida serta tapak jalan pentosa fosfat.
Selain itu, tapak jalan metabolisme askorbat dan aldarat serta metabolisme
glioksilat dan dikarboksilat juga menunjukkan kadar impak yang tinggi dengan
penglibatan tiga ke empat metabolit. Tapak jalan pentosa fosfat, biosintesis lisin dan
metabolisme inositol fosfat menunjukkan penglibatan metabolit paling rendah iaitu
antara satu dan dua metabolit sahaja. Seterusnya, kajian tapak jalan juga dilakukan
dengan merujuk kepada kajian-kajian lepas (Aizat et al. 2014; Mazlan et al. 2019).
Ekoran daripada maklumat mengenai proses pemasakan buah manggis yang masih
terhad, masih tiada tapak jalan yang lengkap untuk proses pemasakan bagi buah ini.
Oleh itu, kajian kepustakaan difokuskan kepada tapak jalan metabolit dalam tumbuhan
dan buah berdasarkan pangkalan data KEGG.
Selain daripada terlibat dalam beberapa tapak jalan utama seperti metabolisme
galaktosa dan metabolisme askorbat dan aldarat, metabolit gula seperti sukrosa, D-
glukosa dan D-fruktosa juga berkait rapat dengan tapak jalan glikolisis melalui
penghasilan glukosa-6-fosfat dan fruktosa-6-fosfat. Tapak jalan glikolisis berperanan
membekalkan substrat bagi pelbagai tapak jalan lain termasuklah kitaran asid
trikarboksilik (TCA) melalui penghasilan asetil-CoA dan metabolisme sisteina dan
metionina serta biosintesis etilena melalui penghasilan asid aspartik. Selain itu,
glikolisis juga menghasilkan substrat yang terlibat dalam tapak jalan metabolit sekunder
melalui penghasilan fosfoenol piruvat yang menyumbang kepada penghasilan alkaloid,
terpenoid, flavonoid dan juga xanton (Rajah 4.16).
Berpandukan hasil analisis MetPA, metabolit yang dikenalpasti melalui analisis
GC-MS dan LC-MS telah dipetakan ke dalam gabungan tapak jalan yang telah
diringkaskan seperti yang dapat dilihat di dalam Rajah 4.16 di bawah. Setiap metabolit
yang telah dikenalpasti diwakili dengan peta haba yang menunjukkan nilai purata
kawasan puncak yang telah dinormalisasikan bagi analisis GC-MS dan purata

92
ketinggian puncak yang telah dinormalisasikan bagi analisis LC-MS. Tahap sesuatu
metabolit dapat ditentukan berdasarkan peta haba ini.
Sebagai contoh, α-mangostin didapati tinggi di dalam perikarpa bermula
peringkat 2 pemasakan dan paling tinggi di dalam biji manggis pada peringkat akhir
pemasakan. Metabolit xanton lain seperti gartanin pula didapati tinggi di dalam
perikarpa manggis pada peringkat 4 dan 6 manakala 1,3,5-trihidroksixanton pula adalah
paling tinggi di dalam isi manggis pada peringkat akhir pemasakan (peringkat 6).
Maklumat ini boleh digunakan untuk mendapatkan gambaran mengenai peringkat
pemasakan dan tisu manggis yang paling sesuai untuk digunakan bagi mendapatkan
sebatian bioaktif manggis yang paling tinggi.
Kebanyakan kajian lepas terhadap pengekstrakan xanton seperti mangostin
hanya memfokuskan kepada peringkat 6, iaitu peringkat akhir pemasakan (Aizat et al.
2019; Failla & Gutiérrez-Orozco 2017). Namun, kajian ini mendapati bahawa metabolit
ini telah hadir dalam kadar yang tinggi bermula peringkat 2 pemasakan. Justeru itu,
maklumat ini bukan sahaja akan menyumbang kepada pelbagai kajian mengenai
sebatian bioaktif bernilai tinggi yang terdapat di dalam manggis malah boleh
dimanfaatkan sebagai panduan bagi tujuan penuaian buah manggis. Tapak jalan
pemasakan manggis ini adalah yang pertama dilaporkan menggabungkan beberapa
tapak jalan yang berlainan seperti tapak jalan kanji dan sukrosa, glikolisis serta
metabolisme sisteina dan metionina yang merupakan hasil yang signifikan bagi kajian
ini.
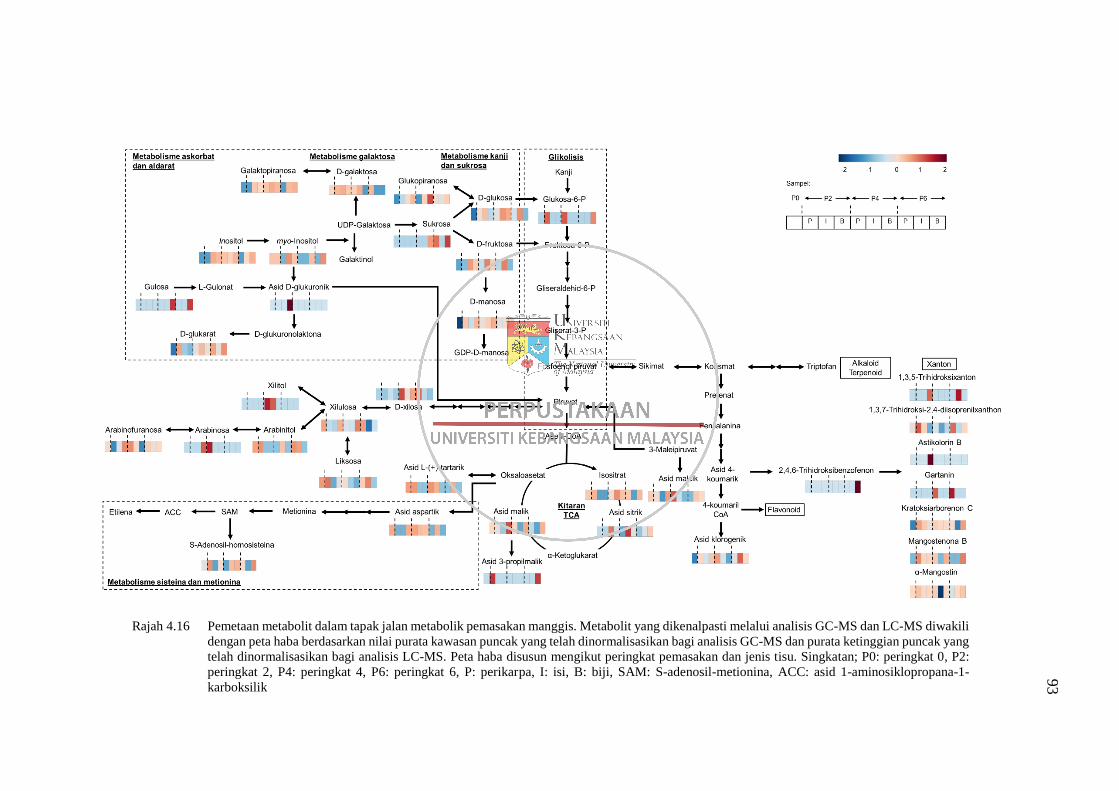
93
Rajah 4.16 Pemetaan metabolit dalam tapak jalan metabolik pemasakan manggis. Metabolit yang dikenalpasti melalui analisis GC-MS dan LC-MS diwakili
dengan peta haba berdasarkan nilai purata kawasan puncak yang telah dinormalisasikan bagi analisis GC-MS dan purata ketinggian puncak yang
telah dinormalisasikan bagi analisis LC-MS. Peta haba disusun mengikut peringkat pemasakan dan jenis tisu. Singkatan; P0: peringkat 0, P2:
peringkat 2, P4: peringkat 4, P6: peringkat 6, P: perikarpa, I: isi, B: biji, SAM: S-adenosil-metionina, ACC: asid 1-aminosiklopropana-1-
karboksilik

BAB V
PERBINCANGAN
5.1 PERBANDINGAN KAEDAH PENGEKSTRAKAN METABOLIT
5.1.1 Perbandingan keberkesanan derivatisasi menggunakan MSTFA dan BSTFA
terhadap pengesanan dan pengenalpastian metabolit
Dalam analisis pengoptimuman kaedah pengekstrakan ini, proses derivatisasi sampel telah
dijalankan melalui kaedah sililasi dengan menggunakan N-metil-N-(trimetilsilil)
trifluoroasetamina (MSTFA) atau N,O-bistrifluoroasetamina (BSTFA) untuk
menghasilkan metabolit dengan ciri-ciri yang sesuai bagi analisis GC-MS iaitu meruap dan
stabil haba. Keberkesanan tahap penderma sililasi bagi MSTFA telah dilaporkan setanding
dengan BSTFA dan N,O-bistrimetillsililasetamida (BSA), namun kadar kemeruapan
MSTFA didapati lebih tinggi, menyebabkannya lebih sesuai digunakan untuk analisis GC
(Halket et al. 2005). Penambahan 1% trimetilklorosilana (TMCS) ke dalam MSTFA pula
telah dilaporkan mampu menghasilkan fragmentasi ion yang stabil dan tidak menyebabkan
sebarang gangguan kepada sistem kromatografi semasa analisis dijalankan (Kushnir &
Komaromy-Hiller 2000). Hal ini kerana MSTFA kebiasaannya akan menghasilkan produk
sampingan atau artifak yang boleh mengganggu sistem kromatografi (Little 1999).
Manakala, BSTFA pula telah dilaporkan mampu bertindak balas dengan molekul
sebatian terutamanya asid organik dengan cepat, seterusnya memberikan hasil derivatisasi
yang tinggi (Orata 2012). Sifat BSTFA dan produk sampingannya yang mempunyai kadar
kemeruapan yang tinggi menyebabkan puncak terelusi awal dan pemisahan berlaku pada

95
puncak ini. Produk yang terhasil daripada derivatisasi BSTFA mempunyai tahap hingar
yang rendah dan tidak tercemar seterusnya menyebabkan penghasilan puncak yang tajam
dan jelas (Orata 2012). Campuran BSTFA dengan 1% TMCS pula boleh mengurangkan
gangguan (kesan matriks) semasa analisis GC-MS dijalankan (Azizan et al. 2012; Deng et
al. 2005). Proses derivatisasi yang berkesan akan menghasilkan puncak metabolit yang
tajam, bersimetri, jelas dan mempunyai resolusi yang tinggi pada kromatogram (Orata
2012), seterusnya memudahkan proses pengenalpastian bagi setiap individu metabolit.
Pada awalnya, metabolit daripada perikarpa manggis diekstrak menggunakan lima
kaedah pengekstrakan berbeza (kaedah 1, 2, 3, 4 dan 5) dan diderivatisasi menggunakan
MSTFA (Jadual 4.1). Berdasarkan kromatogram ion jumlah (TIC) yang diperoleh
(Lampiran A), campuran MSTFA dan 1% TMCS didapati sesuai digunakan bagi kelima-
lima kaedah pengekstrakan terutamanya kaedah 3 dan kaedah 5 yang menunjukkan
pemisahan puncak yang lebih jelas, tajam dan menghasilkan bilangan puncak yang lebih
banyak berbanding kaedah yang lain (kaedah 1, 2 dan 5). Selain itu, berdasarkan analisis
pengenalpastian metabolit, kaedah 3 (metanol/kloroform/air dengan nisbah 2:1:2) dan
kaedah 5 (metanol/kloroform/air dengan nisbah 3:1:1) juga menghasilkan jumlah
keseluruhan metabolit tertinggi berbanding kaedah 1, 2 dan 4 iaitu sebanyak 31 metabolit
bagi kaedah 3 dan 34 metabolit bagi kaedah 5 (Jadual 4.1).
Kaedah 3 dan 5 kemudiannya telah dipilih untuk menilai keupayaan kedua-dua
kaedah ini untuk mengekstrak metabolit daripada tisu yang berbeza iaitu perikarpa, isi dan
biji manggis. Bagi analisis pengoptimuman kaedah pengekstrakan terhadap tisu manggis
yang berbeza ini, sampel manggis telah diderivatisasi menggunakan BSTFA. Berdasarkan
TIC yang diperoleh daripada analisis GC-MS, kaedah 3 (Lampiran D) dan 5 (Lampiran E)
menunjukkan hasil pemisahan puncak yang jelas dan tajam bagi ketiga-tiga jenis tisu
manggis (perikarpa, isi dan biji), menandakan bahawa campuran BSTFA dengan 1%
TMCS sesuai digunakan untuk kedua-dua kaedah pengekstrakan bagi analisis GC-MS.
Berdasarkan analisis pengenalpastian metabolit pula, kaedah 3 dan 5 masing-masing
menunjukkan keberkesanan dalam pengekstrakan metabolit dengan penghasilan jumlah
metabolit keseluruhan yang sama iaitu sebanyak 36 metabolit bagi setiap kaedah (Jadual

96
4.3). Namun, kaedah 3 menghasilkan jumlah metabolit primer yang lebih rendah (31
metabolit) berbanding kaedah 5 (32 metabolit).
Sementara itu, apabila dibandingkan hasil pengekstrakan berdasarkan kaedah
pengekstrakan yang sama (kaedah 3 dan 5) dengan hasil analisis pengoptimuman
menggunakan MSTFA yang dijalankan sebelum ini, didapati bahawa bilangan metabolit
yang dikenalpasti menggunakan derivatisasi BSTFA adalah lebih banyak berbanding
MSTFA. Tisu manggis yang diekstrak menggunakan kaedah 3 dan 5 dan diderivatisasi
menggunakan BSTFA menghasilkan 1.41%-7.46% lebih tinggi metabolit berbanding tisu
yang diekstrak menggunakan kaedah yang sama tetapi diderivatisasi menggunakan
MSTFA. Oleh itu, kaedah derivatisasi menggunakan BSTFA telah dipilih untuk
menjalankan analisis pemprofilan metabolit daripada perikarpa, isi dan biji manggis pada
peringkat pemasakan yang berbeza (peringkat 0, 2, 4 dan 6). Tambahan pula, proses
derivatisasi sampel menggunakan BSTFA memerlukan masa penyediaan yang lebih
pendek iaitu selama satu jam berbanding MSTFA yang mengambil masa penyediaan
selama dua jam (Bab 3.2.3). Ini membolehkan proses penyediaan sampel dijalankan dengan
lebih cepat dan berkesan.
5.1.2 Perbandingan jenis dan nisbah pelarut serta teknik lisis terhadap pengekstrakan
metabolit menggunakan lima kaedah pengekstrakan berbeza dan derivatisasi
MSTFA
Pengoptimuman kaedah pengekstrakan metabolit adalah satu proses yang sangat penting
dalam kajian metabolomik kerana kaedah pengekstrakan akan mempengaruhi jenis,
bilangan serta kadar kepekatan metabolit yang boleh diekstrak daripada mana-mana sampel
biologi (Cevallos-Cevallos et al. 2009; Kim & Verpoorte 2010; Mushtaq et al. 2014).
Kaedah pengekstrakan yang berkesan akan memberikan hasil yang maksimum kepada
kajian yang dijalankan. Bagi kajian metabolomik bukan sasaran, identiti serta sifat
kebanyakan metabolit yang terkandung di dalam sampel adalah tidak diketahui (Cevallos-
Cevallos et al. 2009). Oleh itu, beberapa faktor yang mempengaruhi keberkesanan kaedah
pengekstrakan haruslah diuji, dibandingkan serta dinilai dengan teliti.

97
Dalam kajian ini, terdapat tiga faktor yang merangkumi faktor kimia serta faktor
fizikal iaitu, 1) jenis larutan, 2) nisbah larutan serta 3) kesan teknik sonikasi terhadap hasil
pengekstrakan metabolit telah dibandingkan (Jadual 4.1). Terdapat dua jenis campuran
larutan telah digunakan iaitu metanol: kloroform: air dengan nisbah berbeza (2:1:2 dan
3:1:3), disertai teknik sonikasi atau tanpa teknik sonikasi serta campuran larutan metanol:
asid formik (599:1) disertai teknik sonikasi telah digunakan. Campuran larutan metanol,
kloroform dengan air telah dipilih kerana campuran larutan ini dipercayai berkesan untuk
memberikan hasil analisis yang memberangsangkan dan campuran ini seringkali digunakan
dalam kajian metabolomik untuk mengekstrak pelbagai jenis sampel biologi tumbuhan
seperti Solanum tuberosum (kentang) (Dobson et al. 2008), Camellia sinensis (teh hijau
Jepun), Fagus sylvatica L. (bic) (Cadahía et al. 2015) dan Cucurbita maxima (labu)
(Okazaki et al. 2016). Nisbah 2:1:2 dan 3:1:3 juga telah terbukti berkesan dalam
mengekstrak pelbagai jenis metabolit berkutub, tidak berkutub, hidrofobik dan hidrofilik
(Cadahía et al. 2015; Lisec et al. 2006; Okazaki et al. 2016).
Hasil daripada analisis perbandingan yang telah dijalankan, gabungan beberapa
pelarut seperti metanol dengan asid formik serta metanol dengan kloroform dan air dapat
dilihat menunjukkan perbezaan dari segi jenis metabolit yang berjaya diekstrak (Rajah 4.1).
Gabungan pelarut metanol, kloroform dan air menunjukkan hasil pengekstrakan yang lebih
baik kerana kaedah ini berjaya mengekstrak pelbagai jenis metabolit yang merangkumi
kumpulan gula, alkohol dan gula alkohol, asid organik dan juga metabolit sekunder. Hal
ini kerana campuran ketiga-tiga pelarut ini sangat sesuai digunakan untuk mengekstrak
metabolit daripada kumpulan berkutub sehingga ke metabolit kumpulan tidak berkutub
daripada pelbagai sampel biologi (Kim & Verpoorte 2010; Mushtaq et al. 2014)
termasuklah sampel tumbuhan seperti Arabidopsis thaliana (Gullberg et al. 2004) dan
Cucurbita maxima (labu) (Okazaki et al. 2016).
Kaedah pengekstrakan yang diiringi dengan teknik sonikasi iaitu kaedah 1, 3 dan 5
juga berjaya mengenalpasti bilangan metabolit yang lebih tinggi iaitu masing-masing
sebanyak 28, 31 dan 34 metabolit berbanding kaedah pengekstrakan yang tidak diiringi
dengan teknik ini iaitu kaedah 2 dan 4 yang masing-masing menghasilkan sebanyak 22 dan

98
14 metabolit sahaja (Jadual 4.1). Hal ini membuktikan bahawa teknik sonikasi adalah
berkesan untuk menggalakkan pengeluaran metabolit daripada sel perikarpa manggis tanpa
dipengaruhi oleh jenis pelarut yang digunakan. Teknik sonikasi dilakukan dengan cara
mendedahkan sampel kepada gelombang bunyi berfrekuensi tinggi di dalam air rendaman
dengan suhu yang sesuai. Gelombang bunyi ini akan menyebabkan sel pecah seterusnya
menggalakkan pembebasan metabolit daripada dalam sel tersebut. Teknik sonikasi telah
terbukti berjaya meningkatkan kadar pengekstrakan metabolit daripada pelbagai tisu
sampel biologi seperti Glycine max (kacang soya) (Rostagno et al. 2003) dan turunan sel
karsinoma endometrium manusia (ECCI) (Matheus et al. 2014). Oleh itu, teknik ini sesuai
untuk diaplikasikan dalam setiap kaedah pengekstrakan metabolit daripada mana-mana
sampel biologi.
Kaedah 1 yang menggunakan campuran pelarut metanol dengan asid formik disertai
teknik sonikasi turut berjaya mengekstrak jumlah keseluruhan metabolit yang ketiga
tertinggi selepas kaedah 3 dan kaedah 5 yang menggunakan campuran metanol, kloroform
dan air disertai dengan teknik sonikasi. De Vos et al. (2007) melaporkan bahawa campuran
pelarut metanol dengan asid formik adalah sesuai digunakan untuk mengekstrak julat
metabolit yang besar merangkumi pelbagai jenis metabolit sekunder daripada tisu
tumbuhan. Kaedah ini telah berjaya mengekstrak jumlah metabolit sekunder yang lebih
tinggi iaitu sebanyak lima metabolit berbanding kaedah-kaedah pengekstrakan yang lain
(Rajah 4.1). Kaedah 3 dan 5 masing-masing berjaya mengekstrak satu metabolit sekunder
manakala tiada metabolit sekunder dapat diekstrak menggunakan kaedah 2 dan 4. Namun
begitu, jumlah keseluruhan metabolit sekunder yang dapat diekstrak menggunakan kelima-
lima kaedah pengekstrakan adalah terlalu rendah. Hal ini mungkin disebabkan oleh limitasi
yang terdapat pada sistem GC-MS yang lebih sensitif terhadap metabolit primer seperti
gula, alkohol, asid organik dan asid amino (Gullberg et al. 2004; Lee et al. 2013; Okazaki
et al. 2016; t’Kindt et al. 2009). Pengenalpastian metabolit sekunder pula dilaporkan lebih
sesuai dijalankan dengan menggunakan analisis LC-MS yang lebih sensitif terhadap
metabolit sekunder seperti flavonoid, alkaloid, saponin, fenilpropanoid dan xanton (De Vos
et al. 2007; Lee et al. 2013; t’Kindt et al. 2009).

99
5.1.3 Kesan nisbah pelarut terhadap pengekstrakan metabolit menggunakan dua kaedah
pengekstrakan berbeza dan derivatisasi BSTFA
Daripada analisis GC-MS yang telah dijalankan dengan derivatisasi BSTFA, kedua-dua
kaedah pengekstrakan (kaedah 3 dan kaedah 5) berjaya mengekstrak jumlah metabolit yang
sama iaitu sebanyak 36 metabolit bagi setiap kaedah (Jadual 4.3). Meskipun kaedah 3 dan
kaedah 5 menghasilkan jumlah metabolit yang sama, terdapat sebilangan metabolit yang
diekstrak adalah berbeza antara dua kaedah pengekstrakan ini yang seterusnya membawa
kepada jumlah keseluruhan metabolit yang dapat dikesan daripada kedua-dua kaedah
adalah sebanyak 43 metabolit (Lampiran B). Perbezaan dapat dilihat dari segi jenis
metabolit yang diekstrak dan bilangan metabolit bagi jenis metabolit tertentu antara kaedah
3 dan 5 (Rajah 4.5).
Berdasarkan analisis multivariat yang telah dijalankan pula, plot skor PCA dalam
Rajah 4.6 menunjukkan pemisahan antara kelompok metabolit lebih cenderung bergantung
kepada jenis tisu berbanding kaedah pengekstrakan. Pertindihan antara kelompok metabolit
hanya berlaku antara kelompok metabolit daripada sampel isi bagi kedua-dua kaedah
pengekstrakan (kaedah 3: metanol/kloroform/air, 2:1:2 dan kaedah 5:
metanol/kloroform/air, 3:1:1). Kelompok metabolit daripada sampel perikarpa dan biji
daripada kaedah 3 dan 5 masing-masing menunjukkan pemisahan yang jelas. Pemisahan
ini mungkin berlaku disebabkan sifat biologi setiap jenis tisu itu sendiri di mana setiap tisu
iaitu perikarpa, isi dan biji memainkan fungsi metabolisme yang berbeza dalam tumbuhan
dan buah.
Seterusnya, analisis perbandingan melalui analisis gambar rajah Venn telah
dijalankan untuk mengkaji dengan lebih teliti persamaan dan perbezaan hasil
pengekstrakan menggunakan dua kaedah pengekstrakan berbeza daripada tiga jenis tisu
manggis (Rajah 4.8 dan Jadual 4.4). Hasil analisis mendapati bahawa darjah kekutuban
pelarut mempengaruhi jenis dan bilangan metabolit yang diekstrak seperti yang telah
dilaporkan dalam kajian lepas (Azizan et al. 2015; Mushtaq et al. 2014). Seperti yang
dibincangkan di dalam bahagian pengoptimuman lima kaedah pengekstrakan sebelum ini

100
(Bab 5.1.2), gabungan pelarut metanol, kloroform dan air digunakan kerana keberkesanan
gabungan pelarut ini untuk mengekstrak pelbagai jenis metabolit berkutub, tidak berkutub,
hidrofilik dan hidrofobik serta tahap kebolehulangan yang tinggi dalam kajian berkaitan
metabolomik tumbuhan (De Vos et al. 2007; Lisec et al. 2006; Mushtaq et al. 2014; Okazaki
et al. 2016).
Berdasarkan Jadual 4.4, kaedah 3 didapati berjaya mengekstrak lebih banyak
metabolit unik yang berkutub berbanding kaedah 5. Contohnya, metabolit berkutub seperti
arabinofuranosa, galakto-heksodialdosa, D-xilofuranosa dan β-D-glukopiranosida hanya
dapat dikesan menggunakan kaedah 3. Hal ini mungkin disebabkan nisbah pelarut berkutub
yang digunakan dalam kaedah 3 (dua nisbah air berbanding satu nisbah air dalam kaedah
5). Tambahan lagi, dua alkohol iaitu 2,3-butanediol dan treitol merupakan metabolit unik
bagi kaedah 5 sama ada daripada sampel perikarpa atau biji. Ini membuktikan bahawa
nisbah metanol yang digunakan dalam kaedah 5 (nisbah 3 berbanding nisbah 2 bagi kaedah
3) menyumbang kepada pengenalpastian metabolit alkohol dalam kajian ini. Hasil kajian
menjelaskan bahawa gabungan pelarut yang berbeza nisbah boleh mempengaruhi
pengekstrakan metabolit, seterusnya menekankan kepentingan kaedah pengekstrakan yang
optimum dalam mana-mana kajian berasakan metabolomik.
Secara kesimpulannya, kedua-dua kaedah pengekstrakan telah menunjukkan hasil
yang memberangsangkan. Kaedah 3 dan 5 sesuai digunakan untuk mengekstrak metabolit
dari perikarpa, isi dan manggis serta melalui proses derivatisasi sampel menggunakan
BSTFA. Namun, kaedah 5 telah dipilih untuk analisis seterusnya iaitu analisis pemprofilan
metabolit daripada perikarpa, isi dan biji manggis pada peringkat pemasakan yang berbeza
(peringkat 0, 2, 4 dan 6) kerana kaedah ini mampu mengekstrak metabolit primer dengan
lebih berkesan berbanding kaedah 3 meskipun perbezaan antara kedua-dua kaedah ini
adalah tidak begitu ketara.

101
5.1.4 Perbandingan kaedah pengekstrakan dan mod pengionan LC-MS terhadap
pengenalpastian metabolit
Keberkesanan kaedah 1 (metanol/asid formik dengan nisbah 599:1) dan kaedah 5
(metanol/kloroform/air dengan nisbah 3:1:1) terhadap pengekstrakan metabolit turut dikaji
menggunakan analisis LC-MS. Kaedah 1 dipilih kerana kaedah ini memberikan hasil
pengekstrakan metabolit sekunder yang paling tinggi berdasarkan analisis pengoptimuman
menggunakan GC-MS yang dijalankan sebelum ini iaitu sebanyak lima metabolit sekunder
berbanding kaedah 2 dan 4 yang tidak dapat mengekstrak metabolit sekunder serta kaedah
3 dan 5 yang masing-masing hanya menghasilkan satu metabolit sekunder sahaja (Rajah
4.1). Sementara itu, kaedah 5 pula dipilih kerana berjaya mengekstrak jumlah keseluruhan
metabolit yang paling tinggi berbanding kaedah lain (Jadual 4.1 dan Jadual 4.3).
Selain daripada kaedah pengekstrakan, mod pengionan (positif atau negatif) juga
mempengaruhi hasil analisis LC-MS dari segi jenis dan bilangan metabolit yang akan
dikenalpasti (Theodoridis et al. 2012b). Satu mod (positif atau negatif) tidak dapat meliputi
semua jenis metabolit (berkutub, tidak berkutub, ionik, neutral dan sebagainya) kerana
sesetengah jenis metabolit mempunyai kadar pengionan yang tinggi dalam salah satu mod
analisis, manakala sesetengah jenis metabolit yang lain pula mempunyai kadar pengionan
tinggi di dalam mod pengionan yang lain. Mod positif seringkali digunakan dalam analisis
LC-MS kerana keberkesanannya untuk mengionkan pelbagai metabolit berkutub dan tidak
berkutub manakala mod negatif dilaporkan berprestasi tinggi mengionkan kelas metabolit
yang tertentu seperti metabolit gula dan asid organik (Theodoridis et al. 2012b).
Bagi analisis perbandingan ini, kaedah 1 dan kaedah 5 dinilai berdasarkan
penghasilan bilangan puncak pada kromatogram yang diwakili dengan pasangan RT: m/z
pada kedua-dua mod positif dan negatif tanpa analisis pengenalpastian metabolit yang
kompleks dan memerlukan masa kajian yang lebih lama. Berdasarkan hasil yang diperolehi
(Jadual 4.5), kaedah 1 menghasilkan sebanyak 46 pasangan RT: m/z pada mod positif dan
58 pasangan RT: m/z pada mod negatif manakala kaedah 5 pula menghasilkan bilangan
pasangan RT: m/z yang lebih tinggi pada mod positif iaitu 55 pasangan RT: m/z dan

102
bilangan puncak yang lebih rendah pada mod negatif iaitu sebanyak 41 pasangan RT: m/z.
Secara kesimpulannya, kaedah 1 menunjukkan hasil yang lebih memberangsangkan
dengan jumlah pasangan RT: m/z yang lebih tinggi berbanding kaedah 5 terutamanya bagi
mod pengionan negatif.
Kaedah 1 seterusnya telah dipilih untuk digunakan bagi analisis pemprofilan
metabolit daripada pelbagai tisu (perikarpa, isi dan biji) manggis pada peringkat pemasakan
yang berbeza (peringkat 0, 2, 4 dan 6). Manakala, mod pengionan pula ditetapkan kepada
mod negatif. Tambahan pula, mod pengionan negatif ESI-MS juga dikatakan sesuai untuk
mengkaji metabolit xanton yang merupakan kumpulan bioaktif utama dalam manggis. Hal
ini kerana mod negatif mampu meningkatkan fragmentasi metabolit xanton, sekaligus
memudahkan proses pengenalpastian dan penentuan struktur xanton (Wittenauer et al.
2012).
5.2 PENGAWALATURAN METABOLISME METABOLIT PRIMER DAN
SEKUNDER SEMASA PEMASAKAN MANGGIS
5.2.1 Analisis multivariat mendedahkan pemisahan kelompok metabolit mengikut
peringkat pemasakan berdasarkan jenis tisu yang berbeza
Analisis multivariat yang merangkumi analisis PCA dan PLS-DA telah dijalankan secara
berasingan bagi data yang diperoleh daripada analisis GC-MS (Rajah 4.9) dan LC-MS
(Rajah 4.12). Terlebih dahulu, analisis PCA telah dijalankan mengikut jenis tisu (perikarpa,
isi dan biji) untuk melihat pemisahan antara peringkat pemasakan yang berbeza iaitu
peringkat 0, 2, 4 dan 6. Kemudian, analisis PLS-DA turut dijalankan untuk mengenalpasti
pembolehubah yang berkepentingan dalam unjuran (VIP) atau metabolit berpengaruh
tinggi yang berkemungkinan memainkan peranan penting dalam pemasakan manggis.
Bagi analisis GC-MS, hasil kajian menunjukkan bahawa taburan metabolit dalam
tisu manggis yang berbeza iaitu perikarpa (Rajah 4.9a dan d), isi (Rajah 4.9b dan e) dan
biji (Rajah 4.9c dan f) pada empat peringkat pemasakan berjaya didiskriminasikan
menggunakan kedua-dua analisis PCA dan PLS-DA ini. Pemisahan yang jelas antara

103
kelompok sampel yang dibandingkan menerangkan bahawa taburan metabolit adalah
dipengaruhi oleh jenis tisu serta peringkat pemasakan dalam manggis. Namun, terdapat
sedikit pertindihan antara kelompok metabolit daripada peringkat 0 dengan perikarpa
(peringkat 2) (Rajah 4.9a dan d), menunjukkan terdapat persamaan antara dua kelompok
metabolit ini. Hal ini mungkin terjadi disebabkan metabolit primer daripada peringkat 0
dan 2 tidak mempunyai perbezaan yang ketara kerana kedua-dua peringkat ini dianggap
sebagai peringkat awal pemasakan atau belum-masak (Osman & Milan 2006; Parijadi et al.
2017).
Manakala bagi data yang diperoleh daripada analisis LC-MS, pemisahan yang jelas
berjaya diperoleh bagi semua plot skor PCA dan PLS-DA, menunjukkan bahawa variasi
antara kelompok metabolit daripada peringkat pemasakan yang berbeza adalah tinggi
mengikut jenis tisu. Sementara itu, ketepatan yang tinggi antara replikat biologi bagi setiap
kelompok metabolit yang dapat dilihat di dalam semua plot skor PCA dan PLS-DA bagi
kedua-dua analisis GC-MS dan LC-MS menandakan tahap kebolehulangan yang tinggi
antara sampel mengikut peringkat dan jenis tisu. Ini sekaligus memberikan gambaran
bahawa pensampelan dan pemprosesan sampel telah dilakukan dengan teliti dan tepat.
5.2.2 Pengawalaturan metabolik gula, asid amino, asid organik serta fitohormon
menyumbang kepada ciri-ciri pemasakan manggis
Pemasakan buah berkait rapat dengan beberapa fenomena utama seperti degradasi dinding
sel yang membawa kepada pelembutan buah serta diikuti dengan perkembangan rasa,
aroma dan bau. Fenomena ini melibatkan biosintesis dan modifikasi pada tahap sesetengah
metabolit termasuklah gula, asid gula, asid organik dan sebatian meruap (Osorio & Fernie
2014; Osorio et al. 2013; Sharon‐Asa et al. 2003). Galaktosa dan manosa adalah antara
metabolit yang telah dikenalpasti sebagai komponen dinding sel yang terlibat dengan
pelembutan buah semasa pemasakan berlaku seperti yang dilaporkan bagi Solanum
lycopersicum (tomato) (Oms-Oliu et al. 2011). Dalam kajian ini, metabolit terbitan kepada
manosa iaitu L-manopiranosa dikenalpasti sebagai metabolit berpengaruh tinggi di dalam
perikarpa manggis dengan nilai VIP ≥1.00 dan p ≤0.05 (Rajah 4.11a) . Pengumpulan L-

104
manopiranosa di dalam perikarpa menunjukkan bahawa degradasi dinding sel berlaku
secara aktif di dalam perikarpa manggis semasa proses pemasakan. Tambahan lagi, analisis
tapak jalan metabolomik (MetPA) menyatakan bahawa metabolisme galaktosa merupakan
tapak jalan paling dominan dalam proses pemasakan manggis di mana manosa dan
galaktosa juga terlibat dalam tapak jalan ini (Jadual 4.9).
Selain itu, myo-inositol juga didapati turut terlibat dalam metabolisme galaktosa
serta metabolisme askorbat dan aldarat yang juga dikenalpasti sebagai tapak jalan utama di
dalam pemasakan manggis (Jadual 4.9). Kehadiran myo-inositol daripada perikarpa
manggis serta kadar metabolit ini yang tinggi di dalam biji manggis pada peringkat 2 dan
6 (Rajah 4.11d) mungkin berkait dengan fungsinya sebagai pelopor yang penting kepada
biosintesis pelbagai polisakarida dinding sel termasuklah pektin dan hemiselulosa (Oms-
Oliu et al. 2011). Oleh itu, apabila ketegaran dinding sel buah menurun apabila
menghampiri peringkat akhir pemasakan, myo-inositol berkumpul di dalam kedua-dua tisu
perikarpa dan biji manggis. myo-Inositol juga berperanan dalam pengisyaratan (Shi et al.
2005; Valluru & Van den Ende 2011) dan kawalan tindak balas tekanan di dalam tumbuhan
dan buah (Nkang 2002; Valluru & Van den Ende 2011; Yildizli et al. 2018). Proses
pemasakan diketahui turut melibatkan proses pengisyaratan dan kawalan tindak balas
tekanan yang kompleks, yang mana kehadiran myo-inositol mungkin terlibat dalam proses
ini.
Gula lain yang turut dikenalpasti dalam kajian ini adalah arabinofuranosa (Rajah
4.11a). Arabinofuranosa sering hadir sebagai arabinogalaktan, komponen utama rantaian
sampingan bagi pektin di dalam dinding sel (Denman & Morris 2015; Robinson & Davies
2000). Penurunan yang signifikan bagi arabinofuranosa di dalam tisu perikarpa dan biji
manggis daripada peringkat pertengahan pemasakan (peringkat 2 atau 4) kepada peringkat
akhir pemasakan (peringkat 6) menunjukkan bahawa berlakunya modifikasi terhadap
struktur pektin di dalam buah manggis. Pektin adalah polisakarida dinding sel yang kaya
dengan asid galakturonik dan merupakan polisakarida yang mempunyai struktur dan fungsi
yang paling kompleks dalam dinding sel tumbuhan (Caffall & Mohnen 2009; Mohnen
2008). Kehadiran asid galakturonik pada kadar yang tinggi semasa peringkat 0 dan

105
peringkat 2 (perikarpa) pemasakan tetapi tidak pada peringkat yang seterusnya (Rajah
4.11c) membuktikan bahawa berlakunya degradasi pektin di dalam struktur dinding sel
manggis yang seterusnya membawa kepada pelembutan buah semasa proses pemasakan
(Osorio & Fernie 2014). Degradasi polisakarida pektin semasa buah masak dipercayai
membawa kepada proses pemasakan buah yang sebenar (Prasanna et al. 2007).
Modifikasi pada kandungan metabolit gula adalah salah satu perubahan utama yang
berlaku semasa pemasakan bagi kebanyakan buah tidak kira sama ada buah klimakterik
atau bukan-klimakterik (Aizat et al. 2014; Fait et al. 2008; Osorio et al. 2012). Kadar yang
tinggi bagi D-glukosa 6-fosfat di dalam perikarpa manggis pada peringkat 2 dan 4 (Rajah
4.14c) didapati selaras dengan aras D-glukosa sewaktu proses pemasakan (Lampiran K).
D-glukosa 6-fosfat merupakan salah satu metabolit yang terlibat dalam bahagian awal tapak
jalan glikolisis (Rajah 4.16) yang merupakan tapak jalan penjanaan tenaga daripada proses
pembubaran glukosa (Aizat et al. 2014; Paul & Pellny 2003). Kadar D-glukosa 6-fosfat
yang tinggi mencadangkan bahawa aktiviti glikolitik di dalam perikarpa manggis adalah
aktif semasa peringkat pertengahan pemasakan (peringkat 2 dan 4). Ini mungkin
disebabkan oleh peranan tapak jalan glikolisis dalam membekalkan pelbagai pelopor
penting kepada tapak jalan yang lain seperti kitaran asid trikarboksilik (TCA), biosintesis
etilena dan biosintesis metabolit sekunder (Rajah 4.16) sewaktu proses pemasakan.
Metabolit berpengaruh tinggi (VIP ≥1.00 dan p ≤0.05) yang lain merangkumi
metabolit gula dan asid organik seperti L(-)-fukosa (Rajah 4.11a), asid ribonik (Rajah 4.11c)
dan asid L-(+)-tartarik (Rajah 4.11f). Kajian oleh Mahmood et al. (2012) melaporkan
kehadiran asid askorbik (vitamin C) yang menyumbang kepada rasa masam berasid dalam
buah strawberi dan ceri manis yang masak. Asid askorbik tidak dapat dikesan di dalam
kajian terhadap manggis ini, namun metabolit terbitan bagi asid askorbik iaitu asid L-
askorbik-2-glukosida dikenalpasti sebagai metabolit berpengaruh tinggi semasa proses
pemasakan (Rajah 4.14d). Metabolit ini dikesan pada kadar yang tinggi pada peringkat
pertengahan dan akhir pemasakan, memberikan gambaran bahawa ianya mungkin
menyumbang kepada rasa masam manggis selepas dituai. Selain itu, asid tartarik turut
dilaporkan berkumpul pada peringkat akhir pemasakan bagi buah mulberi panjang, mulberi

106
kecil dan strawberi, seperti juga di dalam perikarpa manggis di dalam kajian ini (Rajah
4.11f) (Mahmood et al. 2012).
Sebatian meruap yang merangkumi alkohol, ester dan aldehid pula merupakan
kumpulan metabolit utama yang menyumbang kepada ciri-ciri aroma bagi buah masak
(Defilippi et al. 2009; Dixon & Hewett 2000; Osorio et al. 2013; Yang et al. 2016). Buah
manggis yang telah masak mempunyai aroma yang tersendiri. Aroma yang menyenangkan
dalam buah ini dipercayai berkait rapat dengan kadar arabinitol dalam perikarpa dan isi
manggis (pada peringkat 6) (Rajah 4.11) dan 2,3-butanadiol dalam isi manggis (pada
peringkat 4 dan 6) (Rajah 4.11e). Selain itu, peningkatan pada kadar penghasilan butanal
dilihat meningkat di dalam sampel isi dan biji manggis daripada peringkat 2 kepada
peringkat 4 pemasakan dan seterusnya menurun di dalam tisu yang sama pada peringkat 6
(Rajah 4.11). Perubahan ini mungkin berlaku disebabkan penukaran metabolit aldehid ini
kepada alkohol aromatik atau ester yang sepadan (Dixon & Hewett 2000; Yamashita et al.
1976) di dalam manggis sewaktu proses pemasakan.
Asid aspartik dan S-adenosilhomosisteina pula dipercayai terlibat dalam proses
pemasakan manggis melalui metabolisme sisteina dan metionina serta biosintesis etilena
(Rajah 4.16). Metionina adalah molekul utama kepada biosintesis fitohormon etilena yang
bertindak merangsang proses pemasakan buah klimakterik termasuklah manggis (Hesse et
al. 2004; Ravanel et al. 1998). Metionina boleh disintesis daripada aspartat dan
kemudiannya ditukarkan kepada S-adenosil-metionina (SAM) dan seterusnya kepada asid
1-aminosiklopropana-1-karboksilik (ACC) sebelum terhasilnya hormon etilena. Selain itu,
SAM juga boleh membentuk S-adenosilhomosisteina dengan pertambahan molekul tenaga
ATP (Bürstenbinder et al. 2007; Gellekink et al. 2005; Hesse et al. 2004; Ravanel et al.
1998). Dalam kajian ini, metabolit asid aspatik dan S-adenosilhomosisteina yang berkait
rapat dengan metabolisme metionina menunjukkan perubahan semasa pemasakan manggis.
Kadar asid aspartik yang tinggi di dalam ketiga-tiga tisu manggis sama ada pada peringkat
pertengahan atau akhir pemasakan (Rajah 4.14g) menandakan bahawa metabolisme
sisteina dan metionina serta biosintesis hormon etilena adalah aktif pada peringkat ini.
Kadar S-adenosilhomosisteina pula didapati menurun di dalam perikarpa sepanjang proses

107
pemasakan manggis (Rajah 4.14g). Hal ini mungkin berlaku disebabkan pelopornya iaitu
SAM mungkin digunakan dalam penghasilan hormon etilena (Rajah 4.16) yang diketahui
tinggi sewaktu pemasakan manggis (Paul et al. 2012).
Asid absisik (ABA) turut dilaporkan bertindak secara sinergi dengan hormon
etilena untuk menggalakkan pelembutan tisu buah dengan cara merangsang aktiviti
hidrolase dinding sel terutamanya pektin metil esterase (PME) dan pektat liase (PL) di
dalam buah pisang (Lohani et al. 2004). ABA juga berperanan dalam pengawalaturan
aktiviti proses pemasakan yang lain seperti pengumpulan gula dan perkembangan warna
(Nicolas et al. 2014). Dalam kajian ini, peningkatan ABA dalam biji manggis pada
peringkat akhir pemasakan (Rajah 4.14j) didapati selari dengan peningkatan sukrosa di
dalam tisu yang sama (Rajah 4.14a). Penemuan ini adalah sepadan dengan kajian oleh Jia
et al. (2011) yang melaporkan bahawa pengumpulan ABA boleh dirangsang oleh gula
eksogenus terutamanya sukrosa dan glukosa. Selain itu, Kondo et al. (2002) dalam
kajiannya pula turut mencadangkan bahawa ABA mungkin mencetuskan atau memulakan
proses pematangan manggis. Hal ini mungkin menjelaskan kehadiran ABA pada peringkat
0 pemasakan dan meningkat di dalam perikarpa dan biji apabila memasuki peringkat 2
(Rajah 4.14j).
5.2.3 Pengawalaturan dinamik metabolisme sekunder manggis melibatkan metabolit
fenolik, terpenoid, alkaloid dan xanton
Fenolik adalah kumpulan metabolit sekunder yang mempunyai taburan paling meluas di
dalam tumbuhan dan penghasilan kumpulan metabolit ini adalah bergantung kepada
spesies dan varieti tumbuhan (Lattanzio et al. 2008). Dalam kajian ini, metabolit fenolik
yang berpengaruh tinggi (VIP ≥1.00 dan p ≤0.05) dikenalpasti dengan jumlah yang paling
banyak iaitu 19 metabolit (Rajah 4.15a) berbanding kumpulan metabolit sekunder yang lain.
Salah satu fungsi utama kumpulan fenolik adalah untuk melindungi tumbuhan daripada
sinaran ultraungu (UV) merbahaya yang boleh merosakkan tumbuhan melalui penghasilan
kumpulan spesies oksigen aktif (ROS) (Lattanzio et al. 2008). Metabolit fenolik yang
merangkumi flavonoid, koumarin, fenolik glikosida dan kuersetin dikesan dalam jumlah

108
yang berbeza-beza daripada sampel perikarpa, isi dan biji manggis. Sebagai contoh,
peningkatan kadar beberapa fenolik yang dikenalpasti sebagai flavonoid seperti
metilofiogoponona B (perikarpa), leufolin A, prunin 6''-O-galat (isi) dan soiuflavon A (biji)
serta lignan seperti neoisostegana (biji) antara peringkat pemasakan yang tertentu (Rajah
4.15a) menunjukkan bahawa tapak jalan fenilpropanoid berlaku pada tahap yang berbeza
dalam setiap tisu manggis semasa proses pemasakan berlaku. Pengenalpastian leufolin A
turut disokong oleh data daripada analisis MS/MS dengan penghasilan fragmentasi puncak
[M-H]- pada m/z 579, sepadan dengan hasil kajian yang dilaporkan oleh Noor et al. (2007).
Perubahan kadar beberapa fenolik di dalam manggis boleh dikaitkan dengan
pelbagai fungsi kumpulan metabolit ini terhadap tumbuhan. Contohnya, lignan yang
dikategorikan bawah kumpulan fenolik mempunyai bioaktiviti anti-oksida dan
sitopelindung (cytoprotective) seperti yang telah dilaporkan di dalam Euterpe oleracea
(buah acai) (Chin et al. 2008). Peningkatan kadar neoisostegana iaitu metabolit lignan di
dalam biji manggis daripada peringkat 2 kepada peringkat 4 dan 6 (Rajah 4.15a)
menunjukkan bahawa metabolit ini mungkin dihasilkan secara aktif untuk perlindungan
manggis terhadap jangkitan patogen atau serangan daripada serangga (Harmatha & Dinan
2003). Sesetengah lignan turut dilaporkan berupaya untuk mempengaruhi perkembangan
serangga dengan mengubah pelbagai fungsi fisiologi serangga tersebut melalui aktiviti
antagonistik ekdisteroid (Harmatha & Dinan 2003). Hal ini juga mungkin menjelaskan
kadar neoisostegana yang tinggi di dalam perikarpa manggis pada peringkat 2 (Rajah 4.15a)
yang mungkin menunjukkan bahawa aktiviti pertahanan telah bermula dari sejak awal
proses pemasakan lagi bagi menjamin pemasakan buah yang sempurna sehingga peringkat
akhir pemasakan.
Sesetengah fenolik memainkan peranan dalam perkembangan warna, rasa dan nilai
nutrisi bagi buah segar (Zarena & Sankar 2012). Perkembangan warna manggis berkait
rapat dengan proses pemasakan di mana kulit manggis akan berubah daripada warna hijau
pada peringkat awal pemasakan kepada warna ungu gelap apabila menghampiri peringkat
akhir pemasakan (Osman & Milan 2006; Palapol et al. 2009b). Perubahan warna kulit buah
bertindak sebagai penunjuk utama untuk menilai tahap kematangan manggis terutamanya

109
bagi tujuan penuaian. Flavonoid terutamanya antosianin dan flavonol berperanan penting
dalam perkembangan warna bagi buah dan tumbuhan (Lancaster & Dougall 1992; Palapol
et al. 2009b). Salah satu metabolit yang berperanan sebagai pelopor bagi biosintesis
flavonoid telah dikenalpasti dalam kajian ini sebagai asid kuinik (Albertini et al. 2006;
Leuschner et al. 1995). Pengenalpastian asid kuinik di dalam perikarpa manggis
terutamanya pada peringkat terakhir pemasakan iaitu peringkat 6 (Rajah 4.14e) mungkin
disebabkan biosintesis kumpulan metabolit flavonoid adalah aktif pada peringkat ini.
Kuersetin 3,3'-dimetil eter 4'-glukosida yang dikategorikan sebagai kuersetin
(sejenis bioflavonoid) dikenalpasti sebagai metabolit berpengaruh tinggi (VIP ≥1.00 dan p
≤0.05) daripada manggis dan merupakan metabolit unik kepada sampel peringkat 0 dan
sampel perikarpa daripada peringkat 2 (Rajah 4.15a). Kuersetin merupakan flavonoid
berwarna kuning yang terhasil daripada tapak jalan biosintesis yang sama dengan
antosianin (Lancaster & Dougall 1992). Kuersetin telah dilaporkan menunjukkan aktiviti
anti-oksida dalam pelbagai buah dan tumbuhan (Miean & Mohamed 2001; Sultana &
Anwar 2008). Menurut kajian oleh Lancaster dan Dougall (1992), kuersetin dan kuersetin
glikosida banyak didapati di dalam kulit buah epal hijau seperti kultivar ”Granny Smith”
dan “Golden Delicious”. Manakala di dalam manggis, kehadiran metabolit ini pada
peringkat awal pemasakan mungkin berkait dengan warna hijau kekuningan pada perikarpa
manggis muda. Metabolit ini tidak dapat dikesan pada peringkat pertengahan (peringkat 4)
dan peringkat akhir pemasakan (peringkat 6) setelah manggis bertukar warna kepada merah
keperangan dan ungu gelap (Rajah 4.15a). Perubahan warna ini mungkin disebabkan
kehadiran antosianin yang lain seperti sianidin-3-soforosida dan sianidin-3-glukosida yang
dilaporkan meningkat seiring dengan proses pemasakan manggis (Palapol et al. 2009b).
Fungsi individu bagi kebanyakan fenolik yang telah dikenalpasti adalah masih
belum diketahui secara jelas dan terperinci disebabkan tahap metabolit ini yang berbeza-
beza dan tidak konsisten di dalam tisu manggis sepanjang proses pemasakan. Sebagai
contoh, epirobinatinidol-(4β,8)-katesin adalah proantosianidin yang telah ditemui dalam
batang pokok Acacia mearnsii (Kusano et al. 2010). Metabolit ini belum pernah dilaporkan
dalam manggis, namun, unit kumpulan berfungsi katesin telah dilaporkan dalam beberapa

110
kajian lepas sebagai komponen utama dalam proantosianidin yang ditemui di dalam
manggis (Fu et al. 2007; Yoshimura et al. 2015). Secara amnya pula, proantosianidin dan
fenolik secara keseluruhannya adalah berfungsi sebagai pemangsa radikal bebas yang
sangat kuat. Fungsi kumpulan metabolit ini secara langsung menyumbang kepada
kepentingan serta kelebihan buah dan tumbuhan terhadap aspek kesihatan manusia (Fu et
al. 2007; Naczk & Shahidi 2006; Prior & Gu 2005). Namun, mekanisme spesifik bagi
metabolit epirobinatinidol-(4β,8)-katesin ini dan metabolit fenolik yang lain memerlukan
kajian lanjutan yang lebih mendalam dan fokus terhadap fungsi di dalam pemasakan
manggis.
Kumpulan terpenoid seperti timol-β-D-glukopiranosida (Rajah 4.11h), giberelin A3,
ginggolida C, katalposida, montanol dan salvinorin A (Rajah 4.14j) memainkan pelbagai
fungsi di dalam tumbuhan dan buah termasuklah sebagai komponen hormon fotosintesis,
pengawalaturan tumbuhan serta pertahanan terhadap bakteria dan patogen kulat (Croteau
et al. 2000; McGarvey & Croteau 1995). Mazlan et al. (2018b) melaporkan bahawa timol-
β-D-glukopiranosida mungkin terlibat secara signifikan dalam pertahanan manggis
terhadap serangan patogen kerana kumpulan berfungsi timol telah dilaporkan mempunyai
ciri-ciri anti-mikrob (Nabavi et al. 2015) dan anti-fungal (Yang et al. 2011). Timol-β-D-
glukopiranosida yang merupakan monoterpena ini mungkin memainkan peranan yang
sama di dalam manggis semasa peringkat awal pemasakan di mana kadar metabolit ini
adalah tinggi di dalam perikarpa manggis pada peringkat 2 (Rajah 4.11h). Metabolit ini
juga mungkin menyumbang kepada perkembangan rasa dan aroma manggis (Ikan et al.
1993).
Mod pengionan negatif ESI-MS yang digunakan bagi menjalankan analisis LC-MS
dalam kajian ini telah dilaporkan mampu meningkatkan fragmentasi metabolit xanton,
sekaligus memudahkan proses pengenalpastian dan penentuan struktur xanton (Wittenauer
et al. 2012). Namun, hanya tujuh xanton berjaya dikenalpasti daripada kajian ini (Lampiran
K) dan empat daripadanya (α-mangostin, astikolorin B, gartanin dan kratoksiarborenon C)
dikenalpasti sebagai metabolit berpengaruh tinggi dalam pemasakan manggis (Rajah
4.14k). Hal ini mungkin kerana jenis pelarut yang digunakan dalam kaedah pengekstrakan

111
metabolit memfokuskan kepada pengekstrakan keseluruhan metabolit secara global
merentasi pelbagai kumpulan metabolit terutamanya metabolit sekunder dan tidak spesifik
kepada hanya xanton.
α-Mangostin adalah salah satu xanton utama dalam manggis dan metabolit ini telah
dilaporkan mempunyai pelbagai bioaktiviti dalam tumbuhan iaitu anti-oksida (Fang et al.
2016; Thong et al. 2015), anti-tumor (Hafeez et al. 2014; Shibata et al. 2011), anti-bakteria
(Koh et al. 2013) dan anti-radang (Mohan et al. 2018). Pengenalpastian α-mangostin di
dalam kajian ini turut disokong oleh data analisis MS/MS serta metabolit piawai.
Berdasarkan Jadual 4.8 dan Lampiran O, puncak pradominan bagi α-mangostin
dikenalpasti pada m/z 351 adalah terhasil melalui kehilangan radikal C3H7 (-43 Da)
manakala fragmentasi pada m/z 339 pula terhasil daripada reaksi ion radikal daripada
kumpulan fenil yang menyebabkan kehilangan radikal C4H7 (-55 Da) (Wittenauer et al.
2012). Dalam kajian ini, α-mangostin dikenalpasti menunjukkan kandungan yang paling
tinggi di dalam tisu perikarpa pada peringkat 2 dan 4 namun pada peringkat 6, kadar
metabolit ini menurun sehingga menjadi lebih rendah berbanding isi dan biji manggis
(Rajah 4.14k). Sebaliknya kajian lepas oleh Wittenauer et al. (2012) menyatakan bahawa
kandungan α-mangostin adalah lebih tinggi di dalam perikarpa berbanding isi dan biji
manggis. Perbezaan ini mungkin disebabkan penggunaan peringkat manggis yang lebih
awal oleh Wittenauer et al. (2012). Selain itu, metabolit ini mungkin larut ke dalam tisu
manggis pada peringkat akhir pemasakan, menyebabkan pengumpulan pada tisu isi dan
sekaligus memberikan manfaat kepada kesihatan.
Sementara itu, fragmentasi ion bagi gartanin (m/z 395) pula dilaporkan berlaku pada
m/z 271, 283 dan 339 pada masa penahanan minit ke 15.47, iaitu sebelum α-mangostin
pada minit ke 15.83, sepadan dengan kajian yang dijalankan oleh Wittenauer et al. (2012)
dan Zarena dan Sankar (2009). Gartanin hanya dikenalpasti di dalam perikarpa sahaja
dengan kadar yang meningkat daripada peringkat 4 ke peringkat 6, sejajar dengan laporan
oleh Wittenauer et al. (2012). Peningkatan pada kadar kandungan α-mangostin, gartanin
dan xanton yang lain iaitu astikolorin B dan kratoksiarborenon C di dalam biji manggis
apabila menghampiri peringkat akhir pemasakan (Rajah 4.14k) mungkin berkait dengan

112
fungsi utama xanton sebagai anti-oksida dan anti-mikrob dalam mekanisme perlindungan
buah terhadap jangkitan patogen dan tekanan biotik (Gutierrez-Orozco & Failla 2013;
Thong et al. 2015; Wittenauer et al. 2012). Radikal bebas boleh terhasil sebagai produk
sampingan daripada proses metabolisme buah dan boleh meningkat apabila buah
mengalami tekanan persekitaran (Fernie et al. 2004; Van Dongen et al. 2011).
Selain itu, terdapat tiga xanton yang belum pernah dilaporkan di dalam manggis
telah dikenalpasti melalui kajian ini iaitu astikolorin B, kratoksiarborenon C dan 1,3,5-
trihidroksixanton. Pengenalpastian dua daripada tiga xanton ini turut disokong dengan data
daripada analisis LC-MS/MS seperti yang dinyatakan di dalam Jadual 4.8 bagi
kratoksiarborenon C dan Lampiran N bagi 1,3,5-trihidroksixanton. Berdasarkan kajian
lepas, kehadiran kratoksiarborenon C telah dilaporkan di dalam Cratoxylum sumatranum
iaitu sejenis tumbuhan berbunga daripada famili Hypericaceae (Seo et al. 2002). Metabolit
ini mempunyai aktiviti sitotoksik sederhana terhadap turunan sel epidermoid oral manusia
(Seo et al. 2002). Manakala, 1,3,5-trihidroksixanton pula dilaporkan hadir dalam Garcinia
smeathmannii (Komguem et al. 2005) dan Garcinia assigu (Ito et al. 1998). Metabolit ini
turut dilaporkan mempunyai aktiviti anti-malaria terhadap parasit Plasmodium falciparum
(Ignatushchenko et al. 2000) dan aktiviti anti-virus (Ito et al. 1998). Penentuan struktur
molekul keempat-empat xanton ini daripada manggis menggunakan analisis NMR
mungkin diperlukan sebagai kajian lanjutan pada masa akan datang untuk pengenalpastian
identiti sebatian ini dengan lebih tepat.
Terdapat sebanyak 102 metabolit yang tidak dapat dikenalpasti telah diperoleh
daripada analisis LC-MS (Lampiran L). Analisis LC-MS/MS turut dijalankan dengan
menyasarkan kepada pengenalpastian metabolit ini namun tiada pengenalpastian yang tepat
diperoleh (tiada padanan antara fragmentasi metabolit dengan pangkalan data yang dirujuk
atau fragmentasi didapati sepadan dengan lebih daripada satu metabolit). Oleh itu, kajian
lanjutan iaitu kajian metabolomik sasaran yang melibatkan pemencilan metabolit
menggunakan kaedah pengekstrakan yang lebih spesifik dan analisis metabolit
menggunakan NMR mungkin diperlukan untuk menentukan struktur dan identiti metabolit

113
ini. Selain itu, metabolit ini juga boleh dikaji dari segi bioaktivitinya yang mungkin
berpotensi dalam aplikasi nutraseutikal dan farmaseutikal pada masa hadapan.

114
BAB VI
KESIMPULAN DAN CADANGAN
6.1 KESIMPULAN
Kajian komprehensif terhadap perubahan metabolik semasa pemasakan buah manggis
telah dijalankan menggunakan platform analitikal GC-MS dan LC-MS. Awalnya,
beberapa kaedah pengekstrakan telah dikaji dan dioptimumkan untuk analisis
metabolomik ini. Kaedah pengekstrakan menggunakan campuran larutan
metanol/kloroform/air dengan nisbah 3:1:1 disertai teknik sonikasi (kaedah 5) dan
diikuti dengan derivatisasi sampel menggunakan BSTFA telah digunakan untuk
memprofil kandungan metabolit manggis menggunakan GC-MS. Ini adalah kerana
kaedah ini telah menghasilkan jumlah metabolit tertinggi berbanding kaedah yang lain.
Kaedah ini juga menunjukkan hasil pemisahan puncak yang jelas, tajam serta
merangkumi bilangan puncak yang lebih banyak berbanding kaedah lain. Manakala,
kaedah pengekstrakan menggunakan larutan metanol/asid formik dengan nisbah 599:1
(kaedah 1) pula digunakan bagi pemprofilan metabolit menggunakan analisis LC-MS
manakala mod pengionan sistem LC-MS pula ditetapkan kepada mod negatif. Hal ini
kerana kaedah dan protokol ini dilihat lebih berkesan dalam pengekstrakan metabolit
sekunder berbanding kaedah lain yang dibandingkan.
Daripada analisis yang dijalankan merangkumi sampel perikarpa, isi dan biji
manggis pada empat peringkat pemasakan yang berbeza (peringkat 0, 2, 4 dan 6),
analisis GC-MS berjaya memprofil sebanyak 57 metabolit manakala 98 metabolit pula
berjaya dikenalpasti melalui analisis LC-MS. Metabolit yang dikenalpasti merangkumi
pelbagai kelas metabolit termasuklah gula dan terbitan, asid amino dan terbitan, asid
organik, fenolik, terpenoid dan alkaloid dan xanton. Taburan metabolit antara peringkat
pemasakan dan jenis tisu manggis yang berbeza dapat dilihat dan dikaji menggunakan

115
analisis statistik multivariat termasuklah analisis PCA dan PLS-DA. Analisis
pembolehubah berkepentingan dalam unjuran (VIP) disertai dengan analisis statistik
univariat (analisis varians dua-hala) pula mendedahkan kehadiran 21 metabolit
berpengaruh tinggi (VIP ≥1.00 and p ≤0.05) daripada analisis GC-MS dan 55 metabolit
berpengaruh tinggi daripada analisis LC-MS yang terlibat dalam pelbagai tapak jalan
biosintesis. Antaranya adalah tapak jalan metabolisme galaktosa, metabolisme askorbat
dan aldarat, metabolisme glioksilat dan dikarboksilat, penyalingtukaran pentosa dan
glukuronat dan kitaran asid trikarboksilik (TCA). Metabolit yang berjaya dikenalpasti
dalam kajian ini telah dipetakan ke dalam tapak jalan metabolomik pemasakan manggis
yang menggabungkan beberapa tapak jalan berlainan.
Kajian ini adalah yang pertama mengkaji tentang pemasakan manggis melalui
pemprofilan metabolit perikarpa, isi dan biji manggis pada peringkat pemasakan yang
berbeza menggunakan gabungan platform GC-MS dan LC-MS. Tapak jalan pemasakan
manggis juga adalah yang pertama dilaporkan menggabungkan beberapa tapak jalan
yang berlainan seperti tapak jalan kanji dan sukrosa, glikolisis serta metabolisme
sisteina dan metionina. Pemetaan metabolit ke dalam tapak jalan pemasakan manggis
ini merupakan hasil yang berimpak besar diperoleh daripada kajian ini. Secara
keseluruhannya, pendekatan metabolomik khususnya menggunakan platform analitikal
GC-MS dan LC-MS disertai analisis data seperti analisis univariat dan multivariat
berupaya merungkai perubahan metabolit di dalam pelbagai tisu manggis pada
peringkat pemasakan yang berbeza. Teknik ini boleh digunakan untuk mengkaji
pengawalaturan, biosintesis serta fungsi metabolit yang membawa kepada pemahaman
yang lebih baik terhadap biokimia tumbuhan dalam konteks pemasakan buah. Platform
serta teknik bagi kajian metabolomik yang digunakan dalam kajian ini adalah bersifat
universal dan sesuai untuk diaplikasikan terhadap mana-mana sampel tumbuhan.
Pemahaman mengenai perubahan metabolit semasa proses pemasakan manggis yang
diperoleh melalui kajian ini diharapkan dapat menjadi asas kepada kajian akan datang
untuk merungkai dengan lebih mendalam terhadap manggis yang mempunyai pelbagai
potensi dan kelebihan dari segi kesihatan mahupun prospek ekonomi negara.

116
6.2 CADANGAN KAJIAN LANJUTAN
Antara cabaran atau limitasi yang dihadapi dalam kajian ini adalah proses
pengenalpastian metabolit bagi data yang diperoleh daripada analisis LC-MS yang
kompleks dan melibatkan masa kajian yang lama. Hal ini kerana analisis LC-MS tidak
mempunyai pangkalan data yang khusus dan kukuh seperti analisis GC-MS yang
mempunyai perpustakaan sebatian rujukan yang sangat teratur dan tepat seperti
National Institute of Standards and Technology (NIST). Oleh itu, pembangunan
pangkalan data yang khusus adalah sangat penting bagi melancarkan serta
mengurangkan masa yang diperlukan bagi proses pengenalpastian metabolit bagi
analisis LC-MS seterusnya menghasilkan dapatan kajian yang lebih tepat dan berkesan.
Kajian pada masa akan datang mungkin boleh difokuskan kepada pembangunan
pangkalan data dalaman (in-house) yang seterusnya boleh digunakan bagi analisis
metabolomik tumbuhan yang lain.
Selain itu, kajian yang lebih mendalam terhadap metabolit yang telah berjaya
dikenalpasti melalui kajian ini dipercayai dapat merungkai dengan lebih banyak dan
lebih terperinci mengenai fungsi metabolit ini sama ada terhadap pemasakan manggis
sendiri atau fungsi lain sebagai metabolit bioaktif. Sebagai contoh, kajian ini telah
melaporkan kehadiran lignan (neoisostegana) dalam manggis. Sesetengah lignan telah
dilaporkan mempunyai keupayaan untuk mempengaruhi perkembangan serangga
dengan mengubah pelbagai fungsi fisiologi serangga tersebut melalui aktiviti
antagonistik ekdisteroid (Harmatha & Dinan 2003). Kajian terhadap metabolit lignan
ini mungkin boleh diaplikasikan untuk menghasilkan racun serangga biologi pada masa
akan datang seterusnya membantu dalam sektor pertanian negara.
Pengenalpastian tiga metabolit xanton iaitu 1,3,5-trihidroksixanton, astikolorin
B dan kratoksiarborenon C yang belum pernah dilaporkan dalam manggis juga adalah
sangat menarik untuk dikaji dengan lebih mendalam. Xanton 1,3,5-trihidroksixanton
telah dilaporkan mempunyai aktiviti anti-malaria (Ignatushchenko et al. 2000) dan anti-
virus (Ito et al. 1998) manakala kratoksiarborenon C pula telah dilaporkan mempunyai
aktiviti sitotoksik (Seo et al. 2002). Metabolit ini perlu dipencilkan terlebih dahulu
menggunakan kaedah pengekstrakan yang spesifik sebelum dianalisis untuk

117
mendapatkan maklumat mengenai struktur kimia molekul. Penerokaan terhadap
bioaktiviti metabolit ini serta metabolit xanton yang lain akan meningkatkan lagi nilai
manggis sebagai salah satu sumber sebatian bioaktif yang berkepentingan tinggi dalam
industri farmaseutikal dan perubatan.
Selain itu, sebanyak 102 metabolit yang tidak dapat dikenalpasti telah diperoleh
daripada analisis LC-MS dan telah dilaporkan dalam Lampiran L. Kajian metabolomik
sasaran menggunakan kaedah pengekstrakan yang lebih spesifik untuk memencilkan
metabolit ini dan diikuti dengan analisis analitik menggunakan NMR mungkin
diperlukan untuk menentukan struktur molekul dan identiti metabolit ini. Seterusnya,
kajian bioaktiviti juga boleh dilakukan sekiranya terdapat molekul yang berpotensi
sebagai sebatian bioaktif daripada manggis.

118
RUJUKAN
Abdel-Farid, I. B., Kim, H. K., Choi, Y. H. & Verpoorte, R. 2007. Metabolic
characterization of Brassica rapa leaves by NMR spectroscopy. Journal of
Agricultural and Food Chemistry 55(19): 7936-7943.
Abdul-Rahman, A., Goh, H.-H., Loke, K.-K., Noor, N. M. & Aizat, W. M. 2017. RNA-
seq analysis of mangosteen (Garcinia mangostana L.) fruit ripening. Genomics
Data 12: 159-160.
Abeles, F. B., Morgan, P. W. & Saltveit Jr, M. E. 2012. Ethylene in Plant Biology. Ed.
ke-2. New York: Elsevier Science.
Abu Dardak, R., Ab. Halim, N., Kasa, J. & Mahmood, Z. 2011. Challenges and prospect
of mangosteen industry in Malaysia. Economic and Technology Management
Review 6: 19-31.
Adnyana, I. K., Abuzaid, A. S., Isk, E. Y. & Kurniati, N. F. 2016. Pancreatic lipase and
a-amylase inhibitory potential of mangosteen (Garcinia mangostana Linn.)
pericarp extract. International Journal of Medical Research & Health Sciences
5(1): 23-28.
Aizat, W. M., Able, J. A., Stangoulis, J. C. & Able, A. J. 2013. Characterisation of
ethylene pathway components in non-climacteric capsicum. BMC Plant Biology
13(1): 191.
Aizat, W. M., Dias, D. A., Stangoulis, J. C., Able, J. A., Roessner, U. & Able, A. J.
2014. Metabolomics of capsicum ripening reveals modification of the ethylene
related-pathway and carbon metabolism. Postharvest Biology and Technology
89: 19-31.
Aizat, W. M., Ismail, I. & Noor, N. M. 2018. Recent Development in Omics Studies.
Dlm. Aizat, W. M., Goh, H.-H. & Baharum, S. N. (pnyt.). Omics Applications
for Systems Biology. Advances in Experimental Medicine and Biology, 1102.
hlm. 1-9. Bangi: Springer, Cham.
Aizat, W. M., Jamil, I. N., Ahmad-Hashim, F. H. & Noor, N. M. 2019. Recent updates
on metabolite composition and medicinal benefits of mangosteen plant. PeerJ
7: e6324.
Albertini, M.-V., Carcouet, E., Pailly, O., Gambotti, C., Luro, F. & Berti, L. 2006.
Changes in organic acids and sugars during early stages of development of
acidic and acidless citrus fruit. Journal of Agricultural and Food Chemistry
54(21): 8335-8339.
Anon. 2017. Liquid chromatography–mass spectrometry.
https://en.wikipedia.org/wiki/Liquid_chromatography%E2%80%93mass_spec
trometry [9 April 2019]

119
Apiratikorn, S., Sdoodee, S., Lerslerwong, L. & Rongsawat, S. 2012. The impact of
climatic variability on phenological change, yield and fruit quality of
mangosteen in Phatthalung province, Southern Thailand. Kasetsart Journal
(Natural Science) 46: 1-9.
Aretz, I. & Meierhofer, D. 2016. Advantages and pitfalls of mass spectrometry based
metabolome profiling in systems biology. International Journal of Molecular
Sciences 17(5): 632.
Ayman, E., Hassan, S. M. & Osman, H.-E. H. 2019. Mangosteen (Garcinia mangostana
L.). Dlm. Nabavi, S. M. N. & Silva, A. S. (pnyt.). Nonvitamin and Nonmineral
Nutritional Supplements, hlm. 313-319. Elsevier.
Azizan, K. A., Baharum, S. N., Ressom, H. W. & Noor, N. M. 2012. GC-MS analysis
and PLS-DA validation of the trimethyl silyl-derivatization techniques.
American Journal of Applied Sciences 9(7): 1124-1136.
Azizan, K. A., Ghani, N. H. A. & Nawawi, M. F. 2015. GC-MS based metabolomics
and multivariate statistical analysis of Wedelia trilobata extracts for the
identification of potential phytochemical properties. Plant Omics 8(6): 537-543.
Azizan, K. A., Ibrahim, S., Ghani, N. H. A. & Nawawi, M. F. 2016. LC-MS based
metabolomics analysis to identify potential allelochemicals in Wedelia
trilobata. Records of Natural Products 10(6): 788.
Baharum, S. N. & Azizan, K. A. 2018. Metabolomics in systems biology. Dlm. Aizat,
W. M., Goh, H.-H. & Baharum, S. N. (pnyt.). Omics Applications for Systems
Biology. Advances in Experimental Medicine and Biology, 1102. hlm. 51-68.
Springer.
Baharum, S. N., Bunawan, H., Ghani, M. a. A., Mustapha, W. a. W. & Noor, N. M.
2010. Analysis of the chemical composition of the essential oil of Polygonum
minus Huds. using two-dimensional gas chromatography-time-of-flight mass
spectrometry (GC-TOF MS). Molecules 15(10): 7006-7015.
Baldi, P., Orsucci, S., Moser, M., Brilli, M., Giongo, L. & Si-Ammour, A. 2018. Gene
expression and metabolite accumulation during strawberry (Fragaria x
ananassa) fruit development and ripening. Planta 248(5): 1143-1157.
Behrends, V., Tredwell, G. D. & Bundy, J. G. 2011. A software complement to AMDIS
for processing GC-MS metabolomic data. Analytical Biochemistry 415(2): 206-
208.
Bingol, K. 2018. Recent advances in targeted and untargeted metabolomics by NMR
and MS/NMR methods. High-Throughput 7(2): 9.
Blankenship, S. M. & Dole, J. M. 2003. 1-Methylcyclopropene: a review. Postharvest
Biology and Technology 28(1): 1-25.

120
Broeckling, C. D., Reddy, I. R., Duran, A. L., Zhao, X. & Sumner, L. W. 2006. MET-
IDEA: data extraction tool for mass spectrometry-based metabolomics.
Analytical Chemistry 78(13): 4334-4341.
Brummell, D. A. 2006. Cell wall disassembly in ripening fruit. Functional Plant
Biology 33(2): 103-119.
Brunner, B. R. & Morales-Payan, J. P. 2010. Soils,plant growth and crop production-
Mangosteen and rambutan. Encyclopedia of Life Support Systems 1-10.
Bürstenbinder, K., Rzewuski, G., Wirtz, M., Hell, R. & Sauter, M. 2007. The role of
methionine recycling for ethylene synthesis in Arabidopsis. The Plant Journal
49(2): 238-249.
Cadahia, E., Fernandez De Simon, B., Aranda, I., Sanz, M., Sanchez‐Gomez, D. &
Pinto, E. 2015. Non-targeted metabolomic profile of Fagus sylvatica L. leaves
using liquid chromatography with mass spectrometry and gas chromatography
with mass spectrometry. Phytochemical Analysis 26(2): 171-182.
Cadahía, E., Fernández De Simón, B., Aranda, I., Sanz, M., Sánchez‐Gómez, D. &
Pinto, E. 2015. Non‐targeted metabolomic profile of Fagus sylvatica L. leaves
using liquid chromatography with mass spectrometry and gas chromatography
with mass spectrometry. Phytochemical Analysis 26(2): 171-182.
Caffall, K. H. & Mohnen, D. 2009. The structure, function, and biosynthesis of plant
cell wall pectic polysaccharides. Carbohydrate Research 344(14): 1879-1900.
Cambiaghi, A., Ferrario, M. & Masseroli, M. 2017. Analysis of metabolomic data:
tools, current strategies and future challenges for omics data integration.
Briefings in Bioinformatics 18(3): 498-510.
Capitani, D., Sobolev, A. P., Tomassini, A., Sciubba, F., De Salvador, F. R., Mannina,
L. & Delfini, M. 2012. Peach fruit: metabolic comparative analysis of two
varieties with different resistances to insect attacks by NMR spectroscopy.
Journal of Agricultural and Food Chemistry 61(8): 1718-1726.
Cevallos-Cevallos, J. M., Reyes-De-Corcuera, J. I., Etxeberria, E., Danyluk, M. D. &
Rodrick, G. E. 2009. Metabolomic analysis in food science: a review. Trends in
Food Science & Technology 20(11-12): 557-566.
Chaijaroenkul, W., Mubaraki, M. A., Ward, S. A. & Na-Bangchang, K. 2014.
Metabolite footprinting of Plasmodium falciparum following exposure to
Garcinia mangostana Linn. crude extract. Experimental Parasitology 145(1):
80-86.
Chen, H.-H., Tseng, Y. J., Wang, S.-Y., Tsai, Y.-S., Chang, C.-S., Kuo, T.-C., Yao, W.-
J., Shieh, C.-C., Wu, C.-H. & Kuo, P.-H. 2015. The metabolome profiling and
pathway analysis in metabolic healthy and abnormal obesity. International
Journal of Obesity 39(8): 1241.

121
Chen, L.-G., Yang, L.-L. & Wang, C.-C. 2008. Anti-inflammatory activity of
mangostins from Garcinia mangostana. Food and Chemical Toxicology 46(2):
688-693.
Cheng, A. X., Lou, Y. G., Mao, Y. B., Lu, S., Wang, L. J. & Chen, X. Y. 2007. Plant
terpenoids: biosynthesis and ecological functions. Journal of Integrative Plant
Biology 49(2): 179-186.
Chin, Y.-W., Chai, H.-B., Keller, W. J. & Kinghorn, A. D. 2008. Lignans and other
constituents of the fruits of Euterpe oleracea (acai) with antioxidant and
cytoprotective activities. Journal of Agricultural and Food Chemistry 56(17):
7759-7764.
Crookes, P. R. & Grierson, D. 1983. Ultrastructure of tomato fruit ripening and the role
of polygalacturonase isoenzymes in cell wall degradation. Plant Physiology
72(4): 1088-1093.
Croteau, R., Kutchan, T. M. & Lewis, N. G. 2000. Natural products (secondary
metabolites). Dlm. Buchanan, B. B., Gruissem, W. & Jones, R. L. (pnyt.).
Biochemistry and Molecular Biology of Plants, 24. hlm. 1250-1318. Rockville:
American Society of Plant Physiologists.
Dai, J. & Mumper, R. J. 2010. Plant phenolics: extraction, analysis and their antioxidant
and anticancer properties. Molecules 15(10): 7313-7352.
Das, P. R. & Eun, J.-B. 2018. A comparative study of ultra-sonication and agitation
extraction techniques on bioactive metabolites of green tea extract. Food
Chemistry 253: 22-29.
De Vos, R. C., Moco, S., Lommen, A., Keurentjes, J. J., Bino, R. J. & Hall, R. D. 2007.
Untargeted large-scale plant metabolomics using liquid chromatography
coupled to mass spectrometry. Nature Protocols 2(4): 778-791.
Defilippi, B. G., Manríquez, D., Luengwilai, K. & González-Agüero, M. 2009. Aroma
volatiles: biosynthesis and mechanisms of modulation during fruit ripening.
Advances in Botanical Research 50: 1-37.
Deng, C., Yin, X., Zhang, L. & Zhang, X. 2005. Development of microwave‐assisted
derivatization followed by gas chromatography/mass spectrometry for fast
determination of amino acids in neonatal blood samples. Rapid
Communications in Mass Spectrometry 19(16): 2227-2234.
Denman, L. J. & Morris, G. A. 2015. An experimental design approach to the chemical
characterisation of pectin polysaccharides extracted from Cucumis melo
Inodorus. Carbohydrate Polymers 117: 364-369.
Devlieghere, F., Vermeulen, A. & Debevere, J. 2004. Chitosan: antimicrobial activity,
interactions with food components and applicability as a coating on fruit and
vegetables. Food Microbiology 21(6): 703-714.

122
Dixon, J. & Hewett, E. W. 2000. Factors affecting apple aroma/flavour volatile
concentration: a review. New Zealand Journal of Crop and Horticultural
Science 28(3): 155-173.
Dobson, G., Shepherd, T., Verrall, S. R., Conner, S., Mcnicol, J. W., Ramsay, G.,
Shepherd, L. V., Davies, H. V. & Stewart, D. 2008. Phytochemical diversity in
tubers of potato cultivars and landraces using a GC-MS metabolomics approach.
Journal of Agricultural and Food Chemistry 56(21): 10280-10291.
Drouin, N., Rudaz, S. & Schappler, J. 2018. Sample preparation for polar metabolites
in bioanalysis. Analyst 143(1): 16-20.
El-Seedi, H. R., El-Barbary, M., El-Ghorab, D., Bohlin, L., Borg-Karlson, A.-K.,
Goransson, U. & Verpoorte, R. 2010. Recent insights into the biosynthesis and
biological activities of natural xanthones. Current Medicinal Chemistry 17(9):
854-901.
El-Seedi, H. R., El-Ghorab, D. M., El-Barbary, M. A., Zayed, M. F., Goransson, U.,
Larsson, S. & Verpoorte, R. 2009. Naturally occurring xanthones; latest
investigations: isolation, structure elucidation and chemosystematic
significance. Current Medicinal Chemistry 16(20): 2581-2626.
Failla, M. L. & Gutiérrez-Orozco, F. 2017. Mangosteen xanthones: Bioavailability and
bioactivities. Fruit and Vegetable Phytochemicals: Chemistry and Human
Health 1(2): 165-182.
Fairchild, D. 1915. The mangosteen “Queen of Fruits” now almost confined to Malayan
archipelago, but can be acclimated in many parts of tropics-experiments in
America-desirability of widespread cultivation. Journal of Heredity 6(8): 339-
347.
Fait, A., Hanhineva, K., Beleggia, R., Dai, N., Rogachev, I., Nikiforova, V. J., Fernie,
A. R. & Aharoni, A. 2008. Reconfiguration of the achene and receptacle
metabolic networks during strawberry fruit development. Plant Physiology
148(2): 730-750.
Fang, Y., Su, T., Qiu, X., Mao, P., Xu, Y., Hu, Z., Zhang, Y., Zheng, X., Xie, P. & Liu,
Q. 2016. Protective effect of alpha-mangostin against oxidative stress induced-
retinal cell death. Scientific Reports 6: 21018.
Fernie, A. R., Carrari, F. & Sweetlove, L. J. 2004. Respiratory metabolism: glycolysis,
the TCA cycle and mitochondrial electron transport. Current Opinion in Plant
Biology 7(3): 254-261.
Fiehn, O. 2002. Metabolomics-The link between genotypes and phenotypes. Plant
Molecular Biology 48(1-2): 155-171.

123
Fiehn, O. 2008. Extending the breadth of metabolite profiling by gas chromatography
coupled to mass spectrometry. TrAC Trends in Analytical Chemistry 27(3): 261-
269.
Fiehn, O. 2016. Metabolomics by gas chromatography–mass spectrometry: combined
targeted and untargeted profiling. Current Protocols in Molecular Biology
114(1): 1-32.
Fu, C., Loo, A. E. K., Chia, F. P. P. & Huang, D. 2007. Oligomeric proanthocyanidins
from mangosteen pericarps. Journal of Agricultural and Food Chemistry
55(19): 7689-7694.
Garg, N., Kapono, C. A., Lim, Y. W., Koyama, N., Vermeij, M. J., Conrad, D., Rohwer,
F. & Dorrestein, P. C. 2015. Mass spectral similarity for untargeted
metabolomics data analysis of complex mixtures. International Journal Of
Mass Spectrometry 377: 719-727.
Gellekink, H., Van Oppenraaij-Emmerzaal, D., Van Rooij, A., Struys, E. A., Den
Heijer, M. & Blom, H. J. 2005. Stable-isotope dilution liquid chromatography–
electrospray injection tandem mass spectrometry method for fast, selective
measurement of S-adenosylmethionine and S-adenosylhomocysteine in plasma.
Clinical Chemistry 51(8): 1487-1492.
Giovannoni, J. 2001. Molecular biology of fruit maturation and ripening. Annual
Review of Plant Biology 52(1): 725-749.
Giovannoni, J., Nguyen, C., Ampofo, B., Zhong, S. & Fei, Z. 2017. The epigenome and
transcriptional dynamics of fruit ripening. Annual Review of Plant Biology 68:
61-84.
Gondokesumo, M. E., Pardjianto, B., Sumitro, S. B. & Widowati, W. 2019. Xanthones
analysis and antioxidant activity analysis (applying ESR) of six different
maturity levels of mangosteen rind extract (Garcinia mangostana Linn.).
Pharmacognosy Journal 11(2): 369-373.
Goodacre, R., Vaidyanathan, S., Dunn, W. B., Harrigan, G. G. & Kell, D. B. 2004.
Metabolomics by numbers: acquiring and understanding global metabolite data.
Trends in Biotechnology 22(5): 245-252.
Grassmann, J. 2005. Terpenoids as plant antioxidants. Vitamins & Hormones 72: 505-
535.
Griffiths, W. J., Koal, T., Wang, Y., Kohl, M., Enot, D. P. & Deigner, H. P. 2010.
Targeted metabolomics for biomarker discovery. Angewandte Chemie
International Edition 49(32): 5426-5445.
Gullberg, J., Jonsson, P., Nordstrom, A., Sjostrom, M. & Moritz, T. 2004. Design of
experiments: an efficient strategy to identify factors influencing extraction and

124
derivatization of Arabidopsis thaliana samples in metabolomic studies with gas
chromatography/mass spectrometry. Analytical Biochemistry 331(2): 283-295.
Gutierrez-Orozco, F. & Failla, M. L. 2013. Biological activities and bioavailability of
mangosteen xanthones: a critical review of the current evidence. Nutrients 5(8):
3163-3183.
Hafeez, B. B., Mustafa, A., Fischer, J. W., Singh, A., Zhong, W., Shekhani, M. O.,
Meske, L., Havighurst, T., Kim, K. & Verma, A. K. 2014. α-Mangostin: a
dietary antioxidant derived from the pericarp of Garcinia mangostana L.
inhibits pancreatic tumor growth in xenograft mouse model. Antioxidants &
Redox Signaling 21(5): 682-699.
Halket, J. M., Waterman, D., Przyborowska, A. M., Patel, R. K., Fraser, P. D. &
Bramley, P. M. 2005. Chemical derivatization and mass spectral libraries in
metabolic profiling by GC/MS and LC/MS/MS. Journal of Experimental
Botany 56(410): 219-243.
Hallett, I. C., Macrae, E. A. & Wegrzyn, T. F. 1992. Changes in kiwifruit cell wall
ultrastructure and cell packing during postharvest ripening. International
Journal of Plant Sciences 153(1): 49-60.
Hamid, M. A., Sarmidi, M. R. & Park, C. S. 2012. Mangosteen leaf extract increases
melanogenesis in B16F1 melanoma cells by stimulating tyrosinase activity in
vitro and by up-regulating tyrosinase gene expression. International Journal Of
Molecular Medicine 29(2): 209-217.
Hapsari, D. P., Poerwanto, R., Sopandie, D. & Santosa, E. 2018. Partial root-zone
irrigation effects on growth, metabolism and calcium status of Mangosteen
seedling (Garcinia mangostana L.). Advances in Horticultural Science 32(1):
49-59.
Harbourne, N., Marete, E., Jacquier, J. C. & O'riordan, D. 2009. Effect of drying
methods on the phenolic constituents of meadowsweet (Filipendula ulmaria)
and willow (Salix alba). LWT-Food Science and Technology 42(9): 1468-1473.
Harker, F. & Sutherland, P. 1993. Physiological changes associated with fruit ripening
and the development of mealy texture during storage of nectarines. Postharvest
Biology and Technology 2(4): 269-277.
Harmatha, J. & Dinan, L. 2003. Biological activities of lignans and stilbenoids
associated with plant-insect chemical interactions. Phytochemistry Reviews
2(3): 321-330.
Harrison, L. J. & Nilar. 2002. Xanthones from the heartwood of Garcinia mangostana.
Phytochemistry 60(5): 541-548.

125
Hesse, H., Kreft, O., Maimann, S., Zeh, M. & Hoefgen, R. 2004. Current understanding
of the regulation of methionine biosynthesis in plants. Journal of Experimental
Botany 55(404): 1799-1808.
Horai, H., Arita, M., Kanaya, S., Nihei, Y., Ikeda, T., Suwa, K., Ojima, Y., Tanaka, K.,
Tanaka, S. & Aoshima, K. 2010. MassBank: a public repository for sharing
mass spectral data for life sciences. Journal of Mass Spectrometry 45(7): 703-
714.
Huhman, D. V. & Sumner, L. W. 2002. Metabolic profiling of saponins in Medicago
sativa and Medicago truncatula using HPLC coupled to an electrospray ion-trap
mass spectrometer. Phytochemistry 59(3): 347-360.
Hurtado-Fernández, E., Pacchiarotta, T., Mayboroda, O., Fernández-Gutiérrez, A. &
Carrasco-Pancorbo, A. 2015. Metabolomic analysis of avocado fruits by GC-
APCI-TOF MS: effects of ripening degrees and fruit varieties. Analytical and
Bioanalytical Chemistry 407(2): 547-555.
Hussain, S. Z. & Maqbool, K. 2014. GC-MS: Principle, technique and its application in
food science. International Journal of Current Science 13: 116-126.
Ibrahim, M. Y., Hashim, N. M., Mariod, A. A., Mohan, S., Abdulla, M. A.,
Abdelwahab, S. I. & Arbab, I. A. 2016. α-Mangostin from Garcinia mangostana
Linn.: an updated review of its pharmacological properties. Arabian Journal of
Chemistry 9(3): 317-329.
Ignatushchenko, M. V., Winter, R. W. & Riscoe, M. 2000. Xanthones as antimalarial
agents: stage specificity. The American Journal of Tropical Medicine and
Hygiene 62(1): 77-81.
Ikan, R., Weinstein, V., Milner, Y., Bravdo, B., Shoseyov, O., Segal, D., Altman, A. &
Chet, I. 1993. Natural glycosides as potential odorants and flavorants.
International Symposium on Medicinal and Aromatic Plants hlm. 17-28.
Ito, C., Itoigawa, M., Furukawa, H., Rao, K. S., Enjo, F., Bu, P., Takayasu, J., Tokuda,
H. & Nishino, H. 1998. Xanthones as inhibitors of Epstein–Barr virus
activation1. Cancer Letters 132(1-2): 113-117.
Jabatan Pertanian Negeri Pulau Pinang. 2018. Keterangan am.
http://jpn.penang.gov.my/index.php/teknologi-tanaman-2/buah-buahan/67-
manggis-sp-17566 [15 Mei 2019]
Jarvis, M., Briggs, S. & Knox, J. 2003. Intercellular adhesion and cell separation in
plants. Plant, Cell & Environment 26(7): 977-989.
Ji, X., Avula, B. & Khan, I. A. 2007. Quantitative and qualitative determination of six
xanthones in Garcinia mangostana L. by LC–PDA and LC–ESI-MS. Journal
of Pharmaceutical and Biomedical Analysis 43(4): 1270-1276.

126
Jia, H.-F., Chai, Y.-M., Li, C.-L., Lu, D., Luo, J.-J., Qin, L. & Shen, Y.-Y. 2011.
Abscisic acid plays an important role in the regulation of strawberry fruit
ripening. Plant Physiology 157(1): 188-199.
Jianglian, D. & Shaoying, Z. 2013. Application of chitosan based coating in fruit and
vegetable preservation: a review. Journal of Food Processing & Technology
4(5): 227.
Johnson, C. H., Ivanisevic, J. & Siuzdak, G. 2016. Metabolomics: beyond biomarkers
and towards mechanisms. Nature Reviews Molecular Cell biology 17(7): 451.
Jolliffe, I. 2011. Principal component analysis. Dlm. Lovric, M. (pnyt.). International
Encyclopedia Of Statistical Science, hlm. 1094-1096. Berlin: Springer.
Kabera, J. N., Semana, E., Mussa, A. R. & He, X. 2014. Plant secondary metabolites:
biosynthesis, classification, function and pharmacological properties. Journal of
Pharmacy and Pharmacology 2: 377-392.
Kampa, M., Alexaki, V.-I., Notas, G., Nifli, A.-P., Nistikaki, A., Hatzoglou, A.,
Bakogeorgou, E., Kouimtzoglou, E., Blekas, G. & Boskou, D. 2004.
Antiproliferative and apoptotic effects of selective phenolic acids on T47D
human breast cancer cells: potential mechanisms of action. Breast Cancer
Research 6(2): R63.
Kanehisa, M. & Goto, S. 2000. KEGG: kyoto encyclopedia of genes and genomes.
Nucleic Acids Research 28(1): 27-30.
Kang, J.-S. 2012. Principles and applications of LC-MS/MS for the quantitative
bioanalysis of analytes in various biological samples. Tandem Mass
Spectrometry–Applications and Principles 441-492.
Kaufmann, B. & Christen, P. 2002. Recent extraction techniques for natural products:
microwave-assisted extraction and pressurised solvent extraction.
Phytochemical Analysis 13(2): 105-113.
Kim, H.-S., Park, S. J., Hyun, S.-H., Yang, S.-O., Lee, J., Auh, J.-H., Kim, J.-H., Cho,
S.-M., Marriott, P. J. & Choi, H.-K. 2011. Biochemical monitoring of black
raspberry (Rubus coreanus Miquel) fruits according to maturation stage by 1H
NMR using multiple solvent systems. Food Research International 44(7): 1977-
1987.
Kim, H. K. & Verpoorte, R. 2010. Sample preparation for plant metabolomics.
Phytochemical Analysis 21(1): 4-13.
Kind, T., Tolstikov, V., Fiehn, O. & Weiss, R. H. 2007. A comprehensive urinary
metabolomic approach for identifying kidney cancer. Analytical Biochemistry
363(2): 185-195.

127
Kitano, H. 2002. Systems biology: a brief overview. Science 295(5560): 1662-1664.
Klaser Cheng, D. M. 2017. Ethnobotany: A Phytochemical Perspective. Wiley-
Blackwell.
Klipp, E., Liebermeister, W., Wierling, C., Kowald, A. & Herwig, R. 2016. Systems
Biology: A Textbook. Berlin: John Wiley & Sons.
Knapp, D. R. 1979. Handbook of analytical derivatization reactions. New York: John
Wiley & Sons.
Koh, J.-J., Qiu, S., Zou, H., Lakshminarayanan, R., Li, J., Zhou, X., Tang, C.,
Saraswathi, P., Verma, C. & Tan, D. T. 2013. Rapid bactericidal action of alpha-
mangostin against MRSA as an outcome of membrane targeting. Biochimica et
Biophysica Acta (BBA)-Biomembranes 1828(2): 834-844.
Komguem, J., Meli, A., Manfouo, R., Lontsi, D., Ngounou, F., Kuete, V., Kamdem, H.
W., Tane, P., Ngadjui, B. T. & Sondengam, B. L. 2005. Xanthones from
Garcinia smeathmannii (oliver) and their antimicrobial activity. Phytochemistry
66(14): 1713-1717.
Kondo, S., Ponrod, W., Kanlayanarat, S. & Hirai, N. 2002. Abscisic acid metabolism
during fruit development and maturation of mangosteens. Journal of the
American Society for Horticultural Science 127(5): 737-741.
Kou, X. & Wu, M. 2018. Characterization of climacteric and non-climacteric fruit
ripening. Dlm. Guo, Y. (pnyt.). Plant Senescence. Methods in Molecular
Biology, 1744. hlm. 89-102. New York: Humana Press.
Krug, D., Zurek, G., Schneider, B., Garcia, R. & Müller, R. 2008. Efficient mining of
myxobacterial metabolite profiles enabled by liquid chromatography–
electrospray ionisation-time-of-flight mass spectrometry and compound-based
principal component analysis. Analytica Chimica Acta 624(1): 97-106.
Kühnel, E., Laffan, D. D., Lloyd‐Jones, G. C., Martínez Del Campo, T., Shepperson, I.
R. & Slaughter, J. L. 2007. Mechanism of methyl esterification of carboxylic
acids by trimethylsilyldiazomethane. Angewandte Chemie International Edition
46(37): 7075-7078.
Kusano, R., Ogawa, S., Matsuo, Y., Tanaka, T., Yazaki, Y. & Kouno, I. 2010. α-
Amylase and lipase inhibitory activity and structural characterization of acacia
bark proanthocyanidins. Journal of Natural Products 74(2): 119-128.
Kushnir, M. M. & Komaromy-Hiller, G. 2000. Optimization and performance of a rapid
gas chromatography–mass spectrometry analysis for methylmalonic acid
determination in serum and plasma. Journal of Chromatography B: Biomedical
Sciences and Applications 741(2): 231-241.
Lan, L. 1989. The embryology of Garcinia mangostana L. (Clusiaceae). Gard Bull
Singap 37: 97-103.

128
Lancaster, J. & Dougall, D. K. 1992. Regulation of skin color in apples. Critical
Reviews in Plant Sciences 10(6): 487-502.
Lattanzio, V., Kroon, P. A., Quideau, S. & Treutter, D. 2008. Plant phenolics-secondary
metabolites with diverse functions. Dlm. Daayf, F. & Lattanzio, V. (pnyt.).
Recent Advances in Polyphenol Research, 1. hlm. 1-35. United Kingdom: John
Wiley & Sons.
Lee, D.-K., Yoon, M. H., Kang, Y. P., Yu, J., Park, J. H., Lee, J. & Kwon, S. W. 2013.
Comparison of primary and secondary metabolites for suitability to discriminate
the origins of Schisandra chinensis by GC/MS and LC/MS. Food Chemistry
141(4): 3931-3937.
Lee, S., Choi, H.-K., Cho, S. K. & Kim, Y.-S. 2010. Metabolic analysis of guava
(Psidium guajava L.) fruits at different ripening stages using different data-
processing approaches. Journal of Chromatography B 878(29): 2983-2988.
Lei, Z., Li, H., Chang, J., Zhao, P. X. & Sumner, L. W. 2012. MET-IDEA version 2.06;
improved efficiency and additional functions for mass spectrometry-based
metabolomics data processing. Metabolomics 8(1): 105-110.
Lembaga Pemasaran Pertanian Persekutuan. 2017. Statistik utama pemasaran FAMA
2017.
Lerslerwong, L., Rugkong, A., Imsabai, W. & Ketsa, S. 2013. The harvest period of
mangosteen fruit can be extended by chemical control of ripening-a proof of
concept study. Scientia Horticulturae 157: 13-18.
Leuschner, C., Herrmann, K. M. & Schultz, G. 1995. The metabolism of quinate in pea
roots (purification and partial characterization of a quinate hydrolyase). Plant
Physiology 108(1): 319-325.
Li, L., Guo, M., Wang, X., Zhang, X. & Liu, T. 2017. Effects and mechanism of 1‐
methylcyclopropene and ethephon on softening in Ailsa Craig tomato fruit.
Journal of Food Processing and Preservation 41(3): e12883.
Li, X., Xu, Z., Lu, X., Yang, X., Yin, P., Kong, H., Yu, Y. & Xu, G. 2009.
Comprehensive two-dimensional gas chromatography/time-of-flight mass
spectrometry for metabonomics: biomarker discovery for diabetes mellitus.
Analytica Chimica Acta 633(2): 257-262.
Lieberman, M. 1979. Biosynthesis and action of ethylene. Annual Review of Plant
Physiology 30(1): 533-591.
Lisec, J., Schauer, N., Kopka, J., Willmitzer, L. & Fernie, A. R. 2006. Gas
chromatography mass spectrometry-based metabolite profiling in plants. Nature
Protocols 1(1): 387.

129
Little, J. L. 1999. Artifacts in trimethylsilyl derivatization reactions and ways to avoid
them. Journal of Chromatography A 844(1-2): 1-22.
Liu, Q.-Y., Wang, Y.-T. & Lin, L.-G. 2015. New insights into the anti-obesity activity
of xanthones from Garcinia mangostana. Food & Function 6(2): 383-393.
Lohani, S., Trivedi, P. K. & Nath, P. 2004. Changes in activities of cell wall hydrolases
during ethylene-induced ripening in banana: effect of 1-MCP, ABA and IAA.
Postharvest Biology and Technology 31(2): 119-126.
Lombardo, V. A., Osorio, S., Borsani, J., Lauxmann, M. A., Bustamante, C. A., Budde,
C. O., Andreo, C. S., Lara, M. V., Fernie, A. R. & Drincovich, M. F. 2011.
Metabolic profiling during peach fruit development and ripening reveals the
metabolic networks which underpin each developmental stage. Plant
Physiology 157: 1696-1710.
Mahmood, T., Anwar, F., Abbas, M., Boyce, M. C. & Saari, N. 2012. Compositional
variation in sugars and organic acids at different maturity stages in selected
small fruits from Pakistan. International Journal of Molecular Sciences 13(2):
1380-1392.
Marco-Ramell, A., Palau-Rodriguez, M., Alay, A., Tulipani, S., Urpi-Sarda, M.,
Sanchez-Pla, A. & Andres-Lacueva, C. 2018. Evaluation and comparison of
bioinformatic tools for the enrichment analysis of metabolomics data. BMC
Bioinformatics 19(1): 1.
Matheus, N., Hansen, S., Rozet, E., Peixoto, P., Maquoi, E., Lambert, V., Noel, A.,
Frederich, M., Mottet, D. & Tullio, P. 2014. An easy, convenient cell and tissue
extraction protocol for nuclear magnetic resonance metabolomics.
Phytochemical Analysis 25(4): 342-349.
Matsumoto, K., Akao, Y., Kobayashi, E., Ohguchi, K., Ito, T., Tanaka, T., Iinuma, M.
& Nozawa, Y. 2003. Induction of apoptosis by xanthones from mangosteen in
human leukemia cell lines. Journal of Natural Products 66(8): 1124-1127.
Mazlan, O., Aizat, W. M., Baharum, S. N., Azizan, K. A. & Noor, N. M. 2018a. Mass
spectrometry dataset for LC-MS metabolomics analysis of Garcinia
mangostana L. seed development. Data in Brief 21: 548-551.
Mazlan, O., Aizat, W. M., Baharum, S. N., Azizan, K. A. & Noor, N. M. 2018b.
Metabolomics analysis of developing Garcinia mangostana seed reveals
modulated levels of sugars, organic acids and phenylpropanoid compounds.
Scientia Horticulturae 233: 323-330.
Mazlan, O., Aizat, W. M., Zuddin, N. S. A., Baharum, S. N. & Noor, N. M. 2019.
Metabolite profiling of mangosteen seed germination highlights metabolic
changes related to carbon utilization and seed protection. Scientia Horticulturae
243: 226-234.

130
Mcgarvey, D. J. & Croteau, R. 1995. Terpenoid metabolism. The Plant Cell 7(7): 1015.
Metz, T. O., Baker, E. S., Schymanski, E. L., Renslow, R. S., Thomas, D. G., Causon,
T. J., Webb, I. K., Hann, S., Smith, R. D. & Teeguarden, J. G. 2017. Integrating
ion mobility spectrometry into mass spectrometry-based exposome
measurements: what can it add and how far can it go? Bioanalysis 9(1): 81-98.
Miean, K. H. & Mohamed, S. 2001. Flavonoid (myricetin, quercetin, kaempferol,
luteolin, and apigenin) content of edible tropical plants. Journal of Agricultural
and Food Chemistry 49(6): 3106-3112.
Moco, S., Bino, R. J., Vorst, O., Verhoeven, H. A., De Groot, J., Van Beek, T. A.,
Vervoort, J. & De Vos, C. R. 2006. A liquid chromatography-mass
spectrometry-based metabolome database for tomato. Plant Physiology 141(4):
1205-1218.
Mohamed, G. A., Ibrahim, S. R., Shaaban, M. I. & Ross, S. A. 2014.
Mangostanaxanthones I and II, new xanthones from the pericarp of Garcinia
mangostana. Fitoterapia 98: 215-221.
Mohan, S., Abdelwahab, S. I. & Thangavel, N. 2018. Anti-inflammatory molecular
mechanism of action of α-mangostin, the major xanthone from the pericarp of
Garcinia mangostana; an in silico, in vitro and in vivo approach. Food &
Function 9: 3860-3871.
Mohnen, D. 2008. Pectin structure and biosynthesis. Current Opinion in Plant Biology
11(3): 266-277.
Moing, A., Aharoni, A., Biais, B., Rogachev, I., Meir, S., Brodsky, L., Allwood, J. W.,
Erban, A., Dunn, W. B. & Kay, L. 2011. Extensive metabolic cross‐talk in
melon fruit revealed by spatial and developmental combinatorial metabolomics.
New Phytologist 190(3): 683-696.
Msaada, K., Hosni, K., Taarit, M. B., Chahed, T., Kchouk, M. E. & Marzouk, B. 2007.
Changes on essential oil composition of coriander (Coriandrum sativum L.)
fruits during three stages of maturity. Food Chemistry 102(4): 1131-1134.
Mushtaq, M. Y., Choi, Y. H., Verpoorte, R. & Wilson, E. G. 2014. Extraction for
metabolomics: access to the metabolome. Phytochemical Analysis 25(4): 291-
306.
Mworia, E. G., Yoshikawa, T., Salikon, N., Oda, C., Asiche, W. O., Yokotani, N., Abe,
D., Ushijima, K., Nakano, R. & Kubo, Y. 2012. Low-temperature-modulated
fruit ripening is independent of ethylene in ‘Sanuki Gold’ kiwifruit. Journal of
Experimental Botany 63(2): 963-971.
Nabavi, S. M., Marchese, A., Izadi, M., Curti, V., Daglia, M. & Nabavi, S. F. 2015.
Plants belonging to the genus Thymus as antibacterial agents: From farm to
pharmacy. Food Chemistry 173: 339-347.

131
Naczk, M. & Shahidi, F. 2006. Phenolics in cereals, fruits and vegetables: Occurrence,
extraction and analysis. Journal of Pharmaceutical and Biomedical Analysis
41(5): 1523-1542.
Nakatani, K., Atsumi, M., Arakawa, T., Oosawa, K., Shimura, S., Nakahata, N. &
Ohizumi, Y. 2002. Inhibitions of histamine release and prostaglandin E2
synthesis by mangosteen, a Thai medicinal plant. Biological and
Pharmaceutical Bulletin 25(9): 1137-1141.
Negi, J., Bisht, V., Singh, P., Rawat, M. & Joshi, G. 2013. Naturally occurring
xanthones: chemistry and biology. Journal of Applied Chemistry 2013: 1-9.
Nguyen, L.-H. D., Venkatraman, G., Sim, K.-Y. & Harrison, L. J. 2005. Xanthones and
benzophenones from Garcinia griffithii and Garcinia mangostana.
Phytochemistry 66(14): 1718-1723.
Nicolas, P., Lecourieux, D., Kappel, C., Cluzet, S., Cramer, G., Delrot, S. &
Lecourieux, F. 2014. The basic leucine zipper transcription factor ABSCISIC
ACID RESPONSE ELEMENT-BINDING FACTOR2 is an important
transcriptional regulator of abscisic acid-dependent grape berry ripening
processes. Plant Physiology 164(1): 365-383.
Niessen, W. 1999. State-of-the-art in liquid chromatography–mass spectrometry.
Journal of Chromatography A 856(1-2): 179-197.
Nkang, A. 2002. Carbohydrate composition during seed development and germination
in two sub-tropical rainforest tree species (Erythrina caffra and Guilfoylia
monostylis). Journal of Plant Physiology 159(5): 473-483.
Noor, A., Fatima, I., Ahmad, I., Malik, A., Afza, N., Iqbal, L., Latif, M. & Khan, S. B.
2007. Leufolins A and B, potent butyrylcholinesterase-inhibiting flavonoid
glucosides from Leucas urticifolia. Molecules 12(7): 1447-1454.
Okazaki, K., Kimura, Y., Sugiyama, K., Kami, D., Nakamura, T. & Oka, N. 2016.
Discovering metabolic indices for early detection of squash (Cucurbita maxima)
storage quality using GC–MS-based metabolite profiling. Food Chemistry 196:
1150-1155.
Oms-Oliu, G., Hertog, M., Van De Poel, B., Ampofo-Asiama, J., Geeraerd, A. &
Nicolai, B. 2011. Metabolic characterization of tomato fruit during preharvest
development, ripening, and postharvest shelf-life. Postharvest Biology and
Technology 62(1): 7-16.
Orata, F. 2012. Derivatization reactions and reagents for gas chromatography analysis.
Masinde Muliro University of Science and Technology, Kenya: INTECH Open
Access Publisher Rijeka.

132
Osman, M. & Milan, A. R. 2006. Fruits for the future 9: Mangosteen-Garcinia
mangostana. Southampton Centre for Underutilised Crops, University of
Southampton, Southampton: Crops for the Future.
Osorio, S., Alba, R., Nikoloski, Z., Kochevenko, A., Fernie, A. R. & Giovannoni, J. J.
2012. Integrative comparative analyses of transcript and metabolite profiles
from pepper and tomato ripening and development stages uncovers species-
specific patterns of network regulatory behaviour. Plant Physiology 159: 1713-
1729.
Osorio, S. & Fernie, A. R. 2014. Fruit ripening: primary metabolism. Dlm. Nath, P.,
Bouzayen, M., Mattoo, A. & Pech, J. (pnyt.). Fruit Ripening: Physiology,
Signalling and Genomics, hlm. 15-27. Wallingford: CAB International.
Osorio, S., Scossa, F. & Fernie, A. 2013. Molecular regulation of fruit ripening.
Frontiers in Plant Science 4: 1-7.
Palapol, Y., Ketsa, S., Lin-Wang, K., Ferguson, I. B. & Allan, A. C. 2009a. A MYB
transcription factor regulates anthocyanin biosynthesis in mangosteen (Garcinia
mangostana L.) fruit during ripening. Planta 229(6): 1323-1334.
Palapol, Y., Ketsa, S., Stevenson, D., Cooney, J., Allan, A. & Ferguson, I. 2009b.
Colour development and quality of mangosteen (Garcinia mangostana L.) fruit
during ripening and after harvest. Postharvest Biology and Technology 51(3):
349-353.
Pan, Z., Li, Y., Deng, X. & Xiao, S. 2014. Non-targeted metabolomic analysis of orange
(Citrus sinensis [L.] Osbeck) wild type and bud mutant fruits by direct analysis
in real-time and HPLC-electrospray mass spectrometry. Metabolomics 10(3):
508-523.
Panche, A. N., Diwan, A. D. & Chandra, S. R. 2016. Flavonoids: an overview. Journal
of Nutritional Science 5(47): 1-15.
Parijadi, A. A., Putri, S. P., Ridwani, S., Dwivany, F. M. & Fukusaki, E. 2017.
Metabolic profiling of Garcinia mangostana (mangosteen) based on ripening
stages. Journal of Bioscience and Bioengineering 125(2): 238-244.
Parveen, M. & Khan, N. U.-D. 1988. Two xanthones from Garcinia mangostana.
Phytochemistry 27(11): 3694-3696.
Patti, G. J., Yanes, O. & Siuzdak, G. 2012. Innovation: Metabolomics: the apogee of
the omics trilogy. Nature Reviews Molecular Cell Biology 13(4): 263.
Paul, M. J. & Pellny, T. K. 2003. Carbon metabolite feedback regulation of leaf
photosynthesis and development. Journal of Experimental Botany 54(382): 539-
547.

133
Paul, V., Pandey, R. & Srivastava, G. C. 2012. The fading distinctions between classical
patterns of ripening in climacteric and non-climacteric fruit and the ubiquity of
ethylene-an overview. Journal of Food Science and Technology 49(1): 1-21.
Pedraza-Chaverri, J., Cárdenas-Rodríguez, N., Orozco-Ibarra, M. & Pérez-Rojas, J. M.
2008. Medicinal properties of mangosteen (Garcinia mangostana). Food and
Chemical Toxicology 46(10): 3227-3239.
Pedreschi, R., Muñoz, P., Robledo, P., Becerra, C., Defilippi, B. G., Van Eekelen, H.,
Mumm, R., Westra, E. & De Vos, R. C. 2014. Metabolomics analysis of
postharvest ripening heterogeneity of ‘Hass’ avocadoes. Postharvest Biology
and Technology 92: 172-179.
Pitt, J. J. 2009. Principles and applications of liquid chromatography-mass spectrometry
in clinical biochemistry. The Clinical Biochemist Reviews 30(1): 19.
Pluskal, T., Castillo, S., Villar-Briones, A. & Orešič, M. 2010. MZmine 2: modular
framework for processing, visualizing, and analyzing mass spectrometry-based
molecular profile data. BMC Bioinformatics 11(1): 395.
Poerwanto, R., Efendi, D. & Suhartanto, R. 2006. Improving productivity and quality
of Indonesian mangosteen. ISHS Acta Horticulturae 769: XXVII International
Horticultural Congress - IHC2006: International Symposium on Asian Plants
with Unique Horticultural Potential, hlm. 285-288.
Pothitirat, W., Chomnawang, M. T., Supabphol, R. & Gritsanapan, W. 2009.
Comparison of bioactive compounds content, free radical scavenging and anti-
acne inducing bacteria activities of extracts from the mangosteen fruit rind at
two stages of maturity. Fitoterapia 80(7): 442-447.
Prasanna, V., Prabha, T. & Tharanathan, R. 2007. Fruit ripening phenomena-an
overview. Critical Reviews in Food Science and Nutrition 47(1): 1-19.
Prior, R. L. & Gu, L. 2005. Occurrence and biological significance of
proanthocyanidins in the American diet. Phytochemistry 66(18): 2264-2280.
Qin, Y., Sun, Y., Li, J., Xie, R., Deng, Z., Chen, H. & Li, H. 2017. Characterization and
antioxidant activities of procyanidins from lotus seedpod, mangosteen pericarp,
and camellia flower. International Journal of Food Properties 20(7): 1621-
1632.
Rangarajan, R. & Ghosh, P. 2011. Role of water contamination within the GC column
of a GasBench II peripheral on the reproducibility of 18O/16O ratios in water
samples. Isotopes in Environmental and Health Studies 47(4): 498-511.
Ravanel, S., Gakière, B., Job, D. & Douce, R. 1998. The specific features of methionine
biosynthesis and metabolism in plants. Proceedings of the National Academy of
Sciences 95(13): 7805-7812.

134
Ren, S., Hinzman, A. A., Kang, E. L., Szczesniak, R. D. & Lu, L. J. 2015.
Computational and statistical analysis of metabolomics data. Metabolomics
11(6): 1492-1513.
Rhodes, M. J. & Price, K. R. 1996. Analytical problems in the study of flavonoid
compounds in onions. Food Chemistry 57(1): 113-117.
Roberts, L. D., Souza, A. L., Gerszten, R. E. & Clish, C. B. 2012. Targeted
metabolomics. Current Protocols in Molecular Biology 98: 1-24.
Robertson, D. G., Watkins, P. B. & Reily, M. D. 2011. Metabolomics in toxicology:
preclinical and clinical applications. Toxicological Sciences 120(suppl 1): S146-
S170.
Robinson, S. P. & Davies, C. 2000. Molecular biology of grape berry ripening.
Australian Journal of Grape and Wine Research 6(2): 175-188.
Rosli, M. a. F., Azizan, K. A., Baharum, S. N. & Goh, H.-H. 2017. Mass spectrometry
data of metabolomics analysis of Nepenthes pitchers. Data in Brief 14(295-297.
Rostagno, M. A., Palma, M. & Barroso, C. G. 2003. Ultrasound-assisted extraction of
soy isoflavones. Journal of Chromatography A 1012(2): 119-128.
Ryan, D. & Robards, K. 2006. Metabolomics: the greatest omics of them all? Analytical
Chemistry 78(23): 7954-7958.
Saladié, M., Cañizares, J., Phillips, M. A., Rodriguez-Concepcion, M., Larrigaudière,
C., Gibon, Y., Stitt, M., Lunn, J. E. & Garcia-Mas, J. 2015. Comparative
transcriptional profiling analysis of developing melon (Cucumis melo L.) fruit
from climacteric and non-climacteric varieties. BMC Genomics 16(1): 440.
Saladié, M., Matas, A. J., Isaacson, T., Jenks, M. A., Goodwin, S. M., Niklas, K. J.,
Xiaolin, R., Labavitch, J. M., Shackel, K. A. & Fernie, A. R. 2007. A
reevaluation of the key factors that influence tomato fruit softening and
integrity. Plant Physiology 144(2): 1012-1028.
Saleem, M. 2009. Lupeol, a novel anti-inflammatory and anti-cancer dietary triterpene.
Cancer Letters 285(2): 109-115.
Sandrang, A. K., Megat Amaddin, P. A., Idris, S., Mahmood, Z. & Hasan, M. R. H.
2015. Pembiakan mesta berskala besar dengan kaedah keratan mikro. Buletin
Teknologi MARDI 63-69.
Schmid, W. 1855. Ueber das mangostin. Justus Liebigs Annalen der Chemie 93(1): 83-
88.
Seo, E.-K., Kim, N.-C., Wani, M. C., Wall, M. E., Navarro, H. A., Burgess, J. P.,
Kawanishi, K., Kardono, L. B., Riswan, S. & Rose, W. C. 2002. Cytotoxic

135
prenylated xanthones and the unusual compounds anthraquinobenzophenones
from Cratoxylum sumatranum. Journal of Natural Products 65(3): 299-305.
Shackel, K. A., Greve, C., Labavitch, J. M. & Ahmadi, H. 1991. Cell turgor changes
associated with ripening in tomato pericarp tissue. Plant Physiology 97(2): 814-
816.
Shan, T., Ma, Q., Guo, K., Liu, J., Li, W., Wang, F. & Wu, E. 2011. Xanthones from
mangosteen extracts as natural chemopreventive agents: potential anticancer
drugs. Current Molecular Medicine 11(8): 666-677.
Sharon‐Asa, L., Shalit, M., Frydman, A., Bar, E., Holland, D., Or, E., Lavi, U.,
Lewinsohn, E. & Eyal, Y. 2003. Citrus fruit flavor and aroma biosynthesis:
isolation, functional characterization, and developmental regulation of Cstps1,
a key gene in the production of the sesquiterpene aroma compound valencene.
The Plant Journal 36(5): 664-674.
Shi, J., Wang, H., Hazebroek, J., Ertl, D. S. & Harp, T. 2005. The maize low-phytic
acid 3 encodes a myo-inositol kinase that plays a role in phytic acid biosynthesis
in developing seeds. The Plant Journal 42(5): 708-719.
Shibata, M.-A., Iinuma, M., Morimoto, J., Kurose, H., Akamatsu, K., Okuno, Y., Akao,
Y. & Otsuki, Y. 2011. α-Mangostin extracted from the pericarp of the
mangosteen (Garcinia mangostana Linn.) reduces tumor growth and lymph
node metastasis in an immunocompetent xenograft model of metastatic
mammary cancer carrying a p53 mutation. BMC Medicine 9(1): 69.
Shulaev, V. 2006. Metabolomics technology and bioinformatics. Briefings in
Bioinformatics 7(2): 128-139.
Sinaga, R. N. & Siregar, N. S. 2016. Phytochemical screening and test of antioxidant
activity in the extract of mangosteen rind. International Conference on Public
Health 2016, hlm. 124-124.
Smith, C. A., Want, E. J., O'maille, G., Abagyan, R. & Siuzdak, G. 2006. XCMS:
processing mass spectrometry data for metabolite profiling using nonlinear peak
alignment, matching, and identification. Analytical Chemistry 78(3): 779-787.
Soga, T., Ohashi, Y., Ueno, Y., Naraoka, H., Tomita, M. & Nishioka, T. 2003.
Quantitative metabolome analysis using capillary electrophoresis mass
spectrometry. Journal of Proteome Research 2(5): 488-494.
Son, H.-S., Hwang, G.-S., Kim, K. M., Ahn, H.-J., Park, W.-M., Van Den Berg, F.,
Hong, Y.-S. & Lee, C.-H. 2009. Metabolomic studies on geographical grapes
and their wines using 1H NMR analysis coupled with multivariate statistics.
Journal of Agricultural and Food Chemistry 57(4): 1481-1490.

136
Sugita, P., Arya, S., Ilmiawati, A. & Arifin, B. 2017. Characterization, antibacterial and
antioxidant activity of mangosteen (Garcinia mangostana L.) pericarp
nanosized extract. Rasayan Journal of Chemistry 10(3): 707-715.
Sukatta, U., Takenaka, M., Ono, H., Okadome, H., Sotome, I., Nanayama, K.,
Thanapase, W. & Isobe, S. 2013. Distribution of major xanthones in the
pericarp, aril, and yellow gum of mangosteen (Garcinia mangostana Linn.) fruit
and their contribution to antioxidative activity. Bioscience, Biotechnology, and
Biochemistry 77(5): 984-987.
Suksamrarn, S., Suwannapoch, N., Phakhodee, W., Thanuhiranlert, J., Ratananukul, P.,
Chimnoi, N. & Suksamrarn, A. 2003. Antimycobacterial activity of prenylated
xanthones from the fruits of Garcinia mangostana. Chemical and
Pharmaceutical Bulletin 51(7): 857-859.
Sultana, B. & Anwar, F. 2008. Flavonols (kaempeferol, quercetin, myricetin) contents
of selected fruits, vegetables and medicinal plants. Food Chemistry 108(3): 879-
884.
Sumner, L. W., Mendes, P. & Dixon, R. A. 2003. Plant metabolomics: large-scale
phytochemistry in the functional genomics era. Phytochemistry 62(6): 817-836.
T’kindt, R., Morreel, K., Deforce, D., Boerjan, W. & Van Bocxlaer, J. 2009. Joint GC–
MS and LC–MS platforms for comprehensive plant metabolomics:
Repeatability and sample pre-treatment. Journal of Chromatography B 877(29):
3572-3580.
Tautenhahn, R., Cho, K., Uritboonthai, W., Zhu, Z., Patti, G. J. & Siuzdak, G. 2012. An
accelerated workflow for untargeted metabolomics using the METLIN
database. Nature Biotechnology 30(9): 826.
Theodoridis, G., Gika, H., Franceschi, P., Caputi, L., Arapitsas, P., Scholz, M.,
Masuero, D., Wehrens, R., Vrhovsek, U. & Mattivi, F. 2012a. LC-MS based
global metabolite profiling of grapes: solvent extraction protocol optimisation.
Metabolomics 8(2): 175-185.
Theodoridis, G. A., Gika, H. G., Want, E. J. & Wilson, I. D. 2012b. Liquid
chromatography–mass spectrometry based global metabolite profiling: a
review. Analytica Chimica Acta 711: 7-16.
Tholl, D. 2015. Biosynthesis and biological functions of terpenoids in plants. Dlm.
Schrader, J. & Bohlmann, J. (pnyt.). Biotechnology of Isoprenoids. Advances
in Biochemical Engineering/Biotechnology, 148. hlm. 63-106. Springer.
Thong, N. M., Quang, D. T., Bui, N. H. T., Dao, D. Q. & Nam, P. C. 2015. Antioxidant
properties of xanthones extracted from the pericarp of Garcinia mangostana
(mangosteen): A theoretical study. Chemical Physics Letters 625: 30-35.

137
Tjitrosemito, S. & Poerwanto, R. 2008. secretory duct structure and phytochemistry
compounds of yellow latex in mangosteen fruit. HAYATI Journal of Biosciences
15(3): 99-104.
Tohge, T., Alseekh, S. & Fernie, A. R. 2014. On the regulation and function of
secondary metabolism during fruit development and ripening. Journal of
Experimental Botany 65(16): 4599-4611.
Valluru, R. & Van Den Ende, W. 2011. Myo-inositol and beyond-emerging networks
under stress. Plant Science 181(4): 387-400.
Van Dongen, J. T., Gupta, K. J., Ramírez-Aguilar, S. J., Araújo, W. L., Nunes-Nesi, A.
& Fernie, A. R. 2011. Regulation of respiration in plants: a role for alternative
metabolic pathways. Journal of Plant Physiology 168(12): 1434-1443.
Vieira, L. & Kijjoa, A. 2005. Naturally-occurring xanthones: recent developments.
Current Medicinal Chemistry 12(21): 2413-2446.
Vinaixa, M., Samino, S., Saez, I., Duran, J., Guinovart, J. J. & Yanes, O. 2012. A
guideline to univariate statistical analysis for LC/MS-based untargeted
metabolomics-derived data. Metabolites 2(4): 775-795.
Vinatoru, M. 2001. An overview of the ultrasonically assisted extraction of bioactive
principles from herbs. Ultrasonics Sonochemistry 8(3): 303-313.
Vondras, A. M., Commisso, M., Guzzo, F. & Deluc, L. G. 2017. Metabolite profiling
reveals developmental inequalities in pinot noir berry tissues late in ripening.
Frontiers in Plant Science 8(1108).
Wang, J., Sun, L., Xie, L., He, Y., Luo, T., Sheng, L., Luo, Y., Zeng, Y., Xu, J. & Deng,
X. 2016. Regulation of cuticle formation during fruit development and ripening
in ‘Newhall’navel orange (Citrus sinensis Osbeck) revealed by transcriptomic
and metabolomic profiling. Plant Science 243: 131-144.
Wansi, J. D., Devkota, K. P., Tshikalange, E. & Kuete, V. 2013. Alkaloids from the
medicinal plants of Africa. Dlm. Kuete, V. (pnyt.). Medicinal Plant Research
in Africa, hlm. 557-605. Oxford: Elsevier.
Watanapokasin, R., Jarinthanan, F., Jerusalmi, A., Suksamrarn, S., Nakamura, Y.,
Sukseree, S., Uthaisang-Tanethpongtamb, W., Ratananukul, P. & Sano, T.
2010. Potential of xanthones from tropical fruit mangosteen as anti-cancer
agents: caspase-dependent apoptosis induction in vitro and in mice. Applied
Biochemistry and Biotechnology 162(4): 1080-1094.
Watkins, C. B. 2008. Overview of 1-methylcyclopropene trials and uses for edible
horticultural crops. HortScience 43(1): 86-94.

138
Wei, R., Wang, J., Su, M., Jia, E., Chen, S., Chen, T. & Ni, Y. 2018. Missing value
imputation approach for mass spectrometry-based metabolomics data. Scientific
Reports 8(1): 663.
Wink, M. & Schimmer, O. 2018. Modes of action of defensive secondary metabolites.
Annual Plant Reviews 18-137.
Wishart, D. S., Jewison, T., Guo, A. C., Wilson, M., Knox, C., Liu, Y., Djoumbou, Y.,
Mandal, R., Aziat, F. & Dong, E. 2012. HMDB 3.0-the human metabolome
database in 2013. Nucleic Acids Research 41(D1): D801-D807.
Wittenauer, J., Falk, S., Schweiggert-Weisz, U. & Carle, R. 2012. Characterisation and
quantification of xanthones from the aril and pericarp of mangosteens (Garcinia
mangostana L.) and a mangosteen containing functional beverage by HPLC–
DAD–MSn. Food Chemistry 134(1): 445-452.
Xia, J., Sinelnikov, I. V., Han, B. & Wishart, D. S. 2015a. MetaboAnalyst 3.0-making
metabolomics more meaningful. Nucleic Acids Research 43(W1): W251-W257.
Xia, J., Sinelnikov, I. V., Han, B. & Wishart, D. S. 2015b. MetaboAnalyst 3.0-making
metabolomics more meaningful. Nucleic Acids Research 43(W1): W251-W257.
Yaacob, O. & Tindall, H. D. 1995. Mangosteen cultivation. Food & Agriculture Org.
Yamashita, I., Nemoto, Y. & Yoshikawa, S. 1976. Formation of volatile alcohols and
esters from aldehydes in strawberries. Phytochemistry 15(11): 1633-1637.
Yang, R., Li, P., Li, N., Zhang, Q., Bai, X., Wang, L., Xiao, Y., Sun, L., Yang, Q. &
Yan, J. 2017. Xanthones from the pericarp of Garcinia mangostana. Molecules
22(5): 683.
Yang, S. F. & Hoffman, N. E. 1984. Ethylene biosynthesis and its regulation in higher
plants. Annual Review of Plant Physiology 35(1): 155-189.
Yang, T., Stoopen, G., Yalpani, N., Vervoort, J., De Vos, R., Voster, A., Verstappen,
F. W., Bouwmeester, H. J. & Jongsma, M. A. 2011. Metabolic engineering of
geranic acid in maize to achieve fungal resistance is compromised by novel
glycosylation patterns. Metabolic Engineering 13(4): 414-425.
Yang, X., Song, J., Du, L., Forney, C., Campbell-Palmer, L., Fillmore, S., Wismer, P.
& Zhang, Z. 2016. Ethylene and 1-MCP regulate major volatile biosynthetic
pathways in apple fruit. Food Chemistry 194: 325-336.
Yi, L., Dong, N., Yun, Y., Deng, B., Ren, D., Liu, S. & Liang, Y. 2016. Chemometric
methods in data processing of mass spectrometry-based metabolomics: a
review. Analytica Chimica Acta 914: 17-34.

139
Yildizli, A., Çevik, S. & Ünyayar, S. 2018. Effects of exogenous myo-inositol on leaf
water status and oxidative stress of Capsicum annuum under drought stress. Acta
Physiologiae Plantarum 40(6): 122.
Yoshimura, M., Ninomiya, K., Tagashira, Y., Maejima, K., Yoshida, T. & Amakura,
Y. 2015. Polyphenolic constituents of the pericarp of mangosteen (Garcinia
mangostana L.). Journal of Agricultural and Food Chemistry 63(35): 7670-
7674.
Zadernowski, R., Czaplicki, S. & Naczk, M. 2009. Phenolic acid profiles of mangosteen
fruits (Garcinia mangostana). Food Chemistry 112(3): 685-689.
Zarena, A., Sachindra, N. & Sankar, K. U. 2012. Optimisation of ethanol modified
supercritical carbon dioxide on the extract yield and antioxidant activity from
Garcinia mangostana L. Food Chemistry 130(1): 203-208.
Zarena, A. & Sankar, K. U. 2009. Supercritical carbon dioxide extraction of xanthones
with antioxidant activity from Garcinia mangostana: Characterization by
HPLC/LC–ESI-MS. The Journal of Supercritical Fluids 49(3): 330-337.
Zarena, A. & Sankar, K. U. 2012. Phenolic acids, flavonoid profile and antioxidant
activity in mangosteen (Garcinia Mangostana L.) pericarp. Journal of Food
Biochemistry 36(5): 627-633.
Zhang, A., Sun, H., Yan, G., Wang, P. & Wang, X. 2016. Mass spectrometry‐based
metabolomics: applications to biomarker and metabolic pathway research.
Biomedical Chromatography 30(1): 7-12.
Zhou, Y., Han, Q.-B., Song, J.-Z., Qiao, C.-F. & Xu, H.-X. 2008. Characterization of
polyprenylated xanthones in Garcinia xipshuanbannaensis using liquid
chromatography coupled with electrospray ionization quadrupole time-of-flight
tandem mass spectrometry. Journal of Chromatography A 1206(2): 131-139.
Zierer, J., Menni, C., Kastenmüller, G. & Spector, T. D. 2015. Integration of ‘omics’
data in aging research: from biomarkers to systems biology. Aging Cell 14(6):
933-944.
Zulak, K. G., Liscombe, D. K., Ashihara, H. & Facchini, P. J. 2006. Alkaloids. Dlm.
Crozier, A. C., Clifford, M. N. & Ashihara, H. (pnyt.). Plant Secondary
Metabolites: Occurrence, Structure and Role in the Human Diet, hlm. 102-136.
Oxford: Blackwell Publishing Ltd.
Zwenger, S. & Basu, C. 2008. Plant terpenoids: applications and future potentials.
Biotechnology and Molecular Biology Reviews 3(1): 1-7.
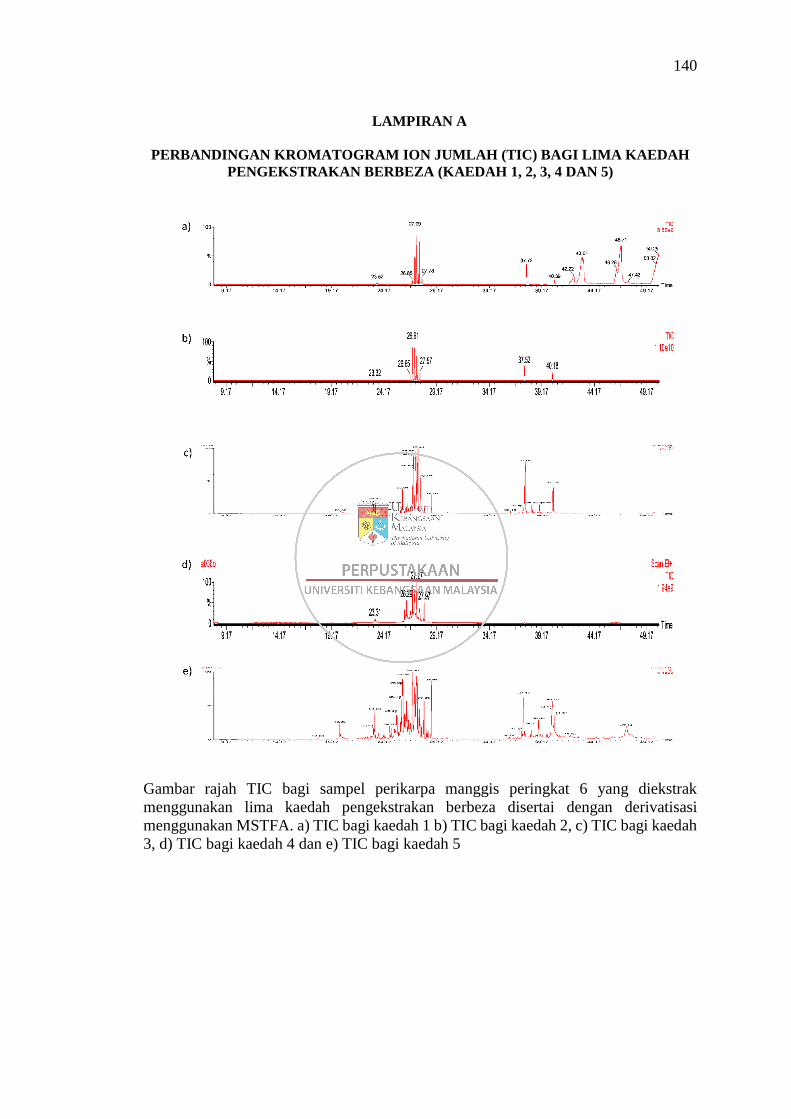
140
LAMPIRAN A
PERBANDINGAN KROMATOGRAM ION JUMLAH (TIC) BAGI LIMA KAEDAH
PENGEKSTRAKAN BERBEZA (KAEDAH 1, 2, 3, 4 DAN 5)
Gambar rajah TIC bagi sampel perikarpa manggis peringkat 6 yang diekstrak
menggunakan lima kaedah pengekstrakan berbeza disertai dengan derivatisasi
menggunakan MSTFA. a) TIC bagi kaedah 1 b) TIC bagi kaedah 2, c) TIC bagi kaedah
3, d) TIC bagi kaedah 4 dan e) TIC bagi kaedah 5
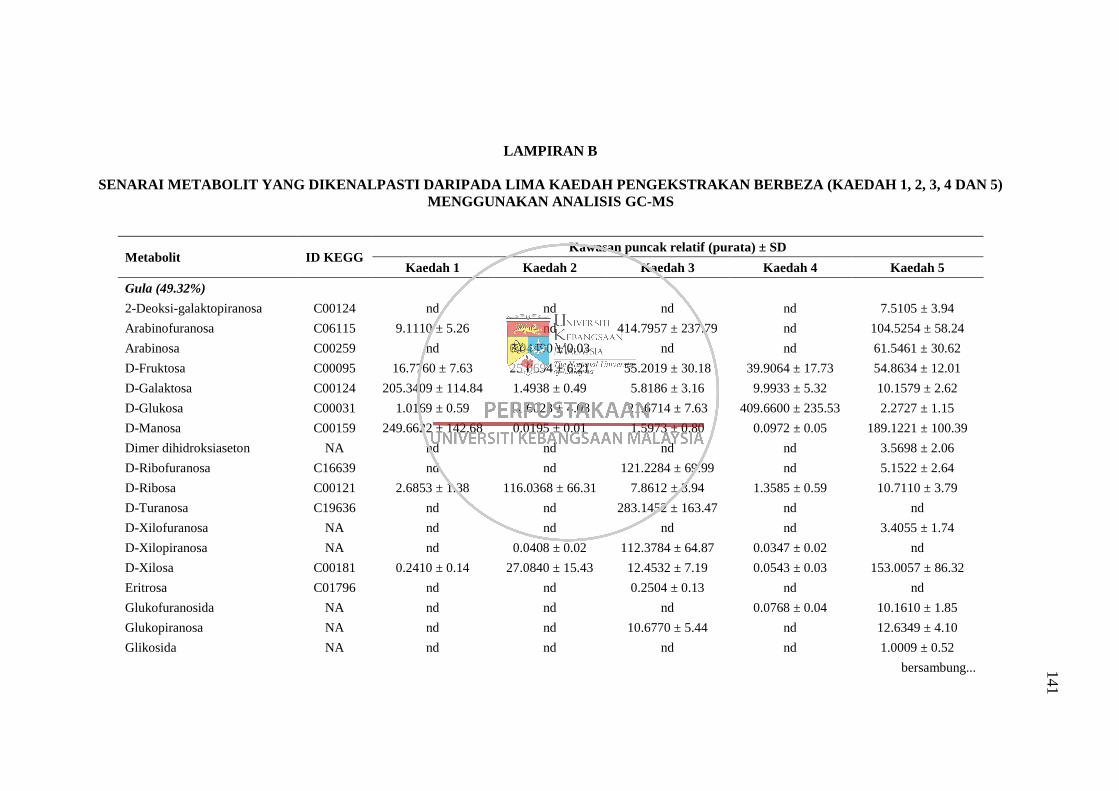
141
LAMPIRAN B
SENARAI METABOLIT YANG DIKENALPASTI DARIPADA LIMA KAEDAH PENGEKSTRAKAN BERBEZA (KAEDAH 1, 2, 3, 4 DAN 5)
MENGGUNAKAN ANALISIS GC-MS
Metabolit ID KEGG Kawasan puncak relatif (purata) ± SD
Kaedah 1 Kaedah 2 Kaedah 3 Kaedah 4 Kaedah 5
Gula (49.32%)
2-Deoksi-galaktopiranosa C00124 nd nd nd nd 7.5105 ± 3.94
Arabinofuranosa C06115 9.1110 ± 5.26 nd 414.7957 ± 237.79 nd 104.5254 ± 58.24
Arabinosa C00259 nd 0.04490 ± 0.03 nd nd 61.5461 ± 30.62
D-Fruktosa C00095 16.7760 ± 7.63 25.0694 ± 6.21 55.2019 ± 30.18 39.9064 ± 17.73 54.8634 ± 12.01
D-Galaktosa C00124 205.3409 ± 114.84 1.4938 ± 0.49 5.8186 ± 3.16 9.9933 ± 5.32 10.1579 ± 2.62
D-Glukosa C00031 1.0169 ± 0.59 11.6023 ± 4.08 21.6714 ± 7.63 409.6600 ± 235.53 2.2727 ± 1.15
D-Manosa C00159 249.6622 ± 142.68 0.0195 ± 0.01 1.5973 ± 0.80 0.0972 ± 0.05 189.1221 ± 100.39
Dimer dihidroksiaseton NA nd nd nd nd 3.5698 ± 2.06
D-Ribofuranosa C16639 nd nd 121.2284 ± 69.99 nd 5.1522 ± 2.64
D-Ribosa C00121 2.6853 ± 1.38 116.0368 ± 66.31 7.8612 ± 3.94 1.3585 ± 0.59 10.7110 ± 3.79
D-Turanosa C19636 nd nd 283.1452 ± 163.47 nd nd
D-Xilofuranosa NA nd nd nd nd 3.4055 ± 1.74
D-Xilopiranosa NA nd 0.0408 ± 0.02 112.3784 ± 64.87 0.0347 ± 0.02 nd
D-Xilosa C00181 0.2410 ± 0.14 27.0840 ± 15.43 12.4532 ± 7.19 0.0543 ± 0.03 153.0057 ± 86.32
Eritrosa C01796 nd nd 0.2504 ± 0.13 nd nd
Glukofuranosida NA nd nd nd 0.0768 ± 0.04 10.1610 ± 1.85
Glukopiranosa NA nd nd 10.6770 ± 5.44 nd 12.6349 ± 4.10
Glikosida NA nd nd nd nd 1.0009 ± 0.52
bersambung...
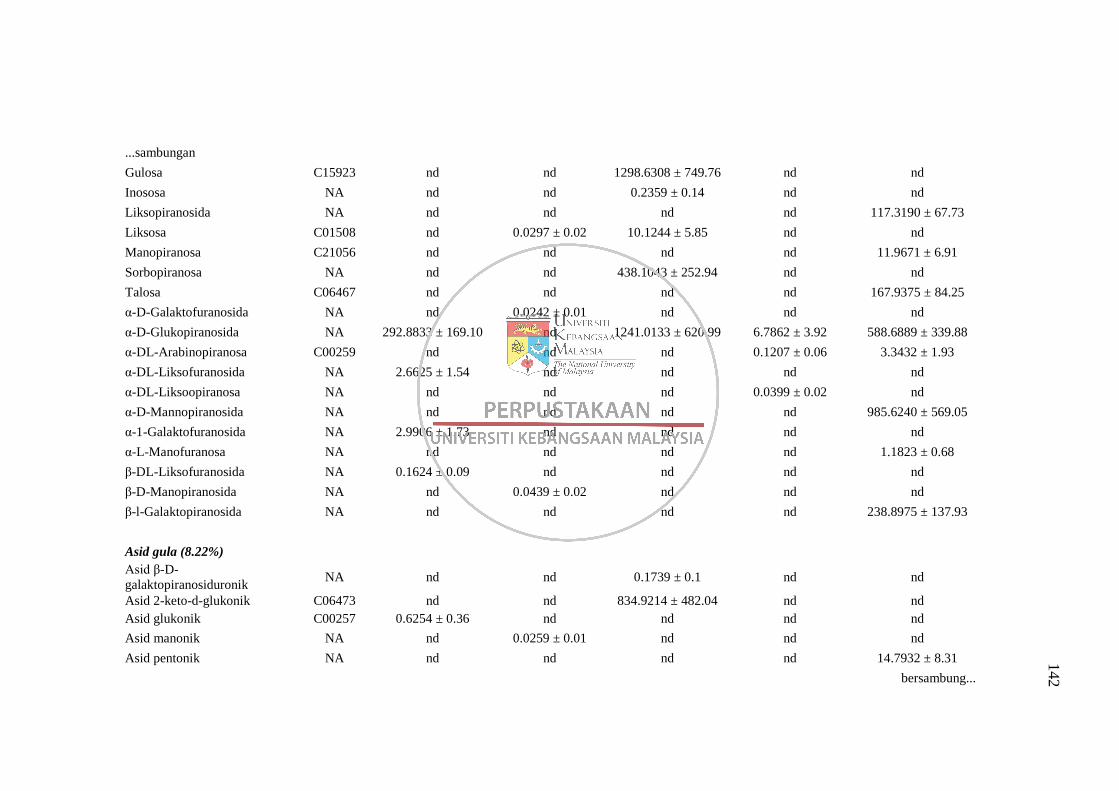
142
...sambungan
Gulosa C15923 nd nd 1298.6308 ± 749.76 nd nd
Inososa NA nd nd 0.2359 ± 0.14 nd nd
Liksopiranosida NA nd nd nd nd 117.3190 ± 67.73
Liksosa C01508 nd 0.0297 ± 0.02 10.1244 ± 5.85 nd nd
Manopiranosa C21056 nd nd nd nd 11.9671 ± 6.91
Sorbopiranosa NA nd nd 438.1043 ± 252.94 nd nd
Talosa C06467 nd nd nd nd 167.9375 ± 84.25
α-D-Galaktofuranosida NA nd 0.0242 ± 0.01 nd nd nd
α-D-Glukopiranosida NA 292.8833 ± 169.10 nd 1241.0133 ± 620.99 6.7862 ± 3.92 588.6889 ± 339.88
α-DL-Arabinopiranosa C00259 nd nd nd 0.1207 ± 0.06 3.3432 ± 1.93
α-DL-Liksofuranosida NA 2.6625 ± 1.54 nd nd nd nd
α-DL-Liksoopiranosa NA nd nd nd 0.0399 ± 0.02 nd
α-D-Mannopiranosida NA nd nd nd nd 985.6240 ± 569.05
α-1-Galaktofuranosida NA 2.9906 ± 1.73 nd nd nd nd
α-L-Manofuranosa NA nd nd nd nd 1.1823 ± 0.68
β-DL-Liksofuranosida NA 0.1624 ± 0.09 nd nd nd nd
β-D-Manopiranosida NA nd 0.0439 ± 0.02 nd nd nd
β-l-Galaktopiranosida NA nd nd nd nd 238.8975 ± 137.93
Asid gula (8.22%)
Asid β-D-
galaktopiranosiduronik NA nd nd 0.1739 ± 0.1 nd nd
Asid 2-keto-d-glukonik C06473 nd nd 834.9214 ± 482.04 nd nd
Asid glukonik C00257 0.6254 ± 0.36 nd nd nd nd
Asid manonik NA nd 0.0259 ± 0.01 nd nd nd
Asid pentonik NA nd nd nd nd 14.7932 ± 8.31
bersambung...
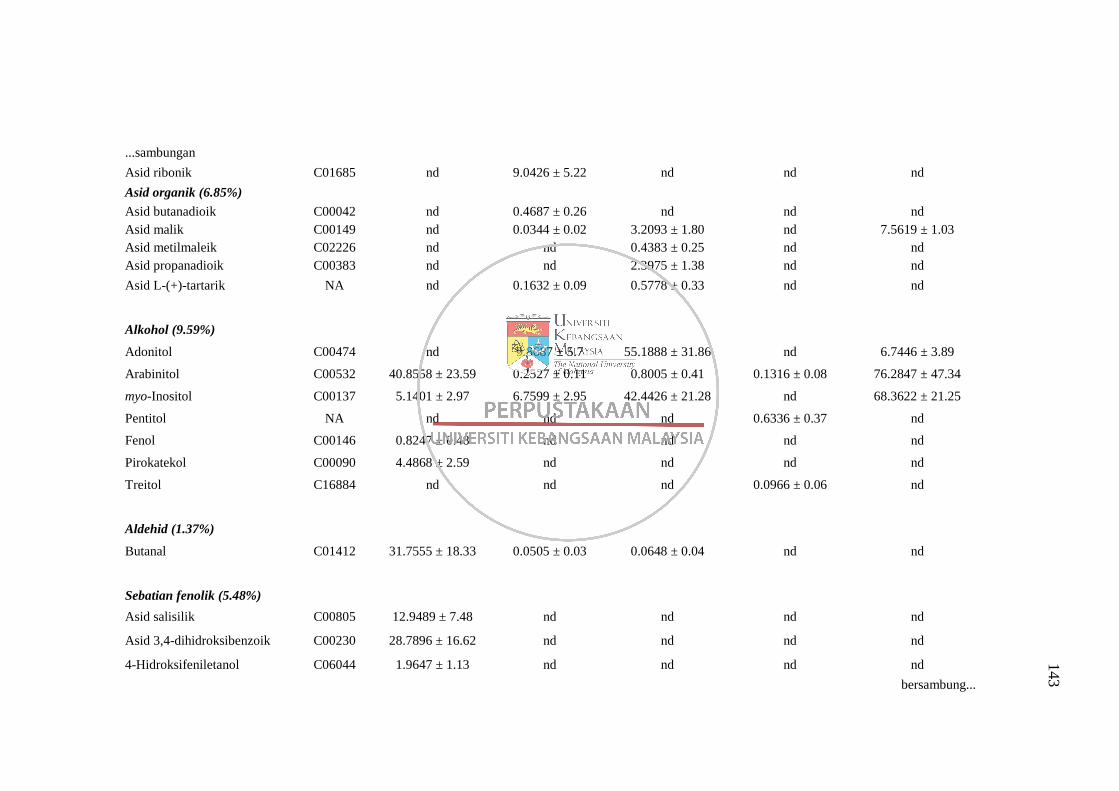
143
...sambungan
Asid ribonik C01685 nd 9.0426 ± 5.22 nd nd nd
Asid organik (6.85%)
Asid butanadioik C00042 nd 0.4687 ± 0.26 nd nd nd
Asid malik C00149 nd 0.0344 ± 0.02 3.2093 ± 1.80 nd 7.5619 ± 1.03
Asid metilmaleik C02226 nd nd 0.4383 ± 0.25 nd nd
Asid propanadioik C00383 nd nd 2.3975 ± 1.38 nd nd
Asid L-(+)-tartarik NA nd 0.1632 ± 0.09 0.5778 ± 0.33 nd nd
Alkohol (9.59%)
Adonitol C00474 nd 9.8687 ± 5.7 55.1888 ± 31.86 nd 6.7446 ± 3.89
Arabinitol C00532 40.8558 ± 23.59 0.2527 ± 0.11 0.8005 ± 0.41 0.1316 ± 0.08 76.2847 ± 47.34
myo-Inositol C00137 5.1401 ± 2.97 6.7599 ± 2.95 42.4426 ± 21.28 nd 68.3622 ± 21.25
Pentitol NA nd nd nd 0.6336 ± 0.37 nd
Fenol C00146 0.8247 ± 0.48 nd nd nd nd
Pirokatekol C00090 4.4868 ± 2.59 nd nd nd nd
Treitol C16884 nd nd nd 0.0966 ± 0.06 nd
Aldehid (1.37%)
Butanal C01412 31.7555 ± 18.33 0.0505 ± 0.03 0.0648 ± 0.04 nd nd
Sebatian fenolik (5.48%)
Asid salisilik C00805 12.9489 ± 7.48 nd nd nd nd
Asid 3,4-dihidroksibenzoik C00230 28.7896 ± 16.62 nd nd nd nd
4-Hidroksifeniletanol C06044 1.9647 ± 1.13 nd nd nd nd
bersambung...
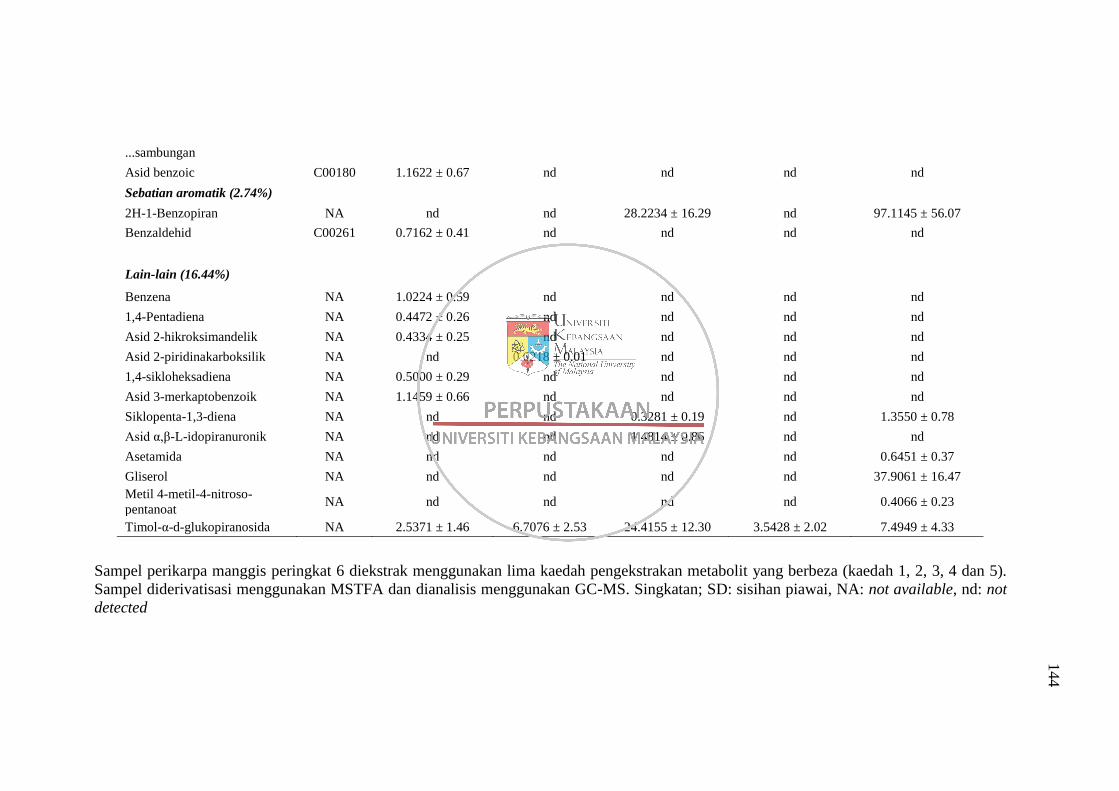
144
...sambungan
Asid benzoic C00180 1.1622 ± 0.67 nd nd nd nd
Sebatian aromatik (2.74%)
2H-1-Benzopiran NA nd nd 28.2234 ± 16.29 nd 97.1145 ± 56.07
Benzaldehid C00261 0.7162 ± 0.41 nd nd nd nd
Lain-lain (16.44%)
Benzena NA 1.0224 ± 0.59 nd nd nd nd
1,4-Pentadiena NA 0.4472 ± 0.26 nd nd nd nd
Asid 2-hikroksimandelik NA 0.4334 ± 0.25 nd nd nd nd
Asid 2-piridinakarboksilik NA nd 0.0218 ± 0.01 nd nd nd
1,4-sikloheksadiena NA 0.5000 ± 0.29 nd nd nd nd
Asid 3-merkaptobenzoik NA 1.1459 ± 0.66 nd nd nd nd
Siklopenta-1,3-diena NA nd nd 0.3281 ± 0.19 nd 1.3550 ± 0.78
Asid α,β-L-idopiranuronik NA nd nd 1.4814 ± 0.86 nd nd
Asetamida NA nd nd nd nd 0.6451 ± 0.37
Gliserol NA nd nd nd nd 37.9061 ± 16.47
Metil 4-metil-4-nitroso-
pentanoat NA nd nd nd nd 0.4066 ± 0.23
Timol-α-d-glukopiranosida NA 2.5371 ± 1.46 6.7076 ± 2.53 24.4155 ± 12.30 3.5428 ± 2.02 7.4949 ± 4.33
Sampel perikarpa manggis peringkat 6 diekstrak menggunakan lima kaedah pengekstrakan metabolit yang berbeza (kaedah 1, 2, 3, 4 dan 5).
Sampel diderivatisasi menggunakan MSTFA dan dianalisis menggunakan GC-MS. Singkatan; SD: sisihan piawai, NA: not available, nd: not
detected
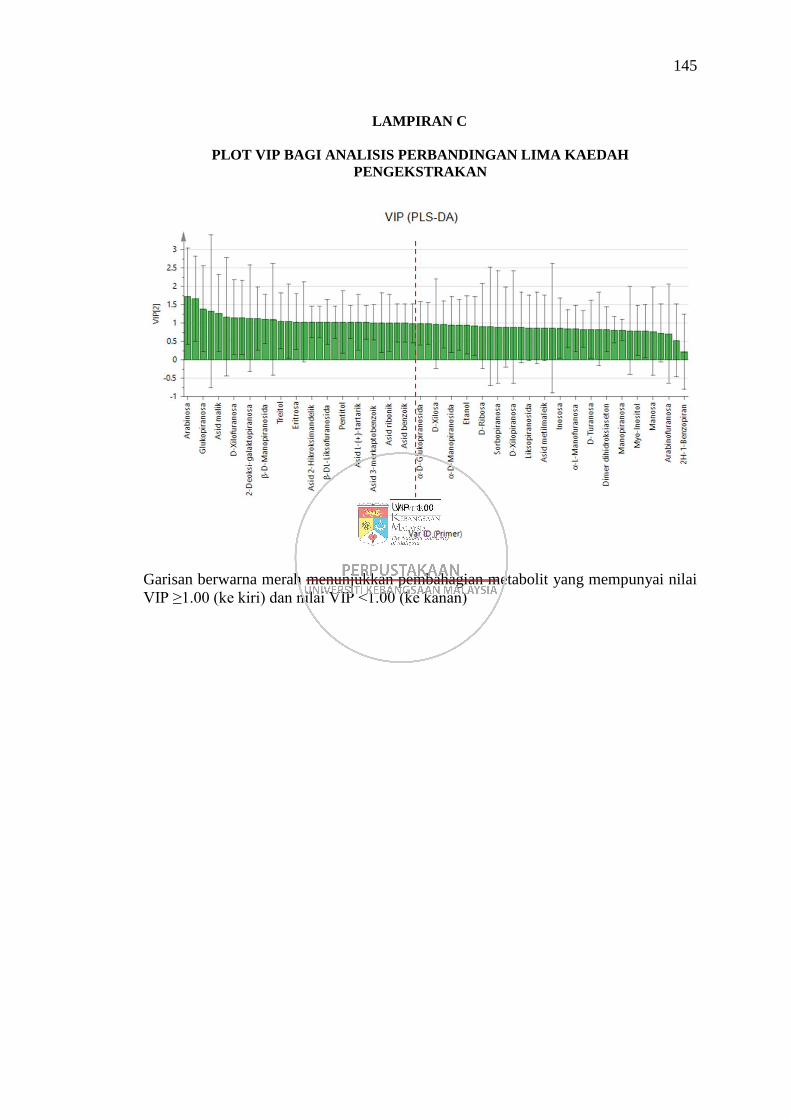
145
LAMPIRAN C
PLOT VIP BAGI ANALISIS PERBANDINGAN LIMA KAEDAH
PENGEKSTRAKAN
Garisan berwarna merah menunjukkan pembahagian metabolit yang mempunyai nilai
VIP ≥1.00 (ke kiri) dan nilai VIP <1.00 (ke kanan)
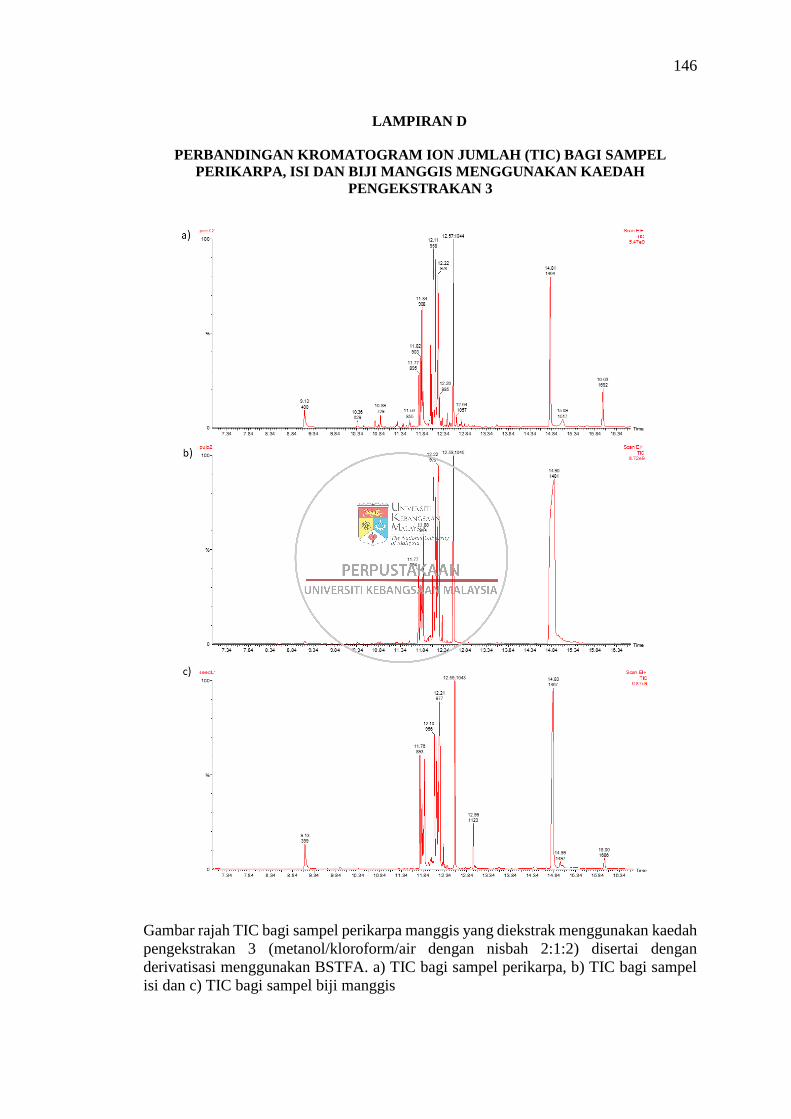
146
LAMPIRAN D
PERBANDINGAN KROMATOGRAM ION JUMLAH (TIC) BAGI SAMPEL
PERIKARPA, ISI DAN BIJI MANGGIS MENGGUNAKAN KAEDAH
PENGEKSTRAKAN 3
Gambar rajah TIC bagi sampel perikarpa manggis yang diekstrak menggunakan kaedah
pengekstrakan 3 (metanol/kloroform/air dengan nisbah 2:1:2) disertai dengan
derivatisasi menggunakan BSTFA. a) TIC bagi sampel perikarpa, b) TIC bagi sampel
isi dan c) TIC bagi sampel biji manggis
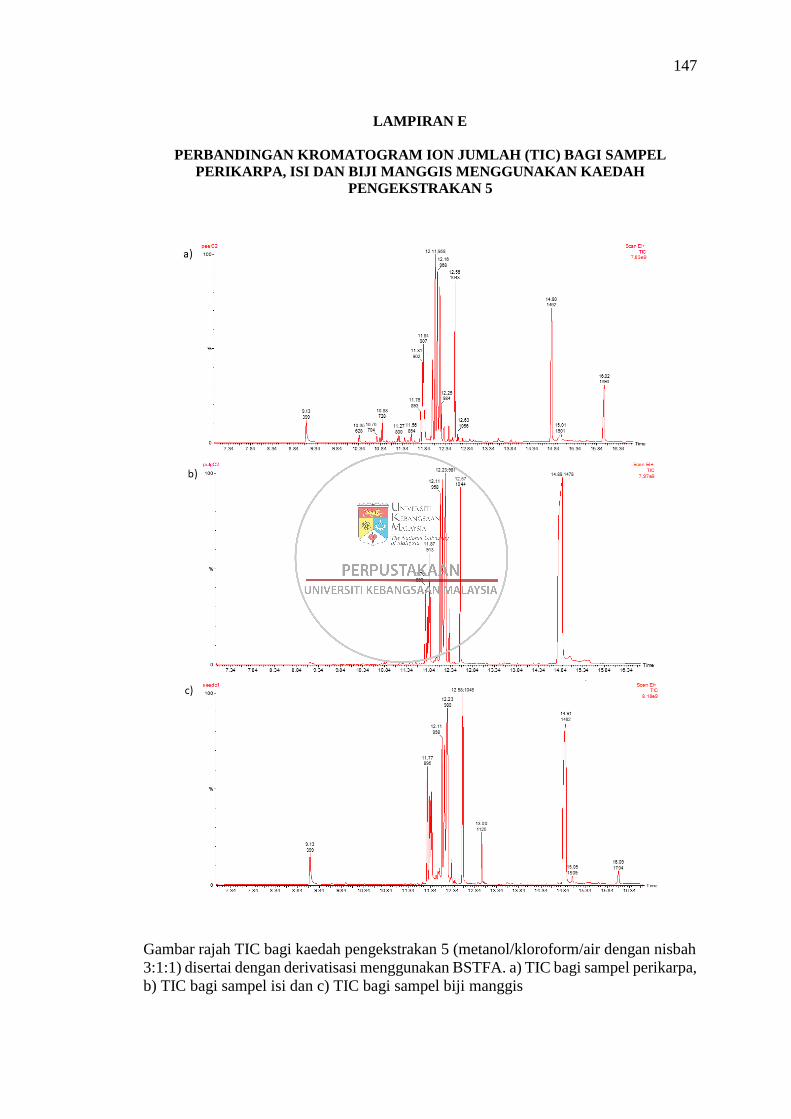
147
LAMPIRAN E
PERBANDINGAN KROMATOGRAM ION JUMLAH (TIC) BAGI SAMPEL
PERIKARPA, ISI DAN BIJI MANGGIS MENGGUNAKAN KAEDAH
PENGEKSTRAKAN 5
Gambar rajah TIC bagi kaedah pengekstrakan 5 (metanol/kloroform/air dengan nisbah
3:1:1) disertai dengan derivatisasi menggunakan BSTFA. a) TIC bagi sampel perikarpa,
b) TIC bagi sampel isi dan c) TIC bagi sampel biji manggis
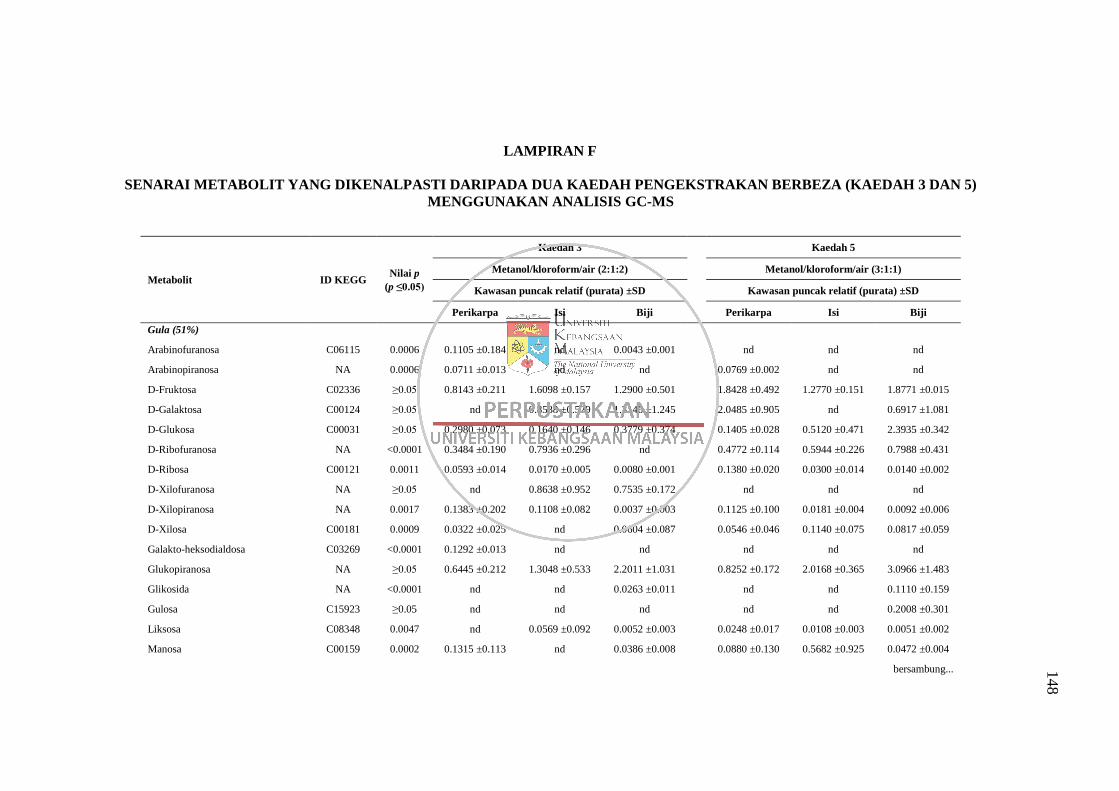
148
LAMPIRAN F
SENARAI METABOLIT YANG DIKENALPASTI DARIPADA DUA KAEDAH PENGEKSTRAKAN BERBEZA (KAEDAH 3 DAN 5)
MENGGUNAKAN ANALISIS GC-MS
Metabolit ID KEGG Nilai p
(p ≤0.05)
Kaedah 3 Kaedah 5
Metanol/kloroform/air (2:1:2) Metanol/kloroform/air (3:1:1)
Kawasan puncak relatif (purata) ±SD Kawasan puncak relatif (purata) ±SD
Perikarpa Isi Biji Perikarpa Isi Biji
Gula (51%)
Arabinofuranosa C06115 0.0006 0.1105 ±0.184 nd 0.0043 ±0.001 nd nd nd
Arabinopiranosa NA 0.0006 0.0711 ±0.013 nd nd 0.0769 ±0.002 nd nd
D-Fruktosa C02336 ≥0.05 0.8143 ±0.211 1.6098 ±0.157 1.2900 ±0.501 1.8428 ±0.492 1.2770 ±0.151 1.8771 ±0.015
D-Galaktosa C00124 ≥0.05 nd 0.8538 ±0.529 1.3146 ±1.245 2.0485 ±0.905 nd 0.6917 ±1.081
D-Glukosa C00031 ≥0.05 0.2980 ±0.073 0.1640 ±0.146 0.3779 ±0.374 0.1405 ±0.028 0.5120 ±0.471 2.3935 ±0.342
D-Ribofuranosa NA <0.0001 0.3484 ±0.190 0.7936 ±0.296 nd 0.4772 ±0.114 0.5944 ±0.226 0.7988 ±0.431
D-Ribosa C00121 0.0011 0.0593 ±0.014 0.0170 ±0.005 0.0080 ±0.001 0.1380 ±0.020 0.0300 ±0.014 0.0140 ±0.002
D-Xilofuranosa NA ≥0.05 nd 0.8638 ±0.952 0.7535 ±0.172 nd nd nd
D-Xilopiranosa NA 0.0017 0.1383 ±0.202 0.1108 ±0.082 0.0037 ±0.003 0.1125 ±0.100 0.0181 ±0.004 0.0092 ±0.006
D-Xilosa C00181 0.0009 0.0322 ±0.025 nd 0.0604 ±0.087 0.0546 ±0.046 0.1140 ±0.075 0.0817 ±0.059
Galakto-heksodialdosa C03269 <0.0001 0.1292 ±0.013 nd nd nd nd nd
Glukopiranosa NA ≥0.05 0.6445 ±0.212 1.3048 ±0.533 2.2011 ±1.031 0.8252 ±0.172 2.0168 ±0.365 3.0966 ±1.483
Glikosida NA <0.0001 nd nd 0.0263 ±0.011 nd nd 0.1110 ±0.159
Gulosa C15923 ≥0.05 nd nd nd nd nd 0.2008 ±0.301
Liksosa C08348 0.0047 nd 0.0569 ±0.092 0.0052 ±0.003 0.0248 ±0.017 0.0108 ±0.003 0.0051 ±0.002
Manosa C00159 0.0002 0.1315 ±0.113 nd 0.0386 ±0.008 0.0880 ±0.130 0.5682 ±0.925 0.0472 ±0.004
bersambung...
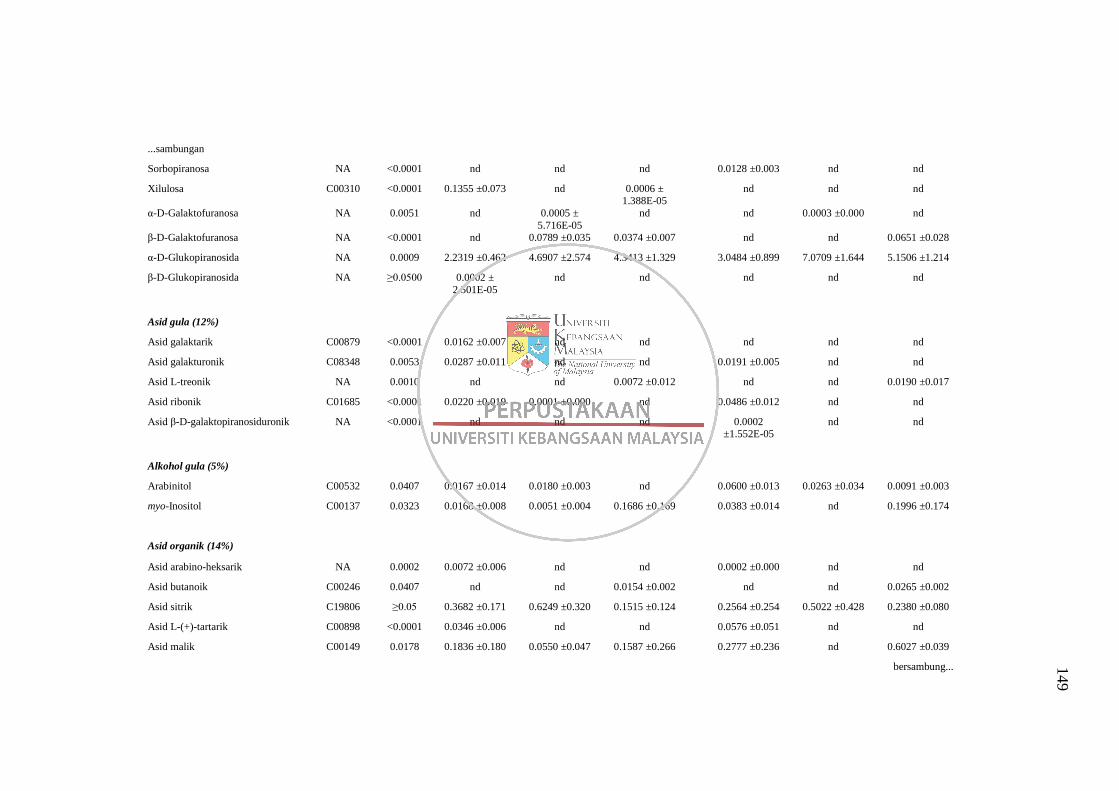
149
...sambungan
Sorbopiranosa NA <0.0001 nd nd nd 0.0128 ±0.003 nd nd
Xilulosa C00310 <0.0001 0.1355 ±0.073 nd 0.0006 ± 1.388E-05
nd nd nd
α-D-Galaktofuranosa NA 0.0051 nd 0.0005 ±
5.716E-05
nd nd 0.0003 ±0.000 nd
β-D-Galaktofuranosa NA <0.0001 nd 0.0789 ±0.035 0.0374 ±0.007 nd nd 0.0651 ±0.028
α-D-Glukopiranosida NA 0.0009 2.2319 ±0.462 4.6907 ±2.574 4.3413 ±1.329 3.0484 ±0.899 7.0709 ±1.644 5.1506 ±1.214
β-D-Glukopiranosida NA ≥0.0500 0.0002 ±
2.501E-05
nd nd nd nd nd
Asid gula (12%)
Asid galaktarik C00879 <0.0001 0.0162 ±0.007 nd nd nd nd nd
Asid galakturonik C08348 0.0053 0.0287 ±0.011 nd nd 0.0191 ±0.005 nd nd
Asid L-treonik NA 0.0010 nd nd 0.0072 ±0.012 nd nd 0.0190 ±0.017
Asid ribonik C01685 <0.0001 0.0220 ±0.019 0.0001 ±0.000 nd 0.0486 ±0.012 nd nd
Asid β-D-galaktopiranosiduronik NA <0.0001 nd nd nd 0.0002 ±1.552E-05
nd nd
Alkohol gula (5%)
Arabinitol C00532 0.0407 0.0167 ±0.014 0.0180 ±0.003 nd 0.0600 ±0.013 0.0263 ±0.034 0.0091 ±0.003
myo-Inositol C00137 0.0323 0.0168 ±0.008 0.0051 ±0.004 0.1686 ±0.169 0.0383 ±0.014 nd 0.1996 ±0.174
Asid organik (14%)
Asid arabino-heksarik NA 0.0002 0.0072 ±0.006 nd nd 0.0002 ±0.000 nd nd
Asid butanoik C00246 0.0407 nd nd 0.0154 ±0.002 nd nd 0.0265 ±0.002
Asid sitrik C19806 ≥0.05 0.3682 ±0.171 0.6249 ±0.320 0.1515 ±0.124 0.2564 ±0.254 0.5022 ±0.428 0.2380 ±0.080
Asid L-(+)-tartarik C00898 <0.0001 0.0346 ±0.006 nd nd 0.0576 ±0.051 nd nd
Asid malik C00149 0.0178 0.1836 ±0.180 0.0550 ±0.047 0.1587 ±0.266 0.2777 ±0.236 nd 0.6027 ±0.039
bersambung...
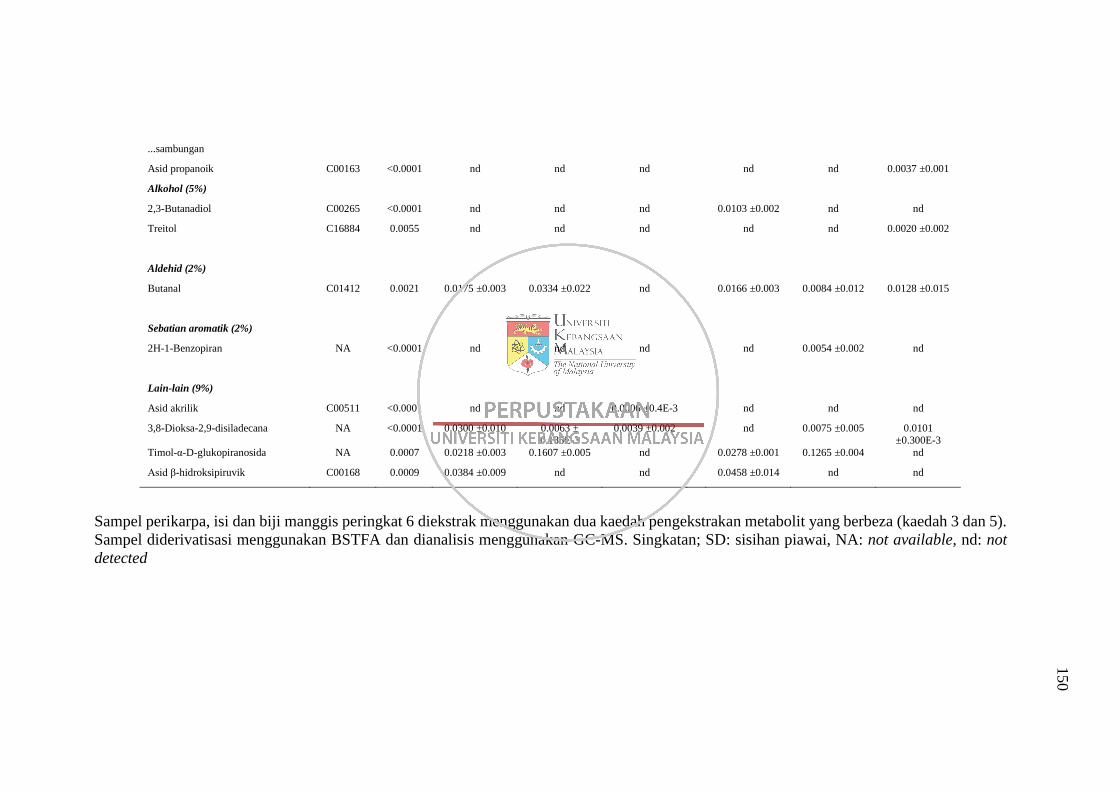
150
...sambungan
Asid propanoik C00163 <0.0001 nd nd nd nd nd 0.0037 ±0.001
Alkohol (5%)
2,3-Butanadiol C00265 <0.0001 nd nd nd 0.0103 ±0.002 nd nd
Treitol C16884 0.0055 nd nd nd nd nd 0.0020 ±0.002
Aldehid (2%)
Butanal C01412 0.0021 0.0175 ±0.003 0.0334 ±0.022 nd 0.0166 ±0.003 0.0084 ±0.012 0.0128 ±0.015
Sebatian aromatik (2%)
2H-1-Benzopiran NA <0.0001 nd nd nd nd 0.0054 ±0.002 nd
Lain-lain (9%)
Asid akrilik C00511 <0.0001 nd nd 0.0006 ±0.4E-3 nd nd nd
3,8-Dioksa-2,9-disiladecana NA <0.0001 0.0300 ±0.010 0.0063 ± 0.185E-3
0.0039 ±0.002 nd 0.0075 ±0.005 0.0101 ±0.300E-3
Timol-α-D-glukopiranosida NA 0.0007 0.0218 ±0.003 0.1607 ±0.005 nd 0.0278 ±0.001 0.1265 ±0.004 nd
Asid β-hidroksipiruvik C00168 0.0009 0.0384 ±0.009 nd nd 0.0458 ±0.014 nd nd
Sampel perikarpa, isi dan biji manggis peringkat 6 diekstrak menggunakan dua kaedah pengekstrakan metabolit yang berbeza (kaedah 3 dan 5).
Sampel diderivatisasi menggunakan BSTFA dan dianalisis menggunakan GC-MS. Singkatan; SD: sisihan piawai, NA: not available, nd: not
detected
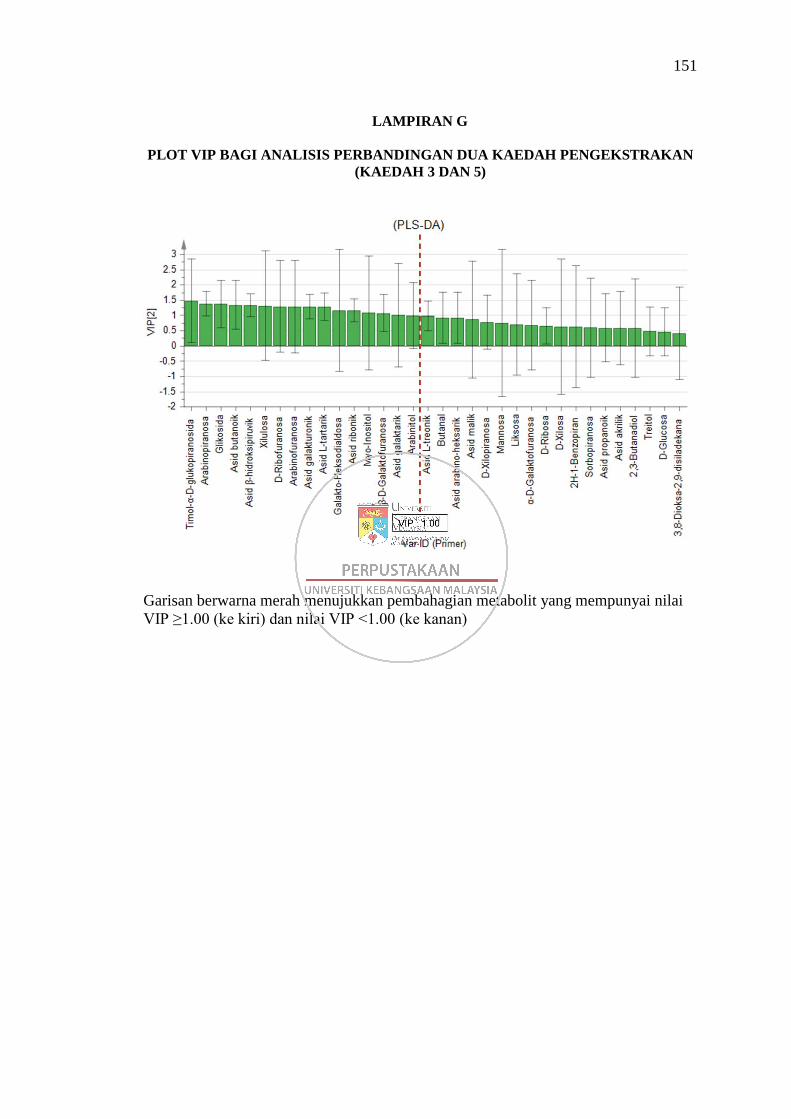
151
LAMPIRAN G
PLOT VIP BAGI ANALISIS PERBANDINGAN DUA KAEDAH PENGEKSTRAKAN
(KAEDAH 3 DAN 5)
Garisan berwarna merah menujukkan pembahagian metabolit yang mempunyai nilai
VIP ≥1.00 (ke kiri) dan nilai VIP <1.00 (ke kanan)
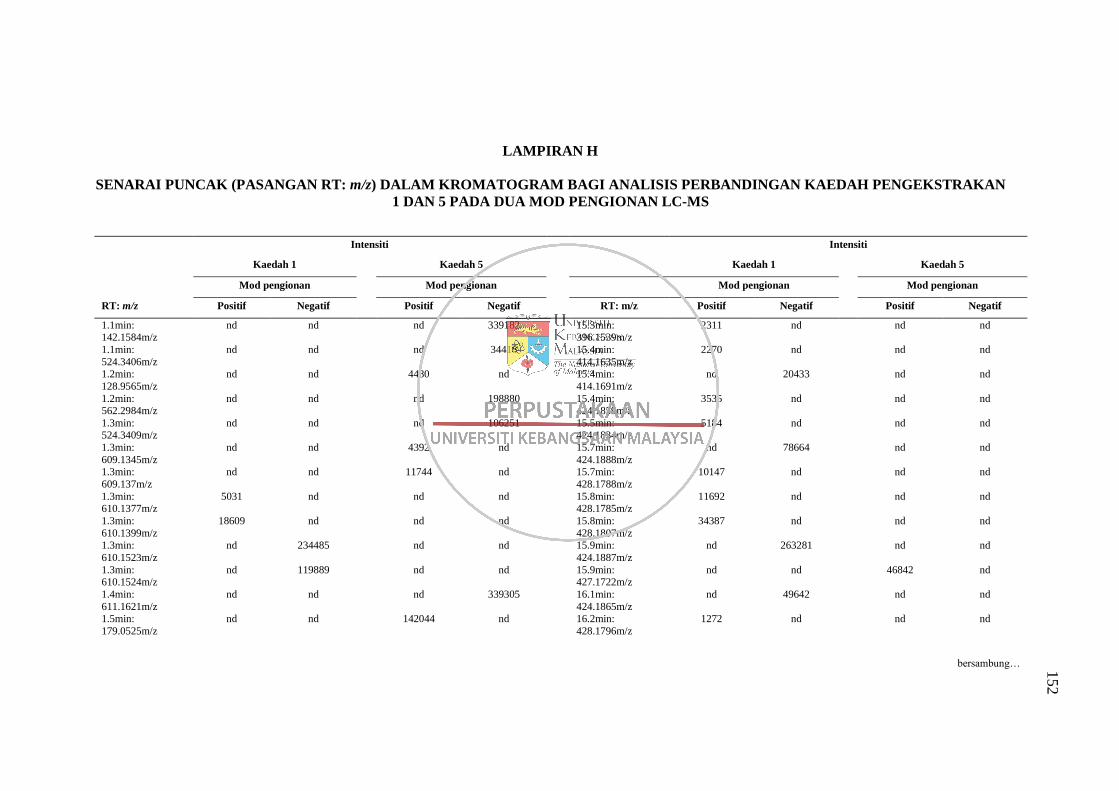
152
LAMPIRAN H
SENARAI PUNCAK (PASANGAN RT: m/z) DALAM KROMATOGRAM BAGI ANALISIS PERBANDINGAN KAEDAH PENGEKSTRAKAN
1 DAN 5 PADA DUA MOD PENGIONAN LC-MS
Intensiti
Intensiti
Kaedah 1
Kaedah 5
Kaedah 1
Kaedah 5
Mod pengionan
Mod pengionan
Mod pengionan
Mod pengionan
RT: m/z Positif Negatif
Positif Negatif
RT: m/z Positif Negatif
Positif Negatif
1.1min: 142.1584m/z
nd nd
nd 339182
15.3min: 396.1539m/z
2311 nd
nd nd
1.1min:
524.3406m/z
nd nd
nd 34418
15.4min:
414.1635m/z
2270 nd
nd nd
1.2min:
128.9565m/z
nd nd
4480 nd
15.4min:
414.1691m/z
nd 20433
nd nd
1.2min: 562.2984m/z
nd nd
nd 198880
15.4min: 424.1859m/z
3535 nd
nd nd
1.3min: 524.3409m/z
nd nd
nd 106251
15.5min: 424.1834m/z
5184 nd
nd nd
1.3min:
609.1345m/z
nd nd
4392 nd
15.7min:
424.1888m/z
nd 78664
nd nd
1.3min:
609.137m/z
nd nd
11744 nd
15.7min:
428.1788m/z
10147 nd
nd nd
1.3min: 610.1377m/z
5031 nd
nd nd
15.8min: 428.1785m/z
11692 nd
nd nd
1.3min:
610.1399m/z
18609 nd
nd nd
15.8min:
428.1807m/z
34387 nd
nd nd
1.3min:
610.1523m/z
nd 234485
nd nd
15.9min:
424.1887m/z
nd 263281
nd nd
1.3min: 610.1524m/z
nd 119889
nd nd
15.9min: 427.1722m/z
nd nd
46842 nd
1.4min:
611.1621m/z
nd nd nd 339305 16.1min:
424.1865m/z
nd 49642 nd nd
1.5min:
179.0525m/z
nd nd 142044 nd 16.2min:
428.1796m/z
1272 nd nd nd
bersambung…
153
sambungan…
1.5min: 179.0526m/z
nd nd
53733 nd
16.6min: 414.1629m/z
4933 nd
nd nd
1.5min:
325.1118m/z
nd nd
nd 331720
16.8min:
414.1627m/z
7942 nd
nd nd
1.5min:
362.1249m/z
23149 nd
nd nd
16.8min:
414.1647m/z
10720 nd
nd nd
1.5min: 387.109m/z
nd nd
170237 nd
16.8min: 414.1678m/z
nd 66117
nd nd
1.5min:
388.1116m/z
24739 nd
nd nd
16.9min:
413.1553m/z
nd nd
18508 nd
1.5min:
388.1134m/z
103795 nd
nd nd
16.9min:
413.1556m/z
nd nd
24722 nd
1.6min:
134.0457m/z
nd nd
nd 132995
16min:
425.1937m/z
nd nd
nd 150550
1.6min: 866.1859m/z
58196 nd
nd nd
17.1min: 415.1731m/z
nd nd
nd 71524
1.6min:
866.2023m/z
nd 95559
nd nd
18.2min:
407.1468m/z
nd nd
5518 nd
1.7min:
524.3411m/z
nd nd
nd 104985
2.1min:
354.0882m/z
13103 nd
nd nd
1.7min: 578.1271m/z
26084 nd
nd nd
2.1min: 354.0943m/z
nd 69590
nd nd
1.7min:
578.1293m/z
30194 nd
nd nd
2.2min:
354.0946m/z
nd 56961
nd nd
1.7min:
578.1388m/z
nd 117294
nd nd
2.2min:
417.3582m/z
nd 24234
nd nd
1.7min: 578.1415m/z
nd 204343
nd nd
2.3min: 191.0524m/z
nd nd
24640 nd
1.7min:
866.1836m/z
13344 nd nd nd 2.3min:
193.0319m/z
nd nd 57304 nd
1.7min:
866.1857m/z
24892 nd nd nd 2.3min:
291.0868m/z
nd nd nd 592602
1.7min: 866.2049m/z
nd 334654 nd nd 2.3min: 405.0618m/z
30385 nd nd nd
1.8min:
524.3438m/z
nd nd nd 128838 2.3min:
965.8846m/z
nd 49249 nd nd
1.8min:
578.1391m/z
nd 90857 nd nd 2.4min:
133.0112m/z
nd nd 111733 nd
10.1min: 524.1491m/z
8890 nd nd nd 2.4min: 291.0869m/z
nd nd nd 548387
bersambung…
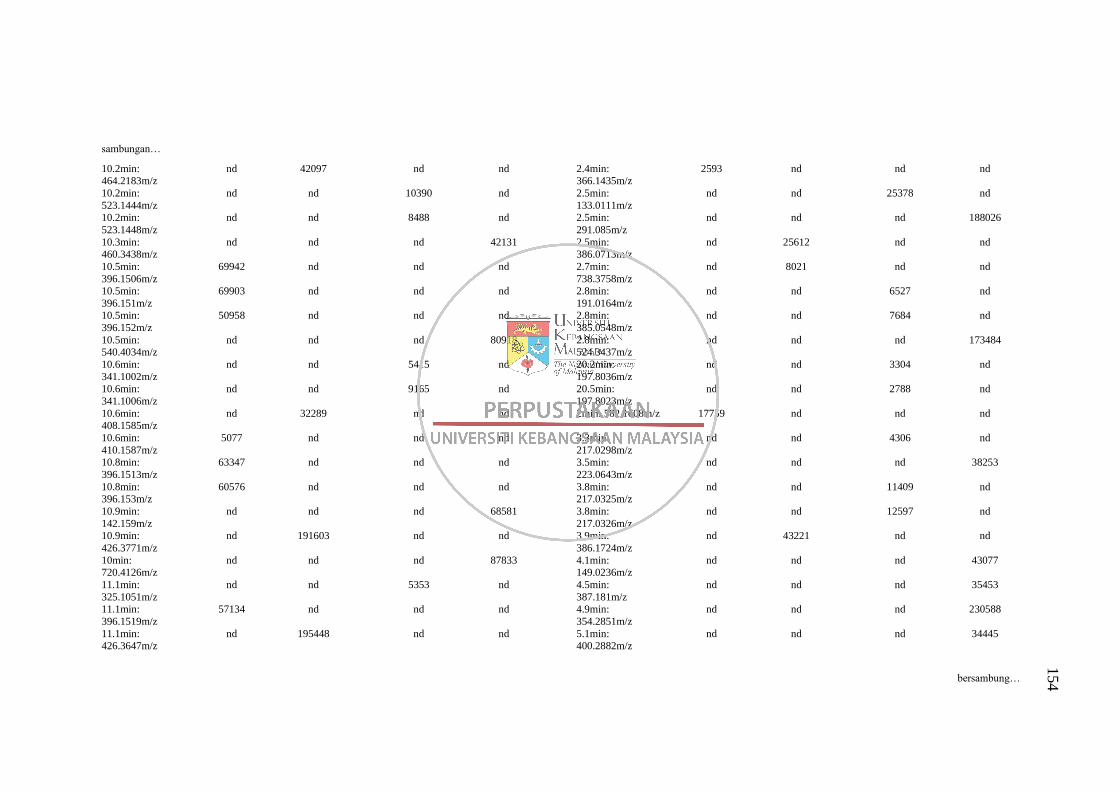
154
sambungan…
10.2min: 464.2183m/z
nd 42097
nd nd
2.4min: 366.1435m/z
2593 nd
nd nd
10.2min:
523.1444m/z
nd nd
10390 nd
2.5min:
133.0111m/z
nd nd
25378 nd
10.2min:
523.1448m/z
nd nd
8488 nd
2.5min:
291.085m/z
nd nd
nd 188026
10.3min: 460.3438m/z
nd nd
nd 42131
2.5min: 386.0713m/z
nd 25612
nd nd
10.5min:
396.1506m/z
69942 nd
nd nd
2.7min:
738.3758m/z
nd 8021
nd nd
10.5min:
396.151m/z
69903 nd
nd nd
2.8min:
191.0164m/z
nd nd
6527 nd
10.5min:
396.152m/z
50958 nd
nd nd
2.8min:
385.0548m/z
nd nd
7684 nd
10.5min: 540.4034m/z
nd nd
nd 80918
2.8min: 524.3437m/z
nd nd
nd 173484
10.6min:
341.1002m/z
nd nd
5415 nd
20.2min:
197.8036m/z
nd nd
3304 nd
10.6min:
341.1006m/z
nd nd
9165 nd
20.5min:
197.8023m/z
nd nd
2788 nd
10.6min: 408.1585m/z
nd 32289
nd nd
2min: 582.1608m/z 17759 nd
nd nd
10.6min:
410.1587m/z
5077 nd
nd nd
3.3min:
217.0298m/z
nd nd
4306 nd
10.8min:
396.1513m/z
63347 nd
nd nd
3.5min:
223.0643m/z
nd nd
nd 38253
10.8min: 396.153m/z
60576 nd
nd nd
3.8min: 217.0325m/z
nd nd
11409 nd
10.9min:
142.159m/z
nd nd
nd 68581
3.8min:
217.0326m/z
nd nd
12597 nd
10.9min:
426.3771m/z
nd 191603
nd nd
3.9min:
386.1724m/z
nd 43221
nd nd
10min: 720.4126m/z
nd nd
nd 87833
4.1min: 149.0236m/z
nd nd
nd 43077
11.1min:
325.1051m/z
nd nd
5353 nd
4.5min:
387.181m/z
nd nd
nd 35453
11.1min:
396.1519m/z
57134 nd
nd nd
4.9min:
354.2851m/z
nd nd
nd 230588
11.1min: 426.3647m/z
nd 195448
nd nd
5.1min: 400.2882m/z
nd nd
nd 34445
bersambung…
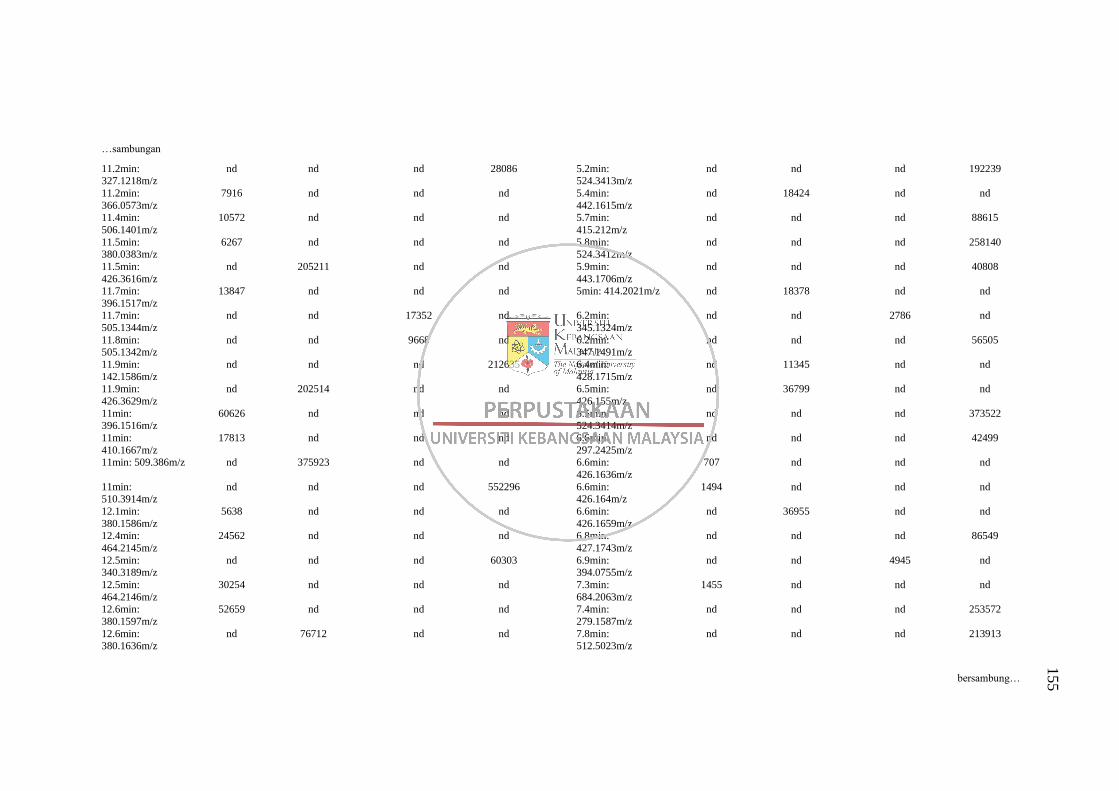
155
…sambungan
11.2min: 327.1218m/z
nd nd
nd 28086
5.2min: 524.3413m/z
nd nd
nd 192239
11.2min:
366.0573m/z
7916 nd
nd nd
5.4min:
442.1615m/z
nd 18424
nd nd
11.4min:
506.1401m/z
10572 nd
nd nd
5.7min:
415.212m/z
nd nd
nd 88615
11.5min: 380.0383m/z
6267 nd
nd nd
5.8min: 524.3412m/z
nd nd
nd 258140
11.5min:
426.3616m/z
nd 205211
nd nd
5.9min:
443.1706m/z
nd nd
nd 40808
11.7min:
396.1517m/z
13847 nd
nd nd
5min: 414.2021m/z nd 18378
nd nd
11.7min:
505.1344m/z
nd nd
17352 nd
6.2min:
345.1324m/z
nd nd
2786 nd
11.8min: 505.1342m/z
nd nd
9668 nd
6.2min: 347.1491m/z
nd nd
nd 56505
11.9min:
142.1586m/z
nd nd
nd 212635
6.4min:
428.1715m/z
nd 11345
nd nd
11.9min:
426.3629m/z
nd 202514
nd nd
6.5min:
426.155m/z
nd 36799
nd nd
11min: 396.1516m/z
60626 nd
nd nd
6.5min: 524.3414m/z
nd nd
nd 373522
11min:
410.1667m/z
17813 nd
nd nd
6.6min:
297.2425m/z
nd nd
nd 42499
11min: 509.386m/z nd 375923
nd nd
6.6min:
426.1636m/z
707 nd
nd nd
11min: 510.3914m/z
nd nd
nd 552296
6.6min: 426.164m/z
1494 nd
nd nd
12.1min:
380.1586m/z
5638 nd
nd nd
6.6min:
426.1659m/z
nd 36955
nd nd
12.4min:
464.2145m/z
24562 nd
nd nd
6.8min:
427.1743m/z
nd nd
nd 86549
12.5min: 340.3189m/z
nd nd
nd 60303
6.9min: 394.0755m/z
nd nd
4945 nd
12.5min:
464.2146m/z
30254 nd
nd nd
7.3min:
684.2063m/z
1455 nd
nd nd
12.6min:
380.1597m/z
52659 nd
nd nd
7.4min:
279.1587m/z
nd nd
nd 253572
12.6min: 380.1636m/z
nd 76712
nd nd
7.8min: 512.5023m/z
nd nd
nd 213913
bersambung…

156
sambungan…
12.7min: 379.1522m/z
nd nd
50039 nd
7.8min: 524.3404m/z
nd nd
nd 319644
12.7min:
379.1525m/z
nd nd
35857 nd
7.9min:
598.422m/z
nd nd
nd 122319
12.8min:
381.1682m/z
nd nd
nd 153048
7min: 394.0738m/z nd nd
3903 nd
12.8min: 426.379m/z
nd 219337
nd nd
7min: 400.1812m/z 1492 nd
nd nd
12.9min:
178.8739m/z
nd nd
10293 nd
7min: 442.1551m/z 2718 nd
nd nd
12min:
523.4022m/z
nd 136850
nd nd
8.2min:
736.4094m/z
nd nd
nd 45746
12min:
524.4078m/z
nd nd
nd 343737
8.3min:
382.1732m/z
5077 nd
nd nd
13.1min: 142.1574m/z
nd nd
nd 238988
8.3min: 382.173m/z
12795 nd
nd nd
13.1min:
178.8755m/z
nd nd
8612 nd
8.3min:
382.1773m/z
nd 31518
nd nd
13.4min:
142.1573m/z
nd nd
nd 236968
8.4min:
381.1683m/z
nd nd
5118 nd
13.4min: 178.8757m/z
nd nd
6755 nd
8.5min: 396.1568m/z
nd 62421
nd nd
13.5min:
380.1581m/z
3455 nd
nd nd
8.6min:
525.3797m/z
nd 90022
nd nd
13.8min:
560.4319m/z
nd nd
nd 36480
8.7min:
526.3762m/z
nd nd
nd 162718
14.2min: 396.1528m/z
46896 nd
nd nd
8.8min: 397.1638m/z
nd nd
nd 136688
14.2min:
396.1549m/z
63970 nd
nd nd
9.1min:
352.2563m/z
3537 nd
nd nd
14.3min:
395.1466m/z
nd nd
39374 nd
9.2min:
539.3933m/z
nd 35547
nd nd
14.3min: 395.1468m/z
nd nd
55016 nd
9.3min: 410.165m/z
2803 nd
nd nd
14.3min:
396.1583m/z
nd 167506
nd nd
9.3min:
410.1716m/z
nd 149347
nd nd
14.4min:
397.1637m/z
nd nd nd 220530 9.3min:
540.4042m/z
nd nd nd 86162
14.4min: 424.1901m/z
nd 95203 nd nd 9.4min: 510.3893m/z
nd nd nd 48170
bersambung…
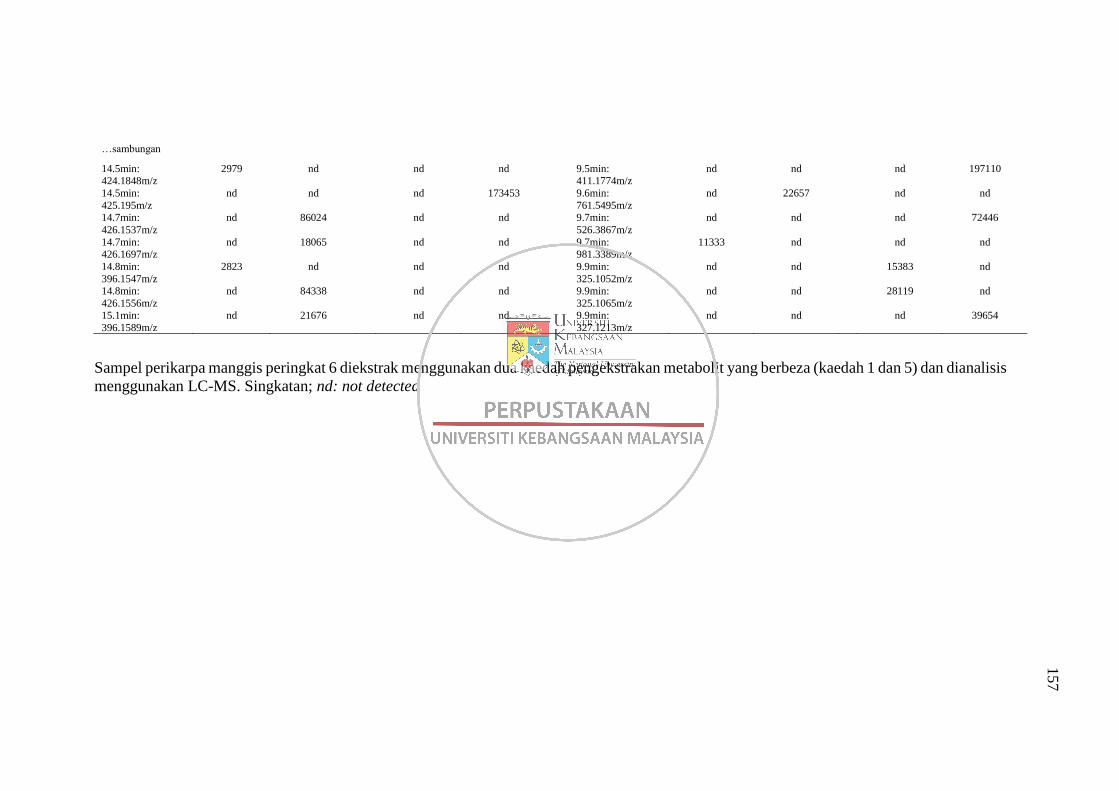
157
…sambungan
14.5min: 424.1848m/z
2979 nd nd nd 9.5min: 411.1774m/z
nd nd nd 197110
14.5min:
425.195m/z
nd nd nd 173453 9.6min:
761.5495m/z
nd 22657 nd nd
14.7min:
426.1537m/z
nd 86024
nd nd
9.7min:
526.3867m/z
nd nd
nd 72446
14.7min: 426.1697m/z
nd 18065
nd nd
9.7min: 981.3389m/z
11333 nd
nd nd
14.8min:
396.1547m/z
2823 nd
nd nd
9.9min:
325.1052m/z
nd nd
15383 nd
14.8min:
426.1556m/z
nd 84338
nd nd
9.9min:
325.1065m/z
nd nd
28119 nd
15.1min:
396.1589m/z
nd 21676
nd nd
9.9min:
327.1213m/z
nd nd
nd 39654
Sampel perikarpa manggis peringkat 6 diekstrak menggunakan dua kaedah pengekstrakan metabolit yang berbeza (kaedah 1 dan 5) dan dianalisis
menggunakan LC-MS. Singkatan; nd: not detected
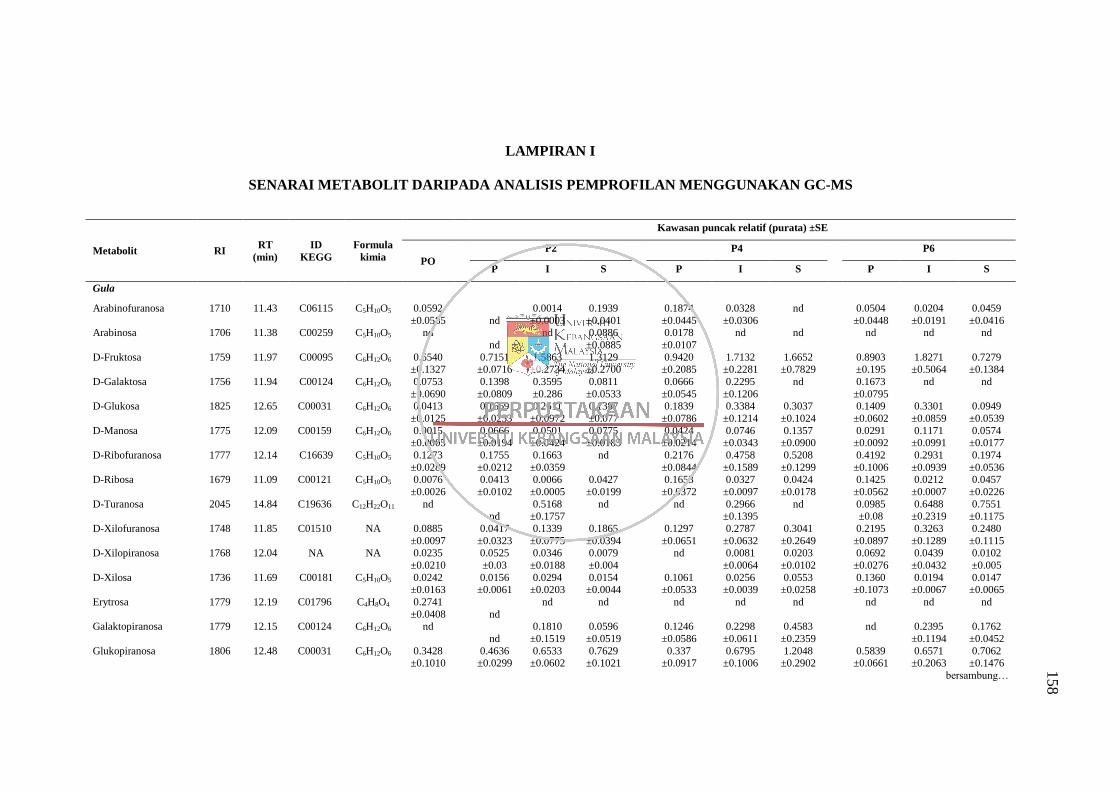
158
LAMPIRAN I
SENARAI METABOLIT DARIPADA ANALISIS PEMPROFILAN MENGGUNAKAN GC-MS
Metabolit RI RT
(min)
ID
KEGG
Formula
kimia
Kawasan puncak relatif (purata) ±SE
PO
P2 P4 P6
P I S P I S P I S
Gula
Arabinofuranosa 1710 11.43 C06115 C5H10O5 0.0592
±0.0585
nd
0.0014
±0.0003
0.1939
±0.0401
0.1874
±0.0445
0.0328
±0.0306
nd 0.0504
±0.0448
0.0204
±0.0191
0.0459
±0.0416 Arabinosa 1706 11.38 C00259 C5H10O5 nd
nd
nd 0.0886
±0.0885
0.0178
±0.0107
nd nd nd nd nd
D-Fruktosa 1759 11.97 C00095 C6H12O6 0.6540 ±0.1327
0.7151 ±0.0716
1.5863 ±0.2734
1.3129 ±0.2700
0.9420 ±0.2085
1.7132 ±0.2281
1.6652 ±0.7829
0.8903 ±0.195
1.8271 ±0.5064
0.7279 ±0.1384
D-Galaktosa 1756 11.94 C00124 C6H12O6 0.0753
±0.0690
0.1398
±0.0809
0.3595
±0.286
0.0811
±0.0533
0.0666
±0.0545
0.2295
±0.1206
nd 0.1673
±0.0795
nd nd
D-Glukosa 1825 12.65 C00031 C6H12O6 0.0413
±0.0125
0.1669
±0.0253
0.2611
±0.0972
0.1387
±0.077
0.1839
±0.0786
0.3384
±0.1214
0.3037
±0.1024
0.1409
±0.0602
0.3301
±0.0859
0.0949
±0.0539
D-Manosa 1775 12.09 C00159 C6H12O6 0.0015 ±0.0005
0.0666 ±0.0194
0.0501 ±0.0424
0.0775 ±0.0185
0.0424 ±0.0214
0.0746 ±0.0343
0.1357 ±0.0900
0.0291 ±0.0092
0.1171 ±0.0991
0.0574 ±0.0177
D-Ribofuranosa 1777 12.14 C16639 C5H10O5 0.1273
±0.0289
0.1755
±0.0212
0.1663
±0.0359
nd 0.2176
±0.0844
0.4758
±0.1589
0.5208
±0.1299
0.4192
±0.1006
0.2931
±0.0939
0.1974
±0.0536 D-Ribosa 1679 11.09 C00121 C5H10O5 0.0076
±0.0026
0.0413
±0.0102
0.0066
±0.0005
0.0427
±0.0199
0.1653
±0.0372
0.0327
±0.0097
0.0424
±0.0178
0.1425
±0.0562
0.0212
±0.0007
0.0457
±0.0226
D-Turanosa 2045 14.84 C19636 C12H22O11 nd nd
0.5168 ±0.1757
nd nd 0.2966 ±0.1395
nd 0.0985 ±0.08
0.6488 ±0.2319
0.7551 ±0.1175
D-Xilofuranosa 1748 11.85 C01510 NA 0.0885
±0.0097
0.0417
±0.0323
0.1339
±0.0775
0.1865
±0.0394
0.1297
±0.0651
0.2787
±0.0632
0.3041
±0.2649
0.2195
±0.0897
0.3263
±0.1289
0.2480
±0.1115
D-Xilopiranosa 1768 12.04 NA NA 0.0235
±0.0210
0.0525
±0.03
0.0346
±0.0188
0.0079
±0.004
nd 0.0081
±0.0064
0.0203
±0.0102
0.0692
±0.0276
0.0439
±0.0432
0.0102
±0.005
D-Xilosa 1736 11.69 C00181 C5H10O5 0.0242 ±0.0163
0.0156 ±0.0061
0.0294 ±0.0203
0.0154 ±0.0044
0.1061 ±0.0533
0.0256 ±0.0039
0.0553 ±0.0258
0.1360 ±0.1073
0.0194 ±0.0067
0.0147 ±0.0065
Erytrosa 1779 12.19 C01796 C4H8O4 0.2741
±0.0408
nd
nd nd nd nd nd nd nd nd
Galaktopiranosa 1779 12.15 C00124 C6H12O6 nd
nd
0.1810
±0.1519
0.0596
±0.0519
0.1246
±0.0586
0.2298
±0.0611
0.4583
±0.2359
nd 0.2395
±0.1194
0.1762
±0.0452
Glukopiranosa 1806 12.48 C00031 C6H12O6 0.3428 ±0.1010
0.4636 ±0.0299
0.6533 ±0.0602
0.7629 ±0.1021
0.337 ±0.0917
0.6795 ±0.1006
1.2048 ±0.2902
0.5839 ±0.0661
0.6571 ±0.2063
0.7062 ±0.1476
bersambung…
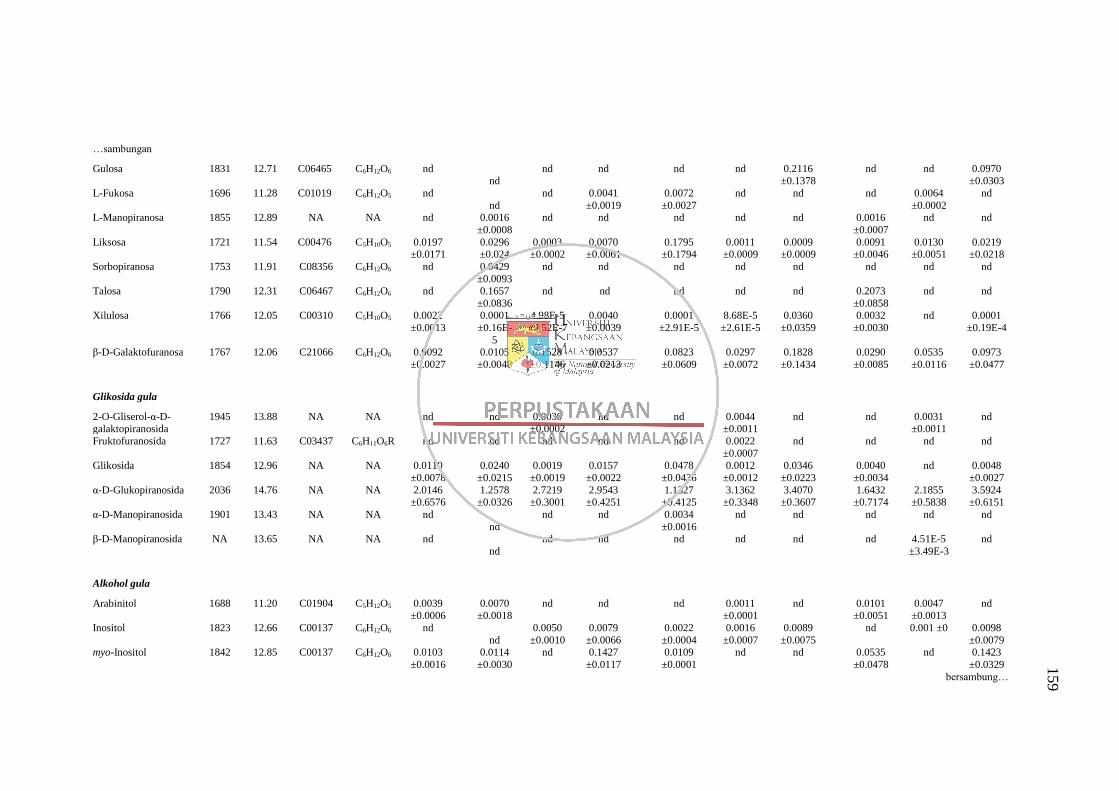
159
…sambungan
Gulosa 1831 12.71 C06465 C6H12O6
nd
nd nd nd nd nd 0.2116
±0.1378 nd nd 0.0970
±0.0303
L-Fukosa 1696 11.28 C01019 C6H12O5 nd
nd
nd 0.0041
±0.0019
0.0072
±0.0027
nd nd nd 0.0064
±0.0002
nd
L-Manopiranosa 1855 12.89 NA NA nd 0.0016
±0.0008
nd nd nd nd nd 0.0016
±0.0007
nd nd
Liksosa 1721 11.54 C00476 C5H10O5 0.0197 ±0.0171
0.0296 ±0.024
0.0003 ±0.0002
0.0070 ±0.0061
0.1795 ±0.1794
0.0011 ±0.0009
0.0009 ±0.0009
0.0091 ±0.0046
0.0130 ±0.0051
0.0219 ±0.0218
Sorbopiranosa 1753 11.91 C08356 C6H12O6 nd 0.0429
±0.0093
nd nd nd nd nd nd nd nd
Talosa 1790 12.31 C06467 C6H12O6 nd 0.1657
±0.0836
nd nd nd nd nd 0.2073
±0.0858
nd nd
Xilulosa 1766 12.05 C00310 C5H10O5 0.0022
±0.0013
0.0001
±0.16E-
5
4.98E-5
±9.52E-7
0.0040
±0.0039
0.0001
±2.91E-5
8.68E-5
±2.61E-5
0.0360
±0.0359
0.0032
±0.0030
nd 0.0001
±0.19E-4
β-D-Galaktofuranosa 1767 12.06 C21066 C6H12O6 0.0092
±0.0027
0.0105
±0.0048
0.1528
±0.1146
0.0537
±0.0213
0.0823
±0.0609
0.0297
±0.0072
0.1828
±0.1434
0.0290
±0.0085
0.0535
±0.0116
0.0973
±0.0477
Glikosida gula
2-O-Gliserol-α-D-galaktopiranosida
1945 13.88 NA NA nd nd 0.0030 ±0.0002
nd nd 0.0044 ±0.0011
nd nd 0.0031 ±0.0011
nd
Fruktofuranosida 1727 11.63 C03437 C6H11O6R nd nd nd nd nd 0.0022
±0.0007
nd nd nd nd
Glikosida 1854 12.96 NA NA 0.0110
±0.0078
0.0240
±0.0215
0.0019
±0.0019
0.0157
±0.0022
0.0478
±0.0436
0.0012
±0.0012
0.0346
±0.0223
0.0040
±0.0034
nd 0.0048
±0.0027
α-D-Glukopiranosida 2036 14.76 NA NA 2.0146 ±0.6576
1.2578 ±0.0326
2.7219 ±0.3001
2.9543 ±0.4251
1.1327 ±0.4125
3.1362 ±0.3348
3.4070 ±0.3607
1.6432 ±0.7174
2.1855 ±0.5838
3.5924 ±0.6151
α-D-Manopiranosida 1901 13.43 NA NA nd
nd
nd nd 0.0034
±0.0016
nd nd nd nd nd
β-D-Manopiranosida NA 13.65 NA NA nd
nd
nd nd nd nd nd nd 4.51E-5
±3.49E-3
nd
Alkohol gula
Arabinitol 1688 11.20 C01904 C5H12O5 0.0039 ±0.0006
0.0070 ±0.0018
nd nd nd 0.0011 ±0.0001
nd 0.0101 ±0.0051
0.0047 ±0.0013
nd
Inositol 1823 12.66 C00137 C6H12O6 nd
nd
0.0050
±0.0010
0.0079
±0.0066
0.0022
±0.0004
0.0016
±0.0007
0.0089
±0.0075
nd 0.001 ±0 0.0098
±0.0079 myo-Inositol 1842 12.85 C00137 C6H12O6
0.0103
±0.0016
0.0114
±0.0030
nd 0.1427
±0.0117
0.0109
±0.0001
nd nd 0.0535
±0.0478
nd 0.1423
±0.0329
bersambung…
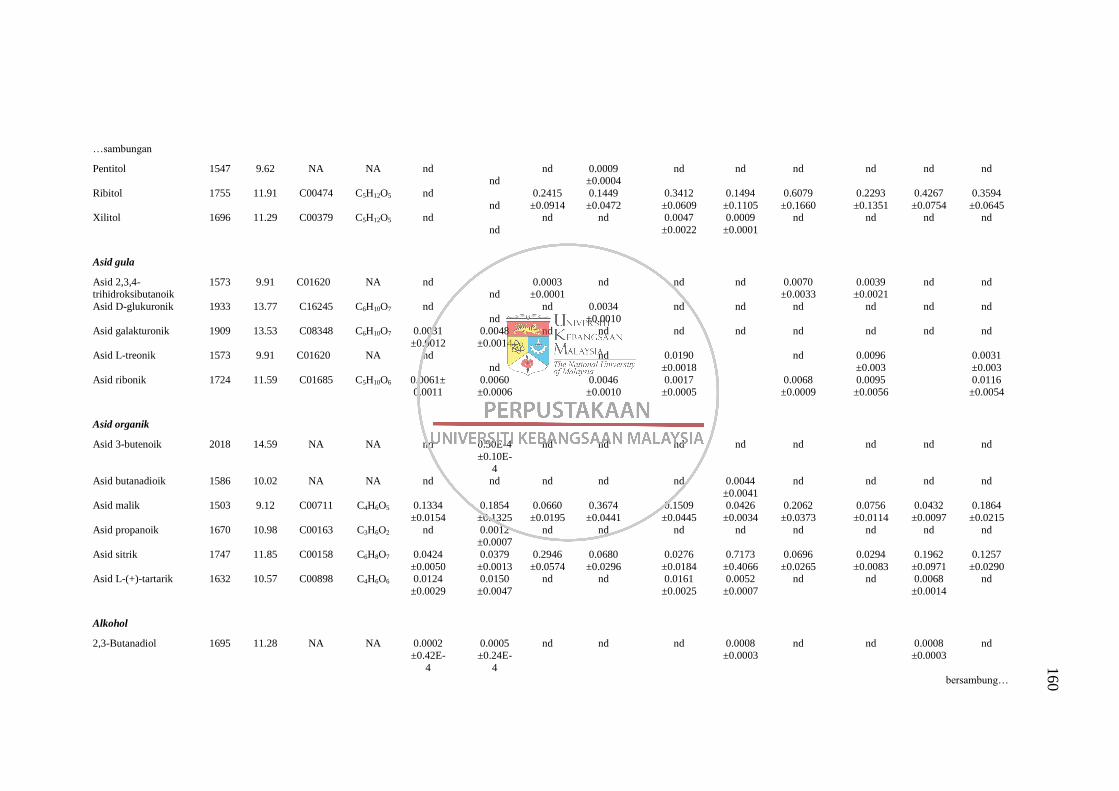
160
…sambungan
Pentitol 1547 9.62 NA NA nd nd
nd 0.0009 ±0.0004
nd nd nd nd nd nd
Ribitol 1755 11.91 C00474 C5H12O5
nd
nd
0.2415
±0.0914
0.1449
±0.0472
0.3412
±0.0609
0.1494
±0.1105
0.6079
±0.1660
0.2293
±0.1351
0.4267
±0.0754
0.3594
±0.0645 Xilitol 1696 11.29 C00379 C5H12O5
nd
nd
nd nd 0.0047
±0.0022
0.0009
±0.0001
nd nd nd nd
Asid gula
Asid 2,3,4-trihidroksibutanoik
1573 9.91 C01620 NA nd nd
0.0003 ±0.0001
nd nd nd 0.0070 ±0.0033
0.0039 ±0.0021
nd nd
Asid D-glukuronik 1933 13.77 C16245 C6H10O7 nd
nd
nd 0.0034
±0.0010
nd nd nd nd nd nd
Asid galakturonik 1909 13.53 C08348 C6H10O7 0.0031
±0.0012
0.0048
±0.0014
nd nd nd nd nd nd nd nd
Asid L-treonik 1573 9.91 C01620 NA nd
nd
nd 0.0190
±0.0018
nd 0.0096
±0.003
0.0031
±0.003
Asid ribonik 1724 11.59 C01685 C5H10O6 0.0061±
0.0011
0.0060
±0.0006
0.0046
±0.0010
0.0017
±0.0005
0.0068
±0.0009
0.0095
±0.0056
0.0116
±0.0054
Asid organik
Asid 3-butenoik 2018 14.59 NA NA nd 0.50E-4
±0.10E-4
nd nd nd nd nd nd nd nd
Asid butanadioik 1586 10.02 NA NA nd nd nd nd nd 0.0044
±0.0041
nd nd nd nd
Asid malik 1503 9.12 C00711 C4H6O5 0.1334
±0.0154
0.1854
±0.1325
0.0660
±0.0195
0.3674
±0.0441
0.1509
±0.0445
0.0426
±0.0034
0.2062
±0.0373
0.0756
±0.0114
0.0432
±0.0097
0.1864
±0.0215
Asid propanoik 1670 10.98 C00163 C3H6O2 nd 0.0012 ±0.0007
nd nd nd nd nd nd nd nd
Asid sitrik 1747 11.85 C00158 C6H8O7 0.0424
±0.0050
0.0379
±0.0013
0.2946
±0.0574
0.0680
±0.0296
0.0276
±0.0184
0.7173
±0.4066
0.0696
±0.0265
0.0294
±0.0083
0.1962
±0.0971
0.1257
±0.0290 Asid L-(+)-tartarik 1632 10.57 C00898 C4H6O6 0.0124
±0.0029
0.0150
±0.0047
nd nd 0.0161
±0.0025
0.0052
±0.0007
nd nd 0.0068
±0.0014
nd
Alkohol
2,3-Butanadiol 1695 11.28 NA NA 0.0002 ±0.42E-
4
0.0005 ±0.24E-
4
nd nd nd 0.0008 ±0.0003
nd nd 0.0008 ±0.0003
nd
bersambung…

161
…sambungan
Sebatian aromatik
2H-1-Benzopiran 2170 15.99 C05851 C9H6O2 nd 0.0665 ±0.0648
nd 0.0030 ±0.0012
0.0094 ±0.0035
nd 0.0360 ±0.0265
0.0670 ±0.0451
0.0043 ±0.0011
0.0208 ±0.0162
Aldehid
Butanal 1669 10.99 C01412 C4H8O 0.0052
±0.0016
0.0062
±0.0016
0.0033
±0.0021
0.0013
±0.0001
nd 0.0116
±0.0042
0.0051
±0.0023
0.0057
±0.0026
nd 0.0008
±0.0002
Lain-lain
3,8-Dioksa-2,9-
disiladekana
1698 11.31 NA NA 0.0283
±0.0270
0.0535
±0.0483
0.0008
±0.0003
nd 4.81E-5
±3.11E-6
0.0012
±0.0011
0.0021
±0.0009
0.0122
±0.0110
0.0080
±0.0072
0.0007
±0.0002
Asid β-hydroksipiruvik
1781 12.20 NA NA nd nd
nd 0.0149 ±0.0075
nd nd nd 0.0192 ±0.0053
nd nd
Asid akrilik 1786 12.27 C00511 C3H4O2 nd
nd
nd 0.0002
±0.0001
nd nd nd nd nd 0.0075
±0.0074
Benzena 1717 11.48 C01407 C6H6 nd
nd
nd 0.0032
±0.0015
nd nd nd nd nd 0.0070
±0.0036 Butana 1718 11.49 C21390 C4H10 nd
nd
nd nd 0.0088
±0.0027
nd nd nd 0.0463
±0.0405
nd
Gliserol 1743 11.80 NA NA nd nd
nd 0.6271 ±0.0551
0.1480 ±0.0916
0.3880 ±0.0612
nd nd nd nd
Isoxantopterin 2160 15.90 C03975 C6H5N5O2 nd
nd
0.0004
±2.87E-5
nd nd nd nd nd nd nd
Timol-β-D-
glukopiranosida
2107 15.41 NA NA nd 0.0033
±0.0005
nd nd nd nd nd nd nd nd
Sampel perikarpa, isi dan biji manggis diekstrak menggunakan metanol/kloroform/air (3:1:1) diikuti dengan derivatisasi menggunakan BSTFA
dan analisis GC-MS. Singkatan; NA: not available, nd: not detected, RI: indeks penahanan, RT: masa penahanan, SE: ralat piawai, P0: peringkat
0, P2: peringkat 2, P4: peringkat 4, P6: peringkat 6, P: perikarpa, I: isi dan B: biji
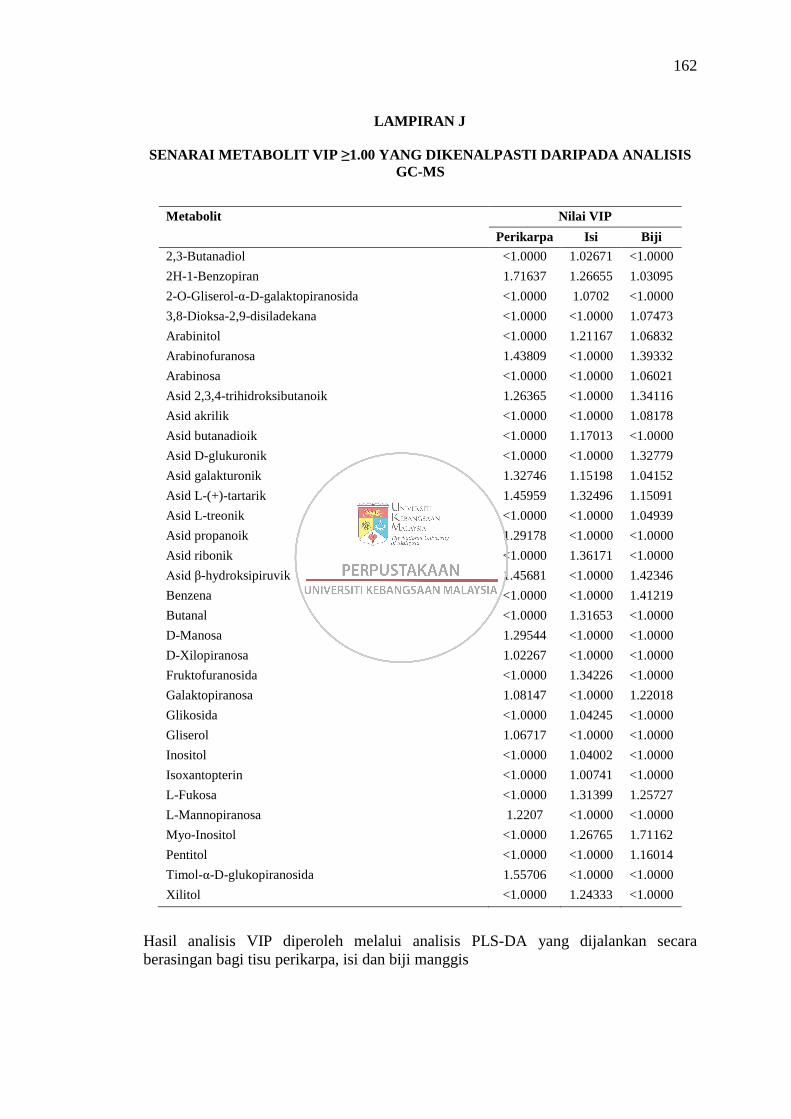
162
LAMPIRAN J
SENARAI METABOLIT VIP ≥1.00 YANG DIKENALPASTI DARIPADA ANALISIS
GC-MS
Metabolit Nilai VIP
Perikarpa Isi Biji
2,3-Butanadiol <1.0000 1.02671 <1.0000
2H-1-Benzopiran 1.71637 1.26655 1.03095
2-O-Gliserol-α-D-galaktopiranosida <1.0000 1.0702 <1.0000
3,8-Dioksa-2,9-disiladekana <1.0000 <1.0000 1.07473
Arabinitol <1.0000 1.21167 1.06832
Arabinofuranosa 1.43809 <1.0000 1.39332
Arabinosa <1.0000 <1.0000 1.06021
Asid 2,3,4-trihidroksibutanoik 1.26365 <1.0000 1.34116
Asid akrilik <1.0000 <1.0000 1.08178
Asid butanadioik <1.0000 1.17013 <1.0000
Asid D-glukuronik <1.0000 <1.0000 1.32779
Asid galakturonik 1.32746 1.15198 1.04152
Asid L-(+)-tartarik 1.45959 1.32496 1.15091
Asid L-treonik <1.0000 <1.0000 1.04939
Asid propanoik 1.29178 <1.0000 <1.0000
Asid ribonik <1.0000 1.36171 <1.0000
Asid β-hydroksipiruvik 1.45681 <1.0000 1.42346
Benzena <1.0000 <1.0000 1.41219
Butanal <1.0000 1.31653 <1.0000
D-Manosa 1.29544 <1.0000 <1.0000
D-Xilopiranosa 1.02267 <1.0000 <1.0000
Fruktofuranosida <1.0000 1.34226 <1.0000
Galaktopiranosa 1.08147 <1.0000 1.22018
Glikosida <1.0000 1.04245 <1.0000
Gliserol 1.06717 <1.0000 <1.0000
Inositol <1.0000 1.04002 <1.0000
Isoxantopterin <1.0000 1.00741 <1.0000
L-Fukosa <1.0000 1.31399 1.25727
L-Mannopiranosa 1.2207 <1.0000 <1.0000
Myo-Inositol <1.0000 1.26765 1.71162
Pentitol <1.0000 <1.0000 1.16014
Timol-α-D-glukopiranosida 1.55706 <1.0000 <1.0000
Xilitol <1.0000 1.24333 <1.0000
Hasil analisis VIP diperoleh melalui analisis PLS-DA yang dijalankan secara
berasingan bagi tisu perikarpa, isi dan biji manggis
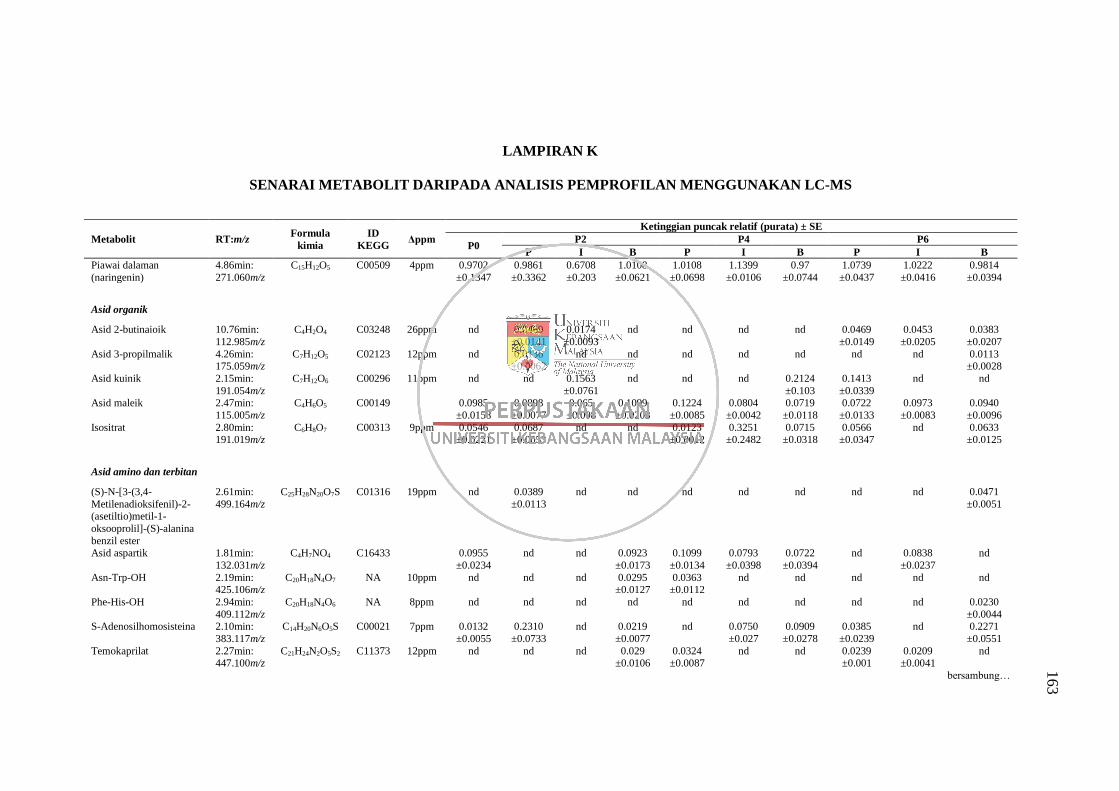
163
LAMPIRAN K
SENARAI METABOLIT DARIPADA ANALISIS PEMPROFILAN MENGGUNAKAN LC-MS
Metabolit RT:m/z Formula
kimia
ID
KEGG Δppm
Ketinggian puncak relatif (purata) ± SE
P0 P2 P4 P6
P I B P I B P I B
Piawai dalaman
(naringenin)
4.86min:
271.060m/z
C15H12O5 C00509 4ppm 0.9702
±0.1347
0.9861
±0.3362
0.6708
±0.203
1.0108
±0.0621
1.0108
±0.0698
1.1399
±0.0106
0.97
±0.0744
1.0739
±0.0437
1.0222
±0.0416
0.9814
±0.0394
Asid organik
Asid 2-butinaioik 10.76min:
112.985m/z
C4H2O4 C03248 26ppm nd 0.0469
±0.0141
0.0174
±0.0093
nd nd nd nd 0.0469
±0.0149
0.0453
±0.0205
0.0383
±0.0207
Asid 3-propilmalik 4.26min: 175.059m/z
C7H12O5 C02123 12ppm nd 0.0136 ±0.0062
nd nd nd nd nd nd nd 0.0113 ±0.0028
Asid kuinik 2.15min:
191.054m/z
C7H12O6 C00296 11ppm nd nd 0.1563
±0.0761
nd nd nd 0.2124
±0.103
0.1413
±0.0339
nd nd
Asid maleik 2.47min:
115.005m/z
C4H6O5 C00149
0.0985
±0.0158
0.0898
±0.0077
0.065
±0.008
0.1099
±0.0203
0.1224
±0.0085
0.0804
±0.0042
0.0719
±0.0118
0.0722
±0.0133
0.0973
±0.0083
0.0940
±0.0096
Isositrat 2.80min: 191.019m/z
C6H8O7 C00313 9ppm 0.0546 ±0.0221
0.0687 ±0.0053
nd nd 0.0123 ±0.0012
0.3251 ±0.2482
0.0715 ±0.0318
0.0566 ±0.0347
nd 0.0633 ±0.0125
Asid amino dan terbitan
(S)-N-[3-(3,4-
Metilenadioksifenil)-2-(asetiltio)metil-1-
oksooprolil]-(S)-alanina
benzil ester
2.61min:
499.164m/z
C25H28N20O7S C01316 19ppm nd 0.0389
±0.0113
nd nd nd nd nd nd nd 0.0471
±0.0051
Asid aspartik 1.81min:
132.031m/z
C4H7NO4 C16433
0.0955
±0.0234
nd nd 0.0923
±0.0173
0.1099
±0.0134
0.0793
±0.0398
0.0722
±0.0394
nd 0.0838
±0.0237
nd
Asn-Trp-OH 2.19min: 425.106m/z
C20H18N4O7 NA 10ppm nd nd nd 0.0295 ±0.0127
0.0363 ±0.0112
nd nd nd nd nd
Phe-His-OH 2.94min:
409.112m/z
C20H18N4O6 NA 8ppm nd nd nd nd nd nd nd nd nd 0.0230
±0.0044 S-Adenosilhomosisteina 2.10min:
383.117m/z
C14H20N6O5S C00021 7ppm 0.0132
±0.0055
0.2310
±0.0733
nd 0.0219
±0.0077
nd 0.0750
±0.027
0.0909
±0.0278
0.0385
±0.0239
nd 0.2271
±0.0551
Temokaprilat 2.27min: 447.100m/z
C21H24N2O5S2 C11373 12ppm nd nd nd 0.029 ±0.0106
0.0324 ±0.0087
nd nd 0.0239 ±0.001
0.0209 ±0.0041
nd
bersambung…
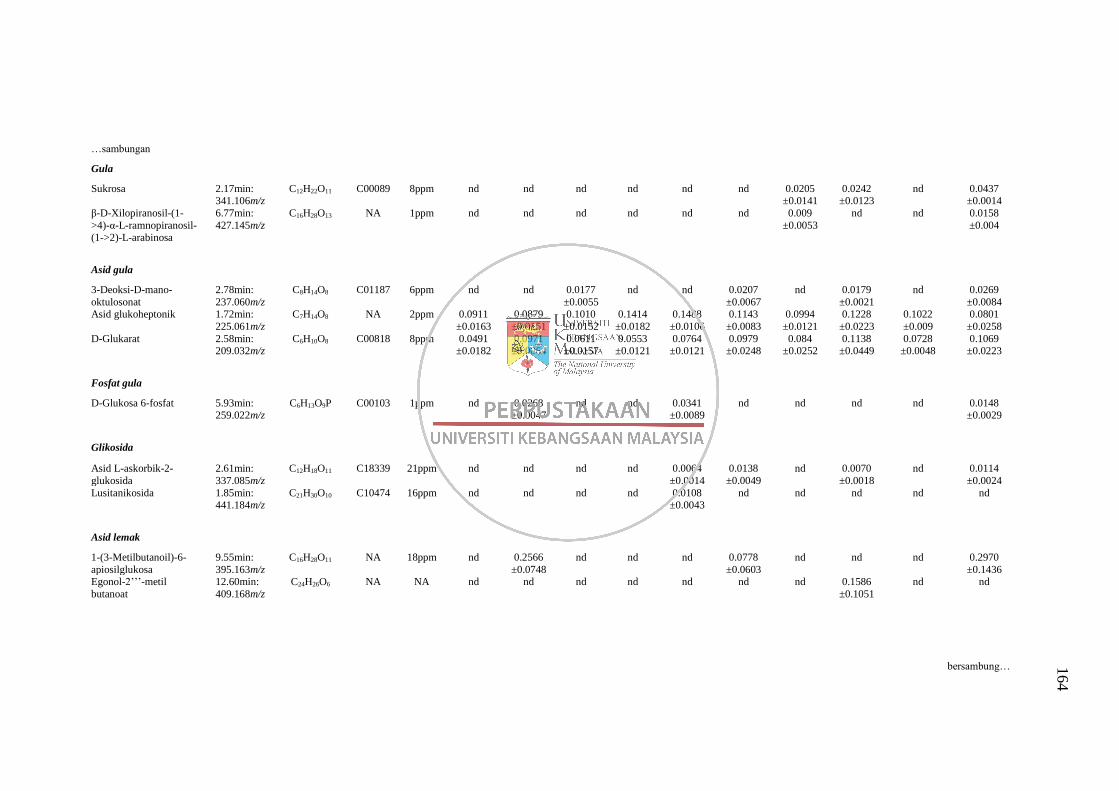
164
…sambungan
Gula
Sukrosa 2.17min: 341.106m/z
C12H22O11 C00089 8ppm nd nd nd nd nd nd 0.0205 ±0.0141
0.0242 ±0.0123
nd 0.0437 ±0.0014
β-D-Xilopiranosil-(1-
>4)-α-L-ramnopiranosil-
(1->2)-L-arabinosa
6.77min:
427.145m/z
C16H28O13 NA 1ppm nd nd nd nd nd nd 0.009
±0.0053
nd nd 0.0158
±0.004
Asid gula
3-Deoksi-D-mano-
oktulosonat
2.78min:
237.060m/z
C8H14O8 C01187 6ppm nd nd 0.0177
±0.0055
nd nd 0.0207
±0.0067
nd 0.0179
±0.0021
nd 0.0269
±0.0084 Asid glukoheptonik 1.72min:
225.061m/z
C7H14O8 NA 2ppm 0.0911
±0.0163
0.0879
±0.0151
0.1010
±0.0152
0.1414
±0.0182
0.1488
±0.0106
0.1143
±0.0083
0.0994
±0.0121
0.1228
±0.0223
0.1022
±0.009
0.0801
±0.0258
D-Glukarat 2.58min: 209.032m/z
C6H10O8 C00818 8ppm 0.0491 ±0.0182
0.0971 ±0.0064
0.0611 ±0.0157
0.0553 ±0.0121
0.0764 ±0.0121
0.0979 ±0.0248
0.084 ±0.0252
0.1138 ±0.0449
0.0728 ±0.0048
0.1069 ±0.0223
Fosfat gula
D-Glukosa 6-fosfat 5.93min: 259.022m/z
C6H13O9P C00103 1ppm nd 0.0268 ±0.0047
nd nd 0.0341 ±0.0089
nd nd nd nd 0.0148 ±0.0029
Glikosida
Asid L-askorbik-2-
glukosida
2.61min:
337.085m/z
C12H18O11 C18339 21ppm nd nd nd nd 0.0064
±0.0014
0.0138
±0.0049
nd 0.0070
±0.0018
nd 0.0114
±0.0024
Lusitanikosida 1.85min: 441.184m/z
C21H30O10 C10474 16ppm nd nd nd nd 0.0108 ±0.0043
nd nd nd nd nd
Asid lemak
1-(3-Metilbutanoil)-6-
apiosilglukosa
9.55min:
395.163m/z
C16H28O11 NA 18ppm nd 0.2566
±0.0748
nd nd nd 0.0778
±0.0603
nd nd nd 0.2970
±0.1436 Egonol-2ʼʼʼ-metil
butanoat
12.60min:
409.168m/z
C24H26O6 NA NA nd nd nd nd nd nd nd 0.1586
±0.1051
nd nd
bersambung…
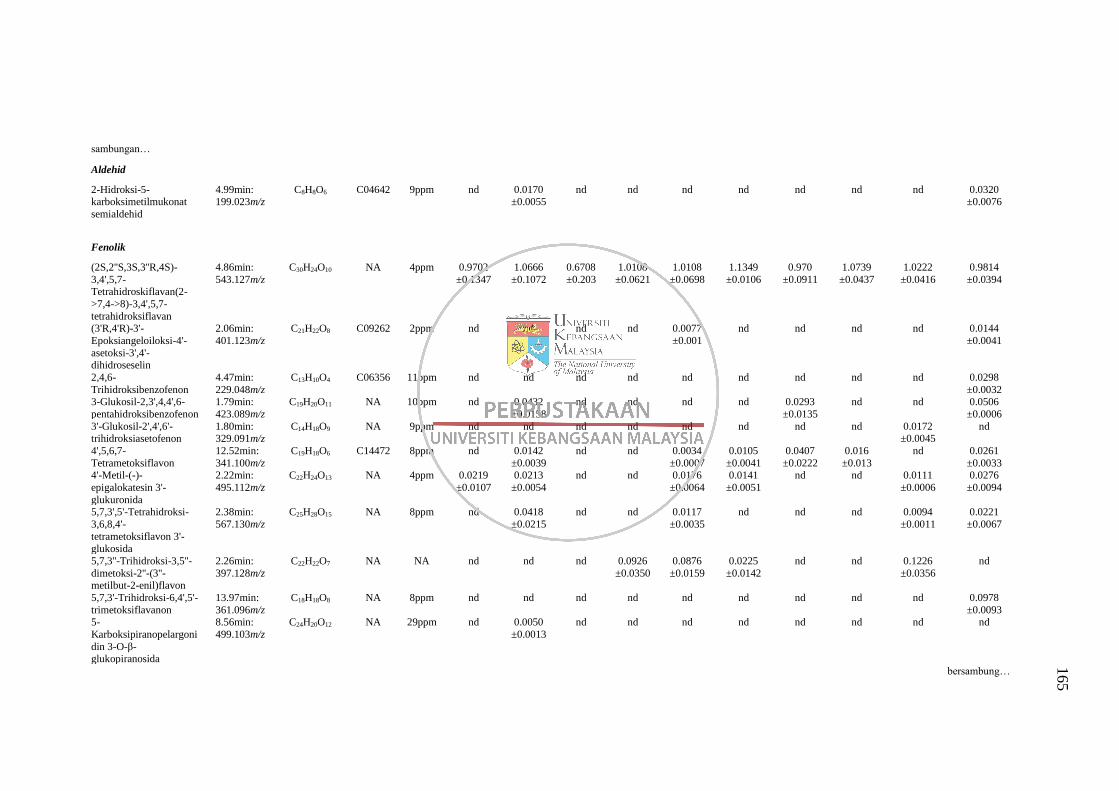
165
sambungan…
Aldehid
2-Hidroksi-5-karboksimetilmukonat
semialdehid
4.99min: 199.023m/z
C8H8O6 C04642 9ppm nd 0.0170 ±0.0055
nd nd nd nd nd nd nd 0.0320 ±0.0076
Fenolik
(2S,2''S,3S,3''R,4S)-
3,4',5,7-Tetrahidroskiflavan(2-
>7,4->8)-3,4',5,7-
tetrahidroksiflavan
4.86min:
543.127m/z
C30H24O10 NA 4ppm 0.9702
±0.1347
1.0666
±0.1072
0.6708
±0.203
1.0108
±0.0621
1.0108
±0.0698
1.1349
±0.0106
0.970
±0.0911
1.0739
±0.0437
1.0222
±0.0416
0.9814
±0.0394
(3'R,4'R)-3'-
Epoksiangeloiloksi-4'-
asetoksi-3',4'-dihidroseselin
2.06min:
401.123m/z
C21H22O8 C09262 2ppm nd nd nd nd 0.0077
±0.001
nd nd nd nd 0.0144
±0.0041
2,4,6-
Trihidroksibenzofenon
4.47min:
229.048m/z
C13H10O4 C06356 11ppm nd nd nd nd nd nd nd nd nd 0.0298
±0.0032 3-Glukosil-2,3',4,4',6-
pentahidroksibenzofenon
1.79min:
423.089m/z
C19H20O11 NA 10ppm nd 0.0432
±0.0158
nd nd nd nd 0.0293
±0.0135
nd nd 0.0506
±0.0006
3'-Glukosil-2',4',6'-trihidroksiasetofenon
1.80min: 329.091m/z
C14H18O9 NA 9ppm nd nd nd nd nd nd nd nd 0.0172 ±0.0045
nd
4',5,6,7-
Tetrametoksiflavon
12.52min:
341.100m/z
C19H18O6 C14472 8ppm nd 0.0142
±0.0039
nd nd 0.0034
±0.0007
0.0105
±0.0041
0.0407
±0.0222
0.016
±0.013
nd 0.0261
±0.0033 4'-Metil-(-)-
epigalokatesin 3'-
glukuronida
2.22min:
495.112m/z
C22H24O13 NA 4ppm 0.0219
±0.0107
0.0213
±0.0054
nd nd 0.0176
±0.0064
0.0141
±0.0051
nd nd 0.0111
±0.0006
0.0276
±0.0094
5,7,3',5'-Tetrahidroksi-
3,6,8,4'-
tetrametoksiflavon 3'-
glukosida
2.38min:
567.130m/z
C25H28O15 NA 8ppm nd 0.0418
±0.0215
nd nd 0.0117
±0.0035
nd nd nd 0.0094
±0.0011
0.0221
±0.0067
5,7,3''-Trihidroksi-3,5''-
dimetoksi-2''-(3''-metilbut-2-enil)flavon
2.26min:
397.128m/z
C22H22O7 NA NA nd nd nd 0.0926
±0.0350
0.0876
±0.0159
0.0225
±0.0142
nd nd 0.1226
±0.0356
nd
5,7,3'-Trihidroksi-6,4',5'-
trimetoksiflavanon
13.97min:
361.096m/z
C18H18O8 NA 8ppm nd nd nd nd nd nd nd nd nd 0.0978
±0.0093 5-
Karboksipiranopelargoni
din 3-O-β-glukopiranosida
8.56min:
499.103m/z
C24H20O12 NA 29ppm nd 0.0050
±0.0013
nd nd nd nd nd nd nd nd
bersambung…
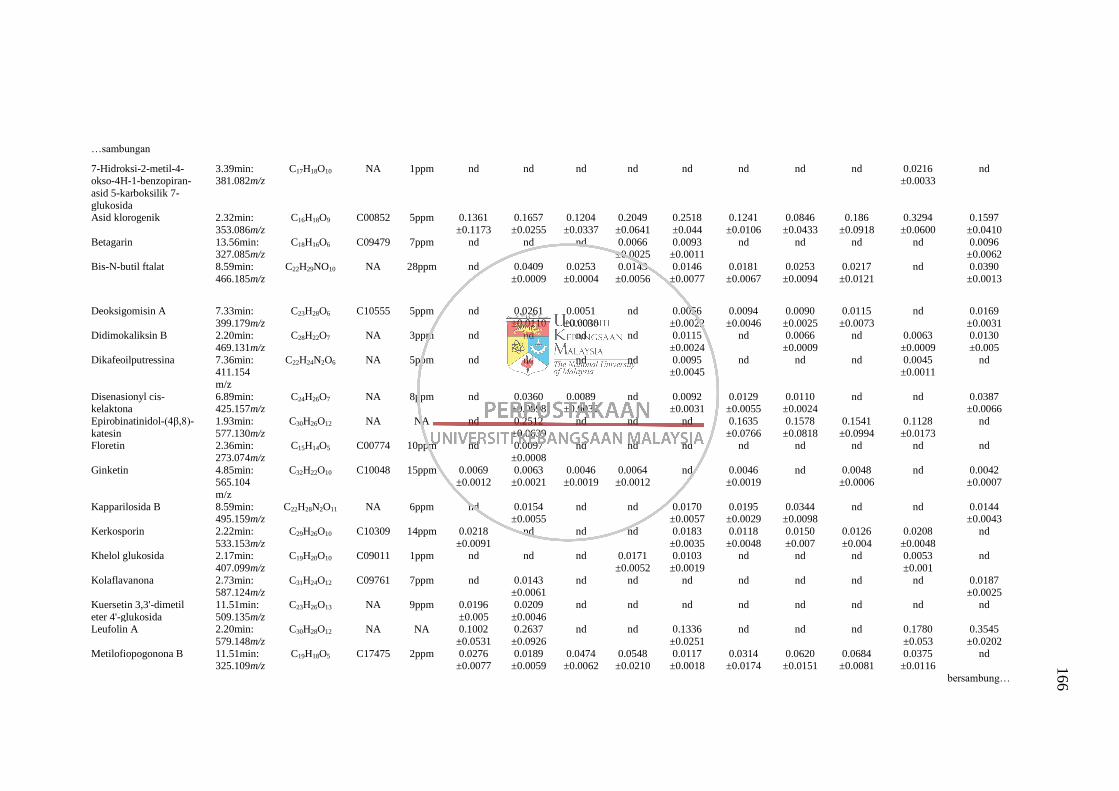
166
…sambungan
7-Hidroksi-2-metil-4-okso-4H-1-benzopiran-
asid 5-karboksilik 7-
glukosida
3.39min: 381.082m/z
C17H18O10 NA 1ppm nd nd nd nd nd nd nd nd 0.0216 ±0.0033
nd
Asid klorogenik 2.32min:
353.086m/z
C16H18O9 C00852 5ppm 0.1361
±0.1173
0.1657
±0.0255
0.1204
±0.0337
0.2049
±0.0641
0.2518
±0.044
0.1241
±0.0106
0.0846
±0.0433
0.186
±0.0918
0.3294
±0.0600
0.1597
±0.0410
Betagarin 13.56min: 327.085m/z
C18H16O6 C09479 7ppm nd nd nd 0.0066 ±0.0025
0.0093 ±0.0011
nd nd nd nd 0.0096 ±0.0062
Bis-N-butil ftalat 8.59min:
466.185m/z
C22H29NO10 NA 28ppm nd 0.0409
±0.0009
0.0253
±0.0004
0.0143
±0.0056
0.0146
±0.0077
0.0181
±0.0067
0.0253
±0.0094
0.0217
±0.0121
nd 0.0390
±0.0013
Deoksigomisin A 7.33min:
399.179m/z
C23H28O6 C10555 5ppm nd 0.0261
±0.0110
0.0051
±0.0030
nd 0.0056
±0.0022
0.0094
±0.0046
0.0090
±0.0025
0.0115
±0.0073
nd 0.0169
±0.0031 Didimokaliksin B 2.20min:
469.131m/z
C28H22O7 NA 3ppm nd nd nd nd 0.0115
±0.0024
nd 0.0066
±0.0009
nd 0.0063
±0.0009
0.0130
±0.005
Dikafeoilputressina 7.36min: 411.154
m/z
C22H24N2O6 NA 5ppm nd nd nd nd 0.0095 ±0.0045
nd nd nd 0.0045 ±0.0011
nd
Disenasionyl cis-kelaktona
6.89min: 425.157m/z
C24H26O7 NA 8ppm nd 0.0360 ±0.0098
0.0089 ±0.0032
nd 0.0092 ±0.0031
0.0129 ±0.0055
0.0110 ±0.0024
nd nd 0.0387 ±0.0066
Epirobinatinidol-(4β,8)-
katesin
1.93min:
577.130m/z
C30H26O12 NA NA nd 0.2512
±0.0639
nd nd nd 0.1635
±0.0766
0.1578
±0.0818
0.1541
±0.0994
0.1128
±0.0173
nd
Floretin 2.36min:
273.074m/z
C15H14O5 C00774 10ppm nd 0.0097
±0.0008
nd nd nd nd nd nd nd nd
Ginketin 4.85min: 565.104
m/z
C32H22O10 C10048 15ppm 0.0069 ±0.0012
0.0063 ±0.0021
0.0046 ±0.0019
0.0064 ±0.0012
nd 0.0046 ±0.0019
nd 0.0048 ±0.0006
nd 0.0042 ±0.0007
Kapparilosida B 8.59min: 495.159m/z
C22H28N2O11 NA 6ppm nd 0.0154 ±0.0055
nd nd 0.0170 ±0.0057
0.0195 ±0.0029
0.0344 ±0.0098
nd nd 0.0144 ±0.0043
Kerkosporin 2.22min:
533.153m/z
C29H26O10 C10309 14ppm 0.0218
±0.0091
nd nd nd 0.0183
±0.0035
0.0118
±0.0048
0.0150
±0.007
0.0126
±0.004
0.0208
±0.0048
nd
Khelol glukosida 2.17min:
407.099m/z
C19H20O10 C09011 1ppm nd nd nd 0.0171
±0.0052
0.0103
±0.0019
nd nd nd 0.0053
±0.001
nd
Kolaflavanona 2.73min: 587.124m/z
C31H24O12 C09761 7ppm nd 0.0143 ±0.0061
nd nd nd nd nd nd nd 0.0187 ±0.0025
Kuersetin 3,3'-dimetil
eter 4'-glukosida
11.51min:
509.135m/z
C23H26O13 NA 9ppm 0.0196
±0.005
0.0209
±0.0046
nd nd nd nd nd nd nd nd
Leufolin A 2.20min:
579.148m/z
C30H28O12 NA NA 0.1002
±0.0531
0.2637
±0.0926
nd nd 0.1336
±0.0251
nd nd nd 0.1780
±0.053
0.3545
±0.0202
Metilofiopogonona B 11.51min: 325.109m/z
C19H18O5 C17475 2ppm 0.0276 ±0.0077
0.0189 ±0.0059
0.0474 ±0.0062
0.0548 ±0.0210
0.0117 ±0.0018
0.0314 ±0.0174
0.0620 ±0.0151
0.0684 ±0.0081
0.0375 ±0.0116
nd
bersambung…
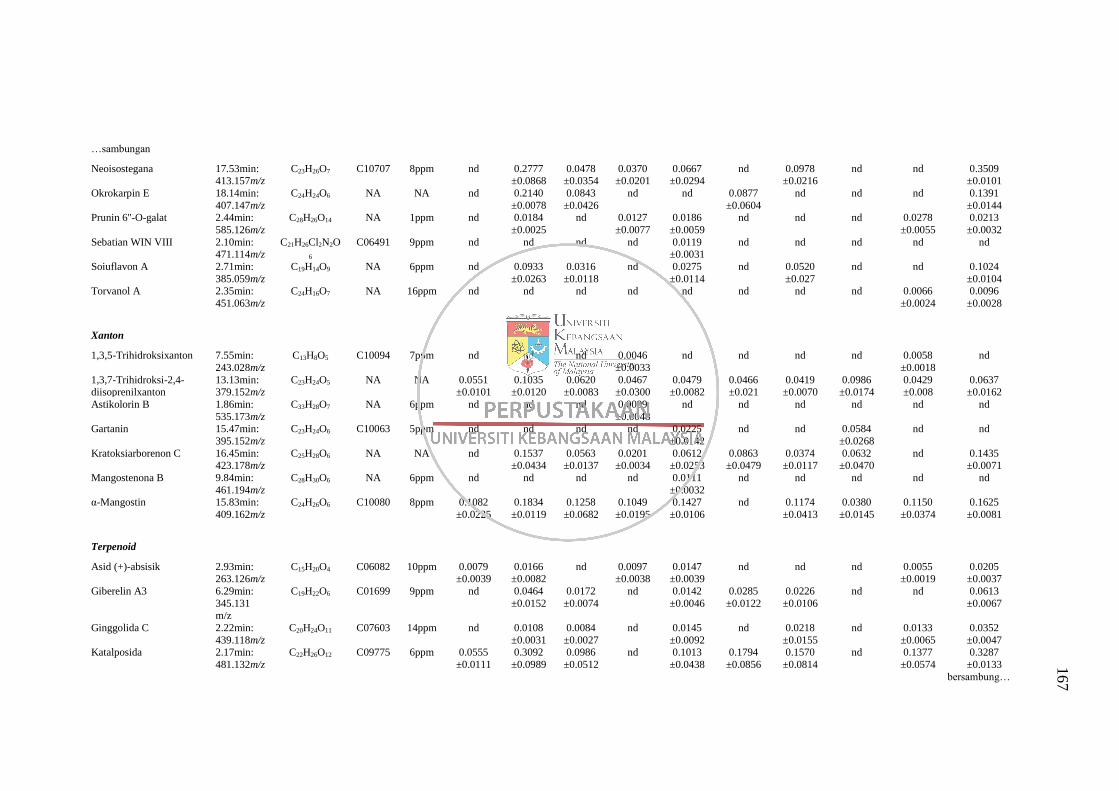
167
…sambungan
Neoisostegana 17.53min: 413.157m/z
C23H26O7 C10707 8ppm nd 0.2777 ±0.0868
0.0478 ±0.0354
0.0370 ±0.0201
0.0667 ±0.0294
nd 0.0978 ±0.0216
nd nd 0.3509 ±0.0101
Okrokarpin E 18.14min:
407.147m/z
C24H24O6 NA NA nd 0.2140
±0.0078
0.0843
±0.0426
nd nd 0.0877
±0.0604
nd nd nd 0.1391
±0.0144 Prunin 6''-O-galat 2.44min:
585.126m/z
C28H26O14 NA 1ppm nd 0.0184
±0.0025
nd 0.0127
±0.0077
0.0186
±0.0059
nd nd nd 0.0278
±0.0055
0.0213
±0.0032
Sebatian WIN VIII 2.10min: 471.114m/z
C21H26Cl2N2O
6
C06491 9ppm nd nd nd nd 0.0119 ±0.0031
nd nd nd nd nd
Soiuflavon A 2.71min:
385.059m/z
C19H14O9 NA 6ppm nd 0.0933
±0.0263
0.0316
±0.0118
nd 0.0275
±0.0114
nd 0.0520
±0.027
nd nd 0.1024
±0.0104 Torvanol A 2.35min:
451.063m/z
C24H16O7 NA 16ppm nd nd nd nd nd nd nd nd 0.0066
±0.0024
0.0096
±0.0028
Xanton
1,3,5-Trihidroksixanton 7.55min:
243.028m/z
C13H8O5 C10094 7ppm nd nd nd 0.0046
±0.0033
nd nd nd nd 0.0058
±0.0018
nd
1,3,7-Trihidroksi-2,4-
diisoprenilxanton
13.13min:
379.152m/z C23H24O5 NA NA 0.0551
±0.0101
0.1035
±0.0120
0.0620
±0.0083
0.0467
±0.0300
0.0479
±0.0082
0.0466
±0.021
0.0419
±0.0070
0.0986
±0.0174
0.0429
±0.008
0.0637
±0.0162
Astikolorin B 1.86min:
535.173m/z
C33H28O7 NA 6ppm nd nd nd 0.0099
±0.0048
nd nd nd nd nd nd
Gartanin 15.47min:
395.152m/z
C23H24O6 C10063 5ppm nd nd nd nd 0.0225
±0.0142
nd nd 0.0584
±0.0268
nd nd
Kratoksiarborenon C 16.45min: 423.178m/z
C25H28O6 NA NA nd 0.1537 ±0.0434
0.0563 ±0.0137
0.0201 ±0.0034
0.0612 ±0.0253
0.0863 ±0.0479
0.0374 ±0.0117
0.0632 ±0.0470
nd 0.1435 ±0.0071
Mangostenona B 9.84min:
461.194m/z
C28H30O6 NA 6ppm nd nd nd nd 0.0111
±0.0032
nd nd nd nd nd
α-Mangostin 15.83min:
409.162m/z
C24H26O6 C10080 8ppm 0.1082
±0.0225
0.1834
±0.0119
0.1258
±0.0682
0.1049
±0.0195
0.1427
±0.0106
nd 0.1174
±0.0413
0.0380
±0.0145
0.1150
±0.0374
0.1625
±0.0081
Terpenoid
Asid (+)-absisik 2.93min: 263.126m/z
C15H20O4 C06082 10ppm 0.0079 ±0.0039
0.0166 ±0.0082
nd 0.0097 ±0.0038
0.0147 ±0.0039
nd nd nd 0.0055 ±0.0019
0.0205 ±0.0037
Giberelin A3 6.29min:
345.131 m/z
C19H22O6 C01699 9ppm nd 0.0464
±0.0152
0.0172
±0.0074
nd 0.0142
±0.0046
0.0285
±0.0122
0.0226
±0.0106
nd nd 0.0613
±0.0067
Ginggolida C 2.22min:
439.118m/z
C20H24O11 C07603 14ppm nd 0.0108
±0.0031
0.0084
±0.0027
nd 0.0145
±0.0092
nd 0.0218
±0.0155
nd 0.0133
±0.0065
0.0352
±0.0047 Katalposida 2.17min:
481.132m/z
C22H26O12 C09775 6ppm 0.0555
±0.0111
0.3092
±0.0989
0.0986
±0.0512
nd 0.1013
±0.0438
0.1794
±0.0856
0.1570
±0.0814
nd 0.1377
±0.0574
0.3287
±0.0133
bersambung…
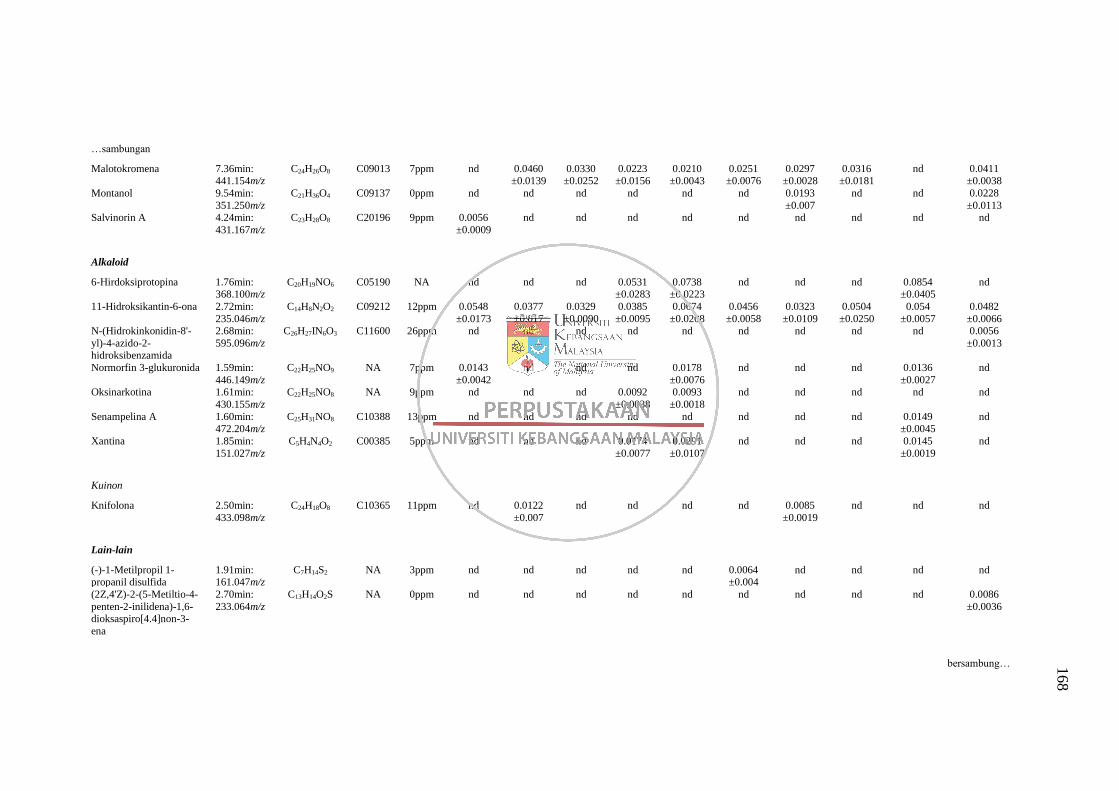
168
…sambungan
Malotokromena 7.36min: 441.154m/z
C24H26O8 C09013 7ppm nd 0.0460 ±0.0139
0.0330 ±0.0252
0.0223 ±0.0156
0.0210 ±0.0043
0.0251 ±0.0076
0.0297 ±0.0028
0.0316 ±0.0181
nd 0.0411 ±0.0038
Montanol 9.54min:
351.250m/z
C21H36O4 C09137 0ppm nd nd nd nd nd nd 0.0193
±0.007
nd nd 0.0228
±0.0113 Salvinorin A 4.24min:
431.167m/z
C23H28O8 C20196 9ppm 0.0056
±0.0009
nd nd nd nd nd nd nd nd nd
Alkaloid
6-Hirdoksiprotopina 1.76min: 368.100m/z
C20H19NO6 C05190 NA nd nd nd 0.0531 ±0.0283
0.0738 ±0.0223
nd nd nd 0.0854 ±0.0405
nd
11-Hidroksikantin-6-ona 2.72min:
235.046m/z
C14H8N2O2 C09212 12ppm 0.0548
±0.0173
0.0377
±0.017
0.0329
±0.0090
0.0385
±0.0095
0.0674
±0.0208
0.0456
±0.0058
0.0323
±0.0109
0.0504
±0.0250
0.054
±0.0057
0.0482
±0.0066 N-(Hidrokinkonidin-8'-
yl)-4-azido-2-hidroksibenzamida
2.68min:
595.096m/z
C26H27IN6O3 C11600 26ppm nd nd nd nd nd nd nd nd nd 0.0056
±0.0013
Normorfin 3-glukuronida 1.59min:
446.149m/z
C22H25NO9 NA 7ppm 0.0143
±0.0042
nd nd nd 0.0178
±0.0076
nd nd nd 0.0136
±0.0027
nd
Oksinarkotina 1.61min:
430.155m/z
C22H25NO8 NA 9ppm nd nd nd 0.0092
±0.0038
0.0093
±0.0018
nd nd nd nd nd
Senampelina A 1.60min: 472.204m/z
C25H31NO8 C10388 13ppm nd nd nd nd nd nd nd nd 0.0149 ±0.0045
nd
Xantina 1.85min:
151.027m/z
C5H4N4O2 C00385 5ppm nd nd nd 0.0174
±0.0077
0.0291
±0.0107
nd nd nd 0.0145
±0.0019
nd
Kuinon
Knifolona 2.50min:
433.098m/z
C24H18O8 C10365 11ppm nd 0.0122
±0.007
nd nd nd nd 0.0085
±0.0019
nd nd nd
Lain-lain
(-)-1-Metilpropil 1-propanil disulfida
1.91min: 161.047m/z
C7H14S2 NA 3ppm nd nd nd nd nd 0.0064 ±0.004
nd nd nd nd
(2Z,4'Z)-2-(5-Metiltio-4-
penten-2-inilidena)-1,6-dioksaspiro[4.4]non-3-
ena
2.70min:
233.064m/z
C13H14O2S NA 0ppm nd nd nd nd nd nd nd nd nd 0.0086
±0.0036
bersambung…
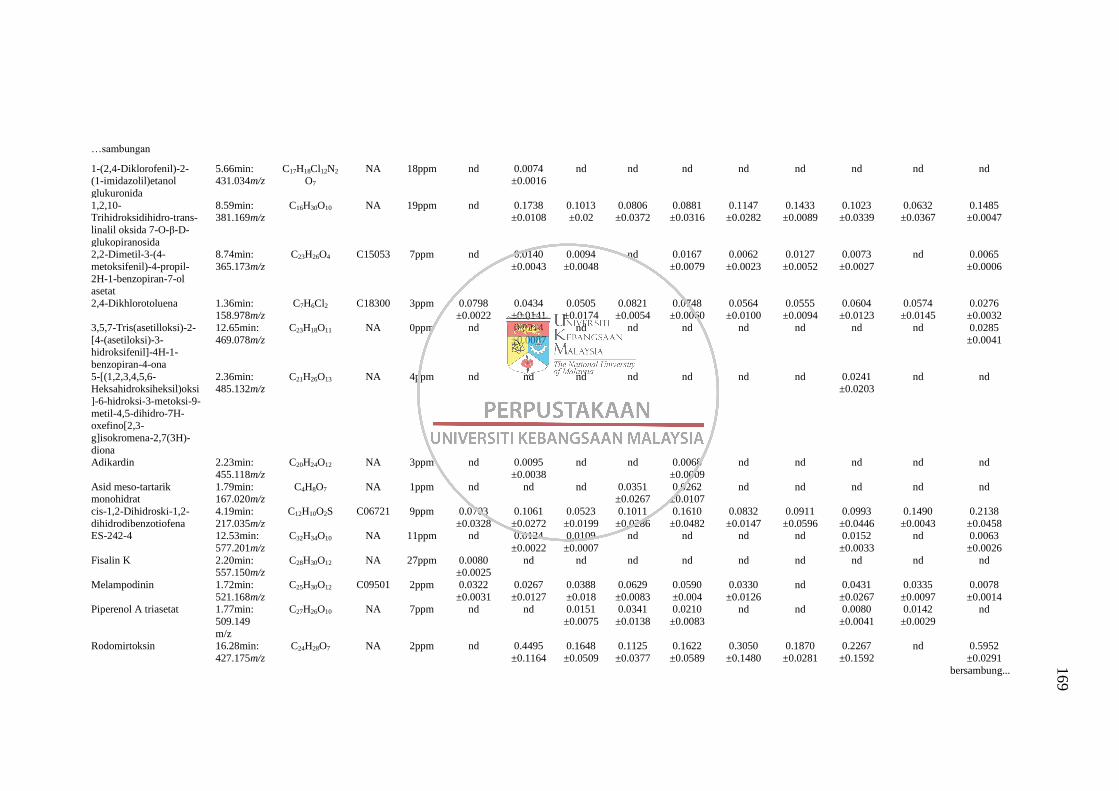
169
…sambungan
1-(2,4-Diklorofenil)-2-(1-imidazolil)etanol
glukuronida
5.66min: 431.034m/z
C17H18Cl12N2
O7
NA 18ppm nd 0.0074 ±0.0016
nd nd nd nd nd nd nd nd
1,2,10-Trihidroksidihidro-trans-
linalil oksida 7-O-β-D-
glukopiranosida
8.59min: 381.169m/z
C16H30O10 NA 19ppm nd 0.1738 ±0.0108
0.1013 ±0.02
0.0806 ±0.0372
0.0881 ±0.0316
0.1147 ±0.0282
0.1433 ±0.0089
0.1023 ±0.0339
0.0632 ±0.0367
0.1485 ±0.0047
2,2-Dimetil-3-(4-
metoksifenil)-4-propil-
2H-1-benzopiran-7-ol asetat
8.74min:
365.173m/z
C23H26O4 C15053 7ppm nd 0.0140
±0.0043
0.0094
±0.0048
nd 0.0167
±0.0079
0.0062
±0.0023
0.0127
±0.0052
0.0073
±0.0027
nd 0.0065
±0.0006
2,4-Dikhlorotoluena 1.36min: 158.978m/z
C7H6Cl2 C18300 3ppm 0.0798 ±0.0022
0.0434 ±0.0141
0.0505 ±0.0174
0.0821 ±0.0054
0.0748 ±0.0060
0.0564 ±0.0100
0.0555 ±0.0094
0.0604 ±0.0123
0.0574 ±0.0145
0.0276 ±0.0032
3,5,7-Tris(asetilloksi)-2-
[4-(asetiloksi)-3-hidroksifenil]-4H-1-
benzopiran-4-ona
12.65min:
469.078m/z
C23H18O11 NA 0ppm nd 0.0214
±0.0087
nd nd nd nd nd nd nd 0.0285
±0.0041
5-[(1,2,3,4,5,6-
Heksahidroksiheksil)oksi
]-6-hidroksi-3-metoksi-9-
metil-4,5-dihidro-7H-oxefino[2,3-
g]isokromena-2,7(3H)-
diona
2.36min:
485.132m/z C21H26O13 NA 4ppm nd nd nd nd nd nd nd 0.0241
±0.0203
nd nd
Adikardin 2.23min:
455.118m/z
C20H24O12 NA 3ppm nd 0.0095
±0.0038
nd nd 0.0068
±0.0009
nd nd nd nd nd
Asid meso-tartarik monohidrat
1.79min: 167.020m/z
C4H8O7 NA 1ppm nd nd nd 0.0351 ±0.0267
0.0262 ±0.0107
nd nd nd nd nd
cis-1,2-Dihidroski-1,2-
dihidrodibenzotiofena
4.19min:
217.035m/z
C12H10O2S C06721 9ppm 0.0703
±0.0328
0.1061
±0.0272
0.0523
±0.0199
0.1011
±0.0286
0.1610
±0.0482
0.0832
±0.0147
0.0911
±0.0596
0.0993
±0.0446
0.1490
±0.0043
0.2138
±0.0458 ES-242-4 12.53min:
577.201m/z
C32H34O10 NA 11ppm nd 0.0124
±0.0022
0.0109
±0.0007
nd nd nd nd 0.0152
±0.0033
nd 0.0063
±0.0026
Fisalin K 2.20min: 557.150m/z
C28H30O12 NA 27ppm 0.0080 ±0.0025
nd nd nd nd nd nd nd nd nd
Melampodinin 1.72min:
521.168m/z
C25H30O12
C09501 2ppm 0.0322
±0.0031
0.0267
±0.0127
0.0388
±0.018
0.0629
±0.0083
0.0590
±0.004
0.0330
±0.0126
nd 0.0431
±0.0267
0.0335
±0.0097
0.0078
±0.0014 Piperenol A triasetat 1.77min:
509.149
m/z
C27H26O10 NA 7ppm nd nd 0.0151
±0.0075
0.0341
±0.0138
0.0210
±0.0083
nd nd 0.0080
±0.0041
0.0142
±0.0029
nd
Rodomirtoksin 16.28min:
427.175m/z
C24H28O7 NA 2ppm nd 0.4495
±0.1164
0.1648
±0.0509
0.1125
±0.0377
0.1622
±0.0589
0.3050
±0.1480
0.1870
±0.0281
0.2267
±0.1592
nd 0.5952
±0.0291
bersambung...
170
...sambungan
Rubriflordilakton B 12.10min: 461.201m/z
C28H30O6 NA NA nd nd nd nd nd nd nd 0.0668 ±0.0331
nd nd
S-(2,5-Dimetil-3-furanil)
2-furankarbotioat
3.78min:
221.028m/z
C11H10O3S NA 0ppm nd 0.0254
±0.0156
nd nd nd nd nd nd nd 0.0273
±0.0088 Sativanina B 12.58min:
517.293m/z
C30H38N4O4 C10013 21ppm nd 0.0093
±0.0027
nd nd nd nd nd nd nd nd
Singkatan; NA: not available, nd: not detected, RT:m/z: masa pengekalan/nisbah jisim kepada caj, Δppm: ralat jisim molekul, SE: ralat piawai,
P0: peringkat 0, P2: peringkat 2, P4: peringkat 4, P6: peringkat 6, P: perikarpa, I: isi dan B: biji
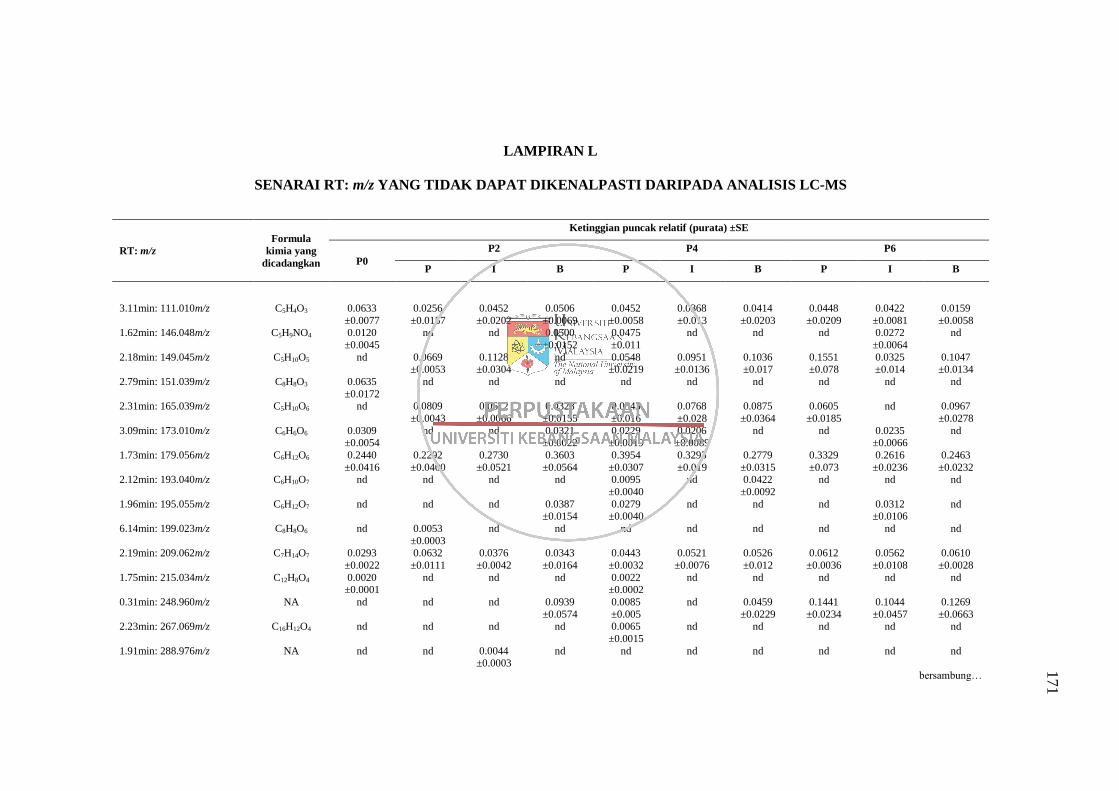
171
LAMPIRAN L
SENARAI RT: m/z YANG TIDAK DAPAT DIKENALPASTI DARIPADA ANALISIS LC-MS
RT: m/z
Formula
kimia yang
dicadangkan
Ketinggian puncak relatif (purata) ±SE
P0
P2 P4 P6
P I B P I B P I B
3.11min: 111.010m/z C5H4O3 0.0633
±0.0077
0.0256
±0.0157
0.0452
±0.0202
0.0506
±0.0069
0.0452
±0.0058
0.0368
±0.013
0.0414
±0.0203
0.0448
±0.0209
0.0422
±0.0081
0.0159
±0.0058 1.62min: 146.048m/z C5H9NO4 0.0120
±0.0045
nd nd 0.0500
±0.0152
0.0475
±0.011
nd nd nd 0.0272
±0.0064
nd
2.18min: 149.045m/z C5H10O5 nd 0.0669 ±0.0053
0.1128 ±0.0304
nd 0.0548 ±0.0219
0.0951 ±0.0136
0.1036 ±0.017
0.1551 ±0.078
0.0325 ±0.014
0.1047 ±0.0134
2.79min: 151.039m/z C8H8O3 0.0635
±0.0172
nd nd nd nd nd nd nd nd nd
2.31min: 165.039m/z C5H10O6 nd 0.0809
±0.0043
0.0612
±0.0066
0.0328
±0.0155
0.0543
±0.016
0.0768
±0.028
0.0875
±0.0364
0.0605
±0.0185
nd 0.0967
±0.0278
3.09min: 173.010m/z C6H6O6 0.0309 ±0.0054
nd nd 0.0321 ±0.0022
0.0229 ±0.0019
0.0206 ±0.0089
nd nd 0.0235 ±0.0066
nd
1.73min: 179.056m/z C6H12O6 0.2440
±0.0416
0.2292
±0.0400
0.2730
±0.0521
0.3603
±0.0564
0.3954
±0.0307
0.3293
±0.019
0.2779
±0.0315
0.3329
±0.073
0.2616
±0.0236
0.2463
±0.0232 2.12min: 193.040m/z C6H10O7 nd nd nd nd 0.0095
±0.0040
nd 0.0422
±0.0092
nd nd nd
1.96min: 195.055m/z C6H12O7 nd nd nd 0.0387 ±0.0154
0.0279 ±0.0040
nd nd nd 0.0312 ±0.0106
nd
6.14min: 199.023m/z C8H8O6 nd 0.0053
±0.0003
nd nd nd nd nd nd nd nd
2.19min: 209.062m/z C7H14O7 0.0293
±0.0022
0.0632
±0.0111
0.0376
±0.0042
0.0343
±0.0164
0.0443
±0.0032
0.0521
±0.0076
0.0526
±0.012
0.0612
±0.0036
0.0562
±0.0108
0.0610
±0.0028
1.75min: 215.034m/z C12H8O4 0.0020 ±0.0001
nd nd nd 0.0022 ±0.0002
nd nd nd nd nd
0.31min: 248.960m/z NA nd nd nd 0.0939
±0.0574
0.0085
±0.005
nd 0.0459
±0.0229
0.1441
±0.0234
0.1044
±0.0457
0.1269
±0.0663 2.23min: 267.069m/z C16H12O4 nd nd nd nd 0.0065
±0.0015
nd nd nd nd nd
1.91min: 288.976m/z NA nd nd 0.0044 ±0.0003
nd nd nd nd nd nd nd
bersambung…
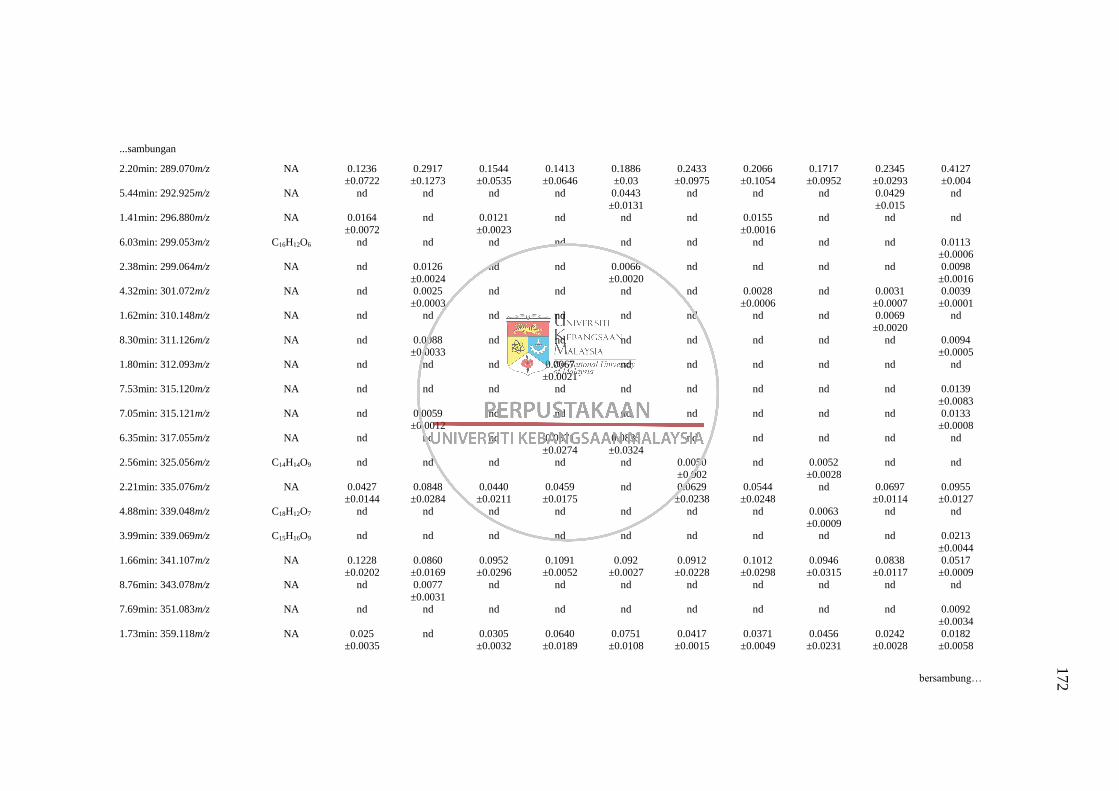
172
...sambungan
2.20min: 289.070m/z NA 0.1236 ±0.0722
0.2917 ±0.1273
0.1544 ±0.0535
0.1413 ±0.0646
0.1886 ±0.03
0.2433 ±0.0975
0.2066 ±0.1054
0.1717 ±0.0952
0.2345 ±0.0293
0.4127 ±0.004
5.44min: 292.925m/z NA nd nd nd nd 0.0443
±0.0131
nd nd nd 0.0429
±0.015
nd
1.41min: 296.880m/z NA 0.0164
±0.0072
nd 0.0121
±0.0023
nd nd nd 0.0155
±0.0016
nd nd nd
6.03min: 299.053m/z C16H12O6 nd nd nd nd nd nd nd nd nd 0.0113 ±0.0006
2.38min: 299.064m/z NA nd 0.0126
±0.0024
nd nd 0.0066
±0.0020
nd nd nd nd 0.0098
±0.0016 4.32min: 301.072m/z NA nd 0.0025
±0.0003
nd nd nd nd 0.0028
±0.0006
nd 0.0031
±0.0007
0.0039
±0.0001 1.62min: 310.148m/z NA nd nd nd nd nd nd nd nd 0.0069
±0.0020
nd
8.30min: 311.126m/z NA nd 0.0088 ±0.0033
nd nd nd nd nd nd nd 0.0094 ±0.0005
1.80min: 312.093m/z NA nd nd nd 0.0067
±0.0021
nd nd nd nd nd nd
7.53min: 315.120m/z NA nd nd nd nd nd nd nd nd nd 0.0139
±0.0083
7.05min: 315.121m/z NA nd 0.0059 ±0.0012
nd nd nd nd nd nd nd 0.0133 ±0.0008
6.35min: 317.055m/z NA nd nd nd 0.0871
±0.0274
0.0833
±0.0324
nd nd nd nd nd
2.56min: 325.056m/z C14H14O9 nd nd nd nd nd 0.0050
±0.002
nd 0.0052
±0.0028
nd nd
2.21min: 335.076m/z NA 0.0427 ±0.0144
0.0848 ±0.0284
0.0440 ±0.0211
0.0459 ±0.0175
nd 0.0629 ±0.0238
0.0544 ±0.0248
nd 0.0697 ±0.0114
0.0955 ±0.0127
4.88min: 339.048m/z C18H12O7 nd nd nd nd nd nd nd 0.0063
±0.0009
nd nd
3.99min: 339.069m/z C15H16O9 nd nd nd nd nd nd nd nd nd 0.0213
±0.0044
1.66min: 341.107m/z NA 0.1228 ±0.0202
0.0860 ±0.0169
0.0952 ±0.0296
0.1091 ±0.0052
0.092 ±0.0027
0.0912 ±0.0228
0.1012 ±0.0298
0.0946 ±0.0315
0.0838 ±0.0117
0.0517 ±0.0009
8.76min: 343.078m/z NA nd 0.0077
±0.0031
nd nd nd nd nd nd nd nd
7.69min: 351.083m/z NA nd nd nd nd nd nd nd nd nd 0.0092
±0.0034
1.73min: 359.118m/z NA 0.025 ±0.0035
nd 0.0305 ±0.0032
0.0640 ±0.0189
0.0751 ±0.0108
0.0417 ±0.0015
0.0371 ±0.0049
0.0456 ±0.0231
0.0242 ±0.0028
0.0182 ±0.0058
bersambung…
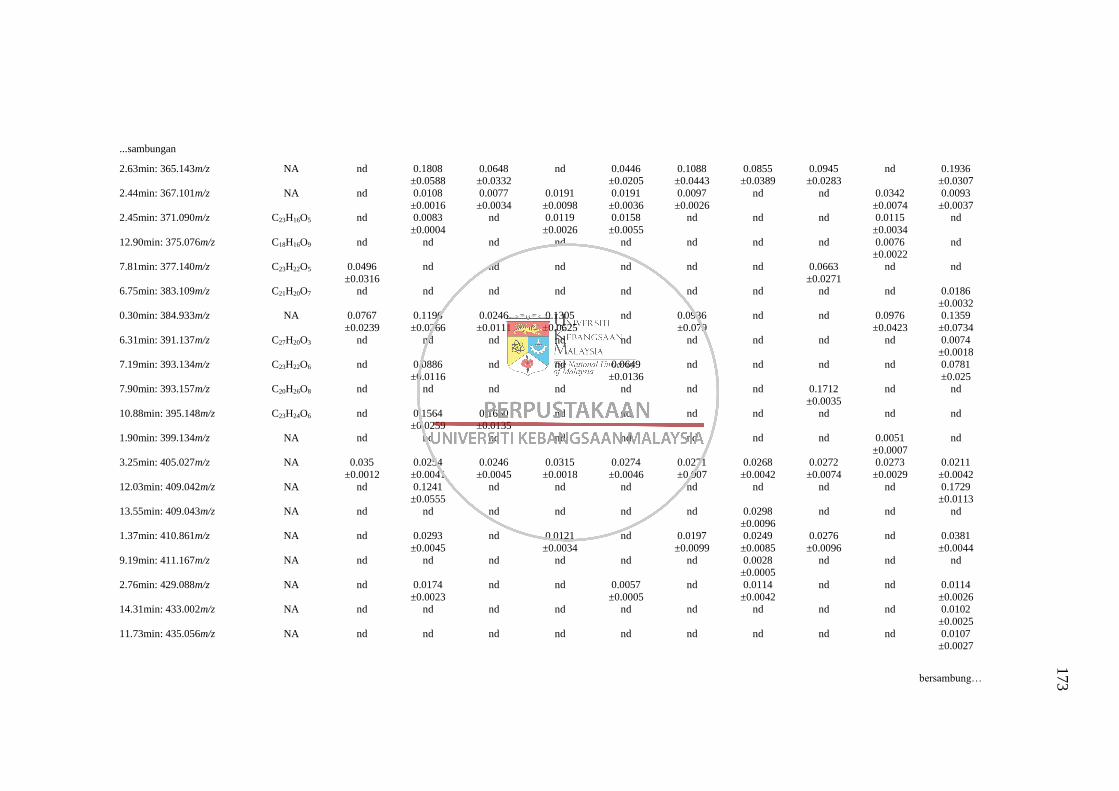
173
...sambungan
2.63min: 365.143m/z NA nd 0.1808 ±0.0588
0.0648 ±0.0332
nd 0.0446 ±0.0205
0.1088 ±0.0443
0.0855 ±0.0389
0.0945 ±0.0283
nd 0.1936 ±0.0307
2.44min: 367.101m/z NA nd 0.0108
±0.0016
0.0077
±0.0034
0.0191
±0.0098
0.0191
±0.0036
0.0097
±0.0026
nd nd 0.0342
±0.0074
0.0093
±0.0037 2.45min: 371.090m/z C23H16O5 nd 0.0083
±0.0004
nd 0.0119
±0.0026
0.0158
±0.0055
nd nd nd 0.0115
±0.0034
nd
12.90min: 375.076m/z C18H16O9 nd nd nd nd nd nd nd nd 0.0076 ±0.0022
nd
7.81min: 377.140m/z C23H22O5 0.0496
±0.0316
nd nd nd nd nd nd 0.0663
±0.0271
nd nd
6.75min: 383.109m/z C21H20O7 nd nd nd nd nd nd nd nd nd 0.0186
±0.0032 0.30min: 384.933m/z NA 0.0767
±0.0239
0.1196
±0.0266
0.0246
±0.0111
0.1305
±0.0625
nd 0.0936
±0.079
nd nd 0.0976
±0.0423
0.1359
±0.0734
6.31min: 391.137m/z C27H20O3 nd nd nd nd nd nd nd nd nd 0.0074 ±0.0018
7.19min: 393.134m/z C23H22O6 nd 0.0886
±0.0116
nd nd 0.0649
±0.0136
nd nd nd nd 0.0781
±0.025
7.90min: 393.157m/z C20H26O8 nd nd nd nd nd nd nd 0.1712
±0.0035
nd nd
10.88min: 395.148m/z C23H24O6 nd 0.1564 ±0.0259
0.1660 ±0.0135
nd nd nd nd nd nd nd
1.90min: 399.134m/z NA nd nd nd nd nd nd nd nd 0.0051
±0.0007
nd
3.25min: 405.027m/z NA 0.035
±0.0012
0.0254
±0.0041
0.0246
±0.0045
0.0315
±0.0018
0.0274
±0.0046
0.0271
±0.007
0.0268
±0.0042
0.0272
±0.0074
0.0273
±0.0029
0.0211
±0.0042
12.03min: 409.042m/z NA nd 0.1241 ±0.0555
nd nd nd nd nd nd nd 0.1729 ±0.0113
13.55min: 409.043m/z NA nd nd nd nd nd nd 0.0298
±0.0096
nd nd nd
1.37min: 410.861m/z NA nd 0.0293
±0.0045
nd 0.0121
±0.0034
nd 0.0197
±0.0099
0.0249
±0.0085
0.0276
±0.0096
nd 0.0381
±0.0044
9.19min: 411.167m/z NA nd nd nd nd nd nd 0.0028 ±0.0005
nd nd nd
2.76min: 429.088m/z NA nd 0.0174
±0.0023
nd nd 0.0057
±0.0005
nd 0.0114
±0.0042
nd nd 0.0114
±0.0026 14.31min: 433.002m/z NA nd nd nd nd nd nd nd nd nd 0.0102
±0.0025
11.73min: 435.056m/z NA nd nd nd nd nd nd nd nd nd 0.0107 ±0.0027
bersambung…
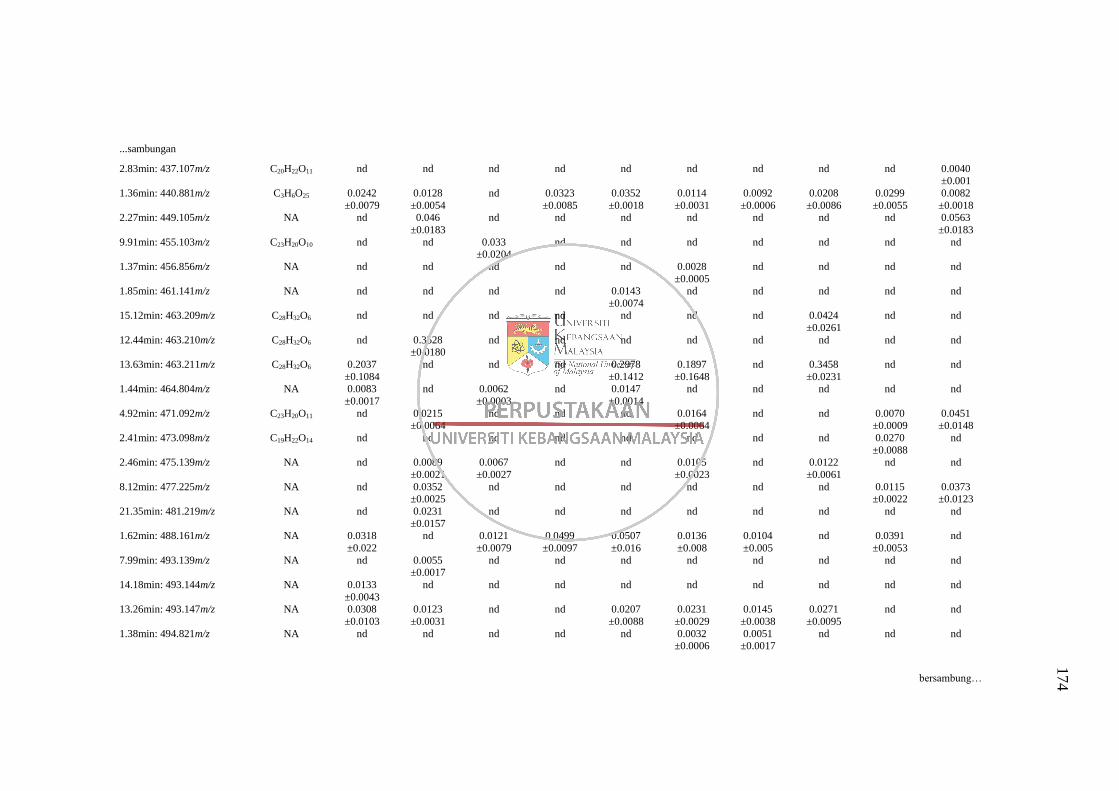
174
...sambungan
2.83min: 437.107m/z C20H22O11 nd nd nd nd nd nd nd nd nd 0.0040 ±0.001
1.36min: 440.881m/z C3H6O25 0.0242
±0.0079
0.0128
±0.0054
nd 0.0323
±0.0085
0.0352
±0.0018
0.0114
±0.0031
0.0092
±0.0006
0.0208
±0.0086
0.0299
±0.0055
0.0082
±0.0018 2.27min: 449.105m/z NA nd 0.046
±0.0183
nd nd nd nd nd nd nd 0.0563
±0.0183
9.91min: 455.103m/z C23H20O10 nd nd 0.033 ±0.0204
nd nd nd nd nd nd nd
1.37min: 456.856m/z NA nd nd nd nd nd 0.0028
±0.0005
nd nd nd nd
1.85min: 461.141m/z NA nd nd nd nd 0.0143
±0.0074
nd nd nd nd nd
15.12min: 463.209m/z C28H32O6 nd nd nd nd nd nd nd 0.0424
±0.0261
nd nd
12.44min: 463.210m/z C28H32O6 nd 0.3528 ±0.0180
nd nd nd nd nd nd nd nd
13.63min: 463.211m/z C28H32O6 0.2037
±0.1084
nd nd nd 0.2978
±0.1412
0.1897
±0.1648
nd 0.3458
±0.0231
nd nd
1.44min: 464.804m/z NA 0.0083
±0.0017
nd 0.0062
±0.0003
nd 0.0147
±0.0014
nd nd nd nd nd
4.92min: 471.092m/z C23H20O11 nd 0.0215 ±0.0064
nd nd nd 0.0164 ±0.0064
nd nd 0.0070 ±0.0009
0.0451 ±0.0148
2.41min: 473.098m/z C19H22O14 nd nd nd nd nd nd nd nd 0.0270
±0.0088
nd
2.46min: 475.139m/z NA nd 0.0089
±0.0021
0.0067
±0.0027
nd nd 0.0105
±0.0023
nd 0.0122
±0.0061
nd nd
8.12min: 477.225m/z NA nd 0.0352 ±0.0025
nd nd nd nd nd nd 0.0115 ±0.0022
0.0373 ±0.0123
21.35min: 481.219m/z NA nd 0.0231
±0.0157
nd nd nd nd nd nd nd nd
1.62min: 488.161m/z NA 0.0318
±0.022
nd 0.0121
±0.0079
0.0499
±0.0097
0.0507
±0.016
0.0136
±0.008
0.0104
±0.005
nd 0.0391
±0.0053
nd
7.99min: 493.139m/z NA nd 0.0055 ±0.0017
nd nd nd nd nd nd nd nd
14.18min: 493.144m/z NA 0.0133
±0.0043
nd nd nd nd nd nd nd nd nd
13.26min: 493.147m/z NA 0.0308
±0.0103
0.0123
±0.0031
nd nd 0.0207
±0.0088
0.0231
±0.0029
0.0145
±0.0038
0.0271
±0.0095
nd nd
1.38min: 494.821m/z NA nd nd nd nd nd 0.0032 ±0.0006
0.0051 ±0.0017
nd nd nd
bersambung…
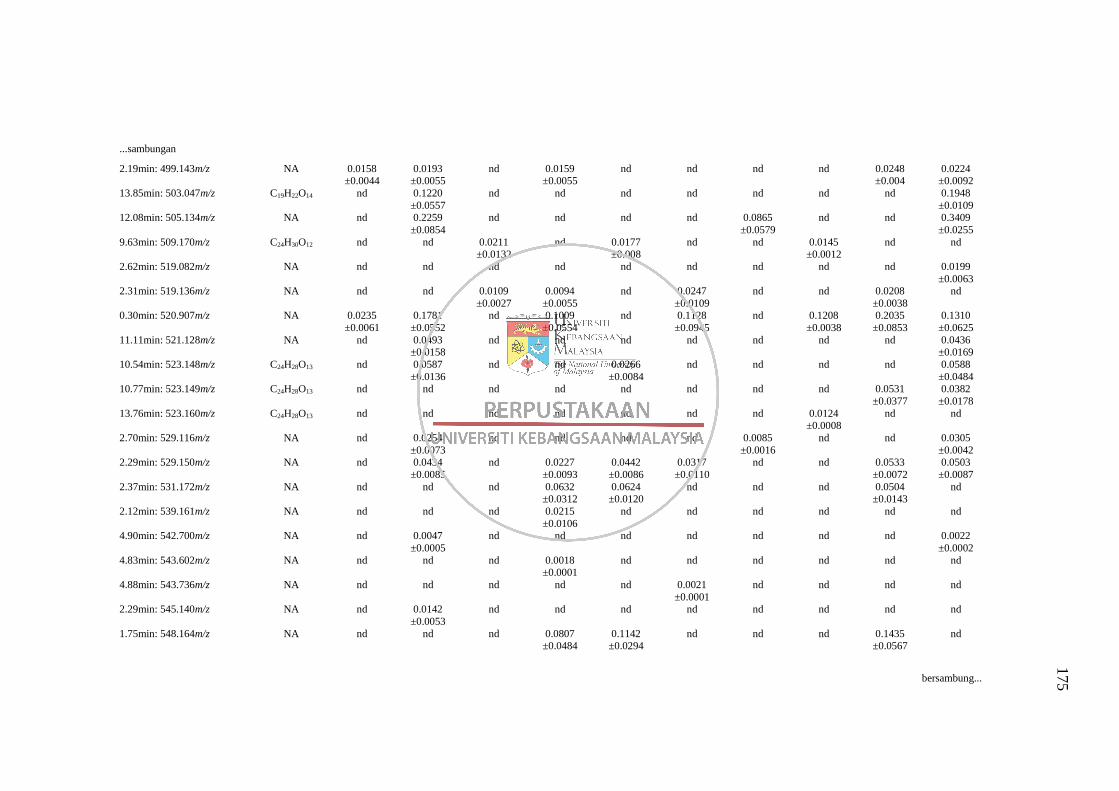
175
...sambungan
2.19min: 499.143m/z NA 0.0158 ±0.0044
0.0193 ±0.0055
nd 0.0159 ±0.0055
nd nd nd nd 0.0248 ±0.004
0.0224 ±0.0092
13.85min: 503.047m/z C19H22O14 nd 0.1220
±0.0557
nd nd nd nd nd nd nd 0.1948
±0.0109 12.08min: 505.134m/z NA nd 0.2259
±0.0854
nd nd nd nd 0.0865
±0.0579
nd nd 0.3409
±0.0255
9.63min: 509.170m/z C24H30O12 nd nd 0.0211 ±0.0132
nd 0.0177 ±0.008
nd nd 0.0145 ±0.0012
nd nd
2.62min: 519.082m/z NA nd nd nd nd nd nd nd nd nd 0.0199
±0.0063 2.31min: 519.136m/z NA nd nd 0.0109
±0.0027
0.0094
±0.0055
nd 0.0247
±0.0109
nd nd 0.0208
±0.0038
nd
0.30min: 520.907m/z NA 0.0235
±0.0061
0.1781
±0.0552
nd 0.1009
±0.0554
nd 0.1128
±0.0945
nd 0.1208
±0.0038
0.2035
±0.0853
0.1310
±0.0625
11.11min: 521.128m/z NA nd 0.0493 ±0.0158
nd nd nd nd nd nd nd 0.0436 ±0.0169
10.54min: 523.148m/z C24H28O13 nd 0.0587
±0.0136
nd nd 0.0266
±0.0084
nd nd nd nd 0.0588
±0.0484
10.77min: 523.149m/z C24H28O13 nd nd nd nd nd nd nd nd 0.0531
±0.0377
0.0382
±0.0178
13.76min: 523.160m/z C24H28O13 nd nd nd nd nd nd nd 0.0124 ±0.0008
nd nd
2.70min: 529.116m/z NA nd 0.0254
±0.0073
nd nd nd nd 0.0085
±0.0016
nd nd 0.0305
±0.0042 2.29min: 529.150m/z NA nd 0.0434
±0.0085
nd 0.0227
±0.0093
0.0442
±0.0086
0.0317
±0.0110
nd nd 0.0533
±0.0072
0.0503
±0.0087
2.37min: 531.172m/z NA nd nd nd 0.0632 ±0.0312
0.0624 ±0.0120
nd nd nd 0.0504 ±0.0143
nd
2.12min: 539.161m/z NA nd nd nd 0.0215
±0.0106
nd nd nd nd nd nd
4.90min: 542.700m/z NA nd 0.0047
±0.0005
nd nd nd nd nd nd nd 0.0022
±0.0002
4.83min: 543.602m/z NA nd nd nd 0.0018 ±0.0001
nd nd nd nd nd nd
4.88min: 543.736m/z NA nd nd nd nd nd 0.0021
±0.0001
nd nd nd nd
2.29min: 545.140m/z NA nd 0.0142
±0.0053
nd nd nd nd nd nd nd nd
1.75min: 548.164m/z NA nd nd nd 0.0807 ±0.0484
0.1142 ±0.0294
nd nd nd 0.1435 ±0.0567
nd
bersambung...
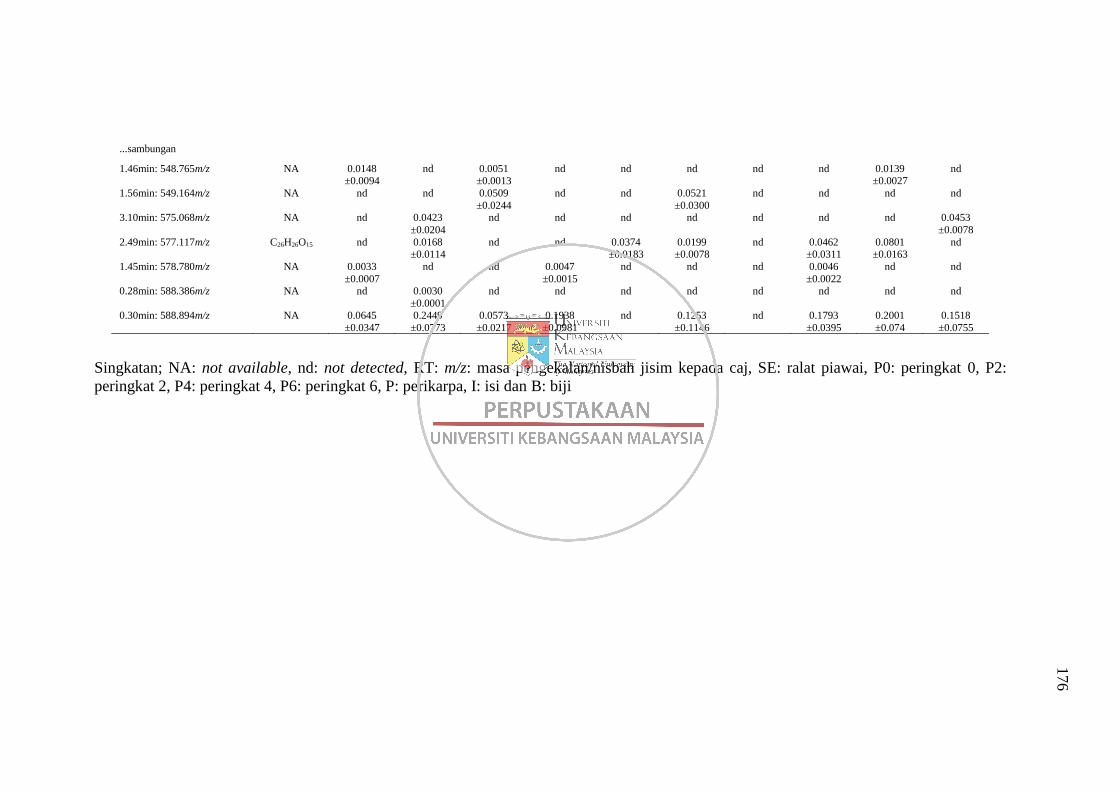
176
...sambungan
1.46min: 548.765m/z NA 0.0148 ±0.0094
nd 0.0051 ±0.0013
nd nd nd nd nd 0.0139 ±0.0027
nd
1.56min: 549.164m/z NA nd nd 0.0509
±0.0244
nd nd 0.0521
±0.0300
nd nd nd nd
3.10min: 575.068m/z NA nd 0.0423
±0.0204
nd nd nd nd nd nd nd 0.0453
±0.0078
2.49min: 577.117m/z C26H26O15 nd 0.0168 ±0.0114
nd nd 0.0374 ±0.0183
0.0199 ±0.0078
nd 0.0462 ±0.0311
0.0801 ±0.0163
nd
1.45min: 578.780m/z NA 0.0033
±0.0007
nd nd 0.0047
±0.0015
nd nd nd 0.0046
±0.0022
nd nd
0.28min: 588.386m/z NA nd 0.0030
±0.0001
nd nd nd nd nd nd nd nd
0.30min: 588.894m/z NA 0.0645
±0.0347
0.2449
±0.0773
0.0573
±0.0217
0.1938
±0.0981
nd 0.1253
±0.1146
nd 0.1793
±0.0395
0.2001
±0.074
0.1518
±0.0755
Singkatan; NA: not available, nd: not detected, RT: m/z: masa pengekalan/nisbah jisim kepada caj, SE: ralat piawai, P0: peringkat 0, P2:
peringkat 2, P4: peringkat 4, P6: peringkat 6, P: perikarpa, I: isi dan B: biji
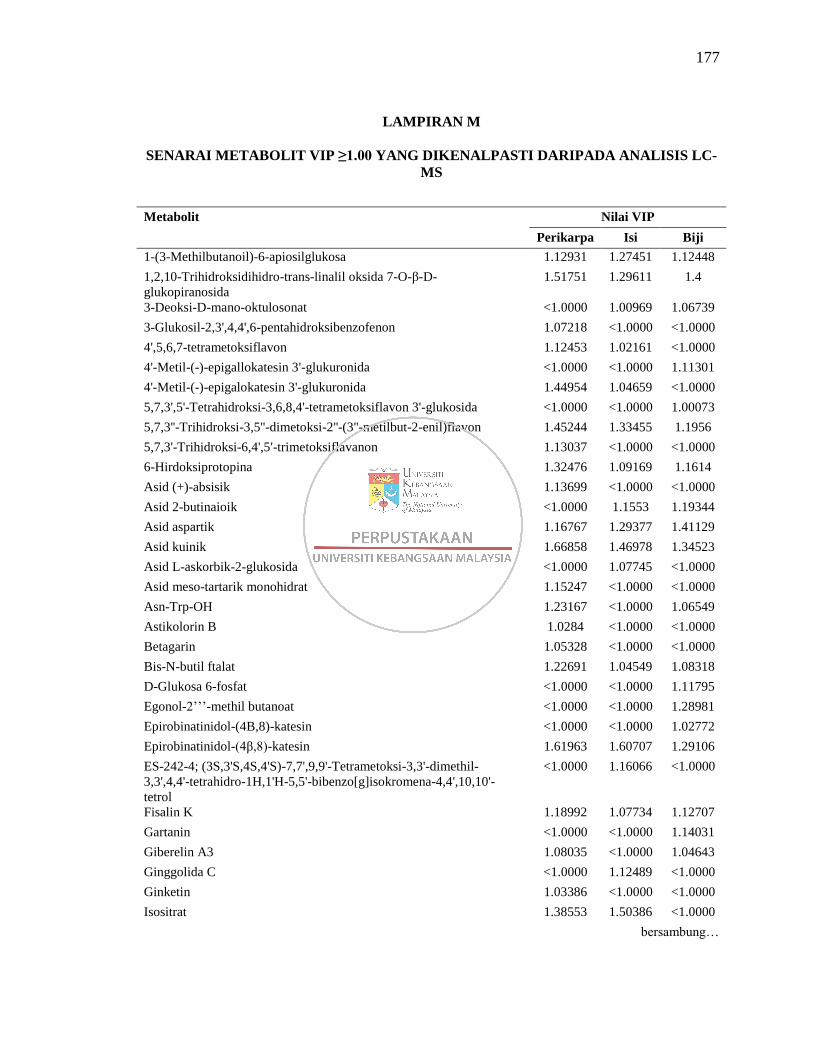
177
LAMPIRAN M
SENARAI METABOLIT VIP ≥1.00 YANG DIKENALPASTI DARIPADA ANALISIS LC-
MS
Metabolit Nilai VIP
Perikarpa Isi Biji
1-(3-Methilbutanoil)-6-apiosilglukosa 1.12931 1.27451 1.12448
1,2,10-Trihidroksidihidro-trans-linalil oksida 7-O-β-D-
glukopiranosida
1.51751 1.29611 1.4
3-Deoksi-D-mano-oktulosonat <1.0000 1.00969 1.06739
3-Glukosil-2,3',4,4',6-pentahidroksibenzofenon 1.07218 <1.0000 <1.0000
4',5,6,7-tetrametoksiflavon 1.12453 1.02161 <1.0000
4'-Metil-(-)-epigallokatesin 3'-glukuronida <1.0000 <1.0000 1.11301
4'-Metil-(-)-epigalokatesin 3'-glukuronida 1.44954 1.04659 <1.0000
5,7,3',5'-Tetrahidroksi-3,6,8,4'-tetrametoksiflavon 3'-glukosida <1.0000 <1.0000 1.00073
5,7,3''-Trihidroksi-3,5''-dimetoksi-2''-(3''-metilbut-2-enil)flavon 1.45244 1.33455 1.1956
5,7,3'-Trihidroksi-6,4',5'-trimetoksiflavanon 1.13037 <1.0000 <1.0000
6-Hirdoksiprotopina 1.32476 1.09169 1.1614
Asid (+)-absisik 1.13699 <1.0000 <1.0000
Asid 2-butinaioik <1.0000 1.1553 1.19344
Asid aspartik 1.16767 1.29377 1.41129
Asid kuinik 1.66858 1.46978 1.34523
Asid L-askorbik-2-glukosida <1.0000 1.07745 <1.0000
Asid meso-tartarik monohidrat 1.15247 <1.0000 <1.0000
Asn-Trp-OH 1.23167 <1.0000 1.06549
Astikolorin B 1.0284 <1.0000 <1.0000
Betagarin 1.05328 <1.0000 <1.0000
Bis-N-butil ftalat 1.22691 1.04549 1.08318
D-Glukosa 6-fosfat <1.0000 <1.0000 1.11795
Egonol-2ʼʼʼ-methil butanoat <1.0000 <1.0000 1.28981
Epirobinatinidol-(4B,8)-katesin <1.0000 <1.0000 1.02772
Epirobinatinidol-(4β,8)-katesin 1.61963 1.60707 1.29106
ES-242-4; (3S,3'S,4S,4'S)-7,7',9,9'-Tetrametoksi-3,3'-dimethil-
3,3',4,4'-tetrahidro-1H,1'H-5,5'-bibenzo[g]isokromena-4,4',10,10'-
tetrol
<1.0000 1.16066 <1.0000
Fisalin K 1.18992 1.07734 1.12707
Gartanin <1.0000 <1.0000 1.14031
Giberelin A3 1.08035 <1.0000 1.04643
Ginggolida C <1.0000 1.12489 <1.0000
Ginketin 1.03386 <1.0000 <1.0000
Isositrat 1.38553 1.50386 <1.0000
bersambung…
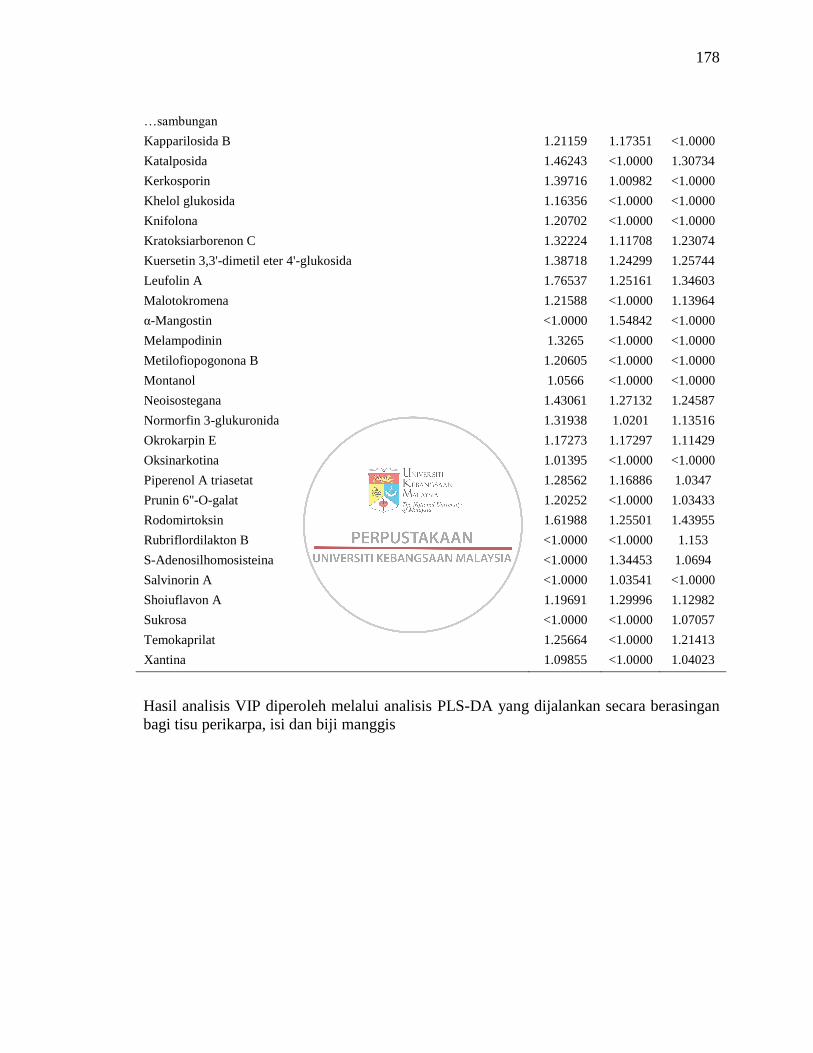
178
…sambungan
Kapparilosida B 1.21159 1.17351 <1.0000
Katalposida 1.46243 <1.0000 1.30734
Kerkosporin 1.39716 1.00982 <1.0000
Khelol glukosida 1.16356 <1.0000 <1.0000
Knifolona 1.20702 <1.0000 <1.0000
Kratoksiarborenon C 1.32224 1.11708 1.23074
Kuersetin 3,3'-dimetil eter 4'-glukosida 1.38718 1.24299 1.25744
Leufolin A 1.76537 1.25161 1.34603
Malotokromena 1.21588 <1.0000 1.13964
α-Mangostin <1.0000 1.54842 <1.0000
Melampodinin 1.3265 <1.0000 <1.0000
Metilofiopogonona B 1.20605 <1.0000 <1.0000
Montanol 1.0566 <1.0000 <1.0000
Neoisostegana 1.43061 1.27132 1.24587
Normorfin 3-glukuronida 1.31938 1.0201 1.13516
Okrokarpin E 1.17273 1.17297 1.11429
Oksinarkotina 1.01395 <1.0000 <1.0000
Piperenol A triasetat 1.28562 1.16886 1.0347
Prunin 6''-O-galat 1.20252 <1.0000 1.03433
Rodomirtoksin 1.61988 1.25501 1.43955
Rubriflordilakton B <1.0000 <1.0000 1.153
S-Adenosilhomosisteina <1.0000 1.34453 1.0694
Salvinorin A <1.0000 1.03541 <1.0000
Shoiuflavon A 1.19691 1.29996 1.12982
Sukrosa <1.0000 <1.0000 1.07057
Temokaprilat 1.25664 <1.0000 1.21413
Xantina 1.09855 <1.0000 1.04023
Hasil analisis VIP diperoleh melalui analisis PLS-DA yang dijalankan secara berasingan
bagi tisu perikarpa, isi dan biji manggis
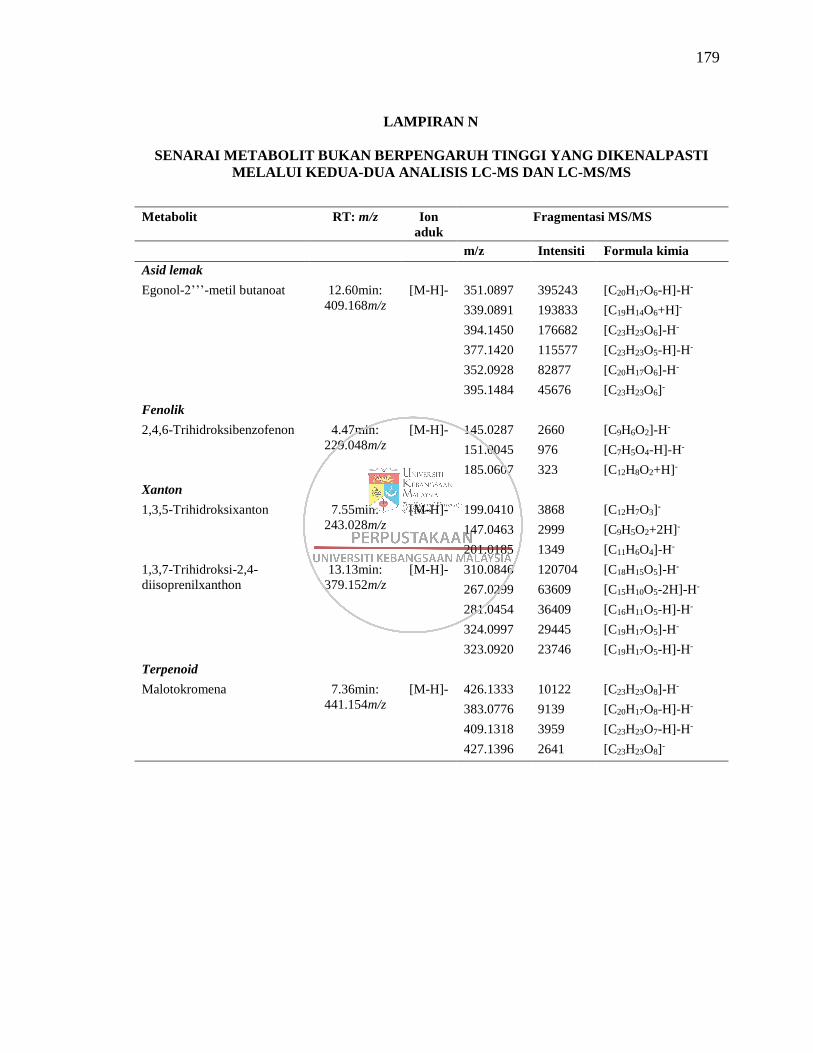
179
LAMPIRAN N
SENARAI METABOLIT BUKAN BERPENGARUH TINGGI YANG DIKENALPASTI
MELALUI KEDUA-DUA ANALISIS LC-MS DAN LC-MS/MS
Metabolit RT: m/z Ion
aduk
Fragmentasi MS/MS
m/z Intensiti Formula kimia
Asid lemak
Egonol-2ʼʼʼ-metil butanoat 12.60min:
409.168m/z
[M-H]- 351.0897 395243 [C20H17O6-H]-H-
339.0891 193833 [C19H14O6+H]-
394.1450 176682 [C23H23O6]-H-
377.1420 115577 [C23H23O5-H]-H-
352.0928 82877 [C20H17O6]-H-
395.1484 45676 [C23H23O6]-
Fenolik
2,4,6-Trihidroksibenzofenon 4.47min:
229.048m/z
[M-H]- 145.0287 2660 [C9H6O2]-H-
151.0045 976 [C7H5O4-H]-H-
185.0607 323 [C12H8O2+H]-
Xanton
1,3,5-Trihidroksixanton 7.55min:
243.028m/z
[M-H]- 199.0410 3868 [C12H7O3]-
147.0463 2999 [C9H5O2+2H]-
201.0185 1349 [C11H6O4]-H-
1,3,7-Trihidroksi-2,4-
diisoprenilxanthon
13.13min:
379.152m/z
[M-H]- 310.0846 120704 [C18H15O5]-H-
267.0299 63609 [C15H10O5-2H]-H-
281.0454 36409 [C16H11O5-H]-H-
324.0997 29445 [C19H17O5]-H-
323.0920 23746 [C19H17O5-H]-H-
Terpenoid
Malotokromena 7.36min:
441.154m/z
[M-H]- 426.1333 10122 [C23H23O8]-H-
383.0776 9139 [C20H17O8-H]-H-
409.1318 3959 [C23H23O7-H]-H-
427.1396 2641 [C23H23O8]-
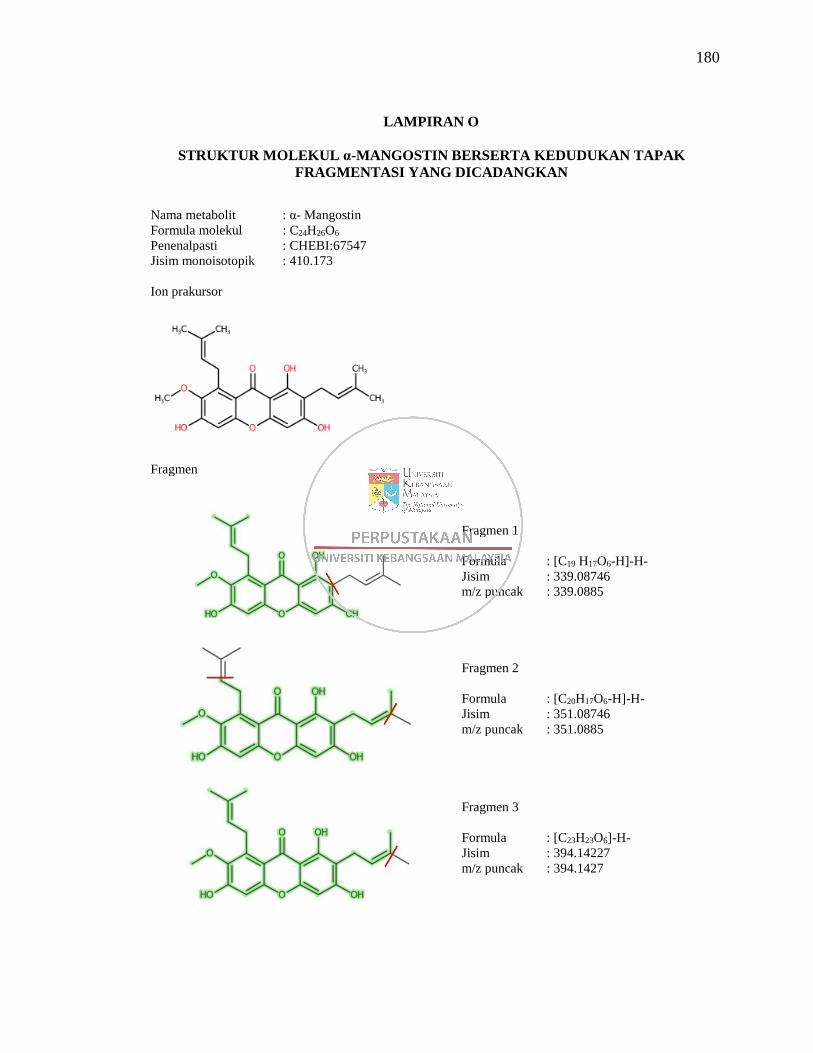
180
LAMPIRAN O
STRUKTUR MOLEKUL α-MANGOSTIN BERSERTA KEDUDUKAN TAPAK
FRAGMENTASI YANG DICADANGKAN
Nama metabolit : α- Mangostin
Formula molekul : C24H26O6
Penenalpasti : CHEBI:67547
Jisim monoisotopik : 410.173
Ion prakursor
Fragmen
Fragmen 1
Formula : [C19 H17O6-H]-H-
Jisim : 339.08746
m/z puncak : 339.0885
Fragmen 2
Formula : [C20H17O6-H]-H-
Jisim : 351.08746
m/z puncak : 351.0885
Fragmen 3
Formula : [C23H23O6]-H-
Jisim : 394.14227
m/z puncak : 394.1427







