fen bİlİmlerİ enstİtÜsÜ doktora tezİye’den halofİk...
TRANSCRIPT

ANKARA ÜNİVERSİTESİ
FEN BİLİMLERİ ENSTİTÜSÜ
DOKTORA TEZİ
TÜRKİYE’DEN HALOFİLİK ARKEBAKTERİLERİN İZOLASYONU VE
KARAKTERİZASYONU
Birgül ÖZCAN
BİYOLOJİ ANABİLİM DALI
ANKARA
2004
Her hakkı saklıdır

Prof Dr. Cumhur ÇÖKMÜŞ danışmanlığında, Birgül ÖZCAN tarafından hazırlanan bu
çalışma 29/06/2004 tarihinde aşağıdaki jüri tarafından Biyoloji Anabilim Dalı’nda
doktora tezi olarak kabul edilmiştir.
Başkan :Prof. Dr. Şevki YAZGAN
Üye :Prof. Dr. Cumhur ÇÖKMÜŞ
Üye :Prof. Dr. Gülay ÖZCENGİZ
Üye :Prof. Dr. Mustafa AKÇELİK
Üye :Prof. Dr. Yavuz BEYATLI
Yukarıdaki sonucu onaylarım
Prof. Dr. Metin OLGUN
Enstitü Müdürü

İÇİNDEKİLER
ÖZET............................................................................................................................ i
ABSTRACT................................................................................................................. ii
TEŞEKKÜR................................................................................................................. iii
SİMGELER DİZİNİ.................................................................................................... vi
ŞEKİLLER DİZİNİ..................................................................................................... vii
ÇİZELGELER DİZİNİ................................................................................................ x
1. GİRİŞ............................................................................................................... 1
2. KAYNAK ÖZETLERİ................................................................................... 3
2.1. Mikrobiyal Filogenetik.................................................................................. 3
2.2. Halofilik Arkelerin Taksonomisi ve Teşhisi................................................. 7
2.2.1. Halobacteriales ordosunun özellikleri...................................................... 10
2.3. Halofilik Arkelerin Habitatları ve Ekolojisi................................................. 12
2.4. Halofilik Arkelerde Ozmotik Adaptasyon.................................................... 19
2.5. Halofilik Arkelerin Yapısal, Genetik ve Fizyolojik Özellikleri................... 20
2.5.1. Polar ve nötral lipitler................................................................................ 20
2.5.2. Karotenoid ve retinal pigmentler............................................................... 22
2.5.3. Diğer yapısal özellikleri............................................................................. 23
2.5.4. Genetik yapıları......................................................................................... 23
2.5.5. Fizyolojileri............................................................................................... 24
2.6. Halofilik Arkelerin Antimikrobiyal Maddelere Duyarlılıkları..................... 25
2.7. Halofilik Arkelerin Uygulama Alanları........................................................ 25
3. MATERYAL VE YÖNTEM........................................................................ 28
3.1. Materyal........................................................................................................ 28
3.1.1. Halofilik arkeler......................................................................................... 28
3.1.2 Besiyeri ...................................................................................................... 28
3.1.3. Mikroskobik inceleme............................................................................... 29
3.1.4. Antibiyotikler............................................................................................. 29
3.1.5. İnce tabaka kromatografisi........................................................................ 29
3.1.6. Elektroforez............................................................................................... 29
3.2. Yöntem......................................................................................................... 30
iv

3.2.1. Halofilik arkelerin izolasyonu ve kültürü.................................................. 30
3.2.2. Koloni morfolojisi ve pigmentasyon......................................................... 30
3.2.3. Gram boyama ve faz-kontrast mikroskobu............................................... 31
3.2.4. Biyokimyasal testler.................................................................................. 31
3.2.5. Antibiyogram testi..................................................................................... 33
3.2.6. İnce tabaka kromatografisi........................................................................ 33
3.2.7. Protein ekstraksiyonu................................................................................ 33
3.2.7.1. Sodyum dodesil sülfat poliakrilamid jel elektroforezi (SDS-PAGE)..... 34
3.2.7.2. Proteinlerin moleküler kütlelerinin belirlenmesi.................................... 34
3.2.7.3. Benzerlik matriksi ve protein ilişkisine göre benzerlik dendogramı...... 34
3.2.8. Plazmid izolasyonu.................................................................................... 35
3.2.8.1. Agaroz Jel elektroforezi.......................................................................... 36
3.2.8.2. Plazmid büyüklüklerinin saptanması...................................................... 36
4. ARAŞTIRMA BULGULARI VE TARTIŞMA.......................................... 38
4.1. Halofilik Arke İzolatları............................................................................... 38
4.2. Halofilik Arke İzolatlarının Gram Reaksiyonu, Hareket ve Morfolojik
Özellikleri..................................................................................................... 39
4.3. Halofilik arke İzolatlarının Antibiyotik Duyarlılıkları................................. 56
4.4. Halofilik Arke İzolatlarının Hücre zarı Eter-Lipit Analizleri....................... 62
4.5. Halofilik Arke İzolatlarının Biyokimyasal Özellikleri................................. 69
4.6. Halofilik Arke İzolatalarının Plazmid İçerikleri........................................... 75
4.7. Halofilik Arke İzolatlarının Protein profilleri ve Benzerlik Dendogramı.... 89
KAYNAKLAR.......................................................................................................... 105
EKLER...................................................................................................................... 123
EK 1.................................................................................................................... 124
EK 2.................................................................................................................... 126
ÖZGEÇMİŞ............................................................................................................... 128
v

SİMGELER DİZİNİ
APS Amonyum Persülfat
cccDNA Kovalent bağlı dairesel deoksiribonükleik asit
dk Dakika
DNA Deoksiribonükleik Asit
EDTA Etilen Diamin Tetraasetik Asit
FeSO4.H2O Demir sülfat
g Gram
H2O2 Hidrojen peroksit
kb Kilobaz
kDa Kilodalton
KNO3 Potasyum nitrat
L Litre
mA Miliamper
MDa Megadalton
Me-PGP Fosfatidilgliserol Fosfat-Metilester
mg Miligram
MgSO4.7H2O Magnezyum sülfat
mL Mililitre
M Molar
Na2S2O3.5H2O Sodyum tiyosülfat
RNA Ribonükleik Asit
rpm Dakikadaki dönüş sayısı
SDS Sodyum Dodesil Sülfat
SDS-PAGE Sodyum Dodesil Sülfat-Poliakrilamid Jel Elektroforezi
SG Sehgal-Gibbons Besiyeri
TEMED Tetrametil Etilen Diamin
V Volt
µg Mikrogram
µL Mikrolitre
vi

ŞEKİLLER DİZİNİ
Şekil 2.1. Ribozomal küçük alt birimde bulunan 16S ve 18S rRNA dizilerine göre
oluşturulmuş filogenetik ............................................................................. 5
Şekil2.2. Halobacteriaceae familyasının 16S rRNA dizi analizlerine göre
oluşturulan filogenetik ağacı........................................................................ 8
Şekil 2.3. Halofilik arkelerde bulunan temel lipitleri................................................. 21
Şekil 4.1. %25 NaCl içeren SG agar besiyerinde 37 °C’de 7 gün geliştirilen A317
izolatı ve Haloferax mediterranei standart suşunun koloni morfolojisi..... 45
Şekil 4.2. %25 NaCl içeren SG agar besiyerinde 37 °C’de 7 gün geliştirilen A128
izolatı ve Haloferax volcanii standart suşunun koloni morfolojisi............. 46
Şekil 4.3. %25 NaCl içeren SG sıvı besiyerinde 37 °C’de 7 gün geliştirilen
halofilik arke izolatları ve standart suşun üremesi..................................... 47
Şekil 4.4. SG sıvı besiyerinde 37 °C’de 120 devir/dk.’da 6 gün geliştirilen
Halobacterium salinarum ve Natrialba asiatica standart suş hücrelerinin
faz-kontrast mikroskobik görünümleri ....................................................... 49
Şekil 4.5. SG sıvı besiyerinde 37 °C’de 120 devir/dk.’da 6 gün geliştirilen
Haloferax volcanii ve Haloferax mediterranei standart suş hücrelerinin
faz-kontrast mikroskobik görünümleri....................................................... 50
Şekil 4.6. SG sıvı besiyerinde 37 °C’de 120 devir/dk.’da 6 gün geliştirilen
Haloarcula vallismortis DSM 3756 ve Haloarcula marismortui
standart suş hücrelerinin Faz-kontrast mikroskobik görünümleri............... 51
Şekil 4.7. SG sıvı besiyerinde 37 °C’de 120 devir/dk.’da 6 gün geliştirilen Halococcus
morrhuae CCM 537 standart suşu ve E133 numaralı izolat hücrelerinin
faz-kontrast mikroskobik görünümleri....................................................... 52
Şekil 4.8. SG sıvı besiyerinde 37 °C’de 120 devir/dk.’da 6 gün geliştirilen E22
ve A29 numaralı izolat hücrelerinin faz-kontrast mikroskobik
görünümleri................................................................................................ 53
Şekil 4.9. SG sıvı besiyerinde 37 °C’de 120 devir/dk.’da 6 gün geliştirilen D107
ve A317 numaralı izolat hücrelerinin faz-kontrast mikroskobik
görünümleri................................................................................................ 54
vii

Şekil 4.10. SG sıvı besiyerinde 37 °C’de 120 devir/dk.’da 6 gün geliştirilen A347
ve B44A numaralı izolat hücrelerinin faz-kontrast mikroskobik
görünümleri.............................................................................................. 55
Şekil 4.11. SG agar besiyerinde 37°C’de 14 gün geliştirilen Haloferax
mediterranei ve A440 numaralı halofilik arke izolatının antibiyotik
duyarlılığı................................................................................................... 58
Şekil 4.12. SG agar besiyerinde 37°C’de 14 gün geliştirilen F89 ve A128 numaralı
halofilik arke izolatlarının antibiyotik duyarlılığı.................................... 59
Şekil 4.13. Halofilik arke izolat ve standart suşlarına ait hücre metanolizatlarının
ince tabaka kromatogramı ....................................................................... 63
Şekil 4.14. Halofilik arke izolat ve standart suşlarına ait hücre metanolizatlarının
ince tabaka kromatogramı ....................................................................... 64
Şekil 4.15. Halofilik arke izolat ve standart suşlarına ait hücre metanolizatlarının
ince tabaka kromatogramı ....................................................................... 65
Şekil 4.16. Halofilik arke izolatlarına ait hücre metanolizatlarının ince tabaka
kromatogramı............................................................................................ 66
Şekil 4.17. Halofilik arke izolatlarına ait hücre metanolizatlarının ince tabaka
kromatogramı........................................................................................... 67
Şekil 4.18. Halofilik arke izolatlarına ait hücre metanolizatlarının ince tabaka
Kromatogramı........................................................................................... 68
Şekil 4.19. Halofilik arke izolatları ve standart suşların plazmid içerikleri............... 76
Şekil 4.20. Halofilik arke izolatlarının plazmid profilleri.......................................... 77
Şekil 4.21. Halofilik arke izolatlarının plazmid profilleri.......................................... 78
Şekil 4.22. Halofilik arke izolatlarının plazmid profilleri.......................................... 79
Şekil 4.23. Halofilik arke izolatlarının plazmid profilleri.......................................... 80
Şekil 4.24. Halofilik arke izolatlarının plazmid profilleri.......................................... 81
Şekil 4.25. Halofilik arke izolatlarının plazmid profilleri.......................................... 82
Şekil 4.26. Halofilik arke izolatlarının plazmid profilleri.......................................... 83
Şekil 4.27. Halofilik arke izolat ve standart suşlarının SDS-PAGE protein
profillerine göre oluşturulan benzerlik dendogramı................................. 90
Şekil 4.28. Halofilik arke izolatlarının SDS-PAGE protein profilleri........................ 91
Şekil 4.29. Halofilik arke izolatlarının SDS-PAGE protein profilleri........................ 92
viii

Şekil 4.30. Halofilik arke izolatlarının SDS-PAGE protein profilleri........................ 93
Şekil 4.31. Halofilik arke izolatlarının SDS-PAGE protein profilleri........................ 94
Şekil 4.32. Halofilik arke izolatlarının SDS-PAGE protein profilleri........................ 95
Şekil 4.33. Halofilik arke izolat ve standart suşlarının SDS-PAGE protein
profilleri................................................................................................... 96
Şekil 4.34. Halofilik arke izolatlarının SDS-PAGE protein profilleri........................ 89
ix

ÇİZELGELER DİZİNİ
Çizelge 2.1. Bakteri, Arke ve Ökaryotlar arasındaki bazı temel farklılıklar............... 4
Çizelge 2.2. Halobacteriales ordosuna ait cinsler ve türler...................................... 11
Çizelge 2.3. Halobacterium, Halobaculum, Halogeometricum, Halococcus ve
Halorhabdus cinslerine ait türlerin özellikleri ....................................... 13
Çizelge 2.4. Halorubrum cinsine ait türlerin özellikleri ........................................... 14
Çizelge 2.5. Halorubrum, Haloarcula ve Halomicrobium cinsine ait türlerin
özellikleri ............................................................................................... 15
Çizelge 2.6. Haloferax ve Halosimplex cinslerine ait türlerin özellikleri................ 16
Çizelge 2.7. Halobioforma, Natronococcus, Natrinema ve Natronorubrum
cinslerine ait türlerin özellikleri ........................................................... 17
Çizelge 2.8. Natrialba, Haloterrigena, Natronomonas ve Natronobacterium cinslerine
cinslerine ait türlerin özellikleri............................................................. 18
Çizelge 4.1. Halofilik Arke izolatlarının bölge ve örneğe göre dağılımı................... 39
Çizelge 4.2. Halofilik arke izolatlarının Gram reaksiyonu, koloni morfolojisi,
pigmentasyon, hücre morfolojisi ve hareket testi sonuçları................... 40
Çizelge 4.3. Halofilik arke izolatlarının Gram reaksiyonu, mukoid koloni özelliği,
hareket ve hücre şekillerinin genel sayısal sonuçları............................. 43
Çizelge 4.4. Halofilik arkebakteri izolat ve standart suşlarının genel antibiyotik
duyarlılıkları........................................................................................... 56
Çizelge 4.5. Halofilik arke izolatlarının biyokimyasal özellikleri.............................. 70
Çizelge 4.6. Halofilik arke izolatlarının sayısal biyokimyasal (+) test sonuçları....... 73
Çizelge 4.7. Halofilik arkebakteri izolat ve suşlarının plazmid içerikleri.................. 84
Çizelge 4.8. Halofilik arke izolat ve standart suşlarının SDS-PAGE protein göre
profillerine oluşturulan benzerlik matriksi............................................. 89
x

ÖZET
Doktora Tezi
TÜRKİYE’DEN HALOFİLİK ARKEBAKTERİLERİN İZOLASYONU VE KARAKTERİZASYONU
Birgül ÖZCAN
Ankara Üniversitesi
Fen Bilimleri Enstitüsü Biyoloji Anabilim Dalı
Danışman: Prof. Dr. Cumhur ÇÖKMÜŞ
Tuz gölü (Ankara), Acı göl (Denizli), Salda gölü (Burdur), Seyfe gölü (Kırşehir), Tuzla gölü (Kayseri) ve Bolluk gölü (Konya) olmak üzere Türkiye’nin altı farklı tuzlu bölgesinden toplanan toprak, su ve tuz kütlesi örneklerinden %25 (w/v) NaCl içeren kompleks besi yeri kullanılarak 95 adet halofilik arke izole edilmiştir. Antibiyotik duyarlılığı ve zar-eter lipit analizleri ile arke olarak tanımlanan izolatlar, hareket, pigmentasyon, Gram reaksiyonları, koloni ve hücre morfolojisi, biyokimyasal testler, elektroforetik protein profilleri ve plazmid içerikleri yönünden incelenmiştir. Buna göre izolatların %83.2’sinin hareketli, %96.8’inin Gram (-), %62’sinin çubuk ve %19’unun pleomorfik hücre morfolojisine sahip olduğu görülmüştür. İzolatların, genelde yuvarlak, düz kenarlı, konveks koloni morfolojisine sahip oldukları ve açık pembeden kırmızıya kadar değişen renk tonlarında pigmentasyon gösterdikleri bulunmuştur. Çeşitli biyokimyasal testler sonucunda halofilik arke izolatlarının tümünün katalaz ve oksidaz aktivitesine ve sadece 2 izolatın L-arjininde anaerobik üreme yeteneğine sahip oldukları tespit edilmiştir. Bunların dışındaki biyokimyasal testlerde %85.3-2.1 aralığında pozitif sonuçlar elde edilmiştir. Antibiyotik duyarlılık testi sonucuna göre tüm izolatların bazitrasin, novobiyosin ve rifampisin’e duyarlı oldukları saptanmıştır. İzolatlardan 60’nın 1-6 arasında değişen sayıda ve 1.0-36.9 kb arasında değişen farklı moleküler büyüklüklerde plazmid içerdikleri belirlenmiştir. Ayrıca izolatların tüm hücre protein profilleri SDS-PAGE ile çıkarılarak MVSP analiz programı ve UPGMA metodu kullanılarak karşılaştırılmış, benzerlik dendogramı çıkarılmıştır. Buna göre izolatlar 8 gruba ayrılmış, 10 adet izolatın standart suşlarla birlikte tek bir grup içinde toplandıkları ve bu grup içerisinde 4 izolatın diğerleri farklı gruplar oluşturduğu halde standart suşlara benzerlik oranlarının yüksek olduğu görülmüştür. 2004, 128 sayfa ANAHTAR KELİMELER: Halofilik arkebakteriler, izolasyon, karakterizasyon, plazmid, SDS-PAGE, Türkiye
i

ABSTRACT
Ph. D. Thesis
ISOLATION AND CHARACTERIZATION OF HALOPHILIC ARCHAEBACTERIA FROM TURKEY
Birgül ÖZCAN
Ankara Universty
Graduate School of Natural and Applied Sciences Department of Biology
Supervisor: Prof. Dr. Cumhur ÇÖKMÜŞ
Ninetyfive halophilic archaeal isolates were obtained from saline soil, brine and halit samples collected from six distinct regions of Turkey by using complex medium containing 25% NaCl (w/v). The isolates identified as archae by antibiotic suscebtibility and membrane ether-lipid analysis, were tested for motility, Gram reaction, pigmentation, cell and colony morphology, biochemical reactions, electrophoretic protein profiles and plasmid contents. The results indicated that 83,2% of the isolates were motile, 96.8% were Gram (-), 62% were rod shaped and 19% were in pleomorphic cell morphology. The most of the isolates had circular, entire and convex shaped colony morphology and their pigmentation were observed from light pink to red color. As a result of biochemical tests all halophilic archaea isolates were determined to be catalase and oxidase-positive, and only 2 isolates had anaerobic growth ability on L-argine. To other biochemical tests, the positive reactions were observed in 2.1-85.3% range. According to the antibiotic susceptibility tests, all isolates were susceptible to bacitracin, novobiocine and rifampicine. It was established that the 60 halophilic archaeal isolates contained plasmids varied in the numbers from 1 to 6 and in different molecular sizes ranged from 1 to 36.9 kb. On the other hand, whole-cell protein profiles of all isolates were obtained by SDS-PAGE, and the similarity dendograme was constructed from protein profiles by means of UPGMA method using MVSP analyses programme. Standard strains were clustured in 8 groups, and ten of our isolates were placed in the same group with standard strains while others formed distinct groups.
2004, 128 pages Key Words: Halofilik archaebacteria, isolation, characterization, plasmids, SDS- PAGE, Turkey
ii

TEŞEKKÜR
Bana bu konuda çalışma imkanı sunan ve çalışmalarım aşamasında önerileri ile beni
yönledirip destek olan danışman hocam Sayın Prof. Dr. Cumhur ÇÖKMÜŞ’e;
Çalışmalarım sırasında değerli düşünce ve önerilerinden yararlandığım Tez izleme
komitesi üyeleri Sayın Prof. Dr. Gülay Özcengiz’e (Ortadoğu Teknik Üniversitesi Fen-
Edebiyat Fakültesi) ve Sayın Prof. Dr. Mustafa AKÇELİK’e (Ankara Ünivesitesi Fen
Fakültesi);
Her konuda yanımda bulunarak, destek ve yardımlarını esirgemeyen Sayın Doç. Dr.
Mahmut Çalışkan’a (Mustafa Kemal Üniversitesi Fen-Edebiyat Fakültesi);
Bana standart suşlar ve kaynak sağlayarak yardımcı olan ve bilgilerinden faydalandığım
Sayın Prof. Dr. Aharon OREN’e (Hebrew Universty Jerusalem İsrail);
Yardım ve desteğinden dolayı laboratuvar arkadaşım sayın Arş. Gör. Arzu ÇÖLERİ’e;
Laboratuvarında çalışma olanağı sağlayarak yardımcı olan Sayın Prof. Dr. Ongun
ONARAN’a (Ankara Üniversitesi Tıp Fakültesi), verilerin analizi aşamasında
yardımlarını gördüğüm Sayın Yrd. Doç. Dr. İrfan KANDEMİR (Karaelmas Üniversitesi
Fen-Edebiyat Fakültesi) ve Yrd. Doç. Dr. Volkan Çevik’e (Mustafa Kemal Üniversitesi
Fen-Edebiyat Fakültesi);
Bana herkonuda destek olan ve hiçbir konuda fedakarlıklarını esirgemeyen annem,
babam ve kardeşlerime;
En içten saygı ve teşekkürlerimi sunarım.
Birgül ÖZCAN
Ankara, Haziran 2004
iii

1. GİRİŞ
Bindokuyüzotuzlu yıllarda Edward Chatton, tüm canlıları evrimsel gelişmişlik
düzeyleri, benzerlikleri ve hücre morfolojileri yönünden prokayot ve ökaryot olmak
üzere iki gruba ayırmıştır. İlerleyen zaman içerisinde ribozomal RNA dizi analizlerini
kapsayan moleküler tekniklerin de kullanılmaya başlanması ile hücresel yaşamın üç ayrı
dalda evrimleştiği kabul edilmiş ve yaşamın üçüncü formu olan Arkebakteriler (arkeler)
1970’lerin sonunda tanımlanmıştır. Böylece canlılar prokaryotik olan Bakteri ve
Arkelerle Ökaryotlar olmak üzere 3 domaine ayrılmıştır.
Tuz, günlük yaşamımızda önemli bir madde olup daha çok yemeklerde kullanılmasının
yanısıra, yiyeceklerin ve deri, et gibi ürünlerin saklanması ve korunması için de
kullanılagelmiştir. Tuzlanmış, fakat renkleri kırmızıya dönerek bozulan balık ve
derilerde, organizmaların rolü ve bu zararların önlenmesi için çıkış yollarının bulunması
amacıyla yüksek tuzlulukta yaşayan organizmalara karşı ilk bilimsel merak ortaya
çıkmıştır. Buna dönük araştırmalarda ekstrem halofilik mikroorganizmaların sorumlu
oldukları gösterilmiştir. Daha sonra bu mikroorganizmalar Arke domainine bağlı
Euryarchaeota filumuna ait Halobacteriales ordosu içerisinde toplanmıştır.
Yüksek tuzlu ortamlardan izole edilen halofilik arkebakteri (halofilik arke) izolatlarının
Halobacteriales ordosuna dahil edilebilmesi için kültürel, morfolojik, fizyolojik ve
besinsel özellikleri, antibiyotiklere duyarlılıkları, hücre zarı lipitleri, 16S rRNA dizi
analizleri, %G+C oranları, protein profilleri gibi testlerin yapılması gerekmektedir.
Canlılıkları için minimum 1.5 M NaCl’e gereksinim duyan ve tuzlalar, yüksek tuzlu
topraklar, deniz ve tuz gölleri gibi doğal ortamların yanısıra tuzlanmış ürünlerden de
izole edilebilen halofilik arkeler; çubuktan üçgene, kareye kadar değişen hücre
morfolojilerine ve katalaz ve oksidaz aktivitelerine sahip, aerobik, Gram (-),
kemoorganotrofik karakterdedirler. Ayrıca bu organizmalar, dış proton pompası olan
bakteriodopsin ve iç klor pompası olan halorodopsin pigmentlerini içermeleriyle de
diğer canlılardan farklılık gösterirler.
1

Halobacteriales ordosunun ekstrem halofilik arkeleri organik maddelerin parçalanması
ve petrolün bioremediasyonu dahil olmak üzere elektronik, gıda, tıp ve birçok
biyokimyasal ve biyofiziksel çalışmalarda biyoteknolojik potansiyele ve uygulamalara
sahiptirler.
Günümüzde dünyanın değişik tuzlu bölgelerinden izole edilen halofilik arke
izolatlarının tanımlanması halen devam etmekte ve bu izolatlar Halobacteriales ordosu
içerisinde yeni cins ve türler olarak yerlerini almaktadırlar. Ülkemizde ise, halofilik
arkelerin izolasyonu ve tanımlanmaları ile ilgili olarak çok sınırlı düzeyde çalışma
mevcuttur.
Tüm bu verilen bilgiler doğrultusunda bu çalışma ile, Türkiye’nin 6 farklı tuzlu
bölgesinden toplanan toprak ve su örneklerinden halofilik arkelerin izolasyon ve
karakterizasyonlarının yapılması amaçlanmış olup bu izolatların kültürel ve
biyokimyasal özellikleri, hücre morfolojileri, antibiyotik duyarlılıkları, arkeal lipit ve
plazmid içerikleri, elektroforetik tüm hücre protein profilleri incelenerek izolatların
standart halofilik arke suşları ve birbirleriyle olan benzerlik ve farklılıkları
belirlenmiştir.
2

2. KAYNAK ÖZETLERİ
2.1. Mikrobiyal Filogenetik
Genel anlamdaki bitki/hayvan ayırımı 1866 yılında resmi olarak Haeckel tarafından
değiştirilmiş olup her iki kategoriye yerleştirilmemiş olan ve hem hayvan hem de
bitkilerden ayrılan özelliklere sahip olan tek hücreli formlar da (protistler) ele alınarak
canlılar 3’e ayrılmıştır. Daha sonraları 1930 yılında Edward Chatton canlıları ökaryot ve
prokaryot olmak üzere 2 kategoriye ayırmış ancak önerisi rededilmiştir. İleri sürdüğü bu
ayırıma karşı çıkılma nedeni temelde prokaryot ve ökaryot kavramının sitolojik temele
dayalı olmasıydı. Daha sonra Coepland, bakterileri de ekleyerek canlıları 4 dala
ayırmıştır. 1969 yılında ise Whitakker, Bitki, Hayvan, Protist, Monera (bakteri)
gruplamasına Fungi (mantarlar) alemini de ekleyerek canlıları 5 dala ayırmıştır. Bu
sınıflama yakın zamana kadar en çok kabul gören yaşamın temel organizasyon şeması
olmuştur (Woese et al. 1990, Woese 1999).
Özellikle mikroorganizmalar açısından moleküler yapılar ve dizilerin, klasik fenotipik
özelliklere göre evrimsel ilişkileri daha iyi ortaya çıkardığı görülmüştür. Bundan dolayı
taksonların tanımlanmasında temel alınan organizmal→hücresel seviye, moleküler
düzeye (16S ve 18S rRNA dizi karşılaştırmaları) kaydırılmıştır. Moleküler
karşılaştırmalar sonucu dünyadaki yaşam, 1977 yılında Carl Woese tarafından 3 ana
gruba ayrılmıştır: Bakteriler, Ökaryotlar ve Arkeler. Bu sınıflandırmada herbir kategori
"Domain" olarak adlandırılmış olup herbir domain 2 ya da daha fazla alem
içermektedir. Arke, Bakteri ve Ökaryotlara bağlı farklı alemler bulunduğu için daha
esnek ve daha açıklayıcı olmalarından dolayı en üst hiyerarşi seviyesini adlandırmak
amacıyla en yüksek taksona Domain adının verilmesi önerilmiştir (Woese et al. 1990).
Çizelge 2.1.’de Bakteri, Arke ve Ökaryotlar arasındaki bazı karakteristik farklılıklar
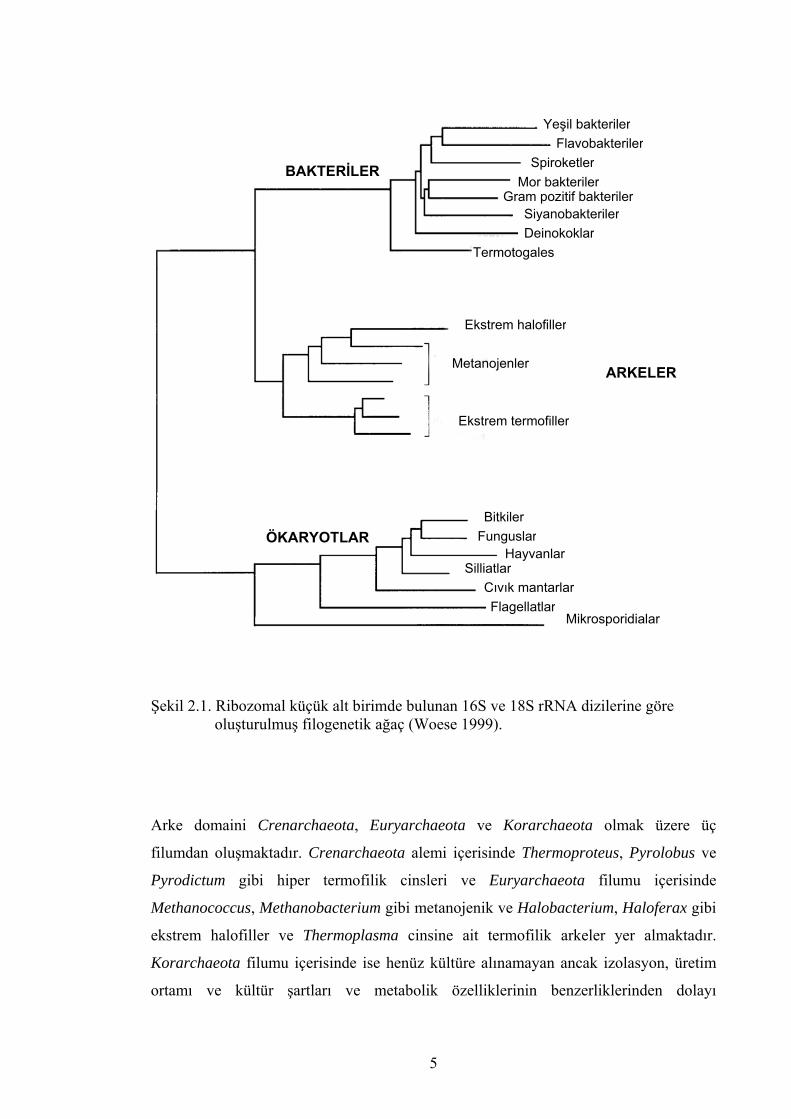
özetlenmiştir (Krieg 2001, Madigan et al. 2000, Woese 1999). Şekil 2.1.’de ise ribozom
küçük alt biriminde bulunan 16 ve 18S rRNA dizilerinin karşılaştırılmasıyla
oluşturulmuş evrensel filogenetik ağaç verilmiştir (Woese 1999).
3

Çizelge 2.1. Bakteri, Arke ve Ökaryotlar arasındaki bazı temel farklılıklar ÖZELLİKLER BAKTERİLER ARKELER ÖKARYOTLARMoleküler Özellikler Halkasal kromozom yapısı + + − Plazmid bulunuşu + + Nadir Ribozom sedimentasyon sabiti 70 S 70 S 80 S1
Ribozomal RNA sedimentasyon sabitleri
16S, 23S, 5S 16S, 23S, 5S 18S, 28S, 5.85S, 5S
RNA polimeraz sayısı 1adet(4 alt birim)
Birkaç adet(Herbiri 8-12 alt birim)
3 adet(Herbiri 12-14 alt birim)
Trinskripsiyon faktörü (EF-2) gerekliliği
− + +
Promotör yapıları TATAAT (Pribnow) kutusu
(-10 ve -35 sekansları)
TATA kutusu (-38 ve -25 sekansları)
TATA kutusu (-30 ve -25 sekansları)
Metabolik Özellikler Azot fiksasyonu + + − Denitrifikasyon + + − Metan oluşturma − + − Klorofille fotosentez + − + Kemolitotrofik metabolizma + + − 80°C üzerindeki sıcaklıkta ve 1.5 M’ın üzerindeki tuz konsantras- yonlarında üreme
+ + −
Kimyasal Analizlere Dayalı Özellikler
PHB üretimi + + − Peptidoglikan bulunuşu +2 − − Sterol bulunuşu (hücre zarlarında) + − + Zar lipitleri Gliserole ester bağlı yağ
asitleriGliserole eter bağlı
ftanil zincirleri Gliserole ester bağlı yağ
asitleriSitolojik Özellikler Prokaryotik hücre yapısı + + − Histon proteinleri − +3 9+2 düzeninde flagella yapısı − − + Başlangıç amino asiti N-formilmetiyonin Metiyonin Metiyonin 5'CAP ve poli-A uzantısı − − + Mitoz − − + Antibiyotiklere Duyarlılık Β-laktam, kloramfenikol, streptomisin
+ − −
Siklohekzimit − − + 1: Mitokondri ve kloroplast hariç, 2: Protein yapıda hücre duvarına sahip Chlamydiae ve Planctomycet’ler hariç, 3:Bazı tRNA genleri intron proteinleri içermektedir
Bakteri, Arke ve Ökaryot domainlerine ait 37 farklı türün total genom dizilerinin ve
ortolog genlerin bulunma yüzdelerinin karşılaştırılmasıyla Bakteri ve Arkelerin
birbirlerinden çok fazla ayrılmadıkları saptanmıştır (Bansal ve Meyer 2002).
4
Yeşil bakterilerFlavobakteriler
Spiroketler BAKTERİLERMor bakteriler
Gram pozitif bakterilerSiyanobakterilerDeinokoklar
Termotogales
Ekstrem halofiller
Metanojenler ARKELER
Ekstrem termofiller
BitkilerFunguslar
HayvanlarrSilliatla
ÖKARYOTLAR
Cıvık mantarlarFlagellatlar
Mikrosporidialar
Şekil 2.1. Ribozomal küçük alt birimde bulunan 16S ve 18S rRNA dizilerine göre oluşturulmuş filogenetik ağaç (Woese 1999).
Arke domaini Crenarchaeota, Euryarchaeota ve Korarchaeota olmak üzere üç
filumdan oluşmaktadır. Crenarchaeota alemi içerisinde Thermoproteus, Pyrolobus ve
Pyrodictum gibi hiper termofilik cinsleri ve Euryarchaeota filumu içerisinde
Methanococcus, Methanobacterium gibi metanojenik ve Halobacterium, Haloferax gibi
ekstrem halofiller ve Thermoplasma cinsine ait termofilik arkeler yer almaktadır.
Korarchaeota filumu içerisinde ise henüz kültüre alınamayan ancak izolasyon, üretim
ortamı ve kültür şartları ve metabolik özelliklerinin benzerliklerinden dolayı
5

hipertermofilik olabilecekleri düşünülmektedir (Woese ve Wolfe 1985, Madigan et al.
2000, Wolfang ve Klenk 2001).
Ekstrem halofillerin gerçek bakteri olup olmadıkları sorusuna, 1970 lerin sonunda yeni
tanımlanmış olan Arke domainine ait Halobacterium salinarum ve ilgili diğer
halofillerin tanımı yapıldıktıktan sonra, cevap olumsuz olarak verilmiştir. Halofilik
arkelerin prokaryotik, ancak Bakteri sınıflamasına girmeyen canlılar olduğu
belirtilmiştir (Shand ve Perez 1999). Euryarchaeota filumunda sadece Halobacteriales
ordosu ve Halobacteriaceae familyası bulunmaktadır (Grant et al. 2001).
Halobacteriales, Euryarchaeota filumu içerisinde Methanomicrobiales/Methanosar-
cinales’e (metanojenlere) yakın bir dallanma göstermektedir (Oren 2001, Woese 1999)
(şekil 2.1.).
İlk halofilik arkelere ait bilgilere 5000 yıllık Çin eserlerinde rastlanılmış olup denizden
tuz elde edilmesi esnasında suyun kızardığı belirtilmiştir. Günümüzde bu sonucun
halofilik bakterilerden kaynaklandığı bilinmektedir. Halobacteriales ordosunun, kırmızı
halofilik arkeleriyle ilgili ilk çalışmaların büyük bir kısmı, tuzlanmış balık ve deriye
zarar veren etmenin anlaşılması amacıyla yapılmıştır. Bindoküzyüzondokuz yılında
Klebahn raporuna göre muhtemelen halobakterilerin ilk doğru tanımlamaları yapılmış
ve ilk bakteri Bacillus halobius ruber olarak isimlendirilmiştir. Bunu takiben 1922’de
tuzlanmış konserve morina balığı üzerindeki kırmızı lekelenmeye neden olan etken
Pseudomonas salinaria izole edilmiş, kültürü günümüzde mevcut olmayıp bu
organizmanın daha sonraları izole edilen H. salinarum’a muhtemelen çok benzediği
kabul edilmektedir. Peter, 1931 yılında tuzlamış balıklar üzerindeki halofilik arklerle
ilgili çalışmalara devam etmiştir. Tuzlanmış derilerin bakteriyel bozunumu olayı
1934’de Lochead tarafından belgelenmiştir. 1930’lu yıllarda solar tuzla ve tuz göllerinin
ilk mikrobiyolojik çalışmaları yapılmış ve bu tür ekosistemlerdeki kırmızı halofilik
bakterilerin önemi açıklanmıştır (Woese ve Wolfe 1985, Tindall 1992, Oren 2000).
6

2.2. Halofilik Arkelerin Taksonomisi ve Teşhisi
Halobacteriaceae familyasına ait cins ve türlerin teşhisi ve sınıflandırılması; hücre
morfolojisi, üreme özellikleri, spesifik polar lipitlerin analizine dayanan kemotak-
sonomik çalışmalar ve nükleik asit dizi verileri gibi özellikleri kapsayan polifazik bir
yaklaşıma göre yapılmaktadır (Oren et al. 1997, Oren 2001). Halofilik arkelerin
taksonomik sıralaması aşağıdaki gibidir (Brenner et al. 2001, Grant et al. 2001):
Domain...................... Archaea
Filum II..................... Euryarchaeota
Sınıf III ..................... Halobacteria
Ordo I........................ Halobacteriales
Familya I................... Halobacteriaceae
Son yıllarda 16S rDNA dizi karşılaştırmaları, pek çok yeni cinsin tanımlanmasına,
mevcut cinslerin bölünmesine ve hatta daha önce farklı cinslere yerleştirilen iki türün
bir cinste birleştirilmesine neden olan yeni bir sınıflandırmaya yol açmıştır (Kamekura
ve Seno 1993, Kamekura 1998). 16S rRNA nükleotid dizi karşılaştırmaları halofilik
arkelerin alkalifik üyelerinin farklı bir filogenetik grup oluşturmadığını ortaya
koymuştur. Günümüzde, halofilik arke üyelerinin filogenetik sınıflamasında önemli
olan 16S rRNA dizi analizleri, polar lipit kompozisyonunun önemini oldukça
azaltmıştır. Çünkü bazı tanımlayıcı glikolipitler birden fazla cinste bulunabilmekte ve
bazı cinslerin üyeleri farklı glikolipitler içerebilmektedirler (Kamekura ve Kates 1999).
Şekil 2.2.’de Halobacteriaceae ailesi üyelerinin 16S rRNA dizi analizlerine dayanılarak
yapılmış filogenetik ağacı görülmektedir.
Halobacteraiales ordosuna ait yeni bir taksonun tanımlanması ile ilgili minimum
standartlar Oren et al. (1997) tarafından belirtilmiştir. Buna göre;
• Koloni büyüklüğü, şekli ve pigmentasyon
• Hücre morfolojisi ve hareket
• Gram reaksiyonu
7
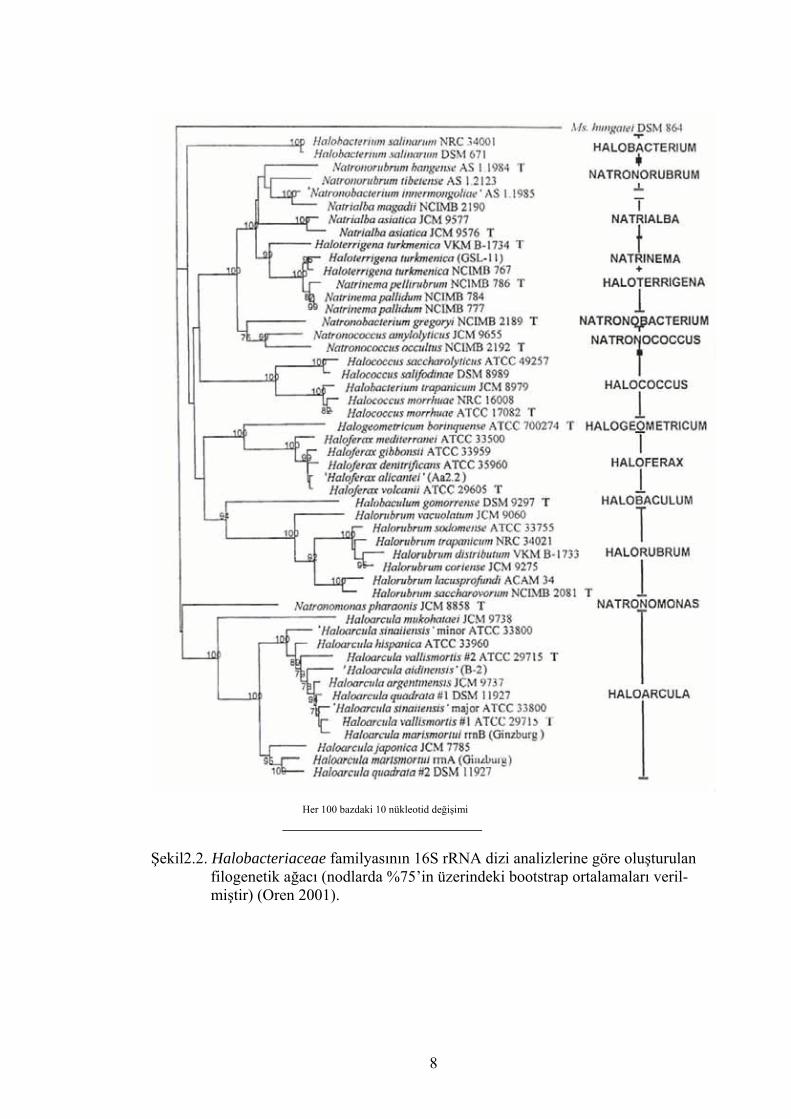
Her 100 bazdaki 10 nükleotid değişimi _________________________ Şekil2.2. Halobacteriaceae familyasının 16S rRNA dizi analizlerine göre oluşturulan filogenetik ağacı (nodlarda %75’in üzerindeki bootstrap ortalamaları veril- miştir) (Oren 2001).
8

• Minimum ve optimum NaCl ve MgCI2 konsantrasyonları
• Sıcaklık ve pH aralığı
• Nitrat varlığında anaerobik üreme yeteneği ve L-arjininde anaerobik üreme
• Karbohidratlardan asit üretimi
• Tek karbon kaynağında üreyebilme
• Katalaz ve oksidaz aktivitesi, indol üretimi ve nişasta, jelatin, kazein ve Tween
80 hidrolizi
• Antimikrobiyal maddelere duyarlılık
• Mevcut glikolipit tiplerini ve PGS’nin varlığını veya yokluğunu kapsayacak
şekilde polar lipit karakterizasyonu
• DNA’nın G+C içeriği
• 16S rRNA dizi
bilgisini içermektedir.
Mevcut cinslere ait yeni türlerin tanımlanması ve yakınlık derecelerinin saptanmasında
DNA-DNA hibridizasyonunun yanısıra DNA-16SrRNA hibridizasyonu çalışmaları da
kullanılmıştır (Ross ve Grant 1985, Gutierrez et al. 1989, Gutierrez et al. 1990). Diğer
yandan yakın suşların karşılaştırılmasında Rastgele Çoğaltılmış Polimorfik DNA
analizleri (RAPD) kullanılarak, suşların tanımlanmasında ortaya çıkan problemlerin
çözümünde güçlü bir araç olduğu gösterilmiştir (Martinez-Murcia ve Rodriguez-Valera
1994, Martinez-Murcia et al. 1995). Ayrıca suşların karşılaştırılması ve doğrulamak
amacyla protein profillerinin çıkarıldığı SDS poliakrilamid jel elektroforezi kullanılan
diğer bir yöntemdir (Hesselberg ve Vreeland 1995, McGenity et al. 1998, Asker ve
Ohta 2002, Stan-Lotter et al. 2002).
Yeni bir taksonun (cins veya tür) tam ve doğru bir tanımının yapılabilmesi için, suşların
fizyolojik ve biyokimyasal karakterizasyonuyla birlikte herhangi bir kemotaksonomik
verinin de kullanılması gerektiği belirtilmektedir (Oren 2001).
Halobacteriales ordosu içerisinde günümüze kadar tanımlanmış 18 cins ve 51 tür
bulunmaktadır (çizelge 2.2.).
9

2.2.1. Halobacteriales ordosunun özellikleri
Halobacteriales ordosu, çubuk, kok, düz extrem pleomorfik hücreler, kusursuz düz kare
hücrelerden (Oren et al. 1996, Oren 1999) üçgen ve trapezoid (Takashina et al. 1990,
Horikoshi et al. 1993) şekilli hücrelere kadar çeşitli morfolojik tipler içermektedir. Bazı
suşlarda halokist olarak belirtilen yapıların varlığı rapor edilmesine rağmen (Kostrikina
et al. 1991), dinlenme evrelerinin varlığı ve spor yapıları rapor edilememiştir.
Hareketsiz veya demet şeklindeki flagellumları ile aktif hareketlidirler. Gram negatif
veya pozitif boyanabilmektedirler (Dussault 1955, Grant et al. 2001). Aerobik olup,
oksidaz ve katalaz aktiviteleri pozitif, bazı suşlar nitrat varlığında yada
bakteriyorodopsin yardımıyla anaerobik olarak üreyebilmektedirler. Ayrıca arjinin
varlığında fermentatif üreyebilen bazı suşları bulunmaktadır (Hartmann et al. 1980,
Oesterhelt 1998, Oren ve Litchfield 1999). Üremeleri için en az 1,5 M NaCl’e ihtiyaç
duymakta olup suşların çoğunluğu 3,5-4,5 M NaCl konsantrasyonunda en iyi üreme
göstemektedir (Oren 2000). Pek çok suşun koloni rengi, içerdikleri C50 ve C40
karotenoidlerinden dolayı kırmızı ve tonlarındadır. Optimum üreme sıcaklıkları 35-
50°C arasındadır. Kemoorganotrofiktirler, karbon kaynağı olarak karbohidratları veya
amino asitleri kullanırlar. Kompleks ortamlarda üremelerine rağmen bazı türler tek
karbon kaynağı içeren inorganik ortamlarda üreyebilmektedirler (Grant et al. 2001).
Fitanil (C20) veya sesterpanil (C25) yan zincirleri olan di-izoprenoid gliserol eter türevli
polar lipitleri mevcuttur (Kates 1993). Ordo üyelerinin DNA’ları genellikle bir majör ve
bir minör komponent içermektedir. Minör komponent total DNA’nın %10-30’nu
oluşturmaktadır. DNA’nın %mol G+C oranı majör komponentde 59-71 ve minör
komponentde 51-59 arasındadır (Pfeifer et al. 1988, Gutierrez et al. 1986).
Halobacteriaceae familyasının özellikleri ordonunki ile aynıdır. Familyanın cins tipi
Halobacterium’dur (Tindall 1992, Grant et al. 2001). Çizelge 2.3., 2.4., 2.5., 2.6., 2.7.
ve 2.8.’de tanımlanmış halofilik arke türlerinin özellikleri verilmiştir.
10

Çizelge 2.2. Halobacteriales ordosuna ait cinsler ve türler
FAMİLYA CİNS TÜR Halobacteriaceae HalobacteriumT Halobacterium salinarumT
Haloarcula Haloarcula vallismortisT
Haloarcula marismortui Haloarcula hispanica Haloarcula japonica Haloarcula argentinensis Haloarcula quadrata Haloarcula californiae Haloarcula sinaiensis Haloarcula aidinensis Halobaculum Halobaculum gomorrenseT
Halococcus Halococcus morrhuaeT
Halococcuss saccharolyticus Halococcus salifodinae Halococcus dombrowskii Haloferax Haloferax volcaniiT Haloferax denitrificans Haloferax gibbonsii Haloferax mediterranei Haloferax alicantei Haloferax lucentensis Haloferax alexandrinus Halogeometricum Halogeometricum borinquenseT
Halorubrum Halorubrum saccharovorumT
Halorubrum coriense Halorubrum distributum Halorubrum lacusprofundi Halorubrum sodomonse Halorubrum trapanicum Halorubrum vacuolatum Halorubrum tebenquichense Halorubrum terrestre Haloterrigena Haloterrigena turkmenicaT
Haloterrigena thermotolerans Natrialba Natrialba asiaticaT
Natrialba magadii Natrialba taiwanensis Natrialba aegytiaca Halosimplex Halosimplex carlbadanseT
Halomicrobium Halomicrobium mukohataeiT
Halorhabdus Halorhabdus utahensisT
Halobioforma Halobioforma haloterrestrisT
Natrinema Natrinema pellirubrumT
Natrinema pallidium Natronobacterium Natronobacterium gregoryiT
Natronococcus Natronococcus occultusT
Natronococcus amylolyticus Natronomonas Natronomonas pharaonisT
Natronorubrum Natronorubrum bangenseT
Natronorubrum tibetense
T= Familyanın cins tipi veya Cinsin tür tipi
11

2.3. Halofilik Arkelerin Habitatları ve Ekolojisi
Doğada halofilik arkelerin dağılımını belirleyen temel faktörler; total tuz
konsantrasyonu, tuzların iyonik kompozisyonu ve mevcut besinlerdir. Dünya üzerinde
halofilik arkeler, evaporasyon ile denizden orijinlenen ve dominant iyon kompozisyonu
Na+ ve CI- olan denizel (thalassohaline) ortamlarda ve oldukça farklı iyonik
kompozisyona sahip denizel olmayan (athalassohaline) sularda da yaşamaktadırlar.
Denizel olmayan sulardan olan İsraildeki Ölü Deniz, divalent katyonlar yönünden
zengindir ve yaklaşık olarak 1.9 M Mg+2, 0.4 M Ca+2, 1.7 M Na+ ve 0.14 M K+
içermektedir. Magadi gölü (Kenya), Wadi Naturn gölü (Mısır) gibi hiper tuzlu, yüksek
pH değerlerine sahip soda göllerinde de halofilik arkeler bol miktarda bulunmaktadırlar
(Rodriguez-Valera et al. 1979, Rodriguez-Valera 1988, Tindall 1988, Oren 1994,
Kamekura 1999). Halofilik arkelerin çoğu yüksek karotenoid içermelerinden dolayı
yüksek kommunite yoğunluğuna ulaştıkları zaman nötral ve alkali hiper tuzlu suların
kırmızı renk tonlarında gözükmelerine neden olmaktadırlar (Arahal et al. 1996, Oren ve
Rodriguez-Valera 2001, Oren 2002). Asidofilik halofilik arke tipleri henüz rapor
edilmemiştir. Ölü Deniz’in (İsrail) pH’ı yaklaşık 6.0 civarında olup bu alanın
muhtemelen halofilik arkelerin kitleler halinde üredikleri en asidik ortam olduğu
belirtilmiştir (Oren ve Gurevich 1993). Halofilik arkelerin çoğunluğu optimum olarak
35-50 ºC arasındaki ve bazen daha yüksek sıcaklıklarda üremektedirler (Shand ve perez
1999). Buna karşılık su sıcaklığı +11.5 ile 0’ın altındaki aralıklarda değişen su
sıcaklığına sahip Antartika’daki Deep Lake’den Halorubrum lacusprofundi, izole
edilmiştir ve optimum üreme sıcaklığının 31-37 ºC olduğu, ancak 4ºC gibi düşük
sıcaklıklarda üremesinin yavaşladığı belirtilmiştir (Franzmann et al. 1988). Halofilik
arkeler sulu ortamların yanı sıra tuzlu topraklardan da izole edilmişlerdir. İspanyanın
Akdeniz kıyıları (Queseda et al. 1982), Eski SSCB’in yüksek dağ ve düzlüklerindeki
(Kulicheuskaya et al. 1992, Zyvagintseva ve Tarasov 1988) çorak alanlardan da izole
edilmişlerdir. Halobacteriaceae üyeleri ayrıca, korunması için tuzlanmış yiyecek, deri
ve diğer ürünlerde de bulunabilmektedirler (Rodriguez-Valera 1988).
12
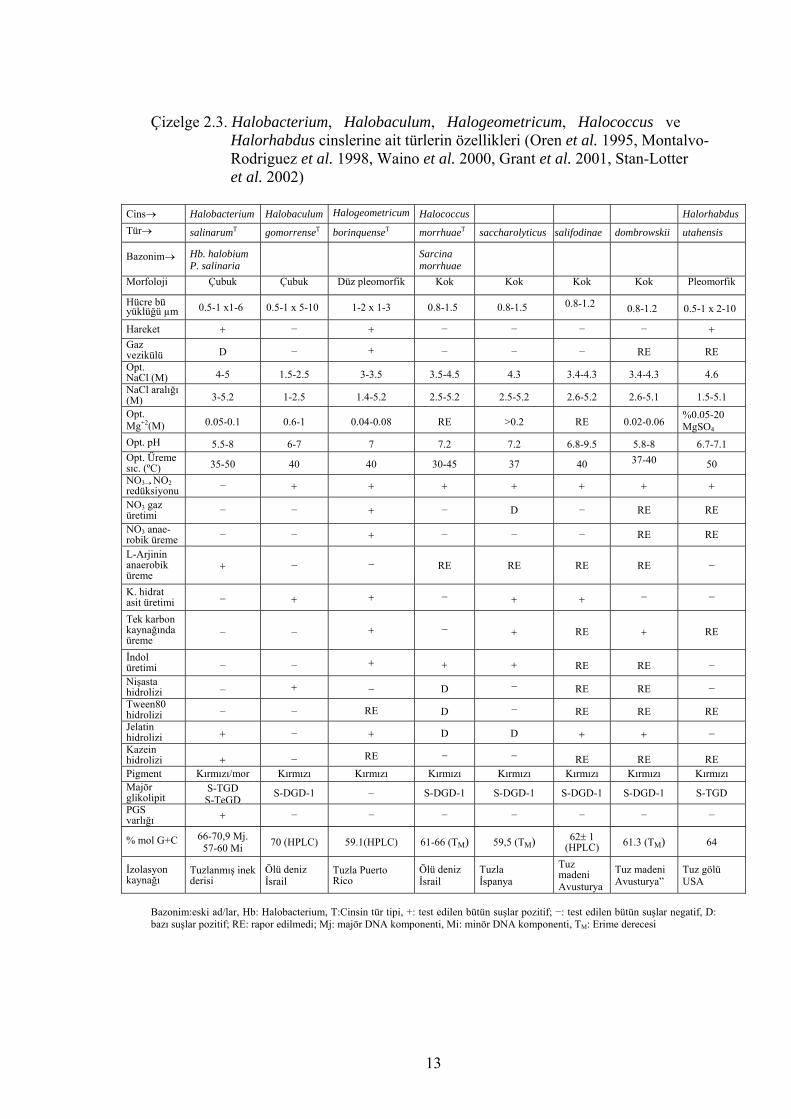
Çizelge 2.3. Halobacterium, Halobaculum, Halogeometricum, Halococcus ve Halorhabdus cinslerine ait türlerin özellikleri (Oren et al. 1995, Montalvo- Rodriguez et al. 1998, Waino et al. 2000, Grant et al. 2001, Stan-Lotter et al. 2002)
Cins→ Halobacterium Halobaculum Halogeometricum Halococcus Halorhabdus Tür→ salinarumT gomorrenseT borinquenseT morrhuaeT saccharolyticus salifodinae dombrowskii utahensis
Bazonim→ Hb. halobium P. salinaria
Sarcina morrhuae
Morfoloji Çubuk Çubuk Düz pleomorfik Kok Kok Kok Kok Pleomorfik
Hücre bü yüklüğü µm 0.5-1 x1-6 0.5-1 x 5-10 1-2 x 1-3 0.8-1.5 0.8-1.5 0.8-1.2
0.8-1.2 0.5-1 x 2-10
Hareket + − + − − − − + Gaz vezikülü D − + − − − RE RE Opt. NaCl (M) 4-5 1.5-2.5 3-3.5 3.5-4.5 4.3 3.4-4.3 3.4-4.3 4.6 NaCl aralığı (M) 3-5.2 1-2.5 1.4-5.2 2.5-5.2 2.5-5.2 2.6-5.2 2.6-5.1 1.5-5.1 Opt. Mg+2(M) 0.05-0.1 0.6-1 0.04-0.08 RE >0.2 RE 0.02-0.06
%0.05-20 MgSO4
Opt. pH 5.5-8 6-7 7 7.2 7.2 6.8-9.5 5.8-8 6.7-7.1 Opt. Üreme sıc. (ºC) 35-50 40 40 30-45 37 40 37-40 50 NO3→ NO2 redüksiyonu − + + + + + + +
NO3 gaz üretimi − − + − D − RE RE
NO3 anae-robik üreme − − + − − − RE RE
L-Arjinin anaerobik üreme
+ − − RE RE RE RE −
K. hidrat asit üretimi − + + − + + − −
Tek karbon kaynağında üreme
− − + − + RE + RE
İndol üretimi − − + + + RE RE − Nişasta hidrolizi − + − D − RE RE − Tween80 hidrolizi − − RE D − RE RE RE Jelatin hidrolizi + − + D D + + − Kazein hidrolizi + − RE − − RE RE RE Pigment Kırmızı/mor Kırmızı Kırmızı Kırmızı Kırmızı Kırmızı Kırmızı Kırmızı Majör glikolipit
S-TGD S-TeGD
S-DGD-1 − S-DGD-1 S-DGD-1 S-DGD-1 S-DGD-1 S-TGD PGS varlığı + − − − − − − −
% mol G+C
66-70,9 Mj. 57-60 Mi 70 (HPLC) 59.1(HPLC) 61-66 (TM) 59,5 (TM) 62± 1
(HPLC) 61.3 (TM) 64
İzolasyon kaynağı
Tuzlanmış inek derisi
Ölü deniz İsrail
Tuzla Puerto Rico
Ölü deniz İsrail
Tuzla İspanya
Tuz madeni Avusturya
Tuz madeni Avusturya”
Tuz gölü USA
Bazonim:eski ad/lar, Hb: Halobacterium, T:Cinsin tür tipi, +: test edilen bütün suşlar pozitif; −: test edilen bütün suşlar negatif, D: bazı suşlar pozitif; RE: rapor edilmedi; Mj: majör DNA komponenti, Mi: minör DNA komponenti, TM: Erime derecesi
13

Çizelge 2.4. Halorubrum cinsine ait türlerin özellikleri (Tomlinson ve Hochstein 1976, Franzmann et al. 1988, McGenity ve Grant 1995, Kamekura ve Dyall- Smith 1995, Grant et al. 2001, Lizama et al. 2002)
Cins→ Halorubrum
Tür→ saccharovorumT sodomonse lacusprofundi coriense distributum vacuolatum trapanicum tebenquichense
Bazonim→ Hb. saccharovorum
Hb. sodomonse
Hb. lacusprofundi
Hrb. coriense
Hb. distributum
Natro. Vacuolatum (vacuolata)
Hb. trapanicum
Morfoloji Çubuk Çubuk Çubuk Pleomorfik çubuk
Pleomorfik çubuk
Çubuk Çubuk Düzensiz disk
Hücre bü yüklüğü µm
0.6-1.2 x2.5
0.5 x2.5-5 12’e kadar 0.5-1 x
0.5-5 0.8-1 x2.5-7 0.5-0.7 x 1.5-3
0.7-1 x 1.5-3 1-2 x0.8-1.5
Hareket + + D + + − − R.E
Gaz vezikülü − − − − − + − R.E
Opt. NaCl (M) 3.5-4.5 1.7-2.5 2.5-3.5 2.2-2.7 2.5-4.3 3.5 2.5-5.2 5.2
NaCl aralığı (M) 1.5-5.2 0.5-4.3 1.5-5.2 2-5.2 1.7-5.2 2.5-5.2 RE 2.5-5.2
Opt. Mg+2(M)
>0.005 0.6-1.2 0.005 >0.005 RE >0.001 RE −
Opt. pH Nötral Nötral Nötral Nötral Nötral 9.5 Nötral R.E Opt. Üreme sıc. (ºC) 50 40 31-36 50 37-45 35-40 37 R.E
NO3→ NO2 redüksiyonu
+ ± + veya ± RE + + + +
NO3 gaz üretimi
− − − RE − − + R.E
NO3 anae-robik üreme
− − − RE − − − R.E
L-Arjinin anaerobik üreme
− − RE RE RE RE RE R.E
K. hidrat asit üretimi + + + RE + RE + −
Tek karbon kaynağında üreme
RE − + + − RE RE +
İndol üretimi − − − − − RE − −
Nişasta hidrolizi − + − RE − − − − Tween80 hidrolizi RE − RE RE RE RE RE − Jelatin hidrolizi − − − RE − − − − Kazein hidrolizi − RE − RE RE RE RE R.E
Pigment Turuncu-kırmızı Turuncu-Kırmızı
Turuncu-kırmızı Kırmızı Turuncu-
kırmızı Parlak pembeı Kırmızı Turuncu-
kırmızı Majör glikolipit
S-DGD-3 veya S-DGD-1 S-DGD-3 S-DGD-3 S-DGD-3 S-DGD-3 − S-DGD-5 S-DGD-3
PGS varlığı + + + + + − + −
% mol G+C
71.2 (Bd) 68 (Bd) 65.3-65.8 Mj 54.6-65.3 Mi RE 63.6 Mj
54.6 Mi 62.7 (TM) 64.3 (Bd) 63.2 (TM)
İzolasyon kaynağı
Tuzla Kalifornia
Ölü deniz İsrail
Deep lake Antarktika
Tuzla, Avustralya
Tuzlu top-rak, USSR
Magadi gö-lü, Kenya
Solar tuz İtalya
Tebenquiche gölü, Şili
Bazonim:eski ad/lar, Hb: Halobacterium, Hrb:Halorubrobacterium, Natro: Natronobacterium, T:Cinsin tür tipi, +: test edilen bütün suşlar pozitif; −: test edilen bütün suşlar negatif, ±: zayıf reaksiyon, D: bazı suşlar pozitif; RE: rapor edilmedi; Mj: majör DNA komponenti, Mi: minör DNA komponenti, TM: Erime derecesi, Bd:Buoyant density
14
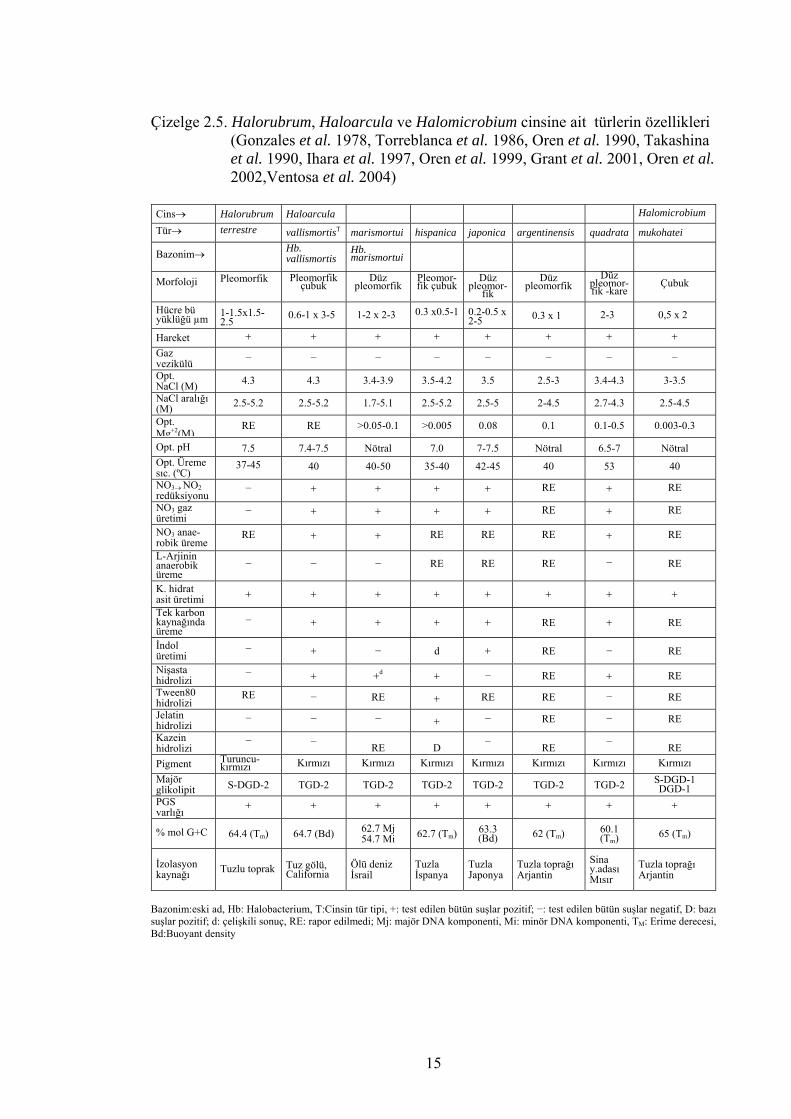
Çizelge 2.5. Halorubrum, Haloarcula ve Halomicrobium cinsine ait türlerin özellikleri (Gonzales et al. 1978, Torreblanca et al. 1986, Oren et al. 1990, Takashina et al. 1990, Ihara et al. 1997, Oren et al. 1999, Grant et al. 2001, Oren et al. 2002,Ventosa et al. 2004) Cins→ Halorubrum Haloarcula Halomicrobium
Tür→ terrestre vallismortisT marismortui hispanica japonica argentinensis quadrata mukohatei
Bazonim→ Hb. vallismortis
Hb. marismortui
Morfoloji Pleomorfik Pleomorfik çubuk
Düz pleomorfik
Pleomor-fik çubuk
Düz pleomor-
fik Düz
pleomorfik Düz
pleomor-fik -kare
Çubuk
Hücre bü yüklüğü µm 1-1.5x1.5-
2.5 0.6-1 x 3-5 1-2 x 2-3 0.3 x0.5-1 0.2-0.5 x
2-5 0.3 x 1 2-3 0,5 x 2
Hareket + + + + + + + + Gaz vezikülü − − − − − − − −
Opt. NaCl (M) 4.3 4.3 3.4-3.9 3.5-4.2 3.5 2.5-3 3.4-4.3 3-3.5
NaCl aralığı (M) 2.5-5.2 2.5-5.2 1.7-5.1 2.5-5.2 2.5-5 2-4.5 2.7-4.3 2.5-4.5
Opt. Mg+2(M)
RE RE >0.05-0.1 >0.005 0.08 0.1 0.1-0.5 0.003-0.3
Opt. pH 7.5 7.4-7.5 Nötral 7.0 7-7.5 Nötral 6.5-7 Nötral Opt. Üreme sıc. (ºC)
37-45 40 40-50 35-40 42-45 40 53 40
NO3→ NO2 redüksiyonu
− + + + + RE + RE
NO3 gaz üretimi
− + + + + RE + RE
NO3 anae-robik üreme
RE + + RE RE RE + RE
L-Arjinin anaerobik üreme
− − − RE RE RE − RE
K. hidrat asit üretimi + + + + + + + +
Tek karbon kaynağında üreme
− + + + + RE + RE
İndol üretimi
− + − d + RE − RE
Nişasta hidrolizi
− + +d + − RE + RE Tween80 hidrolizi
RE − RE + RE RE − RE
Jelatin hidrolizi
− − − + − RE − RE
Kazein hidrolizi
− − RE D − RE − REPigment Turuncu-
kırmızı Kırmızı Kırmızı Kırmızı Kırmızı Kırmızı Kırmızı Kırmızı Majör glikolipit S-DGD-2 TGD-2 TGD-2 TGD-2 TGD-2 TGD-2 TGD-2 S-DGD-1
DGD-1 PGS varlığı
+ + + + + + + +
% mol G+C
64.4 (Tm) 64.7 (Bd) 62.7 Mj 54.7 Mi 62.7 (Tm) 63.3
(Bd) 62 (Tm) 60.1 (Tm) 65 (Tm)
İzolasyon kaynağı Tuzlu toprak Tuz gölü,
California Ölü deniz İsrail
Tuzla İspanya
Tuzla Japonya
Tuzla toprağı Arjantin
Sina y.adası Mısır
Tuzla toprağı Arjantin
Bazonim:eski ad, Hb: Halobacterium, T:Cinsin tür tipi, +: test edilen bütün suşlar pozitif; −: test edilen bütün suşlar negatif, D: bazı suşlar pozitif; d: çelişkili sonuç, RE: rapor edilmedi; Mj: majör DNA komponenti, Mi: minör DNA komponenti, TM: Erime derecesi, Bd:Buoyant density
15

Çizelge 2.6. Haloferax ve Halosimplex cinslerine ait türlerin özellikleri (Rodriguez- Valera et al. 1983, Torreblanca et al. 1986, Tindall et al. 1989, Grant et al. 2001, Gutierrez et al. 2002, Vreeland et al. 2002, Asker ve Ohta 2002)
Cins→ Haloferax Halosimp-lex
Tür→ volcaniiT gibbonsii denitrificans mediterranei lucentensis alexandri-nus carlbadense
Bazonim→ Hb. volcanii
Hb. denitrificans
Hb. mediterranei
Morfoloji Düz pleomorfik
Pleomorfik çubuk Pleomorfik Pleomorfik Pleomorfik
çubuk Pleomorfik- çubuk Çubuk
Hücre bü yüklüğü µm 1-2 x 2-3 0.4x0.5-2.5 0.8-1 x 2-3 0.5-1 x 2-3 0.6 x 2.5 1-1.5x1.6-2 0.95 x 5.0
Hareket − + − + + − + Gaz vezikülü − − − + RE − RE
Opt. NaCl (M) 1.5-2.5 3-4 4.3 2-3 4.3 4.3 4.3
NaCl aralığı (M) 1-4.5 1.5-5.2 1.5-4.5 1-5.2 1.7-5.1 1.7-5.2 RE
Opt. Mg+2(M)
0.2 >0.02-0.04 RE 0.02-0.04 RE %4 MgSO4 %1-2w/v
Opt. pH Nötral 6.5-7 6.7 6.5 7.5 7.2 RE Opt. Üreme sıc. (ºC) 45 35-40 50 47-54 37 37 37-40
NO3→ NO2 redüksiyonu
+ D + + − + RE
NO3 gaz üretimi
− D + + − RE
NO3 anae-robik üreme −d RE + + RE − −
L-Arjinin anaerobik üreme
− − − − − − −
K. hidrat asit üretimi
+ + + + + + RE
Tek karbon kaynağında üreme
+ + + + + + +
İndol üretimi
+ + − + + RE RE
Nişasta hidrolizi
− − − + − − −
Tween 80 hidrolizi − + − + RE + RE
Jelatin hidrolizi − − + + − + −
Kazein hidrolizi − ± RE RE − − RE
Pigment Kırmızı Kırmızı Turuncu-kırmızı Açık pembe Pembe Kırmızı Pembe-
kırmızı Majör glikolipit S-DGD-1 S-DGD-1 S-DGD-1 S-DGD-1 S-DGD-1 S-DGD-1 S2-DGD
S-TeGD PGS varlığı − − − − − − −
% mol G+C
63.4 Mj T m55.3 Mi.Bd 61.8 (Tm) 64.2 (Tm) 60 (Tm) 64.5 (Tm) 59.5± 0.3
(HPLC) 64.4
(HPLC)
İzolasyon kaynağı
Ölü deniz İsrail
Tuzla İspanya
Tuzla California
Tuzla İspanya
Tuzla İspanya Tuzla, Mısır
Tuz kristaliNew
mexico Bazonim:eski ad, Hb: Halobacterium, T:Cinsin tür tipi, +: test edilen bütün suşlar pozitif; −: test edilen bütün suşlar negatif, D: bazı suşlar pozitif; d: çelişkili sonuç, RE: rapor edilmedi; Mj: majör DNA komponenti, Mi: minör DNA komponenti, TM: Erime derecesi, Bd:Buoyant density
16
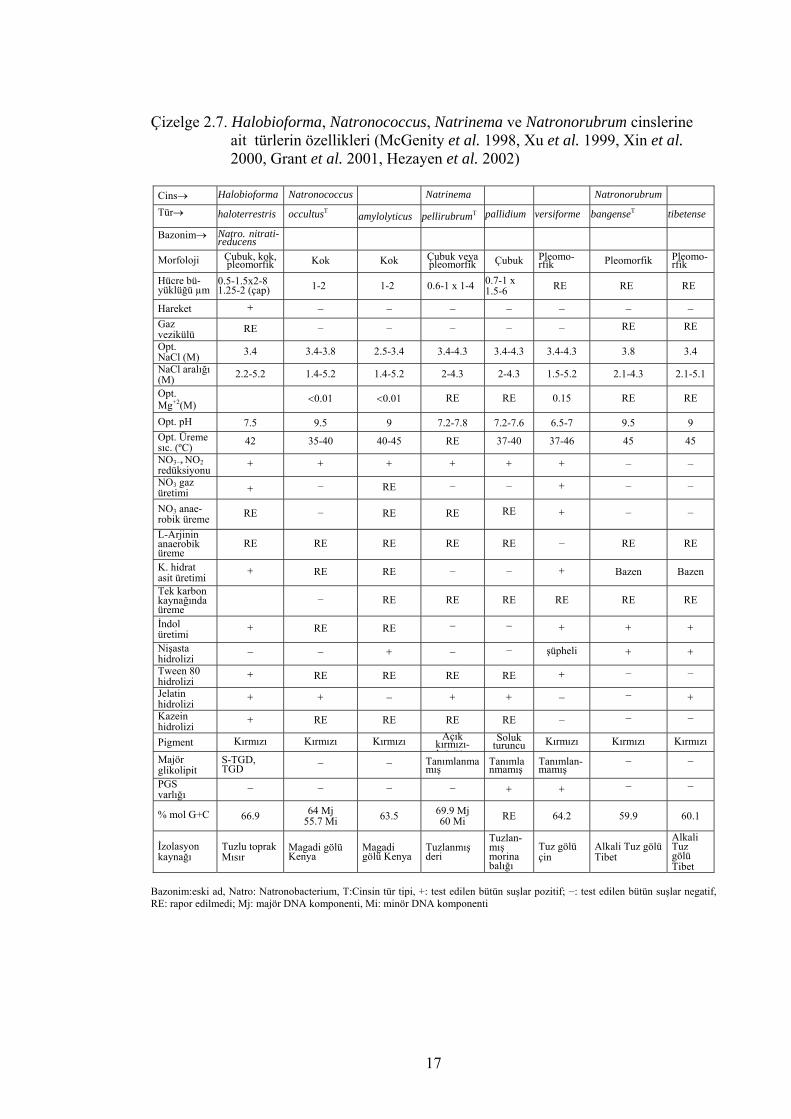
Çizelge 2.7. Halobioforma, Natronococcus, Natrinema ve Natronorubrum cinslerine ait türlerin özellikleri (McGenity et al. 1998, Xu et al. 1999, Xin et al. 2000, Grant et al. 2001, Hezayen et al. 2002)
Cins→ Halobioforma Natronococcus Natrinema Natronorubrum
Tür→ haloterrestris occultusT amylolyticus pellirubrumT pallidium versiforme bangenseT tibetense
Bazonim→ Natro. nitrati-reducens
Morfoloji Çubuk, kok, pleomorfik Kok Kok Çubuk veya
pleomorfik Çubuk Pleomo-rfik Pleomorfik Pleomo-
rfik Hücre bü-yüklüğü µm
0.5-1.5x2-8 1.25-2 (çap) 1-2 1-2 0.6-1 x 1-4 0.7-1 x
1.5-6 RE RE RE
Hareket + − − − − − − − Gaz vezikülü RE − − − − − RE RE
Opt. NaCl (M) 3.4 3.4-3.8 2.5-3.4 3.4-4.3 3.4-4.3 3.4-4.3 3.8 3.4
NaCl aralığı (M) 2.2-5.2 1.4-5.2 1.4-5.2 2-4.3 2-4.3 1.5-5.2 2.1-4.3 2.1-5.1
Opt. Mg+2(M)
<0.01 <0.01 RE RE 0.15 RE RE
Opt. pH 7.5 9.5 9 7.2-7.8 7.2-7.6 6.5-7 9.5 9 Opt. Üreme sıc. (ºC) 42 35-40 40-45 RE 37-40 37-46 45 45
NO3→ NO2 redüksiyonu
+ + + + + + − −
NO3 gaz üretimi + − RE − − + − −
NO3 anae-robik üreme RE − RE RE RE + − −
L-Arjinin anaerobik üreme
RE RE RE RE RE − RE RE
K. hidrat asit üretimi
+ RE RE − − + Bazen Bazen
Tek karbon kaynağında üreme
− RE RE RE RE RE RE
İndol üretimi
+ RE RE − − + + +
Nişasta hidrolizi − − + − − şüpheli + +
Tween 80 hidrolizi
+ RE RE RE RE + − −
Jelatin hidrolizi
+ + − + + − − +
Kazein hidrolizi
+ RE RE RE RE − − −
Pigment Kırmızı Kırmızı Kırmızı Açık kırmızı-t r nc
Soluk turuncu Kırmızı Kırmızı Kırmızı
Majör glikolipit
S-TGD, TGD − − Tanımlanma
mış Tanımlanmamış
Tanımlan-mamış
− −
PGS varlığı − − − − + + − −
% mol G+C
66.9 64 Mj 55.7 Mi 63.5 69.9 Mj
60 Mi RE 64.2 59.9 60.1
İzolasyon kaynağı
Tuzlu toprak Mısır
Magadi gölü Kenya
Magadi gölü Kenya
Tuzlanmış deri
Tuzlan-mış morina balığı
Tuz gölü çin
Alkali Tuz gölü Tibet
Alkali Tuz gölü Tibet
Bazonim:eski ad, Natro: Natronobacterium, T:Cinsin tür tipi, +: test edilen bütün suşlar pozitif; −: test edilen bütün suşlar negatif, RE: rapor edilmedi; Mj: majör DNA komponenti, Mi: minör DNA komponenti
17
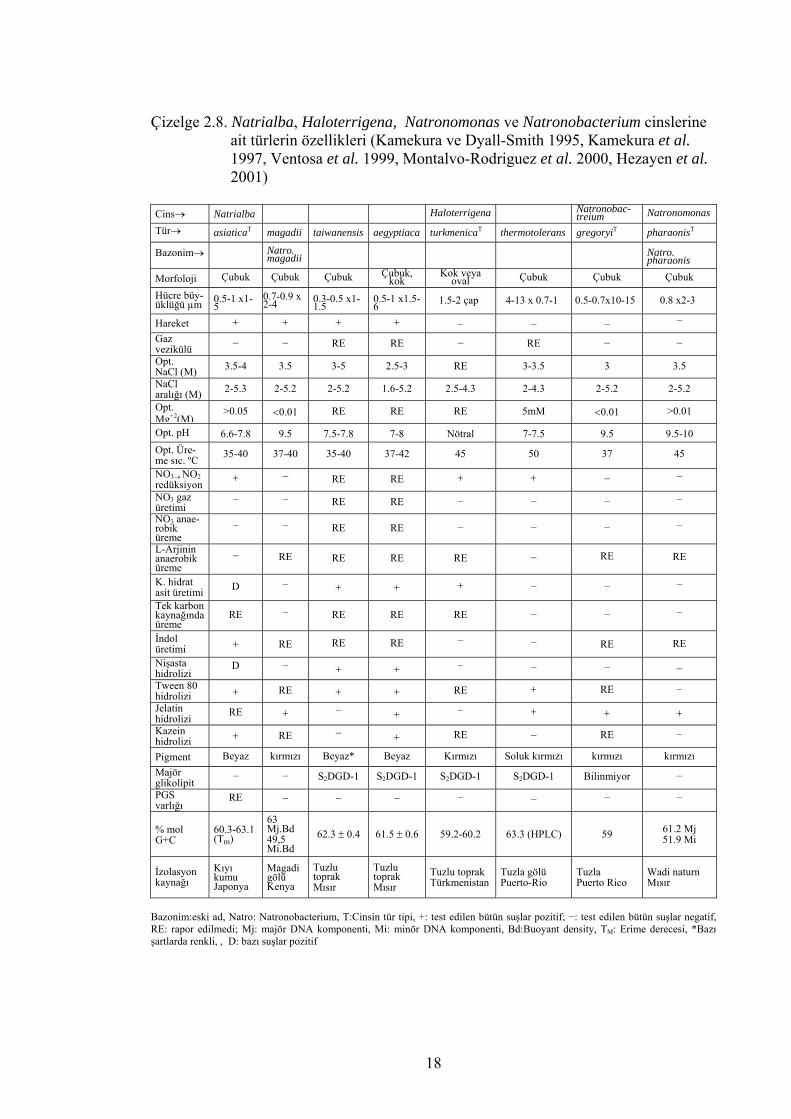
Çizelge 2.8. Natrialba, Haloterrigena, Natronomonas ve Natronobacterium cinslerine ait türlerin özellikleri (Kamekura ve Dyall-Smith 1995, Kamekura et al. 1997, Ventosa et al. 1999, Montalvo-Rodriguez et al. 2000, Hezayen et al. 2001) Cins→ Natrialba Haloterrigena Natronobac-
treium Natronomonas
Tür→ asiaticaT magadii taiwanensis aegyptiaca turkmenicaT thermotolerans gregoryiT pharaonisT
Bazonim→ Natro. magadii
Natro.
pharaonis
Morfoloji Çubuk Çubuk Çubuk Çubuk, kok
Kok veya oval Çubuk Çubuk Çubuk
Hücre büy-üklüğü µm 0.5-1 x1-
5 0.7-0.9 x 2-4 0.3-0.5 x1-
1.5 0.5-1 x1.5-6 1.5-2 çap 4-13 x 0.7-1 0.5-0.7x10-15 0.8 x2-3
Hareket + + + + − − − −
Gaz vezikülü − − RE RE − RE − −
Opt. NaCl (M) 3.5-4 3.5 3-5 2.5-3 RE 3-3.5 3 3.5
NaCl aralığı (M) 2-5.3 2-5.2 2-5.2 1.6-5.2 2.5-4.3 2-4.3 2-5.2 2-5.2
Opt. Mg+2(M)
>0.05 <0.01 RE RE RE 5mM <0.01 >0.01
Opt. pH 6.6-7.8 9.5 7.5-7.8 7-8 Nötral 7-7.5 9.5 9.5-10 Opt. Üre-me sıc. ºC 35-40 37-40 35-40 37-42 45 50 37 45
NO3→ NO2 redüksiyon
+ − RE RE + + − −
NO3 gaz üretimi
− − RE RE − − − −
NO3 anae-robik üreme
− − RE RE − − − −
L-Arjinin anaerobik üreme
− RE RE RE RE − RE RE
K. hidrat asit üretimi D − + + + − − −
Tek karbon kaynağında üreme
RE − RE RE RE − − −
İndol üretimi + RE RE RE − − RE RE
Nişasta hidrolizi
D − + + − − − − Tween 80 hidrolizi + RE + + RE + RE −
Jelatin hidrolizi RE + − + − + + +
Kazein hidrolizi + RE − + RE − RE −
Pigment Beyaz kırmızı Beyaz* Beyaz Kırmızı Soluk kırmızı kırmızı kırmızı Majör glikolipit − − S2DGD-1 S2DGD-1 S2DGD-1 S2DGD-1 Bilinmiyor −
PGS varlığı
RE − − − − − − −
% mol G+C
60.3-63.1 (Tm)
63 Mj.Bd 49,5 Mi.Bd
62.3 ± 0.4 61.5 ± 0.6 59.2-60.2 63.3 (HPLC) 59 61.2 Mj 51.9 Mi
İzolasyon kaynağı
Kıyı kumu Japonya
Magadi gölü Kenya
Tuzlu toprak Mısır
Tuzlu toprak Mısır
Tuzlu toprak Türkmenistan
Tuzla gölü Puerto-Rio
Tuzla Puerto Rico
Wadi naturn Mısır
Bazonim:eski ad, Natro: Natronobacterium, T:Cinsin tür tipi, +: test edilen bütün suşlar pozitif; −: test edilen bütün suşlar negatif, RE: rapor edilmedi; Mj: majör DNA komponenti, Mi: minör DNA komponenti, Bd:Buoyant density, TM: Erime derecesi, *Bazı şartlarda renkli, , D: bazı suşlar pozitif
18

Halobacteriaceae içinde tuz gereksinimi ve toleransına bağlı olarak farklı cins ve türler
arasında büyük farklılık görülür. Halobacterium cinsine ait tiplerin aksine türlerin çoğu,
özellikle Haloferax cinsine ait olanlar daha düşük tuz konsantrasyonlarında
gelişebilmektedirler (Rodriguez-Valera et al. 1985). Ortamdaki Mg+2 konsantrasyonu da
ekolojik bir öneme sahiptir. Nötrofil halofilik arkelerin çoğunluğu diğerlerine göre daha
fazla Mg+2 konsantrasyonuna (min 50-100mM) gereksinim duymaktadırlar. Suboptimal
Mg+2 konsantrasyonlarında türlere göre değişmek üzere çoğunlukla hücresel şekillerini
kaybederek canlıklarını sürdürebilmekte (Brown and Gibbons 1995) ya da
yitirmektedirler (Cohen et al. 1983). Alkalifilik üyeler ise yüksek Mg+2
konsantrasyonuna gerek duymamaktadırlar (Tindall 1988).
Halofilik arkeler yüksek tuz konsantrasyonunda yaşadıkları için bu ortamlarda
gelişebilecek predatörleri az olduğundan dolayı 107-108 hücre/ml yoğunluğa kadar
ulaşabilmektedirler. Halofilik arkelerin bakteriofajlarla lizisinin kommünite büyüklüğü
üzerinde etkili olabildiği belirtilmiştir (Oren et al. 1997). Halosinler laboratuvar
ortamında diğer halobakterilerin üremesini emgelleyen proteinlerdir. Halobakteriler
arasında oldukça yaygın olan bakteriyosin (halosin) üretiminin, hiper tuzlu akuatik
ortamlardaki rekabette katkısının önemli olmadığı gösterilmiştir (Rodriguez-Valera et
al. 1981, Meseguer et al. 1986, Torreblanca et al. 1994, Kis-Papo ve Oren 2000).
2.4. Halofilik Arkelerde Ozmotik Adaptasyon
Doygun tuz konsantrasyonunda (5.2 M NaCl) yaşayan Halobacteriales ordosu üyeleri,
ortamdaki yüksek ozmotik basınca karşı koyabilmek için stoplazmalarında başta K+ ve
Cl- olmak üzere yüksek oranlarda iyonlar biriktirmektedirler ("salt-in" stratejisi). 3.5 M
NaCl’den daha yüksek olan hücre içi tuz konsantrasyonunun (yaklaşık 5M) büyük
kısmını KCl ve daha az olmak üzere NaCl oluşturmaktadır (Kushner 1985, Hough ve
Danson 1989). Halofilik arkeler, iç ortamlarında ozmolit çözünür madde olarak dış
ortamdakinin 100 katı oranında KCl biriktirdikleri için, hücresel bileşenleri de bu tuz
konsantrasyonuna adapte olup proteinlerin stabilitesi ve fonksiyonu için bu tuz oranı
gereklidir. Halofilik arkeler, protein yapısında, diğer canlılara göre negatif yüklü amino
asit sıklığını arttırarak (%20 daha fazla) ve ortamın salting-out etkisini kompanse etmek
19

için polar olmayan amino asit miktarını azaltarak adapte olmuşlar ve halofilik enzimler
yaklaşık 1 M’ın altındaki NaCl/KCl konsantrasyonlarında hızla denatüre olmaktadırlar
(Jaenicke ve Zavodszky 1990, Danson ve Hough 1997, Sellek ve Chaudhuri 1999,
Hough ve Danson 1999, Mevarech et al. 2000, Kunte et al. 2001). H. salinarum NRC-1
proteomunun oldukça asidik karakterde olduğu (izolektrik noktası 4.9) ve diğer
canlıların proteinlerinin denatüre olduğu doygunluk derecesindeki hücre içi tuz
konsantrasyonunda stabil kaldığı belirtilmiştir. Bazı halofilik arkelerin (Natronococcus
occultus, Natronobacterium gregoryi v.b.) sitoplazmalarında yüksek tuz
konsantrasyonunun yanısıra organik ozmolit çözünür madde olarak 2-sülfo trehaloz da
saptanmıştır. Enzimatik mekanizmanın adaptasyonunda organik ozmolit
biriktirilmesinin enerjetik yönden KCl’e göre ekonomik olmadığı belirtilmiştir. Salt-in
sitoplazma stratejisiyle hücre içerisinde dışarıya göre daha yüksek oranda K+ biriktiren
halofilik arkeler, Na+ iyonlarını Na+/H+ antiport sistemi ile dışarı atmaktadırlar (Oren
2000, Oren 1999, Roeßler ve Müller 2001, Oren 2002). Potasyum ile birlikte karşıt iyon
olarak Cl- da stoplazmada yüksek oranda bulunmakta ve enerji gereksinimi olan aktif
Cl- alımının büyüme ve bölünme aşamalarında hücrelerin hacimlerini arttırmaları için
gerekli olduğu belirtilmiştir. Halofilik arkelerde Cl- alımı ile ilgi 2 sistem
tanımlanmıştır; ışıkla çalışan klor pompası rodopsin ve ışığa bağımlı olmayan transport
sistemi (Kunte et al. 2001, Müller ve Oren 2003). Ayrıca KCl’ün H. salinarum’da
DNA’da zarar oluşturan oksidatif letal faktörlere karşı organizmayı koruduğu da rapor
edilmiştir (Shahmohommadi et al. 1998).
2.5. Halofilik Arkelerin Yapısal, Genetik ve Fizyolojik Özellikleri
2.5.1. Polar ve nötral lipitler
Halobacteriales ordosu üyelerinin hücre zarlarında, gliserole eter bağlı dallanmış 20-
karbonlu (fitanil) ve bazen de 25-karbonlu (sesterpanil) zincirleri bulunmaktadır. Nötral
lipitlerden farklı olarak fosfolipit, sülfolipit ve glikolipitlerden oluşan polar lipit tipleri
suşların taksonomisinde önemli bir özellik olarak kullanılmaktadır (Torreblanca et al.
1986, Kates 1993). Ayrıca tuzlu bölgelerdeki biyomasın polar lipit kompozisyonları ile
ilgili bilgiler, doğal kommunitelerdeki mevcut halofilik arke tip veya tiplerinin
20

belirlenmesi için de kullanılmaktadır (Oren ve Gurevich 1993, Oren et al. 1996,
Litchfield et al. 2000). Polar lipitlerin karakterizasyonu ince tabaka kromatoğrafisi,
kütle spektrometresi, NMR spektroskopisi gibi yöntemler ile yapılabilmektedir (Ross et
al. 1981, Kates 1993). Halobacteriaceae üyelerinde bulunan polar lipitlerin büyük bir
kısmı C20C20 temel lipit (2,3-di-O-fitanil-sn-gliserol) yapısındadır (şekil 2.3.a).
Alkalifilik türlerin büyük bir kısmı ve bazı nötrofilik türler farklı oranlarda C25C20 (2-O-
sesterpanil-3-O-fitanil-sn-gliserol içermektedirler (şekil 2.3.b) (Kamekura ve Kates
1988, Kamekura ve Dyall-Smith 1995, Mcgenity et al. 1998, Morita et al. 1998). Bazı
halofillerde minör bileşen olarak C25C25’e (2,3-di-O-sesterpanil-sn-gliserol)
rastlanılmakta (şekil 2.3.c) (Kamekura ve Kates 1999), C40 şeklindeki bidiftanil tetraeter
yapısına bugüne kadar rastlanılmamıştır (Grant et al. 2001).
CH2OH
CH2OH
CH2OH
CH
CH
CH
CH2
CH2
CH2
O
O
O
O
O
O
Şekil 2.3. Halofilik arkelerde bulunan temel lipitler (Kates 1992).
a: C20C20, b: C25C20, c: C25C25
Tüm halofilik arkeler fosfatidil gliserol (PG) ve metil ester fosfatidil gliserofosfat (Me-
PGP) dieter türevleri içerirler. Fosfatidil gliserosülfat (PGS), nötrofilik türlerin çoğunda
mevcut iken bazı nötrofilik üyelerin yanısıra, özellikle alkalifilik üyeler PGS
içermeyişleri ile karakterize edilebilmektedirler. Halobacteriales’in farklı üyelerinde
21

di-, tri- veya tetraglikozil dieter yapısında çeşitli glikolipitler tanımlanmıştır: mannozil-
(1-2)-glukozil-gliserodieter (DGD-1), mannozil-glukozil-gliserodieter (DGD-2
tanımlanmamış), glukozil-(1-6)-glukozil-gliserodieter (DGD-4), mannozil-6-Sülfat-(1-
2)-glukozil-gliseroldieter (S-DGD-1), mannozil-2-sülfat-(1-4)-glukozil-gliseroldieter
(S-DGD-3), mannozil-2-sülfat-(1-2)-glukozil-gliseroldieter (S-DGD-5), mannozil-2,6-
bisülsat-(1-2)-glukozil-gliseroldieter (S2-DGD-1), galaktozil-(1-6)-mannozil-(1-2)-
glukozil-gliserodieter (TGD-1), glukozil-(1-6)-mannozil-(1-2)-glukozil-gliserodieter
(TGD-2), galaktozil-3-sülfat-(1-6)-mannozil-(1-2)-glukozil-gliserodieter (S-TGD-1),
galaktozil-(1-6)-mannozil-(3-1)-galaktozil-(1-2)-glukozil-gliserodieter (TeGD), galakto-
zil-3-sülfat-(1-6)-mannozil-(3-1)-(galaktozil)-(1-2)-glukozil-gliserodieter (S-TeGD)
(Hancock ve Kates 1973, Kates ve Deroo 1973, Kamekura ve Kates 1988, Trincone et
al. 1990, Kates 1993, Upasani et al. 1994, Kates 1996, Kamekura ve Kates 1999).
Halofilik arkelerin total lipit içeriklerinin yaklaşık %10’unu nötral lipitlerin oluşturduğu
rapor edilmiş olup başlıca tipleri şunlardır: C20 isoprenoid lipidlerden geranilgeraniol,
gliserolün nötral fitanil eterlerinden DL-O-fitanil-sn-gliserol ve 2,3-di-O-fitanil-sn-
gliserol, C30-isoprenoid bileşiklerinden squalen, dihidrosqualen, tetrahidrosqualen ve
dehidrosqualen (Kamekura ve Kates 1988, Oren 2001).
2.5.2. Karotenoid ve retinal pigmentler
Halobacteriales ordosunun pek çok üyesi hücre zarlarında yüksek oranda içerdikleri α-
bakterioruberin’in C50 düz-zincir türevli karotenoid pigmentlerinden ve daha düşük
miktarda içerdikleri C40 tipi karotenoid olan likopen ve β-karotenden dolayı parlak
kırmızıdan turuncuya kadar değişen renk tonlarında görülürler. Karotenoid
pigmentlerinin hücreyi iyonize radyasyon ve UV radyasyonu gibi DNA’ya zara veren
letal faktörlere karşı korudukları ve fotoreaktivasyona yardım ettikleri ileri sürülmüştür
(Wu et al. 1983, Tindall 1992).
Halofilik arkelerde 4 adet retinal pigment içeren protein tanımlanmıştır:
bakteriyorodopsin (ışıkla çalışan dış proton pompası), halorodopsin (ışıkla çalışan iç CI−
pompası) ve 2 adet alıcı rodopsin (ışığa duyarlı fototaksis olayına katılırlar). Mor
22

pigmentli bakterioyrodopsin, H. salinarum ve Halorubrum sodomonse’de bulunur
(Oesterhelt 1998, Oren 1983, Oren 2001) ve H. salinarum’daki bakteriyorodopsin
kromofor (retinal) içeren tek bir polipeptit zincirinden oluşmuş integral bir zar
proteinidir ve hücre dışına protonların ışığa bağlı translokasyonuyla transmembran
elektrokimyasal gradienti oluşturur ve ATP’nin sentezlenmesini sağlar (Kushner 1985,
Yatsunami et al. 2000).
2.5.3. Diğer yapısal özellikleri
Halobacteriales’in bir çok üyesi, çözünür oksijen miktarı düşük olan tuzlu sularda
oksijen ve ışık alımını arttıran ve yüzme olanağı sağlayan gaz vezikülleri içermek-
edirler. Bunların UV ışınlarının zararlı etkilerine karşı koruyucu kalkan oldukları belir-
tilmiştir (DasSarma 1993, Beard et al. 1997).
Diğer prokaryotik gruplarda taksonomik marker olarak kullanılan ve halofilik arkelerde
konsantrasyonları düşük olan poliaminlerin taksonomik olarak önemli olmadığı rapor
edilmiştir (Kamekura et al. 1986, Hamara et al. 1995).
Halobacteriales ordosunun non-kokkoid temsilcilerinin yüksek molekül ağırlığına sahip
glikoproteinlerden oluşmuş hücre duvarına sahip oldukları belirtilmiştir. Halococcus
türleri sülfatlı heteropolisakkarit yapısında kalın bir hücre duvarına sahiptirler. Natrono-
coccus occultus, farklı olarak tekrar eden poli L-glutamin ünitelerine sahip kalın bir
hücre duvarı içermektedir (Kushner 1985, Niemetz et al. 1997, Messner et al. 1997,
Tindall 1992, Oren 2001).
2.5.4. Genetik yapıları
Joshi et al. (1963) CsCl buoyant density satrifügasyon yöntemini kullanarak
Halobacteriaceae familyasındaki pek çok türün DNA’nın majör "FI DNA" ve minör
"FII DNA" (kromozomal DNA daki A+T’ce zengin adalar veya satellit DNA) bölgeleri
içerdiklerini belirtmişlerdir. FII fraksiyonu, total DNA’nın %11-36’lık bölümünü
oluşturmakta olup daha çok heterojen bir plazmid populasyonundan olştuğu, ancak kro-
23

mozom içerisinde de FII adalarının bulunduğu ve bunların daha çok insersiyon
sekansları olduğu ve H. salinarum’da yüksek miktarda bulunduğu, dolayısıyla yüksek
sıklıkta (10-2) meydana gelen spontan mutasyonların nedeni oldukları rapor edilmiş olup
(Pfeifer et al. 1988, Pfeifer ve Blaseio 1989, Pfeifer et al. 1989, DasSarma 1993, Ng et
al. 1998) Halobacteriales’in diğer üyelerinin daha stabil bir genoma sahip oldukları
belirtilmiştir (Lopez-Garcia et al. 1995). H. salinarum NRC-1’in genomik dizisi tam
olarak çıkarılmış olup 2Mb (2.014 kb) büyüklüğünde halkasal bir kromozom, pNRC200
(365 kb) ve pNRC100 (191 kb) şeklinde iki adet minikromozom içerdiği ve genomunun
yaklaşık 2630 adet protein geni bulundurduğu belirtilmiştir (Ng et al. 1998, Ng et al.
2000, Kennedy et al. 2001, Goo et al. 2003). Halofilik arkeler küçük molekül ağırlıklı
plazmidlerin yanısıra 100-300 kb arasında değişik plazmidler (megaplazmidler)
içermektedirler (Pfeifer et al. 1981a, Pfeifer et al. 1981b, Pfeifer et al. 1988, Ross ve
Grant 1985, Gutierrez et al. 1986, Rosenshine ve Mevarech 1989, Hackett et al. 1989,
Akhmanova et al. 1993). Plazmidlerin fonksiyonları tam olarak bilinmemesine rağmen
bazı halofilik arkelerde gaz keseciği ve bakteriyoruberin sentezi ile ilgili olabildiği rapor
edilmiştir (Pfeifer et al. 1981a, DasSarma ve Arora 1997).
2.5.5. Fizyolojileri
Halofilik arkeler aerobik heterotrofik prokoryatlar olup karbon kaynaklarının aerobik
yıkımı, glioksilat döngüsü ve sitokrom zinciri içeren elektron transport sistemi
kombinasyonu ile trikarboksilik asit döngüsü temelinde gerçekleşmektedir. Embden
Meyerhof metabolik yolu, hekzokinaz ve fosfofruktokinaz aktivitelerinin düşüklüğü ve
tuzla güçlü bir şekilde inhibe edilmeleri nedeniyle Halobacteriaceae’de işlevsel değildir
(Rawal et al. 1988). Bunun yerine muhtemelen glukozun yıkımı, fosforilasyon
aşamasının atlandığı bir modifiye Entner-Doudoroff metabolik yol ile
gerçekleştirilmektedir. Aminoasitler, şekerler ve organik asitler gibi basit bileşiklere ek
olarak bazı polimerik maddeler de halofilik arkeler tarafından yıkılabilmektedir.
Halobacteriales’in birçok türü proteaz, lipaz, DNaz ve amilaz gibi ekzoenzimleri de
üretmektedir (Izotova et al. 1983, Kushner 1985, Johnsen et al. 2001, Oren 2001). Tuza
doymuş ortamlarda oksijen çözünürlüğünün az olmasından dolayı halofilik arkelerin
çoğu anaerobik olarak üreme yeteneğine sahip olup bu amaçla fumarat, trimetilamin N-
24

oksit (TMAO), dimetilsülfoksit veya nitrat gibi alternatif elektron alıcıları, arjinin
fermentasyonu veya bakteriyorodopsin kullanırlar (Hartmann et al. 1980, Mancinelli ve
Hochstein 1986, Oren ve Trüper 1990, Oren 1991, Ruepp ve Soppa 1996, Oren ve
Litchfield 1999, Goo et al. 2003).
2.6. Halofilik Arkelerin Antimikrobiyal Maddelere Duyarlılıkları
Halofilik arkeal suşların karakterizasyonu veya karşılaştırılmasıyla ilgili taksonomik
çalışmalarda antibiyotikler ve diğer antimikrobiyal madellere duyarlılık oldukça fazla
kullanılmaktadır (Oren et al. 1997). Mürein yapıda hücre duvarına sahip olmayan
halofilik arkeler penisilin, ampisilin, sikloserin, kanamisin, neomisin, polimiksin,
karbenisillin, sefatoksim ve streptomisin gibi bakterilere spesifik antibiyotiklere
dirençlidirler. Çelişkili olmasına rağmen, yapılan çalışmaların çoğunda bu canlıların
kloramfenikol ve tetrasikline duyarlı oldukları rapor edilmiştir. Bunun dışında halofilik
arkelerin novobiosin, bazitrasin, rifampisin ile ökaryotlarda protein sentez inhibitörü
olan anizomisin ve quinolon türevlerine karşı duyarlı oldukları tespit edilmiştir (Hilpert
et al. 1981, Pecher ve Böck 1981, Bonelo et al. 1984, Böck ve Kandler 1985, Sioud et
al. 1988, Oren 1996). Ayrıca bu canlıların Streptomyces venazuale subsp.
xanthophaeus’ca üretilen halokinon ile DNA polimeraz inhibitörü olan afidikolin ve
safra tuzlarına karşı duyarlı oldukları belirtilmiştir (Oren 1990, Kamekura et al. 1988,
Oren 2001).
2.7. Halofilik Arkelerin Uygulama Alanları
Potansiyel biyoteknolojik uygulamalara sahip olan kırmızı halofilik arkelerin, tuzla-
kristalizasyon göllerinde solar ısınmayı ve evaporasyonu arttırarak tuz üretiminin
arttırılmasında kullanılabilecekleri ileri sürülmüştür (Jones et al. 81, Grant et al. 1998,
Litchfield 2002). Bakteriyorodpsinin, güneş ışığından elektrik üretimi, deniz suyundan
tuz giderimi, kimyasal ve biyosensörlerde kullanılması ve ultrahızda ışık saptanması
gibi alanlarda da potansiyel olarak kullanılabileceği rapor edilmiştir (Kushner 1985,
Hough ve Danson 1989, Margesin ve Schinner 2001).
25

Ayrıca Haloferax ve Haloarcula cinslerine ait bazı türlerin, ürettikleri ekzopolisakkarit
yoluyla vizkozite stabilizasyonunda, jelleştirme ajanı ve emülsifiyer olarak ve petrolün
mikrobiyolojik geri kazanımını artırmak için kullanılabilecekleri belirtilmiştir (Ventosa
ve Nieto 1995, Parolis et al. 1996, Margesin ve Schinner 2001). Bazı halofilik arke poli-
β-hidroksi alkonat (PHA) ürettikleri belirtilmiştir (Lillo ve Rodriguez-Valera 1990,
Hezayen et al. 2000). Yağsız katkı maddesi olarak gliserol dieter lipitlerin besin katkı
maddesi olarak kullanılabileceği ileri sürülmüştür (Post ve Collins 1982). İlaç ve
kozmetiklerin yapısında kullanılan lipozomlar’ın yapımında halofilik arkelerin
esterazlara karşı yüksek bir kimyasal stabiliteye sahip olan eter bağlı lipitleri
kullanılmaktadır. Amilaz, amiloglukozidaz, proteaz ve lipaz gibi yüksek tuzlulukta
işlevsel olan ekzoenzimleri, yüksek tuz konsantrasyonunda makromoleküllerin yıkımı
ile ilgili biyoteknolojik proseslerde kullanılabilirler (Chaga et al. 1993, Ventosa ve
Nieto 1995, Margesin ve Schinner 2001).
Düşük su aktivitesine adapte olan halofilik enzimlerin, endüstriyel organik sentezlerdeki
yüksek özgüllükleri, stereoözgüllükleri ve etkinliklerinden dolayı önemleri artmaktadır
(Hough ve Danson 1989, Danson ve Hough 1997, Sellek ve Chaudhuri 1999). Diğer
yandan hidrokarbonları parçalama yeteneğine sahip halofilik arkeler petrol kazalarında
dökülen petrolün bioremediasyonunda kullanılabilirler (Bertrand et al. 1990, Emerson
et al. 1994). İnsektisit, lindan, DDT veya triklorofenoller gibi halojenli organik
bileşiklerrin yüksek konsantrasyonlarında üreyebilen halofilik arkelerin patentli
proseslerde de kullanıldığı belirtilmiştir (Margesin ve Schinner 2001).
H. salinarum NRC-1 suşunun, lipit içermeyen protein yapısında zara sahip olan ve suya
geçirgen olmayıp gazlara geçirgen olan, yüksek oranda stabilite gösteren ve proteolitik
yıkıma dayanıklı non-toksik gaz keseciklerinin epitop düzenlenmesi amacıyla yeni bir
sistem olarak geliştirilmesi düşünülmüştür (Stuart et al. 2001).
Halosinlerin, tuzlanmış yiyeceklerin veya derilerin tabaklanması esnasında meydana
gelebilen bakteriyel bozunumun kontrolü amacıyla kullanılabilecekleri ve bunlardan
HalH7 halosininin halobakterilerdeki gibi memelilerde de benzer etkiye sahip olduğu
belirtilmiştir (Litchfield 2002). H. salinarum’daki 84 kD luk bir protein, kanser
26

hastalarının serumlarındaki insan c-myc onkogen ürününe karşı antikor elde etmek için
bir antijen olarak kullanılmış ve bu proteinin bazı kanser tiplerinin saptanmasında
kullanılabileceği belirtilmiştir (Rodriguez-Valera 1992).
27

3. MATERYAL ve YÖNTEM
3.1. Materyal
3.1.1. Halofilik arkeler
Türkiye’den halofilik arkelerin izolasyonu amacı ile 6 farklı tuzlu bölge belirlenmiştir:
1) Tuz gölü-Şereflikoçhisar-Ankara (Anonim 1999), 2) Acıgöl-Denizli (M.T.A. derleme
rapor 1955, Yurdakulol et al. 1996), 3) Salda gölü-Burdur (Braithwaite ve Zedef 1994),
4) Seyfe gölü-Kırşehir (Eyüboğlu 1995, Sözeri 2000), 5) Tuzla gölü-Kayseri (Kibar
1999), 6) Bolluk gölü-Konya (Canik 1988, Anonim 1999). Yukarıdaki 6 farklı gölden
2000-2001 yıllarında yaz-sonbahar döneminde toprak, su ve tuz kütlesi örnekleri steril
plastik kaplar içerisine alınmış, laboratuvara bekletmeden transfer edilmiştir. Standart
halofilik arke suşu olarak kullanılan Halobacterium salinarum, Haloarcula
marismortui, Haloarcula vallismortis, Haloferax mediterranei, Haloferax volcanii ve
Natrialba asiatica Prof. Dr. Aharon Oren’den (Division of Microbial and Molecular
Biology, The Institute of Life Sciences and the Moshe Shilo Minerva center for Marine
Biochemistry, The Hebrew Universty of Jerusalem, Jerusalem, Israil), Halococcus
morrhuae CCM 537 Çek Mikroorganizma Kolleksiyonundan (Czech Collection of
Microorganisms, Masaryk Universty Brno, Çek Cumhuriyeti) ve Haloarcula
vallismortis DSM 3756 Alman Mikroorganizma Kültür Kolleksiyonu’dan temin
edilmiştir. Öbakteriyel standart suş olarak da Enterococcus faecium CDC NJ-1 Ankara
Üniversitesi Fen Fakültesi Biyoloji Bölümü Bakteriyoloji Laboratuvarından temin
edilmiştir. Türkiye’den izole edilen halofilik arkebakteri izolatları ve standart suşlar
%20 gliserollü ortamda -70°C’de saklanmıştır (Xin et al. 2000).
3.1.2 Besiyeri
Tuz, toprak ve su örneklerinden halofilik arkebakterilerin izolasyonu ve standart
suşların kültürü amacıyla Sehgal-Gibbons besiyeri (SG) (Gonzalez et al. 1978,
Montalvo-Rodriguez et al. 2000) kullanılmış olup 250g/L NaCl, 20g/L MgSO4.7H2O,
28

2g/L KCl, 3g/L Sodyum sitrat, 0.023g/L FeSO4.7H2O, 7.5g/L Casamino asit, 1g/L
Yeast extract, içeren besiyerinin pH’ı 1N KOH ile 7.3’e ayarlanmıştır.
3.1.3. Mikroskobik inceleme
İzolatların hücre morfolojisi ve hareket yeteneklerinin belirlenmsi ve fotoğraf
çekimlerinde Olympus Vanox Mikroskop (faz kontrast ataçmanlı) ve Olympus Pm-10A
fotoğraf ataçmanı, Koloni morfolojisinin belirlenmesinde ise Stereo mikroskop
(Euromex Arnhem) kullanılmıştır.
3.1.4. Antibiyotikler
İzolatların duyarlılıklarını belirlemek için Norfloksasin (10µg), Tetrasiklin (30µg),
Bazitrasin (10 IU), Rifampisin (5µg), Azitromisin (15µg), Ampisilin (10µg), Neomisin
(30µg), Kloramfenikol (30µg), Penisilin G (10 IU), Vankomisin (30µg), Novobiyosin
(30µg) (OXOID) antibiyotikleri kullanılmıştır.
3.1.5. İnce tabaka kromatografisi
İnce tabaka kromatografisi için Liyofilizasyon cihazı (Edwards), metanol, toluen,
sülfürik asit, hekzan, silikajel plakaları (Merck, 60F254, 20x20, alüminyum plaka), petrol
eteri, dietil eter, dodekamolibdofosforik asit, etanol kullanılmıştır.
3.1.6. Elektroforez
Protein elektroforezi için, sodyum dodesil sülfat (SDS), merkaptoetanol, bromofenol
mavisi, akrilamid-bisakrilamid monomerleri, trizma base, amonyum per sülfat (APS),
tetra metil etilen diamin (TEMED), glisin, coomassie brillant mavisi R250 (Sigma B-
0149), molekül ağırlık standardı (Sigma MW-SDS-200) ve gliserol, izopropil alkol,
asetik asit, bütanol, hidroklorik asit (Merck) kimyasalları ile dikey elektroforez tankı,
güç kaynağı, ışıklı tabla kullanılmıştır.
29

Agaroz jel elektroforez için, Tris, Etilen diamin tetra asetik asit (EDTA), Sodyum
klorür, Tris-HCl, SDS, Sodyum asetat, bromofenol blue, etanol, RNaz A, Agaroz,
etidyum bromür ve supercoiled ccc DNA marker (Sigma D5292) ile yatay elektroforez
tankı, güç kaynağı, Ray Test görüntüleme ve analiz cihazı ve Diana Densitometrik
Analiz Programı kullanılmıştır.
3.2. Yöntem
3.2.1. Halofilik arkelerin izolasyonu ve kültürü
Tuzlu bölgelerden toplanan toprak ve tuz örnekleri steril şartlar korunarak 10-13 g
tartılmış, 50 mL steril tuz çözeltisi ile (SG besiyerinin inorganik kısmı) süspanse
edilmiştir. Süspansiyonlardan ve su örneklerinden katı ve sıvı SG ortamlarına ekim
yapılarak, sıvı ortamlar 120 rpm’de çalkalamalı olarak 37 °C’de üretilmiştir. Hem
toprak hem de sıvı örneklerinden ekim yapılan gelişmiş kültürlerden tekrar katı
ortamlara ekim yapılmıştır. İnkübasyonun yaklaşık olarak 3. gününden itibaren
besiyerlerinde pigmentsiz koloniler gözlenmiş, 10-15 gün sonra ise pembe-kırmızı
renkte koloniler gözlenmiştir. Farklı morfoloji gösteren kolonilerden seçim yapılarak
besiyerlerine tekrar ekilmiş, saf kültürler hazırlanarak saflık kontrolleri koloni
morfolojisi ve faz kontrast mikroskobisi çalışmaları ile yapılmıştır.
3.2.2. Koloni morfolojisi ve pigmentasyon
İzolatların koloni morfolojisi, pigmentasyonu ve mukoid koloni özelliği standart
kriterlere göre, katı ortamda 7 gün geliştirilmiş kültürler kullanılarak belirlenmiştir.
Koloni morfolojileri mikrometrik okülerli, 4X büyütmeli stereo mikroskop kullanılarak
yapılmış pigmentasyon ise görsel olarak belirlenmiştir.
30

3.2.3. Gram boyama ve faz-kontrast mikroskobu
Gram boyama Dussault (1955)’e göre yapılmış 7 günlük gelişmiş sıvı kültürlerden
hazırlanan preparatlar %2 asetik asitle ile fikse edilip boyama işlemi yapılarak
immersiyon objektifi ile ışık mikroskobunda incelenmiştir.
Hücre morfolojleri ve hareket 7 gün geliştirilen sıvı kültürlerden hazırlanan preparatlar
immersiyon büyütmeli faz kontrast mikroskobunda incelenerek belirlenmiştir.
3.2.4. Biyokimyasal testler
Biyokimyasal testlerde Oren et al. (1997) , Colwell et al. (1979) ve Tindall (1992)’a
göre belirlenen biyokimyasal testler yapılmıştır. Yedi günlük katı ve sıvı kültürler
kullanılmış olup testler 37 °C’ de ve sıvı ortamda 120 rpm’de yapılmıştır. Testler 5
farklı cinse ait 7 farklı Halofilik arke türü ve pozitif kontrol bakteriler kullanılarak 3
tekrarlı olarak yapılmıştır.
Katalaz aktivitesi, SG agarda geliştirilmiş 7 günlük kültür üzerine %3’lük H2O2
dökülerek belirlenmiştir (Holding ve Collee 1971, Rodriguez-Valera et al. 1983,
Franzmann et al. 1988, Takashina et al. 1990, Arda 1997, Montalvo-Rodriguez et al.
1998, Montalvo-Rodriguez et al. 2000).
Oksidaz aktivitesi, SG agarda geliştirilmiş 7 günlük kültürlerle %1 tetrametil-p-
fenilendiamin ayıracı kullanılarak saptanmıştır (Holding ve Collee 1971, Gonzalez et al.
1978, Franzmann et al. 1988, Takashina et al. 1990, Montalvo-Rodriguez et al. 2000).
Jelatin ve Tween 80 hidrolizi, Gutierrez ve Gonzalez (1972)’e göre yapılmış olup 14
günlük inkübasyon sonrası jelatin hidrolizini belirlemek için Frazier’s ayracı
kullanılmıştır.
31

Nişasta hidrolizi, %1 oranında çözünür nişasta içeren SG agarda geliştirilmiş 14 günlük
kültürler ve Lugol ayracı kullanılarak belirlenmiştir (Holding ve Collee 1971, Takashina
et al. 1990, Temiz 1994, Arda 1997, Montalvo-Rodriguez et al. 1998).
Kazein hidrolizi, %0,15 oranında Skim Milk Powder içeren SG agar besiyerinde
geliştirilmiş 14 günlük kültürler ve %1 HCI kullanılarak belirlenmiştir (Tomlinson ve
Hochstein 1976, Gonzalez et al. 1978, Arda 1997).
İndol oluşumu, %1 Tripton içeren SG sıvı besiyerinde geliştirilmiş 10 günlük kültürler
üzerine Kovac’s ayracı eklenerek tayin edilmiştir (Holding ve Collee 1971, Rodriguez-
Valera et al. 1983, Temiz 1994, Arda 1997).
Sodyum tiyosülfattan (Na2S2O3.5H2O) H2S üretimi, izolatların %0.5 sodyum tiyosülfat
ve kurşun asetat emdirilmiş kağıt şeritler içeren SG sıvı besiyerinde geliştirilmesiyle
saptanmıştır (Holding ve Collee 1971, Franzmann et al. 1988, Takashina et al. 1990,
Arda 1997).
Nitratın nitrite indirgenmesi ve gaz oluşumu, %1 oranında KNO3 içeren Durham tüpü
bulunan SG broth besiyerindeki kullanılmıştır. 10 günlük kültürlere %1 sülfanilik asit
ve %0.6 α-naftilamin ayraçları eklenmesiyle belirlenmiştir (Holding ve Collee 1971,
Tomlinson ve Hochstein 1976, Gonzalez et al. 1978, Arda 1997, Montalvo-Rodriguez
et al. 1998, Xin et al. 2000).
Şekerlerden asit üretiminin belirlenmesi amacıyla 7 farklı şeker kullanılmıştır: D(+)-
glukoz, D(-)-fruktoz, L(+)-arabinoz, D(+)-galaktoz, sükroz, D(+)-ksiloz, maltoz.
Şekerler filtre edilerek, %1 oranında olacak şekilde steril besiyerine eklenmiş 12-14
günlük inkübasyon sonrası kültürlere 1 damla fenol red damlatılarak asit üretimi
saptanmıştır (Rodriguez-Valera et al. 1983, Torreblanca et al. 1986, Oren et al. 1999,
Xin et al. 2000)
Arjinin varlığında anaerobik üremenin belirlenmesi amacıyla, 5g/L L-arjinin içeren SG
besiyeri vida kapaklı kültür tüplerine doldurularak ekim yapılmış ve 5-7 gün karanlıkta
32

inkübe edilmiştir. İnkübasyon sonrası üremenin varlığı 600 nm’de turbidite okunarak
saptanmıştır. Kontrol için arginin içermeyen besiyerlerine ekim yapılmıştır (Oren ve
Litchfield 1999, Hartmann et al. 1980).
3.2.5. Antibiyogram testi
İzolatların antibiyotik duyarlılık testleri disk difüzyon yöntemi ile yapılmıştır (Bonelo et
al. 1984, Montalvo-Rodriguez et al. 2000). Sıvı ortamda üretilen kültürlerin
Spektrofotometrede 520 nm dalga boyunda absorbansları 0,8’e ayarlanmış
süspansiyondan steril eküvyonla SG agar yüzeyine ekim yapılmış ve antibiyotik diskleri
steril pens ile besiyeri yüzeyine yerleştirildikten sonra plaklar 37 °C’de 12-14 gün
inkübe edilmiştir.
3.2.6. İnce tabaka kromatografisi
Arkeal lipit varlığının belirlenmesi amacı ile, izolatların tüm hücre asit
metanolizatlarının ince tabaka kromatoğrafik analizi yapılmıştır (Ross et al. 1981). SG
sıvı besiyerinde üretilen kültürlerden hücre peleti santrifüjle toplanarak liyofilize edilen
pelet üzerine 3mL metanol, 3mL toluen ve 0,1 mL konsantre sülfürik asit eklenerek 50
°C’de 15-18 saat bekletilmiştir. Hücre zarı lipitlerinin ekstraktsiyonu için karışım
üzerine 1,5 mL hekzan eklenmiş ve ekstraklar silika gel plakalarına yüklenerek petrol
eteri/dietil eter (85:15) karışımında ayırımları gerçekleştirilmiştir. Plakalar,
dodekamolibdofosforik asit çözeltisi püskürtülerek lipit lekeleri görünür hale gelinceye
kadar 150 °C’de ısıtılmıştır. Lipit yapıları sarı zemin üzerinde koyu mavi renkli spotlar
halinde görünür hale gelmiştir. Standart olarak ester bağlı öbakteriyel yağ asit yapısı ile
karşılaştırmak amacıyla Enteroccus faecium CDC NJ-1 ve Tuz gölünden izole edilen
halofilik öbakteri izolatı kullanılmıştır.
3.2.7. Protein ekstraksiyonu
Toplam 95 halofilik arke izolatı ve 7 standart suşun tüm hücre SDS-PAGE analizleri
Hesselberg ve Vreeland (1995) ve Laemmli (1970)’ye göre yapılmıştır. İzolatların 37
33

°C’de 3-4 gün süreyle 3 kez çalkalamalı olarak inkübe edilmesiyle senkron kültürleri
hazırlanmıştır. Üç mL’lik sıvı kültürlerden santrifüjle toplanan peletler -70°C’de
saklanmış peletlerin üzerine SDS-örnek tamponundan (Laemmli 1970) 50µL eklenerek,
7 dakika kaynar su banyosunda tutulmuştur. Ekstraklardaki protein miktarının yarı
kantitatif tayini için spot test yapılmıştır (Esen 1978).
3.2.7.1. Sodyum dodesil sülfat poliakrilamid jel elektroforezi (SDS-PAGE)
Laemmli (1970)’e göre yapılan SDS-PAGE’de %4’lük yığma jeli ve %10’luk ayırma
jeli kullanılmış, örnekler yüklendikten sonra sırasıyla 25 ve 30 mA’de elektroforez
yürütülmüş, jel bir gece Coomassie Brillant mavisinde boyandıktan sonra dekolorize
edilmiş ve fotoğrafı çekilmiştir.
3.2.7.2. Proteinlerin moleküler kütlelerinin belirlenmesi
Molekül kütleleri hesaplanacak olan protein bantlarının Rf (oransal hız) değerleri,
standart proteinlerin Rf değerleri kullanılarak Excel programında çizilen yarı logaritmik
grafiğin kalibrasyon eğrisi aracılığı ile hesaplanmıştır.
3.2.7.3. Benzerlik matriksi ve protein ilişkisine göre benzerlik dendogramı
Bu amaçla, majör protein bantları değerlendirmeye alınmış bulunma veya bulunmama
durumuna göre bantlar "1" veya "0"şeklinde değerlendirilme yapılarak benzerlik
matriksi oluşturulmuştur.
Bu amaçla Nei ve Li (1979) tarafından geliştirilen ve aşağıda verilen Nei ve Li
Katsayısı (coefficient-NLcij) kullanılarak iki izolat arasındaki benzerlik
karşılaştırılmaları yapılmıştır.
(Nei ve Li Katsayısı) NLcij = [2N11/(2N11+N10+N01)]
N11 : i ve j bireylerinin her ikisindede bulunan bant sayısı
34

N10 : sadece i bireyinde bulunan bant sayısı
N01 : sadece j bireyinde bulunan bant sayısı
İzolatlar arasındaki protein profili benzerliklerine dayalı dendogram verileri MVSP
Version 3.1 (Multi-Variate Statistical Package) programı kullanılarak "UPGMA" analizi
ile belirlenmiştir (Sneath ve Sokal 1973).
3.2.8. Plazmid izolasyonu
SG broth besiyerinde 37 °C’de çalkalamalı olarak 4 gün geliştirilen kültürlerden 1,5 mL
alınarak 13000 devir/dk.’da 3 dakika santrifüj edilerek pelete 330 µl Tris-EDTA
(50mM:1mM, pH:8) ve 48.2 µL Tris-EDTA-1 (50mM:0.25mM, pH:8) tamponlarından
ilave edilerek karıştırılmıştır. Bunun üzerine %20’lik SDS çözeltisinden 27,6 µL
eklenerek karıştırılmış ve 37 °C’lik su banyosunda 10 dakika bekletilmiştir. Bu süre
sonunda tüpler 30 saniye yüksek devirde vortekslenmiş taze hazırlanmış 3 N NaOH
çözeltisinden 27,6 µL ilave edilerek tüpler yavaşça 10 dakika süresince alt üst
edilmiştir. Daha sonra 49,6 µL 2M Tris-HCl (2M, pH:7) çözeltisi eklenerek 5 dakika
tekrar alt üst edilerek karıştırılmıştır. Tüplere +4°C’deki 5M NaCl çözeltisinden 71.7
µLve %3 NaCl ile doyurulmuş Fenol çözeltisinden 700 µL ilave edilerek +4°C’de
15000 devir/dk.’da 15 dakika santrifüj edilmiştir. Oluşan üst faz tüplere alınarak üzerine
700 µL kloroform/izoamil alkol (24:1) çözeltisinden eklenmiş ve +4°C’de 15000
devir/dk.’da 7 dakika santrifüj edileerek üst faz tüplere alınmış ve üzerine eşit hacimde
soğuk etanol eklenmiştir. Bir gece -20°C’de bekletildikten sonra plazmid DNA 15000
devir/dk.’da 20 dakika santrifüj edilerek çöktürülmüştür. Üst faz dökülerek DNA peleti
havada kurutulmuştur. Plazmid DNA pelletleri elektroforez uygulamasından önce 20
µL Tris-EDTA-2 (10mM:1mM, pH:7.5) çözeltisi ile süspanse edilmiş ve 2 µL RNaz
stok çözeltisinden ilave edilerek 37 °C’lik su banyosunda 45 dakika inkübe edilmiştir
(Anderson ve McKay 1983, Dyall-Smith 2001).
35

3.2.8.1. Agaroz Jel elektroforezi
Plazmid DNA örneklerinin elektroforezi, yatay elektroforez tankında, %0.7’lik agaroz
jel ve Tris-Asetat tamponu kullanılarak yapılmıştır. RNaz’la 37 °C’de inkübe edilen
plazmid DNA örnekleri üzerine 2 µL bromofenol mavisi izleme boyası ilave edilerek
karıştırılmış ve mikropipet aracılığı ile 20 µL olacak şekilde kuyucuklara yüklenmiştir.
Elektroforez 120V’da sabit akımla 3.5-4 saat yürütülmüş işlem bittikten sonra jel, 0,2
µg/mL etidyum bromür içeren Tris-asetat tamponunda 1 saat bekletilmiştir. Boyama
işlemi sonrası jeller, Jel görüntüleme sistemi ile bilgisayar ortamında incelenerek
görüntüleri kaydedilmiştir (Meyers et al. 1976).
3.2.8.2. Plazmid büyüklüklerinin saptanması
Plazmidlerin büyüklüklerinin saptanmasında, 2.067-16.210 baz çifti arasında değişen 11
süpersarmal parça içerikli ccc DNA marker (Sigma D-5292) kullanılmıştır. Plazmidlerin
moleküler büyüklükleri hesaplanırken marker DNA’ların jel üzerindeki hareketi ile
büyüklüklerinin logaritmaları arasında oluşturulan doğrusal ilişkiden yararlanılmıştır.
Süpersarmal marker DNA moleküllerinin agaroz jel fotoğrafları üzerindeki göç
aralıkları ile bunlara ait büyüklüklerin logaritması alınarak eğriler oluşturulmuştur. Her
farklı jel fotoğrafı için istatistiki analizler kullanılarak eğrinin eğimi ile korelasyon
katsayısı hesaplanmış ve halofilik arke izolat ve standartlarından elde edilen plazmid
DNA’ların molekül ağırlıkları saptanmıştır (Macrina et al. 1978).
Moleküler büyüklük (W)= Antilog10[ I. (α-G)+H)]
X= Marker DNA moleküllerinin Agaroz jel üzerindeki göç aralığı (mm)
Y= Marker DNA moleküllerinin büyüklüğü (kb)
A= X1 + X2 + X3 +.......................................................................+ Xn
B= X12 + X2
2 + X32 +...................................................................+ Xn
2
C= log10Y1 + log10Y2 +log10Y3 +.............................................+log10Yn
D= (log10Y1)2 + (log10Y2)2 +(log10Y3)2 +.................................+(log10Yn)2
E= X1(log10Y1) +X2( log10Y2) +X3(log10Y3) +.........................+Xn(log10Yn)
36

G=Ortalama X=A/N
H=Ortalama Y=C/N
α= Moleküler büyüklüğü bilinmeyen plazmidin jel üzerinde göçü (mm)
Moleküler büyüklük(W)= ( )[ ]HGI +−α.
Eğrinin Eğimi (I)= ( )( )AGB
CGE..
−−
Korelasyon katsayısı(J)= ( )( )[ ] ( )[ ]AGBCHD
CGE...
.−−
−
37

UPGMA
Nei & Li's Katsayısı
Halobacter ium salinar um A263A A263BHaloar cula m ar ism or tuiHaloar cula vallism or tis DSMZ Halofer ax volcanii Natr ialba aisatica Halofer ax m editer r anei A317 A440Halococcus m or r huae CCM 5 E 133 A82 C13 C15 B19 B65A29F100F89F56A312E36C22B88C35D22F92F73E22F23F36D58AA75A182B71A87A407E20A206BA206AC37A405F42C36B54AA62AE57E49AB38BB38AB77AB77BB36A471E92BE92AA337B44AB44BD83BD83AD113A283A235A420B85A212A85F98E107D34E7D118AB53D50D91E55D25D107F84C23A43A347B74D6D19D74D108D31A137A181AA181BF4AF75AF5A138B49B45A191F30AA128
0,04 0,2 0,36 0,52 0,68 0,84 1
6
4
1
Şekil 4.34. Halofilik arke izolat ve standart suşlarının SDS-PAGE protein profillerine göre oluşturulan benzerlik dendogramı
2
3
5
7
8
− −−−−
−−
−−
−
−−
−
−
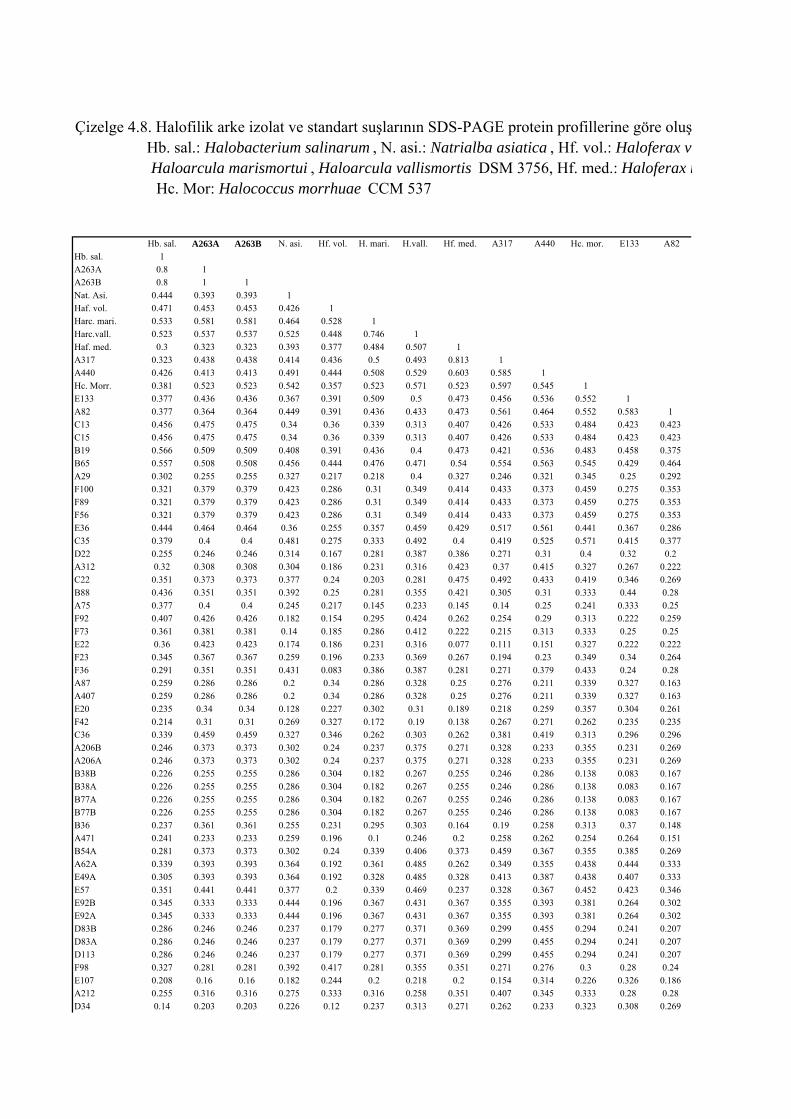
Çizelge 4.8. Halofilik arke izolat ve standart suşlarının SDS-PAGE protein profillerine göre olu Hb. sal.: Halobacterium salinarum , N. asi.: Natrialba asiatica , Hf. vol.: Haloferax v Haloarcula marismortui , Haloarcula vallismortis DSM 3756, Hf. med.: Haloferax Hc. Mor: Halococcus morrhuae CCM 537
Hb. sal. A263A A263B N. asi. Hf. vol. H. mari. H.vall. Hf. med. A317 A440 Hc. mor. E133Hb. sal. 1A263A 0.8 1A263B 0.8 1 1Nat. Asi. 0.444 0.393 0.393 1Haf. vol. 0.471 0.453 0.453 0.426 1Harc. mari. 0.533 0.581 0.581 0.464 0.528 1Harc.vall. 0.523 0.537 0.537 0.525 0.448 0.746 1Haf. med. 0.3 0.323 0.323 0.393 0.377 0.484 0.507 1A317 0.323 0.438 0.438 0.414 0.436 0.5 0.493 0.813 1A440 0.426 0.413 0.413 0.491 0.444 0.508 0.529 0.603 0.585 1Hc. Morr. 0.381 0.523 0.523 0.542 0.357 0.523 0.571 0.523 0.597 0.545 1E133 0.377 0.436 0.436 0.367 0.391 0.509 0.5 0.473 0.456 0.536 0.552 1A82 0.377 0.364 0.364 0.449 0.391 0.436 0.433 0.473 0.561 0.464 0.552 0.583C13 0.456 0.475 0.475 0.34 0.36 0.339 0.313 0.407 0.426 0.533 0.484 0.423C15 0.456 0.475 0.475 0.34 0.36 0.339 0.313 0.407 0.426 0.533 0.484 0.423B19 0.566 0.509 0.509 0.408 0.391 0.436 0.4 0.473 0.421 0.536 0.483 0.458B65 0.557 0.508 0.508 0.456 0.444 0.476 0.471 0.54 0.554 0.563 0.545 0.429A29 0.302 0.255 0.255 0.327 0.217 0.218 0.4 0.327 0.246 0.321 0.345 0.25F100 0.321 0.379 0.379 0.423 0.286 0.31 0.349 0.414 0.433 0.373 0.459 0.275F89 0.321 0.379 0.379 0.423 0.286 0.31 0.349 0.414 0.433 0.373 0.459 0.275F56 0.321 0.379 0.379 0.423 0.286 0.31 0.349 0.414 0.433 0.373 0.459 0.275E36 0.444 0.464 0.464 0.36 0.255 0.357 0.459 0.429 0.517 0.561 0.441 0.367C35 0.379 0.4 0.4 0.481 0.275 0.333 0.492 0.4 0.419 0.525 0.571 0.415D22 0.255 0.246 0.246 0.314 0.167 0.281 0.387 0.386 0.271 0.31 0.4 0.32A312 0.32 0.308 0.308 0.304 0.186 0.231 0.316 0.423 0.37 0.415 0.327 0.267C22 0.351 0.373 0.373 0.377 0.24 0.203 0.281 0.475 0.492 0.433 0.419 0.346B88 0.436 0.351 0.351 0.392 0.25 0.281 0.355 0.421 0.305 0.31 0.333 0.44A75 0.377 0.4 0.4 0.245 0.217 0.145 0.233 0.145 0.14 0.25 0.241 0.333F92 0.407 0.426 0.426 0.182 0.154 0.295 0.424 0.262 0.254 0.29 0.313 0.222F73 0.361 0.381 0.381 0.14 0.185 0.286 0.412 0.222 0.215 0.313 0.333 0.25E22 0.36 0.423 0.423 0.174 0.186 0.231 0.316 0.077 0.111 0.151 0.327 0.222F23 0.345 0.367 0.367 0.259 0.196 0.233 0.369 0.267 0.194 0.23 0.349 0.34F36 0.291 0.351 0.351 0.431 0.083 0.386 0.387 0.281 0.271 0.379 0.433 0.24A87 0.259 0.286 0.286 0.2 0.34 0.286 0.328 0.25 0.276 0.211 0.339 0.327A407 0.259 0.286 0.286 0.2 0.34 0.286 0.328 0.25 0.276 0.211 0.339 0.327E20 0.235 0.34 0.34 0.128 0.227 0.302 0.31 0.189 0.218 0.259 0.357 0.304F42 0.214 0.31 0.31 0.269 0.327 0.172 0.19 0.138 0.267 0.271 0.262 0.235C36 0.339 0.459 0.459 0.327 0.346 0.262 0.303 0.262 0.381 0.419 0.313 0.296A206B 0.246 0.373 0.373 0.302 0.24 0.237 0.375 0.271 0.328 0.233 0.355 0.231A206A 0.246 0.373 0.373 0.302 0.24 0.237 0.375 0.271 0.328 0.233 0.355 0.231B38B 0.226 0.255 0.255 0.286 0.304 0.182 0.267 0.255 0.246 0.286 0.138 0.083B38A 0.226 0.255 0.255 0.286 0.304 0.182 0.267 0.255 0.246 0.286 0.138 0.083B77A 0.226 0.255 0.255 0.286 0.304 0.182 0.267 0.255 0.246 0.286 0.138 0.083B77B 0.226 0.255 0.255 0.286 0.304 0.182 0.267 0.255 0.246 0.286 0.138 0.083B36 0.237 0.361 0.361 0.255 0.231 0.295 0.303 0.164 0.19 0.258 0.313 0.37A471 0.241 0.233 0.233 0.259 0.196 0.1 0.246 0.2 0.258 0.262 0.254 0.264B54A 0.281 0.373 0.373 0.302 0.24 0.339 0.406 0.373 0.459 0.367 0.355 0.385A62A 0.339 0.393 0.393 0.364 0.192 0.361 0.485 0.262 0.349 0.355 0.438 0.444E49A 0.305 0.393 0.393 0.364 0.192 0.328 0.485 0.328 0.413 0.387 0.438 0.407E57 0.351 0.441 0.441 0.377 0.2 0.339 0.469 0.237 0.328 0.367 0.452 0.423E92B 0.345 0.333 0.333 0.444 0.196 0.367 0.431 0.367 0.355 0.393 0.381 0.264E92A 0.345 0.333 0.333 0.444 0.196 0.367 0.431 0.367 0.355 0.393 0.381 0.264D83B 0.286 0.246 0.246 0.237 0.179 0.277 0.371 0.369 0.299 0.455 0.294 0.241D83A 0.286 0.246 0.246 0.237 0.179 0.277 0.371 0.369 0.299 0.455 0.294 0.241D113 0.286 0.246 0.246 0.237 0.179 0.277 0.371 0.369 0.299 0.455 0.294 0.241F98 0.327 0.281 0.281 0.392 0.417 0.281 0.355 0.351 0.271 0.276 0.3 0.28E107 0.208 0.16 0.16 0.182 0.244 0.2 0.218 0.2 0.154 0.314 0.226 0.326A212 0.255 0.316 0.316 0.275 0.333 0.316 0.258 0.351 0.407 0.345 0.333 0.28D34 0.14 0.203 0.203 0.226 0.12 0.237 0.313 0.271 0.262 0.233 0.323 0.308
ş
m
A82
10.4230.4230.3750.4640.2920.3530.3530.3530.2860.377
0.20.2220.2690.280.250.2590.250.2220.2640.280.1630.1630.2610.2350.2960.2690.2690.1670.1670.1670.1670.1480.1510.2690.3330.3330.3460.3020.3020.2070.2070.2070.240.1860.280.269

A85 0.314 0.264 0.264 0.213 0.273 0.189 0.31 0.264 0.255 0.37 0.214 0.13A283 0.508 0.525 0.525 0.327 0.308 0.361 0.455 0.295 0.286 0.355 0.344 0.37A235 0.508 0.525 0.525 0.327 0.308 0.361 0.455 0.295 0.286 0.355 0.344 0.37B44A 0.25 0.241 0.241 0.269 0.122 0.207 0.254 0.241 0.233 0.271 0.328 0.196B44B 0.25 0.241 0.241 0.269 0.122 0.207 0.254 0.241 0.233 0.271 0.328 0.196A420 0.345 0.333 0.333 0.444 0.471 0.433 0.462 0.367 0.387 0.426 0.349 0.34B85 0.345 0.333 0.333 0.444 0.471 0.433 0.462 0.367 0.387 0.426 0.349 0.34A137 0.321 0.241 0.241 0.269 0.286 0.31 0.381 0.31 0.267 0.339 0.295 0.392A181A 0.321 0.241 0.241 0.269 0.286 0.31 0.381 0.31 0.267 0.339 0.295 0.392A181B 0.321 0.241 0.241 0.269 0.286 0.31 0.381 0.31 0.267 0.339 0.295 0.392F4A 0.321 0.241 0.241 0.269 0.286 0.31 0.381 0.31 0.267 0.339 0.295 0.392F75A 0.321 0.241 0.241 0.269 0.286 0.31 0.381 0.31 0.267 0.339 0.295 0.392B45 0.231 0.222 0.222 0.292 0.311 0.222 0.203 0.222 0.25 0.218 0.351 0.298D58A 0.464 0.379 0.379 0.423 0.204 0.276 0.317 0.276 0.267 0.271 0.262 0.235F5 0.259 0.357 0.357 0.2 0.17 0.357 0.262 0.214 0.31 0.281 0.441 0.408A138 0.421 0.441 0.441 0.302 0.32 0.339 0.406 0.271 0.328 0.3 0.452 0.423B49 0.379 0.4 0.4 0.407 0.392 0.333 0.4 0.3 0.355 0.295 0.508 0.453D50 0.2 0.192 0.192 0.261 0.233 0.231 0.281 0.269 0.222 0.453 0.109 0.311E7 0.113 0.182 0.182 0.163 0.217 0.255 0.333 0.327 0.316 0.286 0.276 0.208D118A 0.37 0.357 0.357 0.24 0.255 0.321 0.361 0.214 0.207 0.351 0.305 0.204B53 0.32 0.308 0.308 0.087 0.14 0.192 0.175 0.192 0.222 0.264 0.255 0.133D19 0.28 0.308 0.308 0.261 0.326 0.385 0.246 0.192 0.296 0.302 0.218 0.311D74 0.28 0.308 0.308 0.261 0.326 0.385 0.246 0.192 0.296 0.302 0.218 0.311D108 0.28 0.308 0.308 0.261 0.326 0.385 0.246 0.192 0.296 0.302 0.218 0.311D31 0.255 0.281 0.281 0.275 0.375 0.316 0.29 0.281 0.203 0.379 0.267 0.32E55 0.302 0.291 0.291 0.163 0.217 0.327 0.367 0.291 0.246 0.321 0.31 0.25A43 0.29 0.281 0.281 0.172 0.291 0.313 0.232 0.344 0.273 0.4 0.239 0.211A347 0.286 0.338 0.338 0.237 0.321 0.369 0.286 0.338 0.328 0.424 0.294 0.31A182 0.31 0.3 0.3 0.148 0.275 0.267 0.308 0.2 0.258 0.393 0.19 0.302B71 0.321 0.31 0.31 0.192 0.163 0.241 0.349 0.241 0.2 0.169 0.328 0.235C37 0.327 0.421 0.421 0.431 0.333 0.421 0.452 0.386 0.339 0.379 0.4 0.24A405 0.327 0.351 0.351 0.314 0.25 0.281 0.355 0.351 0.373 0.31 0.433 0.32A337 0.305 0.295 0.295 0.364 0.192 0.23 0.273 0.295 0.286 0.29 0.375 0.185D25 0.154 0.222 0.222 0.125 0.222 0.222 0.203 0.222 0.214 0.291 0.281 0.298D107 0.321 0.379 0.379 0.269 0.163 0.276 0.317 0.241 0.233 0.305 0.295 0.314F84 0.259 0.286 0.286 0.28 0.213 0.214 0.23 0.25 0.172 0.211 0.237 0.082C23 0.276 0.3 0.3 0.222 0.196 0.267 0.277 0.3 0.258 0.295 0.222 0.302D91 0.308 0.259 0.259 0.333 0.311 0.37 0.373 0.407 0.357 0.436 0.281 0.383F30A 0.246 0.305 0.305 0.264 0.24 0.339 0.281 0.305 0.262 0.233 0.258 0.231D6 0.169 0.23 0.23 0.218 0.423 0.295 0.303 0.361 0.349 0.323 0.375 0.333A191 0.231 0.222 0.222 0.292 0.4 0.37 0.339 0.296 0.393 0.4 0.246 0.383A128 0.211 0.203 0.203 0.264 0.24 0.237 0.281 0.271 0.164 0.3 0.226 0.269
B74 0.2 0.29 0.29 0.25 0.264 0.452 0.358 0.419 0.406 0.413 0.338 0.4
Hb. sal. A263A A263B N. asi. Hf. vol. H. mari. H.vall. Hf. med. A317 A440 Hc. mor. E133
0.1740.2960.2960.2350.2350.3020.3020.4310.4310.4310.4310.4310.2550.2750.3270.3460.4150.1780.2080.1630.2220.2220.2220.2220.240.2080.2110.2070.2260.1570.280.320.2590.170.2350.2040.2640.340.1920.2220.3830.231
0.255
A82

şturulan benzerlik matriksivolcanii , H. mari:mediterranei ,
C13 C15 B19 B65 A29 F100 F89 F56 E36 C35 D22 A312 C22
11 1
0.654 0.654 10.667 0.667 0.857 10.308 0.308 0.333 0.357 10.364 0.364 0.471 0.475 0.627 10.364 0.364 0.471 0.475 0.627 1 10.364 0.364 0.471 0.475 0.627 1 1 10.453 0.453 0.531 0.526 0.408 0.654 0.654 0.654 10.386 0.386 0.377 0.393 0.453 0.5 0.5 0.5 0.556 10.37 0.37 0.32 0.345 0.52 0.566 0.566 0.566 0.431 0.655 10.408 0.408 0.444 0.415 0.4 0.667 0.667 0.667 0.652 0.4 0.511 10.321 0.321 0.346 0.333 0.385 0.618 0.618 0.618 0.566 0.491 0.407 0.531 10.37 0.37 0.4 0.379 0.48 0.604 0.604 0.604 0.392 0.4 0.5 0.468 0.630.346 0.346 0.375 0.357 0.417 0.51 0.51 0.51 0.408 0.264 0.36 0.444 0.4230.172 0.172 0.259 0.258 0.407 0.316 0.316 0.316 0.291 0.373 0.357 0.353 0.414
0.2 0.2 0.179 0.219 0.357 0.339 0.339 0.339 0.316 0.393 0.448 0.415 0.40.245 0.245 0.267 0.264 0.311 0.375 0.375 0.375 0.261 0.36 0.511 0.381 0.3270.281 0.281 0.302 0.295 0.302 0.393 0.393 0.393 0.333 0.414 0.545 0.44 0.5260.296 0.296 0.32 0.345 0.4 0.528 0.528 0.528 0.431 0.473 0.5 0.468 0.4070.302 0.302 0.327 0.281 0.204 0.269 0.269 0.269 0.32 0.296 0.275 0.13 0.3020.302 0.302 0.327 0.281 0.204 0.269 0.269 0.269 0.32 0.296 0.275 0.13 0.3020.24 0.24 0.348 0.296 0.217 0.286 0.286 0.286 0.426 0.314 0.25 0.233 0.240.291 0.291 0.275 0.339 0.314 0.296 0.296 0.296 0.308 0.286 0.189 0.125 0.2180.379 0.379 0.37 0.355 0.185 0.351 0.351 0.351 0.436 0.271 0.143 0.275 0.310.393 0.393 0.385 0.367 0.308 0.4 0.4 0.4 0.34 0.281 0.259 0.286 0.2860.393 0.393 0.385 0.367 0.308 0.4 0.4 0.4 0.34 0.281 0.259 0.286 0.2860.154 0.154 0.333 0.286 0.375 0.392 0.392 0.392 0.327 0.264 0.2 0.222 0.3460.154 0.154 0.333 0.286 0.375 0.392 0.392 0.392 0.327 0.264 0.2 0.222 0.3460.154 0.154 0.333 0.286 0.375 0.392 0.392 0.392 0.327 0.264 0.2 0.222 0.3460.154 0.154 0.333 0.286 0.375 0.392 0.392 0.392 0.327 0.264 0.2 0.222 0.3460.241 0.241 0.296 0.258 0.333 0.316 0.316 0.316 0.218 0.271 0.286 0.118 0.2410.351 0.351 0.264 0.262 0.189 0.214 0.214 0.214 0.333 0.31 0.182 0.24 0.3510.25 0.25 0.269 0.3 0.231 0.436 0.436 0.436 0.453 0.386 0.296 0.327 0.4640.31 0.31 0.296 0.323 0.37 0.421 0.421 0.421 0.473 0.407 0.321 0.275 0.4480.276 0.276 0.259 0.29 0.407 0.421 0.421 0.421 0.436 0.441 0.321 0.275 0.4830.321 0.321 0.308 0.333 0.385 0.436 0.436 0.436 0.453 0.386 0.259 0.286 0.4290.211 0.211 0.264 0.328 0.377 0.321 0.321 0.321 0.296 0.379 0.291 0.2 0.3860.211 0.211 0.264 0.328 0.377 0.321 0.321 0.321 0.296 0.379 0.291 0.2 0.3860.355 0.355 0.276 0.364 0.276 0.328 0.328 0.328 0.441 0.317 0.367 0.327 0.3230.355 0.355 0.276 0.364 0.276 0.328 0.328 0.328 0.441 0.317 0.367 0.327 0.3230.355 0.355 0.276 0.364 0.276 0.328 0.328 0.328 0.441 0.317 0.367 0.327 0.3230.222 0.222 0.28 0.31 0.28 0.302 0.302 0.302 0.235 0.255 0.192 0.255 0.4070.383 0.383 0.279 0.275 0.233 0.217 0.217 0.217 0.136 0.25 0.178 0.1 0.2130.444 0.444 0.32 0.379 0.24 0.302 0.302 0.302 0.235 0.218 0.192 0.17 0.370.214 0.214 0.231 0.3 0.308 0.255 0.255 0.255 0.075 0.316 0.222 0.163 0.321
B88 A75
10.52 10.321 0.370.31 0.4290.213 0.3560.473 0.4150.308 0.280.314 0.2450.314 0.2450.167 0.3480.151 0.3140.214 0.4810.259 0.3460.259 0.346
0.2 0.2920.2 0.2920.2 0.2920.2 0.292
0.286 0.3330.291 0.3020.37 0.3460.393 0.4070.357 0.370.37 0.4620.364 0.3020.364 0.3020.233 0.310.233 0.310.233 0.310.346 0.320.311 0.2330.346 0.280.37 0.385

0.32 0.32 0.261 0.222 0.391 0.327 0.327 0.327 0.298 0.235 0.208 0.233 0.520.379 0.379 0.37 0.484 0.37 0.351 0.351 0.351 0.364 0.339 0.25 0.275 0.310.379 0.379 0.37 0.484 0.37 0.351 0.351 0.351 0.364 0.339 0.25 0.275 0.310.291 0.291 0.314 0.407 0.314 0.407 0.407 0.407 0.385 0.25 0.264 0.333 0.40.291 0.291 0.314 0.407 0.314 0.407 0.407 0.407 0.385 0.25 0.264 0.333 0.40.421 0.421 0.34 0.426 0.302 0.321 0.321 0.321 0.407 0.345 0.327 0.32 0.3160.421 0.421 0.34 0.426 0.302 0.321 0.321 0.321 0.407 0.345 0.327 0.32 0.3160.327 0.327 0.392 0.339 0.431 0.296 0.296 0.296 0.346 0.393 0.264 0.25 0.1820.327 0.327 0.392 0.339 0.431 0.296 0.296 0.296 0.346 0.393 0.264 0.25 0.1820.327 0.327 0.392 0.339 0.431 0.296 0.296 0.296 0.346 0.393 0.264 0.25 0.1820.327 0.327 0.392 0.339 0.431 0.296 0.296 0.296 0.346 0.393 0.264 0.25 0.1820.327 0.327 0.392 0.339 0.431 0.296 0.296 0.296 0.346 0.393 0.264 0.25 0.1820.196 0.196 0.255 0.255 0.213 0.2 0.2 0.2 0.083 0.308 0.204 0.136 0.1570.255 0.255 0.314 0.305 0.353 0.333 0.333 0.333 0.308 0.393 0.377 0.375 0.40.377 0.377 0.327 0.281 0.163 0.231 0.231 0.231 0.16 0.222 0.118 0.174 0.2260.25 0.25 0.346 0.3 0.192 0.182 0.182 0.182 0.264 0.351 0.222 0.163 0.2140.316 0.316 0.377 0.393 0.226 0.214 0.214 0.214 0.185 0.345 0.255 0.16 0.2110.367 0.367 0.222 0.264 0.133 0.208 0.208 0.208 0.304 0.24 0.085 0.238 0.2860.269 0.269 0.292 0.321 0.167 0.235 0.235 0.235 0.327 0.34 0.2 0.178 0.3080.415 0.415 0.408 0.386 0.204 0.231 0.231 0.231 0.28 0.296 0.196 0.217 0.2640.367 0.367 0.311 0.302 0.089 0.125 0.125 0.125 0.174 0.16 0.128 0.19 0.2040.327 0.327 0.311 0.34 0.089 0.292 0.292 0.292 0.304 0.12 0.128 0.238 0.3270.327 0.327 0.311 0.34 0.089 0.292 0.292 0.292 0.304 0.12 0.128 0.238 0.3270.327 0.327 0.311 0.34 0.089 0.292 0.292 0.292 0.304 0.12 0.128 0.238 0.3270.259 0.259 0.2 0.241 0.2 0.302 0.302 0.302 0.235 0.255 0.231 0.298 0.2960.269 0.269 0.25 0.25 0.208 0.235 0.235 0.235 0.204 0.264 0.36 0.311 0.2690.459 0.459 0.421 0.4 0.281 0.3 0.3 0.3 0.241 0.226 0.237 0.37 0.2620.419 0.419 0.379 0.364 0.207 0.295 0.295 0.295 0.339 0.286 0.2 0.327 0.3230.351 0.351 0.264 0.295 0.264 0.286 0.286 0.286 0.333 0.241 0.109 0.24 0.2460.291 0.291 0.275 0.271 0.353 0.37 0.37 0.37 0.308 0.286 0.302 0.292 0.3640.407 0.407 0.44 0.448 0.28 0.415 0.415 0.415 0.314 0.291 0.308 0.383 0.2590.333 0.333 0.28 0.31 0.4 0.34 0.34 0.34 0.275 0.4 0.269 0.255 0.370.31 0.31 0.259 0.258 0.407 0.386 0.386 0.386 0.291 0.407 0.321 0.275 0.4830.353 0.353 0.17 0.182 0.085 0.16 0.16 0.16 0.167 0.269 0.204 0.273 0.2350.291 0.291 0.314 0.339 0.157 0.296 0.296 0.296 0.346 0.321 0.151 0.25 0.2910.264 0.264 0.204 0.246 0.204 0.269 0.269 0.269 0.12 0.259 0.235 0.217 0.2640.281 0.281 0.226 0.262 0.151 0.25 0.25 0.25 0.185 0.207 0.291 0.2 0.2810.235 0.235 0.298 0.291 0.17 0.32 0.32 0.32 0.375 0.269 0.163 0.364 0.3140.179 0.179 0.308 0.433 0.231 0.291 0.291 0.291 0.264 0.175 0.259 0.245 0.1790.241 0.241 0.222 0.355 0.333 0.281 0.281 0.281 0.182 0.441 0.393 0.196 0.2410.196 0.196 0.255 0.327 0.213 0.2 0.2 0.2 0.333 0.308 0.204 0.227 0.1960.179 0.179 0.231 0.233 0.269 0.255 0.255 0.255 0.34 0.246 0.333 0.204 0.25
0.237 0.237 0.327 0.349 0.255 0.276 0.276 0.276 0.357 0.333 0.281 0.269 0.271
C13 C15 B19 B65 A29 F100 F89 F56 E36 C35 D22 A312 C22
0.333 0.3480.321 0.370.321 0.370.302 0.3140.302 0.3140.218 0.3770.218 0.3770.34 0.2750.34 0.2750.34 0.2750.34 0.2750.34 0.2750.286 0.2130.491 0.3530.235 0.1630.296 0.1920.291 0.2260.34 0.3560.16 0.2080.235 0.2860.128 0.2670.255 0.2670.255 0.2670.255 0.2670.385 0.40.24 0.250.271 0.2110.233 0.2410.255 0.4910.453 0.4310.308 0.320.269 0.280.429 0.2960.286 0.2550.302 0.3140.275 0.2450.364 0.2640.286 0.2130.222 0.4230.179 0.2220.122 0.2130.259 0.423
0.211 0.218
B88 A75

F92 F73 E22 F23 F36 A87 A407 E20 F42 C36 A206B A206A
10.839 10.431 0.566 10.475 0.557 0.68 10.429 0.517 0.511 0.473 10.218 0.211 0.174 0.333 0.118 10.218 0.211 0.174 0.333 0.118 1 10.192 0.185 0.186 0.275 0.25 0.596 0.596 10.175 0.169 0.167 0.179 0.113 0.385 0.385 0.449 10.333 0.323 0.235 0.237 0.25 0.4 0.4 0.5 0.632 10.276 0.233 0.245 0.316 0.222 0.491 0.491 0.56 0.4 0.552 10.276 0.233 0.245 0.316 0.222 0.491 0.491 0.56 0.4 0.552 1 10.222 0.107 0.222 0.226 0.2 0.327 0.327 0.435 0.471 0.444 0.5 0.50.222 0.107 0.222 0.226 0.2 0.327 0.327 0.435 0.471 0.444 0.5 0.50.222 0.107 0.222 0.226 0.2 0.327 0.327 0.435 0.471 0.444 0.5 0.50.222 0.107 0.222 0.226 0.2 0.327 0.327 0.435 0.471 0.444 0.5 0.50.167 0.226 0.353 0.339 0.214 0.436 0.436 0.385 0.456 0.467 0.414 0.4140.136 0.164 0.2 0.345 0.145 0.481 0.481 0.353 0.321 0.407 0.456 0.4560.241 0.233 0.245 0.351 0.296 0.34 0.34 0.36 0.4 0.552 0.429 0.4290.333 0.29 0.392 0.339 0.429 0.364 0.364 0.462 0.386 0.533 0.448 0.4480.333 0.29 0.353 0.339 0.429 0.291 0.291 0.462 0.316 0.533 0.483 0.4830.31 0.267 0.367 0.351 0.407 0.302 0.302 0.48 0.436 0.552 0.429 0.4290.339 0.295 0.24 0.31 0.436 0.222 0.222 0.314 0.286 0.339 0.281 0.2810.339 0.295 0.24 0.31 0.436 0.222 0.222 0.314 0.286 0.339 0.281 0.2810.375 0.424 0.255 0.349 0.433 0.203 0.203 0.357 0.295 0.375 0.387 0.3870.375 0.424 0.255 0.349 0.433 0.203 0.203 0.357 0.295 0.375 0.387 0.3870.375 0.424 0.255 0.349 0.433 0.203 0.203 0.357 0.295 0.375 0.387 0.3870.214 0.138 0.128 0.291 0.192 0.353 0.353 0.333 0.302 0.286 0.407 0.4070.122 0.196 0.1 0.208 0.178 0.409 0.409 0.195 0.261 0.245 0.255 0.2550.143 0.103 0.085 0.218 0.269 0.314 0.314 0.333 0.302 0.357 0.37 0.370.345 0.267 0.204 0.351 0.333 0.264 0.264 0.16 0.109 0.138 0.321 0.321
B38B B38A B77A
11 11 1 11 1 1
0.556 0.556 0.5560.491 0.491 0.4910.385 0.385 0.3850.407 0.407 0.4070.407 0.407 0.4070.385 0.385 0.3850.377 0.377 0.3770.377 0.377 0.3770.31 0.31 0.310.31 0.31 0.310.31 0.31 0.310.32 0.32 0.320.14 0.14 0.140.24 0.24 0.240.115 0.115 0.115

0.423 0.333 0.14 0.353 0.208 0.255 0.255 0.227 0.245 0.308 0.28 0.280.4 0.323 0.353 0.305 0.393 0.327 0.327 0.385 0.246 0.367 0.448 0.4480.4 0.323 0.353 0.305 0.393 0.327 0.327 0.385 0.246 0.367 0.448 0.448
0.351 0.339 0.25 0.286 0.415 0.269 0.269 0.327 0.333 0.316 0.327 0.3270.351 0.339 0.25 0.286 0.415 0.269 0.269 0.327 0.333 0.316 0.327 0.3270.271 0.23 0.2 0.276 0.327 0.333 0.333 0.353 0.321 0.339 0.316 0.3160.271 0.23 0.2 0.276 0.327 0.333 0.333 0.353 0.321 0.339 0.316 0.3160.281 0.271 0.208 0.214 0.226 0.269 0.269 0.286 0.296 0.351 0.291 0.2910.281 0.271 0.208 0.214 0.226 0.269 0.269 0.286 0.296 0.351 0.291 0.2910.281 0.271 0.208 0.214 0.226 0.269 0.269 0.286 0.296 0.351 0.291 0.2910.281 0.271 0.208 0.214 0.226 0.269 0.269 0.286 0.296 0.351 0.291 0.2910.281 0.271 0.208 0.214 0.226 0.269 0.269 0.286 0.296 0.351 0.291 0.2910.189 0.145 0.136 0.154 0.163 0.375 0.375 0.222 0.36 0.189 0.196 0.1960.351 0.305 0.292 0.429 0.264 0.231 0.231 0.122 0.222 0.211 0.145 0.1450.327 0.246 0.13 0.185 0.235 0.28 0.28 0.17 0.231 0.291 0.377 0.3770.31 0.267 0.245 0.211 0.185 0.34 0.34 0.32 0.218 0.379 0.357 0.3570.305 0.262 0.2 0.207 0.145 0.333 0.333 0.275 0.286 0.373 0.386 0.3860.157 0.226 0 0.24 0.17 0.261 0.261 0.186 0.208 0.392 0.327 0.3270.222 0.25 0.222 0.302 0.28 0.204 0.204 0.261 0.157 0.296 0.308 0.3080.327 0.351 0.217 0.296 0.275 0.36 0.36 0.34 0.154 0.291 0.377 0.3770.314 0.34 0.19 0.28 0.213 0.217 0.217 0.14 0.167 0.314 0.163 0.1630.157 0.189 0.095 0.24 0.213 0.304 0.304 0.326 0.417 0.392 0.286 0.2860.157 0.189 0.095 0.24 0.213 0.304 0.304 0.326 0.417 0.392 0.286 0.2860.157 0.189 0.095 0.24 0.213 0.304 0.304 0.326 0.417 0.392 0.286 0.2860.179 0.207 0.085 0.327 0.231 0.314 0.314 0.375 0.34 0.321 0.222 0.2220.296 0.357 0.356 0.415 0.4 0.286 0.286 0.348 0.157 0.259 0.346 0.3460.286 0.308 0.259 0.323 0.305 0.276 0.276 0.182 0.3 0.286 0.328 0.3280.281 0.333 0.218 0.222 0.333 0.373 0.373 0.321 0.328 0.469 0.387 0.3870.237 0.295 0.2 0.138 0.255 0.333 0.333 0.431 0.357 0.508 0.351 0.3510.316 0.305 0.208 0.393 0.226 0.423 0.423 0.367 0.185 0.281 0.4 0.40.214 0.207 0.34 0.4 0.385 0.275 0.275 0.417 0.226 0.393 0.556 0.5560.321 0.276 0.298 0.4 0.423 0.392 0.392 0.417 0.189 0.393 0.481 0.481
0.4 0.355 0.314 0.373 0.429 0.255 0.255 0.192 0.211 0.3 0.276 0.2760.264 0.291 0.091 0.308 0.245 0.375 0.375 0.4 0.12 0.189 0.314 0.3140.246 0.305 0.125 0.214 0.34 0.385 0.385 0.449 0.296 0.386 0.436 0.4360.218 0.246 0.261 0.444 0.353 0.28 0.28 0.298 0.192 0.255 0.302 0.3020.271 0.295 0.12 0.379 0.364 0.333 0.333 0.392 0.321 0.373 0.386 0.3860.151 0.109 0.091 0.269 0.286 0.375 0.375 0.4 0.16 0.34 0.353 0.3530.241 0.233 0.122 0.211 0.259 0.264 0.264 0.32 0.364 0.345 0.25 0.250.267 0.355 0.275 0.271 0.25 0.255 0.255 0.231 0.421 0.267 0.241 0.2410.226 0.255 0.182 0.192 0.286 0.292 0.292 0.222 0.24 0.302 0.235 0.2350.276 0.333 0.245 0.351 0.296 0.226 0.226 0.28 0.255 0.31 0.179 0.179
0.295 0.317 0.154 0.2 0.316 0.286 0.286 0.302 0.241 0.295 0.305 0.305
F92 F73 E22 F23 F36 A87 A407 E20 F42 C36 A206B A206A
0.348 0.348 0.3480.37 0.37 0.370.37 0.37 0.370.392 0.392 0.3920.392 0.392 0.3920.34 0.34 0.340.34 0.34 0.340.353 0.353 0.3530.353 0.353 0.3530.353 0.353 0.3530.353 0.353 0.3530.353 0.353 0.3530.17 0.17 0.170.353 0.353 0.3530.204 0.204 0.2040.269 0.269 0.2690.264 0.264 0.2640.222 0.222 0.2220.333 0.333 0.3330.204 0.204 0.2040.089 0.089 0.0890.178 0.178 0.1780.178 0.178 0.1780.178 0.178 0.1780.16 0.16 0.160.25 0.25 0.250.316 0.316 0.3160.241 0.241 0.2410.34 0.34 0.340.353 0.353 0.3530.36 0.36 0.360.28 0.28 0.280.37 0.37 0.370.213 0.213 0.2130.196 0.196 0.1960.327 0.327 0.3270.264 0.264 0.2640.383 0.383 0.3830.346 0.346 0.3460.333 0.333 0.3330.213 0.213 0.2130.346 0.346 0.346
0.291 0.291 0.291
B38B B38A B77A

B77B B36 A471 B54A A62A E49A E57 E92B E92A D83B D83A
10.556 10.491 0.644 10.385 0.517 0.561 10.407 0.5 0.441 0.69 10.407 0.433 0.407 0.655 0.9 10.385 0.483 0.386 0.679 0.931 0.862 10.377 0.407 0.345 0.456 0.441 0.441 0.456 10.377 0.407 0.345 0.456 0.441 0.441 0.456 1 10.31 0.281 0.317 0.323 0.406 0.406 0.419 0.476 0.476 10.31 0.281 0.317 0.323 0.406 0.406 0.419 0.476 0.476 1 10.31 0.281 0.317 0.323 0.406 0.406 0.419 0.476 0.476 1 10.32 0.214 0.291 0.296 0.25 0.25 0.222 0.4 0.4 0.3 0.30.14 0.286 0.25 0.298 0.286 0.204 0.298 0.25 0.25 0.34 0.340.24 0.179 0.218 0.222 0.25 0.357 0.259 0.327 0.327 0.333 0.3330.115 0.207 0.246 0.286 0.345 0.379 0.357 0.316 0.316 0.258 0.258
D113 F98 E107 A212
10.3 1
0.34 0.444 10.333 0.385 0.4 10.258 0.296 0.298 0.407

0.348 0.192 0.196 0.2 0.308 0.385 0.36 0.392 0.392 0.429 0.4290.37 0.333 0.271 0.345 0.5 0.467 0.483 0.475 0.475 0.438 0.4380.37 0.333 0.271 0.345 0.5 0.467 0.483 0.475 0.475 0.438 0.4380.392 0.246 0.286 0.327 0.351 0.351 0.364 0.464 0.464 0.426 0.4260.392 0.246 0.286 0.327 0.351 0.351 0.364 0.464 0.464 0.426 0.4260.34 0.169 0.207 0.246 0.305 0.305 0.316 0.379 0.379 0.476 0.4760.34 0.169 0.207 0.246 0.305 0.305 0.316 0.379 0.379 0.476 0.4760.353 0.351 0.321 0.255 0.351 0.316 0.327 0.357 0.357 0.197 0.1970.353 0.351 0.321 0.255 0.351 0.316 0.327 0.357 0.357 0.197 0.1970.353 0.351 0.321 0.255 0.351 0.316 0.327 0.357 0.357 0.197 0.1970.353 0.351 0.321 0.255 0.351 0.316 0.327 0.357 0.357 0.197 0.1970.353 0.351 0.321 0.255 0.351 0.316 0.327 0.357 0.357 0.197 0.1970.17 0.34 0.269 0.275 0.264 0.189 0.275 0.269 0.269 0.14 0.140.353 0.246 0.357 0.255 0.281 0.246 0.255 0.321 0.321 0.197 0.1970.204 0.364 0.333 0.302 0.218 0.218 0.226 0.296 0.296 0.136 0.1360.269 0.414 0.421 0.321 0.379 0.379 0.321 0.281 0.281 0.194 0.1940.264 0.407 0.379 0.316 0.339 0.339 0.281 0.241 0.241 0.159 0.1590.222 0.314 0.44 0.408 0.275 0.275 0.286 0.16 0.16 0.327 0.3270.333 0.222 0.377 0.385 0.296 0.333 0.269 0.34 0.34 0.31 0.310.204 0.218 0.259 0.226 0.291 0.327 0.34 0.222 0.222 0.373 0.3730.089 0.196 0.28 0.163 0.157 0.157 0.204 0.24 0.24 0.327 0.3270.178 0.235 0.2 0.408 0.314 0.275 0.327 0.24 0.24 0.218 0.2180.178 0.235 0.2 0.408 0.314 0.275 0.327 0.24 0.24 0.218 0.2180.178 0.235 0.2 0.408 0.314 0.275 0.327 0.24 0.24 0.218 0.2180.16 0.214 0.109 0.296 0.321 0.321 0.37 0.327 0.327 0.367 0.3670.25 0.333 0.34 0.308 0.333 0.296 0.308 0.264 0.264 0.379 0.3790.316 0.286 0.323 0.23 0.127 0.063 0.098 0.226 0.226 0.299 0.2990.241 0.313 0.317 0.258 0.25 0.188 0.194 0.222 0.222 0.294 0.2940.34 0.373 0.379 0.421 0.407 0.407 0.421 0.276 0.276 0.286 0.2860.353 0.456 0.464 0.4 0.421 0.386 0.364 0.214 0.214 0.164 0.1640.36 0.321 0.291 0.296 0.357 0.393 0.37 0.218 0.218 0.267 0.2670.28 0.357 0.436 0.407 0.429 0.5 0.37 0.364 0.364 0.267 0.2670.37 0.4 0.475 0.414 0.367 0.333 0.31 0.576 0.576 0.25 0.250.213 0.302 0.385 0.196 0.113 0.113 0.157 0.231 0.231 0.316 0.3160.196 0.351 0.357 0.364 0.316 0.316 0.364 0.357 0.357 0.393 0.3930.327 0.364 0.407 0.34 0.218 0.218 0.264 0.37 0.37 0.203 0.2030.264 0.441 0.345 0.386 0.305 0.237 0.316 0.414 0.414 0.381 0.3810.383 0.264 0.385 0.431 0.34 0.377 0.314 0.346 0.346 0.316 0.3160.346 0.31 0.246 0.321 0.31 0.31 0.357 0.316 0.316 0.258 0.2580.333 0.3 0.305 0.379 0.333 0.333 0.31 0.203 0.203 0.281 0.2810.213 0.264 0.308 0.314 0.302 0.302 0.275 0.269 0.269 0.211 0.2110.346 0.241 0.246 0.179 0.276 0.276 0.286 0.14 0.14 0.258 0.258
0.291 0.361 0.3 0.305 0.328 0.361 0.271 0.167 0.167 0.338 0.338
B77B B36 A471 B54A A62A E49A E57 E92B E92A D83B D83A
0.429 0.292 0.341 0.4580.438 0.429 0.327 0.3570.438 0.429 0.327 0.3570.426 0.34 0.304 0.340.426 0.34 0.304 0.340.476 0.436 0.292 0.4730.476 0.436 0.292 0.4730.197 0.189 0.13 0.1890.197 0.189 0.13 0.1890.197 0.189 0.13 0.1890.197 0.189 0.13 0.1890.197 0.189 0.13 0.1890.14 0.286 0.238 0.1630.197 0.151 0.13 0.2260.136 0.157 0.227 0.2750.194 0.222 0.128 0.1850.159 0.291 0.208 0.2180.327 0.213 0.4 0.2980.31 0.32 0.233 0.280.373 0.196 0.409 0.3140.327 0.17 0.35 0.2130.218 0.34 0.25 0.340.218 0.34 0.25 0.340.218 0.34 0.25 0.340.367 0.462 0.267 0.3080.379 0.28 0.186 0.240.299 0.305 0.385 0.3390.294 0.333 0.377 0.3670.286 0.291 0.375 0.3640.164 0.34 0.304 0.3020.267 0.308 0.178 0.4230.267 0.423 0.267 0.4230.25 0.429 0.286 0.2860.316 0.327 0.238 0.2860.393 0.302 0.348 0.340.203 0.275 0.273 0.2750.381 0.291 0.333 0.2910.316 0.367 0.143 0.2860.258 0.185 0.085 0.2590.281 0.286 0.286 0.1790.211 0.327 0.238 0.2860.258 0.111 0.085 0.148
0.338 0.175 0.2 0.211
D113 F98 E107 A212

D34 A85 A283 A235 B44A B44B A420 B85 A137 A181A
1
A181B F4A F75A B45 D58A

0.28 10.379 0.269 10.379 0.269 1 10.327 0.286 0.561 0.561 10.327 0.286 0.561 0.561 1 10.316 0.431 0.576 0.576 0.357 0.357 10.316 0.431 0.576 0.576 0.357 0.357 1 10.218 0.204 0.281 0.281 0.259 0.259 0.214 0.214 10.218 0.204 0.281 0.281 0.259 0.259 0.214 0.214 1 10.218 0.204 0.281 0.281 0.259 0.259 0.214 0.214 1 10.218 0.204 0.281 0.281 0.259 0.259 0.214 0.214 1 1 1 10.218 0.204 0.281 0.281 0.259 0.259 0.214 0.214 1 1 1 1 10.275 0.133 0.075 0.075 0.04 0.04 0.192 0.192 0.36 0.360.218 0.245 0.246 0.246 0.222 0.222 0.321 0.321 0.333 0.3330.302 0.213 0.182 0.182 0.269 0.269 0.111 0.111 0.385 0.3850.214 0.12 0.276 0.276 0.145 0.145 0.175 0.175 0.545 0.5450.246 0.118 0.271 0.271 0.143 0.143 0.207 0.207 0.5 0.50.327 0.279 0.196 0.196 0.208 0.208 0.24 0.24 0.292 0.2920.346 0.217 0.222 0.222 0.353 0.353 0.264 0.264 0.314 0.3140.302 0.383 0.291 0.291 0.154 0.154 0.37 0.37 0.192 0.1920.286 0.326 0.196 0.196 0.208 0.208 0.32 0.32 0.167 0.1670.163 0.186 0.275 0.275 0.333 0.333 0.36 0.36 0.167 0.1670.163 0.186 0.275 0.275 0.333 0.333 0.36 0.36 0.167 0.1670.163 0.186 0.275 0.275 0.333 0.333 0.36 0.36 0.167 0.1670.333 0.25 0.214 0.214 0.151 0.151 0.327 0.327 0.189 0.1890.269 0.13 0.37 0.37 0.353 0.353 0.264 0.264 0.235 0.2350.262 0.291 0.286 0.286 0.367 0.367 0.355 0.355 0.333 0.3330.226 0.286 0.281 0.281 0.295 0.295 0.286 0.286 0.361 0.3610.316 0.196 0.373 0.373 0.321 0.321 0.31 0.31 0.286 0.2860.364 0.286 0.351 0.351 0.37 0.37 0.25 0.25 0.222 0.2220.296 0.125 0.357 0.357 0.264 0.264 0.327 0.327 0.189 0.1890.333 0.25 0.393 0.393 0.226 0.226 0.255 0.255 0.264 0.2640.31 0.308 0.333 0.333 0.491 0.491 0.237 0.237 0.351 0.3510.275 0.222 0.264 0.264 0.24 0.24 0.269 0.269 0.24 0.240.364 0.204 0.386 0.386 0.37 0.37 0.107 0.107 0.296 0.2960.34 0.255 0.291 0.291 0.423 0.423 0.111 0.111 0.231 0.2310.281 0.275 0.305 0.305 0.321 0.321 0.138 0.138 0.214 0.2140.196 0.267 0.226 0.226 0.12 0.12 0.269 0.269 0.4 0.40.25 0.16 0.31 0.31 0.4 0.4 0.316 0.316 0.291 0.2910.207 0.154 0.2 0.2 0.281 0.281 0.305 0.305 0.281 0.2810.196 0.222 0.302 0.302 0.2 0.2 0.385 0.385 0.36 0.360.179 0.2 0.172 0.172 0.218 0.218 0.246 0.246 0.364 0.364
0.271 0.113 0.197 0.197 0.207 0.207 0.267 0.267 0.345 0.345
D34 A85 A283 A235 B44A B44B A420 B85 A137 A181A
1
0.36 0.36 0.36 10.333 0.333 0.333 0.28 10.385 0.385 0.385 0.333 0.2310.545 0.545 0.545 0.392 0.327
0.5 0.5 0.5 0.385 0.2860.292 0.292 0.292 0.182 0.1250.314 0.314 0.314 0.128 0.1180.192 0.192 0.192 0.292 0.2310.167 0.167 0.167 0.182 0.2080.167 0.167 0.167 0.182 0.1670.167 0.167 0.167 0.182 0.1670.167 0.167 0.167 0.182 0.1670.189 0.189 0.189 0.408 0.2260.235 0.235 0.235 0.17 0.2750.333 0.333 0.333 0.321 0.2670.361 0.361 0.361 0.281 0.230.286 0.286 0.286 0.192 0.1790.222 0.222 0.222 0.24 0.2960.189 0.189 0.189 0.245 0.3020.264 0.264 0.264 0.245 0.2640.351 0.351 0.351 0.189 0.4210.24 0.24 0.24 0.391 0.120.296 0.296 0.296 0.32 0.0740.231 0.231 0.231 0.292 0.2690.214 0.214 0.214 0.269 0.25
0.4 0.4 0.4 0.348 0.20.291 0.291 0.291 0.275 0.2550.281 0.281 0.281 0.34 0.2460.36 0.36 0.36 0.348 0.160.364 0.364 0.364 0.196 0.291
0.345 0.345 0.345 0.37 0.207
A181B F4A F75A B45 D58A

F5 A138 B49 D50 E7 D118A B53 D19 D74 D108 D31 E55 A43 A347 A182
10.528 10.556 0.877 10.261 0.204 0.24 10.163 0.154 0.151 0.356 10.28 0.34 0.37 0.478 0.408 10.217 0.327 0.28 0.333 0.356 0.565 10.217 0.204 0.24 0.333 0.311 0.348 0.286 10.217 0.204 0.24 0.333 0.311 0.348 0.286 1 10.217 0.204 0.24 0.333 0.311 0.348 0.286 1 10.118 0.222 0.255 0.298 0.2 0.314 0.213 0.511 0.5110.204 0.346 0.302 0.222 0.292 0.245 0.267 0.311 0.3110.345 0.328 0.323 0.407 0.386 0.379 0.444 0.407 0.4070.339 0.387 0.381 0.436 0.379 0.373 0.4 0.4 0.40.222 0.211 0.207 0.44 0.302 0.333 0.16 0.4 0.40.385 0.4 0.393 0.292 0.196 0.269 0.167 0.208 0.2080.275 0.37 0.4 0.383 0.28 0.471 0.213 0.255 0.2550.275 0.37 0.364 0.34 0.36 0.314 0.17 0.34 0.340.364 0.31 0.305 0.235 0.37 0.145 0.235 0.275 0.2750.333 0.314 0.308 0.409 0.255 0.333 0.227 0.318 0.3180.346 0.291 0.286 0.417 0.235 0.385 0.167 0.25 0.250.24 0.226 0.222 0.304 0.327 0.32 0.261 0.174 0.1740.333 0.316 0.31 0.32 0.151 0.259 0.28 0.36 0.360.292 0.392 0.385 0.455 0.298 0.417 0.182 0.318 0.3180.151 0.214 0.246 0.163 0.308 0.226 0.245 0.327 0.3270.145 0.241 0.305 0.235 0.333 0.145 0.157 0.275 0.2750.25 0.314 0.308 0.273 0.213 0.208 0.182 0.182 0.1820.113 0.286 0.281 0.327 0.231 0.302 0.204 0.163 0.163
0.25 0.305 0.367 0.346 0.327 0.321 0.192 0.308 0.308
F5 A138 B49 D50 E7 D118A B53 D19 D74
10.511 10.311 0.32 10.407 0.237 0.456 1
0.4 0.267 0.414 0.776 10.4 0.4 0.453 0.355 0.444 1
0.208 0.264 0.392 0.367 0.262 0.4290.255 0.346 0.44 0.407 0.4 0.3270.34 0.231 0.4 0.373 0.4 0.4360.275 0.214 0.407 0.444 0.406 0.4070.318 0.408 0.426 0.429 0.351 0.3850.25 0.302 0.275 0.3 0.426 0.50.174 0.314 0.408 0.448 0.407 0.3330.36 0.436 0.566 0.355 0.413 0.4140.318 0.408 0.255 0.321 0.351 0.3850.327 0.333 0.346 0.328 0.29 0.3160.275 0.286 0.259 0.381 0.375 0.3050.182 0.245 0.34 0.25 0.281 0.3460.163 0.333 0.269 0.164 0.258 0.316
0.308 0.316 0.364 0.406 0.492 0.467
D108 D31 E55 A43 A347 A182

B71 C37 A405 A337 D25 D107 F84 C23 D91 F30A D6 A191 A128 B74
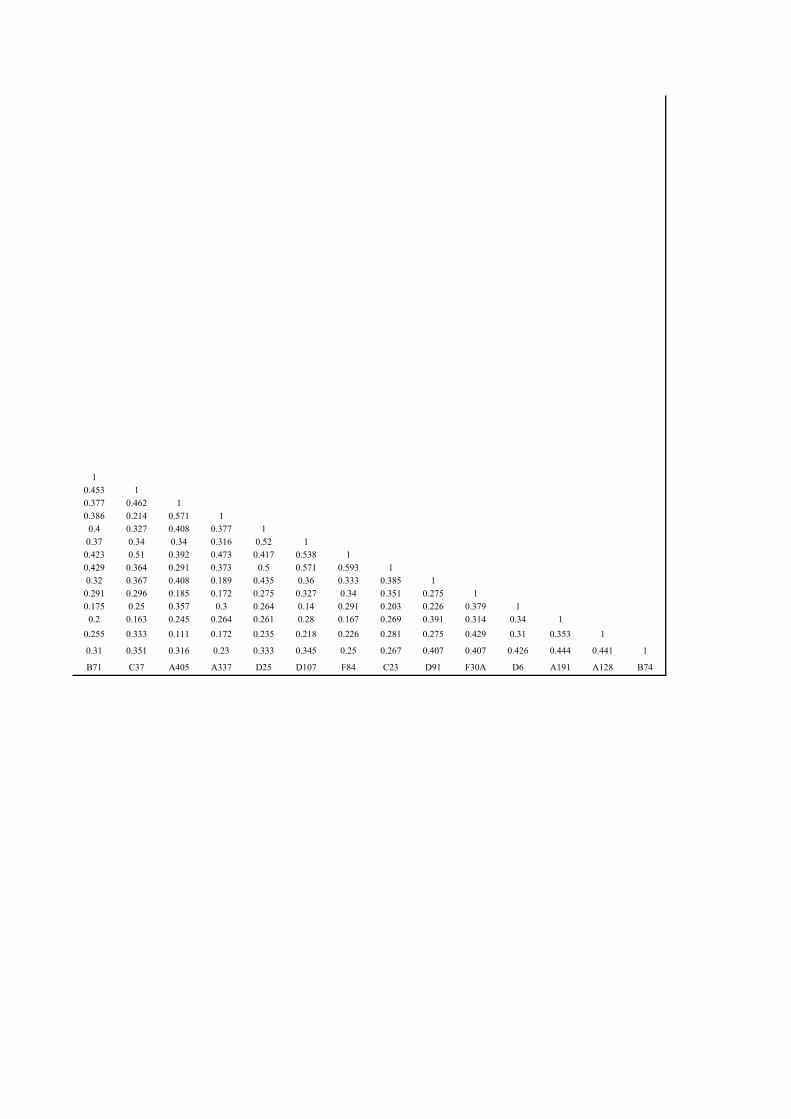
10.453 10.377 0.462 10.386 0.214 0.571 1
0.4 0.327 0.408 0.377 10.37 0.34 0.34 0.316 0.52 10.423 0.51 0.392 0.473 0.417 0.538 10.429 0.364 0.291 0.373 0.5 0.571 0.593 10.32 0.367 0.408 0.189 0.435 0.36 0.333 0.3850.291 0.296 0.185 0.172 0.275 0.327 0.34 0.3510.175 0.25 0.357 0.3 0.264 0.14 0.291 0.203
0.2 0.163 0.245 0.264 0.261 0.28 0.167 0.2690.255 0.333 0.111 0.172 0.235 0.218 0.226 0.281
0.31 0.351 0.316 0.23 0.333 0.345 0.25 0.267
B71 C37 A405 A337 D25 D107 F84 C23
10.275 10.226 0.379 10.391 0.314 0.34 10.275 0.429 0.31 0.353 1
0.407 0.407 0.426 0.444 0.441 1
D91 F30A D6 A191 A128 B74

KAYNAKLAR
Akhmanova, A.S., Kagramanova, V. K. and Mankin, A.S. 1993. Heterogeneity of small
plasmids from halophilic archaea. J. of Bacteriol., 175 (4); 1081-1086.
Anderson, D. G. and MacKay, L. L. 1983. Simple and rapid method for isolating large
plasmid DNA from Lactic streptococci. Appl. And Environ. Microbiol., 46(3);
549-552.
Anonim. 1999. Uluslararası Önemi Olan Sulak Alanların Biyolojik ve Ekolojik Yönden
Araştırılması Projesi, Alt Proje III, Sonuç Raporu. Proje yürütücüsü Prof. Dr. Ali
Demirsoy, T.C. Çevre Bakanlığı, Çevre Koruma Genel Müd., Ankara.
Arahal, D. R., Dewhirst, F. E., Paster, B. J., Volcani, B. E. and Ventosa, A. 1996.
Phylogenetic analyses of some extremely halophilic archaea isolated from Dead
Sea water, determined on the basis of their 16 S rRNA sequences. Appl. And
Environ. Mic., 62(10); 3779-3786).
Arda, M. 1997. Temel Mikrobiyoloji. Medisan yayınevi, Ankara, ISBN:975-7774-22-7.
Asker, D. and Ohta, Y. 2002. Haloferax alexandrinus sp. nov., an exteremely halophilic
cathaxanthin-producing archaeon from a solar saltern in Alexandria (Egypt). Int.
J. Syst. and Evolut. Microbiol, 52; 729-738.
Bansal, A. K. and Meyer, T. E. 2002. Evolutionar analysis by whole-genome
comparisions. J. bacteriol., 184(8); 2260-2272.
Beard, S. J., Hayes, P. K. and Walsby, A. E. 1997. Growth competetion between
Halobacterium salinarum strain PHH1 and mutants affected in gas vesicle
synthesis. Microniology UK, 143; 467-473.
Bertrand, J. C., Almallah, M., Acquaviva, M. and Mille, G. 1990. Biodegredation of
hydrocarbons by an exremely halophilic archaebacterium. Letters in Applied
Microbiolgy, 11; 260-263.
Bonelo, G., Ventosa, A.i Megias, M. and Ruiz-Berraquero, F. 1984. The sensitivity of
halobacteria to antibiotics. FEMS Microbiol. Letters, 21; 341-345.
Böck, A. and Kandler, O. 1985. Antibiotic sensitivity of archaeabacteria In: C.R. Woese
and R. S. Wolfe (Eds), The Bacteria-a treatise on structure and function.
Archaebacteria, Academic Press Orlando, FLVol.III, 525-541.
105

Braithwaite, C. J. R. and Zedef, V. 1994. Living hydromagnesite stromatolites from
Turkey. Sedimentary Geology, 92; 1-5.
Brenner, D. J., Staley, J. T. and Krieg, N. R. 2001. Classification of prokaryotic
organisms and the concept of bacterial speciation In: Bergey’s Manual of
Systematic Bacteriology V.I, The Archaea and Deeply Branching and
Phototrophic Bacteria, Garrity, G. M. (Man. Ed.), 2nd Ed. New York, Springer,
ISBN: 0387987711.
Brown, H. J. and Gibbons, N. E. 1995. The effect of magnesium, potassium and iron on
the growth and morphology of red halophilic bacteria. Can J.Microbiol., 1; 486-
494.
Canik, B. 1988. Bozdağ-Yapalı Toprakkale dolayındaki (Cihanbeyli) sıcak ve mineralli
sular ve oluşukları. Ulusal I. Hidrojeoloji Sempozyumu, Ankara Üniv. Basımevi.
Chaga, G, Porath, J. and Illéni, T. 1993. Isolation and purification of amyloglucosidase
from Halobacterium sodomonse. Biomedical chromatography, 7; 256-261.
Cohen, S., Oren, A. and Shilo, M. 1983. The divalent cation requirement of Dead Sea
halobacteria. Arch. Microbiol., 136; 184-190.
Danson, M. and Hough, D. W. 1997. The structural basis of protein halophilicity.
Comp. Biochem. Physiol., 117A (3); 307-312.
DasSarma, S. 1993. Identification and analysis of the gas vesicle gene cluster on an
unstable plasmid of Halobacterium halobium. Experientia, 49; 482-486.
DasSarma, S. and Arora, P. 1997. Genetic analysis of the gas vesicle gene cluster in
haloarchaea. FEMS mic. Letters, 153; 1-10.
Dussault, H. P. 1955. Am improved technique for staining red halophilic bacteria. J. of
Bacteriol., 70, 484-485.
Dyall-Smith, M. 2001. The Halohandbook: Protocols for halobacterial genetics
(4.5 version, electronic publication).
Emerson, D., Chauhan, S., Oriel, P. and Breznak, J. A. 1994. Haloferax sp. D1227, a
halophilic archaeon capable of growth on aromatic compounds. Arch.
Microbiol., 161; 445-452.
Esen, A. 1978. A simple method for quantitative, semi-quantitative and qualitative
assay of protein. Anal. Biochem., 89; 264-273.
106

Eyüboğlu, Ö. 1995. Seyfe Gölü (Kırşehir) Tabiatı koruma alanının florası. Doktora tezi,
Gazi Üniv., Fen Bilimleri Ens., Ankara.
Franzmann, P.D., Stackebrandt, E., Sanderson, K., Volkman, J. K., Cameron, D. E.,
Stevenson, P. L., McMeekin, T.A. and Burton, H. R. 1988. Halobacterium
lacusprofundi sp. nov., halophilic bacterium isolated from Deep Lake,
Antarctica. System. Appl. Microbiol., 11; 20-27.
Gonzales, C., Gutierrez, C. and Ramirez, C. 1978. Halobacterium vallismortis sp. nov.,
an amylolytic and carbohydrate-metabolizing extremely halophilic bacterium.
Can. J. Microbiol., 24; 710-715.
Goo, Y. A., Yi, E. C., Baliga, N. S., Tao, W. A., Pan, M., Aebersold, R., Goodlett, D.
R., Hood, L. and Ng, W. V. 2003. Proteomic analysis of an extreme halophilic
Archaeon, Halobacterium sp. NRC-1. Molecular and Cellular Proteomics2; 506-
524.
Grant, W. D, Gemmell, R.T. and McGenity, T. J. 1998. Halobacteria: the evidence for
longevity. Extremophiles, 2; 279-287.
Grant, W. D., Kamekura, M., McGenity, T. J. and Ventosa, A. 2001. Order I.
Halobacteriales In: Bergey’s Manual of Systematic Bacteriology V.I, The
Archaea and Deeply Branching and Phototrophic Bacteria, Garrity, G. M. (Man.
Ed.), 2nd Ed. New York, Springer, ISBN: 0387987711.
Gutierrez, C. and Gonzalez, C. 1972. Method for simultaneous detection of proteinase
and esterase activities in extremely halophilic bacteria. Appl. Microbiol., 24;
516-517.
Gutierrez, M. C., Garcia, M. T., Ventosa, A., Nieto, J. J. and Ruiz-Berraquero, F. 1986.
Occurence of megaplasmids in halobacteria. J. of Applied Bacteriology, 61; 67-
71.
Gutierrez, M. C., Ventosa, A. and Berraquero, F. 1989. DNA-DNA homology studies
among strains of Haloferax and other Halobacteria. Curr. Mic., 18; 253-256.
Gutierrez, M. C., ventosa, A. and Ruiz-Berrquero, F. 1990. deoxyribonucleic acid
relatidness among species of Haloarcula and other Halobacteria. Biochem. Cell.
Biol., 68;106-110.
107

Gutierrez, M. C., Kamekura, M., Holmes, M. L., Dyall-Smith, M. L. and Ventosa, A.
2002. Taxonomic characterization of Haloferax sp. ("H. alicantei") strain Aa 2.2:
description of Haloferax lucentensis sp. nov. Extremophiles: 6; 479-483.
Hackett, R.N. and Grant, W. D. 1989. Characterization of the small endogenous plasmid
of Halobacterium strain SB3 and its use in transformation of H. halobium. Can.
J. Microbiol., 35; 86-91.
Hamana, K., Hamana, H. and Itoh, T. 1995. Ubiqutious occurence of agmatine as the
major polyamine within extremely halophilic archaebacteria. J. Gen. Appl.
Microbiol., 41; 153-158.
Hancock, A. J. and Kates, M. 1973. Structure determination of the phophatidygycero-
phosfate (dieter analog) from Halobacterium cutirubrum. J. of lipid research, 14;
422-429.
Hartmann, R., Sickinger, H., D. and Oesterhelt, D. 1980. Anaerobic growth of
halobacteria. Proc. Natl. Acad. Sci. USA, 77 (6); 3821-3825.
Hesselberg, M. and Vreeland, R. H. 1995. Utilization of protein profiles for the
characterization of halophilic bacteria. Current Microbiolgy, 31; 158-162.
Hezayen, F. F., Rehm, B. H. A., Eberhardt, R. and Steinbüchel, A. 2000. Polymer
production by two newly isolatedremely halophilic archaea: application of a
novel corrosion-resistant bioreactor. Appl. Microbiol. Biotechnol., 54; 319-325.
Hezayen, F. F., Rehm, B. H. A., Tindall, B. J. and Steinbüchel, A. 2001. Transfer of
Natrialba asiatica B1T to Natrialba taiwanwnsis sp. nov. and description of
Natrialba aegyptiaca sp. nov., a novel extremely halophilic, aerobic, non-
pigmented member of the Archaea from Egypt that produces extracellular
poly(glutamic acid). Int. J. Syst. and Evolut. Microbiol, 51; 1133-1142.
Hezayen, F. F., Tindall, B. J., Steinbüchel and Rehm, B. H. A. 2002. Characterization of
a novel halophilic archaeon, Halobioforma haloterrestris gen. nov., sp. nov., and
transfer of Natronobacterium nitratireducens to Halobioforma nitratireducens
comb. nov. Int. J. Syst. and Evolut. Microbiol, 52; 2271-2280.
Hilpert, R., Winter, J., Hammes, W. and Kandler, . 1981. The sensitivity of
archaebacteria to antibiotics. Zbl. Bakt. Hyg. I. Abt. Orig. C., 2, 11-20.
Holding, A. J. and Collee, J. G. 1971. Routine biochemical tests. Methods Microbiol.,
6A; 11-20.
108

Holmes, M. L. and Dyall-Smith, M. L. 1991. Mutations in DNA gyrase result in
novobiocin resistance in halophilic archaebacteria. J. of. Bacteriol., 173 (2); 642-
648.
Horikoshi, K., Aono, R. and Nakamura, S. 1993. The triangular halophilic
archaebacterium Haloarcula japonica strain TR-1. Experientia, 49; 497-502.
Hough, D. W. And Danson, D. J. 1999. Extremozymes. Curr. Opin. İn Chem. Biology,
3; 39-46.
Hough, D. W. and Danson, M. J. 1989. Archaebacteria: ancient organisms with
commercial potential. Letters in Applied Microbiolgy, 9; 33-39.
Ihara, K., Watanabe, S. and Tamura, T. 1997. Haloarcula argentinensis sp. nov. and
Haloarcula mukohatei sp. nov., two extremely halophilic archaea collected in
Argentina. Int. J. Syst. Bacteriol., 47 (1); 73-77.
Izotova, L. S., Strongin, A. Y., Chekulaeva, L. N., Sterkin, V. E., Ostoslavskaya, V. I.,
Lyublinskaya, L. A., Timokhina E. A. and Stepanov, V. M. 1983. Purification
and properties of serine protease from Halobacterium halobium. J. of Bact.,
155(2); 826-830.
Jaenicke, R. and Zavodszky. P. 1990. Proteins under extreme physical conditions.
FEBS, 268 (2); 344-349.
Jocklik, W. K., Wilett, H. P., Amos, D. B. and Wilfert C. M. 1992. Zinsser
Microbiology. 20th Ed., Appleton and Lange Publisher, ISBN: 0838599834.
Johnsen, U., Selig, M., Xavier, K. B., Santos, H. and Schönheit, P. 2001. Different
glycolytic pathways for glucose and fructose in the halophilic archaeon
Halococcus saccharolyticus. Arch. Microbiol., 175; 52-61.
Jones, . G., Ewing, C. M. and Melvin, M. V. 1981. Biotechnology of solar saltfield.
Hidrobiologia, 82; 391-406.
Kamekura, M., Bardocz, S., Anderson, P., Wallace, R. and Kushner, D. J. 1986.
Polyamines in moderately and extremely halophilic bacteria. Biochim. Biophys.
Acta., 880, 204-208.
Kamekura, M. and Kates, M. 1988. Lipids of halophilic archaebacteria In. F.
Rodriguez-Valera (Ed.) Halophilic Bacteria Vol. II CRC Press Boca Raton,
Florida, 25-54.
109

Kamekura, M. and Seno, Y. 1993. Partial sequence of the gene for aserine protease
from a halophilic archaeum Haloferax mediterranei R4 and nucleotid sequences
of 16S rRNA encoding genes from several halophilic archaea. Experientia, 49;
503-513.
Kamekura, M. and Dyall-Smith, M. L. 1995. Taxonomy of the family halobacteriaceae
and description of two new genera Halorubrobacterium and Natrialba. 1995. J.
gen. Appl. Microbiol., 41; 333-350.
Kamekura, M. Dyall-Smith, M. L., Upasani, V., Ventosa, A. and Kates, M. 1997.
Diversty of alkaliphilic halobacteria: Proposals for transfer of Natronobacterium
vacuolatum, Natronobacterium magadii, and Natronobacterium pharaonis to
Halorubrum, Natrialba and Natronomonas gen. nov., respectively as
Halorubrum vacuolatum comb. Nov., Natrialba magadii comb. Nov., and
Natronomonas pharaonis comb. Nov., Respectively. I. J. Syst. Bact., 47(3); 853-
857.
Kamekura, M. 1998. Diversty of extremely halophilic bacteria. Extremophiles, 2; 289-
295.
Kamekura, M. 1999. Alkaliphilic Microorganisms: J. Seckbah (ed.), Enigmatic
Microorganisms and life in extrem environments, 479-485, Kluwer Academic
Publishers, Netherland.
Kamekura, M. and Kates, M. 1999. Structural diversty of membrane lipids in members
of Halobacteriaceae. Biosci. Biotechnol. Biochem., 63 (6); 969-972.
Kamekura, M., Oesterhelt, D., Wallace, R., Anderson, P. and Kushner, D. J. 1988. Lysis
of halobacteria in Bacto-peptone by bile acids. App. And Environ. Mic., 54(4);
990-995.
Kates, M. 1992. archaebacterial lipids: structure, biosynthesis and function In: The
Archaebacteria: Biochemistry and Biotechnology, Danson M. J., Hough, D. W.
and Lunt, G. G. (Ed.), The Biochemical Society, Portland Press, London, ISBN:
1-85578-010-0.
Kates, M. 1993. Membrane lipids of extreme halophiles: biosynthesis, function and
evolutionary significans. İn Halophilic Bacteria, Part II. Experientia, 49; 1027-
1036. Edited by R. H. Vreeland. Basel: Birkhäuser Verlag.
110

Kates, M. 1996. Structural analysis of phospholipids and glycolipids in extremely
halophilic archaebacteria. J. of Microbiol. Methods, 25; 113-128.
Kates, M. and Deroo, P. W. 1973. Structure determination of the glycolipid sulfate from
the extreme halophile Halobacterium cutirubrum. J. of lipid research, 14; 438-
445.
Kennedy, S. P., Ng, W. V., Salzberg, S. L., Hood, L. and DasSarma, S. 2001.
Understanding the adaptation of Halobacterium species NRC-1 to its extreme
environment through compututional analysis of its genome sequence. Genome
research, 11; 1641-4650.
Kibar, M. 1999. Güncel Tuzla Gölünün Sedimentalojik İncelemesi (Sarıoğlan,
Kayseri), Doktora tezi, Jeoloji Müh. A.B.D., Ankara Üniv., Ankara.
Kis-Papo, T. and Oren, A. 2000. Halocins: are they involved in the competetion
between halobacteria in saltern ponds?. Extremophiles, 4, 35-41.
Kostrikina, N. A., Zvyagintseva, I. S. and Duda, V. I. 1991. Cytological pecularities of
some extremely halophilic soil archaeobacteria. Arch. Microbiol., 156; 344-349.
Krieg, N.R. 2001. Prokaryotik Domains In: Bergey’s Manual of Systematic
Bacteriology V.I, The Archaea and Deeply Branching and Phototrophic Bacteria,
Garrity, G. M. (Man. Ed.), 2nd Ed. New York, Springer, ISBN: 0387987711.
Kulichevskaya, I. S., Zvyagintseva, I. S., Tarasov, A. L. and Plakunov, V. K. 1992. The
extremely halophilic archaebacteria from some hypersaline ecotops.
Microbiology, 46; 51-56.
Kunte, H. J., Trüper, G. and Stan-Lotter, H. 2001 Halophilic microorganisms.
Astrobiology: The quest for the conditions of life. Gerda Horneck, Christa
Baumstark-Khan (Eds), Heidelberg: Springer-virlag, ISBN: 3-540-42101-7
Kushawa, S. C. and Kates, M. 1979. Effect of glycerol on carotenogenesis in the extrem
halophile, Halobacterium cutirubrum. Can. J. Microbiol., 25, 1288-1372.
Kushner, D. J. 1985. The Halobacteriaceae. In: C.R. Woese and R. S. Wolfe (Eds), The
Bacteria-a treatise on structure and function. Archaebacteria, Academic Press
Orlando, FLVol.III, 171-214.
Kushner, D. J. 1988. What is the “true” internal environment of halophilic and other
bacteria? Can. J. Microbiol., 34; 482-486.
111

Laemmli, U. K. 1970. Clevage of structural proteins during the assembly of the head of
bacteriophage T4. Nature, 227; 680-685.
Lillo, J. G. and Rodriguez-Valera, F. 1990. Effects of culture conditions on poly (β-
hydroxybutiric acid) production by Haloferax mediterranei. Appl. And Environ.
Microbiol., 56 (8); 2517-2521.
Litchfield, C. D. 2002. Halophiles. J. İndustrial microbiol. And biotechnol., 28; 21-22.
Litchfield, C. D., Irby, A., Kis-papo, T. and Oren, A. 2000. Comparisions of the polar
lipd and pigment profiles of two solar salterns located in Newark, California,
U.S.A., and Eliat, Israel. Extremophiles, 4, 259-265.
Lizama, C., Monteoliva-sanchez, M., Suarez-Garcia, A., Rosello-Mora, R., Aguilera,
M., Campos, V. and Ramos-Cormenzana, A. 2002. Halorubrum tebenquichense
sp. nov., a novel halophilic archaeon isolated from the Atacam saltern, Chile. Int.
J. Syst. And Evol. Mic., 52; 149-155.
Lopez-Garcia, P., Jean, A. S., Amilas, R. and Charlebois. R. L. 1995. Genomic stability
in the archaea Haloferax volcanii and Haloferax mediterranei. J. bacteriol.,
177(5); 1405-1408.
M.T.A. derleme rapor. 1955. Denizli Acı göl mevkiinin jeolojisi. Rapor No:2509.
Macrina, F.l., Kobcko, D. J. Jones, K. R., Ayers, D. S. and McCoven, S. M. 1978. A
multiple plasmid containing Escherichia coli strain: convenient source of size
reference plasmid molecules. Plasmid, 1; 417-420.
Madigan, M. T., Martinko, M. J. and Parker, J. 2000. Biology of Microorganisms,
Prentice Hall International, Inc., USA, 9. Edt., ISBN:0-13-085264-3.
Mancinelli, R. L. and Hochstein L. I. 1986. The occurrence of denitrification in
extremely halophilic bacteria. FEMS Microbiol. Letters, 35; 55-58.
Margesin, H. and Schinner, F. 2001. Potential of halotolerant and halophilic
microorganisms for biotechnology. Extremophiles, 5; 73-83.
Martinez-Murcia, A. J. and Rodriguez-Valera, F. 1994. Random amplified polymorfic
DNA of a group of halophilic archaeal isolates. Syst. Appl. Microbiol., 17, 395-
401.
Martinez-Murcia, A. J., Acinas, S. G. and Rodriguez-Valera, F. 1995. Evaluation of
prokaryotic diversty by restrictase digestion of 16S rDNA directly amplified
from hypersaline environmets. FEMS Mic. Ecology, 17; 247-256.
112

Matheson, A T. 1985. Ribosomes of archaebacteria In: C.R. Woese and R. S. Wolfe
(Eds), The Bacteria-a treatise on structure and function. Archaebacteria,
Academic Press Orlando, FLVol.III, 345-372.
McGenity, T. and Grant, W. D. 1995. Transfer of Halobacterium Saccharovorum,
Halobacterium sodomonse, Halobacterium trapanicum NRC 34021 and
Halobacterium lacusprofundi to the genus Halorubrum gen. nov., as
Halorubrum saccharovorum comb. nov., Halorubrum sodomonse comb. nov.,
Halorubrum trapanicum comb. nov., and Halorubrum lacusprofundi comb. nov.
McGenity, T. J., Gemmell, R. and Grant, W. D. 1998. Proposal of a new halobacterial
genus Natrinema gen. nov. with two species Natrinema gen. nov., with two
species Natrinema Pellirubrum nom. Nov. and Natrinema pallidium. Nom. Nov.
Int. J. Syst. Bact., 48; 1187-1196.
Meseguer, I., Rodriguez-Valera, F. and Ventosa, A. 1986. Antagonistic interactions
among halobacteria due to halocine production. FEMS Microbiol. Letters, 36;
177-182.
Messner, P., Allmaier, G., Schäffer, C., wugeditsch, T., Lotal, S., König, H., Niemetz, R
and Dorner, M. 1997. III. Biochemistry of S-layers. FEMS Mic. Reviews, 20;
25-46.
Mevarech, M., Frolow, F. and Gloss, L. M. 2000. Halophilic enzymes: proteins with a
grain of salt. Biophysical chem., 86; 155-164.
Meyers, D. A., Sanches, D., Elwell, L. P. and Falkow, S. 1976. Simple agaros gel
electrophoretic method for the identification and the characterization plasmid
deoxyribonucleic acid. J. Bacteriol., 127(3); 1529-1537.
Montalvo-Rodriguez, R., Vreeland, R. H., Oren, A., Kessel, M., Betancourt, C. and
Lopez-Gariga, J. 1998. Halogeometricum borinquense gen. nov., sp. nov., a
novel halophilic archaeon from Puerto Rico. İnt. J. of Syst. Bacteriol., 48; 1305-
1312.
Montalvo-Rodriguez, R., Lopez-Garriga, J., Vreeland, H., Oren, A., Ventosa, A. and
Kamekura, M. 2000. Haloterrigena thermotolerans sp. nov., a halophilic
archaeon from Puerto Rico. İnt. J. Syst. and Evolut. Microbiol., 50; 1065-1071.
113

Montero, G. C., Ventosa, A., Nieto, J. J. and Ruiz-Berraquero, F. 1988. Isolation and
partial characterization of a plasmid in the extremely halophilic archaebacterium
Halococcus morrhuae CCM 537. J. of General Microbiol., 134; 2959-2963.
Morita, M., Yamaguchi, N., Eguchi, T. and Kakinuma, K. 1998. Structural diversty of
the membrane core lipids extreme halophiles. Biosci. Biotechnol. Biochem., 62;
596-598.
Müller, V. and Oren, A. 2003. Metabolism of chloride in halophilic prokaryotes.
Extremophiles, 7; 261-266
Nei, M. and Li, W. 1979. Mathematical model for studying genetic variation in terms of
restriction endonucleases. Proc.Natl.Acad.Sci.(USA), 76; 5269-5273.
Ng. W. L., Kothakota, S. and DasSarma, S. 1991. structure of the gas vesicle plasmid in
Halobacterium halobium: Inversion isomers, inverted repeats and insertion
squences. J. Bact., 173(6); 1958-1964.
Ng. W. V., Ciufo, S. A., Smith, T. M., Bumgarner, R. E., Baskin, D., Faust, J., Hall, B.,
Loretz, C,. Seto, J., Slagel, J., Hood, l. and DasSarma, S. 1998. Snapshot of a
large Dynamic replicon in a halophilic archaeon: Megaplasmid or
minichromosome?. Genome research, 8, 1131-1141.
Ng. W. V., Kennedy, S. P., ahairas, G. G., Berqu,st, B., Pan, M., Shukla, H. D., Lasky,
S. R., Baliga, N. S., Thorsson, V., Sbrogna, J., Swartzell, S., Weir, D., Hall, J.,
Dahl, T. A., Welti, R., Goo, Y. A., Leithauser, B., Keller, K., Cruz, R,. Danson,
M. J., Hough, D. W., Maddocks, D. G., Jablonski, P. E., Krebs, M. P., Angevine,
C. M., Dale, H., Isanbarger, T. A., Peck, R. F., Pohlschroder, M., Spudich, J. L.,
Jung, K. H., Alam, M., Freitas, T., Hou, S., Danielsi C. J., Dennis, P. P., Omer,
A. D., Ebhardt, H., Lowe, T. ., Liang, P., Riley, M., Hood, L. and DasSarma, S.
2000. Genome sequence of Halobacterium species NRC-1. Proc. Natl. Acad. Sci.
USA., 97(22); 12176-12181.
Nıemetz, R., Kärcher, U., Knadler, O., Tindall, B. J. and König, H. 1997. The cell wall
polymer of the extremely halophilic archaeon Natronococcus occultus. Eur. J.
Biochem., 249; 905-911.
Oesterhelt, D. 1998. The structure and mechanism of the family of retinal proteins from
halophilic archaea. Current opinion in structural biology, 8; 489-500.
114

Oren, A. 1983. Halobacterium sodomonse sp. nov., a Dead sea halobacterium with an
exteremely high magnesium requirement. Int. J. Syst. Bacteriol., 33; 381-386.
Oren, A., Lau, P. P. and Fox, G. E. 1988. The taxonomic status of Halobacterium
marismortui from the Dead sea: a comparision with Halobacterium vallismortis.
Syst. Appl. Microbiol., 10; 251-258.
Oren, A. 1990. Starch counteracts the inhibitory action of bacto peptone and bile salts in
media for the growth of halobacteria. Can. J. Microbiol., 36; 299-301.
Oren, A. and Trüper, H. G. 1990. Anaerobic growth of halophilic archaeobacteria by
reduction of dimethylsulfoxide and trimethylamine N-oxide. FEMS Microbiol.
Letters, 70; 33-36.
Oren, A., Ginzburg, M., Ginzburg, M. Z., Hochstein L. I. and Volcani, B. E. 1990.
Haloarcula marismortui (volcani) sp. Nov., nom, rev., an extremely halophilic
bacterium from Dead Sea. Int. J. of Syst. Bact., 40 (2); 209-210.
Oren, A. 1991. Anaerobic growth of halophilic archaeobacteria by reduction of
fumarate. J. of General Microbiol., 137; 1387-1390.
Oren, A. and Gurevich, P. 1993. Characterization of the dominant halophilic archaea in
a bacterial bloom in the Dead Sea. FEMS microbiology Ecology, 12; 249-256.
Oren, A. 1994. The ecology of the exteremely halophilic archaea. FEMS mic. Reviews,
13, 415-440.
Oren, A. Gurevich, p. 1994. Production of D-lactate, acetate and pyruvate from glycerol
in communities of halophilic archaea in Dead sea and in saltern crystalilzer
ponds. FEMS Microbiol. Ecol., 14, 147-156.
Oren, A., Gurevich, P., Gemmel, R. T. and Teske, A. 1995. Halobaculum gomorrense
gen. nov., sp. nov., a novel extremely halophilic archaeon from dead sea. I. J.
Syst. Bact., 45(4); 747-754.
Oren, A. 1996. Sensitivity of selected members of the family Halobacteriaceae to
quinolone antimicrobial compounds. Arch. Microbiol., 165; 354-358.
Oren, A., Duker, S. and Ritter, S. 1996. The polar lipid composition of Walsby’s square
bacterium. FEMS Microbiol. Letters, 138; 135-140.
Oren, A., Bratbak, G. and Heldal, . 1997. Occurence of virus like particles in the dead
sea. Extremophiles, 1: 143-149.
115

Oren, A., Ventosa, A. and Grant, W.D. 1997. Proposed minimal standarts for
description of new taxa in the order Halobacteriales. İnt. J. of Syst. Bacteriol., 47
(1); 233-238.
Oren, A and Litchfield, D. C. 1999. A procedure for the enrichment and isolation of
Halobacterium. FEMS Microbiology Letters, 173; 353-358.
Oren, A. 1999. Bioenergetic aspects of halophilism. Microbiol. And Molecul. Biology
Rev., 63(2); 334-348.
Oren, A. 1999. The enigma of square and triangular halophilic archaea. In: Enigmatic
Microorganisms and Life in Extreme Environments, 337-355. Edited by J.
Seckbah. Netherland: Kluwer academic publishers.
Oren, A., Ventosa, A., Gutierrez, M. C. and Kamekura, M. 1999. Haloarcula quadrata
sp. nov., A square, motile arcaeon isolated from a brine pool in Sinai (Egypt).
İnt. J. Syst. and Evolut. Microbiol., 49; 1149-1155.
Oren, A. 2000. Biological processes in the Dead sea as influenced by short-term and
long-term salinity changes. Arch. Hydrobiol. Spec. Issues Advanc. Limnol., 55;
531-542.
Oren, A. 2000. Life at High Salt Concentrations. In: The prokaryotes. A handbook on
the biology of Bacteria: Ecophsiology, Isolation, Identifications, Applications,
3rd. Edt., (Dwarkin, M., Falkow, S., Rosenberg, E., Schleifer, K. H. and
Stackebrandt, E., Eds). Springer-Verlag, New York (electronic publication).
Oren, A. 2001. The Order Halobacteriales. In: The prokaryotes. A handbook on the
biology of Bacteria: Ecophsiology, Isolation, Identifications, Applications, 3rd.
Edt., (Dwarkin, M., Falkow, S., Rosenberg, E., Schleifer, K. H. and
Stackebrandt, E., Eds). Springer-Verlag, New York (electronic publication).
Oren, A. and Rodriguez-Valera, F. 2001. The contribution of halophilic bacteria to the
coloration of saltern crystallizer ponds. FEMS Microbiology Ecology, 36; 123-
130.
Oren, A. 2002. Diversty of halophilic microorganisms: Environments, phylogeny,
physiology, and applications. J. Indust. Micro. And Biotech. 28, 56-63.
Oren, A. 2002. Molecular ecology of extremely halophilic arcahaea and bacteria. FEMS
Microbiol. Ecology, 39; 1-7.
116

Oren, A., Elevi, R., Watanabe, S., Ihara, K. and Corcelli, A. 2002. Halomicrobium
mukohatei gen. nov., comb. nov., and emended description of Halomicrobium
mukohatei. Int. J. Syst. and Evolut. Microbiol., 52; 1831-1835.
Parolis, H., Parolis, L. A. S., Boan, I. F., Rodriguez-Valera, F., Widmalm, G., Manca,
M. C., Jansson, P. E. and Sutherland, I. W. 1996. The structure of the
exopolysaccharide producedby the halophilic archaeon Haloferax mediterranei
strain R4 (ATCC 33500). Carbohydr. Res., 295, 147-156.
Pecher, T. and Böck, A. 1981. In vivo susceptibility of halophilic and mathanogenic
organisms to protein synthesis inhibitors. FEMS Microbiol. Letters, 10; 295-297.
Pfeifer. F., Weidinger, G. and Goebel, W. 1981a. Genetic variability in Halobacterium
halobium. J. Bacteriol., 145(1); 375-381.
Pfeifer. F., Weidinger, G. and Goebel, W. 1981b. Characterization of plasmids in
halobacteria. J. Bacteriol., 145(1); 369-374.
Pfeifer, F., Blaseio, U. And ghahraman, P. 1988. Dynamic plasmid populations in
Halobacterium halobium. J. of Bacteriology, 170 (8); 3718-3724.
Pfeifer. F and Blaseio, U. 1989. Insertion elements and deletion formation in a
halophilic archaebacterium. J. bacteriol., 171(9); 5135-5140.
Pfeifer. F., Blaseio, U. and Horne, M. 1989. Genome structure of Halobacterium
halobium: Plasmid dynamics in gas vacuole deficient mutants. Can. J. Mic., 35;
96-100.
Post, F. J. and Collins, N. F. 1982. A preliminary investigations of the membrane lipid
of Halobacterium halobium as a food additive. J. Food Biochem., 6; 25-38.
Prüß, B., Meyer, H. E. and Holldorf, A. W. 1993. Characterization of the
glyceraldehyde 3-phosphate dehyrogenase from the extremely halophilic
archaebacterium Haloarcula vallismortis. Arc. Microbiol., 160, 5-11.
Quesada, E., Ventosa, A., Rodriguez-Valera, F. and Ramos-Cormenzana. 1982. Types
and propoties of some bacteria isolated from hypersaline soils. J. Appl.
Bacteriol., 53, 155-61.
Rawal, n., Kelkar, S. M. and Altekar, W. Alternaticve routs of carbohydrate metabolism
in halophilic archaebacteria. Indian J. Biohemi Biophys., 25, 674-686.
Reese, E. R., Betts, R. F. and Gümüştop, B. 2000. Handbook of Antibiyotics. Third
edition, Lippincott Williams and Willkins, U.S.A.
117

Rodriguez-Valera, F., Ruiz-Berraquero, F. and Ramos-Cormenzana, A. 1979. Isolation
of extreme halophiles from seawater. Appl. And Environ. Microbiol., 38 (1),
164-165.
Rodriguez-Valera, F., Ruiz-Berraquero, F. and Ramos-Cormenzana, A. 1980.
Isolation of extremely halophilic bacteria able to grow in defined inorganic
media with single carbon sources. J. of General Microbiol., 119; 535-538.
Rodriguez-Valera, F., Juez, G. and Kushner, D. J. 1981. Halocins: salt-dependant
bacteriocins produced by extremely halophilic rods. Can. J. Microbiol., 28; 151-
154.
Rodriguez-Valera, F., Ruiz-Berraquero, F. and Ramos-Cormenzana, A. 1981.
Characteristics of the heterotrophic bacterial populations in hypersaline
environments of different salt concentrations. Microb. Ecol., 7; 235-243.
Rodriguez-Valera, F., Juez, G. and Kushner, D.J. 1983. Halobacterium mediterranei
spec. nov., a new carbohydrate-utilizing extreme halophile. System. Appl.
Microbiol., 4; 369-381.
Rodriguez-Valera, F., Ventosa, A., Juez, G. and Imhoff, J. F. 1985. Variation of
environmental features and micobial populations with salt concentrations in a
multi-pond saltern. Microb. Ecol., 11; 107-115.
Rodriguez-Valera, F. 1988. Characteristics and microbiyal ecology of hipersaline
environments. In: Halophilic Bacteria, F. Rodriguez-Valera (Ed.),V.I, CRC.
Press, Boca Raton, Florida, 3-30.
Rodriguez-Valera, F. 1992. Biotechnological potential of halobacteria In: The
Archaebacteria: Biochemistry and Biotechnology, Danson M. J., Hough, D. W.
and Lunt, G. G. (Ed.), The Biochemical Society, Portland Press, London, ISBN:
1-85578-010-0.
Roeßler and Müller, V. 2001. Osmoadaptation in bacteria and archaea: common
principles and differences. Environ. Microbiol., 3 (12); 743-759.
Rosenshine I. and Mevarech, M. 1989. Isolation and partial characterization of plasmids
found in three Halobacterium volcanii isolates. Can. J. Microbiol., 35; 92-95.
Ross, H. N. M., Collins, M. D., Tindall, B. J. and Grant, W. D. 1981. A rapid procedure
foe the detection of archaebacterial lipids in halophilic bacteria. J. of General
Microbiol., 123; 75-80.
118

Ross, H. N. M. and Grant, W. D. 1985. Nucleic acid studies on halophilic
archaebacteria. J. of General Micrbiol., 131; 165-173.
Ruepp, A. and Soppa, J. 1996. Fermentative arginine degredation in Halobacterium
salinarum (formerly Halobacterium halobium): Genes, genes products, and
transcripts of the arcRACB gene cluster. J. bacteriol., 178(8); 4942-4947.
Schiraldi, C., guiliano, M. and DeRosa, M. 2002. Perspectives on biotechnological
applications of archaea. Archaea, 1; 75-86.
Sellek, G. A. and Chaudhuri, J. B. 1999. Biocatalysis in organic media using enzymes
from extremophiles. Enzyme and Microbial Biotech., 25; 471-482.
Shahmohammadi, H. R., Asgarani, E., Teratı, H., Saito, T., Ohyama, Y., Gekko, K.,
Yamomoto, O. And Ide, H. 1998. protective roles of bacterioruberin and
intracellullar KCl in the resistance Halobacterium salinarum against DNA-
damaging agents. J. of Radaiation Research, 39 (4); 251-262.
Shand, R. F. and Perez, A. M. 1999. Haloarchaeal growth physiology IN: J. Seckbah
(Ed.) Enigmatic microorganisms and life in extreme environments, Kluwer
Acadamic Publisher Dordrecht, 414-424.
Sioud, M., Baldacci, G., de Rocondo, A. M. and Forterre, P. 1988. Novobiocin induces
positive supercoiling of small plasmids from halophilic archaebacteria in vivo.
Nucleic Acids Resourch, 16 (4); 1379-1391.
Sneath, P. H. A. and Sokal, R. R. 1973. Numerical taxonomy. Freeman, San Francisco.
Sözeri, S. 2000. Seyfe Gölü (Kırşehir) Çevresi Bitkilerine Katkılar. Türkiye Herbaloji
Dergisi, 3 (2); 19-33.
Stan-Lotter, H., Pfaffenhuemer, M., Legat, A., Busse, H. J., Radax, C. and Gruber, C.
2002. Halococcus dombrowskii sp. nov., an archaeal isolate from a permian
alpine salt deposit. Int. J. syst. And Evolut. Mic., 52; 1807-1814.
Stuart, E. S., Morshed, F, Sremac, M. and DasSarma, s. 2001. Antigen presentation
using novel particulate organelles from halophilic archaea. J. Biotech., 88; 119-
128.
Takashina, T., Hamamoto, T., Otozai, K., Grant, W. D. and Horikoshi, K. 1990.
Haloarcula japonica sp. Nov., a new triangular halophilic archaebacterium.
System. Appl. Microbiol. 13; 177-181.
119

Temiz, A. 1994. Genel mikrobiyoloji uygulama teknikleri. Şafak matbaacılık, Ankara,
ISBN: 975-95834-0-2.
Tindall, B. J. 1988. Prokaryotic life in the alkaline, saline, athalassic environment. In:
Halophilic Bacteria, F. Rodriguez-Valera (Ed.),V.I, CRC. Press, Boca Raton,
Florida, 31-67.
Tindall, B. J., Tomlinson, G. A. and Hochstein, L. I. 1989. Transfer of Halobacterium
denitrificans (Tomlinson, Jahnke, and Hochstein) to the genus Haloferax as
Haloferax denitrificans comb. Nov. Int. J. syst. Bacteriol., 39(3); 359-360.
Tindall, B. J. 1992. The family Halobacteriaceae In: The prokaryotes. A handbook on
the biology of Bacteria: Ecophsiology, Isolation, Identifications, Applications,
Balows, A., Trüper, H. G., Dworkin, M., Harder, W. and Schleifer, K. H. (Ed.),
V.I, Springer Verlag, New York, 768-808.
Tomlinson, G. A. and Hochstein, L. 1976. Halobacterium saccharovorum sp. nov., a
carbohydrate-metabolizing extremely halophilic bacterium. Can. J. Microbiol.,
22; 587-591.
Torreblanca, M., Rodriguez-Valera, F., Juez, G., Ventosa, A, Kamekura, M. and Kates,
M. 1986. Classification of non-alkaliphilic halobacteria based on numerical
taxonomy and polar lipid composition, and description of Haloarcula gen. nov.
and Haloferax gen. nov.. System. Appl. Microbiol., 8; 89-99.
Torreblanca, M., Meseguer, I. and Ventosa, A. 1994. Production of halocin is a
practically universal feature of archaeal halophilic rods. Letters in Appl.
Microbiol., 19; 201-205.
Trincone, A., Nicolaus, B., Lama, L., De Rosa, M., Gambacorta, A. and Grant, W. D.
1990. The glycolipid of Halobacterium sodomonse. J. of General Microbiol.,
136; 2327-2331.
Upasani, V. N., Desai, S. G., Moldoveanu, N. and Kates, M. 1994. Lipids of extremely
halophilic archeaobacteria from saline environments in India: a novel gljcolipid
in Natronobacterium strains. Microbiolgy, 140; 1959-1966.
Ventosa, A. and Nieto, J. J. 1995. Biotechnological applications and potentialities of
halophilic microorganisms. World J. Microbiol. Biotechnol., 11; 85-94.
120

Ventosa, A. Gutierrez, M. C., Kamekura, M. and Dyall-Smith, M. 1999. Proposal to
transfer Halococcus turcmenicus, Halobacterium trapanicum JCM 9743 and
strain GSL-11 to Haloterrigena turkmenica gen. nov, comb. nov. Int. J. of Syst.
Bacteriol., 49; 131-136.
Ventosa, A., Gutierrez, M. C., Kamekura, M., Zvyagintseva, I. S. and Oren, A. 2004.
Taxonomic study of Halorubrum distributum and proposal of Halorubrum
terrestre sp. nov. Int. J. Syst. and Evolut. Microbiol, 54; 389-392.
Vreeland, R. H., Straight, S., Krammes, J, Doughherity, K., Rosenweig, W. D. and
Kamekura, M. 2002. Halosimplex carlbadense gen. nov., sp. nov., a uniqe
halophilic archaeon with three 16S rRNA genes, that grows only in defined
medium with glycerol and acetate or pyruvate. Extremophiles, 6; 445-452.
Waino, M., Tindall, B. J. and Ingvorsen, K. 2000. Halorhabdus utahensis gen. nov., sp.
nov., an aerobic,extremely halophilic member of the Archaea from Great salt
Lake, Utah. Int. J. Syst. and Evolut. Microbiol., 50; 183-190.
Weidinger, G., Klotz, G. and Goebel, W. 1979. A large plasmid from Halobacterium
halobium carrying information for gas vacuole formation. Plasmid, 2; 377-386.
Wieland, F., Lechner, J. and Sumper, M. 1982. The cell wall glycoprotein of
Halobacterium structural, functional and biosynthetic aspects. Zbl. Bakt. Hyg. I.
Abt. Orig., C3; 161-170.
Woese, C. R. and Wolfe, R. S. 1985. The Bacteria, V.II, Academic Press, USA, ISBN:
0-12-307208-5.
Woese, C.R., Kandler, O. And Wheelis, M. L. 1990. Towards a natural system of
organisms: Proposal for the domains Archaea, Bacteria, and Eucarya. Proc. Natl.
Acad. Sci. USA., 87; 4576-4579.
Woese, C. R. 1999. Prokaryote Systematics. The evolulition of a science In: The
prokaryotes. A handbook on the biology of Bacteria: Ecophsiology, Isolation,
Identifications, Applications, 3rd. Edt., (Dwarkin, M., Falkow, S., Rosenberg, E.,
Schleifer, K. H. and Stackebrandt, E., Eds). Springer-Verlag, New York
(electronic publication).
121

Wolfang, L. and Klenk H-P. 2001. Overview: A phylogenetic backbone and taxonomic
framework for prokaryotic systematics In: Bergey’s Manual of Systematic
Bacteriology V.I, The Archaea and Deeply Branching and Phototrophic Bacteria,
Garrity, G. M. (Man. Ed.), 2nd Ed. New York, Springer, ISBN: 0387987711.
Wu, l., Chow, K. and Mark, K. 1983. The rol of pigments in Halobacterium cutirubrum
against uv irradiation. Microbiol. Letter, 24; 85-90.
Xin, H., Itoh, T., Zhou, P., Suzuki, K., Kamekura, M. and Nakase, T. 2000. Natrinema
versiforme sp. nov., an halophilic archaeon from Aibi salt lake. İnt. J. Syst. and
Evolut. Microbiol., 50; 1297-1303.
Xu, Y., Zhou, P. and Tian, X. 1999. Characterization of two haloalkaliphilic archaea
Natronorubrum bangense gen. nov., sp. nov. and Natronorubrum tibetense gen.
nov., sp. nov. İnt. J. Syst. Bac., 49; 261-266.
Yatsunami, R., Kawakami, T. and Ohtani, H. 2000. A novel bacteriorhodopsin like
protein from Haloarcula japonica strain TR-1: gene cloning, sequencing, and
transcript analysis. Extremophiles, 4; 109-114.
Yurdakulol, E., Öncel, I., Demirörs, M., Yıldız, A. ve Yüksel Keleş. 1996. Ecological
and syntaxonomic investigation of salt marshes vegetation in the vicinity of
Burdur and Acıgöl (Denizli/Turkey).Ecologia Mediterranea XXII (1/2), 51-61.
Zillig, W., Stetter, K. O., Schnabel, R. and Thomm, M. 1985. DNA-Dependent RNA
polymerases of the archaebacteria In: C.R. Woese and R. S. Wolfe (Eds), The
Bacteria-a treatise on structure and function. Archaebacteria, Academic Press
Orlando, FLVol.III, 499-521.
Zvyagintseva, I. S. and Tarasov, A. L. 1988. Extreme halophilic bacteria from saline
soils. Microbiology, 56, 664-668.
122

E K L E R
EK 1 SDS-PAGE’inde kullanılan stoklar ve hazırlanışları
EK 2 Plazmid İzolasyonunda Kullanılan Stok solüsyonları
123

EK 1. SDS-PAGE’inde kullanılan stoklar ve hazırlanışları
Akrilamid/Bisakrilamid çözeltisi
Akrilamid 29.2 g
Bisakrilamid 0.8 g
d. su 100 mL
Yığma Jeli (%4)
Akrilamid/Bisakrilamid 0.82 mL
d. su 2.93 mL
0.5 M Tris-HCl (pH 6.8) 1.25 mL
APS 30 µL
TEMED 5 µL
Ayırma jeli (%10)
Akrilamid/Bisakrilamid 5.78 mL
d. su 7.13 mL
1.5 M Tris-HCl (pH 8.6) 4.33 mL
APS (%10) 86.7 µL
TEMED 8.16 µL
Yükleme Tamponu Stoğu (pH 8.3)
Trizma base 1.65g
Glisin 8 g
SDS 1.4 g
d. su 1100 mL
124

Örnek tamponu (4X)
0.5 M Tris-HCl (pH 6.8) 5.12 mL
Gliserol 8 mL
SDS 2 g
d. su 3 mL
Merkaptoetanol 4 mL
Bromofenol blue 2.10-4 g
Coomassie Brillant Blue boya çözeltisi
Coomassie Brillant Blue R250 1.5 g
İzopropil alkol 250 mL
Glasiyel asetik asit 100 mL
d. su 650 mL
Boya Giderme Çözeltisi
İzopropil alkol 250 mL
Glasiyel asetik asit 100 mL
d. su 650 mL
125

EK 2. Plazmid İzolasyonunda Kullanılan Stok solüsyonları
Tris(50mM)-EDTA(0.25mM)-1 (pH 8)
Trizma base 0.605 g
EDTA 9.31 g
d. su 100 mL
Tris(2M)-Cl (pH 7)
Trizma base 24.22 g
d. su 100 mL
SDS Çözeltisi (pH 8)
Trizma base (50 mM) 0.605 g
EDTA (20 mM) 0.74 g
SDS 20 g
d. su 100 mL
Tris(10 mM)-EDTA(1 mM)-2(pH 7.5)-2
Trizma base 0.121 g
EDTA 0.37 g
d. su 100 mL
RNaz A Çözeltisi
5 mL Steril distile su içinde hazırlanan 0.05 M sodyum asetat çözeltisinin pH’ı asetik
asit ile 5’e ayarlanmış ve üzerine 5 mg RNaz A ilave edilmiştir. Kaynar su içinde 5
dakika tutulduktan sonra -20°C’de saklanmıştır.
126

Tris-Asetat Tamponu (10X) (pH 8)
Trizma base 4.84 g
Sodyum asetat 4.08 g
EDTA 0.37
d. su 1000 mL
Bromofenol Mavisi Çözeltisi
Bromofenol Mavisi 0.25 g
Sükroz 40 g
d. su 100 mL
127

ÖZGEÇMİŞ
13.08.1972 Malatya’da doğdu. İlk, orta ve lise öğrenimini Malatya’da tamamladı. 1989
yılında girdiği İnönü Üniversitesi Fen-Edebiyat Fakültesi Biyoloji Bölümü’den
(Malatya) 1993 yılında Biyolog ünvanıyla mezun oldu. 1995 yılında Mustafa Kemal
Üniversitesi Fen Bilimleri Enstitüsü Biyoloji Anabilim Dalı’nda başladığı yüksek lisans
öğrenimini, "Zeytinyağı Fabrikası Atığında Üretilen Lentinus tigrinus ve Laetiporus
sulphureus Funguslarından GA3, ABA ve Sitokinin Üretimi" konulu yüksek lisans
tezini vererek, 1997 yılında tamamladı. 1998 yılında Ankara Üniversitesi Fen bilimleri
Enstitüsü Biyoloji Anabilim Dalı’nda doktora öğrenimine başladı.
1995 yılında Mustafa Kemal Üniversitesi Fen-Edebiyat Fakültesi Biyoloji Bölümünde
Araştırma Görevlisi olarak göreve başlamıştır. 35. madde kapsamında doktora
öğrenimini yapmak üzere Ağustos 1999 yılından bu yana Ankara üniversitesi fen
fakültesi Biyoloji bölümünde görev yapmaktadır.
128