file name: supplementary information description ...10.1038/s41467-017... · o-glycosylation of tsr...
TRANSCRIPT

File name: Supplementary Information Description: Supplementary Figures, Supplementary Table, Supplementary Methods and Supplementary References

1
Supplementary Figure 1. O-glycosylation of TSR domains in metazoans.

2
Supplementary Figure 2. Multiple sequence alignment of putative POFUT2s from Plasmodium
falciparum (PF3D7_0909200.1), P. berghei (PBANKA_0810400.1), P. chabaudi chabaudi (PCHAS_0810700.1), P. cynomolgi (PCYB_071700-t26_1), P. knowlesi (PKNH_0707200.1), P. reichenowi (PRCDC_0907300.1), P. vivax (PVX_098900.1) and P. yoelii yoelii (PYYM_0813400.1). Black highlights indicate identity, gray highlights indicate similarity. For
consensus sequence: uppercase is identity, lowercase is consensus level > 0.5, ! is anyone of IV, $
is anyone of LM, % is anyone of FY, # is anyone of NDQEBZ.
1 10 20 30 40 50 60 70 80
PF3D7_0909200.1 MK C D Y D F K KKY YDV GEGFNLQKE YR L Y LN Y L L I Q I V F R IM NI I L VI DK I FIIVLL F...FFFKVIDRV CVTP KL LKE LG F FL F S N KK IN Y PBANKA_0810400.1 MK C D Y D F K KKY YDV GEGFNLQKE YR L Y LN Y L I I S I I Y K IL NI V M VI DK I NMIYNL SISLYSLIIILTD YAKT SL NKN LN K FL L S Q KR EY Y PCHAS_0810700.1 MK C D Y D F K KKY YDV GEGFNLQKE YR L Y LN Y L I V S I I Y K LL NI V M VI DK I NMTYNL SLCLCSLIIVLTD YTKT SL NKN LS N FL L S Q KQ EY Y PCYB_071700-t26_1 MK C D Y D F K KKY YDV GEGFNLQKE YR L Y LN Y L V L S V V Y K VL NI V V V ER V SRAHIW ALLLACLPPRVKN DKDV SP KTD TG A PF L A G Y QE TH H PKNH_0707200.1 MK C D Y D F K KKY YDV GEGFNLQKE YR L Y LN Y L V L S V V F K IL NL V V AV R V GRTHIW ALLLVCLTPRFKN DKDV SP RTD TG A PF L A Y QEV TQ H PRCDC_0907300.1 MK C D Y D F K KKY YDV GEGFNLQKE YR L Y LN Y L L I I V F IM NI I L VI DK I FIIVLL F...FFFKVIDRV CVTPPKS LKE LG S FL E F S N KK LK Y PVX_098900.1 MK C D Y D F K KKY YDV GEGFNLQKE YR L Y LN Y L V L S V V Y K VL I V V AV ER V GRAHIW ALLLACLPPRFRN DKDV SP RTD TG A PF H L A Y QE TH H PYYM_0813400.1 MK C D Y D F K KKY YDV GEGFNLQKE YR L Y LN Y L I V S I V Y K LL NI V M VI DK I NIIYNL SFSLCSLIIVLTD YAKT SL NKN LN K FL L S Q KR EY Y consensus>80
90 100 110 120 130 140 150 160 170
PF3D7_0909200.1 VLPPWCY HW W F N VIP E Y G D K L C K VT K L IM II Y E Y MIN K IL F E I R NIR .GNN R EF F TD KK E EKL NYS I S Y DNY....KE S LI PF E N NVN F .PBANKA_0810400.1 VLPPWCY HW W F N VIP E Y G D K L C K LS R L II II F D Y IIS K LL Y D I K SNK .HDN Q NI L MN KN S KKL DVT F F Y DSNA..QKA Y HV PF K Y EDY F .PCHAS_0810700.1 VLPPWCY HW W F N VIP E Y G D K L C K LS R L II II F E Y IIS K LL Y D I K ANK .HAN Q DI L TN KN S KKL DVT F F Y NSQA..SKT Y HG PF K P EDY F .PCYB_071700-t26_1 VLPPWCY HW W F N VIP E Y G D K L C K VT K I AL VM Y F ILS VI F D V K GRE THNR K DV F LK QN HQ EEQ SHV Y YQH EDWSKMDEK S QV DL K H KYY K KPKNH_0707200.1 VLPPWCY HW W F N VIP E Y G D K L C K LT K I VL VI Y E F ILS R I F E V K GRE KHNR K SV F MK QN T EKE PHA Y Y HF GDWSKMNEK S QF DI K H ENY L KPRCDC_0907300.1 VLPPWCY HW W F N VIP E Y G D K L C K VT K L IM II Y E Y MIN K IL F E NRK .GNN R EF F TD KK E EKL NYC I S Y DNY....KE S LI PF E NTNVNSF .PVX_098900.1 VLPPWCY HW W F N VIP E Y G D K L C K VT R I AL VM Y E F ILS R II F D V K GRE TNAR K SI F LK QN A EGQ PHT Y Y H GEWPKRGDK S QV KL K Q KGY L KPYYM_0813400.1 VLPPWCY HW W F N VIP E Y G D K L C K LS R L II II F D Y IIS K LL Y D I K TNK .HDN E NI L ID KN S KKL DVT F F Y DSYG..QKG Y HV PF K Y ENY F .consensus>80
180 190 200 210 220 230 240 250 26 0
PF3D7_0909200.1 C KY YSG C K C F S LIK LV EL N ED L L C E NV TTI ISNY IT L L I V T I F N L I FN L GQF KK H L Y NT QSE YSYNM F SI EN L.YN T QS N P V YQS L NK SY PBANKA_0810400.1 C KY YSG C K C F S LIK LV EL N ED L L C D SV TNI VTNY VN V L I I N V F N E I I FN I GIV KN Y T N KG NTL YSDYI L YV HK YIHN N NS V P P F N L EK YN PCHAS_0810700.1 C KY YSG C K C F S LIK LV EL N ED L L C D SV TNI VTSY VN V L I I T I F N E V I FN I GTV NN Y T D KG NVI YGDYI F SV HK YIHN N NG V P Q F N L ER YN PCYB_071700-t26_1 C KY YSG C K C F S LIK LV EL N ED L L C D SV TNV ITSY VS L I V I N E L V YS I GNL KN H V N KG KTE LEIFF F ST SD H.YDNN HGRN ALA V A P KY NE PKNH_0707200.1 C KY YSG C K C F S LIK LV EL N ED L L C D SV NV ITSY VS L I I V S I F N E L V YS I GNL KN Y V N A KG KTE LEFFF F ST SD H.YD D HG N S V V A P ED NE PRCDC_0907300.1 C KY YSG C K C F S LIK LV EL N ED L L C E NI TTI ISNY IT L L I V T I F N E L I FN L QF KK Y I Y NT ESE YSYNM F SI KN L.YD T QS N P V Y N L KK LY APVX_098900.1 C KY YSG C K C F S LIK LV EL N ED L L D SV TNV ITS VS L I A V S I F N D L V YS I GNLRKN H E K KG KME LEFFF HF ST LD Q.YD D HG N A M V A P ED NE PYYM_0813400.1 C KY YSG C K C F S LIK LV EL N ED L L C D SV T I VTSY VN V L I I S V F T E I I FN I GII EN Y T N H KG NTL YGDYI L FV HK YIHN N NS V P P F N L ER YN consensus>80
270 280 290 300 310 320 330 340
PF3D7_0909200.1 K YIS HLRY DF I Y KLLYIMF IF DEK I F Q K FY N H NNYI T N T K S V I I L I I S I V Y K FY SNIL . H S Y R N PP H A L NNCRI A VE QK NKD H H N QNNL EPBANKA_0810400.1 K YIS HLRY DF I Y KLLYIMF IF DEK I F Q K FY N H NNYI N S N K T V V I I M I T V I F H FY KEIL . N C K S D PS Q A L NNKEK S KY QY NKH K F E KDNY EPCHAS_0810700.1 K YIS HLRY DF I Y KLLYIMF IF DEK I F Q K FY N H NNYI S N T K T V V L I M I T V I F H FY KEIL . N C K S D PS Q A F NSKDK S KS QY NKH K F E KDNY EPCYB_071700-t26_1 K YIS HLRY DF I Y KLLYIMF IF DEK I F Q K FY N H NQFI S N T R S V V I I L I T V V F H F EKNF S N C K T D SP R S L RKSDI S KE KK DKQ P F. E G.KL PPKNH_0707200.1 K YIS HLRY DF I Y KLLYIMF IF DEK I F Q K FY N H NQFV S N T R S V I I L L V T V V Y FF EKNF N N C K T D SP E S L RKSSI S KQ KK DRQ S NF E E.KL TPRCDC_0907300.1 K YIS HLRY DF I Y KLLYIMF IF DEK I F Q K FY N H NNYI T N N K S V I I L I I S I V Y K FY SNIL . H S N R N PP H A V NNCRI A LQ QK DQY Y H N QNNL EPVX_098900.1 K YIS HLRY DF I Y KLLYIMF IF DEK I F Q K FY N H QFV S T R S V V I L L V T V V F H FF D EKNF SKN C K T D SP G S L RKSTL S KE KK DSQ P F E E.KL TPYYM_0813400.1 K YIS HLRY DF I Y KLLYIMF IF DEK I F Q K FY N H NNYI N S S K T I V L I M I T V I F H F KEIL . N C K S D PS Q A F NNKEK S KN QY NKH K F H E KDNY Econsensus>80
350 360 370 380 390 400 410 420 430
PF3D7_0909200.1 G I QWIC S F G IFSRF M I WER LI KG N DLC Y I Y K Y S IE I N T I N IK I I AL I I EFS TR YI N N H N QINQ S H DDNDQD NS KK VH .FNHK Q KN DNY D RPBANKA_0810400.1 G I QWIC S F G IFSRF M I WER LI KG N DLC Y I Y K Y S IE V N T I NN LK I I I L I QVA AN ST H I Y T KEHQ G S N..EQ NR KK QY .YDHSS E NN NTY E IPCHAS_0810700.1 G I QWIC S F G IFSRF M I WER LI KG N DLC Y I Y K Y S IE V N T L NN LK I I AI L I QVA AN SI H M Y T KENQ G S N..EK NR KK QH .YDHN E GN NTY E TPCYB_071700-t26_1 G I QWIC S F G IFSRF M I WER LI KG N DLC Y I Y K Y S VD V T S L T LR I A AL L L QVA TR GT H K S G GADH G S G D..QG KK SD QD DLDEE Q HP MSL Q TPKNH_0707200.1 G I QWIC S F G IFSRF M I WER LI KG N DLC Y I Y K Y S ID V N S L NT LR I AL L L QVA IR GT H K S G GQDH G S D..EG KK SD QDTHLDEE Q HP MRL Q TPRCDC_0907300.1 G I QWIC S F G IFSRF M I WER LI KG N DLC Y I Y K Y S IE I N T I NN IK I I AL I I EFS TR HI N N H N QINN S H DNDQD NS KK VH .FNHK Q KN DNY D KPVX_098900.1 G I QWIC S F G IFSRF M I WER LI KG N DLC Y I Y K Y S VD V N S L ST LR V A AL L L QVA AR GT H K S G GPDH G S N..HE KK SD QD HLDEE Q RP MRL Q KPYYM_0813400.1 G I QWIC S F G IFSRF M I WER LI KG N DLC Y I Y K Y S IE V N T L NN LK I I AI L I QVA AN SI H I Y T NENQ G S N..EQ NR KK QH .YDHH E ND NTY E Iconsensus>80
440 450 460
PF3D7_0909200.1 D T C P N SIYR K YI I Y F H N K I K N N LS F R EY TN............T PBANKA_0810400.1 D T C P N SIYR K YI L F F H S L K I K I G PS F KY PF............A PCHAS_0810700.1 D T C P N SIYR K YI L F F H N L K I K I G PS L KY PFA...........E PCYB_071700-t26_1 D T C P N SIYR R YL I Y H Q V R V D R DSAY Y GD PPEVRVVGIGGKRSA PKNH_0707200.1 D T C P N SIYR K FL I Y F H Q V R V K R D AH Y GY PPELWGVSNGGKKNA PRCDC_0907300.1 D T C P N SIYR R YI I Y F H N I K I K N N PS F EY SN............I PVX_098900.1 D T C P N SIYR R FL I Y F Q V R D R D AHYY GERREEGRLM........ PYYM_0813400.1 D T C P N SIYR K YI L F F H N L K I K I G PS L KY PFA...........E
consensus>80

3
Supplementary Figure 3. Multiple sequence alignment of putative POFUT2s from P. falciparum (PF3D7_0909200.1) and P. vivax (PVX_098900.1) with POFUT2 from Homo sapiens (CAC24557.1)
1 and Caenorhabditis elegans (NP_001255070.1)
2. The glutamate residue that
serves as the catalytic base and the arginine residue that binds the pyrophosphate moiety of GDP-
fucose, both highlighted in red, are conserved. Black highlights indicate identity, gray highlights indicate similarity. For consensus sequence: uppercase is identity, lowercase is consensus level > 0.5, ! is anyone of IV, $ is anyone of LM, % is anyone of FY, # is anyone of NDQEBZ.
1 10 20 30 40 50 60 70 80
PF3D7_0909200.1 M L YDV EGFNL R L K VL L F T Q I ED VY L RKKYIM N G KEIF L VI N FI...I FFF KVIDRVICV P ..KL CLK . LGDFFFF K I Q Y SL Y NKKDKIN IPVX_098900.1 M L YDV EGFNL R L K VA LLA S V DD VY KKKYVL G KEV VA AV GRAHIW CLPPRFRNLDKDV ..SP CRT . TGDAFYPFK HI Q LY L YY NQEERTH V
CAC24557.1 M L YDV EGFNL R L LL AV W S Q D I A RRRYLL N P RDVY IA LL T ATLSFVF G S PPA....SA G EFWPGQSAA . LS....GA S P R I S K LKT...E ENP_001255070.1 M L YDV EGFNL R L H I LVL F N V D Y A KKFLL N G RDVY VA V S FF...P Q F AEKI.AFAE SD..QT SRV SNR .....SV AE F R M NT R RDS..GE Nconsensus>70
90 100 110 120 130 140 150 160
PF3D7_0909200.1 LVLPPW HW W FF IP E D Y V L M K II YEEY G MI LD Y KS I Y CY T NIRKG..NN R EF NTDI K V EKLY .NYS I NSKYI N K.........E FL LPFE EPVX_098900.1 LVLPPW HW W FF IP E D V A I L LQ VM Y EY G P IL I W R D KS V HY CY T GRERT.N R K SI N KA NV A EGQF . HT Y SYRHI GE P.....K G K FQ LKLD KCAC24557.1 LVLPPW HW W FF IP E D W L V I L LNK VI YE F G P V AE W K D R I V GR Y QSPDIHQ R P SE D PS N Q IAES G FI Q YVLQSY G KEGTWEE V E PC. D... .NP_001255070.1 LVLPPW HW W FF IP E D Y L V L L LNR VI FEDF P VI AE W K E RS L I GR H KR.M..E A S RL D ES F LDENR. .I Q YLQHY G GT.EYVR F K C. PPA. .
consensus>70
170 180 190 200 210 220 230 240
PF3D7_0909200.1 C L S L K H Y Y GY I KQ S N S I I I VLI NI EL L CNINVNRFKQ.FC KCE K .N.VL S CTT NT SE Y Y MI NYF TS ENLFLYN T KQST VPFVN YQSN ED IPVX_098900.1 C L S L KN H Y Y G V K L T V A VLI H NI EL L CQVKGYKLKKNLR CD K .S.VE S KCTN KG KME EFFFI SHF SST LDIFQYD D K GS VAFMN VDAN ED V
CAC24557.1 C L S L Q H Y F GY N LS Q V L A VML R NL .........QLLYS DK E .YRGW W .EETRGL VS V GSA.SI AP LR..NTS R D AE HDHYGGKEYWDTRR SNP_001255070.1 C L S L KQ F F Y V RN VS Q S L L IMV R TI EV A .........ESHY .VEE KWKGW YS .ED YS FQ I GD .GT KD KHSNFSEST D AE HEHYG DYWK RR Sconsensus>70
250 260 270 280 290 300 310
PF3D7_0909200.1 L Y H R DF L FN L G Y L I L YI R VP I L KL IM I F L NK SY NN ISNI K.........................TNH SS YT K S YN P HIA L LY FINNCRI PVX_098900.1 L Y H R DF L YS I GD F K S I L I V V L KL IM I F P ED NE Q VE NFK ........................SKN SC YT RK STYD SP GIS L LY FLRKSTL CAC24557.1 L Y H R DF M F GD F R L S L L W R VP L V KI LM V F V ARH REV E RS H N TDDADRIPFQEDWMKMKVKLGSALGGP GV RK I GH QD S EGA R RS KTHRLDK NP_001255070.1 L Y H R DF M YS V D F K L S L YA R LP I A L L I Y R ND DVA A RK Y D DDKRDKTKLVDDWTKEKPRR.TAIGGP GI W RR L R AQ T PGT KI QD CKKLDLQK
consensus>70
320 330 340 350 360 370 380 390
PF3D7_0909200.1 D G I Q C F G S F I R C IAS EI VI K F FY Q LHE IE WI R I N T E LI G NQ L EK....V QK NKDFHQYK H Y NN NN. EFS T SYI IF R MN NW H NK QI NID S Y
PVX_098900.1 D G I Q C F G S F I R C V T EV VI H F FY EN LH VA VD WI AR V N S H E LIG G L G S EK....K KK DSQFPQFK F F . EK. T Q SGT IF R M KW S K GPDHNLD YCAC24557.1 D G I Q C F G S F I R C VAT L R W E KD VA ID WI AH I T S R E ILG TT G AV....RKEYEE KKLLPEMV FEPT . ELELY G ARF SV T F HE E LDPK YNRF DNP_001255070.1 D G I Q C F G S F I R C LAT EL LL Y F DT L D IA ID YL AH I S T R D IIG P TT L P APDQEVD KA NGELEV.... R T. QK. N Q AAY YE T F QE E F IS FNR Dconsensus>70
400 410 420 430 440 450 460
PF3D7_0909200.1 Y D QK K I HINDDN QDIKNSYKKIVHIFNHKAL I N DNYSDRDKKYINTICYNFLSHFPNNRSIYRKEYITNT.... PVX_098900.1 Y S E QK R L ISTNH LRKKYSDVQDAHL.DEEAL L P MRLSQKDRDFLRTICYDFAHYYPQNVSIYRRGERREEGRLM CAC24557.1 Y Q E TH K EKAC. .................QP W IT .........................................
NP_001255070.1 Y T E K K V EPTC. .................QPA W I ......................................... consensus>70

4
Supplementary Figure 4. (A) Homology model of P. falciparum POFUT2 (light blue) overlaid on
the structures of H. sapiens POFUT2 (brown, 4AP5, RMS = 0.39 Å) 1 and C. elegans POFUT2
(pale green, 5FOE, RMS = 1.4 Å) 2, and (B) with GDP-fucose (green) and a TSR domain (gold)
docked to give a plausible representation of the Michaelis complex. (C) The relative positions of substrates and catalytic residues within the active site of the P. falciparum POFUT2 homology
model.

5
Supplementary Figure 5. (A) Construct design for expression of P. vivax POFUT2 using
baculovirus in Sf21 insect cells. (B) Construct design for expression of P. falciparum CSP and
TRAP TSR domains in E. coli. (C) Coomassie-stained SDS-PAGE gels of reduced and non-
reduced samples of purified recombinant proteins.
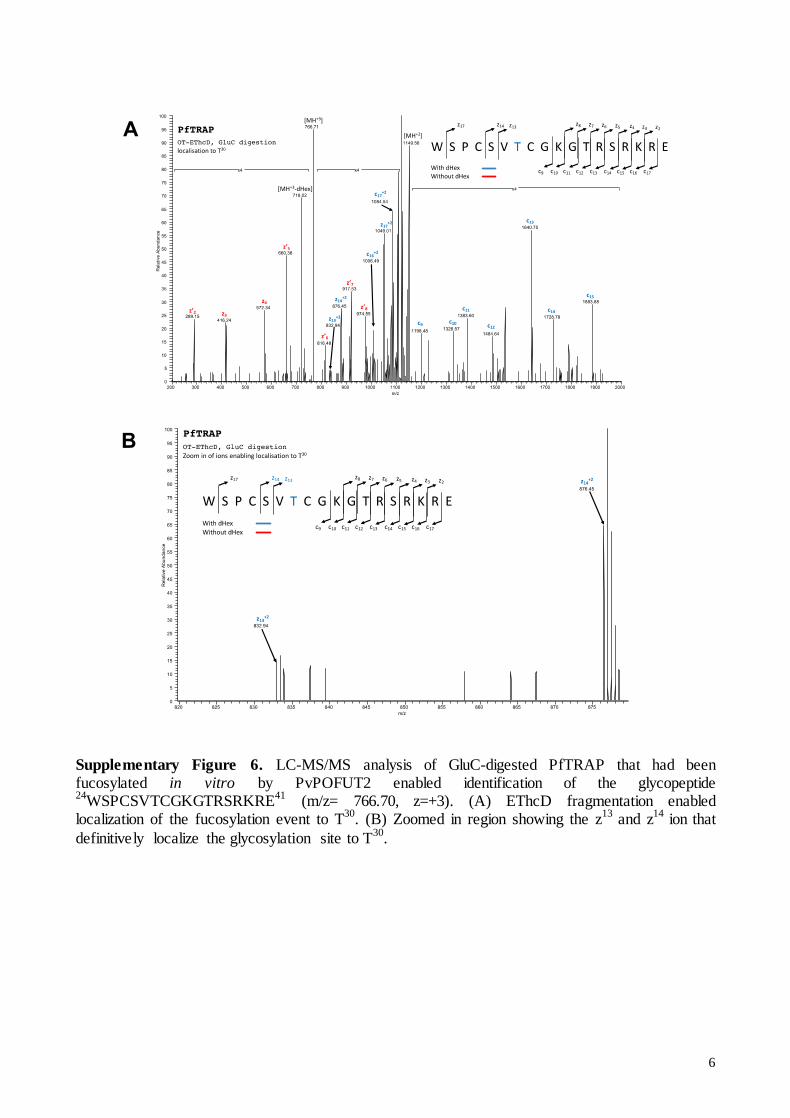
6
Supplementary Figure 6. LC-MS/MS analysis of GluC-digested PfTRAP that had been fucosylated in vitro by PvPOFUT2 enabled identification of the glycopeptide 24
WSPCSVTCGKGTRSRKRE41
(m/z= 766.70, z=+3). (A) EThcD fragmentation enabled localization of the fucosylation event to T
30. (B) Zoomed in region showing the z
13 and z
14 ion that
definitively localize the glycosylation site to T30
.
A
B

7
Supplementary Figure 7. (A) Strategy for inserting a C-terminal triple hemagglutinin (HA)
epitope tag into the P. falciparum POFUT2 locus. (B) Southern blot analysis of parental NF54 and
POFUT2-HA3 parasite cultures. (C) Western blot analysis of NF54 and POFUT2-HA3 trophozoites
and schizonts. Anti-PfAldolase was used as a loading control. (D) Immunofluorescence microscopy
of ring (top) and trophozoite (bottom) stage POFUT2-HA3 parasites stained for the ER-resident
protein plasmepsin V (PMV, red) and POFUT2-HA3 (green) with the nucleus visualized using
DAPI (blue). Punctae of co-localization (yellow) are evident in both micrographs and indicated
with arrowheads in the ring parasite for reference. Scale, 2 m.

8
Supplementary Figure 8. (A) Strategy for disrupting the POFUT2 locus in P. falciparum NF54.
(B) Southern blot analysis of parental NF54 and two clonal POFUT2 parasite cultures (D3 and
G8). (C) Quantitation of the blood stage growth rate of POFUT2 (D3 and G8) parasite cultures
relative to NF54. (D) Quantitation of gametocytogenesis for NF54 and POFUT2 (D3 and G8)
parasite cultures. Data in (C) and (D) are mean ± S.E.M from three independent experiments. No
statistical differences were found relative to NF54 in panel (C) (p=0.2367) or panel (D) (p=0.1762)
using the Kruskal-Wallis one-way ANOVA.

9
Supplementary Table 1. Oligonucleotides used in this study.
*Restriction sites used for cloning are underlined.
Oligonucleotide Sequence (5’ to 3’)*
PFOUTHA_F ATCGGATCCCTTATTTTACCTTTCGAAGA
POFUTHA_R ATCCTGCAGGTGTGTTGGTAATATATTCCT
POFUT2KO_FL1_F ATCCCGCGGCTCTCAATTTGCGAAGCACA
POFUT2KO_FL1_R GATACTAGTAACACCAAGGTGGAAGTACAAGA
POFUT2KO_FL2_F ATCGAATTCTGTTCCTCCTATTCATATCGCTTT
POFUT2KO_FL2_R GATCCTAGGTGTGTTGGTAATATATTCCTTTCG
POFUT2hm_F TTCTTATTTTACCTTTCGAAGAATG
POFUT2hm_R TGAATAACATTCACTCTGTTTTGTG
hDHFRhm_F ACCTAATAGAAATATATCAGGATCC
hDHFRhm_R GGTTGTGGTCATTCTCTGGA
Pf18S_F GTAATTGGAATGATAGGAATTTACAGGT
Pf18S_R TCAACTACGAACGTTTTAACTGCAAC
Pfs25_F CAGATGAGTGGTCATTTGGAA
Pfs25_R CAAGCGTATGAAACGGGATT
PfCTRP_F GAATGGAGTCCCTGTCCTGA
PfCTRP_R TGGTCCTTTCCTTTCCCTTT
Asrps7_F TGCGGCTTCAGATCCGAGTTC
Asrps7_R TTCGTTGTGAACCCAAATAAAAATC
PfTRAP_F GTGTTTGGGACGAATGGTCT
PfTRAP_R CGGGTTCATCTGGAACATCT
PfCSP_F TGGGAAACAGGAAAATTGGT
PfCSP_R CATCCGCTGGTTGCTTTAAT

10
Supplementary Methods
Generating PfPOFUT2 homology model
A homology model for P. falciparum POFUT2 was built using the Phyre2 server 3. This model was
overlaid on the structures of H. sapiens POFUT2 (4AP5) 1 and C. elegans POFUT2 (5FOE)
2 using
PyMOL. The P. falciparum POFUT2 model, GDP-fucose and the structure of the TSR domain
from 5FOE were docked together using the ROSIE server 4-6
. All images for Supplementary Figure
4 were generated using PyMOL.
P. falciparum TSR domain expression and purification
Codon-harmonised dsDNA polynucleotides encoding residues 242–288 of P. falciparum TRAP
(PF3D7_1335900) and residues 307–376 of P. falciparum CSP (PF3D7_0304600) with an N-
terminal Strep-II tag and C-terminal hexahistidine tag were synthesized (Integrated DNA
Technologies) and cloned into pGEX6P3 (Invitrogen) at the BamHI and NotI restriction sites to
give pGEX-PfTRAP and pGEX-PfCSP.
>tags-PfTSR
GGATCCTGGTCACATCCGCAGTTTGAAAAAGGTAGCGCAAGCTGTGGTGTTTGGGATGAATGGTCACCGTGTAG
CGTTACCTGTGGTAAAGGCACCCGTAGCCGTAAACGTGAAATTCTGCATGAAGGTTGTACCAGCGAACTGCAAG
AACAGTGTGAAGAAGAACGTTGTCTGCCTGGTAGCCATCATCATCACCATCATTAAGCGGCCGC
>tags-PfCSPTSR
GGATCCTGGTCACATCCGCAGTTTGAAAAAGGTAGCAATAATGAAGAACCGAGCGATAAACACATCAAAGAATA
TCTGAACAAAATCCAGAATAGCCTGAGCACCGAATGGTCACCGTGTAGCGTTACCTGTGGTAATGGTATTCAGG
TTCGTATTAAACCGGGTAGCGCAAATAAACCGAAAGATGAACTGGATTATGCCAACGATATCGAGAAAAAAATC
TGCAAAATGGAAAAATGCAGCAGCGTTGGTAGCCATCATCATCACCATCATTAAGCGGCCGC
Sequence-verified plasmids were transformed into E. coli BL21(DE3) and grown in 1.0 L LB
media (100 μg ml–1
Amp) with shaking (250 rpm) at 37 °C until the culture reached an OD600 of
0.8. The culture was cooled to room temperature, IPTG added to a final concentration of 200 μM,
and incubated with shaking (250 rpm) at 18 °C for 16 h. Cells were harvested by centrifugation
(17,000 × g, 20 min, 4 °C) and the pellet was resuspended in 40 ml of binding buffer (50 mM Tris,
500 mM NaCl, 5 mM imidazole, pH 8) containing protease inhibitor (Roche complete EDTA-free
protease inhibitor cocktail) and lysozyme (0.1 mg/ml) by nutating at 4°C for 30 min. Benzonase (1
μl, 25 U) was added to the mixture and lysis was effected by sonication. The lysate was centrifuged
(17000 × g, 20 min, 4 °C) and the supernatant collected, filtered (0.45 μm) and loaded onto a 1 ml
gravity IMAC column (GE HiTrap TALON). The column was washed with 15 ml binding buffer,
then the protein was eluted using elution buffer (50 mM Tris, 500 mM NaCl, 500 mM imidazole,
pH 8). Fractions containing product (as determined by SDS-PAGE) were combined and further
purified by size exclusion chromatography (GE HiLoad 16/600 Superdex 200) using 50 mM Tris,

11
150 mM NaCl, pH 7.5 buffer. Fractions containing monomeric product (as determined by SDS-
PAGE on non-reduced samples) were concentrated (5 mg ml–1
) and incubated with HRV 3C
protease (50 μg ml–1
), 1 mM EDTA, 2 mM reduced glutathione and 1 mM oxidized glutathione at 4
°C for 16 h. The liberated TSR domains were purified by size exclusion chromatography (GE
HiLoad 16/600 Superdex 75 using 50 mM Tris, 150 mM NaCl, pH 7.5 buffer) and ion exchange
chromatography (GE MonoQ 5/50 GL using 20 mM Tris, pH 7.5 with a NaCl gradient).
P. vivax POFUT2 expression and purification
A codon-harmonised dsDNA polynucleotide encoding residues 22–481 of P. vivax POFUT2
(PVX_098900) with an N-terminal gp67 signal peptide, His10 purification / V5 epitope tag, and
SUMO fusion partner was synthesized (Integrated DNA Technologies) and cloned into pFastBac
(Invitrogen) at the RsrII and XhoI restriction sites to give pFastBac-SUMOPvPOFUT2.
>SS-tags-SUMO-PvPOFUT
CGGTCCGAAACCATGCTACTAGTAAATCAGTCACACCAAGGCTTCAATAAGGAACACACAAGCAAGATGGTAAG
CGCTATTGTTTTATATGTGCTTTTGGCGGCGGCGGCGCATTCTGCCTTTGCGGCGGATCTTGGATCCCATCACC
ACCACCACCATCACCATCATCACGGTAAACCGATACCAAATCCCCTGCTCGGCCTCGACTCGACTATCGACGGT
CGCTCCGATTCCGAAGTAAACCAGGAGGCCAAACCTGAGGTCAAACCGGAAGTGAAACCCGAAACTCACATCAA
CTTGAAGGTCAGCGACGGCAGTTCGGAAATCTTCTTCAAAATCAAAAAAACTACCCCTTTGCGTAGACTTATGG
AAGCTTTCGCTAAGCGTCAGGGTAAAGAGATGGACTCCCTGACCTTTCTGTACGATGGAATTGAAATCCAGGCC
GACCAAACCCCTGAGGACCTCGACATGGAGGACAACGATATAATTGAGGCTCACCGCGAACAGATCGGTGGAGA
ATTCAATCTTGATAAGGATGTAAGTTCCCCCGTTTGTAGAACTGACGACGTATACACTGGAGACGCCTTTTACC
CTTTCAAGAAAAAGAAATACGTACTGTACGATGTGCATATCGGAGAGGGATTCAATCTTCAAAAGGAAGTTCTC
TACAGGGTCGCCCTTGCCGTGTACTATCTGAACCAGGAGGAACGCACACATGTGCACTACCTGGTACTCCCTCC
TTGGTGCTACGTTACGCACTGGGGACGTGAGAGGACTAACGCCAGGATCAAGTGGTCTATCTTTTTCAATCTGA
AAGCCCTGCAGAACGTAATCCCAGTTATGGAGTACGCGGAATACGAAGGACAGTTCGGCCCACATACAGATTAC
ATACTCTCGTACCGTCACATTATCGGCGAATGGCCCAAGCGTGGAGACAAGAAGAGCTTCCAGGTTCTCAAGTT
GGATAAGTGTCAAGTAAAGGGCTACAAGTTGAAGAAGAATCTGAGGAAGAACTGTGACCACAAGTACTCTGTTG
AGTATAGTGGAAAATGTACCAATGTCAAGGGCAAGAAAATGGAGTGCCTCGAATTTTTCTTTATTACATCACAT
TTCGTGTCCTCAACCCTTCTCGATATCTTCCAGTACGACGCTGATTCGGTGCTTATCAAGCACGGTAGTAACAT
CTTGGTGGCCTTCATGAACGAGCTTGTCGATGCTAATCTCGAAGACGTCCTGCCTTACTCCGAAGATCTGATCA
ACGAGGGCGACCAATTCGTCGAGAAAAATTTCAAGAGTTCCAAGAATTATATCAGCTGTCACCTTAGGTACACT
GATTTCCGCAAAATCTCCACTTACGACGTCAGCCCAGTGGGTATTAGCCTTCTGAAGCTGTTGTATATTATGTT
TCTGCGTAAGTCAACGTTGATCTTCGTCTCTACTGACGAAAAGAAGGAAGTGAAAAAAGTGATCGACAGCCAGT
TCCCTCAATTCAAGCATTTCTTCTTCTTCTACGAGAACGAAAAACTGCACACAGGACAAGTCGCAATAGTCGAC
CAGTGGATATGTGCACGCTCCGGAACATTCGTCGGCAACATTTTCAGTCGCTTCTCAATGCACATTAAGTGGGA
AAGATCCTTGATTGGAAAGGGCGGTCCAGATCACAACCTCGACTTGTGCGGCTACTCAATCTCGACTAATCACG
AACTGCGTAAAAAGTACAGTGATGTCCAAGACGCACATTTGGACGAGGAGGCTCTTCAGAAGCTGCGTCCACTC
TACATGCGCCTGTCTCAAAAAGACCGCGACTTCCTTAGGACCATCTGTTACGATTTCGCTCACTACTACCCACA
AAACGTCTCTATATACAGACGTGGAGAGAGGAGGGAGGAAGGCAGGTTGATGTGACTCGAG
Bacmid and baculovirus was prepared in accordance with the manufacturer’s instructions for the
‘Bac-To-Bac Baculovirus Expression System’. Sf21 cells (ThermoFisher Scientific #11497013, 1.8
L, 1.8×106 cells/ml) were inoculated with P3 virus stock (200 ml) and incubated (27 °C, 110 rpm)
for 72 h. The culture was centrifuged (300 × g, 5 min, 4 °C) and the supernatant filtered (0.22 m),
concentrated ten-fold (Amicon stirred cells, 10 kDa NMWL), dialysed (2×2.0 L of 50 mM Tris,
300 mM NaCl, pH 8.2) and loaded onto a 1 ml gravity IMAC column (GE HiTrap TALON). The

12
column was washed with 15 ml buffer (50 mM Tris, 300 mM NaCl, pH 8.2), then the protein was
eluted using elution buffer (50 mM Tris, 300 mM NaCl, 500 mM imidazole, pH 8.2). Fractions
containing product (as determined by SDS-PAGE) were combined and further purified by
immunoaffinity chromatography using anti-V5 tag antibody agarose (Abcam, ab1229).
Intact protein mass spectrometry
Protein samples were prepared for intact protein analysis using a modified SP3 clean-up
procedure7. Briefly, samples were acidified with formic acid (final concentration 2%, 5 mM DTT,
final volume 30 l) and incubated for 1 h at RT with 3 l of SP3 beads and an equal volume of
acetonitrile. Proteins were precipitated on to beads overnight with 240 l ice-cold acetone at –20
C. Beads were pelleted via magnetic capture and washed with 200 l of ice-cold 80% acetone
three times. Proteins were eluted with ice-cold 80% formic acid and rapidly diluted with 270 l
H2O to prevent acid induced modifications. Eluted samples were snap-frozen, dried by
lyophilisation and stored at –20 C. Intact analysis was performed using a Maxis II ETD UHR-
QqTOF mass spectrometer (Bruker Daltonics, Bremen, Germany) equipped with a Captivespray
source and NanoBooster infusing acetonitrile at 0.2 ml.min-1
to enhance MS signal intensity.
Protein samples were resuspended in 2% formic acid and immediately loaded onto a ProSwift RP-
4H 200 mm 0.1 mm column (Thermo Scientific) and washed with buffer A (3% acetonitrile,
0.1% formic acid) for 10 min and then eluted with a linear gradient from 3–85% buffer B (99.9%
acetonitrile, 0.1% formic acid) over 35 min at a flow rate of 1 l.min-1
. MS1 Mass spectra were
acquired at 1 Hz between a mass range of 150–2200 m/z. Intact mass analysis and deconvolution
was performed using DataAnalysis 4.3 (Bruker).
Glycosylation site localization by mass spectrometry
Digestion of O-fucosylated TRAP: In vitro O-fucosylation samples were resuspended in 6 M urea, 2
M thiourea, 40 mM NH4HCO3 and reduced then alkylated8 before being diluted 10-fold into 100
mM ammonium bicarbonate prior to the addition of GluC (1/50 w/w, Promega) and incubation
overnight at 25°C. Digested samples were acidified to a final concentration of 0.5% formic acid and
desalted using C18 stage tips 9,10
before analysis by LC-MS.
MS-MS Identification of O-fucosylation using reversed phase LC-MS: Purified peptides were re-
suspended in buffer (2% acetonitrile, 0.1% TFA) and separated using a two-column
chromatography set up composed of a PepMap100 C18 20 mm × 75 μm trap and a PepMap C18 500
mm × 75 μm analytical column (ThermoScientific). Samples were concentrated onto the trap

13
column (5 μl.min-1
) with buffer A (2% acetonitrile, 0.1% formic acid) for 5 min and infused into an
Orbitrap Fusion™ Lumos™ Tribrid™ mass spectrometer (ThermoScientific) at 300 nl.min-1
via the
analytical column using an Dionex Ultimate 3000 UPLC (ThermoScientific). Sixty min gradients
were run altering the buffer composition from 1% to 28% buffer B (80% acetonitrile, 0.1% formic
acid) over 35 min, then from 28% to 40% buffer B over 10 min, then from 40% to 100% buffer B
over 2 min, the composition was held at 100% buffer B for 3 min, and then dropped to 3% buffer B
over 5 min and held at 3% buffer B for another 10 min. The Lumos™ mass spectrometer was
operated in a data-dependent mode automatically switching between the acquisition of a single
Orbitrap MS scan (120,000 resolution) every 3 seconds and four different MS-MS approaches of
the same precursor: HCD (Normalized collision energy of 35 with a maximum fill time of 80 ms or
AGC of 2×105 analyzed within the Orbitrap at a resolution of 30K, EThcD (Predictive ETD
reaction times were enabled 11
and a normalized collision energy of 25 set, with a maximum fill
time of 120 ms or AGC 2×105 analyzed within the Orbitrap at a resolution of 30K), CID
(Normalized collision energy of 30 with a maximum fill time 80 ms or AGC 2×105 using Ion trap
based detection) and ETD (Predictive ETD reaction times were enabled 11
with a maximum fill
time of 120 ms or AGC 2×105 using Ion trap based detection).
Data Analysis: Identification of O-fucosylated peptides was accomplished using MaxQuant
(v1.5.3.1) 12
. Searches were performed against the amino acid sequence of the PfTRAP protein with
GluC cleavage specificity allowing 2 miss cleavage events and the variable modifications of
oxidation of methionine, O-fucosylation addition to serine or threonine and acetylation of protein
N-termini. The precursor mass tolerance was set to 20 parts-per-million (ppm) for the first search
and 10 ppm for main search, with a maximum false discovery rate (FDR) of 1.0% set for protein
and peptide identifications. The resulting outputs were processed within the Perseus (v1.4.0.6) 13
analysis environment to remove reverse matches and common proteins contaminates prior to
further analysis.
Transgenic parasites
To HA-tag the C-terminus of P. falciparum POFUT2, the 3 of the POFUT2 locus was amplified
from P. falciparum NF54 gDNA using POFUT2HA_F and POFUT2HA_R primers. The amplicon
was cloned into p1.2-SHA 14
via BamHI/PstI. Purified plasmid DNA (80 g, Life Technologies)
was transfected into P. falciparum NF54 and stable transfectants selected as described previously
15. Integration of the cassette was confirmed by Southern blot analysis using the Roche digoxigenin
(DIG) system according to the manufacturer’s instructions.

14
To disrupt the P. falciparum POFUT2 locus, 5 and 3 flanks were amplified from P. falciparum
NF54 gDNA using primers POFUT2KO_FL1_F and POFUT2KO_FL1_R for flank 1 and
POFUT2KO_FL2_F and POFUT2KO_FL2_R for flank 2. Primer sequences are provided in
Supplementary Table 1. Flanks were cloned into pCC1 via SacII/SpeI (5 flank) and EcoRI/AvrII
(3 flank) to make the knockout construct. Transfected lines were cloned by limiting dilution and
analyses were performed on these clonal parasite lines as described above 16
.
Supplementary References
1 Chen, C. I. et al. Structure of human POFUT2: insights into thrombospondin type 1 repeat
fold and O-fucosylation. EMBO J 31, 3183-3197, doi:10.1038/emboj.2012.143 (2012).
2 Valero-Gonzalez, J. et al. A proactive role of water molecules in acceptor recognition by
protein O-fucosyltransferase 2. Nat Chem Biol 12, 240-246, doi:10.1038/nchembio.2019
(2016).
3 Kelley, L. A., Mezulis, S., Yates, C. M., Wass, M. N. & Sternberg, M. J. The Phyre2 web
portal for protein modeling, prediction and analysis. Nature protocols 10, 845-858,
doi:10.1038/nprot.2015.053 (2015).
4 Lyskov, S. & Gray, J. J. The RosettaDock server for local protein-protein docking. Nucleic
Acids Res 36, W233-238, doi:10.1093/nar/gkn216 (2008).
5 Lyskov, S. et al. Serverification of molecular modeling applications: the Rosetta Online
Server that Includes Everyone (ROSIE). PLoS One 8, e63906,
doi:10.1371/journal.pone.0063906 (2013).
6 Combs, S. A. et al. Small-molecule ligand docking into comparative models with Rosetta.
Nature protocols 8, 1277-1298, doi:10.1038/nprot.2013.074 (2013).
7 Hughes, C. S. et al. Ultrasensitive proteome analysis using paramagnetic bead technology.
Mol Syst Biol 10, 757, doi:10.15252/msb.20145625 (2014).
8 Scott, N. E. et al. Simultaneous glycan-peptide characterization using hydrophilic
interaction chromatography and parallel fragmentation by CID, higher energy collisional
dissociation, and electron transfer dissociation MS applied to the N-linked glycoproteome of
Campylobacter jejuni. Mol Cell Proteomics 10, M000031-MCP000201,
doi:10.1074/mcp.M000031-MCP201 (2011).
9 Ishihama, Y., Rappsilber, J. & Mann, M. Modular stop and go extraction tips with stacked
disks for parallel and multidimensional Peptide fractionation in proteomics. J Proteome Res
5, 988-994, doi:10.1021/pr050385q (2006).

15
10 Rappsilber, J., Mann, M. & Ishihama, Y. Protocol for micro-purification, enrichment, pre-
fractionation and storage of peptides for proteomics using StageTips. Nature protocols 2,
1896-1906, doi:10.1038/nprot.2007.261 (2007).
11 Rose, C. M. et al. A calibration routine for efficient ETD in large-scale proteomics. J Am
Soc Mass Spectrom 26, 1848-1857, doi:10.1007/s13361-015-1183-1 (2015).
12 Cox, J. & Mann, M. MaxQuant enables high peptide identification rates, individualized
p.p.b.-range mass accuracies and proteome-wide protein quantification. Nature
biotechnology 26, 1367-1372, doi:10.1038/nbt.1511 (2008).
13 Tyanova, S., Temu, T. & Cox, J. The MaxQuant computational platform for mass
spectrometry-based shotgun proteomics. Nature protocols 11, 2301-2319,
doi:10.1038/nprot.2016.136 (2016).
14 Boddey, J. A. et al. An aspartyl protease directs malaria effector proteins to the host cell.
Nature 463, 627-631, doi:10.1038/nature08728 (2010).
15 Yang, A. S. P. et al. Cell Traversal Activity is Required for Plasmodium falciparum Liver
Infection in Humanized Mice. Cell reports 18, 3105-3116, doi:10.1016/j.celrep.2017.03.017
(2017).
16 Sleebs, B. E. et al. Inhibition of Plasmepsin V activity demonstrates its essential role in
protein export, PfEMP1 display, and survival of malaria parasites. PLoS Biol 12, e1001897,
doi:10.1371/journal.pbio.1001897 (2014).