evaluation of cellular and humoral systemic immune response against giardia duodenalis infection in...
TRANSCRIPT

V
Er
GJa
9b
c
ARRA
KGCICCS
0
ARTICLE IN PRESSG ModelETPAR-7185; No. of Pages 11
Veterinary Parasitology xxx (2014) xxx–xxx
Contents lists available at ScienceDirect
Veterinary Parasitology
jo u r nal homep age: www.elsev ier .com/ locate /vetpar
valuation of cellular and humoral systemic immuneesponse against Giardia duodenalis infection in cattle
.H. Grita, S. Van Coppernollea, B. Devriendta, T. Geurdenc, L. Dreesena,. Hopeb, J. Vercruyssea, E. Coxa, P. Geldhofa, E. Claerebouta,∗
Department of Virology, Parasitology & Immunology, Faculty of Veterinary Medicine, Ghent University, Salisburylaan 133,820 Merelbeke, BelgiumThe Roslin Institute, University of Edinburgh, Easter Bush, Midlothian EH25 9RG, UKZoetis – Veterinary Medicine Research and Development, Mercuriuslaan 20, 1930 Zaventem, Belgium
a r t i c l e i n f o
rticle history:eceived 17 December 2013eceived in revised form 6 March 2014ccepted 8 March 2014
eywords:iardia duodenalisattle
mmune responseytokinesD4+ T-cellserum antibodies
a b s t r a c t
Giardia duodenalis causes diarrhoea in humans and a wide range of mammals, includingcattle. In cattle, the infection often has a chronic character. Infected calves may excretecysts for several months, suggesting that Giardia is able to suppress and evade the immuneresponse.
In this study six calves were infected with G. duodenalis assemblage A and E and housed inan environment that allowed reinfection. Cyst excretion was monitored twice a week andblood was collected every 2 weeks, until decreasing cyst counts indicated the developmentof protective immunity. The kinetics of the circulating memory cells and serum antibodieswere followed up throughout this period. Cyst excretion started 1 week post-infection andremained high until week 14. Low cyst counts from week 15 p.i. onwards indicated that thecalves had developed immunity.
From week 5 p.i. significant proliferation of CD4+ �� T-cells was observed after in vitrostimulation with G. duodenalis antigen. Characterisation of the proliferating CD4+ T-cellsusing real time qPCR showed that at the peak of antigen driven PBMC proliferation themajority of cells were CD4+ T-cells expressing IL-17 and to a lesser extent FoxP3. The cellproliferation was strongly reduced after plastic adhesion of the PBMC, suggesting a rolefor antigen-presenting cells. Failure to restore proliferation of depleted PBMC with Giardia-stimulated monocyte-derived dendritic cells (MoDC) and unchanged proliferation afterdepletion of CD21+ B-cells showed that other antigen-presenting cells than MoDC and B-cells were important for T-cell proliferation. Analysis of the antibody response indicatedthat serum IgG1 and IgA levels against G. duodenalis assemblage A and E increased fromweek 11 post-infection. From the start of the antibody response, all trophozoites stained
Please cite this article in press as: Grit, G.H., et al., Evaluation ofGiardia duodenalis infection in cattle. Vet. Parasitol. (2014), http
positive in an immunofluorescence assay with serum antibodies, indicating that a broadrepertoire of antibodies was produced against all variant-specific surface proteins.
Further research is necessary to determine which effector T-cell subset produces IL-17and which cells play a role in antigen presentation.
© 2014 Elsevier B.V. All rights reserved.
∗ Corresponding author. Tel.: +32 09 2647393; fax: +32 092647496.E-mail address: [email protected] (E. Claerebout).
http://dx.doi.org/10.1016/j.vetpar.2014.03.012304-4017/© 2014 Elsevier B.V. All rights reserved.
cellular and humoral systemic immune response against://dx.doi.org/10.1016/j.vetpar.2014.03.012

ING Model
Parasit
incubated with medium alone were used as negativecontrols. As a positive control 5 �g/ml Concanavalin A
ARTICLEVETPAR-7185; No. of Pages 11
2 G.H. Grit et al. / Veterinary
1. Introduction
Giardia infections are an important cause of diarrhoeain humans and a wide range of mammals, including cat-tle. In cattle Giardia duodenalis infections can cause chronicintermittent diarrhoea and weight loss and have been asso-ciated with significant economical losses (Geurden et al.,2010a,b). Generally calves suffer from different episodesof diarrhoea and develop a chronic stage of giardiosis(O’Handley et al., 1999). Calves can excrete cysts until atleast 112 days after infection (Taminelli et al., 1989) anda peak prevalence of G. duodenalis is seen in animals agedbetween 1 and 6 months (Xiao and Herd, 1994; O’Handleyet al., 1999).
In cattle little is known about the development ofthe immune response against G. duodenalis. It seems thatduring the infection the required adaptive immunity incalves is not complete and hence re-infections often occur.Dreesen et al. (2012) found no inflammation in the smallintestine of calves after 3 weeks of G. duodenalis infection.Microarray results showed a gene pattern correspond-ing to decreased inflammation, immune cell recruitmentand immune response. This anti-inflammatory responseis possibly caused by the nuclear peroxisome proliferator-activated receptors PPAR� and PPAR� acting as regulators(Dreesen et al., 2012). However, after 6 weeks of infectionmoderate to severe diffuse inflammation and cellular infil-tration of the jejunal mucosa has been observed in calves(Taminelli et al., 1989; Ruest et al., 1997). In cattle, toour knowledge, there is no data on the humoral responseexcept for one study where no significant increase of IgGwas found during infection (O’Handley et al., 2003). HenceO’Handley et al. (2003) suggested that chronic infectionsin calves are a result of the failure to develop a humoralresponse.
Current knowledge of immunity against Giardia ismainly based on studies in mice. In mice CD4+ T-cells arecrucial in both the acute and the antibody dependent phase(Singer and Nash, 2000; Zhou et al., 2007). Antibodies alsoplay an important role; IgA is indispensible for protectiveimmunity (Taherkhani et al., 2009). It is thought that theshift of variant specific surface proteins (vsp) is driven bythe antibody response of the host (Prucca et al., 2008). Itis hypothesised that the development of immunity is com-plete when all Giardia vsp, which cover the entire surface ofthe trophozoite, are recognised by the specific antibodies(Lujan and Svärd, 2011).
In this study we investigated the systemic immuneresponse against G. duodenalis infection in a naturalhost, i.e. cattle. The immune response was monitoreduntil decreasing cyst excretion indicated the develop-ment of protective immunity. We studied the kineticsand nature of circulating memory cells and identifiedthe proliferating cells and their cytokine transcriptionlevels. The role of antigen-presenting cells in the initia-tion of the cellular response was assessed. The kineticsof serum antibodies were evaluated and G. duodenalistrophozoites were stained with serum antibodies toinvestigate if the proportion of trophozoites that were
Please cite this article in press as: Grit, G.H., et al., Evaluation ofGiardia duodenalis infection in cattle. Vet. Parasitol. (2014), http
recognised by serum antibodies increased with developingimmunity.
PRESSology xxx (2014) xxx–xxx
2. Materials and methods
2.1. Animals and infection
Six healthy male Holstein Friesian calves between 10and 14 days old were confirmed to be free of Giardia andCryptosporidium by faecal examinations on 2 consecutivedays, using a direct immunofluorescence assay (Merifluor©
Cryptosporidium/Giardia kit, Meridian Diagnostics Inc.).The calves were fed with colostrum in the first few hoursafter birth and were subsequently fed with a commercialmilk replacer. The calves were housed two by two in boxeswith straw. At the age of 3 weeks they were given an oralinfection with 105 G. duodenalis cysts, isolated from twonaturally infected calves. The cysts were confirmed to be amixture of assemblage A and E by PCR, targeting the triosephosphate isomerase gene (Sulaiman et al., 2003; Geurdenet al., 2008). The calves were weaned at week 9 and housedtogether in a box with straw bedding. Fresh straw wasadded regularly but the boxes were never cleaned, to allowreinfection from the environment.
During the experiment rectal faecal samples were col-lected two times a week and Giardia cyst excretion wasmonitored by a quantitative direct immunofluorescenceassay (Merifluor© Cryptosporidium/Giardia kit, MeridianDiagnostics Inc.) (Geurden et al., 2004). Geometric meancyst counts were calculated using Excel version 14.0.0. Fae-cal cyst counts were continued until repetitive cyst countsof ≤200 cyst/g of faeces in all calves suggested that the ani-mals had developed protective immunity. The experimentswere approved by the ethical committee of the faculty ofveterinary medicine, Ghent University (approval numberEC 2012/094).
2.2. Isolation of peripheral blood mononuclear cells
Every 2 weeks, venous blood was drawn from thevena jugularis using vacutainer tubes with heparin. Periph-eral blood mononuclear cells (PBMC) were obtained usinga Lymphoprep density gradient (Axis-Shield). The inter-phase was harvested and cells were washed three times inPBS. Cells were cultured in 96-well round-bottomed plates(Nunc) in cell culture medium (RPMI-1640 + Glutamax,Invitrogen) supplemented with 10% foetal calf serum,50 �g/ml gentamycin and 1/1000 �-mercapto-ethanol orwere first depleted for antigen-presenting cells by plasticadhesion.
2.3. Cell proliferation assays
PBMC (2 × 105) were cultured with 2 × 104 live G. duode-nalis trophozoites from an axenic G. duodenalis assemblageE culture (Laboratory of Microbiology, Parasitology andHygiene, University of Antwerp, Belgium) in a 96-wellround-bottom plate (Nunc) in modified TYI-S-33 medium(Keister, 1983). The trophozoites were viable and mobilefor up to 36 h after the start of the co-incubation. Cells
cellular and humoral systemic immune response against://dx.doi.org/10.1016/j.vetpar.2014.03.012
(Sigma–Aldrich) was used. All conditions were performedin triplicate. PBMC depleted for either antigen-presenting

ING ModelV
Parasit
cncwpheaitm
uiPadaamCVwaCBwPsSwMdpttm
2
wftPCawqccTspRcwaR2s
ARTICLEETPAR-7185; No. of Pages 11
G.H. Grit et al. / Veterinary
ells or B-lymphocytes were stimulated with the sameumber of G. duodenalis trophozoites and in the sameonditions as for full PBMC. After 5 days of culture, cellsere pulsed with 1 �Ci [3H] thymidine (Amersham ICN)er well and incubated for 18 h. Subsequently cells werearvested on to glass fibre filters (Perkin Elmer, Life Sci-nces). The incorporated radioactivity was measured using
�-scintillation counter (Perkin Elmer). The stimulationndex was calculated by dividing the counts per minute ofrophozoite-stimulated cells by the count per minute of the
edium control.PKH staining was performed to identify which cell pop-
lations from the PBMC were proliferating. PBMC weresolated as described above and labelled with PKH using theKH26 red fluorescent cell linker mini kit (Sigma–Aldrich)ccording to the manufacturer’s instruction. The same con-itions and cell numbers were used as for the thymidinessay. After 6 days of culture, the cells were harvested and
quadruple staining was performed using the followingouse anti-bovine antibodies: CD3 (IgG1, MM1A VMRD),
D4 (IgG2a, MCA1653G, AbD Serotec), CD8 (IgM, BAQ111AMRD) and TCR�� (IgG2b, GB21A VMRD). Bound mAbere detected by anti-mouse IgG1-V450 (BD Biosciences),
nti-mouse IgG2a-APC (Invitrogen), anti-mouse IgM-APC-y7 (Biolegend) and anti-mouse IgG2b-FITC (Southerniotech). Sytox blue dead cell stain (Live Technologies)as added to distinguish live cells from dead cells. The
KH intensity was measured using the FACSAriaIII (BD bio-ciences). Data were analysed using Flowjo software (Treetar) and the proliferation of different cell populationsas quantified by calculating the stimulation index usingodFit LT software (Verity Software House). To do so the
ifferent populations were gated and for each separate cellopulation a proliferation ratio was calculated based onhe number of cell divisions. The proliferation ratios werehen compared between cell populations stimulated with
edium vs. Giardia.
.4. Characterisation of proliferating CD4+ T-cells
On week 13 of infection, when CD4+ T-cell proliferationas the highest, CD4+ T-cells were sorted from stimulated
ull PBMC after 6 days of stimulation with G. duodenalisrophozoites and characterised by real time quantitativeCR (RT qPCR). For the sorting a quadruple staining forD3, CD4, CD8 and TCR�� was done as described abovend analysed using the FACSAriaIII. Doublet exclusionas performed based on FSC-H and FSC-A, and subse-
uently on SSC-H and SSC-A. Propidium iodide positiveells were excluded from the analysis. The CD3+CD4+ T-ells were sorted. After sorting, cells were washed in PBS.he supernatant was removed and the cell pellets werenap-frozen in liquid nitrogen and stored at −80 ◦C untilrocessing. The RNeasy kit (Qiagen) was used to extractNA following the manufacturer’s instructions. To removeontaminating genomic DNA, on-column DNase digestionas performed using the RNase-free DNase set (Qiagen)
Please cite this article in press as: Grit, G.H., et al., Evaluation ofGiardia duodenalis infection in cattle. Vet. Parasitol. (2014), http
ccording to the manufacturer’s instructions. ObtainedNA concentrations were measured using a Nano-Drop000 spectrophotometer (Thermo Scientific) and first-trand cDNA was generated from 100 ng RNA using the
PRESSology xxx (2014) xxx–xxx 3
iScript cDNA synthesis kit (Bio-rad). The reaction was per-formed in a 20 �l volume, according to the manufacturer’sinstructions.
Real-time quantitative PCR was carried out for a panelof selected cytokines and transcription factors, allowingdifferentiation of pro-inflammatory and anti-inflammatoryor inhibitory cell functions. The transcription of the fol-lowing cytokines and transcription factors was evaluated:T-Bet, Gata-3, FoxP3, RORc, IL-2, IL-4, IL-6, IL-10, IL-15,IL-17, TGF-�, IFN-�, TNF-� and Peroxisome proliferator-activator receptor alpha and gamma (PPAR� and PPAR�).All primer sequences are listed in Table 1. Real-time qPCRwas performed according to Dreesen et al. (2012) using aStepOnePlus Real-Time PCR system (Applied Biosciences),using 10 �l SYBR Green master mix (Applied Biosystems),2 �l of single stranded cDNA (1 ng of the input total RNAequivalent) and 400 nM of each amplification primer in a20 �l reaction volume. Primer efficiencies ranged between1.90 and 2.09. All conditions were performed in duplicate.A relative quantification was carried out using the delta-delta Ct method. Out of a panel of 4 candidate control genes,the ribosomal protein S29 (RSP29) and hypoxanthine phos-phoribosyltransferase 1 (HPRT1) genes were selected fornormalisation, based on gene stability.
2.5. Depletion of PBMC for antigen-presenting cells orB-cells
To determine the contribution of antigen-presentingcells and B-lymphocytes to the lymphocyte prolifera-tion, proliferation assays were also performed with PBMCdepleted for antigen-presenting cells or B-cells, respec-tively. PBMC were depleted for antigen-presenting cells byplastic adhesion. PBMC were cultured in a concentration of2 × 106 cells/ml in cell culture medium in a cell culture flask(75 cm2, Novolab) for 2–3 h to allow antigen-presentingcells to attach to the plastic. Afterwards the non-adheredcells were harvested and cultured. The efficiency of theplastic adhesion was measured by flow cytometry. Non-adherent cells were stained with mouse anti-bovine CD14(IgG1, CCG33), and bound antibodies were detected usinganti-mouse IgG1-V450 (BD Biosciences). The maximum ofremaining CD14+ cells measured in the depleted PBMCfraction was 3%.
PBMC collected 19–21 weeks post-infection weredepleted of CD21+ B-cells or MHCII+ cells (including allCD21+ cells). This experiment was performed using cellsfrom four animals. A double staining was performed usingprimary anti-bovine CD21 (IgM, BAQ15A VMRD) and MHCII(IgG2a, CC158) antibodies. Bound antibodies were detectedby anti-mouse IgG2a-APC (Invitrogen) and anti-mouseIgM-APC-Cy7 (Biolegend). Doublet exclusion was donebased on FSC-H and FSC-A, and subsequently on SSC-Hand SSC-A. Propidium iodide stain (Invitrogen) was addedto distinguish live cells from dead cells, incubated for3 min and unbound propidium iodide was then removedby washing the cells in PBS (BD Biosciences). Propidium
cellular and humoral systemic immune response against://dx.doi.org/10.1016/j.vetpar.2014.03.012
iodide positive cells were excluded from the analysis andMHCIIlow and CD21− cell fractions were gated and sorted.A post-sort measurement was done and the purity ofMHCII− was at least 99% and the purity for CD21− cells

ARTICLE IN PRESSG ModelVETPAR-7185; No. of Pages 11
4 G.H. Grit et al. / Veterinary Parasitology xxx (2014) xxx–xxx
Table 1Genes used in the qRT-PCR assay, indicating GeneBank accession number and primer sequences.
Validation microarray Accession number Primer sequence
RPS29 BC102702 F: GGAGCCATCCGAGAAAATTCGR: CAACTTAATGAAGCCGATGTCCTT
HPRT1 NM 001034035.1 F: CACTGGGAAGACAATGCAGAR: ACACTTCGAGGGGTCCTTTT
RPLP0 NM 001012682.1 F: CTTCATTGTGGGAGCAGACAR: GGCAACAGTTTCTCCAGAGC
HDAC10 NM 001075460.1 F: CCGATGACGGGAGAAATCTAR: CTCAGGAACCCACCAGTTGT
Cytokines Accession number Primer sequence
IL2 NM 180997.1 F: TCCAAGCAAAAACCTGAACCR: CAGCGTTTACTGTTGCATCATC
IL4 NM 173921.2 F: GCGGACTTGACAGGAATCTCR: TCAGCGTACTTGTGCTCGTC
IL-6 NM 173923.2 F: TCCTTGCTGCTTTCACACTCR: CACCCCAGGCAGACTACTTC
IL-10 NM 174088.1 F: TGTTGACCCAGTCTCTGCTGR: GGCATCACCTCTTCCAGGTA
IL15 NM 174090.1 F: AGAGGCTGGCATTCATGTCTR: CTGCATCGCTGTTACTTTGC
IL-17 NM 001008412.1 F: GGACTCTCCACCGCAATGAGR: TGGCCTCCCAGATCACAGA
TGF-�1 NM 001166068.1 F: CTGCTGTGTTCGTCAGCTCTR: TCCAGGCTCCAGATGTAAGG
IFN-� NM 174086.1 F: TTCTTGAATGGCAGCTCTGAR: TTCTCTTCGGCTTTCTGAGG
TNF-� NM 173966.2 F: GCCCTCTGGTTCAGACACTC
Transcription factor Accession number Primer sequence
T-bet TC216567 F: CCTGGACCCAACTGTCAACTR: GAAACTCGGCCTCATAGCTG
FOXP3 NM 001045933.1 F: GACAGCACCCTTTCGACTGTR: CTCCAGAGATTGCACCACCT
GATA3 NM 001076804.1 F: CATCACCACCTACCCACCATR: GCCCCACAGTTCACACACTC
RORC NM 001083451.1 F: CGACCAGATCGTGCTACTCAR: TCATTGTCAGCGTTGTAGGC
PPARA NM 001034036.1 F:TCCCTCTTTGTGGCTGCTAT
PPARG NM 181024.2
was at least 98%. Depleted cell fractions were cultured withG. duodenalis trophozoites and cell proliferation assayswere performed under the same conditions as describedabove.
To ensure that MHCIIhigh cells did not cause the prolif-eration seen in PBMC, a PKH26 staining of CD3− (MHCII+)cells was also performed. On week 11, when the maxi-mum proliferation was seen, PKH staining was performedas described above on full PBMC and CD3− cells were ana-lysed (Fig. 6).
2.6. Generation of bovine monocyte-derived dendriticcells and stimulation of MoDC with Giardia
In a previous study (Grit et al., 2014) it was seenthat G. duodenalis stimulated bovine monocyte-derived
Please cite this article in press as: Grit, G.H., et al., Evaluation ofGiardia duodenalis infection in cattle. Vet. Parasitol. (2014), http
DC (MoDC) were able to cause proliferation of autolo-gous PBMC. To test whether dendritic cells are the leadingcells for induction of the cellular response, PBMC depletedfor antigen-presenting cells were co-cultured with MoDC
R: TCGTCAGGATGGTTGTTCTGF: GATCTTGACGGGAAAGACGAR: ACTGACACCCCTGGAAGATG
stimulated with G. duodenalis trophozoites. One weekbefore PBMC were cultured blood was drawn to gen-erate MoDC. The MoDC were generated from PBMC asdescribed by Werling et al. (1999). Briefly, PBMC were iso-lated as described above and subsequently incubated withanti-human CD14 microbeads (Miltenyi Biotec). CD14+
monocytes were isolated using magnetic cell sorting(MACS, Miltenyi Biotec) according to the manufacturer’sinstructions. The purity of the cells was evaluated by flowcytometry and was always ≥95%. Monocytes were cul-tured (0.5 × 106 cells/ml) for 7 days with 10 ng/ml bovinerIL-4 (Sigma) and 37.5 ng/ml porcine recombinant GM-CSF (Inumaru et al., 1998) in DMEM without phenol-red(Invitrogen) supplemented with 10% foetal calf serum (TCSBiosciences), 500 U/ml penicillin and 500 �g/ml strepto-mycin. Every 3 days the cultures were supplemented withfresh cytokines. After 7 days the cells were harvested using
cellular and humoral systemic immune response against://dx.doi.org/10.1016/j.vetpar.2014.03.012
2 mM EDTA and brought in co-culture with PBMC. A totalof 2 × 105 depleted PBMC were put in a 96-well round-bottomed plate (Nunc) without MoDC or with MoDC ata MoDC/PBMC ratio of 1/10. The cells were immediately

IN PRESSG ModelV
Parasitology xxx (2014) xxx–xxx 5
saf
2
Itdtct−dSuwf0sbbFaI(2t1(auR(r
2
vFa5osIFTDsms
2
mtv
ARTICLEETPAR-7185; No. of Pages 11
G.H. Grit et al. / Veterinary
timulated with 2 × 104 live G. duodenalis trophozoites and cell proliferation assay with depleted PBMC was per-ormed as described above.
.7. Enzyme-linked immune-sorbent assay (ELISA)
An ELISA was performed to detect Giardia specificgG1, IgG2 and IgA in serum at different time points ofhe infection. PBS soluble extract was made of G. duo-enalis assemblage A and E trophozoites by performinghree freeze–thaw cycles The pellet was spun down byentrifuging 5 min at 1400 rpm and the supernatant con-aining the soluble proteins was collected and frozen at70 ◦C. The protein concentration in the supernatant wasetermined using a Pierce BCA protein assay (Thermocientific). The assay was performed following the man-facturer’s instructions. Maxisorp 96 well plates (Nunc)ere coated with 100 �l per well of 5 ng/ml Giardia extract
or 12 h at 4 ◦C. The plates were washed with PBS with.05% Tween 20 (PBST), and blocked using 2% bovineerum albumin (Invitrogen) in PBST solution (blockinguffer). The serum of all animals was diluted 1/50 inlocking buffer and 100 �l was brought in each well.or detection, horseradish-peroxidase conjugated sheepnti-bovine IgG1 (AAI21P, AbD Serotec), sheep anti-bovinegG2 (AAI22P, AbD Serotec) and sheep anti bovine-IgGAAAI20P, AbD Serotec) were used at a concentration of
�g/ml diluted in blocking buffer. To visualise the reac-ion, 67 mU/ml of conjugate, streptavidin-POD (Roche) and
mg/ml of o-phenylenediamine dihydrochloride (OPD)Sigma–Aldrich) was used. The reaction was stopped bydding 50 �l of a 2.5 M HCl solution. The reaction prod-ct was measured with a 492 nm spectrophotometer.esults were analysed using DeltaSOFT JV 2.1.2 softwareBioMetallics) with a five-parameter curve-fitting algo-ithm. All conditions were performed in triplicate.
.8. Trophozoite staining with serum antibodies
Trophozoites from both assemblage A and E were har-ested from axenic cultures and washed three times in PBS.rom each assemblage 35.000 trophozoites were dried on
3-aminopropyl tri-ethoxysilane (APES) coated slide for min. After drying, the slides were incubated with 50 �lf 1:2 diluted serum for 30 min at room temperature. Thelides were washed and incubated with sheep anti-bovinegG1-FITC (AAI21F, AbD Serotec) or sheep anti-bovine IgA-ITC (AAI20F, AbD Serotec) for 30 min at room temperature.he slides were washed and subsequently 50 �l of 1 �g/mlAPI solution was brought on the slides for 5 min. The
lides were examined using a fluorescence microscope atagnification of 200×. Overlays were made with Image J
oftware.
.9. Statistical analysis
Please cite this article in press as: Grit, G.H., et al., Evaluation ofGiardia duodenalis infection in cattle. Vet. Parasitol. (2014), http
For longitudinal data containing repeated measure-ents (Figs. 2, 3, 5 and 7), a non-parametric Friedman
est was used with Dunn’s post hoc test to compare meanalues of stimulation index, proliferation index or optical
Fig. 1. Geometric mean cyst excretion (cysts per gram of faeces) during aperiod of 18 weeks after oral infection of six calves with 105 G. duodenaliscysts.
density pre-infection (week 0) and post-infection. Forthe qPCR results (Fig. 4), Q values of unstimulated cellsand stimulated cells were compared using a two-sidedWilcoxon signed-rank test. The mean stimulation indexof six animals was compared between full PBMC andPBMC depleted for MHCII cells or B-cells (Fig. 6) using aKruskal–Wallis test with Dunn’s post hoc test. P-values ofless than 0.05 were considered statistically significant.
3. Results
3.1. Infection
Seven days after the oral infection excretion of cysts wasobserved. Geometric mean cyst counts reached a maximumof 62,936 cysts per gram faeces (cpg) at week 5 (Fig. 1).From week 7 onwards a decrease was seen, with the lowestcyst excretion at week 10, probably due to change of hous-ing. However, at week 11 the cyst number increased againand a second peak of 11,814 cpg was reached at week 13,followed by a second decline. From week 15 onwards, thecyst excretion stayed below 200 cpg and the calves wereconsidered to be immune. Throughout the experiment, allcalves suffered short spells of mild to watery diarrhoea.During the whole period, the general condition of the calvesremained good and the animals were vivid and alert.
3.2. PBMC proliferation during infection
Proliferation of PBMC re-stimulated with G. duodenalistrophozoites was assessed every 2 weeks during 18 weeksby measuring the incorporation of thymidine in the DNA ofdividing cells. Stimulated PBMC showed an increased pro-liferation from week 6 onwards with two peaks at weeks7 and 11 (Fig. 2). The proliferation was significantly higheron week 7, 9 and 11 compared to pre-infection levels. Thehigh cell proliferation between week 6 and 12 roughly cor-responds with the period of high cyst excretion (Fig. 1).
cellular and humoral systemic immune response against://dx.doi.org/10.1016/j.vetpar.2014.03.012
The maximum stimulation index (SI = 60.6) was measuredat week 11 (Fig. 2).
Every 2 weeks, proliferating cell populations were iden-tified using PKH staining. At week 9 and 13 proliferation of

ARTICLE IN PRESSG ModelVETPAR-7185; No. of Pages 11
6 G.H. Grit et al. / Veterinary Parasitology xxx (2014) xxx–xxx
Fig. 4. Effect of stimulation with G. duodenalis on transcription levels of
Fig. 2. Arithmetic mean stimulation index (SI ± st.dev.) of PBMC stimu-lated with G. duodenalis from six G. duodenalis infected calves. *P ≤ 0.05between pre-infection (week 0) and post-infection samples.
CD4+ �� T-cells was significantly higher compared to pre-infection levels (Fig. 3). The maximum proliferation ratio(4.5) was observed at week 13. Strong proliferation of CD3−
cells was occasionally seen, but this proliferation was neversignificantly different from pre-infection levels (Fig. 3).
3.3. Cytokine profile (qPCR)
The transcription level of mRNA for a panel of transcrip-tion factors and cytokines produced by CD4+ T-cells wasevaluated by RT qPCR on week 13 when CD4+ proliferationwas the highest. Mean fold changes of transcription levels
Please cite this article in press as: Grit, G.H., et al., Evaluation ofGiardia duodenalis infection in cattle. Vet. Parasitol. (2014), http
of cytokines and transcription factors in CD4+ T-cells werecompared to unstimulated cells (Fig. 4). Giardia stimulationresulted in significantly higher transcription of IL-17 (25.8-fold) and FoxP3 (6.4-fold). Transcription levels of TNF-�
Fig. 3. Arithmetic mean proliferation ratio (±st.dev.) of PBMC stimulated with
positive cells were divided in CD4+ and CD8+ T-cells. *P ≤ 0.05 between pre-infec
a panel of cytokines and transcription factors in CD4+ T-cells from six G.duodenalis infected calves. The Q values of stimulated cells were comparedto unstimulated cells. *P ≤ 0.05.
(13.5-fold), IL-15 (2.3-fold) and IL-2 (2.1-fold) were alsoincreased after stimulation with G. duodenalis, but thesedifferences were not statistically significant.
3.4. Thymidine proliferation assay with depleted PBMC
Proliferation of PBMC depleted from antigen-presentingcells, with and without MoDC stimulated with G. duode-nalis trophozoites, was measured every 2 weeks during20 weeks by the incorporation of thymidine in the DNAof dividing cells. No significant proliferation of depletedPBMC with or without MoDC was measured at several timepoints (Fig. 5), except in the co-culture of depleted PBMC
cellular and humoral systemic immune response against://dx.doi.org/10.1016/j.vetpar.2014.03.012
with MoDC at week 18. The proliferation of depleted PBMC,either with or without MoDC, was much lower comparedto the proliferation of PBMC (Fig. 2). A maximum stimula-tion index of depleted PBMC without MoDC (SI = 19.5) was
G. duodenalis from six G. duodenalis infected calves. T-cell receptor ��tion (week 0) and post-infection samples.
ARTICLE ING ModelVETPAR-7185; No. of Pages 11
G.H. Grit et al. / Veterinary Parasit
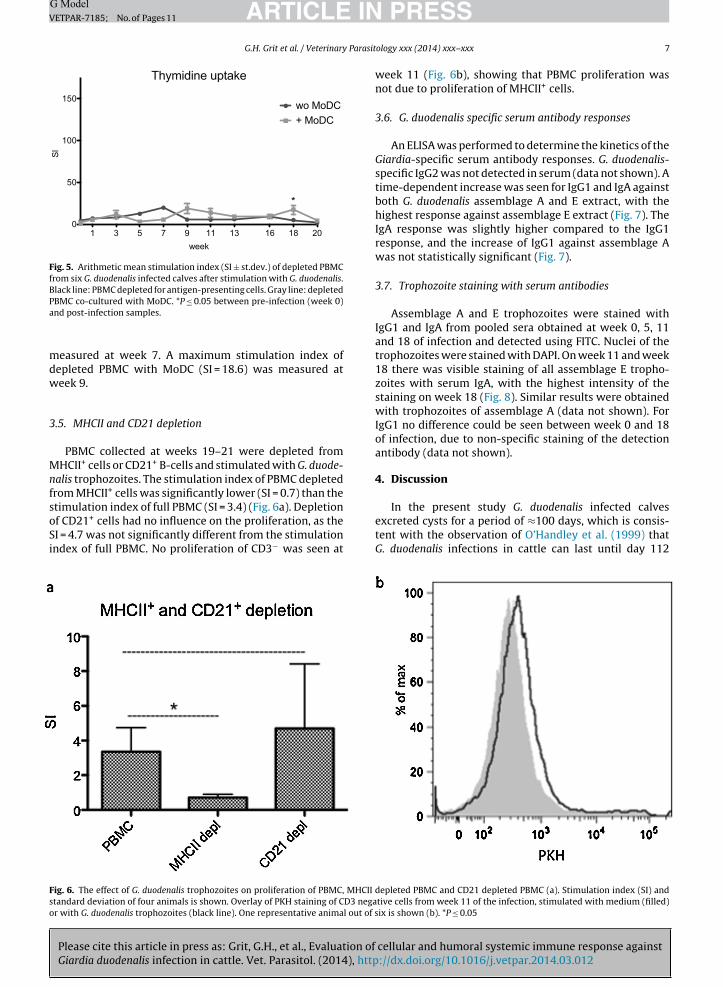
Fig. 5. Arithmetic mean stimulation index (SI ± st.dev.) of depleted PBMCfrom six G. duodenalis infected calves after stimulation with G. duodenalis.Black line: PBMC depleted for antigen-presenting cells. Gray line: depletedPa
mdw
3
MnfsoSi
In the present study G. duodenalis infected calves
Fso
BMC co-cultured with MoDC. *P ≤ 0.05 between pre-infection (week 0)nd post-infection samples.
easured at week 7. A maximum stimulation index ofepleted PBMC with MoDC (SI = 18.6) was measured ateek 9.
.5. MHCII and CD21 depletion
PBMC collected at weeks 19–21 were depleted fromHCII+ cells or CD21+ B-cells and stimulated with G. duode-
alis trophozoites. The stimulation index of PBMC depletedrom MHCII+ cells was significantly lower (SI = 0.7) than thetimulation index of full PBMC (SI = 3.4) (Fig. 6a). Depletion
Please cite this article in press as: Grit, G.H., et al., Evaluation ofGiardia duodenalis infection in cattle. Vet. Parasitol. (2014), http
f CD21+ cells had no influence on the proliferation, as theI = 4.7 was not significantly different from the stimulationndex of full PBMC. No proliferation of CD3− was seen at
ig. 6. The effect of G. duodenalis trophozoites on proliferation of PBMC, MHCII dtandard deviation of four animals is shown. Overlay of PKH staining of CD3 negar with G. duodenalis trophozoites (black line). One representative animal out of s
PRESSology xxx (2014) xxx–xxx 7
week 11 (Fig. 6b), showing that PBMC proliferation wasnot due to proliferation of MHCII+ cells.
3.6. G. duodenalis specific serum antibody responses
An ELISA was performed to determine the kinetics of theGiardia-specific serum antibody responses. G. duodenalis-specific IgG2 was not detected in serum (data not shown). Atime-dependent increase was seen for IgG1 and IgA againstboth G. duodenalis assemblage A and E extract, with thehighest response against assemblage E extract (Fig. 7). TheIgA response was slightly higher compared to the IgG1response, and the increase of IgG1 against assemblage Awas not statistically significant (Fig. 7).
3.7. Trophozoite staining with serum antibodies
Assemblage A and E trophozoites were stained withIgG1 and IgA from pooled sera obtained at week 0, 5, 11and 18 of infection and detected using FITC. Nuclei of thetrophozoites were stained with DAPI. On week 11 and week18 there was visible staining of all assemblage E tropho-zoites with serum IgA, with the highest intensity of thestaining on week 18 (Fig. 8). Similar results were obtainedwith trophozoites of assemblage A (data not shown). ForIgG1 no difference could be seen between week 0 and 18of infection, due to non-specific staining of the detectionantibody (data not shown).
4. Discussion
cellular and humoral systemic immune response against://dx.doi.org/10.1016/j.vetpar.2014.03.012
excreted cysts for a period of ≈100 days, which is consis-tent with the observation of O’Handley et al. (1999) thatG. duodenalis infections in cattle can last until day 112
epleted PBMC and CD21 depleted PBMC (a). Stimulation index (SI) andtive cells from week 11 of the infection, stimulated with medium (filled)ix is shown (b). *P ≤ 0.05

Please cite this article in press as: Grit, G.H., et al., Evaluation of cellular and humoral systemic immune response againstGiardia duodenalis infection in cattle. Vet. Parasitol. (2014), http://dx.doi.org/10.1016/j.vetpar.2014.03.012
ARTICLE IN PRESSG ModelVETPAR-7185; No. of Pages 11
8 G.H. Grit et al. / Veterinary Parasitology xxx (2014) xxx–xxx
Fig. 7. Optical density (OD) values for serum IgG1 and IgA antibodies against G. duodenalis extract of assemblage A and E, of six G. duodenalis infected calvesat six time points, during 18 weeks. *P ≤ 0.05 between pre-infection (week 0) and post-infection samples.
Fig. 8. Trophozoites of assemblage E stained with DAPI (blue) and IgA (green) from sera collected pre-infection (week 0) and week 5, 11 and 18 post-infection.Magnification 200×. (For interpretation of the references to colour in this figure legend, the reader is referred to the web version of this article.)

ING ModelV
Parasit
pwrbwic
l(iTws1ini
tTsiapAnl2iCalir2wcbiokbccadl(teirGR�etfTl
ARTICLEETPAR-7185; No. of Pages 11
G.H. Grit et al. / Veterinary
ost-infection. The calves suffered from periods of mild toatery diarrhoea but the general condition of the calves
emained well. It is possible that the diarrhoea was causedy G. duodenalis infection, given the high cyst counts thatere measured. Diarrhoea has previously been reported
n naturally infected calves with similar G. duodenalis cystounts (Geurden et al., 2006).
The chronic character of the infection might corre-ate with the slow immunological response seen in calvesO’Handley et al., 2003). In the present study systemicmmune responses were only observed after several weeks.he first in vitro cell proliferation of re-stimulated PBMCas seen after 5 weeks, while G. duodenalis specific
erum antibodies only increased significantly from week1 post-infection. This slow development of immunity is
n agreement with previous observations, where no intesti-al immune response was seen in calves after 3 weeks of
nfection (Dreesen et al., 2012).After 15 weeks of infection, a repeatedly low cyst excre-
ion indicated the development of protective immunity.his protective immunity may be associated with thetrong cellular responses that were observed during thenfection, although other mechanisms have a role in par-site clearance as well and it is possible that CD4+ T-cellslay a role in the initiation of these additional mechanisms.
long-term cellular immune response against G. duode-alis has also been reported in exposed humans, and was
argely driven by CD4+ T-cell activation (Hanevik et al.,011). Similarly, the proliferation of re-stimulated PBMC
n our experiment was mainly caused by CD4+ �� T-cells.D4+ T-cells can actively direct immune responses and playn important role in immunity against Giardia in mice. Miceacking CD4+ T lymphocytes are not able to clear infection,n particular CD4+ �� T-cells are required to control Gia-dia infection in mice (Singer and Nash, 2000; Zhou et al.,007). Therefore the cytokine mRNA profile of CD4+ T-cellsas evaluated. The fact that obvious up-regulation of some
ytokines (e.g. TNF-�) was not statistically significant, maye due to the small sample size and the large variabil-
ty between individual animals’ responses. Transcriptionf IL-17 was significantly (25.8-fold) upregulated. IL-17 isnown for its role in immunity at epithelial and mucosalarriers. Th17 cells have a pro-inflammatory function andan induce production of pro-inflammatory cytokines andhemokines, causing recruitment of effector cells, suchs neutrophils and macrophages (Gaffen, 2008). The pro-uction of IL-17 was described before in spleen cells and
ymph node cells obtained from G. duodenalis infected miceSolaymani-Mohammadi and Singer, 2011) and is essen-ial for clearance of G. muris infections in mice (Dreesent al., in press). Based on those studies and the present data,t can be hypothesised that IL-17 also plays an importantole in the development of protective immunity against. duodenalis in cattle. In most cases IL-17 is produced byOR�t+ Th17 cells, under the influence of IL-6 and TGF-. IL-6 is essential in G. duodenalis clearance in mice (Lit al., 2004). IL-6 causes upregulation of the expression of
Please cite this article in press as: Grit, G.H., et al., Evaluation ofGiardia duodenalis infection in cattle. Vet. Parasitol. (2014), http
he IL-23 receptor and directs T-cell development awayrom Treg towards a Th17 response (Kamda et al., 2012).he potential role of IL-17 in parasite clearance would ateast partially explain the necessity of IL-6. The fact that
PRESSology xxx (2014) xxx–xxx 9
mRNA levels of IL-6 or TGF-� were not upregulated in thisstudy may be due to the time of sampling of CD4+ T-cells,at 13 weeks post-infection. It is likely that IL-6 or TGF-�responses occur earlier in the infection. The transcriptionfactor FoxP3 was also significantly upregulated (6.4-fold).Although FoxP3 is typically expressed by regulatory T-cells,CD4+Foxp3+ IL-17 producing cells have been identified inhuman blood and lymphoid tissue. These cells display thefunctional features of both Treg and Th17 cells (Voo et al.,2009). Although it is possible that IL-17 induced by G. duo-denalis is produced by Foxp3+ Treg cells, it is also possiblethat the CD4+ population consists of both Foxp3+ Treg cellsand Th17 cells.
Dendritic cells (DC) are antigen-presenting cells thatare able to initiate T- and B-cell responses. In a previ-ous study (Grit et al., 2014) it was seen that G. duodenalisstimulated bovine MoDC were able to cause proliferationof autologous PBMC. In contrast, G. duodenalis-stimulatedbone marrow-derived DC from mice were suspected toactively interfere with the host innate immunity (Kamdaand Singer, 2009). Since proliferation was strongly reducedwhen PBMC were depleted by plastic adhesion in thepresent study, antigen presenting cells are required for theinduction of the cellular response. Co-culturing depletedPBMC with autologous MoDC could not restore the reducedproliferation of depleted PBMC to the level of proliferationseen in full PBMC, implying that DC are not the lead-ing cells in the induction of the cellular response. Nor doB-cells play a role in Giardia antigen presentation sincedepletion of B-cells alone (CD21+) from PBMC did notresult in decreased proliferation. However, the depletionof MHCIIhigh cells did cause significantly reduced prolif-eration of PBMC. Since the proliferating PBMC themselveswere not MHCIIhigh cells, the results indicate that Giardiatrophozoites have little or no direct effect on blood lympho-cytes and that antigen-presenting cells, other than DC andB-cells are required to initiate proliferation. Which MHCII+
cells are able to induce lymphocyte proliferation remainsunclear. Possible alternative antigen-presenting cells aremonocytes, macrophages, innate lymphoid cells or mastcells.
Serum antibodies appeared shortly before cyst countsstarted to decline, indicating that antibodies may beinvolved in the development of protective immunity. In thestudy of O’Handley et al. (2003) no significant increase ofserum IgG could be found in calves infected with G. duode-nalis until 8 weeks post-infection. In this study increasingG. duodenalis-specific serum IgG1 and IgA was measuredfrom week 11 post-infection and IgA responses seemedto be most important. B-cells and IgA are also requiredfor parasite clearance in mice. IgA was shown to be themost important isotype in the clearance of Giardia and isthe major isotype present in intestinal mucosal secretions(Stäger and Müller, 1997; Langford et al., 2002). In infectedhumans and mice, serum IgG and IgA increased as soon as 2weeks after infection (Daniels and Belosevic, 1994; Solimanet al., 1998; Velazques et al, 2005), which might explainwhy the eradication of the parasite in these hosts is much
cellular and humoral systemic immune response against://dx.doi.org/10.1016/j.vetpar.2014.03.012
faster compared to cattle.It was hypothesised that immunity is only developed
when all various surface proteins (vsp) on the Giardia

ING Model
Parasit
Velazques, C., Beltran, M., Ontiveros, N., Rascon, L., Figueroa, D.C.,
ARTICLEVETPAR-7185; No. of Pages 11
10 G.H. Grit et al. / Veterinary
surface are recognised (Faubert, 2000), and that the slowdevelopment of immunity against Giardia is the result ofthis antigen variation (Müller and Von Allmen, 2005). How-ever, in our study all trophozoites, and not only a subset,were recognised by serum IgG1 and IgA, and the numberof binding antibodies increased over time during the infec-tion. This suggests that a broad repertoire of antibodies isinduced against all vsp or that the antibody response incattle is at least partly directed against invariable surfaceantigens. This data implies that parasite clearance mightdepend on the antibody concentration.
In conclusion, the slow induction of cellular andhumoral responses was in line with the chronic characterof G. duodenalis infections in cattle. The IL-17 production byCD4+ T-cells might play a role in the development of pro-tective immunity against G. duodenalis. Further research isnecessary to determine the role of IL-17 and to determinewhether IL17 is produced by FoxP3+ cells or by a differ-ent CD4+ T-cell population. Giardia-specific IgG1 and IgA inthe serum increased from week 11 post-infection and alltrophozoites were stained by serum antibodies from week11 p.i. onward, suggesting that antibodies are directedagainst all variant-specific surface proteins or that the anti-body response in cattle is at least partly directed againstinvariable surface antigens.
Acknowledgements
G.H. Grit was funded by a Ph.D. grant from the Insti-tute for the Promotion of Innovation through Science andTechnology in Flanders (IWT-Vlaanderen). S. Van Copper-nolle is supported by EU FP7 project 265862 (Paravac). TheHercules Foundation is acknowledged for its financial sup-port to purchase research equipment (FACSAriaIII). AxenicG. duodenalis assemblage E cultures were obtained from L.Maes and E. Bénéré, from the laboratory of Microbiology,Parasitology and Hygiene, University of Antwerp. PorcinerGM-CSF was a gift from S. Inumaru from the Labora-tory of Bioengineering, National Institute of Animal Health,Tsukuba, Ibaraki, Japan.
References
Daniels, C.W., Belosevic, M., 1994. Serum antibody responses by male andfemale C57BI/6 mice infected with Giardia muris. Clin. Exp. Immunol.97, 424–429.
Dreesen, L., Rinaldi, M., Chiers, K., Li, R., Geurden, T., Van den Broeck,W., Goddeeris, B., Vercruysse, J., Claerebout, E., Geldhof, P., 2012.Microarray analysis of the intestinal host response in Giardia duode-nalis assemblage E infected calves. PLoS One 7 (7), e40985.
Dreesen, L., De Bosscher, K., Grit, G., Staels, B., Lubberts, E., Bauge,E., Geldhof, P., 2014. Giardia muris infection in mice is associatedwith a protective IL-17A response and activation of the peroxisomeproliferator-activated receptor alpha. Infect. Immun. (in press).
Faubert, G., 2000. Immune response to Giardia duodenalis. Clin. Microbiol.Rev. 13, 35–54.
Gaffen, S.L., 2008. An overview of IL-17 function and signaling. Cytokine43, 402–407.
Geurden, T., Claerebout, E., Vercruysse, J., Berkvens, D., 2004. Estimationof diagnostic test characteristics and prevalence of Giardia duodenalisin dairy calves in Belgium using a Bayesian approach. Int. J. Parasitol.
Please cite this article in press as: Grit, G.H., et al., Evaluation ofGiardia duodenalis infection in cattle. Vet. Parasitol. (2014), http
34, 1121–1127.Geurden, T., Claerebout, E., Dursin, L., Deflandre, A., Bernay, F., Kalt-
satos, V., Vercruysse, J., 2006. The efficacy of an oral treatmentwith paramomycin against an experimental infection with Giardia incalves. Vet. Parasitol. 135, 241–247.
PRESSology xxx (2014) xxx–xxx
Geurden, T., Geldhof, P., Levecke, B., Martens, C., Berkvens, D., Casaert, S.,Vercruysse, J., Claerebout, E., 2008. Mixed Giardia duodenalis assem-blage A and E infections in calves. Int. J. Parasitol. 38, 259–264.
Geurden, T., Vercruysse, J., Claerebout, E., 2010a. Is Giardia a significantpathogen in production animals? Exp. Parasitol. 124, 98–106.
Geurden, T., Vandenhoute, E., Pohle, H., Cassaert, S., De Wilde, N.,Vercruysse, J., Claerebout, E., 2010b. The effect of a fenbendazoletreatment on cyst excretion and weight gain in calves experimentallyinfected with Giardia duodenalis. Vet. Parasitol. 169, 18–23.
Grit, G.H., Van Coppernolle, S., Devriendt, B., Geurden, T., Hope, J., Ver-cruysse, J., Cox, E., Geldhof, P., Claerebout, E., 2014. Giardia duodenalisstimulates partial maturation of bovine dendritic cells associated withaltered cytokine secretion and induction of T-cell proliferation. Para-site Immunol 36, 157–169.
Hanevik, K., Kristoffersen, E., Svard, S., Bruserud, O., Ringqvist, E., Sørnes, S.,Langeland, N., 2011. Human cellular immune response against Giardialamblia 5 years after acute giardiasis. J. Infect. Dis. 204, 1779–1789.
Inumaru, S., Kohuho, T., Denham, S., Deyer, M.S., Momotani, E., Kita-mura, S., Corteyn, A., Brookes, S., Parkhouse, R.M., Takamatsu, H., 1998.Expression of biologically active recombinant porcine GM-CSF by bac-ulovirus gene expression system. Immunol. Cell Biol. 76, 195–201.
Kamda, J.D., Singer, S.M., 2009. Phosphoinositide 3-kinase-dependentinhibition of dendritic cell interleukin-12 production by Giardia lam-blia. Infect. Immun. 77, 685–693.
Kamda, J.D., Nash, T.E., Singer, S.M., 2012. Giardia duodenalis: dendritic celldefects in IL-6 deficient mice contribute to susceptibility to intestinalinfection. Exp. Parasitol. 130, 288–291.
Keister, D.B., 1983. Axenic culture of Giardia lamblia in TYI-S-33 mediumsupplemented with bile. Trans. R. Soc. Trop. Med. Hyg. 77, 487–488.
Langford, T.D., Housley, M.P., Boes, M., Chen, J., Kagnoff, M.F., Gilin, F.D.,Eckmann, L., 2002. Central importance of immunoglobulin A in hostdefence against Giardia spp. Infect. Immun. 70, 11–18.
Li, E., Zhou, P., Petrin, Z., Singer, S.M., 2004. Mast cell-dependent controlof Giardia lamblia infections in mice. Infect. Immun. 72, 6642–6649.
Lujan, H.D., Svärd, S., 2011. Giardia—A Model Organism. Springer, NewYork, pp. 420.
Müller, N., Von Allmen, N., 2005. Recent insights into the mucosal reac-tions associated with Giardia lamblia infections. Int. J. Parasitol. 35,1339–1347.
O’Handley, R.M., Cockwill, C., McAllister, T.A., Jelinski, M., Morck, D., Olson,M.E., 1999. Duration of naturally acquired giardiasis and cryptosporid-iosis in dairy calves and their association with diarrhea. J. Am. Vet.Med. Assoc. 214, 391–396.
O’Handley, R.M., Ceri, H., Anette, C., Olson, M.E., 2003. Passive immu-nity and serological immune response in dairy calves associated withnatural Giardia duodenalis infections. Vet. Parasitol. 133, 89–98.
Prucca, C.G., Slavin, I., Quiroga, R., Elias, E.V., Rivero, F.D., Sauro, A., Car-ranza, P.G., Lujan, H.D., 2008. Antigenic variation in Giardia lamblia isregulated by RNA interference. Nature 456, 750–754.
Ruest, N., Couture, Y., Faubert, G.M., Girard, C., 1997. Morphologicalchanges in the jejunum of calves naturally infected with Giardia spp.and Cryptosporidium spp. Vet. Parasitol. 69, 177–186.
Singer, S.M., Nash, T.E., 2000. T-cell-dependent control of acute Giardialamblia infections in mice. Infect. Immun. 68, 170–175.
Solaymani-Mohammadi, S., Singer, S.M., 2011. Host immunity andpathogen strain contributes to intestinal disaccharidase impairmentfollowing gut infection. J. Immunol. 187, 3769–3775.
Soliman, M.M., Taghi-Kilani, R., Abou-Shady, A.F., El-Mageid, S.A., Han-douse, A.A., Hegazi, M.M., Belosevic, M., 1998. Comparison of serumantibody responses to Giardia lamblia of symptomatic and asymp-tomatic patients. Am. J. Trop. Med. Hyg. 58, 232–239.
Stäger, S., Müller, N., 1997. Giardia lamblia infections in B-cell-deficienttransgenic mice. Infect. Immun. 65, 3944–3946.
Sulaiman, I.M., Fayer, R., Bern, C., Gilman, R.H., Trout, J.M., Schantz, P.M.,Das, P., Lal, A.A., Xiao, L., 2003. Triosephosphate isomerase gene char-acterization and potential zoonotic transmission of Giardia duodenalis.Emerg. Infect. Dis. 9, 1444–1452.
Taherkhani, H., Hajilooi, M., Fallah, M., Khyanbanchi, O., Haidari, M., 2009.Gene polymorphism in transforming growth factor-beta codon 10 isassociated with susceptibility to giardiasis. Int. J. Immuogenet. 36,345–349.
Taminelli, V., Ecker, J., Sydler, T., Gottstein, B., Corboz, L., Hofmann, M.,1989. Experimental infection of calves and sheep with bovine Giardiaisolates. Schweiz. Arch. Tierheilkd. 131, 551–564.
cellular and humoral systemic immune response against://dx.doi.org/10.1016/j.vetpar.2014.03.012
Granados, A.J., Hernandez-Martinez, J., Hernandez, J., Astiazaran-Garcia, H., 2005. Giardia lamblia infection induces different secretoryand systemic antbody responses in mice. Parasite Immunol. 27,351–356.

ING ModelV
Parasit
V
W
Xiao, L., Herd, R.P., 1994. Infection pattern of Cryptosporidium and Giardiain calves. Vet. Parasitol. 55, 257–262.
ARTICLEETPAR-7185; No. of Pages 11
G.H. Grit et al. / Veterinary
oo, K.S., Wang, Y., Santori, F.R., Boggiano, C., Wang, Y., Arima, K., Bover,L., Hanabuchi, S., Khalili, J., Marinnova, E., Zheng, B., Littman, D.R., Liu,
Please cite this article in press as: Grit, G.H., et al., Evaluation ofGiardia duodenalis infection in cattle. Vet. Parasitol. (2014), http
Y., 2009. Identification of IL-17 producing FOXP3+ regulatory T cellsin humans. Proc. Natl. Acad. Sci. U.S.A. 106, 4793–4798.
erling, D., Hope, J.C., Chaplin, P., Collins, R.A., Taylor, G., Howard, C.J.,1999. Involvement of caveolae in the uptake of respiratory syncytialvirus antigen by dendritic cells. J. Leukoc. Biol. 66, 50–58.
PRESSology xxx (2014) xxx–xxx 11
cellular and humoral systemic immune response against://dx.doi.org/10.1016/j.vetpar.2014.03.012
Zhou, P., Li, E., Shea-Donohue, T., Singer, S.M., 2007. Tumor necrosis factoralpha contributes to protection against Giardia lamblia infection inmice. Parasite Immunol. 29, 367–374.