exploring third metacarpal capitate facet shape in …pages.nycep.org/ed/download/pdf/nmg...
TRANSCRIPT

Exploring Third Metacarpal CapitateFacet Shape in Early Hominins
THOMAS R. REIN* AND KATERINA HARVATIDepartment of Early Prehistory and Quaternary Ecology, Paleoanthropology Section,
Senckenberg Center for Human Evolution and Paleoecology, Eberhard Karls Universit€atTubingen, Rumelinstr. 23, 72070 Tubingen, Germany
ABSTRACTThe joint between the capitate and third metacarpal plays an impor-
tant role in stabilizing the manus during hand use in great apes andhumans. Researchers have examined the morphology of this region inhumans, our fossil relatives, and other extant primates to try to under-stand the importance of this joint in human evolution. The first goal ofour research was to explore shape variation of the third metacarpal capi-tate facet across extant anthropoids, including hominoids, cercopithecoids,and platyrrhines. This analysis allowed us to examine the range of varia-tion in the capitate facet and the degree to which locomotor behavior,phylogeny, and size explained shape variation. We also examined capitatefacet shape in the early hominin fossil record in order to explore how theshape of this articular surface has changed during early hominin evolu-tion. We captured six landmark coordinates on the edge of the capitatefacet in extant anthropoids and fossil specimens to quantify and visualizeshape variation in this region. We used principal components analysis,Procrustes distances, and multivariate regression analysis to investigatedifferent possible influences on shape variation. We found that shape var-iation corresponded to function, phylogeny, and size. With the exceptionof brachiation, shape variation did not clearly correspond with anyspecific locomotor behavior. However, we identified a shift in the relativemediolateral breadth of the capitate facet during early hominin evolution,which is most likely one of several adaptations for a more stable jointsurface. Anat Rec, 296:240–249, 2013. VC 2012 Wiley Periodicals, Inc.
Key words: geometric morphometrics; carpometacarpal joint;manus
INTRODUCTION
The central joint complex (CJC) described by Lovejoyet al. (2009) provides key evidence for the evolution of thehominin hand. This complex includes articulationsbetween the trapezoid, capitate, and the second and third(Mc3) metacarpals (Lovejoy et al., 2009). The CJC offersimportant information regarding the degree of wristmobility or stability during limb activity, and studies ofthis anatomical region have played an important role ininferring hominin hand use (e.g., Lewis, 1973, 1977;Marzke, 1983; McHenry, 1983; Marzke and Marzke,1987, 2000; Ricklan, 1987; Panger et al., 2002; Young,2003; Tocheri et al., 2008). Furthermore, researchershave examined aspects of the CJC in modern humansand earlier hominins in an attempt to infer the behav-ioral and morphological character states of the last
Grant sponsor: National Science Foundation; Grant numbers:BCS-0849204, IGERT 0333415; Grant sponsor: DeutscherAkademischer Austausch Dienst; Grant number: A0981106;Grant sponsor: L.S.B. Leakey Foundation; Grant sponsor:Wenner-Gren Foundation; Grant sponsor: New York University.
*Correspondence to: Thomas R. Rein, PhD, Department ofEarly Prehistory and Quaternary Ecology, PaleoanthropologySection, Senckenberg Center for Human Evolution andPaleoecology, Eberhard Karls Universit€at Tubingen,Rumelinstr. 23, 72070 Tubingen, Germany. Fax: þ49 (0) 7071/29-5717. E-mail: [email protected]
Received 24 October 2011; Accepted 22 October 2012.
DOI 10.1002/ar.22635Published online 10 December 2012 in Wiley Online Library(wileyonlinelibrary.com).
THE ANATOMICAL RECORD 296:240–249 (2013)
VVC 2012 WILEY PERIODICALS, INC.

common ancestor of humans and chimpanzees (e.g.,Richmond et al., 2001; Orr, 2005; Tocheri et al., 2008;Lovejoy et al., 2009). Within the CJC, articular surfacemorphology of the Mc3-capitate joint is unique in humanscompared to other extant hominoids, and differencesbetween humans and other hominoids have beenhypothesized to correspond with functional differences inhand use, grip capabilities, and wrist stability (Lewis,1977; Marzke and Marzke, 1987; Young, 2003).
Unlike great apes, human Mc3 bases do not have deepgrooves that house large carpometacarpal ligaments.Pongo, Gorilla, and Pan are characterized by large inter-osseus carpometacarpal ligaments on either side of thethird metacarpal. These ligaments require expandedgrooves that separate the dorsal and palmar aspects of theintermetacarpal facets on each side of the third metacar-pal base (Lewis, 1977; Marzke, 1983; Lovejoy et al., 2009).Additionally, the joint surfaces of the human capitate andthe Mc3 are relatively flat compared to great apes. Alongwith the large interosseous ligaments, the uneven jointsurfaces of the capitate and Mc3 observed in extant greatapes act as a locking mechanism that reduces axial rota-tion and sliding at the joint (Marzke and Marzke, 1987;Lovejoy et al., 2009). Modern humans, however, have theirown skeletal adaptations for stability at the Mc3-capitatejoint. Homo is the only extant hominoid with a styloid pro-cess on the dorsal aspect of the third metacarpal base thatarticulates with the capitate, not allowing any movementat the Mc3-capitate joint (Marzke and Marzke, 1987;Marzke and Marzke, 2000). Sarmiento (1994, 1998) hasreported that this feature is found in some gorilla metacar-pals as well (however, see Marzke and Marzke [2000] for adifferent view on this morphology in African apes). On thebasis of biomechanical and comparative analyses, Marzkeand Marzke (1987, 2000) proposed that the dorsal positionof a styloid process, coupled with the palmar attachmentof the pisometacarpal III ligament, might stabilize thehuman third metacarpal from forces acting on the palmaraspect of the Mc3 distal end. Marzke and Marzke (1987,2000) suggested that the evolution of this morphologycould be due to repeated use of hard hammer percussionduring tool manufacture. In accordance with this hypothe-sis, Williams et al.’s (2012) recent study of forces acting onthe hand during Oldowan tool manufacture found that thesecond and third digits experienced higher joint reactionforces during swing and strike phases of tool productionthan the first digit. Ricklan (1987) suggested a differentexplanation for the appearance of a styloid process duringhuman evolution in which its presence offers joint stabilityin humans by creating a larger attachment site for liga-ments that limit dorsal and palmar movement at thecarpometacarpal joint. In either case, Mc3-capitate jointstability appears to be an important aspect of the humanCJC, but the adaptations allowing increased joint stabilitydiffer from those of great apes.
Homo is not the only hominoid genus that differs fromextant great apes in Mc3 base morphology. Early homi-nins, including Ardipithecus and Australopithecus, aresimilar to Homo in lacking the deep ligament grooves onthe radial and ulnar side of the third metacarpal base(Lovejoy et al., 2009). Humans also share this aspect oftheir CJC morphology with an inferred arboreal quadru-ped, Proconsul, which lacks the suspensory and knuckle-walking specializations in its hand skeleton that
characterize extant great apes (e.g., Rose, 1994; Begunet al., 1994; Ward, 1998; Lovejoy et al., 2009; Rein et al.,2011). In addition, Homo sapiens and Australopithecusafarensis share relatively flat and palmarly narrow capi-tate facets to the exclusion of extant great apes (Bushet al., 1982; Drapeau et al., 2005, Lovejoy et al., 2009).The palmar end of this joint surface in two Australopi-thecus afarensis specimens, A.L. 438-1d and A.L. 333w-6, is relatively narrower in the mediolateral plane thanit is in modern humans, whereas other specimens havea palmar end that is wider than that of humans but stillnarrower than great apes (Drapeau et al., 2005). Inmany ways, early hominins (such as Australopithecusafarensis) and modern humans share a capitate facetmorphology that distinguishes them from extant apes.
Beyond these similarities, there are also differences inproximal articular surface morphology between modernhumans and early hominins such as Australopithecusafarensis. For example, Australopithecus afarensis lacksa styloid process on the third metacarpal, a trait thatmost likely evolved in the human lineage to enhance sta-bility at the carpometacarpal joint (Bush et al., 1982;Marzke, 1983; McHenry, 1983; Marzke and Marzke,1987, 2000; Drapeau et al., 2005). However, researchershave reported the presence of short or slight styloid proc-esses on the third metacarpal bases of Australopithecusafricanus (Stw 64) and the unidentified SKX 3646 speci-men (Ricklan, 1987; Susman, 1989). Although the styloidprocess morphology observed in these early homininsfrom South Africa is not equivalent to that of modernhumans, it might signal a shift during early homininevolution to a Mc3-capitate joint that is more adapted forjoint stability and to oppose forces acting on the thirddigit. We were interested in exploring early homininproximal Mc3 shape and examining whether or not therewere other changes to the capitate facet that mightcorrespond to enhanced stability of this joint.
The first goal of our research was to explore shape var-iation in the capitate facet across extant anthropoids,including hominoids, cercopithecoids, and platyrrhines.This analysis allowed us to examine the range of varia-tion in this joint surface and the degree to whichlocomotor behavior, phylogeny, and size explained shapevariation. We expected both locomotor function and phy-logeny to be significantly related to capitate facet shapesince closely related taxa perform similar locomotorbehaviors in many cases. Next, we incorporated measure-ments on the capitate facets of early hominins, includingAustralopithecus afarensis, Australopithecus africanus,Australopithecus sediba, and an unidentified homininmetacarpal from Member 2 of the Swartkrans Formation.We explored how the shape of the capitate facet haschanged during early hominin evolution and we testedthe following hypotheses:
1. Shape variation in the capitate facet will provideboth a functional and phylogenetic signal.a. There will be significant correspondence between
published proportions of locomotor behaviors(knuckle-walking, vertical climbing, brachiation,quadrupedalism, and overall substrate usage) andshape of the capitate facet border in extantanthropoids.
b. Furthermore, there will be a significant relationshipbetween phylogeny and articular surface shape.
HOMININ THIRD METACARPAL CAPITATE FACET SHAPE 241

2. Mc3 capitate facet shape changed during early homi-nin evolution.a. Australopithecus afarensis will be characterized by
a capitate facet shape that is different from laterhominins in South Africa.
MATERIALS AND METHODS
We measured the proximal articular surface of 203third metacarpals representing extant anthropoids, Aus-tralopithecus afarensis, Australopithecus africanus,Australopithecus sediba, and SKX 3646 (Table 1). Meas-urements were collected on extant specimens frommuseums and research institutions in the United States,Europe, and East Africa (see Rein [2010] for a completelist of museum collections). We attempted to measure anequal number of male and female adult specimenswhenever possible. In addition, we measured the Mc3from the left manus when available. If the left was notavailable, we measured the right side.
Locomotor proportion data were based on severalpublished sources (Table 2). We chose five locomotor cat-egories to examine, including palmigrade/digitigradequadrupedalism (QUAD), knuckle-walking (KW), bra-chiation (BRAC), vertical climbing (VC), and overallsubstrate usage (SUB), and assigned taxa values basedon published reports of the proportion of time speciesspend performing different behaviors. Palmigrade anddigitigrade hand postures used during quadrupedalismwere grouped into the same category since recent stud-ies have shown variation in use of these hand postureswithin a species, and metacarpal traits do not alwaysclearly distinguish between ‘‘palmigrade’’ and ‘‘digiti-grade’’ quadrupeds (Patel, 2009, 2010; Patel and Polk,2010). Extant taxa were not assigned to a single locomo-tor category. Instead, we assigned values for eachlocomotor category to species in order to account for the
diverse locomotor repertoires of these taxa. While thesesix categories are quite general and do not account forsubtle differences between species, they are useful sincethey allow the incorporation of a larger number ofspecies into the analysis for which published locomotordata are reported using these general categories.
One of us (TRR) collected three-dimensional coordi-nate data with a portable Microscribe digitizer on allspecimens representing extant taxa and Australopithe-cus afarensis. TRR also collected landmark coordinatedata on scans of fossil metacarpals using the EVAN Tool-box, including those of the original specimens ofAustralopithecus africanus (Stw 64 and Stw 68) andSKX 3646. A surface scan of a cast of UW 88-116 (Aus-tralopithecus sediba third metacarpal described byKivell et al. [2011]) from the American Museum of Natu-ral History was measured as well. The Stw 64, Stw 68,and the cast of UW 88-116 fossil specimens werescanned with a NextEngine desktop laser scanner. Asurface model of the SKX 3646 specimen was createdfrom CT scan data. We chose six landmarks to representvariation in capitate facet shape (Fig. 1; Table 3). To vis-ualize the shape recorded by our landmark set, we builta wireframe model in MorphoJ (version 1.05a; Klingen-berg, 2011). Each specimen that was microscribed wasplaced in plasticine in a position that allowed all land-marks to be captured at one time.
To examine intraobserver error, a modern humanthird metacarpal was digitized ten times by TRR with aMicroscribe digitizer and another ten times with theEvan Toolbox over separate 2 week periods. A surfacemodel of the metacarpal was made by TRR using a Nex-tEngine desktop laser scanner and ScanStudio HD Pro.We assessed the impact of measurement error on ourresults by comparing Procrustes distances calculated inthe Evan Toolbox between repeated measurements ofthe same specimen to those between 20 individuals fromthe Hamann-Todd Collection. To test for measurementerror due to the use of different measurement
TABLE 1. Sample sizes of extant and fossilspecimensa
Taxon Male Female Indeterminate Total
Homo sapiensb 10 10 9 29Pan troglodytes 16 7 0 23Gorilla gorilla 10 10 0 20Pongo pygmaeus 9 9 0 18Hylobates lar 10 10 0 20Papio anubis 13 5 1 19Macaca fascicularis 9 9 0 18Cercopithecus mitis 8 11 0 19Cebus apella 9 10 0 19Ateles geoffroyi 3 7 0 10Australopithecus afarensis – – 4 4Australopithecus africanus – – 2 2Australopithecus sediba – – 1 1SKX 3646c – – 1 1
aAustralopithecus afarensis specimens include A.L. 438–1d,A.L. 333w-6, A.L. 333–16, A.L. 333–65. Australopithecusafricanus specimens include Stw 64 and Stw 68. Australopi-thecus sediba specimen measured in this study was a castof UW 88–116.bIsolated human metacarpal specimens from the skeletalcollections at the University of Tubingen did not have infor-mation regarding sex of the individual.cThird metacarpal proximal base from Swartkrans Member2 not identified to a specific taxon.
TABLE 2. Locomotor proportion data used in cova-riation analysesa
Taxon QUAD KW BRAC VC SUBb
Homo sapiens 0.0 0.0 0.0 0.0 99.0Pan troglodytes 2.0 87.0 1.0 5.0 90.0Gorilla gorilla 2.0 91.0 1.0 1.0 95.0Pongo pygmaeus 12.0 0.0 10.0 25.0 1.0Hylobates lar 0.0 0.0 50.0 17.0 0.0Papio anubis 98.0 0.0 0.0 1.0 77.0Macaca fascicularis 68.0 0.0 0.0 7.0 20.0Cercopithecus mitis 54.0 0.0 0.0 3.0 5.0Cebus apella 54.0 0.0 0.0 10.0 5.0Ateles geoffroyi 24.0 0.0 25.0 7.0 0.0
aLocomotor categories include palmigrade/digitigrade quad-rupedalism (QUAD), knuckle-walking (KW), brachiation(BRAC), vertical climbing (VC), and substrate usage (SUB).Brachiation includes specialized suspensory behaviorsdescribed by Hunt et al. (1996). Locomotor proportions arebased on those reported in Gebo (1996), Youlatos (1998),Manfreda et al. (2006), and Wright (2007). Substrate usageproportions are based on Fleagle (1980), Crockett and Wil-son (1980), Hunt (1989), Cant (1987), Fontaine (1990), Geboand Chapman (1995), Youlatos (1998), and Carlson (2005).bHigher proportion values for overall substrate usage corre-spond to greater amounts of time moving on the ground.
242 REIN AND HARVATI

techniques, we compared Procrustes distances betweenrepeated measurements with a Microscribe and repeatedmeasurements with the Evan Toolbox to those betweenthe 20 individuals from the Hamann-Todd Collection.This method has been used in previous studies to inves-tigate the effect of measurement error on results(Lockwood et al., 2002; Noback et al., 2011). Procrustesdistances were calculated from the Procrustes coordinatedata for all six landmarks.
We superimposed the three-dimensional coordinates ofthe six capitate facet landmarks using generalized Pro-crustes analysis (GPA; Rohlf, 1990; O’Higgins and Jones,1998) in MorphoJ (Klingenberg, 2011). Generalized Pro-crustes analysis allows exploration of shape variationacross different specimens by removing the effects of
translation and rotation and scaling objects to a commoncentroid size (O’Higgins and Jones, 1998). Once GPA hasbeen performed, the resulting Procrustes coordinatescan be used in further statistical analyses.
Most analyses were performed in MorphoJ (Klingen-berg, 2011). Principal components analysis of thecovariance matrix of tangent projected coordinates wasperformed to explore capitate facet shape variationacross extant and extinct specimens (O’Higgins andCollard, 2002; Klingenberg, 2011). We also computedProcrustes distances between the mean taxon shapes forboth extant and fossil specimens using the EvanToolbox. We did this in order to examine whether specieswere most similar due to function or phylogeny and toexamine which species most closely resemble the fossilspecimens.
We also performed multivariate regression analysis inMorphoJ to examine the relationship between shapevariation and different variables (Frost et al., 2003; Tur-ley et al., 2011). The Procrustes coordinates were usedas dependent variables and locomotor data, log centroidsize, and phylogeny were employed as independent vari-ables. Centroid size is the square root of the sum ofsquared distances from each landmark coordinate in ashape configuration to their mean position (Slice, 2007).This is the value used to scale specimens during general-ized Procrustes analysis. It is important to examineallometry (i.e., the effect of size on shape) in this type offunctional analysis since many times differences in bodysize can account for differences in shape (Manfredaet al., 2006). In addition, phylogenetic history can influ-ence skeletal shape. Therefore, we assigned species toeither the Catarrhini or Platyrrhini taxonomic groupusing dummy variables similar to Frost et al. (2003) tosee if shared ancestry corresponded with Mc3 capitatefacet shape. Next, we calculated the proportion of thetotal variance that was explained by each independentvariable in MorphoJ. We then examined the relationshipbetween the independent variables by calculating theangles among the shape vectors for each variable inMorphoJ as the arccosine of the vector dot products (seeTurley et al. [2011]). A parallel angle (0 degrees) repre-sents correlations, whereas perpendicular angles (90degrees) show independence between the vectors.Finally, we visualized how these different variables cor-responded with shape variation by adding andsubtracting the regression coefficients to the consensusshape (see Turley et al. [2011]) and comparing wireframemodels in MorphoJ.
RESULTS
Results from the error test are shown in Figure 2.Three of the 190 Procrustes distances between individu-als of the inter-individual sample (1.6%) overlapped withProcrustes distances between repeated measurementstaken on the same metacarpal. The results showed mini-mal overlap, demonstrating that variation accounted forby intraobserver error and the use of a Microscribe ver-sus data collection from a surface scan was relativelysmall compared to the level of variation between individ-uals. The level of variation across the entire sample ofspecimens included in the study is much greater thanthe level of variation within just the modern humansample tested here. Thus, measurement error accounts
Fig. 1. Landmarks collected on capitate facet shown on human leftthird metacarpal.
TABLE 3. Definitions of six capitate facet landmarksused in this study
Landmark Definition Type
1 Most medial point on thedorsal border of the capitate facet
3
2 Most lateral point on the medialborder of the capitate facet
3
between the dorsal and the palmaraspects of the metacarpal 4 facet
3 Most medial point on the palmarborder of the capitate facet
3
4 Most lateral point on the palmarborder of the capitate facet
3
5 Most medial point on the lateralborder of the capitate facet
3
between the dorsal and the palmar aspectsof the metacarpal 2 facet
6 Most lateral point on the dorsal borderof the capitate facet
3
HOMININ THIRD METACARPAL CAPITATE FACET SHAPE 243

for a relatively small proportion of the total variationacross the interspecies sample.
We first explored shape variation among extantanthropoids using principal components analysis(Fig. 3). As shown in Figure 3, taxa cluster along thefirst two principal components of shape variation in bothphylogenetic and functional groups. Quadrupedal OldWorld monkeys and Cebus apella were characterized bypositive values along the first principal component(PC1), whereas the overwhelming majority of knuckle-walking African apes, suspensory orangutans, andbipedal humans had negative PC1 scores. The greatapes and humans were characterized by mediolaterallyexpanded capitate facets compared to highly quadrupe-dal cercopithecines and platyrrhines. In addition, thelateral aspect of the palmar border of the capitate facetextended farther in the palmar direction compared tothe medial aspect in great apes and humans. The medialand lateral aspects of the palmar border extendedequally in the palmar direction in quadrupedal monkeys.The two brachiating taxa, gibbons and spider monkeys,clustered along the second principal component based ona shared medial constriction of the lateral border of thecapitate facet not observed to the same extent in anyother taxon.
We next examined Procrustes distances between meantaxon shape configurations in order to quantify the over-all similarities and differences between taxa (Table 4).We will only discuss results for the extant taxa in thissection. In most cases, the lowest Procrustes distances
Fig. 2. Analysis of measurement error. Black columns representfrequency of Procrustes distances between interindividual humansample from Hamann-Todd Collection (20 individuals). The other col-umns represent frequency distribution of distances between repeatedmeasurements on a single specimen by TRR with a Microscribedigitizer and using the Evan Toolbox and surface scan data.
Fig. 3. Scatterplot of principal component scores one and two for shape variation of the capitate facetin extant anthropoids. The image on the right shows how capitate facet shape deviates from the samplemean shape along each principal component.
244 REIN AND HARVATI
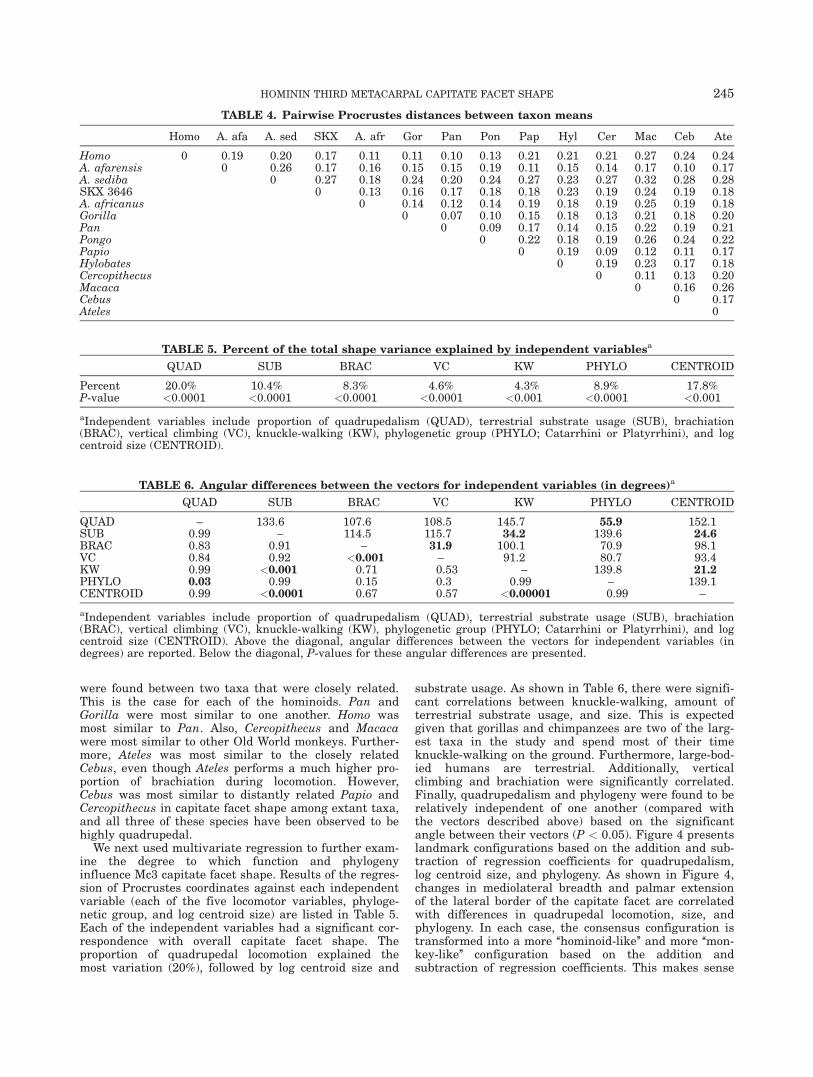
were found between two taxa that were closely related.This is the case for each of the hominoids. Pan andGorilla were most similar to one another. Homo wasmost similar to Pan. Also, Cercopithecus and Macacawere most similar to other Old World monkeys. Further-more, Ateles was most similar to the closely relatedCebus, even though Ateles performs a much higher pro-portion of brachiation during locomotion. However,Cebus was most similar to distantly related Papio andCercopithecus in capitate facet shape among extant taxa,and all three of these species have been observed to behighly quadrupedal.
We next used multivariate regression to further exam-ine the degree to which function and phylogenyinfluence Mc3 capitate facet shape. Results of the regres-sion of Procrustes coordinates against each independentvariable (each of the five locomotor variables, phyloge-netic group, and log centroid size) are listed in Table 5.Each of the independent variables had a significant cor-respondence with overall capitate facet shape. Theproportion of quadrupedal locomotion explained themost variation (20%), followed by log centroid size and
substrate usage. As shown in Table 6, there were signifi-cant correlations between knuckle-walking, amount ofterrestrial substrate usage, and size. This is expectedgiven that gorillas and chimpanzees are two of the larg-est taxa in the study and spend most of their timeknuckle-walking on the ground. Furthermore, large-bod-ied humans are terrestrial. Additionally, verticalclimbing and brachiation were significantly correlated.Finally, quadrupedalism and phylogeny were found to berelatively independent of one another (compared withthe vectors described above) based on the significantangle between their vectors (P < 0.05). Figure 4 presentslandmark configurations based on the addition and sub-traction of regression coefficients for quadrupedalism,log centroid size, and phylogeny. As shown in Figure 4,changes in mediolateral breadth and palmar extensionof the lateral border of the capitate facet are correlatedwith differences in quadrupedal locomotion, size, andphylogeny. In each case, the consensus configuration istransformed into a more ‘‘hominoid-like’’ and more ‘‘mon-key-like’’ configuration based on the addition andsubtraction of regression coefficients. This makes sense
TABLE 4. Pairwise Procrustes distances between taxon means
Homo A. afa A. sed SKX A. afr Gor Pan Pon Pap Hyl Cer Mac Ceb Ate
Homo 0 0.19 0.20 0.17 0.11 0.11 0.10 0.13 0.21 0.21 0.21 0.27 0.24 0.24A. afarensis 0 0.26 0.17 0.16 0.15 0.15 0.19 0.11 0.15 0.14 0.17 0.10 0.17A. sediba 0 0.27 0.18 0.24 0.20 0.24 0.27 0.23 0.27 0.32 0.28 0.28SKX 3646 0 0.13 0.16 0.17 0.18 0.18 0.23 0.19 0.24 0.19 0.18A. africanus 0 0.14 0.12 0.14 0.19 0.18 0.19 0.25 0.19 0.18Gorilla 0 0.07 0.10 0.15 0.18 0.13 0.21 0.18 0.20Pan 0 0.09 0.17 0.14 0.15 0.22 0.19 0.21Pongo 0 0.22 0.18 0.19 0.26 0.24 0.22Papio 0 0.19 0.09 0.12 0.11 0.17Hylobates 0 0.19 0.23 0.17 0.18Cercopithecus 0 0.11 0.13 0.20Macaca 0 0.16 0.26Cebus 0 0.17Ateles 0
TABLE 5. Percent of the total shape variance explained by independent variablesa
QUAD SUB BRAC VC KW PHYLO CENTROID
Percent 20.0% 10.4% 8.3% 4.6% 4.3% 8.9% 17.8%P-value <0.0001 <0.0001 <0.0001 <0.0001 <0.001 <0.0001 <0.001
aIndependent variables include proportion of quadrupedalism (QUAD), terrestrial substrate usage (SUB), brachiation(BRAC), vertical climbing (VC), knuckle-walking (KW), phylogenetic group (PHYLO; Catarrhini or Platyrrhini), and logcentroid size (CENTROID).
TABLE 6. Angular differences between the vectors for independent variables (in degrees)a
QUAD SUB BRAC VC KW PHYLO CENTROID
QUAD – 133.6 107.6 108.5 145.7 55.9 152.1SUB 0.99 – 114.5 115.7 34.2 139.6 24.6BRAC 0.83 0.91 – 31.9 100.1 70.9 98.1VC 0.84 0.92 <0.001 – 91.2 80.7 93.4KW 0.99 <0.001 0.71 0.53 – 139.8 21.2PHYLO 0.03 0.99 0.15 0.3 0.99 – 139.1CENTROID 0.99 <0.0001 0.67 0.57 <0.00001 0.99 –
aIndependent variables include proportion of quadrupedalism (QUAD), terrestrial substrate usage (SUB), brachiation(BRAC), vertical climbing (VC), knuckle-walking (KW), phylogenetic group (PHYLO; Catarrhini or Platyrrhini), and logcentroid size (CENTROID). Above the diagonal, angular differences between the vectors for independent variables (indegrees) are reported. Below the diagonal, P-values for these angular differences are presented.
HOMININ THIRD METACARPAL CAPITATE FACET SHAPE 245

since these different phylogenetic groups vary in termsof body size and locomotor behavior. Thus, it is difficultto tease apart the degree to which variation in mediolat-eral breadth of the capitate facet is caused by any one ofthese factors based on extant taxa alone. However, asimilar analysis demonstrated a small but significantbrachiation signal in overall capitate facet shape. Asshown in Figure 5, brachiators are characterized byenhanced medial constriction on the lateral border of thecapitate facet. This is similar to the shape reflected bythe second principal component shown in Figure 3, inwhich gibbons and spider monkeys group together to theexclusion of other primates.
Next, we added the early hominin fossils to the analy-sis. Based on the principal components analysis and theProcrustes distances between mean taxon configurations(Fig. 6; Table 4), Australopithecus afarensis was mostsimilar to quadrupedal primates, including Cebus andPapio and characterized by a mediolaterally narrowcapitate facet. Even when the A.L. 333w-6 specimen(characterized by the most narrow facet) was removedfrom the analysis, the Australopithecus afarensis meanconfiguration remained closest to Cebus and Papio (Pro-crustes distance ¼ 0.12) With regard to Procrustesdistances, Australopithecus africanus was found to bemost similar to modern humans. The capitate facetshapes of Australopithecus sediba and the SKX 3646 fos-sil were closest to those of the South AfricanAustralopithecus africanus. The first principal compo-nent axis on Figure 6 shows Australopithecus afarensisspecimens grouping along the right side of the graphwith Old World and New World monkeys. The Stw 64
(Australopithecus africanus) and SKX 3646 specimensare found between modern humans and Australopithe-cus afarensis along this PC axis, and Stw 68(Australopithecus africanus) and UW 88-116 (Australopi-thecus sediba) fall within the modern human range.This principal component represents variation in overallmediolateral breadth of the capitate facet, with modernhumans, great apes, and early hominins from SouthAfrica being characterized by relatively broader capitatefacets than Australopithecus afarensis and quadrupedalmonkeys (Fig. 7). Thus, there is a shift to increasedmediolateral breadth early in hominin evolution.
DISCUSSION
We tested two hypotheses regarding shape variationin the capitate facet. The first hypothesis stated thatcapitate facet shape would correspond to both functionand phylogeny. Our results from the principal compo-nents analysis, Procrustes distances, and multivariateregression support this hypothesis. In each case, bothlocomotor function and evolutionary relationships corre-sponded with shape variation. Furthermore, size wassignificantly correlated with variation in capitate facetshape. These results illustrate the difficulty in teasingapart phylogenetic, functional, and allometric signals inskeletal morphology. Hominoids are generally largerthan other anthropoids, do not perform high proportionsof palmigrade/digitigrade quadrupedalism, and areclosely related to one another. Thus, when great apesand humans group together (as they did in our analysis)to the exclusion of Old World monkeys and platyrrhines,it is difficult to tease apart the different potential signalsfrom the skeletal anatomy. We did find, however, someindications of a functional signal in this morphology.Gibbons and spider monkeys grouped together along thesecond principal component of shape variation, and theirshared capitate facet morphology (i.e., medial constric-tion of the lateral facet border) could correspond withthe high proportions of brachiation performed by eachtaxon. Similarities in capitate facet shape betweenhumans and great apes might be related to an underly-ing functional demand for joint stability during handuse. Perhaps increased mediolateral breadth of thecapitate facet allows enhanced joint stability in these
Fig. 4. Visualization of Mc3 capitate facet shape variation as afunction of three independent variables: quadrupedalism (QUAD), logcentroid size, and phylogeny. We estimated these landmark configura-tions by adding and subtracting regression coefficients to the consen-sus configuration. The quadrupedalism and phylogeny images weremagnified (x5) so that shape changes could be seen more easily.
Fig. 5. Visualization of Mc3 capitate facet shape variation as afunction of brachiation. We estimated these landmark configurationsby adding and subtracting regression coefficients to the consensusconfiguration. The images were magnified (x5) so that shape changescould be seen more easily.
246 REIN AND HARVATI

taxa. Increased Mc3- capitate joint breadth might be anadaptation to the biomechanical demands of locomotionin orangutans, chimpanzees, and gorillas, whereas itmay have provided joint stability for behaviors such astool manufacture in the human lineage. However, wecannot test for the effects of tool manufacture on Mc3morphology in this study.
Our second hypothesis postulated a change in Mc3capitate facet shape during early human evolution.Australopithecus afarensis most closely resembled NewWorld and Old World monkeys in overall capitate facetshape, whereas early hominins from South Africa eithermost closely resembled other hominins from the same
region or modern humans. All of the South Africanfossils were characterized by a relatively greater capi-tate facet mediolateral breadth than Australopithecusafarensis. Furthermore, some of these fossils have beenreported to have a slight styloid process on the proximalthird metacarpal base, a feature missing in the Australo-pithecus afarensis fossils and suggested to enhancestability at the Mc3-capitate carpometacarpal joint inhumans (Marzke and Marzke, 1987, 2000; Ricklan,1987; Susman, 1989). The similarity between Australopi-thecus afarensis and extant monkeys in this morphologycould relate to enhanced joint mobility in each taxon.Marzke and Marzke (1987) described how modern
Fig. 6. Scatterplot of principal component scores one and two for shape variation of the capitate facetin extant and extinct species. The image on the right shows how capitate facet shape deviates from thesample mean shape along each principal component. Individual fossil specimens are labeled on thegraph. The convex hull for modern humans is drawn in gray.
Fig. 7. Mean capitate facet shapes of Homo sapiens, Australopithecus africanus, Australopithecus afar-ensis, and Cebus apella. Individual specimens represented include SKX 3646 and UW 88-116 (Australopi-thecus sediba).
HOMININ THIRD METACARPAL CAPITATE FACET SHAPE 247

humans differ from Old World monkeys in the degree ofmobility at the third carpometacarpal joint. There isbasically no movement at the joint in humans, whereasOld World monkey third metacarpals are able to flexand extend to a greater degree (Marzke and Marzke,1987). It is therefore possible that Australopithecus afar-ensis had greater mobility at the Mc3-capitate joint thanlater hominins.
On the basis of our results, there was a shift in capi-tate facet shape quite early in hominin evolution. Theincrease in mediolateral breadth most likely assisted inincreasing stability of the carpometacarpal joint duringhominin evolution. Whether or not this was an adapta-tion to stone tool manufacture or tool use is beyond thescope of this paper but is an interesting avenue forfuture research. Finally, humans are not unique amongextant primates in the relatively high mediolateralbreadth of the capitate facet. Our species is similar toAfrican apes and orangutans in this aspect of shapevariation. However, based on the differences in capitatefacet morphology observed in early hominins such asArdipithecus ramidus (Lovejoy et al., 2009) and Austral-opithecus afarensis, it is possible that we do not sharethis trait with extant apes due to common ancestry butrather due to a common need for joint stability. It wouldbe interesting to incorporate earlier hominin taxa (e.g.,Ardipithecus ramidus) and Miocene hominoids intofuture geometric morphometric analyses to furtherexplore variation in capitate facet shape and enhanceour understanding of the polarity of this trait.
We have found that the shape of the Mc3 capitatefacet corresponds with both locomotor function and phy-logeny, and differences in capitate facet shape may bedue to varying needs for stability or mobility of the cen-tral joint complex. There was change in capitate facetshape during early hominin evolution, in which earlyhominins from South Africa were characterized by abroader capitate facet than Australopithecus afarensisfrom East Africa. The relatively broad capitate facetmorphology that characterizes great apes and modernhumans today may be due to a shared need for stabilityat the Mc3-capitate joint during hand use.
ACKNOWLEDGEMENTS
The authors greatly appreciate the helpful feedback andcomments of the anonymous reviewer and the Journaleditors. They would like to thank the Authority forResearch and Conservation of Cultural Heritage, theNational Museum of Ethiopia, Mamitu Yilma, andWilliam Kimbel for permission to study the Australopi-thecus afarensis specimens. Thanks to Ian Tattersall,Eric Delson, Gisselle Garcia, and Melissa Tallman of theAmerican Museum of Natural History for providing uswith a surface scan of the Australopithecus sediba cast.Thanks to Jason Hemingway, Bernhard Zipfel, and theInstitute for Human Evolution, University of the Witwa-tersrand for providing us access to surface scans of theAustralopithecus africanus specimens. They would alsolike to thank Tracy Kivell, Stephany Potze, the DitsongNational Museum of Natural History, and the Depart-ment of Human Evolution, Max Planck Institute forEvolutionary Anthropology for access to the SKX 3646CT scan surface model. The extant skeletal specimensexamined in this study were generously provided by
William Stanley (FMNH), Yohannes Haile-Selassie andLyman Jellema (HTC), Judith Chupasko (HMCZ),Marcia Ponce de Le�on (AIM), Linda Gordon (NMNH),Malcolm Harman (PCM), Paula Jenkins and LouiseTomsett (NHML), Frieder Mayer (Humboldt), EmmanuelGilissen and Wim Wendelen (RMCA), Emma Mbua andFredrick Kyalo Manthi (NMK), and Eileen Westwig(AMNH). This is NYCEP Morphometrics contributionnumber 74.
LITERATURE CITED
Begun DR, Teaford MF, Walker A. 1994. Comparative and func-tional anatomy of Proconsul phalanges from Kaswanga PrimateSite, Rusinga Island, Kenya. J Hum Evol 26:89–165.
Bush ME, Lovejoy CO, Johanson DC, Coppens Y. 1982. Hominidcarpal, metacarpal, and phalangeal bones recovered from theHadar Formation: 1974–1977 collections. Am J Phys Anthropol57:651–677.
Cant JGH. 1987. Positional behavior of female Bornean orangutans(Pongo pygmaeus). Am J Primatol 12:71–90.
Carlson KJ. 2005. Investigating the form-function interface inAfrican apes: relationships between principal moments of areaand positional behaviors in femoral and humeral diaphyses. Am JPhys Anthropol 127:312–334.
Crockett CM, Wilson WL. 1980. The ecological separation of Macacanemestrina and Macaca fascicularis in Sumatra. In: LindburgDG, editor. The macaques: studies in ecology, behaviour andevolution. New York: Van Nostrand Reinhold Company. p148–181.
Drapeau MSM, Ward CV, Kimbel WH, Johanson DC, Rak Y. 2005.Associated cranial and forelimb remains attributed to Australopi-thecus afarensis from Hadar, Ethiopia. J Hum Evol 48:593–642.
Fleagle JG. 1980. Locomotion and posture. In: Chivers DJ, editor.Malayan forest primates: ten years’ study in tropical rain forests.New York: Plenum Press. p191–207.
Fontaine R. 1990. Positional behavior in Saimiri boliviensis andAteles geoffroyi. Am J Phys Anthropol 82:485–508.
Frost SR, Marcus LF, Bookstein FL, Reddy DP, Delson E. 2003.Cranial allometry, phylogeography, and systematics of large-bod-ied papionins (Primates: Cercopithecinae) inferred from geometricmorphometric analysis of landmark data. Anat Rec 275A:1048–1072.
Gebo DL. 1996. Climbing, brachiation, and terrestrial quadrupedal-ism: historical precursors of hominid bipedalism. Am J PhysAnthropol 101:55–92.
Gebo DL, Chapman CA. 1995. Positional behavior in five sympatricOld World monkeys. Am J Phys Anthropol 97:49–76.
Hunt KD. 1989. Positional behavior in Pan troglodytes at theMahale Mountains and the Gombe Stream National Parks, Tan-zania. Ph.D. Dissertation: University of Michigan.
Hunt KD, Cant JGH, Gebo DL, Rose MD, Walker SE, Youlatos D.1996. Standardized descriptions of primate locomotor and pos-tural modes. Primates 37:363–387.
Kivell TL, Kibii JM, Churchill SE, Schmid P, Berger LR. 2011. Aus-tralopithecus sediba hand demonstrates mosaic evolution of loco-motor and manipulative abilities. Science 333:1411–1417.
Klingenberg CP. 2011. MorphoJ: an integrated software package forgeometric morphometrics. Mol Ecol Res 11:353–357.
Lewis OJ. 1973. The hominoid os capitatum, with special referenceto the fossil bones from Sterkfontein and Olduvai Gorge. J HumEvol 2:1–11.
Lewis OJ. 1977. Joint remodelling and the evolution of the humanhand. J Anat 123:157–201.
Lockwood CA, Lynch JM, Kimbel WH. 2002. Quantifying temporalbone morphology of great apes and humans: an approach usinggeometric morphometrics. J Anat 201:447–464.
Lovejoy CO, Simpson SW, White TD, Asfaw B, Suwa G. 2009.Careful climbing in the Miocene: the forelimbs of Ardipithecusramidus and humans are primitive. Science 326:70e1–8.
248 REIN AND HARVATI

Manfreda E, Mitteroecker P, Bookstein FL, Schaefer K. 2006. Func-tional morphology of the first cervical vertebra in humans andnonhuman primates. Anat Rec 289B:184–194.
Marzke MW. 1983. Joint functions and grips of the Australopithecusafarensis hand, with special reference to the region of the capi-tate. J Hum Evol 12:197–211.
Marzke MW, Marzke RF. 1987. The third metacarpal styloid processin humans: origin and functions. Am J Phys Anthropol 73:415–431.
Marzke MW, Marzke RF. 2000. Evolution of the human hand:approaches to acquiring, analysing and interpreting the anatomi-cal evidence. J Anat 197:121–140.
McHenry HM. 1983. The capitate of Australopithecus afarensis andA. africanus. Am J Phys Anthropol 62:187–198.
Noback ML, Harvati K, Spoor F. 2011. Climate-related varia-tion of the human nasal cavity. Am J Phys Anthropol 145:599–614.
O’Higgins P, Collard M. 2002. Sexual dimorphism and facial growthin papionin monkeys. J Zool 257:255–272.
O’Higgins P, Jones N. 1998. Facial growth in Cercocebus torquatus:an application of three dimensional geometric morphometric techni-ques to the study of morphological variation. J Anat 193:251–272.
Orr CM. 2005. Knuckle-walking anteater: a convergence test ofadaptation for purported knuckle-walking features of AfricanHominidae. Am J Phys Anthropol 128:639–658.
Panger MA, Brooks AS, Richmond BG, Wood B. 2002. Older thanthe Oldowan? Rethinking the emergence of hominin tool use.Evol Anthropol 11:235–245.
Patel BA. 2009. Not so fast: Speed effects on forelimb kinematics incercopithecine monkeys and implications for digitigrade posturesin primates. Am J Phys Anthropol 140:92–112.
Patel BA. 2010. Functional morphology of cercopithecoid primatemetacarpals. J Hum Evol 58:320–337.
Patel BA, Polk JD. 2010. Distal forelimb kinematics in Erythrocebuspatas and Papio anubis during walking and galloping. Int J Pri-matol 31:191–207.
Rein TR. 2010. Locomotor function and phylogeny: implications forinterpreting the hominoid fossil record. Ph.D. Dissertation: NewYork University.
Rein TR, Harrison T, Zollikofer CPE. 2011. Skeletal correlates ofquadrupedalism and climbing in the anthropoid forelimb: implica-tions for inferring locomotion in Miocene catarrhines. J Hum Evol61:564–574.
Richmond BG, Begun DR, Strait DS. 2001. Origin of human biped-alism: the knuckle-walking hypothesis revisited. Yearb PhysAnthropol 44:70–105.
Ricklan DE. 1987. Functional anatomy of the hand of Australopithe-cus africanus. J Hum Evol 16:643–664.
Rohlf FJ. 1990. Rotational fit (Procrustes) methods. In: Rohlf FJ,Bookstein FL, editors. Proceedings of the Michigan Morphomet-rics Workshop. Ann Arbor: University of Michigan Museum of Zo-ology. p227–236.
Rose MD. 1994. Quadrupedalism in some Miocene catarrhines. JHum Evol 26:387–411.
Sarmiento EE. 1994. Terrestrial traits in the hands and feet ofgorillas. Am Mus Novit 3091:1–56.
Sarmiento EE. 1998. Generalized quadrupeds, committed bipeds,and the shift to open habitats: an evolutionary model of hominiddivergence. Am Mus Novit 3250:1–78.
Slice DE. 2007. Geometric morphometrics. Ann Rev Anthropol 36:261–281.
Susman RL. 1989. New hominid fossils from the Swartkrans For-mation (1979–1986 excavations): postcranial specimens. Am JPhys Anthropol 79:451–474.
Tocheri MW, Orr CM, Jacofsky MC, Marzke MW. 2008. The evolu-tionary history of the hominin hand since the last common ances-tor of Pan and Homo. J Anat 212:544–562.
Turley K, Guthrie EH, Frost SR. 2011. Geometric morphometricanalysis of tibial shape and presentation among Catarrhine taxa.Anat Rec 294:217–230.
Ward CV. 1998. Afropithecus, Proconsul, and the primitive hominoidskeleton. In: Strasser E, Fleagle J, Rosenberger A, McHenry H,editors. Primate Locomotion: Recent Advances. New York: Ple-num Press. p337–352.
Williams EM, Gordon AD, Richmond BG. 2012. Hand pressure dis-tribution during Oldowan stone tool production. J Hum Evol 62:520–532.
Wright KA. 2007. The relationship between locomotor behavior andlimb morphology in brown (Cebus apella) and weeper (Cebus oli-vaceus) capuchins. Am J Primatol 69:736–756.
Young RW. 2003. Evolution of the human hand: the role of throwingand clubbing. J Anat 202:165–174.
Youlatos D. 1998. Positional behavior of two sympatric guianancapuchin monkeys, the brown capuchin (Cebus apella) andthe wedge-capped capuchin (Cebus olivaceus). Mammalia 62:351–365.
HOMININ THIRD METACARPAL CAPITATE FACET SHAPE 249