estudio de la dinámica poblacional de la comunidad...
TRANSCRIPT

1
Fa
cultad d
e C
iencia
s E
xperim
enta
les
UNIVERSIDAD DE JAÉN Facultad de Ciencias Experimentales
Trabajo Fin de Grado
Alumno: Ricoy Llavero Enrique
Septiembre, 2015
Estudio de la dinámica poblacional de la
comunidad de simúlidos en la Sierra de Segura.

2
UNIVERSIDAD DE JAÉN
FACULTAD DE CIENCIAS EXPERIMENTALES
Grado en Biología
Trabajo Fin de Grado
Estudio de la dinámica poblacional de la
comunidad de simúlidos en la Sierra de Segura.
Enrique Ricoy Llavero
Jaén, a Septiembre de 2015

3
INDICE
1. RESUMEN ................................................................................................... 5
2. ABSTRACT ................................................................................................. 5
3. INTRODUCCIÓN ......................................................................................... 6
3.1.Biología .................................................................................................... 6
3.1.1. Ciclo de vida .......................................................................................... 6
3.1.1.1. Ovoposición ........................................................................................ 7
3.1.1.2. Desarrollo larvario ............................................................................... 7
3.1.1.3. Metamorfosis ...................................................................................... 8
3.1.1.4. Rotura de la Pupa y dispersión ........................................................... 8
3.1.1.5. Cópula ................................................................................................. 8
3.2. Alimentación ............................................................................................ 9
3.2.1. Alimentación de las larvas ..................................................................... 9
3.2.2. Alimentación de Adultos ........................................................................ 9
3.3. Morfología ................................................................................................ 9
3.3.1. Huevo ..................................................................................................... 9
3.3.2. Larvas ................................................................................................... 10
3.3.3. Pupas.................................................................................................... 13
3.3.4. Imagos ................................................................................................. 15
3.4. Relevancia del estudio de los simúlidos ……………………….....…… 16
3.4.1. Interés científico ................................................................................... 16
3.4.2. Interés sanitario .................................................................................... 17
3.4.2.1. Directos ............................................................................................ 17
3.4.2.2. Vector ............................................................................................... 17
4. OBJETIVOS ............................................................................................... 19
5. MATERIAL Y MÉTODOS .......................................................................... 19
5.1. Áreas de estudio ................................................................................... 19
5.1.1. Arroyo Orcera ...................................................................................... 19
5.1.2. Arroyo de los Molinos ......................................................................... 20
5.1.3. Río Trujala ........................................................................................... 21
5.2. Método de captura ................................................................................ 25
5.2.1. Muestreo .............................................................................................. 25
5.2.2. Recogida de Muestras ........................................................................ 26
5.4. Tratamiento de las hojas ...................................................................... 27

4
5.5. Determinación taxonómica ................................................................. 28
5.6. Índices de diversidad ........................................................................... 29
5.7. Material .................................................................................................. 30
6. RESULTADOS .......................................................................................... 31
6.1. Resultados generales ....................................................................... 31
6.2. Resultados comparativos ................................................................. 35
6.3. Resultados por periodo de influencia ............................................... 38
6.4. Resultados índices de biodiversidad ............................................... 40
7. DISCUSIÓN ............................................................................................... 41
8. CONCLUSIÓN .......................................................................................... 46
9. BIBLIOGRAFÍA ......................................................................................... 47
9.1 Bibliografía con enlaces ........................................................................ 50

5
1. RESUMEN
Este trabajo contribuye al conocimiento faunístico de la familia Simuliidae en el
noroeste de la provincia de Jaén. Se proporcionan datos, mediante la captura de
ejemplares larvarios y pupas, de distribución y dinámica poblacional de las especies
que se encuentran en la región geográfica Sierra de Segura sur, concretamente en
los siguientes cauces fluviales: Arroyo Molinos, Arroyo Orcera y Río Trujala. Las
especies que encontramos son: Simulium (Eusimulium) velutinum, S. (E.) petricolum,
S. (Nevermania) cryophilum, S. (Tetisimulium) bezzi, S. (Wilhemia) pseudoequinum,
S. (Simulium) xanthinum, S. (S.) argireatum, S. (S.) ornatum y S. (S.) intermedium.
Siendo estas dos últimas especies las más abundantes.
Palabras clave: Simúlidos, Simuliidae, Diptera, Dinámica poblacional, Faunística,
Península Ibérica y Sierra de Segura.
2. ABSTRACT
This essay helps to know some characteristics about Simuliidae family. Some
knowledge are given through the capture of some specimen (larvae and pupae)
concerning several species that are found in south geographical region of Sierra de
Segura (Jaén, Spain), affecting the following fluvial waterways: Molinos stream,
Orcera stream and Trujala river.
The species that we can find in these places are: Simulium (Eusimulium) velutinum,
S. (Eusimulium) petricolum, S. (Nevermania) cryophilum, S. (Tetisimulium) bezzi, S.
(Wilhemia) pseudoequinum, S. (Simulium) xanthinum, S. (S.) argireatum, S. (S.)
ornatum and S. (S.) intermedium. The two last species are the most prevalent ones.
Key words: Black flies, Simuliidae, Diptera, Population dynamics, Faunistic, Iberian
Peninsula and Sierra de Segura.

6
3. INTRODUCCIÓN.
Los Simúlidos representan una familia perteneciente al suborden Nematócera,
englobado dentro del orden Dípteros, que esta taxonómicamente enmarcado en la
clase Insectos.
La morfología del individuo adulto se caracteriza por su aspecto compacto, con
“joroba” en el tórax y de tamaño pequeño, así como por su coloración oscura, lo que
ha servido para denominarlo vulgarmente en inglés como "black -flies" o "bull-flies",
mosca negra o mosca toro.
Esta familia de insectos es especialmente importante por tratarse de vectores de
agentes patógenos causantes de diversas enfermedades (p.e. oncocercosis o
filariosis) (Shelley & Coscarón, 2001). En España, dichas enfermedades no aparecen
de manera normal en humanos, mientras que las enfermedades que se dan en
animales domésticos tienen un menor riesgo para su salud si se tratan
adecuadamente. Esto provoca que a nivel peninsular no haya tantos trabajos sobre
simúlidos en relación a los patógenos que portan, sino más bien son trabajos de
carácter taxonómico y ecológico, que implican la determinación de las especies
presentes en distintas áreas geográficas (Lestón et al., 2013).
El primer trabajo relacionado con estos dípteros lo realizo el entomólogo alema
Johann Wilhelm Meigenen 1800, y en él describió el género Melusina. A pesar de ello
la ICZN (Internacional Comisión of Zoological Nomenclature) acordó no usar dicho
nombre para nombrar a este género y se decidió por el de Simulium, utilizado por
Latreilleen 1802. De este nombre genérico procede el nombre actual de la familia.
Posteriormente, se escriben los primeros monográficos sobre la familia Simuliidae, en
1940 Smart y en 1956 Rubtsov.
3.1. Biología. 3.1.1. Ciclo de vida. El ciclo de vida de los simúlidos está marcado, como en la mayoría de la clase
insecta, por cuatro fases bien diferenciadas: huevo, larva, pupa e imago. La etapa
larvaria suele presentar un número variable de fases larvarias (Moor, 1982). El
número de generaciones que se pueden dar por año varía entre una (género
Prosimulium) y hasta cinco, como aparece descrito para Simulium erytrhocephalum

7
(Post, 1983). El número de generaciones anuales depende de las distintas
condiciones ambientales que se den localmente.
3.1.1.1. Ovoposición. Las hembras depositan los huevos entre siete días u ocho días después de su
fecundación. Las puestas son variables en número, pudiendo oscilar entre los 200 y
los 300 huevos (Rivosecchi, 1978). El tiempo de eclosión es muy variable según la
región donde nos encontremos y en algunos casos se da diapausa (Golini & Davies,
1987).
La puesta puede producirse de varias formas: en sedimentos algo húmedos, el
musgo fresco, en zonas encharcadas, sobre vegetación colgante de las orillas, o
directamente en el agua. Cabe señalar que aquellas puestas que se realizan "fuera
de agua" eclosionan al poco tiempo (Colbo & Wotton, 1981). Podemos distinguir dos
métodos de puesta en el agua, (i) dejarlos caer en la superficie y que estos se
hundan, o (ii) la hembra se sumerge directamente en el agua y coloca la puesta
sobre el sustrato (Golini & Davies, 1987). Las puestas depositadas en agua que
permanecen ancladas al sustrato gracias a la presencia de una cubierta gelatinosa.
Este tipo de puestas es la más frecuente y tienen una característica que los hace
más eficientes: pueden permanecer en diapausa durante un periodo de tiempo largo
hasta que se den las condiciones adecuadas. La diapausa es distinta según la
especie, pudiendo encontrar especies que muestran diapausa en el periodo invernal,
mientras que otras lo presentan en el periodo primo-estival. También se ve
influenciada por la cantidad de alimento disponible para las larvas cuando emerjan
(Zwick & Zwick, 1990).
3.1.1.2. Desarrollo larvario. La eclosión de la larva se produce gracias a la existencia de una espina en la
cápsula cefálica que provoca la ruptura de la cubierta del huevo. Cuando la larva se
encuentra libre su reacción es la de anclarse al sustrato, aquel donde tenga unas
buenas condiciones de corriente y alimentación, la corriente principalmente viene
también favoreciendo la cantidad de alimento (Craig & Chance, 1982).
La larva presenta diferentes estadios que varían entre las distintas especies. Se ha
expuesto que podría existir en torno a 6-9 estadios (Colbo & Wotton, 1981; Moor,
1982). La duración de la fase larvaria no está bien definida y depende de dos puntos
importantes, las condiciones ambientales en las que se encuentran y la especie. Por

8
ello podemos ver como en zonas templadas esta fase puede durar algunas semanas
en periodos favorables y meses en periodos desfavorables, mientras que en zonas
tropicales puede durar incluso una única semana. Se sabe que las larvas además
están afectadas por la temperatura, de este modo aquellas que crecen en invierno
son más grandes que las que se desarrollan en verano y del mismo modo aquellas
que se encuentran más al norte (o más cercanas a los polos) suelen ser más
grandes que las del ecuador. (Crosskey, 1973)
3.1.1.3. Metamorfosis. La larva cuando se encuentra bien desarrollada, significa que presenta ya el
histoblasto (filamentos respiratorios) (Figura 1) bien formado, en los laterales del
tórax y están presentes los ganchos del dorso del abdomen, empieza a segregar
sobre si seda, a la que va dando conformación de estuche. La duración de esta fase
ronda entre los pocos días hasta las dos semanas. (Gracio, 1985).
3.1.1.4. Ruptura de la pupa y dispersión. El imago consigue emerger de la pupa rajando el tórax de esta, acto seguido sale
rápidamente del medio acuático y se posa en un sustrato cercano con el fin de
endurecer su cutícula, en el momento de emerger esta es flexible (Cupp, 1981). Los
machos se suelen agrupar en enjambres, mientras que las hembras son más
solitarias.
3.1.1.5. Cópula. Esta puede realizarse bien en los enjambres de machos formados a tal efecto o en
las áreas de descanso para la especie, aquellos donde descansan o se alimentan.
La cópula suele durar entre medio minuto y dos, es el tiempo necesario para que se
transfiera el espermatóforo (Wenk, 1987).
Según sea necesario o no la previa ingesta de sangre para la maduración de sus
huevos diferenciamos dos tipos de especies: (i) las especies anautógenas, que
necesitan la ingesta de sangre para realizar la puesta de huevos y (ii) las especies
autógenas, donde la maduración de los huevos se puede dar sin ingerir previamente
sangre.

9
3.2. Alimentación. 3.2.1. Alimentación de las larvas. Las larvas de la familia Simuliidae son, en su mayoría, filtradoras gracias a la
presencia de dos abanicos formados por radios, con pequeñas sedas que recuerdan
a un peine, este aparato son las premandíbulas modificadas (Figura 2). Estas son
móviles permitiéndole así su apertura y cierre, suelen estar en dos posiciones: (i)
una primera abierta que permite la captura de nutrientes y (ii) cierre por lo que no se
alimentan. Con ayuda de las mandíbulas pueden limpiar los radios y así alimentarse.
(Ross &Craig, 1980; Craig &Chance, 1982; Currie & Craig, 1987).
Se da el caso de especies que no son filtradoras porque no presentan las
premandíbulas modificadas. En este caso se alimentan de perifiton, materia orgánica
particulada (p.e. especies del genero Prosimulium en los primeros estadios
larvarios). Hay casos en los que actúen como depredadoras de otros organismos
como quironómidos, efemenópteros, o incluso darse canibalismo, pero este tipo de
alimentación no es muy común (Currie & Craig 1987).
3.2.2. Alimentación de Adultos. Los imagos se alimentan del néctar de las flores que les dan el sustento nutritivo
para poder realizar todo su metabolismo vital, tanto hembras como machos se
alimentan de néctar.
Aquellas hembras de especies que requieren la ingesta de sangre para la
maduración de los huevos (anautógenas), presentan las piezas bucales adaptadas
para el rasgado y la ingesta de sangre del huésped, siendo estos aves y mamíferos,
entre ellos los seres humanos (McIver & Sutcliffe, 1987).
3.3. Morfología.
3.3.1. Huevo. El huevo tiene forma subtriangular y su tamaño varía entre 0.1-0.5 mm (Gracio,
1985). Sin adornos en su cubierta y recubiertos de una secreción viscosa que los
adhiere al sustrato (Foto 1).

10
Foto 1. Se puede ver un cúmulo de huevos y su tamaño en comparación a unas pinzas antimagnéticas.
3.3.2. Larvas. Presentan una forma alargada tubular con dos engrosamientos, uno en la parte
anterior del tórax, y otro en la parte posterior del abdomen. Su tamaño oscila entre
aproximadamente un milímetro, y hasta los 15 milímetros aproximadamente
(Martínez, 1996).
Presentan una cabeza de morfología cuadrangular y dura, gracias a la existencia de
escleroproteinas. En la parte superior de esta están las dos premandíbulas o
también denominado abanico cefálico primario (Figura2). Por debajo encontramos la
mandíbula, el palpo maxilar, la maxila, el labio-hipofaringe, el submentón (Figura 2),
el puente postgenal, la: hendidura postgenal, y el palpo tensorial (Figura 2).En la
mandíbula podemos ver una pequeña dentición característica denominada proceso
"tp" (Figura 3). Muchas de estas características son utilizadas para la identificación
de diferentes especies. También poseen dos antenas de cuatro segmentos
(contando la base), siendo el cuarto segmento el que variará entre distintas especies
(Figura 2). En el vientre existe una hendidura ventral (Figura 2) que puede variar en
su profundidad o morfología dependiendo de la especie. En el dorso de los
individuos se encuentra el frontoclípeo (Figura 3), este presenta una coloración
característica, debida a las manchas que provoca la inserción de la musculatura en
la cabeza, es distinta en cada especie.
El tórax se encuentra subdividido en tres segmentos anchos, aunque externamente
no se encuentra bien diferenciadas. Aspecto característico es la existencia de una
prolongación, el denominado pseudópodo ventral (Figura 1), esta está dotada de
una corona de ganchos, cuya finalidad es la de sujeción y desplazamiento. Podemos

11
encontrar en los estadios larvarios más avanzados los histoblastos de los filamentos
respiratorios. Estos se encuentran uno a cada lado del tórax (Figura 1). Constituyen
un elemento anatómico que liga directamente las larvas con las pupas, ya que
ambos siempre presentaran el mismo número de filamentos.
El abdomen aparece dividido en ocho segmentos, que están algo más delimitados y
más visibles que en el tórax, dichos segmentos van aumentando de tamaño según
avanzamos a la parte posterior del individuo. En el extremo inferior del abdomen
están presentes las branquias anales, en el dorso, que dependiendo de la etapa,
presentaran un menor o mayor desarrollo, estas además son variables según la
especie, podemos encontrar desde branquias simples, hasta ramificadas o
compuestas. Debajo de las branquias anales queda el denominado esclerito en "X",
(Figura 1) estructura de quitina que permite el anclado o fijación de la musculatura
abdominal.
En la parte ventral del abdomen existe otro órgano de sujeción, consiste en un
conjunto de ganchos distribuidos de forma cilíndrica, actúan a modo de ventosa.
Además algunas especies presentan las denominadas papilas, que son unas
pequeñas protuberancias en la parte ventral, o desplazada hacia el lateral, del
individuo.
La pigmentación que presenta el cuerpo puede ser muy variable entre distintas
especies, pudiendo variar desde tonos blancos, casi transparentes, hasta oscuros o
marrones. Los tejidos externos o cutícula son translucidos, lo que permiten observar
el interior, siendo apreciable tanto el tubo digestivo como las glándulas salivares.
Además permite visualizar si son portadores de patógenos de cierto tamaño ya que
también pueden ser visibles.

12
Figura 1. Representación de la larva de simúlido, presencia del esclerito
"X".González, 1997.
Figura 2. Representación de la cabeza y sus partes: cf: abanico cefálico primario, sf:
abanico cefálico secundario, m: mandíbula, mp: palpo maxilar, mx: maxila, lh: labio-
hipofaringe, h: hipostomio, pb: puente postgenal, pc: hendidura postgenal, t: palpo
tentorial.Antena, p: proximal, me: media, d: distal. Programa de control de simúlidos
hematófogos en la provincia de Arica”.

13
Figura 2. Representación cabeza de la larva y imagen aumentada de la mandíbula
junto a su proceso "tp". González, 1997.
3.3.3. Pupas. Es la fase predecesora de los adultos. El individuo se encuentra en el estadio
larvario, cuando dicha larva está muy desarrollada empieza a segregar una
sustancia sedosa, a la que da forma de capsula pupal, en torno a su cuerpo y fijada
al sustrato. A esta formación se le suele adherir granos del sustrato.
Presentan una morfología muy característica, en forma de medio cono. El extremo
anterior es amplio, pudiendo presentar rasgos morfológicos propios de cada especie:
rebordes, protuberancias alargadas en la parte dorsal o un talón delantero. La
cápsula puede adquirir diferentes formas según la especie (Lechevalier, 1953)
(Figura 4). Particularmente, las especies del género Prosimulium presenta
morfologías irregulares que las diferencian de otros géneros de la familia.
De la cápsula pupal sobresalen la cabeza y los filamentos respiratorio son originados
a partir del histoblasto larvario. Los filamentos respiratorios varían en número, desde
cuatro hasta incluso más de diez en especies del género Prosimulium. Las pupas se
disponen de espaldas a la corriente con el fin de proteger sus filamentos
respiratorios y de facilitar la emergencia del adulto, una vez completada la
metamorfosis (Martínez, 1996).
La pupa propiamente dicha se diferencia en dos partes: (i) cabeza y tórax, que se
encuentra parcialmente descubiertas por la capsula pupal, (ii) y el abdomen.

14
Del tórax parten los dos filamentos respiratorios, que posteriormente se ramifican
dando lugar a distintas morfologías. Podemos encontrar, no en todas las especies,
en la cutícula dorsal de ambas, pequeños tubérculos, espinas o finos pelos (Gracio,
1985) (Figura 4).
En el abdomen se diferencian 9 segmentos (Gracio, 1985).
Figura 4. Fotografía de una pupa de la familia Simuliidae. www.eutaxa.com
Foto 2. Se puede ver una larva de S. (S.) ornatum sin su estuche.
3.3.4. Imagos. Presentan una morfología muy característica cuando son adultos.

15
Los ojos presentan un dimorfismo sexual, los machos los presentan de mayor
tamaño, lo que provoca que ambos ojos contacten a la altura frontal (definidos como
ojos de tipo holóptico). Por el contrario, en las hembras, estos son de menor tamaño,
no llegando a contactar entre ellos (tipo dicóptico).Mientras que los ojos de las
hembras presentan facetas que siempre son iguales, los machos presentan dos
tipos de facetas, unas de mayor tamaño (situadas en la parte superior) y otras de
menor tamaño (dispuestas en la parte inferior) (Figura 5).
Tienen antenas cortas, pero que si se encuentran muy divididas. Sus partes se
denominan escapo, pedicelo y flagelo, dividiéndose este último en unos siete, ocho o
nueve flagelómeros.
Existe distinción entre la cabeza de machos y de hembras, no solo por lo
anteriormente expuesto sobre ojos, si no también por su aparato bucal. Las hembras
pueden ser picadoras o no, dependiendo de que lo sean presentaran una morfología
bucal distinta, lo que causa un cambio general de la cabeza. Esto no ocurre en los
machos pues no son picadores.
Podemos distinguir claramente dos tipos de especies, aquellas que no son picadoras
(autógenas), y aquellas que si pican (anautógenas).
El escudo del tórax se encuentra muy desarrollado, mientras que partes como el
escutelo y post escutelo, situadas tras el escudo, se encuentran poco desarrolladas.
También en el tórax, parte ventral, presenta partes bien diferenciadas como son el
surco mesoepisternal, robusto, y la membrana pleural. Además tienen en cada
lateral del tórax un espiráculo.
El abdomen se encuentra claramente diferenciado en nueve segmentos. Los más
distinguibles son el primero, es el más delgado, por la presencia en el de un anillo de
pelillos, y por otro lado los últimos, ya que en ellos se encuentra la genitalia o
aparatos copuladores. En las hembras en el octavo segmento, presentan dos
láminas que actúan como lóbulos de ovoposición (gonapófisis). En el interior se
encuentra una pieza quitinizada en forma de "Y", la furca. La genitalia de los machos
se caracteriza por la reducción del noveno segmento, el esternito, existe una gran
variedad de estructuras que pueden variar según la especie (coxitos, estilos,
gonofurca, espinas paramerales y gonosterno o lámina media).
Continuamos con las extremidades o apéndices. Los apéndices terrestres se
encuentran subdivididas en nueve segmentos, coxa y trocánter los basales, los más
desarrollados y largos son el fémur y tibia que les continúan a los anteriormente

16
mencionados, y por último cinco artejos tarsales. Al final presentan unas uñas
simples, diseñadas para aferrarse a los individuos.
Por otro lado podemos encontrar las alas, son transparentes y presentan poca
venación. (González, 1997)
Figura 5. Representación de un imago y sus partes. Diferencia entre macho y adulto
en ojos. González, 1997.
3.4. Relevancia del estudio de los simúlidos. 3.4.1. Interés científico Este trabajo se inscribe en una línea de investigación desarrollada en el Área de
Zoología del Departamento de Biología Animal, Biología Vegetal y Ecología
enfocada al estudio de los artrópodos vectores. En el caso de los simúlidos, en
sentido amplio, las investigaciones que se están desarrollando afectan aspectos
relativos a la taxonomía y distribución de las especies del grupo, filogenia molecular,
papel vectorial en la transmisión de agentes patógenos (bacterias, protozoos,
nematodos filaroideos) que transmiten a distintos grupos de vertebrados y
enfermedades fúngicas y parasitarias que afectan a estos y que pueden ser

17
utilizadas para el control de este grupo de organismos (Gordon et al., 1973;
Gradinarov, 2014).
3.4.2. Interés sanitario 3.4.2.1. Directos. Como se recordará, los simúlidos son insectos que pueden picar. Las propiedades
tóxicas de algunos componentes de la saliva de estos dípteros, determina que la
picadura origine problemas de salud tanto en humanos como en el ganado. De este
modo, la concurrencia de estas picaduras puede provocar la muerte de ganado
(Cupp, 1987; Cupp & Cupp 1997). En el caso del hombre, las picaduras de
determinadas especies, pueden provocar una intoxicación del flujo sanguíneo,
dándose los siguientes síntomas: Aceleración de la circulación, disnea, hipertermia e
hipotermia, trastornos nerviosos e incluso la muerte (Noirtin et al., 1979). Entre
dichas especies se encuentra Simulium (S.) ornatum, una especie habitual que
encontraremos, S. (S.) reptans, S. (Boophthora) erythrocephalum, S. (Wilhelmia)
lineatum y S. (W.) equinum, todas ellas muy agresivas y de amplia distribución en
Europa occidental (Beaucournu-Saguet et al., 1990).
3.4.2.2. Vector. Los simúlidos están relacionados con la transmisión de enfermedades, son vectores
de patógenos. Unos de los patógenos más dañinos que transmite son las filarias,
entre estas la oncocercosis (Onchocerca volvulus) u otras como la Mansonella
ozzardi (Blacklock, 1926; Shelley & Coscarón, 2001).
La oncocercosis es una enfermedad que se da en África Central y del Sur, llegando
hasta Arabia y de igual modo en América Central y del Sur. Es causante de un gran
número de casos de ceguera en África. Existen regiones donde el número de
cegueras asciende a los 100.000 de una población total de 12 millones (Philippon,
1987).
Esto tiene una gran importancia socioeconómica, ya que una alta tasa de población
está desempleada debido a la enfermedad, lo que ello supone un incremento de la
pobreza.

18
También está relacionada con el ganado, ya que a éste le transmite patógenos como
filarias (p.e. Onchocerca linealis en mamíferos y Ornithofilaria fallisensis en aves)
(Grácio, 1985), además de poderles transmitir tripanosomas (Bennet, 1961) o
protozoos como Leucocytozoon (Cupp, 1987).Se puede hablar entonces de un
problema económico importante en aquellas regiones donde se encuentra en
abundancia.
Foto 3. Se puede ver una larva de simúlido con un parasito, en este caso una
especie de nematodo o nematomorfo

19
4. OBJETIVOS. Nuestro objetivo es llevar a cabo un estudio que nos permita establecer la dinámica
poblacional de las distintas especies de la familia Simuliidae, que habitan distintos
arroyos de la comarca Sierra de Segura sur (Jaén).
5. MATERIAL Y MÉTODOS. 5.1. Áreas de estudio. Este trabajo se desarrolla en tramos de los siguientes cauces fluviales: Arroyo
Orcera, Arroyo de los Molinos y Rio Trújala (véase mapa1 y 2). Los tres pertenecen
a la cuenca del Guadalquivir, localizados en el distrito sur de la Sierra de Segura.
En cada cauce fluvial se parametrizaron las siguientes variables: temperatura ºC;
conductividad mS/cm2, mS/cm; mΩcm; solubilidad TDS g/L, Sal; niveles de oxígeno
DO%, DO mg/L y pH con el sistema multisonda 556 MPS (YSI Incorporated).La
velocidad de corriente se tomó con el correntómetro FPIII (Global Water™).
Asimismo, hemos valorado la concentración de fosfato total y nitrógeno con los test
NANOCOLOR (Macherey-Nagel®) y el uso del Photometer PF-11 (Macherey-
Nagel®).En ambos casos hemos seguido los protocolos del fabricante (Anexo 1,
Tablas 2, 3 y 4).
5.1.1. Arroyo Orcera. Se trata de un cauce hídrico de aproximadamente unos 9,08 km de longitud desde
su nacimiento en superficie hasta que confluye con el río Trujala. Se encuentra en el
municipio de Orcera. Se trata de un rio típicamente de montaña. La zona de
muestreo se localiza a unos 2,5 km del nacimiento de éste. El transecto de muestreo
se extiende unos 100 metros de longitud a lo largo del cauce. El cauce es de una
anchura aproximada de 2,3 m. La velocidad media de la corriente es de 0,4m/s (0,6
m/s en superficie y 0,2 m/s en profundidad). Presentaba una temperatura media de
11,29 ºC y un pH medio de 9,87 (Anexo 1, Tabla 2)
La vegetación que le rodea es un Pinar de Pinus halepensis, perteneciente a la serie
edafoxerófila del Rhamno lycioidis-Pineto halepensis sigmetum. En la orla del cauce
podemos encontrar la geoserie riparia supramediterránea ibérica subhúmeda-
húmeda del Salix alba, la podemos encontrar en estadios muy degradados donde la

20
vegetación de mayor porte que encontramos en la zona de muestreo fueron los
zarzales de Rubo-Rosetum corymbiferaey los juncales de Lysimachio-
Holoschoenetum (Rivas Martínez, 2011).
Geológicamente hablando encontramos en la zona la siguiente tipología de suelos:
Cambisoles vérticos, vertisoles crómicos y cambisoles cálcicos con regosoles
calcáreos, esto en particular en la zona donde realizamos nuestro transecto. Cuando
el rio ha trascurrido unos 6km cambia ligeramente la estructura del suelo y tenemos
entonces lluvisoles cálcicos, cambisoles cálcicos, cambisoles dúricos con lluvisoles
crómicos y regosoles calcáreos con litosoles. Se caracteriza porque la mayor parte
del cauce del rio presenta un sustrato de roca calcárea (Mapa 3)
(http://www.juntadeandalucia.es/medioambiente/site/portalweb).
5.1.2. Arroyo de los Molinos. Cauce hídrico de aproximadamente unos 9,26 km desde su nacimiento hasta su
desemboca en el pantano del Tranco de Beas de Segura, recibe el aporte de
diferentes afluentes de menor tamaño y poca importancia como para nombrarlos.
Perteneciente al municipio de Hornos de Segura. Este río se encuentra en una zona
más explotada y agrícola. En sus cercanías podemos encontrar granjas bobinas y
vacunas, además la zona está altamente alterada y en su mayoría alejándonos del
cauce del río solemos encontrar nada más que olivares.
El lugar donde realizamos nuestro muestreo fue aproximadamente a unos 500
metros de su desembocadura, el transecto que realizamos abarcaba unos 100
metros de extensión a lo largo del cauce. El cauce fluvial tenía una anchura media
de 2,5 y una corriente media de 0,51 m/s (presentaba una corriente de 0,83 m/s de
media en superficie y 0,18 m/s de media en profundidad). Tenía una temperatura
media de 9,57ºC y presentaba un pH medio de 9,27 (Anexo 1, tabla 3).
La vegetación que albergaba la zona consistía en un olivar con pequeñas manchas
de pinos y encinas. Pero se puede deducir por las condiciones ambientales que la
serie de vegetación que se daría de modo natural es la Paeonio coriaceae-Querco
rotundifoliae sigmetum. Mientras que en la orla del río nos volvemos a encontrar la
geoserie edafohigrófila mesomediterránea iberolevantina y bética oriental seca
basófila del Ulmus minor, Aro italici-Ulmeto minoris sigmetum. En la zona donde
realizamos nuestra labor de recogida de muestras, la serie se encontraba en todos

21
sus estadios aunque quedaba reducida su extensión, limitándose a un metro del río
aproximadamente la colonización de esta (Rivas Martínez, 2011).
En materia de geología hablar que el cauce hídrico pasa principalmente por dos
agrupaciones distintas de suelos, la primera y es la que nos encontramos en la zona
de muestreo, abarca desde la desembocadura hasta aproximadamente 3,5 km agua
arriba, está conformado el suelo aquí por: Cambisoles dúricos, lluvisoles crómicos y
cambisoles cálcicos con regosolesdúricos y calcáreos y lluvisoles cálcicos. Por otro
lado en la zona alta del río podemos encontrar la agrupación formada por: Lluvisoles
cálcicos, cambisoles cálcicos, cambisoles dúricos con lluvisoles crómicos, regosoles
calcáreos y litosoles. Al menos en la parte en la que llevamos a cabo el muestreo el
río presentaba un sustrato rocoso de dimensiones intermedias y de tipo calcáreo
(Mapa 3) (http://www.juntadeandalucia.es/medioambiente/site/portalweb).
5.1.3 Río Trujala. Se trata de un cauce fluvial que desde su nacimiento hasta que confluye, con el río
Siles, para formar ambos el denominado río Guadalimar trascurren unos 12,27km.
Pasa por los municipios de Segura de la Sierra y Orcera. El tramo donde nosotros
realizamos la toma de muestras se encontraba en buen estado y sin importantes
alteraciones en la zona, existían pequeños parches de campos agrícolas muy
antiguos pero ya en estado naturalizado. Tiene como principal afluente el Arroyo
Orcera, el resto son arroyos temporales que se secan en verano su gran mayoría.
Realizamos nuestra labor de muestreo a 1 km desde el nacimiento del río, en este
caso nuestro tramo de trabajo fue de unos 150 metros a lo largo del río, este
presentaba una anchura de unos 3 m de media. Presenta una corriente media de
0,325 m/s (presenta una corriente media en profundidad de 0,15 m/s y en superficie
de 0,5 m/s). Tenía una temperatura media de 10, 98ºC y también un pH medio de
9,53 (Anexo 1, tabla 4).
La vegetación de la zona es muy compleja, ya que podemos encontrar a un lado un
pinar y en el opuesto tenemos mezcla de cultivos naturalizados, bosque de ribera y
pinares salteados. Podemos decir nos encontramos en una serie de Paeonio
coriaceae-Querco rotundifoliae sigmetum, por la presencia de algunos elementos
como quercus sp. pero de pequeño porte y por las condiciones que allí encontramos.
En la ribera del río encontramos claramente la geoserie riparia mesomediterránea
iberolevantina y bética oriental seco-subhúmeda basófila de Salix purpurea

22
subespecie lambertiana, Saliceton eotrichae sigmetum. Existían todas los estadios
de esta y estaban bien representados, abundando sobre todo la Salix purpurea
(Rivas Martínez, 2011).
Para terminar hablamos de la composición de su suelo, se distinguen tres
composiciones distintas, una primera en la que realizamos nuestra tarea que estaba
formada por: Cambisoles vérticos, vertisoles crómicos y cambisoles cálcicos con
regosoles calcáreos. En el tramo medio de este rio se da un cambio en la
composición y pasamos a tener: Lluvisoles cálcicos, cambisoles cálcicos,
cambisoles dúricos con lluvisoles crómicos, regosoles calcáreos y litosoles. Para
acabar, a la altura de la desembocadura del arroyo Orcera encontramos una nueva
composición formada por: fluvisoles calcáreos. En este rio en concreto el sustrato
del fondo que encontramos variaba a los anteriores, encontrábamos roca caliza de
buen tamaño, pero existía una capa fina de arenisca que prácticamente lo cubría
todo (Mapa 3) (http://www.juntadeandalucia.es/medioambiente/site/portalweb).
Mapa 1. Mapa de la Sierra de Cazorla, Segura y las Villas. www.juntadeandalucia.es

23
Mapa 2. Mapa marcado en azul los ríos en los que se trabajo. sigpac.mapa.es

24
Mapa 3. Mapa de composición de suelos. www.juntadeandalucia.es

25
Mapa 4. Mapa ampliado de la composición de los suelos de nuestra zona de estudio.
5.2 Método de captura. 5.2.1. Muestreo. En cada uno de los cauces, dentro del área prospectada, seleccionamos cuatro
puntos de muestreo distintos, procurando que exista una cierta distancia entre ellos.
En cada punto de muestreo colocamos una varilla de hierro en forma de T que actúa
como soporte de dos series de siete hojas de caña común (Arundo donax), que han
sido fijadas con hilo de nailon (Esquema 1)
El proceso de colocación de los trozos de caña se produce según el siguiente
esquema:
(i) En una primera fecha ponemos la primera caña a la que denominaremos
T0.
(ii) Pasa dos quince días sobre el mismo poste colocaríamos un nuevo trozo
de caña (T1). Procedemos a recolectar 3 hojas de la muestra T0.
(iii) Transcurrida otra quincena procedemos a retirarlo tras tres hojas a T0
(existe una hoja de más por si existieran posibles pérdidas). Cortamos la
sujeción de T0 al soporte siendo sustituida por un nuevo trozo de caña
esta vez denominado T2. En esta misma fecha retiramos 3 hojas de la
muestra al soporte T1.
(iv) Repetimos este proceso cada quinces días hasta llegar a T9.
Todo esto en cada punto de muestreo que como recordamos eran cuatro
por rio (Tabla 1).

26
2
8/0
3/2
01
4
12
/04
/20
14
26
/04
/20
14
10
/05
/20
14
28
/05
/20
14
7/0
6/2
01
4
22
/06
/20
14
6/0
7/2
01
4
21
/07
/20
14
5/0
8/2
01
4
20
/08
/20
14
4/0
9/2
01
4
T0 T0-15 T0-30 T1-30 T2-30 T3-30 T4-30 T5-30 T6-30 T7-30 T8-30 T9-30
T1 T1-15 T2-15 T3-15 T4-15 T5-15 T6-15 T7-15 T8-15 T9-15 T10-15
T2 T3 T4 T5 T6 T7 T8 T9 T10 T11
Dibujo 1. Esquema de como quedaría nuestro sistema de recolección de simúlidos.
5.2.2 Recogida de Muestras Se recogen tres hojas por cada quincena que pasa de cada trozo de caña. El
procedimiento que se sigue es el de cortar cada hoja por su peciolo, seguidamente
retirar y guardar en un frasco que contiene alcohol al 70º. Se marca el código
correspondiente al lote y fecha de recogida, y se almacena. Antes de guardar
removemos con fuerza para conseguir desprender la gran mayoría de insectos.
Una vez recogidas las muestras, se limpian. Hay que tener en cuenta que presentan
fango, restos de hojas y otros residuos que no nos interesan. Se extrae primero la
hoja y se recolectan los insectos que estén aun sujetos a ella, estos son colocados
en un eppendorf si la muestra es pequeña o varios si hiciera falta. Una vez sacada la
hoja, todo el alcohol que queda, en el que esta la gran mayoría de nuestros insectos,
Tabla 1. En color azul tenemos los trozos de caña de nueva colocación, en color naranja se
hace referencia a que tras quince días de su colocación se extraen las tres primeras hojas y
por ultimo en color rojo se hace referencia a que tras un mes se recogen las tres últimas
hojas y se corta dicho trozo de caña. En verde aquellas que finalmente excluimos.

27
se hace colar por una malla de menos de un milímetro de tamaño de poro. Se
eliminan los residuos y se recogen los insectos que se depositan en los eppendorf.
Para terminar este proceso una parte importante es la de la limpieza de hojas y su
posteriormente buena conservación. Fueron limpiadas en una batea con agua y
finalmente conservadas en seco, bien marcadas para más tarde identificar.
5.3. Tratamiento de las hojas. A la hora de trabajar con las hojas lo primero que hicimos fue rehidratarlas para que
tomaran su morfología inicial, para ello se introdujeron en bateas que contenían
agua, tras un periodo de unos 5 minutos las sacamos y secamos con papel de filtro.
A continuación el proceso es sencillo metemos las hojas en un escáner y las
escaneamos. Guardamos con su correcto marcaje. También haremos lo mismo con
un cuadrado de cartulina de 10X10 cm (100𝑐𝑚2) que utilizaremos como control.
Posteriormente pasamos los archivos escaneados por un programa informático el
"Gimp2". El primer archivo que empleamos es nuestro control, y mediante las
herramientas de este programa, pondremos en primer lugar el archivo en escala de
grises, recordar que el control es una cartulina de 10X10 cm negra, y finalizaremos
sacando el número de pixel que existen que no sean blancos. Dicho número será la
cantidad que existe en 100𝑐𝑚2. Continuamos haciendo lo mismo para los archivos
de las hojas escaneadas, se pasan a escala de grises y se contabilizan todos los
pixeles que no sean blancos (ya que los negros y grises son los que marcan la
silueta de la hoja).
Para terminar se pasa de numero de pixel a área en 𝑐𝑚2, teniendo en cuenta que
100 𝑐𝑚2 son"X" pixeles (en nuestro caso 410078), por lo que conseguimos deducir
el área de todas las hojas que se han extraído en las muestras. Tener en cuenta que
el área donde se pueden posar los simúlidos se duplican, porque lo pueden hacer
por las dos caras de las hojas, por lo que el área que hemos calculado habría que
multiplicarla por dos, así obtendríamos el área total que pueden ocupar los
simúlidos.

28
5.4. Determinación taxonómica. Una vez ya en el laboratorio el procedimiento es el siguiente: sacamos de cada
muestra (eppendorf) los insectos recolectados. Los separamos excluyendo todos los
que no pertenezcan a la familia Simuliidae.
Una vez que los tenemos separados los dividimos en dos grupos las pupas y las
larvas. Comenzamos determinando las pupas, ya que son un fuerte indicativo de las
especies de larvas que tuviéramos en las muestras. Para ellos usamos claves
dicotómicas (González, 1997; Crosskey, 2000; Belgat & Dakki, 2004).
Tenemos que tener en cuenta que las pupas, en su mayoría, son fácilmente
identificables, salvo que estén muy deformadas o hayamos cometido el error de
romperlas.
En las larvas tenemos que tener en cuenta dos aspectos para su determinación, el
primero es que no todas presentan el mismo tamaño porque pertenecen a estadios
distintos, en este caso excluimos a aquellos inferiores a un milímetro, debido a que
no son fáciles de manipular. En el siguiente aspecto se hace referencia al desarrollo
de la larva, ya que muchas larvas pese a que presenten estadios larvarios muy
avanzados, no se encuentran bien desarrolladas y aquellos criterios de morfología
por los que se determinan no están presentes, por lo que no es conveniente llevar a
cabo su determinación, ya que la probabilidad de errar en ello es muy alta (Foto 5).
Foto 4. Momento de determinación en el laboratorio.

29
Foto 5. Comparativa de larvas.
5.5. Índices de biodiversidad. Usaremos los índices de biodiversidad α de Margalef, Shannon y Pielou para
calcular la diversidad de especies en cada rio, tanto para larvas como para pupas
(Rodríguez et al., 2013).
- Índice de Margalef:
𝐷𝑀𝐺 =𝑆 − 1
𝑙𝑛𝑁
-Índice de Shannon
𝐻 = − ∑(𝑃𝑖 ∗ 𝑙𝑛𝑃𝑖)
-índice de Pielou
𝑃𝑖𝑒𝑙𝑜𝑢´𝐽 = 𝐻/𝑙𝑛(𝑆)
Usamos también un índice de diversidad β, el índice Sorenson (cualitativo), para
comparar los distintos cauces fluviales entre sí.

30
-Índice de Sorenson
𝑆𝑜𝑟𝑒𝑛𝑠𝑜𝑛 (𝑐𝑢𝑎𝑙. ) = 2𝐶/(𝑆𝐴 + 𝑆𝐵)
5.6. Material
- Alcohol 70º - Lupa olympus C011. Europe
- Photometer PF-11 MACHEREY-NAGEL
- Test NANOCOLOR MACHEREY-NAGEL
- Correntómetro FPIII Global Water™ - Microscopio Olympus CX31 Europe
- Placa Petri - Portaobjetos excavado
- Cubreobjetos - Multisonda 556 MPS YSI Incorporated
- Frascos para muestras - Eppendorf
- Pinzas antimagnéticas - Escáner
- Bateas - Malla de 1mm de poro
- Embudo - Papel de filtro

31
6. RESULTADOS.
Exponemos nuestros resultados expresados como número de individuos
encontrados por unidad de hábitat (200 cm2). Recordamos que en cada una de las
fechas registradas, cada cauce fluvial presentaba 4 puntos de muestreo con tres
réplicas, por lo que cada fecha representa la suma de 12 hojas de caña, con valores
de superficie corregidos. Nuestro protocolo de muestreo nos permite diferenciar
entre hojas que han pasado 15 días sumergidas en el cauce de aquellas que han
estado en contacto con el agua 30 días.
En el caso particular del río Trujala no obtuvimos ninguna captura, a pesar de haber
mantenido el mismo protocolo y esfuerzo de muestreo.
6.1. Resultados generales. Durante el desarrollo de este estudio hemos colectado un total de 2371
determinadas y 4026 no determinadas de larvas y 564 pupas pertenecientes a las
siguientes especies (solo se cuentan las determinadas): Simulium (Eusimulium)
velutinum, S. (E.) petricolum, S. (Nevermania) cryophilum, S. (Tetisimulium) bezzi, S.
(Wilhemia) pseudoequinum, S. (Simulium) xanthinum, S. (S.) argireatum, S. (S.)
ornatum y S. (S.) intermedium.
La distribución de estas capturas, teniendo cuenta que hemos excluido aquellos
ejemplares de < 1mm de longitud, ha resultado en la determinación de de 1200
larvas y 337 pupas pertenecientes a seis especies de simúlidos en el arroyo Molinos
y de de 1171 larvas y 227 pupas pertenecientes a seis especies de simúlidos en el
arroyo Orcera.
Con los resultados obtenidos, se hace la media entre el número de individuos y el
global de las doce hojas del mismo periodo y tiempo de permanencia en agua
(cuatro puntos de muestreo y tres replicas en cada uno), ha dichos valores también
se les hace la desviación estándar para controlar el error que pueda existir. (Anexo
1. Tabla 5, 6, 7 y 8)
A partir de aquí, mostramos las gráficas resultantes del cálculo de la media de las 12
hojas recogidas, para formar un lote a los 15 y 30 días respectivamente, en base a
los valores de superficie corregidos de las hojas muestreadas.

32
Las gráficas 1 y 2 hacen referencia a los resultados obtenidos para larvas en el
arroyo Molinos. Mostramos dos gráficas separadas para realzar los valores de las
especies que aparecen menos representadas. Como puede observarse hemos
capturado 6-7 taxones. Recordamos que las larvas de S. (S.) ornatum y S. (S.)
intermedium son indiferenciables a nivel de larvas. La gráfica 1 pone de manifiesto
que tres especies están poco representadas, mientras que en la gráfica 2 los
ejemplares de ornatum/intermedium son mayoritarios durante todo el periodo,
mientras que S. (N.) cryophilum y S. (E.) velutinum aparecen en menor número.
También en la gráfica 3 se ve la cantidad de pupas de aquellas especies que son
poco abundantes y en la gráfica 4 podemos ver la cantidad de pupas de aquella
especie que ha presentado una mayor abundancia y un ciclo marcado.
Gráfica 1. A. Molinos. Se pueden ver las larvas que se han encontrado de especies poco presentes o puntuales.
Gráfica 2. A. Molinos. Se pueden ver la dinámica larvaria que siguen las especies más representativas.

33
Gráfica 3. A. Molinos. Representación de las pupas menos numerosas.
Gráfica 4. A. Molinos. Se puede ver la especie más presente y como varía según el tiempo que la hoja pertenezca en el agua.
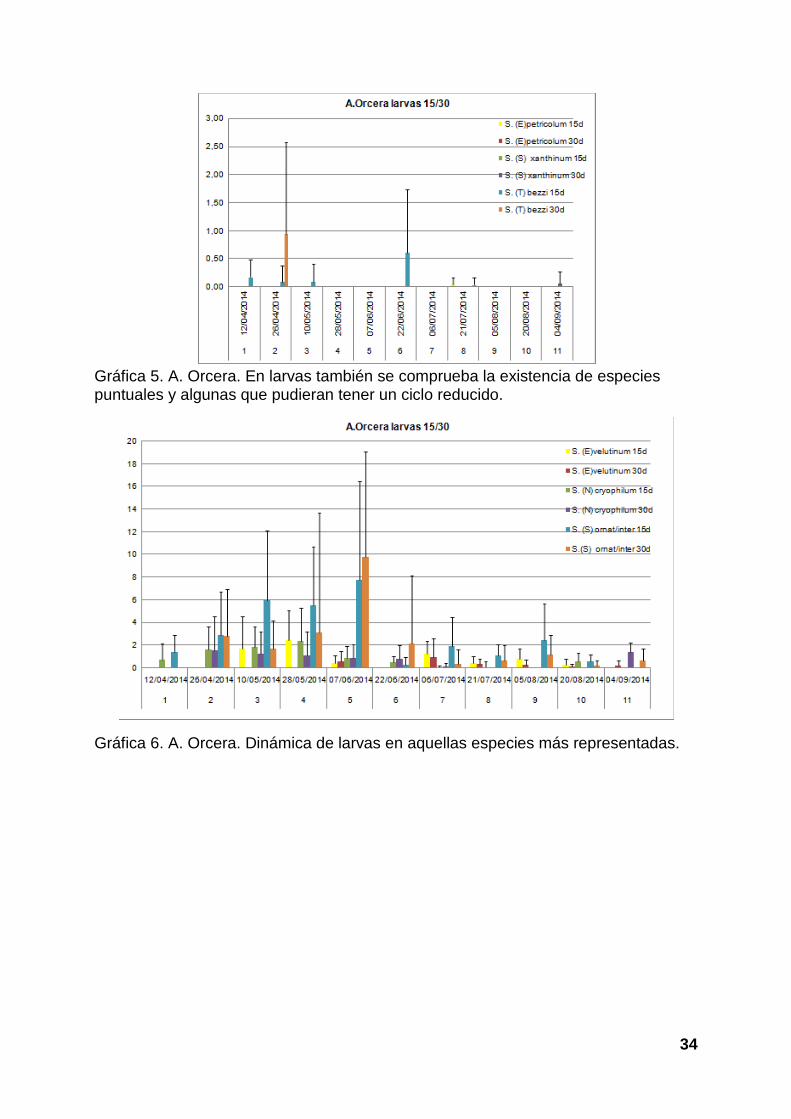
Las gráficas 5 y 6 muestran los resultados obtenidos de larvas en el arroyo Orcera.
Se muestran dos gráficas separadas para facilitar ver las cantidades de especies
que aparecen menos representadas. Como se observa hemos capturado 6-7
taxones. Recordamos que las larvas de S. (S.) ornatum y S. (S.) intermedium son
indiferenciables a nivel de larvas. La gráfica 5 marca que tres especies están poco
representadas, por su parte la gráfica 6 los ejemplares se ve que
ornatum/intermediumson mayoritarios y S. (N.) cryophilum y S. (E.) velutinum
presentan menores cantidades.
En la gráfica 7 se ve la cantidad de pupas de aquellas especies que son poco
abundantes, mientras que y en la gráfica 8 podemos ver la cantidad de pupas de
aquella especie que ha presentado una mayor abundancia y un ciclo marcado
34
Gráfica 5. A. Orcera. En larvas también se comprueba la existencia de especies puntuales y algunas que pudieran tener un ciclo reducido.
Gráfica 6. A. Orcera. Dinámica de larvas en aquellas especies más representadas.

35
Gráfica 7. A. Orcera. Se puede ver la existencia de algunas especies puntuales.
Gráfica 8. A. Orcera. Vemos la dinámica pupar de las especies más representadas.
6.2. Resultados comparativos.
En este caso se comparan datos de aquellas hojas que han estado 30 días en el
agua con la suma de aquellas dos hojas que han estado 15 días (las del mismo

36
periodo de recolecta y las predecesoras).Con el fin de poder comparar la influencia
de estar quince días o treinta días el sustrato colocado.
Volvemos a tener en cuenta los datos de exclusión de individuos (aquellos de< 1mm
de longitud, los datos vienen nueva mente dados en valores de medias como en el
punto anterior, y las desviación de igual modo. (Anexo 1 tablas 9, 10, 11 y 12)
En este apartado solo se procede a exponer las graficas comparativas de aquellas
especies más abundantes, puesto que las que tienen una menor presencia, dan
unos resultados prácticamente idénticos a si no sumásemos los dos periodos de 15
días.
Las gráficas 9 y 10 hacen referencia a los resultados obtenidos para larvas en el
arroyo Molinos. En la gráfica 9 podemos observar un esbozo de un ciclo claro de las
especies y en algunos casos vemos que se igualan los periodos. La gráfica 10 por
su parte demuestra que se alterna la cantidad de individuos capturados en pupas ya
sean con un periodo de 15 días o el de 30 días. Además vemos que la única especie
que muestra una cierta abundancia de pupas es la de S. (S.) ornatum.
En la gráfica 11 se ve un esbozo del ciclo de vida de las especies más abundantes
de larvas en el río Orcera, y en la gráfica 12 queda mucho más patente.
Gráfica 9. A. Molinos. Comparación entre distintos periodos en agua de las especies más numerosas.

37
Gráfica 10. A. Molinos. Se pueden comparar las pupas en ambos periodos.
Gráfica 11. A. Orcera. Comparación entre distintos periodos en agua de las especies más numerosas.
Gráfica 12. A. Orcera. Comparativa de pupas, existen periodos en los que se ve una igualdad.

38
6.3. Resultados por periodo de influencia.
Consiste en la suma de aquellas hojas que se recogen en el mismo periodo y las
que preceden a las de treinta días de la misma caña (del mismo trozo de caña)
(15+15+30d). Con el fin de tener un buen resumen de una dinámica poblacional de
cada especie.
Este apartado nos dará una clara visión de cuál es la dinámica poblacional que sigue
cada especie en cada río, tanto a nivel de larvas como de pupas, todos los datos se
han expuesto en valores de medias para darles mayor fiabilidad y que sean
comparable las especies entre sí.
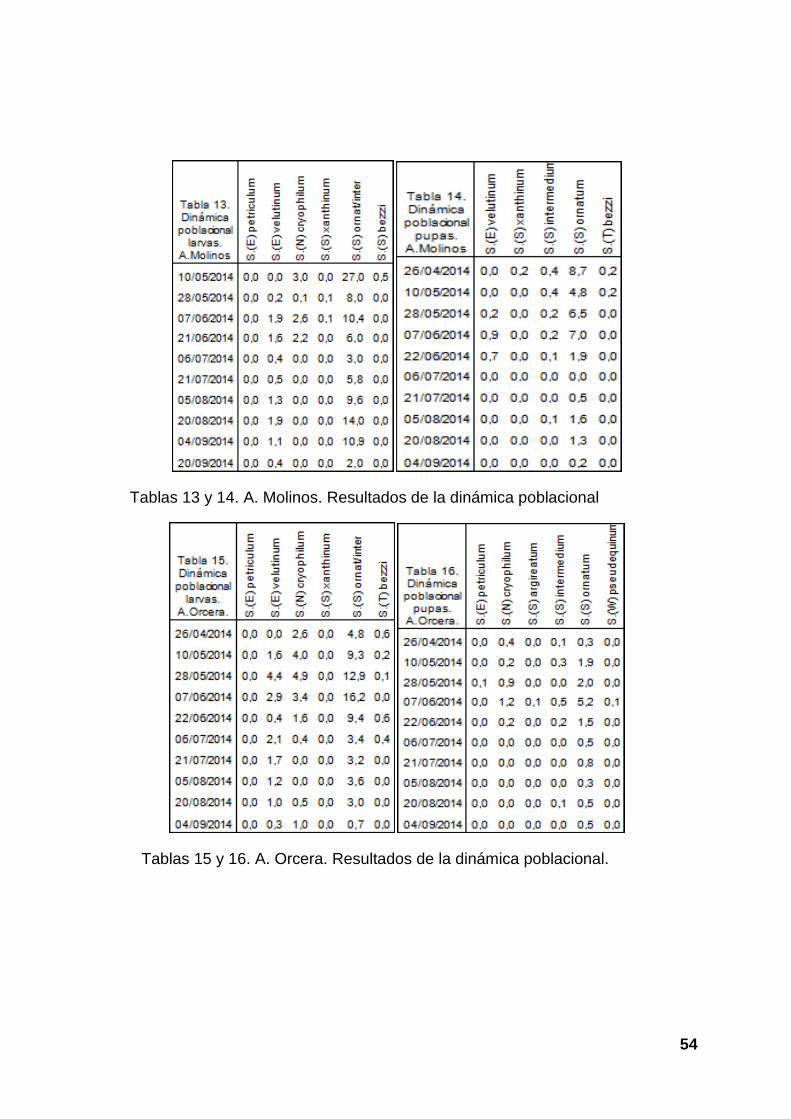
Las gráficas 13 y 14 hacen referencia al arroyo Molinos. En la gráfica 13 vemos las
especies de larvas que hemos encontrado en dicho río y la dinámica que siguen
cada una, en la gráfica 14 podemos ver dicho ciclo en pupas, pero además se ve
cuales son los periodos más favorables para cada especie.
En las gráficas 15 y 16 se reflejan los mismos parámetros pero para arroyo Orcera.
En la gráfica 15 podemos ver la dinámica de las larvas de las diferentes especies
presentes en dicho río, así como comparar cuales son más abundantes y en que
periodo se da una mayor abundancia. En la gráfica 16 podemos ver los mismos
datos pero en pupas y además podemos interpretar cuales son las especies que
presentan una mayor eficacia en el paso de larva a pupa, aunque no es un valor
100% fiable porque actúan otros factores.
Gráfica 13. A. Molinos. Dinámica poblacional larvas.

39
Gráfica 14. A. Molinos. Dinámica poblacional pupas.
Gráfica 15. A Orcera. Dinámica poblacional larvas.
Gráfica 16. A. Orcera. Dinámica poblacional pupas.
40
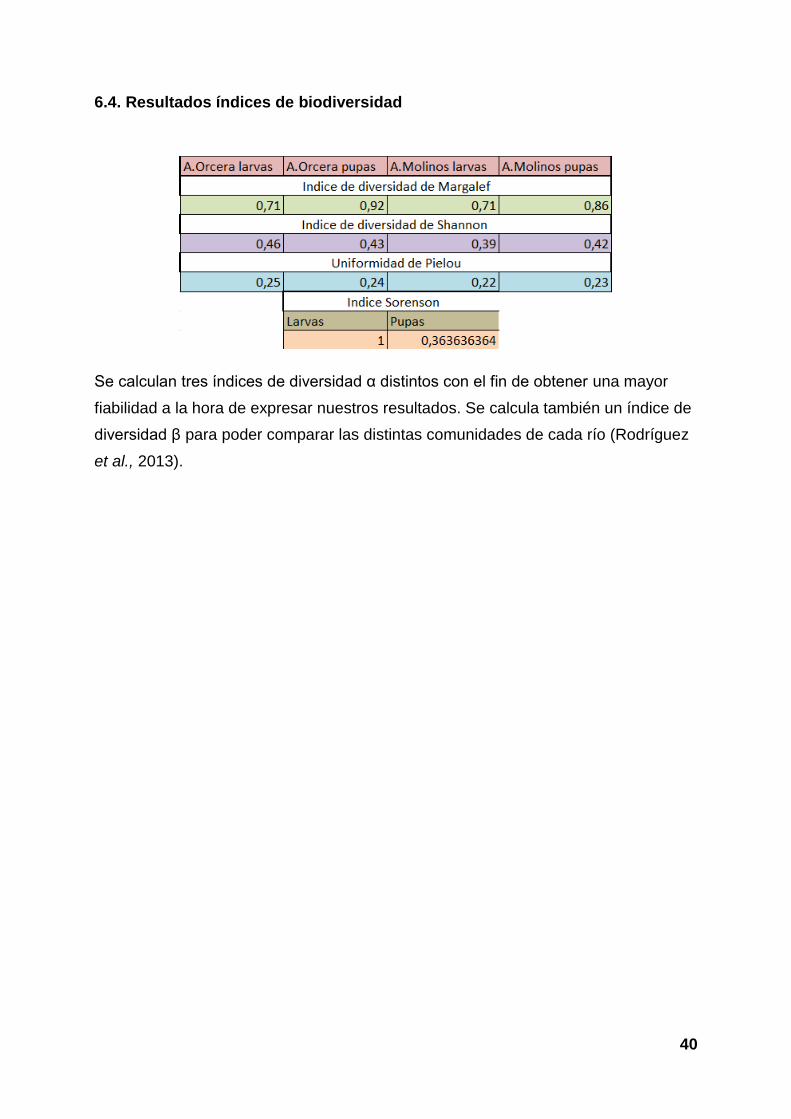
6.4. Resultados índices de biodiversidad
Se calculan tres índices de diversidad α distintos con el fin de obtener una mayor
fiabilidad a la hora de expresar nuestros resultados. Se calcula también un índice de
diversidad β para poder comparar las distintas comunidades de cada río (Rodríguez
et al., 2013).

41
7. DISCUSIÓN Durante el desarrollo de este estudio se ha probado con dos materiales como
soporte: i) el primero consistía en unas láminas de plástico de polietileno rugoso de
30X5 cm, desechamos este procedimiento al ver que se depositaban pocos
individuos en él. De todos modos se dejaron algunos como control, con el fin de ver
si existía alguna especie que se incorporara a dicho soporte y no al que finalmente
usamos; ii) Se trata de hojas de Arundo donax, que dieron mejor resultado,
entendemos por tratase de un soporte natural.
En arroyo Molinos vemos que las especies menos abundantes son S. (E.)
petricolum, S. (S.) xanthinum y S. (T.) bezzi, tanto en número de larvas como de
pupas son poco frecuentes y salteadas en el tiempo. Esto puede deberse a que no
se encuentren en sus condiciones optimas, o que nuestro método de captura no
haya sido el ideal para dichas especies.
La especie S. (E.) velutinum es algo más abundante pero no se consigue tener una
buena abundancia de estos como para determinar su dinámica, principalmente en
pupas. La falta de estas nos complica que podamos afirmar que es una especie que
presenta sus picos de abundancia, en su ciclo, en el periodo final de primavera y al
principio del otoño. Aun así las larvas nos marcan ese modelo.
En este cauce fluvial vemos que la especie S. (N.) cryophilum tiene una aparición
puntual de una buena cantidad de larvas, en alguna fecha muy concreta, y siempre
en los primeros meses de primavera. A la hora de ver su eficiencia del paso de
larvas a pupas podemos comprobar que es muy baja, puesto que no hemos tenido
ninguna pupa, o que por el contrario las larvas pasaban a pupas en otros lugares
donde las condiciones le eran más favorables.
Se ve que el combinado S. (S.) ornatum y S. (S.) intermedium es el que presenta un
mayor número de individuos tanto de larvas como de pupas. Podemos decir que
aunque la especie no esté en su optimo si esta en valores cercanos a él. También
decir que la mayoría de individuos de este bloque pertenecen a la especie S. (S.)
ornatum ya que en pupas (donde podemos diferenciarlos), el número de individuos
es mucho mayor que el de la otra especie.
Esto coincide con lo expuesto en el artículo de (Philipson, 1956) donde ve que las
mayores concentraciones de dicha especie se dan a una velocidad de corriente de
entre 0,8 y 0,9 m/s, a lo que nuestros valores se acercan en determinados puntos en
este cauce fluvial.

42
Por otra parte tenemos el arroyo Orcera, en el cual vemos que existen ocho
especies distintas. Cada una presenta una abundancia
Se ha visto que la especie S. (E.) petricolum ha sido una especie que se ha dado de
manera puntual en muy pocas fechas y en muy baja cantidad, tanto para larvas
como para pupas, esto puede deberse a que la especie no se encuentre en su
optimo o que sus periodos más favorables queden fuera de nuestro periodo de
muestreo. También puede ser causa de que el método de captura no haya sido el
más idóneo.
Tenemos además un bloque de especies formado por: i)en primer lugar las especies
S. (S.) xanthinum, S. (T.) bezzi y S. (E.) velutinum, en las tres obtuvimos capturas
solo de larvas (aunque con una mayor abundancia de captura de esta última), esto
puede deberse a que las larvas buscaran otros lugares más apropiados para realizar
la metamorfosis a pupa.ii) En segundo lugar tenemos las especies S. (S.) argireatum
y S. (W.) pseudoquinum en las que solo se encontraron ejemplares de pupas,
principalmente, esto puede estar causado porque exista un bajo número de
individuos de las especies en la zona y solo se aprecia su existencia en formas fijas.
A esto debemos añadir la existencia de un valor de larvas que no se pudieron
determinar, haciendo pensar que es posible que las de dicha especie entren en ese
conjunto.
Para acabar con este cauce, tenemos algunas especies que nos han dado una clara
línea de captura y que presenta una buena abundancia como para ver cuál es su
dinámica. Se trata de S. (N.) cryophilum y del conjunto S. (S.) intermedium y S. (S.)
ornatum, podemos ver que estas dos últimas están presentes durante todo el año,
dando se su pico de abundancia a finales de verano, y que S. cryophilum es una
especie que abunda en los periodos más fríos como son primavera y otoño,
quedando desapercibida durante los periodos de mayor calor, en verano.
En lo que corresponde a la comparativa de 15+15 días con aquellas hojas que solo
estuvieron 30 días, decir que no existe una relación directa. Por lo que se puede ver
podemos decir que las larvas se ven favorecidas por el periodo en el que las hojas
solo llevan 15 días en el agua. Esto puede deberse a:
- i) La existencia de una fina película de microorganismos en aquellas
hojas que llevaban un periodo de 30 días en el agua. Esto nos hace
pensar que esta circunstancia tenga como consecuencia una disminución
de la capacidad de sujeción al sustrato y probablemente una reducción en
la capacidad de filtración y oxigenación de las larvas.

43
- ii) Que las hojas en un primer momento actúen como un sumidero de
especies, al que las larvas irán para la colonización de nuevos espacios, y
que tras pasado un periodo ocurra el efecto inverso, las hojas actúen
como fuente de larvas hacia otros nuevos espacios que colonizar.
En lo que concierne a poblaciones estables podemos ver que en el arroyo Molinos
las especies S. (E.) velutinum y S. (N.) cryophilum presentan una dinámica
poblacional, la primera de ellas se encaja entre los meses de Mayo y Septiembre,
teniendo un declive de las poblaciones en las épocas más secas, la segunda estuvo
presente en los primeros meses de primavera pero no se consiguió ver un reflejo de
ello en las pupas, esto puedo deberse a que no esté en sus condiciones apropiadas,
su fiabilidad del paso de larva a pupa descienda o que busquen lugares más
idóneos.
Por otro lado tenemos el conjunto formado por las especies S. (S.) intermedium y S.
(S.) ornatum, sabemos que la gran mayoría de individuos de este complejo de
especies pertenecen a la ultima especie mencionado, ya que a nivel de pupas esta
es muchísimo más abúndate, pero como ya se mencionó al no poder distinguir en
larvas no haremos distinción. Esto nos lleva a decir que la gran abundancia de
dichas especies en este río es porque en él se dan condiciones cercanas a su
óptimo (Philipson, 1956).
Por otro lugar en el arroyo Orcera encontramos una situación similar. En la que las
especies que consiguen presentar una dinámica poblacional son S. (E.) velutinum y
S. (N.) cryophilum, en esta ocasión pasa totalmente lo contrario a lo que pasaba en
el anterior cauce fluvial. Pero además Simulium cryophilum presentaba una mayor
abundancia, se podía ver que su ciclo está relegado a primavera y otoño, su ciclo
vital está ligado a temperaturas más suaves, aunque su pico de abundancia se
registro en el mes de mayo, esto probablemente se deba al factor de temperatura
requerida para la eclosión de los huevos (ºC/día). Pero poco después su presencia
escasea hasta el regreso de temperaturas más frescas.
El conjunto de las especie S. (S.) intermedium y S. (S.) ornatum ocurre
prácticamente una situación parecida a la que se comenta en el arroyo anterior.
A nivel de hábitat podemos exponer que actúan como ecosistemas muy distintos. En
arroyo Molinos, dentro de las especies que nos marcan una clara dinámica, vemos
que existen dos picos de abundancia de individuos, tanto para larvas como para
pupas, pudiéndose intuir un tercero al principio de nuestro periodo. Esto nos indica

44
que existen dos ciclos en las especies, pudiendo haber incluso tres. Sin embargo en
el arroyo Orcera, como se puede ver solo se ve un pico de abundancia, ya que
aunque la línea de individuos que se capturan es continua no se deja ver
posteriormente a este ningún otro pico. Por lo que deducimos que habría un único
ciclo, los individuos que encontramos posteriormente puede estar originados por un
retraso en su eclosión o porque se anticipen. Todo esto nos lleva a pensar que las
condiciones de arroyo Molinos son mejores que el resto, provocando un aumento en
la cantidad de simúlidos (Kazanci & Ertunç, 2010).
Como conocemos algunos de los valores de sus parámetros podemos deducir que
los factores que más afectan al caso anterior serían la velocidad de corriente y la
temperatura. La velocidad de corriente, aunque varia no parece que sea la
excluyente de dichos ciclos. Nos centramos en la temperatura, que si varía entre
ellos en dos grados el uno del otro. Podemos decir que estas especies prefieren las
temperaturas más suaves, unas mejores condiciones causan un aumento del
número de ciclos.
Del mismo modo podemos ver que el pico que se da en el único ciclo en arroyo
Orcera, es mucho más pronunciado que el del resto, pudiéndose deber al factor de
temperatura requerida para la eclosión de los huevos (ºC/día). La especie siempre
tiene un valor constante para dicho factor, como hemos visto arroyo Orcera es más
cálido por lo que disminuye los días que transcurren para dicha eclosión.
También de los resultados podemos deducir la ausencia de individuos de la familia
Simuliidae en el río Trujala. Como se ve el arroyo Molinos se encuentra cercano al
optimo de velocidad de corriente para la especie S. (S.) ornatum, por lo que en él
aparece la mayor abundancia de capturas, al pasar al arroyo Orcera vemos que la
velocidad de corriente disminuye y de la misma manera también lo hace el número
de individuos de dicha especie, aunque esto se puede ver influenciado por
temperaturas más bajas en el último. En el río Trujala desciende aún más la
velocidad de corriente, por lo que esto puede suponer que sea un factor limitante no
solo para dicha especie sino para todas las demás.
Se cree que además la ausencia de individuos de la familia Simuliidae, también
puede estar relacionado con que elegimos un tramo de cabecera, donde todavía no
había un proceso de eutrofización de este, y a la presencia de una arenilla
carbonatada que cubría el lecho, pudiendo ser la causante de la ausencia de estos.
Pero aun así no sabemos cuál es el factor limitante, ya que no todas las especies se
ven influenciadas o por el sustrato, o por las temperaturas, debe de existir un

45
parámetro en concreto que nos impida la presencia de dicha familia (Kazanci &
Ertunç, 2010; Bernotiene, 2006; Kazanci, 2006).
Los índices de diversidad α toman valores bajos. Creemos que esto se debe a que
existen pocas especies y que prácticamente entre ellas solo una o dos son
abundantes.
Con respecto al índice de diversidad β, encontramos que a nivel de larvas la similitud
entre los ríos es de 100%. Esto puede deberse a que las especies que los distinguen
o bien no hemos elegido un buen método para capturarlas, o que está dentro de ese
paquete de larvas que no incluimos en el estudio. Los valores obtenidos para estos
índices en el caso de las pupas muestran diferencias que permiten discriminar entre
los arroyos estudiados, poniendo de manifiesto la distinta estructura de la
composición de simúlidos implicados en uno y otro caso.

46
8. CONCLUSIÓN. Se demuestra la existencia de ocho especies distintas en la región biogeografía de
la Sierra de Segura sur que son: Simulium (Eusimulium) velutinum, Simulium (E.)
petricolum,Simulium (Nevermania) cryophilum,Simulium (Tetisimulium) bezzi,
Simulium (Wilhemia) pseudoequinum, Simulium (S.) xanthinum, Simulium (S.)
argireatum, Simulium (S.) ornatum y Simulium (S.) intermedium.
En la mayor parte de los casos hemos podido realizar un seguimiento de la dinámica
poblacional de la especie, con la excepción de aquellas para las que se han
obtenido capturas puntuales [S. (E.) velutinum, S. (N.) cryophilum, S. (S.) ornatum y
S. (S.) intermedium].
Simulium (N.) cryophilum es una especie que presenta una mayor apetencia por
temperaturas más bajas, encontrándose exclusivamente durante primavera y otoño,
interrumpiendo su ciclo en verano.
Simulium (S.) ornatum [y el menos frecuente S. (S.) intermedium] adapta(n) su ciclo
a periodos del año con temperaturas más cálidas, de forma que larvas y pupas
aparecen entre marzo y septiembre.
También vemos que los mejores periodos son primavera y otoño, cuando se
produce menos variabilidad en las condiciones de los cauces fluviales. Los simúlidos
tienen adaptada su dinámica a las condiciones que se dan en los ríos.
La temperatura ha ejercido un efecto limitante en el número de ciclos que se dan en
las especies. Provoca también un aumento del número de eclosiones en un mismo
periodo.

47
9. BIBLIOGRAFÍA.
Beaucournu-Saguez, F., Chevrier, S., Dubois, B., Genete, M., De Lemos, J. B.,
Mutel, P., & Beaucournu, J. C.,1990. Etude preliminaire d'un foyer de
simulidose a Simulium posticatum Meigen, 1838 dans le departement de l'Eure.
Impact medical. Médecine et Maladies Infectieuses, 20(6), 279-283.
Belqat, B., & Dakki, M., 2004. Clés Analytiques des Simulies (Diptera) du
Maroc. Zoologica baetica, (15), 77-138.
Bennet, G.F., 1961. On the specificity and transmision of some avian
trypanosomes. Canadian Journal Zoology 39: 17-33.
Bernotiene, R. A. S. A., 2006. On the distribution of black fly larvae in small
lowland rivers in Lithuania. Acta entomologica serbica, 11, 115-125.
Blacklock, D.B.1926. The developement of Onchocerca volvulus in Simulium
damnosun. Annals of Tropical Medicine and Parasitology, 20: 1-48
Colbo, M.H. & Wotton, R.S., 1981. Preimaginal blackfly bionomics. In: M. Laird
(ed.), Blackflies. The future for biological methods in integrated control.
Academic Press. London, 209-226.
Craig, D. A. & Chance, M. M., 1982. Filter feeding in larvae of Simuliidae (Diptera:
Culicomorpha): aspects of functional morphology and hydrodynamics.
Canadian Journal of Zoology, 60(4), 712-724.
Crosskey, R. W. & Crosskey, M. E., 2000. An investigation of the blackfly fauna of
Andalusia, southern Spain (Diptera: Simuliidae). Journal of Natural
History, 34(6), 895-951.
Crossekey, R.W., 1973. Simuliidae. In: W.G.V. Smith (ed.), Insects and other
Artropods of Medical Importance. London, British Museum (Natural History):
109-153.
Cupp, E.W., 1981. Blackfly physiology. In: M. Laird (ed.), Blackflies: The future for
biological methods in integrated control. Academic Press. London: 199-208.
Cupp, E.W., 1987. The epizootiology of livestock and poultry diseases associated
with black-flies. In: K. Ch. Kin and R.W. Merritt (eds.), Black flies. Ecology,
Population Management and Annotated World List. Pennsylvania State
University: 387-395.
Cupp, E. W. & Cupp, M. S., 1997. Black fly (Diptera: Simuliidae) salivary
secretions importance in vector competence and disease. Journal of medical
entomology, 34(2), 87-94.

48
Currie, D.C. & Craig, D.A., 1987. Feeding strategies of larval black-flies. In:
K.Ch.Kin and R.W. Merritt (eds.), Black flies. Ecology, Population Management
and Annotated World List. Pennsylvania State University: 155-170.
Golini, V.I. & Davies, D.M., 1987. Oviposition of Black Flies. In: K.Ch.Kim and
R.W. Merrit (eds.), Black Flies. Ecology, Population Management and
Annotated World List. Pennsylvania State University. 261-275.
González, G., 1997. Claves para la identificación de las larvas y pupas de los
simúlidos (Diptera) de la Península Ibérica. Asociación Española de
Limnología.
Gordon, R., Ebsary, B. A., & Bennett, G. F., 1973. Potentialities of mermithid
nematodes for the biocontrol of blackflies (Diptera: Simuliidae)—A review.
Experimental parasitology, 33(2), 226-238.
Gracio, A.J. 1985. Estudo sistemático e bioecológico dos simulídeos de Portugal
(Diptera: Simuliidae). Tesis doctoral. Universidad de Lisboa. 796 págs.
Gradinarov, D. (2014). The mermithid species Isomermis lairdi (Nematoda,
Mermithidae), previously only known in Africa, found in Europe. ZooKeys, (454),
1.
Kazanci, N., & Ertunç, Ö., 2010. Use of Simuliidae (Insecta, Diptera) species as
indicators of aquatic habitat quality of Yeşilırmak River Basin (Turkey).Review
of hydrobiology, 3, 27-36.
Kazanci, N., 2006. Ordination of Simuliidae and climate change impact. Acta
entomologica serbica Suplement, 69-76.
Lestón, V., Díaz, J., Quirce, C. & Cobo, F., 2013. Los Simuliidae (Diptera) de la
cuenca del río Serpis (SE, España). Estudio faunístico. Boletín de la Asociación
Española de Entomología, 37(3), 285-300.
McIver, S.B. & Sutcliffe, J.F., 1987. Sensory basis of behavior and strutural
adaptations for feeding in black flies. In: K.Ch. Wim and R.W. Merritt (eds.),
Black flies. Ecology, Population Management and Annotated World List.
Pennsylvania State University, 228-249.
Moor, F. D., 1982. Determination of the number of instars and size variation in the
larvae and pupae of Simulium chutteri Lewis 1965 (Diptera: Simuliidae) and
some possible bionomical implications. Canadian Journal of Zoology, 60(6),
1374-1382.

49
Noirtin, C. & Boiteux, P., 1979. Mort de 25 animaux de ferme (dont 24bovins) par
piqures de simuliesdans les Vosges. Bulletin de la société Vétérinarie Practique
de France, 63: 41-54.
Philippon, B., 1987. Problems in Epidemiology and Control of West Afrrican
Onchocerciasis. In: K.Ch.Kin and R.W.Merrit (eds.), Black flies. Ecology,
Population Management and Annotated World List. Pennsylvania State
University, 363-373.
Phillipson, J., 1956. A study of factors determining the distribution of the larvae of
the blackfly, Simulium ornatum Mg. Bulletin of Entomological Research, 47(02),
227-238.
Post,R.J., 1983. The annual cycle of Simulium erythrocepalum (Diptera-
Simuliidae) at a site in Norfolk. Freshwater Biology, 13: 379-388.
Rivas-Martínez, S., 2011. Mapa de series, geoseries y geopermaseries de
vegetación de España. ItineraGeobotanica 18 (2): 425-800.
Rodríguez, J., Blanco, J. & Rodríguez, V., 2013. Ecología (3ª edición). Pirámide:
Madrid.
Ross,D.H. & Craig, D.A., 1980. Mechanisms of fine particle capture by larval
blackflies (Diptera, Simuliidae). Canadian Journal Zoology, 58: 1186-1192.
Rubtsov, I. A., 1956. Blackflies (Fam. Simuliidae). Fauna of the USSR. (Second
Edition) 6 (6), 860 pp.
Shelley, A. J. & Coscarón, S., 2001. Simuliid blackflies (Diptera: Simuliidae) and
ceratopogonid midges (Diptera: Ceratopogonidae) as vectors of
Mansonellaozzardi (Nematoda: Onchocercidae) in northern
Argentina. Memórias do Instituto Oswaldo Cruz, 96(4), 451-458.
Smart, J., 1940. Simuliidae (Dipt.) from British Guiana and the Lesser
Antilles.Transactions of the Royal Entomological Society of London, 90(1), 1-11.
Wenk, P., 1987. Swarming and mating behavior of black flies. In: K.Ch.Kin and
R.W.Merrit (eds.), Black flies. Ecology, Population Management and Annotated
World List. Pennsylvania State University, 215-227.
Zwick H. & Zwick,P., 1990. Terrestrial mass-oviposition of Prosimulium species
(Diptera-Simuliidae). Aquatic Insects, 12(1).
9.1. Bibliografía con enlaces.

50
MACHEREY-NAGEL Gmbh & Co.KG. NANOCOLOR orto fosfato y fosfato
total 1. ftp://ftp.mn-
net.com/espanol/hoja_de_instrucciones/NANOCOLOR/985076es.pdf
MACHEREY-NAGEL Gmbh & Co.KG. NANOCOLOR Nitrógeno total TNb
22. ftp://ftp.mn-
net.com/espanol/hoja_de_instrucciones/NANOCOLOR/985083es.pdf
MACHEREY-NAGEL Gmbh & Co.KG. Manuals wateranalysis.ftp://ftp.mn-
net.com/english/User%20manuals/Wateranalysis/PF-
11%20User%20Manual%20EN/PF-
11%20test%20pages_NANOCOLOR%20EN.pdf
Mapa de Suelos. Junta de Andalucía. Consejería de Medio Ambiente y
Consejería de Obras Públicas. [http://www.idee.es/web/guest/directorio-de-
servicios?p_p_id=DIRSRVIDEE_WAR_DIRSRVIDEEportlet_INSTANCE_q4BW&p_p_lifecycle=1&p_p_state=normal&p_p
_mode=view&p_p_col_id=column-
1&p_p_col_count=1&_DIRSRVIDEE_WAR_DIRSRVIDEEportlet_INSTANCE_q4BW_tipoServicio=WMS&_DIRSRVIDEE_
WAR_DIRSRVIDEEportlet_INSTANCE_q4BW_supertipo=OGC&_DIRSRVIDEE_WAR_DIRSRVIDEEportlet_INSTANCE_
q4BW_descSrv=VISUALIZACION&_DIRSRVIDEE_WAR_DIRSRVIDEEportlet_INSTANCE_q4BW_tipoAmbito=AUTONO
MICO]

51
Anexo 1
Tabla 1. Índices de diversidad obtenidos.
Tabla 2. A. Molinos. Parámetros fisicoquímicos obtenidos.
Tabla 3. A. Orcera. Parámetros fisicoquímicos obtenidos.
Tabla 4. R. Trujala. Parámetros fisicoquímicos obtenidos.

52
Tablas 5y 6. A.Molinos. Resultados generales.
Tablas 7 y 8. A Orcera. Resultados generales.

53
Tablas 9 y 10. A. Molinos. Resultados comparativos.
Tablas 11 y 12. A. Orcera. Resultados comparativos.
54
Tablas 13 y 14. A. Molinos. Resultados de la dinámica poblacional
Tablas 15 y 16. A. Orcera. Resultados de la dinámica poblacional.