experimental infection of tilapia lake virus (tilv) in ... · challenged fish included massive...
TRANSCRIPT

Contents lists available at ScienceDirect
Veterinary Microbiology
journal homepage: www.elsevier.com/locate/vetmic
Experimental infection of Tilapia Lake Virus (TiLV) in Nile tilapia(Oreochromis niloticus) and red tilapia (Oreochromis spp.)
Puntanat Tattiyaponga,b, Worawan Dachavichitleada,b, Win Surachetponga,b,⁎
a Department of Veterinary Microbiology and Immunology, Faculty of Veterinary Medicine, Kasetsart University, Bangkok, Thailandb Center for Advanced Studies for Agriculture and Food, Kasetsart University Institute for Advanced Studies, Kasetsart University, Bangkok, Thailand
A R T I C L E I N F O
Keywords:Tilapia lake virusIsolationTilapiaKoch’s postulates
A B S T R A C T
Since 2015, a novel orthomyxo-like virus, tilapia lake virus (TiLV) has been associated with outbreaks of diseaseand massive mortality of cultured Nile and red tilapia (Oreochromis niloticus and Oreochromis spp., respectively)in Thailand. In this study, TiLV was isolated from field samples and propagated in the permissive E-11 cell line,with cytopathic effect (CPE) development within 3–5 days post-inoculation. Electron micrographs of infected E-11 cells and fish tissues confirmed the rounded, enveloped virions of 60 to 80 nm with characteristics verysimilar to those of Orthomyxoviridae. In vivo challenge studies showed that high mortality in Nile (86%) and redtilapia (66%) occurred within 4–12 days post-infection. The virus was re-isolated from challenged fish tissues inthe permissive cell line, and PCR analysis confirmed TiLV as a causative pathogen. The distinct histopathology ofchallenged fish included massive degeneration and inflammatory cell infiltration in the liver and brain as well asthe presence of eosinophilic intracytoplasmic inclusions in hepatocytes and splenic cells. Our results fulfilledKoch’s postulates and confirmed that TiLV is an etiologic agent of mass mortality of tilapia in Thailand. Theemergence of this virus in many countries has helped increase awareness that it is a potential threat to tilapiaaquacultured in Thailand, Asia, and worldwide.
1. Introduction
Tilapine cichilds comprise the second most important freshwaterfish aquacultured worldwide, with annual production totaling morethan 3.5 million tonnes (FAO, 2014a; FAO, 2014b). The main tilapiaproducers are China, Indonesia, Egypt, Brazil, the Philippines, andThailand (FAO, 2014a). In Thailand, Nile tilapia (Oreochromis niloticus)and red tilapia (Oreochromis spp.) are commonly raised in cages inrivers, exposing them to multiple pathogens including bacteria, viruses,fungi, and parasites. During 2015–2017, multiple outbreaks of farm-raised Nile and red tilapia with an unknown etiology were observed inThailand. The cumulative mortality rate were in the range 20–100%.The disease has been called “tilapia one month mortality syndrome” asit is frequently reported within one month after fry or juvenile tilapiashave been moved from hatcheries to the grow-out cages. The clinicalsigns and pathology of infected fish include anorexia, poor body con-dition, abnormal swimming, severe anemia, bilateral exophthalmia,skin erosion and congestion, scale protrusion, and abdominal swelling.Recently, an emerging tilapia orthomyxo-like virus named tilapia lakevirus (TiLV) was reported in many moribund fish in Thailand(Surachetpong et al., 2017). TiLV was first identified in farm-raised and
wild tilapia in Israel (Eyngor et al., 2014). Subsequently, the virus wasreported in Ecuador, Colombia, Egypt, and Thailand (Bacharach et al.,2016; Fathi et al., 2017; Kembou Tsofack et al., 2016; Surachetponget al., 2017). TiLV is an enveloped single-stranded RNA virus consistingof ten genomic segments. Of the ten segments, only the first segmentshares weak sequence similarity to viruses in the family Orthomyx-oviridae (Bacharach et al., 2016). In addition, the 3′ and 5′ untranslatedregion repeatable sequences, which are characteristic of the influenzavirus, have been identified in all segments of the TiLV (Bacharach et al.,2016). In 2014, the syncytial hepatitis virus (SHV) was reported inintensive farmed-raised tilapia in Ecuador (Ferguson et al., 2014). Anucleotide sequence analysis suggested that TiLV and SHV could be thesame virus (Del-Pozo et al., 2016).
Although outbreaks of TiLV infection in tilapia have been reportedin Thailand and other countries (Surachetpong et al., 2017), Koch’spostulates have not been fulfilled for this pathogen. It is important todemonstrate that a pathogen from field outbreaks can cause clinicalinfection when used to challenge healthy animals and that the virus canbe re-isolated from the experimentally infected animals. The aims ofthis study were to confirm that TiLV isolated from diseased tilapia couldcause disease and mortality in healthy tilapia after experimental
http://dx.doi.org/10.1016/j.vetmic.2017.06.014Received 31 May 2017; Received in revised form 13 June 2017; Accepted 16 June 2017
⁎ Corresponding author at: Department of Veterinary Microbiology and Immunology, Faculty of Veterinary Medicine, Kasetsart University, Bangkok, Thailand.E-mail address: [email protected] (W. Surachetpong).
Veterinary Microbiology 207 (2017) 170–177
0378-1135/ © 2017 Elsevier B.V. All rights reserved.
MARK

challenge and to develop laboratory-controlled challenge of TiLV in-fection. The identification and isolation of TiLV from clinically diseasedfish, reproduction of clinical symptoms and mortality in healthy fish,and re-isolation of the same pathogen would fulfil Koch’s postulates.
2. Materials and methods
2.1. Clinical samples
Nile tilapia (Oreochromis niloticus) and red tilapia (Oreochromis spp.)were collected from the provinces of Ang Thong, Kanchanaburi, andPathum Thani which are located in central and western Thailand(Table 1). These three separate locations (80 to 180 km apart) sufferedfrom multiple outbreaks with an unidentified etiology and a mortalityrate above 3% per day for 3–5 consecutive days. Twenty Nile and redtilapia were collected from each outbreak (a total of 15 outbreaks) toinvestigate the disease. The experimental protocols and animals usedwere approved by the Institutional Animal Care and Use Committee ofKasetsart University under permit number OACKU00659. The permitallowed the collection of clinical samples of Nile and red tilapia fromnatural outbreaks. In addition, the permit allowed the use of laboratoryanimals for virus isolation and experimental challenge.
2.2. RNA isolation and polymerase chain reaction
Total RNA was extracted from the fish brains or inoculated E-11cells using TRIzol™ reagent (Invitrogen, USA) according to the manu-facturer’s protocol. For cDNA synthesis, reverse transcription (RT) wascarried out using a Viva 2-steps RT-PCR kit (Vivantis, Malaysia) in 20 μlreaction containing 2× mastermix, 4 μM oligo(dT) primer, and 1 mMdNTP. The RT reaction was performed at 42 °C for 60 min and in-activated at 85 °C for 5 min. For PCR reaction, cDNA templates wereamplified with specific primers for TiLV and other viruses (Table 2).Each PCR mastermix contained 2 μl cDNA template, 0.2 μM of forwardand reverse primer, 2 mM MgCl2, 0.2 mM dNTP, and 0.05 U/μl Taq
DNA polymerase (Invitrogen, USA). The PCR cycling conditions weredenaturation at 95 °C for 5 min, followed by 40 cycles at 95 °C for 30 s,58 °C for 30 s, and 72 °C for 30 s, with final elongation at 72 °C for5 min using a T100 PCR thermocycler (Bio-Rad, USA). The PCR pro-ducts were separated on 2% agarose gel and stained with ethidiumbromide. The β-actin gene was used as an internal control gene to showthe quality of the genomic RNA and DNA.
2.3. Virus isolation
The virus was isolated in E-11 cells, a continuous cell line fromsnakehead fish (Ophicephalus striatus). The E-11 cells were purchasedfrom the European Collection of Authenticated Cell Cultures (ECACC),England (catalogue number 01110916) and were propagated inLiebovitz L-15 supplemented with 5% (vol/vol) fetal bovine serum(Sigma, USA) and 2 mM L-glutamine, and maintained at 25 °C withoutCO2. Briefly, brain tissues were homogenized in 10% Hank’s balancedsalt solution (HBSS) and then centrifuged at 3,000×g for 10 min. Thesupernatant was collected and filtered through a 0.22 μm filter mem-brane. Five hundred microliters of the filtered sample were inoculatedinto confluent E-11 cells in a 25 cm2
flask and maintained at 25 °C for14 days. The cells were observed daily. The supernatant was collectedand stored at −80 °C for animal challenge studies.
2.4. Challenge study
A sample of 35 Nile tilapia and red tilapia weighing 30 g± 0.52 geach were housed in the animal facility, Faculty of Veterinary Medicine,Kasetsart University. The water quality parameters of temperature,dissolved oxygen, pH, ammonia, and nitrite were monitored daily. Theanimals were monitored three times per day with all efforts made tominimize animal distress. Prior to the experimental challenge study,five fish were randomly selected to undergo screening for bacterial andviral infections, consisting of Streptococcus agalactiae, Aeromonas spp.,betanodavirus, iridovirus, and TiLV, using a bacterial culture on trypticsoy agar (TSA) and PCR analysis. To prepare for the viral challenge, fishwere sedated in 40 mg/L eugenol solution (Aquanes®, Better Pharma,Thailand). Fish in the control group were injected intraperitoneallywith supernatant from normal E-11 cells, while fish in the infectedgroup were injected with supernatant from E-11 cells infected withTiLV at a dose of 1 × 106 TCID50/fish. The TCID50 was calculatedfollowing the method of Reed and Muench (Muench, 1938). Clinicalsigns after virus inoculation were monitored daily for 14 days and thegross pathology and mortality rate were recorded daily. The decisioncriteria to euthanize animals included two or more clinical signs (poorbody condition, severe skin erosion and hemorrhage, loss of balance,extensive abdominal swelling, scale protrusion, and exophthalmia). At14 dpi, all surviving fish were euthanized using an overdose of eugenolsolution. Brains from the control and virus challenge groups were takenfor PCR examination. Cumulative mortality curves from the challengestudy were generated using the GraphPad Prism software (version5.01).
Table 1Details of clinical isolates.
Sample no. Location Collection date Clinical signs PCR test
1 Ang Thong 02/06/16 + +2 Pathum Thani 22/06/16 + +3 Pathum Thani 28/06/16 + +4 Ang Thong 01/07/16 + +5 Pathum Thani 02/08/16 + +6 Pathum Thani 05/08/16 + +7 Ang Thong 16/08/16 + +8 Ang Thong 22/08/16 + +9 Kanchanaburi 24/08/16 + +10 Pathum Thani 16/09/16 + +11 Pathum Thani 02/10/16 + +12 Pathum Thani 05/10/16 + +13 Pathum Thani 16/10/16 + +14 Ang Thong 25/11/16 + +15 Pathum Thani 26/11/16 + +
Table 2Primers used in this study.
Target name Primer name Sequence (5′-3′) Size (bp) References
Iridovirus 1-F1-R
5′-CTC-AAA-CAC-TCT-GGC-TCA-TC-3′5′-GCA-CCA-ACA-CAT-CTCeCTA-TC-3′
570 bp (Kurita et al., 1998)
Betanodavirus OPVP75R3
5′-CGT-GTC-GGT-GCT-GTG-TCG-CT-3′5′-CGA-GTC-AAC-ACG-GGT-GAA-GA-3′
421 bp (Bigarre et al., 2009)
TiLV Nested ext-2Nested ext-1
5′-TTG-CTC-TGA-GCA-AGA-GTA-CC-3′5′-TAT-GCA-GTA-CTT-TCCeCTG-CC-3′
491 bp (Eyngor et al., 2014)
β-actin β-actin _Fβ-actin _R
5′-TCC-AAT-TTA-TTG-GCC-TTC-GTT-GC-3′5′-CTT-CCA-TTT-TCT-GTG-TGA-GGG-AGG-3′
114 bp (Wang et al., 2014)
P. Tattiyapong et al. Veterinary Microbiology 207 (2017) 170–177
171

2.5. Histopathology
Tissue samples (brain, anterior kidney, liver, and spleen) from thecontrol and infected groups (three per group) were collected for his-tological study at 7 dpi. These samples were preserved in 10% (vol/vol)neutral buffered formalin. After 24 h, the formalin was replaced withfresh 10% formalin solution. Tissue sections were embedded withparaffin and stained with hematoxylin and eosin (H & E) using standardhistological procedures.
2.6. Electron microscopy
For electron microscopy studies, infected cells with CPE were har-vested at 5 dpi. Cells were scraped from culture flasks and centrifuged(3,000×g for 10 min) to separate the supernatant from the cell pellets.The cell pellets were fixed in 2.5% cacodylate buffered glutaraldehydeand post-fixed in 1% cacodylate buffered osmium tetroxide with a pH of7.2 for 2 h, and then dehydrated in ethanol and embedded in resin. Adrop of the sample was placed on a thin copper grid for 15 min andstained with 2% uranyl acetate. To analyze infected brain tissue, sam-ples were collected from infected fish and fixed in 2.5% cacodylatebuffered glutaraldehyde and post-fixed in 1% cacodylate buffered
osmium tetroxide for 45 min, and then washed three times with sodiumcacodylate buffer, dehydrated in acetone and embedded in resin. Tissuesections were cut 1 μm thick and stained with toluidine blue for ob-servation under a light microscope. The ultrathin sections weretrimmed and stained with 2% uranyl acetate. All micrographs weretaken at 80 kV using a Hitachi HT7700 transmission electron micro-scope (Hitachi, Germany) at the Scientific Equipment and ResearchDivision, Kasetsart University, Bangkok, Thailand.
3. Results
3.1. A novel, orthomyxo-like virus, TiLV was identified and isolated fromfield samples
Our previous study indicated that a novel, orthomyxo-like virus,TiLV, could be isolated from field samples associated with massivemortality of both Nile and red tilapia in Thailand. In this study, weinvestigated additional field outbreaks to further confirm TiLV as theprimary cause of tilapia deaths in Thailand (Table 1). From the 15 fieldoutbreaks sampled, TiLV could be detected in all samples with clinicalsigns of infection including distinct skin hemorrhage and erosion, headcongestion and severe anemia (Fig. 1A and B). It was likely that TiLV
Fig. 1. Detection and isolation of TiLV in clinical samples. (A) Gross signs of infected red tilapia; distinct skin redness (black arrow) and skin erosion. (B) Nile tilapia; skin erosion (whitearrow). (C) Infected fish were screened for virus infections using specific primers to TiLV, iridovirus, and betanodavirus. The β-actin was used as an internal control. Viruses were isolatedin E-11 cells. (D) E-11 cells inoculated with normal tilapia brain. (E) E-11 cells inoculated with TiLV-positive brain, CPE with cell shrinkage and syncytial formation (black arrow), 4 dpi ata magnification of 20×.
P. Tattiyapong et al. Veterinary Microbiology 207 (2017) 170–177
172
was the primary cause of such massive mortality as no other virusesincluding iridovirus and betanodavirus could be detected in the fieldsamples (Fig. 1C). Notably, comparison of the nucleotide sequence ofthe PCR products revealed 98% nucleotide sequence identity to TiLV(GenBank accession no. KY381578).
Brain tissues of infected fish were inoculated into confluent E-11cells and at 4 dpi, CPE was clearly observed in E-11 cells inoculatedwith materials prepared from TiLV-positive fish, but not from normalfish (Fig. 1D). The characteristics of infected cells included an increasein cell aggregation, cellular vacuolation, cell shrinkage, and formationof foci of detached, rounded cells (Fig. 1E). More pronounced cell de-tachment and syncytial formation were observed at 5 to 7 dpi.
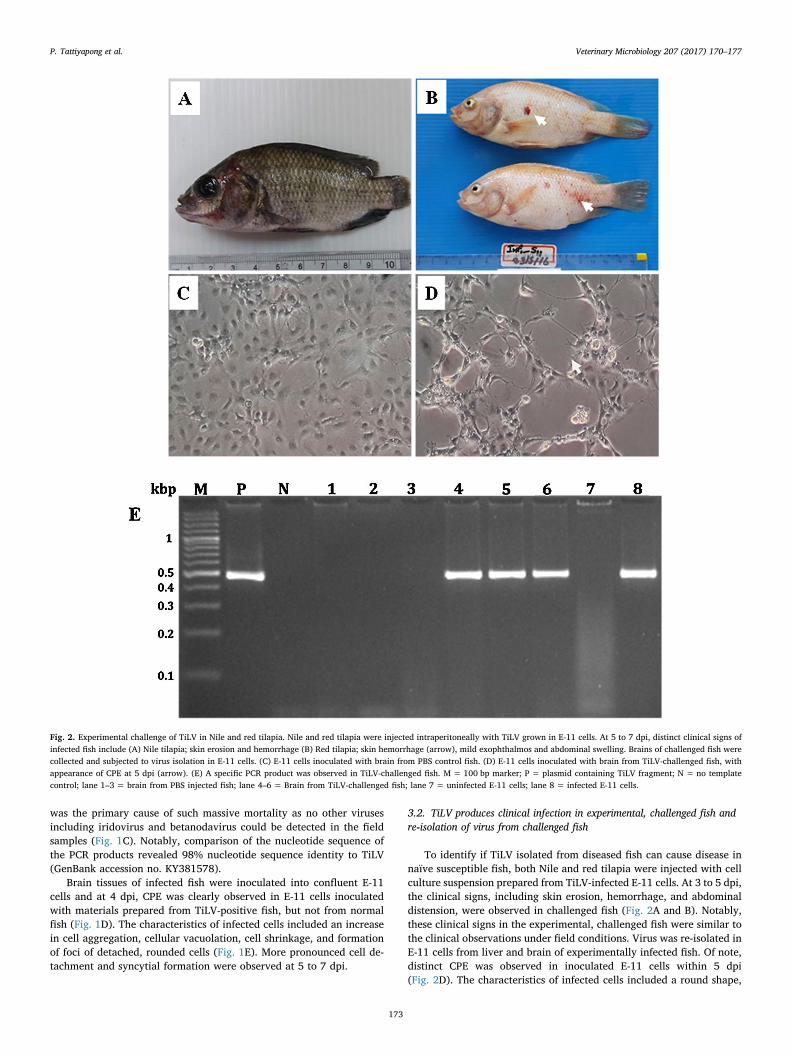
3.2. TiLV produces clinical infection in experimental, challenged fish andre-isolation of virus from challenged fish
To identify if TiLV isolated from diseased fish can cause disease innaïve susceptible fish, both Nile and red tilapia were injected with cellculture suspension prepared from TiLV-infected E-11 cells. At 3 to 5 dpi,the clinical signs, including skin erosion, hemorrhage, and abdominaldistension, were observed in challenged fish (Fig. 2A and B). Notably,these clinical signs in the experimental, challenged fish were similar tothe clinical observations under field conditions. Virus was re-isolated inE-11 cells from liver and brain of experimentally infected fish. Of note,distinct CPE was observed in inoculated E-11 cells within 5 dpi(Fig. 2D). The characteristics of infected cells included a round shape,
Fig. 2. Experimental challenge of TiLV in Nile and red tilapia. Nile and red tilapia were injected intraperitoneally with TiLV grown in E-11 cells. At 5 to 7 dpi, distinct clinical signs ofinfected fish include (A) Nile tilapia; skin erosion and hemorrhage (B) Red tilapia; skin hemorrhage (arrow), mild exophthalmos and abdominal swelling. Brains of challenged fish werecollected and subjected to virus isolation in E-11 cells. (C) E-11 cells inoculated with brain from PBS control fish. (D) E-11 cells inoculated with brain from TiLV-challenged fish, withappearance of CPE at 5 dpi (arrow). (E) A specific PCR product was observed in TiLV-challenged fish. M = 100 bp marker; P = plasmid containing TiLV fragment; N = no templatecontrol; lane 1–3 = brain from PBS injected fish; lane 4–6 = Brain from TiLV-challenged fish; lane 7 = uninfected E-11 cells; lane 8 = infected E-11 cells.
P. Tattiyapong et al. Veterinary Microbiology 207 (2017) 170–177
173

syncytial formation, and cell detachment, while E-11 cells inoculatedwith brain from the control group did not show any CPE (Fig. 2C).Three fish each from the control group and the infected group weresubjected to PCR analysis to confirm the presence of TiLV. All infectedfish showed a positive PCR product on agarose gel (Fig. 2E). In chal-lenged fish, mortality started at 4 dpi and lasted until 10 dpi with ac-cumulative mortality at 86.67% and 66.67% for Nile and red tilapia,respectively. The control fish did not show any clinical signs of infec-tion or mortality (Fig. 3)
3.3. The viral particles were present in infected E-11 cells and brains ofchallenged fish
At 5 dpi, the supernatant of infected E-11 cells and brain tissue fromTiLV-challenged fish was processed for transmission electron
microscopy. The electron micrographs of infected E-11 cell lines re-vealed single, round-shaped, enveloped viral particles (Fig. 4A and B).In the tissue sections, the infected brain tissues contained various en-veloped viral particles located in the cytoplasm of the brain cells(Figs. 4C and 4D). The viral particles were consistently round or oval,60–80 nm in diameter with a central variable electron-dense core.These virus-like particles usually appeared as a single particle or ag-gregate in small groups in the brain cells.
3.4. Distinct histopathological findings in multiple organs of challenged fish
The histological appearance of multiple organs (brain, liver, spleen,and anterior kidney) from the control fish showed normal structures(Fig. 5A, C, E, and G). In TiLV-challenged fish, the pathological findingsincluded multifocal hemorrhages with severe blood congestion and
Fig. 3. Cumulative mortality in TiLV-challenged red tilapia. Nile andred tilapia (n = 30) were injected intraperitoneally with TiLV at1 × 106 TCID50/fish. The control Nile and red tilapia were injectedwith cell culture medium. Survival rate was recorded daily until 14dpi.
Fig. 4. Transmission electron micrograph of infectedE-11 cells and ultrathin sections of infected tilapiabrain tissue. (A and B) High magnification of a freevirion showing a round enveloped viral particle with60 to 80 nm diameter. (C and D) Ultrathin section ofinfected tilapia brain showed multiple viral particlesin the cytoplasm of infected cells (C) marked withsquare box or (D) marked with black arrows. Scalebars: 0.1 μm.
P. Tattiyapong et al. Veterinary Microbiology 207 (2017) 170–177
174

proliferation of glial cells in the brain (Fig. 5B). Notably, syncytial cellformation and massive hepatocellular necrosis with pyknotic and kar-yolytic nuclei were observed in the liver cells of infected fish (Fig. 5D).Moreover, eosinophilic intracytoplasmic inclusion bodies were presentin the liver cells and splenic cells of infected fish (Fig. 5D and F).Multiple necrotic foci were observed in the anterior kidney (Fig. 5H)and increased melanomacrophage centers and dispersion of melaningranules were observed in the spleens of infected fish (Fig. 5F).
4. Discussion
Our attempts to identify the causative agent of massive losses oftilapia at multiple sites throughout Thailand led to the identification ofTiLV infection in tilapia (Surachetpong et al., 2017). Although TiLV hasbeen detected from many field outbreak samples, the reproduction ofdisease and re-isolation of the causative pathogen from challenged fishis an important step to confirm the etiology of disease. In this study, we
Fig. 5. Histopathology in TiLV-challengedtilapia. Section of normal fish tissues: (A)Brain, (C) Liver, (E) Spleen, (G) Anteriorkidney. In infected fish, the pathologicalfindings include (B) Multifocal hemorrhageswith severe blood congestion in the brain,optic tectum (OT), stratum periventriculare(StP). (D) Extensive hepatocellular necrosiswith eosinophilic intracytoplasmic inclusionbody in liver cells (higher magnification).(F) Increased melanomacrophage center(MMC) in the spleen (arrows) with eosino-philic intracytoplasmic inclusion body(arrow head, higher magnification). (H)Multifocal area of necrosis and in-flammatory cells infiltration in the anteriorkidney.
P. Tattiyapong et al. Veterinary Microbiology 207 (2017) 170–177
175

fulfilled Koch’s postulates for TiLV using molecular techniques, histo-pathology, electron microscopy, and a challenge experiment: (i) thevirus was isolated from clinically infected fish but not from normal fish;(ii) inoculation of materials prepared from cell cultures obtained fromisolated, diseased fish resulted in CPE formation; (iii) electron micro-graphs of virus particles from the brains of infected fish revealed round,enveloped virions; (iv) injection of cultured supernatant from infectedE-11 cells into naïve fish reproduced clinical symptoms similar to fieldspecimens; and (v) the virus was re-isolated and detected in challengedfish. Our data indicated that TiLV is sufficient to produce clinical dis-ease and mortality in Nile and red tilapia. In addition to TiLV, otheremerging viruses and bacteria including iridovirus and betanodavirushave been previously identified in diseased tilapia in Thailand(Keawcharoen et al., 2015; Suebsing et al., 2016). However, neither ofthese viruses were detected in any of our clinical specimens. In addi-tion, we did not find other emerging bacterial pathogens such asFrancisella spp. in field samples using PCR analysis (Jantrakajorn andWongtavatchai, 2016; Soto et al., 2009) (data not shown). TiLV is asingle stranded, negative-sense RNA virus containing 10 genomic seg-ments (Bacharach et al., 2016). In addition to TiLV, infectious salmonanemia virus (ISAV) and rainbow trout orthomyxovirus (RbtOV) areviruses in the Orthomyxoviridae family that cause disease in fish (Battset al., 2017; Merour et al., 2011).
To our knowledge, this newly identified virus is very contagious inNile and red tilapia, making it a great threat to tilapia aquaculturedworldwide. Global awareness of this problem must be raised. Previousreported indicated that only tilapia cichlids are highly susceptible toTiLV infection (Eyngor et al., 2014). Thus, it is worthwhile to in-vestigate the infection in other piscine species or to identify the naturalpotential sources of this virus. Our results also suggest that red hybridtilapia (Oreochromis spp.) is susceptible to TiLV infection. Although themortality rates in both Nile and red tilapia were relatively high underlaboratory challenge, our observations from the field outbreaks sug-gested that the mortality rate in red tilapia is 30–100%, while themortality rate of Nile tilapia is below 30%. Other factors in the naturalenvironment may impact clinical outcomes of infection and the chal-lenge route through intraperitoneal injection, which is not the naturalmeans of infection by this virus which may impact on the outcome.Indeed, cohabitation of susceptible fish with infected fish may providean alternative strategy for a TiLV challenge study (Eyngor et al., 2014).It is probable that different genetic backgrounds of Nile and red tilapiamay also influence susceptibility to TiLV as well as concomitant in-fection with other bacteria and viruses. For example, cross-breedingdisease-resistant lines of Nile tilapia with those that are susceptibleproduced more hybrids that were disease-resistant against A. hydrophila(Sarder et al., 2001). Moreover, red tilapia may express different pat-terns of preferred receptors for TiLV. In Atlantic salmon, the tissuedistribution of 4-O-acetylated sialic acid is important for virus entry andcell tropism of ISAV (Aamelfot et al., 2012). All of these researchquestions await further investigation.
The viral morphology of TiLV isolated in Thailand has a close re-semblance to that of viruses in the Orthomyxoviridae family (Fontanaand Steven, 2015; Weli et al., 2013). In our infected tilapia, free andaggregate virions with or without envelopes were observed in thebrains of diseased fish. Previous observation revealed that similar viralparticles of SHV were found in the liver cells of diseased tilapia inEcuador (Ferguson et al., 2014). It has been emphasized that TiLVisolated from tilapia in Israel produces histopathology in the brain(Eyngor et al., 2014), while the closely related SHV isolated fromEcuador mainly causes lesions in the liver (Del-Pozo et al., 2016;Ferguson et al., 2014). For other piscine viruses, distinct genotypes mayresult in different pathological outcomes of infection and infectiontissues, such as in the case of infectious salmon anemia virus (ISAV)(Aamelfot et al., 2014; Godoy et al., 2014). Nevertheless, the differentgenotypes and tissue tropisms of this emerging virus require additionalstudy. In our experiment, extensive histopathological changes were
observed in both liver and brain tissues of fish after experimentalchallenge including severe syncytial hepatitis and meningoencephalitis.Our current study and previous reports indicated that both liver andbrain samples could be used for TiLV detection using RT-PCR or virusisolation in the cell cultures (Kembou Tsofack et al., 2016;Surachetpong et al., 2017).
In summary, the results clearly showed that TiLV isolated inThailand can cause disease in challenged Nile and red tilapia. Our studyprovided evidence that fulfills Koch’s postulates regarding TiLV infec-tion in tilapia. The importance of tilapia culture worldwide highlightsthat effective control measures, biosecurity programs, and the devel-opment of vaccines should be implemented to reduce the impact of thisemerging viral disease.
Acknowledgements
This work was supported by the Center for Advanced Studies forAgriculture and Food, Institute for Advanced Studies, KasetsartUniversity, Bangkok, Thailand Under the Higher Education ResearchPromotion and National Research University Project of Thailand, Officeof the Higher Education Commission, Ministry of Education, Thailand.
References
Aamelfot, M., Dale, O.B., Weli, S.C., Koppang, E.O., Falk, K., 2012. Expression of theinfectious salmon anemia virus receptor on atlantic salmon endothelial cells corre-lates with the cell tropism of the virus. J. Virol. 86, 10571–10578.
Aamelfot, M., Dale, O.B., Falk, K., 2014. Infectious salmon anaemia – pathogenesis andtropism. J. Fish Dis. 37, 291–307.
Bacharach, E., Mishra, N., Briese, T., Zody, M.C., Kembou Tsofack, J.E., Zamostiano, R.,Berkowitz, A., Ng, J., Nitido, A., Corvelo, A., Toussaint, N.C., Abel Nielsen, S.C.,Hornig, M., Del Pozo, J., Bloom, T., Ferguson, H., Eldar, A., Lipkin, W.I., 2016.Characterization of a novel orthomyxo-like virus causing mass die-Offs of tilapia.MBio 7, e00431–00416.
Batts, W.N., LaPatra, S.E., Katona, R., Leis, E., Ng, T.F., Brieuc, M.S., Breyta, R.B., Purcell,M.K., Conway, C.M., Waltzek, T.B., Delwart, E., Winton, J.R., 2017. Molecularcharacterization of a novel orthomyxovirus from rainbow and steelhead trout(Oncorhynchus mykiss). Virus Res. 230, 38–49.
Bigarre, L., Cabon, J., Baud, M., Heimann, M., Body, A., Lieffrig, F., Castric, J., 2009.Outbreak of betanodavirus infection in tilapia Oreochromis niloticus (L.), in freshwater. J. Fish Dis. 32, 667–673.
Del-Pozo, J., Mishra, N., Kabuusu, R., Cheetham, S., Eldar, A., Bacharach, E., Lipkin, W.I.,Ferguson, H.W., 2016. Syncytial hepatitis of tilapia (Oreochromis niloticus L.) isassociated with orthomyxovirus-Like virions in hepatocytes. Vet. Pathol.
Eyngor, M., Zamostiano, R., Kembou Tsofack, J.E., Berkowitz, A., Bercovier, H., Tinman,S., Lev, M., Hurvitz, A., Galeotti, M., Bacharach, E., Eldar, A., 2014. Identification of anovel RNA virus lethal to tilapia. J. Clin. Microbiol. 52, 4137–4146.
FAO, 2014a. Cultured aquatic species information programme, Oreochromis niliticus(Linnaeus, 1758). In on Food and Agriculture Organization of the United Nation,Rome, Italy. http://www.fao.org/fishery/culturedspecies/Oreochromis_niloticus/en.
FAO, 2014b. In: The State of World Fisheries and Aquaculture, on Food and AgricultureOrganization of the United Nation. Rome, Italy.
Fathi, M., Dickson, C., Dickson, M., Leschen, W., Baily, J., Muir, F., Ulrich, K., Weidmann,M., 2017. Identification of Tilapia Lake Virus in Egypt in Nile tilapia affected by‘summer mortality’ syndrome. Aquaculture 473, 430–432.
Ferguson, H.W., Kabuusu, R., Beltran, S., Reyes, E., Lince, J.A., del Pozo, J., 2014.Syncytial hepatitis of farmed tilapia, Oreochromis niloticus (L.): a case report. J. FishDis. 37, 583–589.
Fontana, J., Steven, A.C., 2015. Influenza virus-mediated membrane fusion: structuralinsights from electron microscopy. Arch. Biochem. Biophys. 581, 86–97.
Godoy, M.G., Suarez, R., Lazo, E.S., Llegues, K.O., Kibenge, M.J., Wang, Y., Kibenge, F.S.,2014. Genetic analysis and comparative virulence of infectious salmon anemia virus(ISAV) types HPR7a and HPR7b from recent field outbreaks in Chile. Virol. J. 11,204.
Jantrakajorn, S., Wongtavatchai, J., 2016. Francisella infection in cultured tilapia inThailand and the inflammatory cytokine response. J. Aquat. Anim. Health 28,97–106.
Keawcharoen, J., Techangamsuwan, S., Ponpornpisit, A., Lombardini, E.D., Patchimasiri,T., Pirarat, N., 2015. Genetic characterization of a betanodavirus isolated from aclinical disease outbreak in farm-raised tilapia Oreochromis niloticus (L.) in Thailand.J. Fish Dis. 38, 49–54.
Kembou Tsofack, J.E., Zamostiano, R., Watted, S., Berkowitz, A., Rosenbluth, E., Mishra,N., Briese, T., Lipkin, W.I., Kabuusu, R.M., Ferguson, H., Del Pozo, J., Eldar, A.,Bacharach, E., 2016. Detection of tilapia lake virus (TiLV) in clinical samples byculturing and nested RT-PCR. J. Clin. Microbiol.
Kurita, J., Nakajima, K., Hirono, I., Aoki, T., 1998. Polymerase chain reaction (PCR)amplification of DNA of red sea bream iridovirus (RSIV). Fish Pathol. 33, 17–23.
Merour, E., LeBerre, M., Lamoureux, A., Bernard, J., Bremont, M., Biacchesi, S., 2011.
P. Tattiyapong et al. Veterinary Microbiology 207 (2017) 170–177
176

Completion of the full-length genome sequence of the infectious salmon anemia virus,an aquatic orthomyxovirus-like, and characterization of mAbs. J. Gen. Virol. 92,528–533.
Muench, R.H., 1938. A simple method of estimating fifty percent endpoints. Am. J.Epidemiol. 27, 493–497.
Sarder, M.R., Thompson, K.D., Penman, D.J., McAndrew, B.J., 2001. Immune responses ofnile tilapia (Oreochromis niloticus L.) clones: I. non-specific responses. Dev. Comp.Immunol. 25, 37–46.
Soto, E., Hawke, J.P., Fernandez, D., Morales, J.A., 2009. Francisella sp. an emergingpathogen of tilapia, Oreochromis niloticus (L.), in Costa Rica. J. Fish Dis. 32,713–722.
Suebsing, R., Pradeep, P.J., Jitrakorn, S., Sirithammajak, S., Kampeera, J., Turner, W.A.,
Saksmerprome, V., Withyachumnarnkul, B., Kiatpathomchai, W., 2016. Detection ofnatural infection of infectious spleen and kidney necrosis virus in farmed tilapia byhydroxynapthol blue-loop-mediated isothermal amplification assay. J. Appl.Microbiol. 121, 55–67.
Surachetpong, W., Janetanakit, T., Nonthabenjawan, N., Tattiyapong, P., Sirikanchana,K., Amonsin, A., 2017. Outbreaks of tilapia lake virus infection, Thailand,2015–2016. Emerg. Infect. Dis. 23, 1031–1033.
Wang, Y.T., Huang, H.Y., Tsai, M.A., Wang, P.C., Jiang, B.H., Chen, S.C., 2014.Phosphoglycerate kinase enhanced immunity of the whole cell of Streptococcusagalactiae in tilapia, Oreochromis niloticus. Fish Shellfish Immunol. 41, 250–259.
Weli, S.C., Aamelfot, M., Dale, O.B., Koppang, E.O., Falk, K., 2013. Infectious salmonanaemia virus infection of Atlantic salmon gill epithelial cells. Virol. J. 10, 5.
P. Tattiyapong et al. Veterinary Microbiology 207 (2017) 170–177
177