fasciola hepatica: identification of molecular markers for resistant and susceptible pseudosuccinea...
TRANSCRIPT

Experimental Parasitology 105 (2003) 211–218
www.elsevier.com/locate/yexpr
Fasciola hepatica: identification of molecular markersfor resistant and susceptible Pseudosuccinea columella snail hostsq
Alfredo Guti�errez,a Jean-Pierre Pointier,b Jorge Fraga,a Edouard Jobet,b Sylvain Modat,b
R.T. P�erez,c Mary Yong,a J. Sanchez,a Eric S. Loker,d and Andr�e Th�eronb,*
a Laboratorio de Malacolog�ıa, Instituto ‘‘Pedro Kouri’’, IPK. Apartado 601, Marianao 13, Ciudad Habana, Cubab UMR CNRS-UP 5555, EPHE, Parasitologie Fonctionnelle et Evolutive, CBETM Universit�e 52, Av. Paul Alduy. 66860 Perpignan Cedex, France
c Facultad de Biolog�ıa, Universidad de la Habana, Cubad Department of Biology, University of New Mexico, Albuquerque, NM, USA
Received 8 April 2003; received in revised form 5 November 2003; accepted 11 December 2003
Abstract
Protein electrophoresis, RAPD-PCR and nuclear rDNA ITS sequencing were performed to search for genetic differences between
Pseudosuccinea columella snails susceptible and resistant to Fasciola hepatica infection. Of the 21 enzymatic loci analyzed in both
populations, none of them exhibited neither within- or between-group variation. Such an absence of enzyme polymorphism support
the hypothesis of selfing as the ‘‘prevalent’’ mating system for this hermaphroditic species. Conversely, the RAPD profiles displayed
clear differences between susceptible and resistant isolates for 17 of the 26 primers tested while no within-group variation was
detected. rDNA ITS sequence analysis from snails of each isolates showed only two bases that differed between groups accounting
for a 0.17% of variation confirming that suceptible and resistant snails belong to the same species. This is the first time that a genetic
variation using RAPD markers is demonstrated between susceptible and resistant lymnaeid snails vis-a-vis of F. hepatica infection in
absence of experimental selection.
� 2003 Elsevier Inc. All rights reserved.
Index Descriptors and Abbreviations: DNA, desoxyribonucleic acid; rDNA, ribosomal DNA; ITS, internal transcribed spacer; PCR, polymerase
chain reaction; RAPD, random amplified polymorphic DNA; S, similitude coefficient
Keywords: Fasciola hepatica; Lymnaeidae; Pseudosuccinea columella; Susceptibility; Resistance; Markers
1. Introduction
Fasciola hepatica is widely distributed in temperateand subtropical areas around the world, infecting nu-
merous species of mammals as definitive hosts including
human (Chen and Mott, 1990; Mas-Coma et al., 1999)
and various species of snails of the family Lymnaeidae as
intermediate hosts (Mas-Coma and Bargues, 1997). In
the island of Cuba, two lymnaeid snails, Fossaria cub-
ensis and Pseudosuccinea columella are involved in the
life cycle of F. hepatica and display experimentally a high
qThe sequence data reported herein have been deposited in
GenBank under Accession No. AY186751 (Pseudosuccinea columella).* Corresponding author. Fax: +33-4-68-66-22-81.
E-mail address: [email protected] (A. Th�eron).
0014-4894/$ - see front matter � 2003 Elsevier Inc. All rights reserved.
doi:10.1016/j.exppara.2003.12.006
level of susceptibility to this parasite (Guti�errez et al.,
2000). However, we demonstrated previously that a
population of P. columella, sampled in the outskirts ofLa Palma municipality, province of Pinar del Rio, was
not susceptible to this parasite (Guti�errez et al., 2002,
2003). After repeated attempts to induce infection in
these snails, using high miracidial doses (more than 15
miracidia per snail), no evidence of intramolluscan stages
of F. hepatica was observed in any of the snails exposed
and many of them died a few weeks after exposure.
Studies of Richards (1973, 1975, 1984) and Richardsand Merritt (1972) have demonstrated that snail sus-
ceptibility/resistance to schistosome infection is con-
trolled by genetic factors and genetic variability within
snail and parasite populations may affect parasite
transmission in endemic areas (Sire et al., 2001; Vidigal

212 A. Guti�errez et al. / Experimental Parasitology 105 (2003) 211–218
et al., 1994; Vieira et al., 1991). The present study wasdesigned to differentiate between susceptible and resis-
tant P. columella to F. hepatica infection based on en-
zyme electrophoresis and random amplification of
polymorphic DNA PCR-based technology. Both tech-
niques have been employed before to detect genetic
variations between susceptible and resistant laboratory
selected Biomphalaria snails to schistosome infection
(Abdel-Hamid et al., 1999; Knight et al., 1999; Larsonet al., 1996; Michelson and Dubois, 1981; Mulvey and
Woodruff, 1985). We also sequenced the nuclear rDNA
ITS region in order to detect genetic variation which
might separate the resistant isolate as a different species.
The knowledge about the molecular basis of snail–
trematode interactions and the identification of resis-
tance genes in the snail (Knight et al., 2000; Rollinson
et al., 1998) is of particular interest for a better under-standing of parasitic disease epidemiology and control.
2. Material and methods
2.1. Snails
All snails used in this study were obtained from cul-tures established from isolates of natural populations of
P. columella from Cuba. Susceptible snail cultures were
established with parental snails sampled from two field
populations, one located south (Parque Lenin) and the
other located west (Punta Brava) of Havana City. The
resistant snail culture was established with snails origi-
nating from La Palma population, located north of Pinar
del R�io province. Resistance was histologically demon-strated by the presence of amebocytes surrounding the
miracidium 24 h after infection (Guti�errez et al., 2003).
2.2. Enzyme analysis
Protein extracts: a total of 20 snails (both susceptible
and resistant) were used for the enzyme electrophoresis
Table 1
Number of loci found for 11 isoenzymes tested for the characterization of s
Protein
Code Abbreviation Na
EC 1.15.1.1 SOD Su
EC 3.2.1.1 AMY a-AEC 1.1.1.37 MDH M
EC 4.2.1.2 FUM Fu
EC 1.1.1.204 XDH Xa
EC 2.6.1.1 AAT As
EC 3.1.3.1 AKP Al
EC 3.1.1.1 EST Es
EC 3.1.3.2 ACP Ac
EC 5.3.1.8 MPI M
EC 3.1.4.47 GPI Gl
analysis (10 snails each group). Snails were removedfrom their shells and the soft parts were homogenized in
distilled water and stored at )70 �C until they were to be
used for the electrophoresis. Protein extracts of indi-
vidual snails were run on 10% starch gels for 4 h. After
completion of electrophoresis, bands were revealed by
specific enzymatic staining. Details of the methodology
for protein electrophoresis and enzymatic staining can
be found in Pasteur et al. (1987). Eleven enzymaticsystems were tested and the analysis revealed the pres-
ence of 21 putative loci (Table 1).
2.3. Random amplification of polymorphic DNA analysis
The foot of 14 ethanol-preserved snails from sus-
ceptible and resistant isolate was removed avoiding the
pigmented area and a portion of each foot was takenfor the analysis. Genomic DNA was extracted follow-
ing the methodology of Winnepenninckx et al. (1993)
modified by Bargues and Mas-Coma (1997): foot
samples were put in 400 ll of lysis buffer (10mM Tris–
HCl, pH 8.0; 100mM EDTA; 100mM NaCl; 1% so-
dium dodecyl sulfate [SDS]) containing 500 ll/ml of
proteinase K and digested for 4 h at 55 �C with manual
shaking every 15min. The extraction took place inthree steps: (i) an equal volume (400 ll) phenol was
added; (ii) the aqueous phase obtained (about 400 ll)was mixed with 200 ll phenol and 200 ll chloroform/
isoamyl alcohol (24/1); and (iii) the aqueous phase
obtained (about 400 ll) was mixed with an equal vol-
ume of chloroform/isoamil alcohol (24/1). Each step
was followed by a slow shaking and centrifugation for
15min at 14,000 rpm at room temperature. The aque-ous phase finally obtained was precipitated with 1/10
volume of 3M sodium acetate and 2.5 volumes of
chilled absolute ethanol, then refrigerated for 10min at
)20 �C. The DNA was centrifuged for 30min at
14,000 rpm at 4 �C and the pellet washed with 1ml of
70% ethanol then air dried before resuspending in
110 ll of ultra-pure water.
usceptible and resistant P. columella
me Number of loci
peroxide dismutase 2
mylase 2
alate deshydrogenase 2
marase 1
nthine dehydrogenase 1
partate amino trasferase 1
kaline phosphatase 1
terases 8
id phosphatase 1
annose phosphate isomerase 1
ycosylphosphatidyl inositol 1

A. Guti�errez et al. / Experimental Parasitology 105 (2003) 211–218 213
Twenty six oligonucleotides (Kits A, B and G, Op-eron Technologies, California, USA), were used for the
amplification of random DNA markers to reveal genetic
diversity between individual snails. RAPD reagents and
concentrations were optimized in preliminary experi-
ments (data not shown). DNA amplification was per-
formed in a final volume of 25 ll containing 2.5 ll of10� PCR buffer (100mM Tris–HCl pH 8.3, 15mM
MgCl2, 500mM KCl, and 0.01% gelatin) (Boehringer–Mannheim, Germany), 200 lM each dNTP (Boehrin-
ger–Mannheim, Germany), 5 pmol single primer, 2.5U
Taq DNA polymerase (Boehringer–Mannheim, Ger-
many) and 10 ng template DNA. Negative controls for
each of the 26 primers used contained all of the above
components except sterile distilled water in place of
template DNA. The amplification profile consisted of an
initial denaturation step at 94 �C for 5min followed by45 repetitions of 1min at 94 �C, 1min at 36 �C, and
2min at 72 �C. In the final cycle, the extension step was
for 5min PCR products were analyzed by electropho-
resis in 1.2% agarose gels in TBE buffer containing
ethidium bromide (0.5mg/ml) and visualized using a UV
transilluminator.
A similitude coefficient (S) between groups was calcu-
lated for each primer using the following formula (Dice,1945): S ¼ 2a=ð2aþ bþ cÞ where a represents the num-
ber of bands shared by both groups, b represents the
number of marker bands specific for susceptible snails,
and c represents the number of marker bands specific for
resistant snails. This coefficient was useful to measure the
degree of between-group differentiation among primers.
2.4. rDNA sequencing and analysis
The foot of 13 ethanol-preserved snails of each isolate
was taken for the analysis. Three foot samples of each
Table 2
Size (in base pairs) of the RAPD markers obtained for the characterization
Primer Sequence Suscepti
OPA-01 CAGGCCCTTC 1380
OPA-02 TGCCGAGCTG 250
OPA-04 AATCGGGCTG 480
OPA-06 GGTCCCTGAC 1100
OPA-07 GAAACGGGTG 710, 650
OPA-08 GTGACGTAGG 530
OPA-09 GGGTAACGCC 697, 730
OPA-16 AGCCAGCGAA —
OPA-17 GACCGCTTGT 1489
OPA-18 AGGTGACCGT 697, 740
OPA-19 CAAACGTCGG 421
OPA-20 GTTGCGATCC 1000
OPB-08 GTCCACACGG 500
OPG-05 CTGAGACGGA 350, 450
OPG-10 AGGGCCGTCT —
OPG-17 ACGACCGACA 400, 590
OPG-19 GTCAGGGCAA 240
S, similitude coefficient.
isolate were pooled together for a first sequencing trial,the remaining 10 foot samples were used individually in
a second trial. Genomic DNA extraction protocol was
identical as previously described for the RAPD analysis.
The polymerase chain reaction (PCR) was used to
amplify the rDNA fragment corresponding to the 30 re-gion of the 18S, ITS-1, 5.8S, ITS-2, and the 50 region of
the 28S contained between the forward primer Pc ITS f
50-GTCGTAACAAGGTTTCCGTAG-30 (obtained byaligning 10 sequences of the 18S, found on GenBank,
from 10 snails using the software CLUSTALW) and the
reverse primer Pc ITS r 50-TTCTATGCTTAAATTC
AGGGG-30 (Bargues et al., 2001). PCR were carried out
using 3 ll of genomic DNA for a total reaction volume
of 40 ll. After cloning and sequencing the resulting long
products (containing 1170 bp), other primers were de-
signed in order to amplify and sequence their centralpart where only about 400 bp had been clearly se-
quenced on a first trial at the 50 and 30 regions: Pc ITS r1
50-CGATATCGGTGGAAGCA-30 (position 289) and
Pc ITS f1 50-CAACACGCCTCTCGAGA-30 (position810). Each PCR was programmed for 35 cycles with the
following stages: (i) 95 �C for 3min, (ii) 35 cycles at
95 �C for 1min, 55 �C for 1min, and 72 �C for 2min, and
(iii) 72 �C for 5min to complete extension. The productsobtained were viewed on an ethidium bromide stained
1.5% agarose gel. The amplified products were cloned
using the ‘‘TOPO TA cloning kit’’ (Invitrogen) and se-
quenced using the ‘‘Big Dye Terminator’’ sequencing kit
(Perking–Elmer, Applied Biosystems) which uses the
method described by Sanger et al. (1977). The reaction
products were separated by 6% polyacrylamide gel
electrophoresis using an automatic sequencer ‘‘373ADNA sequencer’’ (Applied Biosystems) and DNA se-
quences were aligned using the software ‘‘Sequencher
3.1.1’’ (Gene Codes).
of susceptible and resistant P. columella
ble snails Resistant snails S
150, 1490 0.5
— 0.75
421 0.6
2300 0.5
510, 810 0.43
600 0.57
710, 820 0.43
1037 0.83
— 0.83
718 0.57
380, 500 0.5
970 0.66
400, 505 0.25
, 780 225, 400 0.36
710 0.83
, 600 500, 605,870 0.22
— 0.75
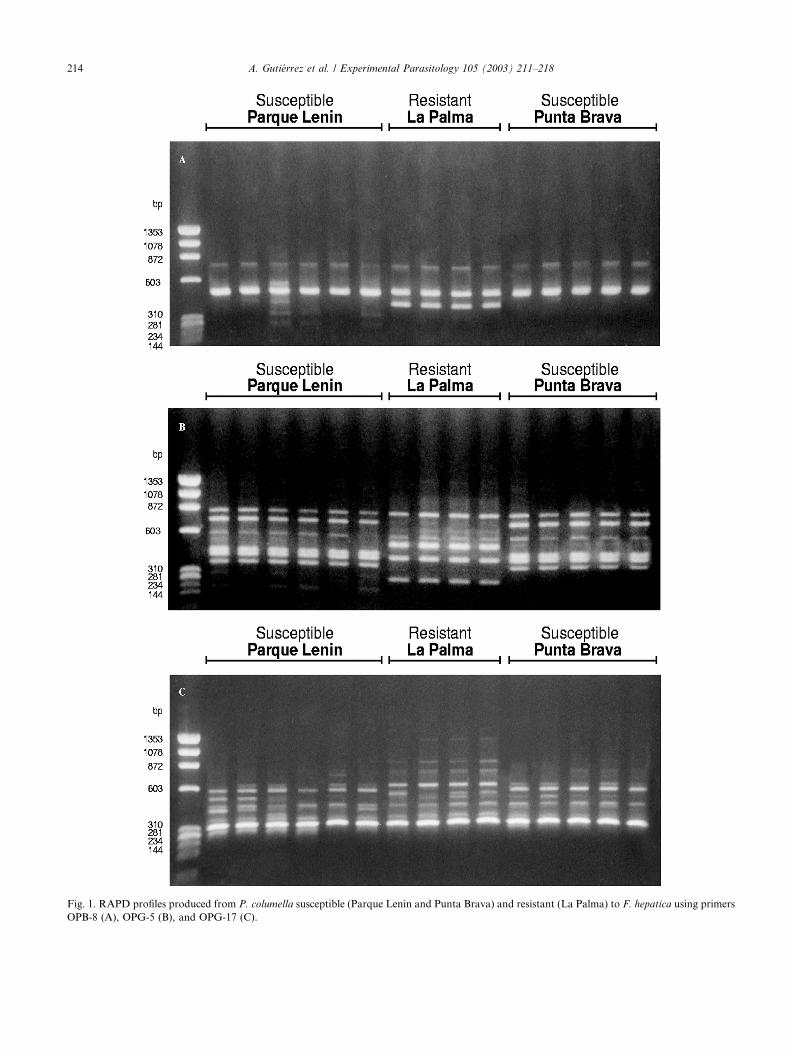
Fig. 1. RAPD profiles produced from P. columella susceptible (Parque Lenin and Punta Brava) and resistant (La Palma) to F. hepatica using primers
OPB-8 (A), OPG-5 (B), and OPG-17 (C).
214 A. Guti�errez et al. / Experimental Parasitology 105 (2003) 211–218

A. Guti�errez et al. / Experimental Parasitology 105 (2003) 211–218 215
3. Results
3.1. Protein electrophoresis
The number of loci for each of the 11 isoenzyme
systems tested is shown in Table 1. Neither within- nor
between-group polymorphism was obtained for any of
the 21 loci revealed in both susceptible and resistant
snails. A high allele fixation was observed for all systemsat every locus. The highest number of loci (8) was found
for the esterase system. The remaining enzymes showed
either one locus or two loci.
3.2. RAPD-PCR analysis for the identification of markers
Of the 26 primers tested, 17 showed notable differ-
ences between resistant and susceptible snails. Allprimers together produced a total of 140 bands and 44
bands were specifically obtained in one group or the
other. Twenty-two bands were found to be markers
for susceptible snails and 22 bands for resistant snails.
Molecular weights of marker bands are shown in
Table 2. Fig. 1, shows the RAPD profiles of three of the
primers (OPB-08, OPG-05, and OPG-17) that produced
the highest between-group differences (e.g., similitudecoefficient of 0.25, 0.36, and 0.22, respectively).
Using primers OPB-8, OPG-5 (Fig. 1), OPG-10,
OPG-16, and OPG-19, no difference was observed be-
tween the two susceptible groups of P. columella (Par-
que Lenin and Punta Brava).
Fig. 2. rDNA sequence of the region comprising the 30 portion of the 18S, the
resistant and susceptible P. columella. X at the position 555 corresponds to ad
725 corresponds to cytosine for susceptible and thymine for resistant snails. H
and Pc ITS r1 used for reamplify the central region.
3.3. rDNA sequencing
The sequence of ribosomal DNA corresponding to
the 30 region of 18S, ITS-1, 5.8S, ITS-2, and the 50 regionof the 28S from pools of three resistant and three sus-
ceptible snails (Fig. 2) shows a difference of only two
bases from a total of 1170, accounting for a 0.17% of
variation between both isolates. These differences cor-
respond to position 555 (adenine for susceptible andguanine for resistant snails) and to position 725 (cyto-
sine for susceptible and thymine for resistant snails).
When comparing this sequence with those published in
GenBank the structure of this fragment may be de-
scribed as follows: bases 1–45, 30 portion of 18S; bases
46–565, ITS1; bases 566–722, 5.8S; bases 723–1122,
ITS2; bases 1123–1170, 50 portion of 28S. A G+C bias
can be observed in the sequence bases composition(57.95% of G+C vs. a 41.88% of A+T).
A second amplification and sequencing trial was
carried out on single rDNA extracted from 10 snails of
each isolate. This fragment comprised a region of about
500 bp containing the bases that had shown variation in
the first trial. Amplification was performed using the
primers Pc ITS f1 and Pc ITS r1 (see sequence in Fig. 2).
Results confirmed those two bases as the only geneticdifference between susceptible and resistant snails. It is
worth noting that one of those bases (position 555 of the
rDNA sequence) is located at the 30 portion of ITS1
whereas the other (position 725 of the same sequence) is
located at the 50 portion of ITS2.
ITS-1, the 5.8S, the ITS-2 and the 50 portion of the 28S of individuals of
enine for susceptible and guanine for resistant snails; Y at the position
ighlighted bases correspond to the sequences of the primers Pc ITS f1

216 A. Guti�errez et al. / Experimental Parasitology 105 (2003) 211–218
4. Discussion
Various biochemical and molecular techniques have
been used to search for genetic markers associated with
susceptible and resistant phenotypes of snails in relation
to digenetic trematode infection (Rollinson et al., 1998).
Most of the studies have concerned the Biomphalaria
glabrata/Schistosoma mansoni combination (see Knight
et al., 2000).From isoenzyme analysis, Mulvey and Woodruff
(1985) showed that biochemical differences existed be-
tween genetically defined resistant and susceptible
snails. Michelson and Dubois (1981), reported the
identification of an isoenzyme marker (AcP2), which
appeared to be associated with susceptibility and re-
fractory (dominant) state of laboratory strains of Bi-
omphalaria glabrata to S. mansoni. In the presentstudy, we were unable to detect biochemical variation
from the analysis of 11 enzymatic systems between and
within groups of P. columella resistant and susceptible
to F. hepatica infection. This absence of enzyme poly-
morphism and the presence of a high allele fixation
support the hypothesis of selfing as the ‘‘prevalent’’
mating system for this hermaphroditic species. Jab-
bour-Zahab et al. (1997) have already observed such amonomorphism within populations of F. cubensis and
L. truncatula, from different geographic sites. They
attributed this absence of variation to the possible role
of the mating system and the population dynamics, as
proposed previously for several monomorphic popula-
tions of Biomphalaria and Bulinus (Jarne, 1995). Con-
versely, the RAPD analysis performed in this study
demonstrated sharp differences between resistant andsusceptible P. columella. This indicates that, despite the
lack of within-group polymorphism observed, genetic
variability does exist at the DNA level between both
isolates. The genetic differences were obtained for 17
out of 26 primers tested and concerned 44 of the 140
bands scored. This is the first time that a genetic var-
iation is demonstrated between susceptible and resis-
tant lymnaeid snails vis-a-vis of F. hepatica infection inabsence of experimental selection. By using RAPD-
PCR analysis Larson et al. (1996) were able to differ-
entiate genetically defined lines of B. glabrata selected
for susceptibility and resistance to S. mansoni. Of the
20 primers they tested, 2 (OPA-1 and OPA-6) were
useful in distinguishing between the two snail lines. The
stable inheritance of these markers in individual prog-
eny was also studied. Using bulked segregant analysisand RAPD-PCR, Knight et al. (1999) identified two
other markers (OPM-04, 1.2 kb and OPZ-11, 1.0 kb)
from 205 primers tested that segregate with the adult
resistant phenotype of B. glabrata snails. RAPDs ge-
netic variation was also detected for Biomphalaria
tenagophila susceptible and resistant laboratory selected
snails (Abdel-Hamid et al., 1999).
One question that may arise is whether resistant iso-lates belong or not to a different species other than P.
columella, given the genetic differences observed with the
RAPDs markers. Despite some phenotypic differences
that have been found between resistant and susceptible
isolates regarding shell morphometry, mantle pigmen-
tation pattern, and egg laying behavior (Guti�errez et al.,2003), no variation has been observed in the general
characteristics that define the species like for instance theshell width and body whorl shape and some specific traits
of the reproductive system (unpublished data). The
rDNA sequencing from snails of each isolate shows
clearly that only a little variation (two bases which ac-
count for 0.17% of the sequenced fragment) is present
between resistant and susceptible snails and this result
should confirm that both isolates correspond to the same
species. Vigo et al. (2000) and Bargues et al. (2001) haveconcluded that the ITS-2 (comprised in the sequenced
rDNA fragment in this study) is an useful marker for
resolving supraspecific, specific, and population rela-
tionships in Lymnaeidae, even in spite of the G+C bias.
Pseudosuccinea columella has been always reported as
highly susceptible to F. hepatica in different parts of the
world (Boray et al., 1985; Cruz-Reyes and Malek, 1987;
Dacal et al., 1988; Rondelaud and Barthe, 1987) and wehave successfully infected several Cuban isolates in our
laboratory for many years. The resistant snails from La
Palma locality are an exception. Regarding life-history
traits, uncommon relationships have been demonstrated
between susceptible infected Cuban specimens of P.
columella and F. hepatica (Guti�errez et al., 2002, 2003).
While F. hepatica is known to castrate its host and to
induce an increase of shell growth in L. truncatula
(Wilson and Denison, 1980), infected susceptible P.
columella showed an increased fecundity during the
patent period of infection and no increase of size (Gu-
ti�errez et al., 2002, 2003). On the other hand, exposed
resistant snails exhibit a reduced survival and fecundity
rates compared to non-exposed resistant controls and to
infected susceptible snails. Furthermore, non-exposed
resistant snails showed lower fecundity rates comparedto non-exposed susceptible P. columella. These observed
differences suggest a possible fitness and physiological
cost associated with resistance of P. columella to dige-
nean infection (Coustau et al., 2000; Minchella and
LoVerde, 1983).
The inheritance pattern of the specific markers iden-
tified for susceptibility and resistance of P. columella
remains to be established. However, mating experimentsmay not resolve this issue if selfing constitutes the al-
most exclusive mode of reproduction of this snail. Re-
cently Meunier et al. (2001) also demonstrated that L.
truncatula was almost exclusive self-crossing, since they
observed a single microsatellite multilocus genotype
among all the individuals sampled from a wide endemic
area of F. hepatica at the Bolivian Altiplano.

A. Guti�errez et al. / Experimental Parasitology 105 (2003) 211–218 217
The origin of the natural population of resistant P.columella is so far unknown and studies to determine the
nature and distribution of the resistant gene(s)
throughout the species range in Cuba are now necessary.
Subsequent studies will be directed toward a better un-
derstanding of the mechanisms involved in the recog-
nition and destruction of the parasite within the snail
(Coustau and Yoshino, 2000; Sapp and Loker, 2000) as
well as the isolation of genes or gene products involvedin the susceptibility or resistance of snails to this para-
sitic infection (Knight et al., 2000).
Acknowledgments
We are grateful to G. Espinosa, Faculty of Biology,
University of Havana, for her valuable help with RAPDtechnique and J.R. Pag�es, University of Perpignan for
his contribution with image management. We thank C.
Coustau from the CNRS and M.D. Bargues from the
University of Valencia (Spain) for reading the manu-
script and helpful comments. This work received finan-
cial support from the CNRS (France)/CITMA (Cuba)
12090-project and CORUS 02 736 087-project.
References
Abdel-Hamid, A.-H.Z., De Molfetta, J.B., Fernandez, V., Rodrigues,
V., 1999. Genetic variation between susceptible and non-suscepti-
ble snails to Schistosoma infection using random amplified poly-
morphic DNA analysis (RAPDs). Revista do Instituto de Medicina
Tropical de Sao Paulo 41, pp 291–295.
Bargues, M.D., Mas-Coma, S., 1997. Phylogenetic analysis of
lymnaeid snails based on 18S rDNA sequences. Molecular Biology
and Evolution 14, 569–577.
Bargues, M.D., Vigo, M., Horak, P., Dvorak, J., Patzner, R.A.,
Pointier, J.P., Jackiewicz, M., Meier-Brook, C., Mas-Coma, S.,
2001. European Lymnaeidae (Mollusca: Gastropoda), intermediate
hosts of trematodiases, based on nuclear ribosomal DNA ITS-2
sequences. Infection, Genetics and Evolution 1, 85–107.
Boray, J.C., Fraser, G.C., Williams, J.D., Wilson, J.M., 1985. The
occurrence of the snail Lymnaea columella on grazing areas in New
South Wales and studies on its susceptibility to Fasciola hepatica.
Australian Veterinary Journal 62, 4–6.
Chen, M.G., Mott, K.E., 1990. Progress in assessment of morbidity
due to Fasciola hepatica infection: a review of recent literature.
Tropical Diseases Bulletin 87, R1–R38.
Coustau, C., Yoshino, T.P., 2000. Flukes without snails: advances in
the in vitro cultivation of intramolluscan stages of trematodes.
Experimental Parasitology 94, 62–66.
Coustau, C., Chevillon, C., ffrench-Constant, R., 2000. Resistance to
xenobiotics and parasites: can we count the cost. Trends in Ecology
and Evolution 15, 378–383.
Cruz-Reyes, A., Malek, E.A., 1987. Suitability of six lymnaeid snails
for infection with Fasciola hepatica. Veterinary-Parasitology 24,
203–210.
Dacal, A.R.C., Costa de Araujo, H.M., Leite, A.C.R., 1988. Suscep-
tibilidade de Lymnaea (Pseudosuccinea) columella (Say, 1817)
exposta a infeccao por miracidios de Fasciola hepatica (Linnaeus,
1758). Revista do Instituto de Medicina Tropical de Sao Paulo. 30,
361–369.
Dice, L.R., 1945. Mesures of the amount of ecological association
between species. Ecology 26, 297–302.
Guti�errez, A., Perera, G., Yong, M., S�anchez, J., Wong, L., 2000. Life-
history traits of Fossaria cubensis (Gastropoda: Lymnaeidae) under
experimental exposure to Fasciola hepatica (Trematoda: Digenea).
Memorias do Instituto Oswaldo Cruz 95, 747–752.
Guti�errez, A., Pointier, J.-P., Yong, M., Sanchez, J., Th�eron, A., 2003.
Evidence of phenotypic differences between resistant and suscep-
tible isolates of Pseudosuccinea columella to Fasciola hepatica in
Cuba. Parasitology Research 40, 129–134.
Guti�errez, A., Yong, M., Perera, G., Sanchez, J., Theron, A., 2002.
Fasciola hepatica (Trematoda: Digenea), its effect on the life-history
traits of Pseudosuccinea columella (Gastropoda: Lymnaeidae). An
uncommon interaction. Parasitology Research 88, 535–539.
Jabbour-Zahab, R., Pointier, J.P., Jourdane, J., Jarne, P., Oviedo,
J.A., Bargues, M.D., Mas-Coma, S., Angl�es, R., Perera, G.,
Balzan, C., Khallayoune, K., Renaud, F., 1997. Phylogeography
and genetic divergence of some lymnaeid snails, intermediate hosts
of human and animal fascioliasis with special reference to
lymnaeids from the Bolivian Altiplano. Acta Tropica 64, 191–
203.
Jarne, P., 1995. Mating system, bottlenecks and genetic polymorphism
in hermaphroditic animals. Genetical Research 65, 193–207.
Knight, M., Miller, A.N., Patterson, C.N., Rowe, C.G., Michaels, G.,
Carr, D., Richards, C.S., Lewis, F.A., 1999. The identification of
markers segregating with resistance to Schistosoma mansoni infec-
tion in the snail Biomphalaria glabrata. Proceedings of the National
Academy of Sciences 96, 1510–1515.
Knight, M., Ongele, E., Lewis, F.A., 2000. Molecular studies of
Biomphalaria glabrata, an intermediate host of Schistosoma man-
soni. International Journal for Parasitology 30, 535–541.
Larson, S.E., Andersen, P.L., Miller, A.N., Cousin, C.E., Richards,
C.S., Lewis, F.A., Knight, M., 1996. Use of RAPD-PCR to
differenciate genetically defined line of an intermediate host of
Schistosoma mansoni, Biomphalaria glabrata. Journal of Parasitol-
ogy 82, 237–244.
Mas-Coma, S., Bargues, M.D., 1997. Human liver flukes: a review.
Reasearch and Reviews in Parasitology 57, 145–218.
Mas-Coma, S., Esteban, J.G., Bargues, M.D., 1999. Epidemiology of
the human fascioliasis: a review and proposed new classification.
Bulletin of the World Health Organisation 77, 340–346.
Meunier, C., Tirard, C., Hurtrez-Bousses, S., Durand, P., Bargues,
M.D., Mas-Coma, S., Pointier, J.-P., Jourdane, J., Renaud, F.,
2001. Lack of molluscan host diversity and the transmission of an
emerging parasitic disease in Bolivia. Molecular Ecology 10, 1333–
1340.
Michelson, E.H., Dubois, L., 1981. An isoenzyme marker possibly
associated with the susceptibility of Biomphalaria glabrata popu-
lations to Schistosoma mansoni. Acta Tropica 38, 419–426.
Minchella, D.J., LoVerde, P.T., 1983. Laboratory comparison of the
relative success of Biomphalaria glabrata stocks which are suscep-
tible and insusceptible to infection with Schistosoma mansoni.
Parasitology 86, 335–344.
Mulvey, M., Woodruff, D.S., 1985. Genetic of Biomphalaria glabrata
linkage analysis of genes for pigmentation, enzymes and resistance
to Schistosoma mansoni. Biochemical Genetics 22, 877–889.
Pasteur, N., Pasteur, G., Bonhomme, F., Catalan, J., Britton-
Davidian, J., 1987. Manuel technique de g�en�etique par �electro-
phor�ese des prot�eines. Lavoisier Editeur. Paris.Richards, C.S., 1973. Susceptibility of adult Biomphalaria glabrata to
Schistosoma mansoni infection. American Journal of Tropical
Medicine and Hygiene 22, 748–756.
Richards, C.S., 1975. Genetic factors in susceptibility of Biomphalaria
glabrata for different strains of Schistosoma mansoni. Parasitology
70, 231–241.

218 A. Guti�errez et al. / Experimental Parasitology 105 (2003) 211–218
Richards, C.S., 1984. Influence of snail age on genetic variations in
susceptibility of Biomphalaria glabrata for infection with Schisto-
soma mansoni. Malacologia 25, 493–502.
Richards, C.S., Merritt, J.W., 1972. Genetic factors in the suscepti-
bility of juvenile Biomphalaria glabrata to Schistosoma mansoni
infection. American Journal of Tropical Medicine and Hygiene 21,
425–434.
Rollinson, D., Stothard, J.R., Jones, C.S., Lockyer, A.E., de Souza,
C.P., Noble, L.R., 1998. Molecular characterisation of intermedi-
ate snail hosts and the search for resistance genes. Memorias do
Instituto Oswaldo Cruz 93, 111–116.
Rondelaud, D., Barthe, D., 1987. Fasciola hepatica L.: Etude du
d�eveloppement des r�edies chez quatre esp�eces de limn�ees.Bulletin de la Soci�et�e Francaise de Parasitologie 5, 99–
104.
Sanger, F., Nicklen, S., Coulson, A.R., 1977. DNA sequencing with
chain-terminating inhibitors. Proceedings of the National Academy
of Sciences USA 74, 5463–5467.
Sapp, K.K., Loker, E.S., 2000. A comparative study of mechanisms
underlying digenean-snail specificity: in vitro interactions between
hemocytes and digenean larvae. Journal of Parasitology 86, 1020–
1029.
Sire, C., Langand, J., Barral, V., Th�eron, A., 2001. Parasite (Schis-
tosoma mansoni) and host (Biomphalaria glabrata) genetic diversity:
population structure in a fragmented landscape. Parasitology 122,
545–554.
Vidigal, T.H.D.A., Dias Neto, E., Carvalho, O.D.S., Simpson, A.J.G.,
1994. Biomphalaria glabrata extensive genetic variation in Brazilian
isolates revealed by random amplified polymorphic DNA analysis.
Experimental Parasitology 79, 187–194.
Vieira, L.Q., Correa-Oliveira, R., Katz, N., De Souza, C.P., Carvalho,
O.S., Araujo, N., Sher, A., Brindley, P.J., 1991. Genomic variabil-
ity in field populations of S. mansoni in Brazil as detected with a
ribosomal gene probe. American Journal of Tropical and Medical
Hygiene 44, 69–78.
Vigo, M., Bargues, M.D., Yong, M., Arenas, J.A., Naquira, C.,
Paraense, W.L., Pointier, J.P., Mas-Coma, S., 2000. Molecular
characterization of a snail species transmitting Fasciolasis, Lym-
naea columella (Gastropoda: Lymaeidae), from Cuba, Guade-
loupe, venezuela, Peru and Brazil. In: XVth Unternational
Congress For Tropical Medicine and Malaria (Cartagena, Colom-
bia). Abstract books 2, 156.
Wilson, R.A., Denison, J., 1980. The parasitic castration and
gigantism of Lymnaea truncatula infected with the larval stages of
Fasciola hepatica. Zeitschrift f€ur Parasitenkunde 61, 109–119.
Winnepenninckx, B., Backeljau, T., DeWatcher, R., 1993. Extraction
of high molecular weight DNA from molluscs. Trends in Genetics
9, 407.