evolutionary history of the metazoa - home | suny geneseobosch/invhistory.pdf · evolutionary...
TRANSCRIPT

1
If we make the time animals have existed to be 1 day, humans have been around for 61 seconds.
Pushing back the origin of animals Oldest traces of animals: ~ 645 mya: 24-isopropylcholestane (2009) Oldest animals sponges, ~650 mya (Science Daily, 2010)
Evolutionary History
of the Metazoa
Mysteries of ancient animal life
Goals:
Consider the central questions about the origin of Metazoa. Review modern evidence about time of origin, patterns of evolution, important processes.
Outline for “History of Life” I. Cambrian Explosion
II. What we knew 15 yrs ago
III. More Recent Discoveries
A. New Fossil Discoveries
B. Molecular Dating
IV. Some explanations for deep divergence
V. Why so many body plans in Cambrian?
First cells with nucleus
Earth formed 4.6 bya
4.5 bya
4 bya
Meteorites bombard the planet 4.6-3.8 bya Making it inhabitable
1st evidence of life Fossil b-g algae 3.6 bya
3 bya
2.0 bya
2.5 bya
First multicelled algae 1.5 bya
Precambrian, 4.6 bya To 543 mya
1.0 bya
600 mya 543 500 mya 400
Cambrian Period 543-510 mya
Source Newsweek
Biology’s “Big Bang”: The Cambrian Fauna (543-510 mya)
Cambrian Explosion
Astonishing array of Multicellular life. All Modern phyla present.
And many More!
Cambrian deposits known since 1909 from >34 locations including the U.S.A., Russia, China, Canada
In Burgess Shale alone, 15 of the 33 modern phyla are recognized
Also 19 unique body forms of Unknown affinity
Morris 1989
n = 40,000

2
• Enigmatic Ediacaran assemblage (565 mya), many of which apparently never survived to the Cambrian
• A 20 my gap in the fossil record before the Cambrian
• Cambrian explosion of body plans
All Modern phyla present and many other types that did not survive.
Skeletized fauna
Summary: ~ 15 years ago Darwin’s Dilemma Why did the Cambrian Fauna present a challenge to the Darwinian view of Evolution? How did Darwin and his contemporary evolutionists respond to this criticism? Why does Sukumaran argue that in evaluating the Cambrian explosion it is important to recognize that the difference between the concepts of diversity and disparity ?
Artist’s rendition of Vendian assemblage as primitive multicelled animals (565 mya)
Different Views on the Exact Nature of the Vendian assemblage:
1946: Martin Glaessner, an Australian paleontologist called them simple soft bodied animals related to jellyfish, corals, segmented worms
1983: German biologist Adolf Seilacher believed they were giant single celled organisms; their own kingdom of life; a failed experiment 1995: Gregory Retalack, Univ. of Oregon, argues they are ancient lichens (structural expts). 2008: Xiao and Laflamme….biota of protists, fungi, algae, and metazoa, but there is growing evidence that among the 100 or so disparate species there were also a few bilateria.
Outline for “History of Life” I. Cambrian Explosion
II. What we knew 15 yrs ago
III. More Recent Discoveries
A. New Fossil Discoveries
B. Molecular Dating
IV. Some explanations for deep divergence
V. Why so many body plans in Cambrian?
A. Dating Techniques 1995: Groetzinger et. al . (Physical Chemistry)
Ediacara Fauna
Precise lead and zircon dating of Vendian-Cambrian boundaries “bridges” the 20 my gap. Some Vendian species survive to the Cambrian; transitional rocks between Vendian and Cambrian explosion bear traces of metazoan life gaining momentum.
3
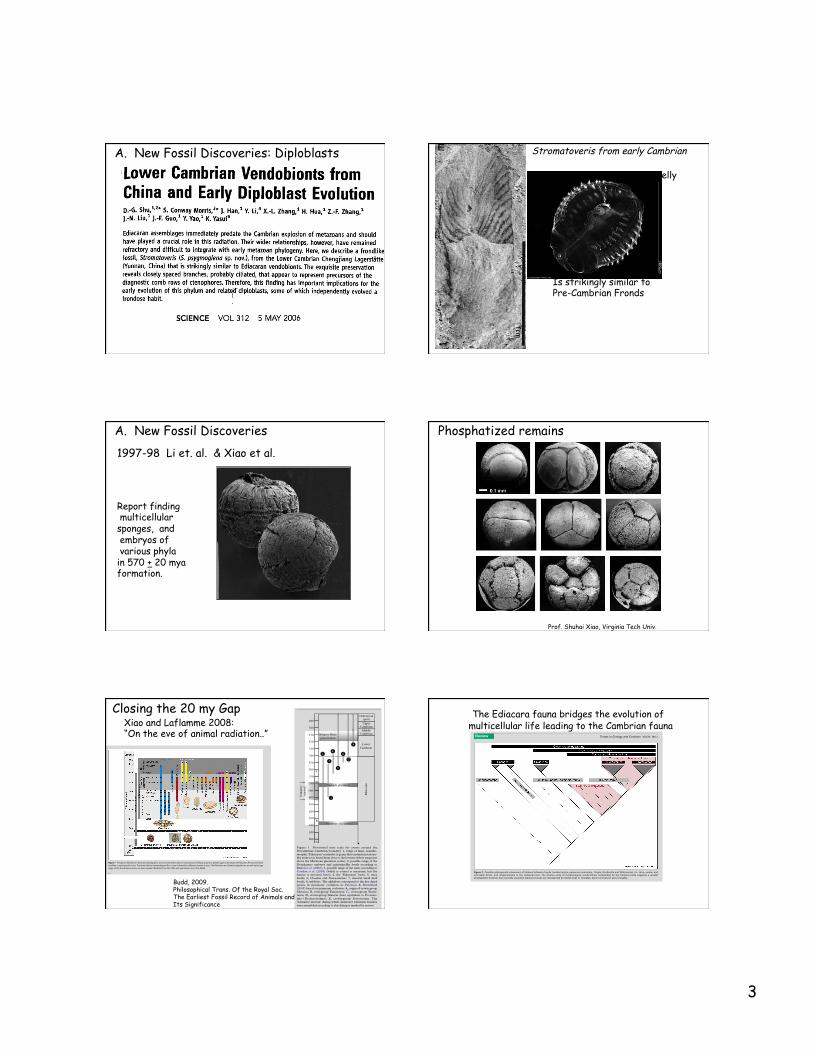
A. New Fossil Discoveries: Diploblasts Stromatoveris from early Cambrian
Modern Comb Jelly
Is strikingly similar to Pre-Cambrian Fronds
1997-98 Li et. al. & Xiao et al. Report finding multicellular sponges, and embryos of various phyla in 570 + 20 mya formation.
A. New Fossil Discoveries Phosphatized remains
Prof. Shuhai Xiao, Virginia Tech Univ.
The phylogenetic uncertainty of Ediacara organisms notonly limits their role in testing hypotheses about the tempoof early animal evolution but also compromises our abilityto interpret their ecology using modern analogs. Fortu-nately, ecological inferences can be independently made onthe basis of trace fossils, functional morphology and taph-onomy. In the past decade, investigation of trace fossilsassociated with Ediacara body fossils has shed importantlight on the autecology of several Ediacara taxa, whereasrecent advances in the paleoecology of Ediacara organisms[9] are of close relevance to the evolutionary radiation inthe Ediacaran–Cambrian transition.
The Ediacaran–Cambrian transition marks a rapidchange in taxonomic diversity, morphological disparityand ecosystem complexity of early animals. Does the earlyevolution of the Ediacara biota share similar patterns withthe Cambrian radiation of animals? Only recently havepaleontologists begun to approach this question usingquantitative methods and a growing database of Ediacarafossils [10,11].
Promising results from emerging research of the phylo-genic affinities, ecological diversity and evolutionary pat-terns of the Ediacara biota prompt this review. Thus, wewill begin with a brief description of the spatial–temporaldistribution and bodyplan diversity of the Ediacara biota,followed by a review of recent advances in the phylogenetic,paleoecological and evolutionary analyses of this biota.
Emphasis is restricted to classical Ediacara fossils thatare soft bodied, macroscopic and morphologically diverse.
The Ediacara biota in space and timeEdiacara fossils are mostly restricted between 575 and541 Ma (Figure 1; Box 1). Discoid fossils from the>635 Ma Twitya Formation in northwestern Canada[12] are similar to some simple forms in the Ediacara biota,but the absence of co-occurring complex forms and theirsignificantly older age suggest that the Twitya discs arepossibly simple forerunners rather than parts (e.g. hold-fasts) of complex Ediacara fossils. A few Cambrian fossilsare interpreted as Ediacara survivors or as phyletic des-cendants [13,14], but with rare exceptions [14] the mosticonic members of the Ediacara biota – the rangeomorphsand erniettomorphs (Box 2), for example – are unknown inthe Cambrian. It has been proposed that the demise of theEdiacara biota might be due to the closure of a uniquetaphonomic window mediated by microbial activities[4,15], and that the Ediacara biota continued to strive afterthe Cambrian radiation but were simply not preserved.However, the scarcity of Ediacara fossils in exceptionallypreserved Cambrian biota such as the Burgess Shale [16]points to a more likely scenario of extinction or at leastecological restriction [2].
The restricted temporal distribution is in contrast witha wide spatial distribution of the Ediacara biota. Ediacara
Figure 1. Temporal distribution (bars) and stratigraphic occurrences (black dots) of representative Ediacara genera, plotted against timescale of Ediacaran Period and fossillocalities or stratigraphic units. The three Ediacara assemblages (Box 1) are indicated by different shades of gray. The Marinoan and Gaskiers glaciations, as well as the agerange of the Doushantuo biota, are also marked. Modified from Ref. [58] with permission from the AAAS.
Review Trends in Ecology and Evolution Vol.24 No.1
32
Closing the 20 my Gap Xiao and Laflamme 2008: “On the eve of animal radiation..”
Doushantuo Fm would have to be incorrect (Barfodet al. 2002), but given the care required to interpret thewhole-rock radiometric dates, this possibility cannotsimply be ruled out.
More recently, the claim has been made that at leastone of the enigmatic acanthomorphic (i.e. spinose)acritarchs (figure 1), which are normally assigned toprotist groups such as the green algae and thedinoflagellates, are actually the hulls of diapause animaleggs (Yin et al. 2004, 2007). Although the fossil inquestion, Tianzhushania, is known to contain embryos
only in the upper part of the Doushantuo Fm, it rangesdown to very close to the base, and thus to 630 Myr agoor so. The claim would be that the oldest animal fossilsof the Doushantuo Fm, dating back to just after theNantuo glaciation (i.e. the Chinese glacial depositsnormally correlated with the Marinoan) are of this age,a time that predates the first Ediacaran fossils by some60 Myr, as well as the more conservative molecularclock estimates for the divergence of the bilaterians.
Despite the obvious uncertainties, the most reason-able interpretation of the data thus is that embryo-forming animals of some sort had evolved by just afterMarinoan time, and that sponges and presumed otheranimals had started to emerge by 580 Myr ago at thelatest, and that the Ediacaran biotas are likely to be alittle younger than the Doushantuo embryos. Theupshot of the new data is that considerably moreconvincing evidence exists in the fossil record for anorigin of the animals considerably before the Cambrianthan it did 10 years ago (Budd & Jensen 2000), with aninferred documented fossil origin of the entire cladebeing datable to just after 635 Myr ago—a significantresult (see figure 2 for summary).
If animals had already evolved at this time, why is itthat the rest of the recorddoes not correlatewith it—whyno macro body fossils and why no (generally accepted)trace fossils? The answer to this question, which onthe face of it seems directly to contradict predictions(Budd & Jensen 2000) that no animals existedsignificantly before the first good trace fossils at ca555 Myr ago,may hinge onwhat sort of organisms theseembryos represent. Given their relatively unusualdevelopment, with large numbers of cell divisions takingplace without the sign of gastrulation or epithelialformation, it has been suggested that they are fromstem-group metazoans, i.e. from the organisms morebasal than any living animals including sponges(Hagadorn et al. 2006). Given that such an organism,lacking muscles and other features of the more derivedbilaterians,wouldbeunlikely readily to formeither bodyor trace fossils, such an assignment is consistent with thehypothesis that bilaterians emerged later, close to
640
630
620
550
540
530
520
610
600
590
580
570
560
510
500
490
Edia
cara
n
LowerCambrian
MiddleCambrian
UpperCambrian
Ordovician(part)
Gaskiers glaciation
A
B
650
660
D
1
E
C
3
4
5
6
7
8
Burgess Shalepreservation
Marinoan glaciation
form
ativ
ein
terv
al
2
Figure 1. Provisional time scale for events around thePrecambrian–Cambrian boundary. 1, range of large, acantho-morphic ‘Ediacaran’ acritarchs (a genus that containsmetazoan-like embryos is found from close to the bottomof their range justabove the Marinoan glaciation rocks); 2, possible range of theDoushantuo embryos and cnidarian-like fossils according toBarfod et al. (2002); 3, possible range of the same according toCondon et al. (2005) (which is correct is uncertain, but theformer is favoured here); 4, the ‘Ediacaran’ biota; 5, tracefossils; 6, Cloudina and Namacalathus; 7, classical small shellfossils; 8, trilobites. The alphabets correspond to the key datedpoints in metazoan evolution in Peterson & Butterfield(2005) based on minimum evolution: A, origin of crown-groupMetazoa; B, total-group Eumetazoa; C, crown-group Eume-tazoa; D, crown-group bilateria (here equivalent to Protosto-miaCDeuterostomia); E, crown-group Protostomia. The‘formative interval’ during which distinctive bilaterian featureswere assembled according to this dating is marked by arrows.
Figure 2. The Ediacaran acanthomorphic acritarch Tanariumpluriprotensum from the Tanana Formation, in the Giles 1drillcore, Officer Basin, Australia; 100!. At least somePrecambrian acanthomorphic acritarchs may be the eggs ofanimals. Courtesy of S. Willman.
Earliest fossil record of animals G. E. Budd 1427
Phil. Trans. R. Soc. B (2008)
on 24 August 2009rstb.royalsocietypublishing.orgDownloaded from
Budd, 2009. Philosophical Trans. Of the Royal Soc. The Earliest Fossil Record of Animals and Its Significance
To summarize, it is important to realize that the Edia-cara biota consists of an assortment of phylogeneticallydiverse taxa, possibly ranging from microbial colonies,algae, fungi and protists to animals, including bilateriananimals [2,39,40]. Just as important, the Ediacara biotalikely comprises stem-group members of various extantclades. These stem groups might have some, but not allfeatures that collectively define extant crown clades.Therefore, extreme approaches to push Ediacara fossilsinto the crown-group Metazoa on the basis of plesiomor-phies, or to relegate them into the phylogenetically distantVendobionta because of the lack of crown-group synapor-morphies, are equally undesirable. Given the phylogeneticuncertainties of many Ediacara fossils, paleoecologists arefacing a daunting task to understand the ecological make-up of Ediacara communities using modern analogs.
Ecological diversitySome Ediacara fossils appear to be preserved where theylived, which offers exceptional opportunities for analysis ofcommunity ecology. Such analyses [41–43] show that mostmembers of the Ediacara biota were epibenthic organisms,with a few possible examples of shallow endobenthic(entirely buried; Pteridinium) and semi-endobenthic
(half-exposed and half-buried; Ernietta) organisms[44,45]. Additional evidence for epibenthic and endo-benthic activities comes from shallow burrows and sedi-ment surface traces [34], but there is no convincingevidence for pelagic or deep endobenthic animals.
Perhaps the most noticeable ecological difference fromPhanerozoic epibenthic communities is the dominance ofsessile organisms in the Ediacara biota. All rangeomorphsappear to have been nonmotile, typically attached to aholdfast (e.g. Charnia) [46] or lying freely on the seafloor(e.g. Fractofusus) [47]. Many other Ediacara fossils, in-cludingPalaeophragmodictya, tri-, tetra-, penta- and octor-adial forms, are also likely to have been attached or freelylying on the seafloor. Although vendomorphs and parvan-corinomorphs were interpreted as relatives of arthropods,there is no convincing evidence for motility. Yorgia andDickinsonia moved intermittently, facultatively and per-haps passively. Only Kimberella seems to have activelypushed sediments during self-powered movement [8,32].The subordinate role of motile animals is also supported bythe lower abundance and diversity of trace fossils in Edia-caran rocks than in Phanerozoic rocks [34].
Although Ediacara communities were likely supportedby cyanobacterial and algal primary producers [48], the
Figure 2. Possible phylogenetic placement of bilateral Ediacara fossils (vendomorphs, parvancorinomorphs, Yorgia, Kimberella and Dickinsonia), tri-, tetra-, penta- andoctoradial forms, and rangeomorphs in the metazoan tree. The diverse array of morphological constructions exemplified by the Ediacara biota suggests a greaterphylogenetic diversity than typically assumed. Ediacara fossils are represented by dotted lines or triangles, extant animals by gray triangles.
Review Trends in Ecology and Evolution Vol.24 No.1
36
The Ediacara fauna bridges the evolution of multicellular life leading to the Cambrian fauna

4
Vertebrates Without Jaws (Extinct) The first Craniates Ca. 530 mya
Yunnanozoon
Myllokunmingia
Haikouella
Ca. 530 mya
New fossil discoveries show the presence of complex (but soft-bodied) animal life before the Cambrian (~570 mya)
Outline for “History of Life” I. Cambrian Explosion
II. What we knew 15 yrs ago
III. More Recent Discoveries
A. Improved Dating Techniques
B. New Fossil Discoveries
C. Molecular Dating
IV. Some explanations for deep divergence
V. Why so many body plans in Cambrian?
3. Explain the concept of disparity and what point does the author try to make aboutthe disparity of the Cambrian fauna?
What does Sukumaran mean by the term “deep divergence” ?
What is the procedure known as “molecular dating” and how is it used to estimate the divergence times for animal phyla?
Fossil Ancestor of
A & B
Calibrating the molecular clock
Age Known From fossil record
Descendant Taxon A
Descendant Taxon B
Determine % difference In mtDNA Gene sequence
% difference years
Molecular Clock Rate
Extant species
years
e.g. nucelotide substitutions Per 10 million years
Genetic Evidence : sequence analyses can provide information on relatedness AND time of divergence
1996. Wray et. al. estimate time of divergence of some phyla at 1 to 1.2 mya
Used vertebrate clock data to estimate divergence times But, Not all molecular clock rates are the same!

5
Copyright ©2004 by the National Academy of Sciences
Peterson, Kevin J. et al. (2004) Proc. Natl. Acad. Sci. USA 101, 6536-6541
Fig. 2. ML tree of the seven concatenated protein sequences from 18 in-group taxa by using Arabidopsis as the outgroup. Shows a slower molecular clock for 8 genes among the
vertebrates, and a faster rate among invertebrates that is consistent between groups
1998 Ayala et al. Added Many genes to the
analysis and discard genes that show variable
molecular clock rates
Copyright ©2004 by the National Academy of Sciences
Peterson, Kevin J. et al. (2004) Proc. Natl. Acad. Sci. USA 101, 6536-6541
Fig. 4. Metazoan divergence estimates with metazoan diversity and phylogeny (7 different a.a. sequences from 23 taxa)
§ What were the ancestors of the Cambrian Fauna? (Ediacaran fauna? 20 my gap?)
§ When did the major lineages appear? (support for cryptic pre-Cambrian history > 600 mya, unique and skeletized body plans appear later.... Referred
to as “deep divergence”)
§ Why was there a burst of body plan diversity starting in the middle Cambrian?
Outline for “History of Life” I. Cambrian Explosion
II. What we knew 15 yrs ago
III. More Recent Discoveries
A. Improved Dating Techniques
B. New Fossil Discoveries
C. Molecular Dating
IV. Some explanations for deep divergence
V. Why so many body plans in Cambrian?
What are HOX genes? Is it possible that the evolution of HOX genes might help explain the rapid origin of diverse body plans and skeletal fauna in the Cambrian? * A simple pair of Hox genes is present in
modern Cnidaria * Basal flatworms have 4 sets * Other Bilateria have 9-11 hox genes

6
Transcription factors, like Hox proteins or Fox proteins, recognize specific sequences in the regulatory regions of downstream target genes, usually motifs present in the 50 or ‘‘upstream’’ region of the gene, and when bound they regulate the transcription of the target
gene. miRNAs, on the other hand, are regulatory RNA molecules that recognize specific sequences in the 30 untranslated region (30UTR) of messenger RNA molecules, and once bound to a target site ultimately prevent the translation of the messenger RNA.(53)
• miRNAs increase genic precision by turning an imprecise number of mRNA transcripts into a more precise number of protein molecules • By reducing gene expression variability they also increase heritability
and allow natural selection to elaborate morphological complexity
• More complex animals have more miRNAs • Origin and elaboration of miRNA systems may have contributed to the high rate of morphological diversification in the Cambrian Explosion
What are HOX genes? Is it possible that the evolution of HOX genes might help explain the rapid origin of diverse body plans and skeletal fauna in the Cambrian? What evidence indicates that the evolution of miRNAs may have played a role in the early evolution of animal life?
Transcription factors, like Hox proteins or Fox proteins, recognize specific sequences in the regulatory regions of downstream target genes, usually motifs present in the 50 or ‘‘upstream’’ region of the gene, and when bound they regulate the transcription of the target
gene. miRNAs, on the other hand, are regulatory RNA molecules that recognize specific sequences in the 30 untranslated region (30UTR) of messenger RNA molecules, and once bound to a target site ultimately prevent the translation of the messenger RNA.(53)
Conclusions:
• Ancestors of Metazoa were around at least 30-80 million years before the Cambrian. ‘Explosion’ may be a more gradual radiation.
• Did the evolution of genetic and developmental complexity contribute to the origin of so many body plans?
Why so many species in the Cambrian? Why a surge of organisms with a skeleton?
• Vendian fauna includes representatives of some modern phyla. They and others that apparently did not leave many fossils contributed to the Cambrian fauna.
Gaskers Ice age Ghaub/Nantuo Ice Age
warm
7
Gaskers Ice age Ghaub/Nantuo Ice Age
warm
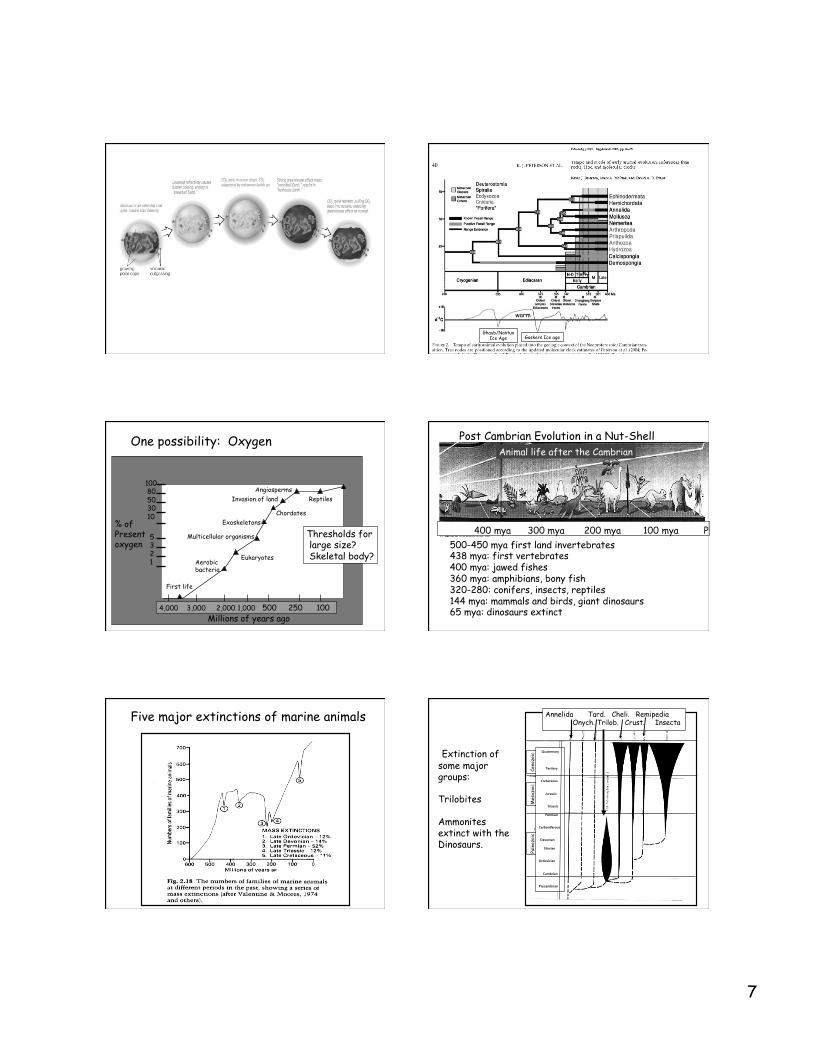
One possibility: Oxygen
4,000 3,000 2,000 1,000 500 250 100 Millions of years ago
% of Present oxygen
100 80 50 30 10 5
3 2 1
First life
Aerobic bacteria
Eukaryotes
Multicellular organisms
Exoskeletons Chordates
Invasion of land Angiosperms
Reptiles
Thresholds for large size? Skeletal body?
Post Cambrian Evolution in a Nut-Shell
400 mya 300 mya 200 mya 100 mya P
Animal life after the Cambrian
500-450 mya first land invertebrates 438 mya: first vertebrates 400 mya: jawed fishes 360 mya: amphibians, bony fish 320-280: conifers, insects, reptiles 144 mya: mammals and birds, giant dinosaurs 65 mya: dinosaurs extinct
Five major extinctions of marine animals
Extinction of some major groups: Trilobites Ammonites extinct with the Dinosaurs.
Annelida Tard. Cheli. Remipedia Onych. Trilob. Crust. Insecta
Pale
ozoi
c M
esoz
oic
Ceno
zoic
Quaternary Tertiary Cretaceous Jurassic Triassic Permian Carboniferous Devonian Silurian Ordovician Cambrian Precambrian

8
Great Mysteries of Early Animal Life
1. What were the ~30 forms in the Ediacara?
Much has been learned in the past 10 years!!
2. Gap of 20 my real? Or were Vendian animals precursors to the Cambrian fauna?
3. Where are the ancestors of the Cambrian Phyla?
4. What conditions (ecological, evolutionary) contributed to great surge of life forms?