exteapyramidal nuclei among mammals. a quantitative study
TRANSCRIPT

EXTRAPYRAMIDAL NUCLEI AMONG 3IAMhlALS
A QUANTITATIVE STUDY
GERHARDT VON B O N N AND QHOUSE AHMED SHARIFF Department of Anatomy, University of Zllinois
FJVE FIOUBEB
The extrapyramidal system still appears as an ill-defined and little understood assembly of gray masses and connecting pathways in the depths of the brain. Prus (’48) coined the term in 1898 to describe experimentally demonstrated path- ways which conducted messages from the cortex to the spinal cord after sections of the pyramids. Spatz (’22) showed that 6 nuclei, viz., the globns pallidus, corpus subthalamicum, nu- cleus ruber, substantia nigra, nucleus dentatus and oliva inferior contained free iron, demonstrable on sections of the fresh human brain by simple, if sometimes malodorous meth- ods (H2S!). W e look nowadays upon the extrapyramidal system as composed of these nuclei as wells as of the “neostri- atum,” the cerebellum, the zona incerta and the reticular substance. The functional role of the last has been brilliantly analyzed in recent years by Magoun and his coworkers (see Magoun and Rhines, ’47), and its connections with the spinal cord were demonstrated by Papez (’26) ; the cerebellum has been brought nearer to our understanding by Snider and his associates (see Snider and Woolsey, ’41). To look upon the extrapyramidal system as a “ servo-mechanism” for the corti- cal control of the final common pathways becomes quite fashionable (see e.g., Bonin, ’50, or Ward, ’50).
Intensity of muscular contraction can be regulated only hp the number of motor units - or of motoneurons - activated
‘Since the original i s difficult of access, we cite the recent translation by Kennard.
427

428 UERHARDT VON BONIN AND GHOUSE A. SHARIFF
at a given moment, and it is reasonable to suppose that the same principle applies to those nervous structures which control the motoneurons in their turn (see Rosenblueth, Wiener and Bigelow, ’43). If we assume further that cell density varies in the same sense and to the same degree in all parts of the brain, it follows that the size of the extra- pyramidal nuclei should be a linear function of the size of the whole brain among all animals in which these nuclei have the same functional role. The brain of such animals could be considered as “models” of each other in the sense in which this term was defined for a theory of similitude by Bonin ( ’46). Of course, we know that matters are not always as simple as that. The nucleus ruber changes considerably in its composition and its connections as we go through the mammals (see e.g., Winkler, ’29 and Hatschek, ’07), and Jelgersma (’34) stated that the inferior olive in aquatic mammals consists alomost entirely of what is called the acces- sory olive in man. No such “evolutionary” changes have been demonstrated, however, for the striatum, i.e., the caudate nucleus + putamen, and the pallidum (we use the nomencla- ture of Foix and Nicolesco, ’25), for the zona incerta, for the subthalamic body or for the substantia nigra. The previous work by Harman ( ’43) and by Harman and Carpenter ( ’50) makes it clear that a linear function may here be expected.
It seemed to promise some sidel’ights on the extrapyramidal system, therefore, if these nuclei were compared quantita- tively in a number of mammals.
MATERIAL AND METHODS
We have used brains of the opossum, of the bat, rabbit, dog, Tarsius, Hapale, Macaque, Chimpanzee and man. We owe several brains of bats to the kindness of Dr. R. L. Webb, Head of the Department of Anatomy, Indiana University, Bloomington, Indiana. The brains of the opossum, and of the primates with the exception of man were put at our disposal through the kindness of Dr. Percival Bailey of the

429 EXTRAPYRAMIDAL NUCLEI
Department of Neurology and Neurological Surgery, and others were prepared by G. A. S. in the Department of Anat- omy of the University of Illinois. Serial sections were used. Outlines were drawn by means of a projector at a suitable magnification (between 2 x and 10 X) and measured by means of a planimeter. The volume was computed by multi- plying the mean of two adjacent pbnes by the thickness of the section, adding up all values and correcting for magni- fication. This differs from the procedure of Harman and Carpenter ('50)' published only after our work had been completed, perhaps explaining in part the different values which were obtained. No allowance for shrinkage was made. I n the case of the caudate nucleus neither the nucleus ac- cumbens nor the amygdaloid complex were included. It was felt that some check was advisable, hence we computed the volume of some nuclei which appear to have a very regular outline, such as the subthalamic nucleus of Luys by assuming it to be a reguhr ellipsoid. Let a, b and c be the three axes of such an ellipsoid, then the volume V is given by the formula:
Several authars, Fore1 (1877), Henle (1879), Kolliker (1896), Dejerine ( 'O l ) , Sano ( 'lo), and Whittier and Mettler ( '49) have given values of a, b and c for the body of Luys; those for the zona incerta have been determined frow our preparations.
The two determinations are compared in detail in table 1. They agree with each other within lo%, and the mean error for all 10 species is less than 1% in both cases.
RESULTS
Figures 1, 2 and 3 show on schematic sagittal sections the structures which were investigated. They were drawn from actual sections, but were then adjusted to show the areas in a way best suited to exhibit the relative size of the ganglia.

430 GERHARDT VON BONIN AND GHOUSE A. SHARIFF
TABLE 1
Coniparison of the volumes, in nwP, obtained b y planimeter ( p ) and by the formula, v (f) 6
CORPUS LUPSlI ZONA INCERTA
100(p-f) f 1XP-f) P P _ _ ~ f
P P
Man Chimpanzee Macaque Marmoset Tarsius Bat
Dog Rabbit Rat Opossum
157.7 150.5 + 4.6 56.0 54.0 + 3.5 13.1 13.4 - 2.3 1.20 1.15 + 4 1.10 .87 + 9 .054 .051 + 5.6 8.20 8.54 -4.5 1.90 2.10 - 10.0 .loo .lo7 -7 .50 .54 -8
57.6 63.3 - 10 51.5 55.3 - 7 15.8 14.6 + 7.6 1.50 1.45 + 3.4 1.70 1.53 + 10
.070 .065 + 7 24.1 26.4 - 9.5 5.10 5.60 - 10 1.13 1.113 + 1 2.50 2.53 - 1
-
RAT
Key to figures 1.2.3. mcahte andPutsmm ~ Z W tnrerto
H h l l t Q U m Sutstantia mQra
CWPUS subMDbmIcum (LuySl
Fig. 1 Schematic sagittal sections through the brainstenis of some lower mammals, indicating size and position of the basal ganglia.

EXTRAPYRAMIDAL NUCLEI 431
This did not require major distortions, a slight flattening or bulging proved sufficient in every case.
The (lissencephalic) brains shown in figure 1 all have a relatively small pallidum, and a remarkably large zona in- certa. The subthalamic body of the bat is larger than any of the other three. Is this corre1,ated with the importance of its upper extremity for locomotion? The position of the anterior commissure is remarkable for its inconstancy. Completely
MACAOUE
Fig. 2 achematic sagittal sections through the brainstems of n carnivore (dog) and some primates.
ventral in the rabbit, it is within the striatum in the bat, and still more dorsal in the rat. The case of the opossum, with its lack of corpus callosum, is, of course, exceptional. All the brains a re almost “straight,” i.e., the axis of the forebrain and the axis of the brain stem are practically parallel to each other. With Spatz (’36, see esp. pp. 478-479) we can refer to these lines as the axis of Fore1 and Meynert, resp. The pallidum, the substantia nigra and the subthalamic body of Luys a re f a r distant from each other.

432 GERHARDT VON BONIN AND G H O U S E A. S H A R I F F
Figure 2 carries the story from the dog to the primates. Perhaps the most noticeable trend is the increase in size of the substantia nigra, accompanied, so it would seem, by a similar increase in the size of the pallidum. Again, the an- terior commissure shows a very varying position. Pallidum and substantia nigra are still as far distant from each other in the dog as they were in the “lower mammals” of figure 1. I n the primates, on the other hand, where Forel’s and Mey- nert’s axes are no longer parallel to each other, the two masses are just opposite each other, on the two banks of the
CHIMPANZEE
Fig. 3 Schematic sagittal sections through the brainstems of chimpanzee and man.
stream of fibers that run from the internal capsule to the pes pedunculi.
In the chimpanzee and man, shown in figure 3, the decreas- ing angle made by the axis of Fore1 with the axis of Meynert has brought about a further shift between substantia nigra and pallidum. The location of the anterior commissure in man corresponds to that shown on page 71 in Jelgersma’s Atlas (Jelgersma, ’31).
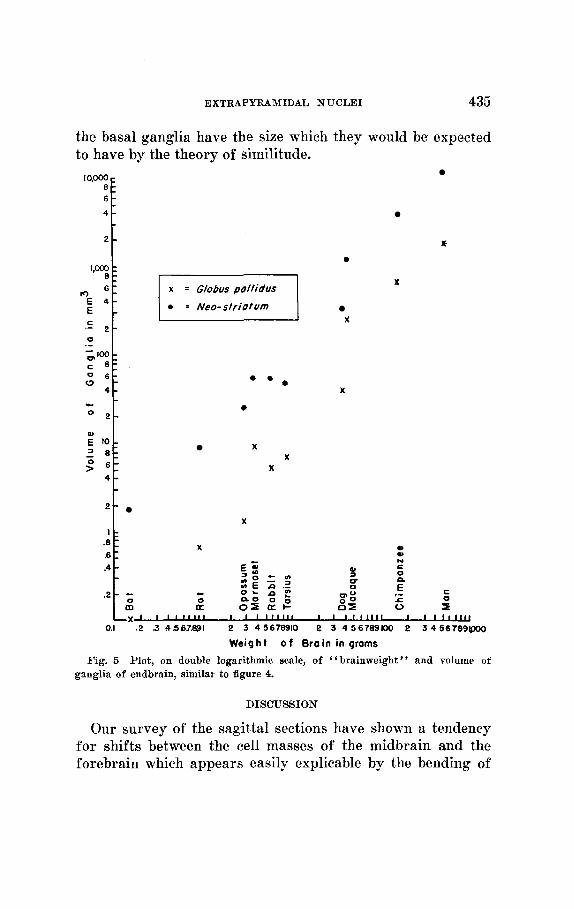
After this rapid topographical survey we turn to the nu- merical data. They are given in table 2. Those for the neostriatum and pallidum are p1,otted on a double logarithmic scale in figure 5, those f o r the zona incerta, the subthalamic body of Luys and the snbstantia nigra are plotted, in the

EXTRAPYRAMIDAL NUCLEI 433
same way, in figure 4. The double logarithmic scale will; as is well known, depict as a straight line the formula of relative growth x = a y b where x and y are two variables, such as brain weight and volume of a nucleus and a and b are two constants. I ts applicability to phylogenetic problems such as the subject of this paper was explained and justified by Huxley ( '31), Bonin ('46) and others.
The discrepancies between our data and those of Harman and Carpenter ( '50) may be partly explicable by the different
TABLE 2
Volumes in rnm3 of basal ganglia of one side and the weights of one-half of the brains
NUCLEUS GLOBUS c08pu~ ZONA BRAIN- CAUDALTUS PALLIDUS INCERTA sgg:y WEIQHT
vm Man 4612 5514.2 1778.2 157.7 57.6 678.5 545' Chimpanzee 1357.1 2274.7 637.8 56 51.5 198.5 172
Marmoset 30.8 20.4 7.4 1.2 1.5 4.3 3.8 Tarsius 26.4 18.6 6.8 1.1 1.7 3.9 9.5 Bat caudate putamen
Dog 259.4 43.8 35.3 8.2 24.1 16.7 39 Rabbit 38.3 18.2 5.4 1.9 5.1 4.5 6.1 Rat caudate putamen
Opossum 16.3 7.0 1.3 .5 2.5 2.1 3.1
Macaque 581.7 632.7 240.2 13.1 15.8 52.2 45
complex 1.8 .1 .054 .07 .16 .16
complex 8.9 .67 .1 1.13 .57 .9
l Weight of brain used, after futation.
methods used to determine the volumes, and may partly be due to the vagaries of random sampling. We know nothing about the variability of the size of the basal ganglia. From a remark by Talairach et al. ( '49) it appears that the volume of the thalamus varies (in psychotic patients) between 8.82 and 15.6 em3 (loc. cit., p. 9). I n any event, it is well to bear in mind that our values for man are lower, and for the chim- panzee and the macaque appreciably higher than those of Harman and Carpenter.

434 aERHARDT VON BONIN AND GHOUSE A. SHABIFF
1,000 8 - 6 -
4 -
2 -
loo rr) 8 -
E 4 -
c .- .- 0 - rn g 10:
5 -
rn
5 0
0 > 1.0
The pallidum exhibits a somewhat curious behavior: the points for the primates are consistently higher than those for the non-primates. The points for the “neostriatum,” on the other hand, fall all into line. I n figure 4, the substantia nigra appears to follow very well a straight regression line
c
- -
6f
2 -
8 : 6 1 4 -
2 -
- -
0.1
.08-
Subsfanfia Nigra t Zona hcerta 0
Corpus Luysii
+ 0 Weight o f broin in g r a m s
0 0 - -
c
0
P 0
+ e
0 + 8
$ 0 0
0 0
in all mammals included in our survey, while the zona incerta of man is definitely smaller than would have been predicted from the rest of the mammals. The corpus Luysii is again remarkably true to type for the majority of mammal’s, but is relatively small in the rat and in Tarsius. By and large, then,
EXTRAPYRAMIDAL N U C L E I 435
10.000 8 - 6 -
4 -
2 -
‘,m8
m 6 :
C
0
E 4 - E -
2 - .-
.- - l3’W c 8 : 0 6 - w -
4 - - O 2 -
; to-
>” 6 1 1 8 : :
4 -
2 -
.8 .6 .4
.2
c
E 0
: - x
X
::
X * * .
0
X X
X
X 1: I 0)
0) N E 0
X I - 0)
U a - E Z
c 0
2 = g - u)
Z E z.2 0
0 0 0 g ,- -g z c 0 - a 0
m a oI at- B s 0 I - -
0
L x 1 1 I ‘ l l l l I I 1 1 1 1 1 1 1 I I I I 1 1 1 1 1 I I I I I l l U
Weight o f B r a i n in grams 0.1 .2 3 4567.891 2 3 45678910 2 3 456789100 2 34567891poo
Fig. 5 Plot, on double logarithmic scale, of “brainweight” and volume of ganglia of endbrain, similar to figure 4.
DISCUSSION
Our survey of the sagittal sections have shown a tendency for shifts between the cell masses of the midbrain and the forebrain which appears easily explicable by the bending of

436 GERHARDT VON BONIN AND GHOUSE A. SH-4RIFF
the forebrain against the brainstem which is well known to occur in primates. I ts effect on the skull, viz., the shift of the foramen magnum, or the change in the pituitary angle has been the subject of numerous investigatjons. The close ap- proach of substantia nigra and pallidum have led Spatz (’21) to assume a diencephalic origin of the latter. While our observations cannot be decisive, evidence from lower mammals would render it unnecessary to consider the pal- lidum as a part of tlie diencephalon. As Rose (’42) pointed out, the boundary between diencephalon an’d telencephalon is arbitrary at best.
The numerical data justify us to consider several extra- pyramidal nuclei, namely the striatum, the body of Luys and the substantia nigra, a s “ functionally isomorpEiic,” if that, expression be permitted, in all mammals. The pallidum on the other hand tends to be larger in primates than in lower mammals and there is in man a reduction of the small zona incerta. An imposing nucleus in the rat (see Gurdjian, ’27). it has shrunk to the insignificance symbolized by its name in man. Is it a part of the facilitatory reticular substance and has it merely become more diffuse in man? Or has its function been taken over by the small-celled part of the red iiucleus? We have to end with these queries, for a merely quantitative study cannot solve the mysteries of the brain, it can only help to formulate problems.
SUMMARY
In the brain clf the opossum, bat, rat, rabbit, dog, Tarsius, Hapale, Macaque, Chimpanzee and of man the volumes of striatum and pallidum. of zona incerta, corpus subthala- micum of Luys and substantia nigra were determined in serial sections. Parasagittal schematic drawings of thew brains were prepared. There is a shift of the relative position of these masses as the forehrain axis becomes inclined against the axis of the brainstem.
The numerical data suggest that the liumari brain is a fairly faithful “model” (Ronin, ’46) of the primate braiii

EXTRAPYRAMIDAL NUCLEI 437
W I T it not for an extraordinary reduction in the size of the zona iiiccrta arid a relative increase in the size of the pallidurn. This latter feature is common to all primates investigated.
L I T E R A T U R E CITED
ROSIN, G . v. . - - 1950 Eswy on the verebral cortex. Rpringfield, Illinois. C. C
I)EJERINE, J. 1901 Amitomie des centrrs nerreaux. Par is , J. Ruff. 2 vols. Foln, n~., AND J. N i c o m s r o 1925 Les noyaux gris ccntraux et In rCgion
niCscnceplialo-sous-optiqne. Paris , Masson et Cie. 581 pp. FORM,, A . 1877 Untersuchungen iiber die H n u l m r e g i o n und ihre oberen
Verkni ipfungm im Oehirne dea Nenschen und einiger SPiigethiere, mit RcitrHgen zu den Methoden der Oel~irnitntersuc~h~ttt~. Arch. f. Psychiat. it . Nwvenkr., 7: 394-495.
GVRDJIAN, E. S. 1027 The dienrephalon of the alliino rat. J. Comp. Neur., I S : 1-111.
I I i R X A S , P. 1943 \'olumes of hasal ganglia and cortex in mammals. Proc.
H A R ~ J A X , P., AND M. H. CAR,PFXTER 1950 Volumetric comparison of thr b n ~ n l g:tnglia of the various primates including ittan. J. Conip. Neur., 9 3 .- 1 5 1 3 7.
~IITS(.HI.:K, H . 1907 Zur rergleichenden Anatontie de5 Nucleus ruher. Arb. 3. d. neur. Inst. a. d. Wien Univ., 75: 89-136.
HESLE. J. 1879 Ilaudburh der Xervenlehre drs Metlsrltrn. Rraunschweig, F. Vicweg. u. Sohn. 3 vols.
l I i . s L m , J. 1931 Problems of relative. growth. New Pork. Lincoln MarVesglt, 2 i 6 pp.
, J E I . G E R S J I . ~ , G. 1931 Atlas anntomicuni cerehri Iiutnani. Amsterdam. Scheltenia & Holkema. 108 Pp.
- - 1934 Das Geltirn der WIssersIugetirre. L e i p i g , J. A. Rartlt. 238 pp.
K ~ ~ J , I . I K L X , A. 1896 Handhwl i der Gewebelehre des JIenschen. Lcipzig, W. Engelniann. 3 vols.
J I A ( ; o c N , H. W., A N D R. RHIXES 194;1 Spasticity. Springfield, Illinois. C. C? Tlioinas, 59 pp.
PAPEZ, J. W. 19% Reticulospinal t rac ts in he eat. Marcahi niethod. 6 . Conrp. Neur., 41 : 365-399.
PKI:S, .T. 1948 On the patliwsys and patliogeneaia of cortical epilepsy. (trans]. by M. Kennard.) Res. Publ. Assn. nerv. nirnt. Uis.. 27: 129-145.
Ross, F:. -1. 1942 The ontogenetie development of the rahbi t diencrphalon. J. Comp. Neur., 77: 61-106.
HOSENBLIIETITII, A., X. WIENER AND J . BIOELOW 1913 Behavior, purpose and telroloyy. I'hilos. of Scienc-e, 1 0 : 7 9-24.
1946 Types ant1 similitudes. Philos. of Scienc-e, 1 3 : 196-20.
TliomaB, I50 pp.
S O C . h J H ' T . R i d . Med., 54 .' 297-298.

438 GERHARDT YON BONIN AND GROUSE A. SHARIFF
SANO, T. 1910 Beitrag zur vergleichenden Anatomie der substantia nigra, des corpus Luysii und der zona incerta. Monatschr. f . Psychiat. and Neur., 873 110-127.
Extensor rigidity in eats produced by simultaneous ablation of the anterior lobe of the cerebellum and the pericrueiate areas of the cerebral hemisphercs. Am. J. Physiol., 133: 154.
SPATZ, H. 1921 Zur Anatomie der Zentren des Streifenhiigels. Munch. Med. Wochschr., 68 3 1441-1446.
Uber den Eisennachweis im Gehirn, besonders in Zentren des extrapyramidal-motorisehen Systems. Z. f. d. ges. New. u. Psychiat.,
1936 Anatomie des Mittelhirns. In : Handbuch der Neurologie, ed. 0. Bumke und 0. Foerster, Bd., 1: 474-540. Berlin, J. Springer.
TALAIRACH, J., H. HECAHN, M. DAVID, M. MONNIER AND J. DE AJURIAWEIRRA 1949 Recherches sur la coagulation thbrapeutiqne des structnres sous- eortieales chez I’homme. Rev. Neurologique, 81 : 4-24.
WARD, A. A., JR., 1950 Somatic functions of the nervous system. Annual Re- view of Physiology, 421-444.
WHITTIER, 5. R., AND F. A. MEITLER Studies on the subthalamus of the rhesus monkey. I. Anatomy and fiber connections of the subthalamic nucleus of Luys. J. Comp. Neur., 90: 281-317.
WINKLER, C. 1929 Anatomie du s y s t h e nerveux. Haarlem, de erven Bohu. 5 vols.
SNPDU, R. S., ANTI C. N. WOOLSEY 1941
1922
773 263-381.
1949