estudio de la dinÁmica poblacional de … · ... de bacterias aceticas en la producciÓn de...
TRANSCRIPT

UNIVERSITAT ROVIRA I VIRGILI
DEPARTAMENT DE BIOQUIMICA I BIOTECNOLOGIA
FACULTAT D’ENOLOGIA
ESTUDIO DE LA DINÁMICA POBLACIONAL DE
BACTERIAS ACÉTICAS EN LA PRODUCCIÓN DE
VINAGRE POR EL MÉTODO TRADICIONAL
Trabajo presentado por:
CARLOS ALFREDO VEGAS PÉREZ
para la defensa del Diploma de Estudios Avanzados (DEA)

2
ESTUDIO DE LA DINÁMICA POBLACIONAL DE BACTERIAS ACETICAS EN
LA PRODUCCIÓN DE VINAGRE POR EL METODO TRADICIONAL
Carlos Vegas, Estibaliz Mateo, Carla Jara, Ángel González, José Manuel Guillamón,
Montse Poblet, Albert Mas y Mª Jesús Torija
UE-CeRTA. Dept. Bioquímica i Biotecnologia, Facultat d‘Enologia. Universitat Rovira i
Virgili. C/ Marcel.lí Domingo s/n. 43007 Tarragona. Tlf 977558855; Fax: 977558232. e-
mail: [email protected]
Palabras claves: Método tradicional, A. pasteurianus, RFLPs-16S rDNA, (GTG)5-PCR

3
RESUMEN
La producción del vinagre de vino por el método tradicional se encuentra en peligro debido
a los elevados riesgos inherentes al tiempo excesivo de producción, principalmente, por el
aporte limitado de oxígeno a las barricas y por el desconocimiento de la microbiota
presente durante la acetificación. El objetivo de este trabajo es realizar un estudio ecológico
de diferentes acetificaciones llevadas a cabo por el método tradicional en dos vinagrerías
(La Guinelle (Banyuls, Francia) y Mas d’en Gil (Priorat, España)) utilizando para ello
barricas nuevas de distintas maderas y grosores. Para la identificación de las bacterias
acéticas (BA) se emplearon técnicas de biología molecular, RFLPs-PCR del 16S rDNA a
nivel de especie y (GTG)5-PCR a nivel de cepa. Los resultados mostraron que, las
acetificaciones fueron más rápidas en la vinagrería de Banyuls, principalmente en el
arranque, lo cual puede ser debido a una mayor adaptación de las cepas autóctonas a estas
condiciones. En la vinagrería de Priorat hubo una parada de acetificación alargando
notablemente este proceso.
A nivel microbiológico, no se encontraron diferencias ni entre maderas en entre grosores.
En ambas vinagrerías, se impuso una cepa (AVF2 en Banyuls y AVT2 en Priorat) a lo largo
de todo el proceso. Estas cepas podrían ser buenas candidatas como cultivos iniciadores en
la producción de vinagre por el método tradicional.

4
INTRODUCCION
La producción de vinagre de vino por el método tradicional (cultivos superficiales de
bacterias acéticas, método Orleáns) se encuentra en peligro debido a los elevados riesgos
inherentes al tiempo excesivo de acetificación. La reducción del tiempo de acetificación
puede conseguirse de manera efectiva mediante aireación continua, hecho en el que se
fundamentan los métodos sumergidos de producción de vinagres industriales. El
inconveniente que conlleva este tipo de métodos es la reducción cualitativa del producto
final debido en parte a la pérdida aromática así como a la falta de contribución del
metabolismo de las bacterias acéticas, convertidas en biorreactores (Tesfaye et al., 2002).
En la actualidad, existe una gran demanda de vinagres de vino de alta calidad pero su
producción anual es baja, lo que encarece el producto. Además, es escaso el conocimiento
del proceso biotecnológico que se lleva a cabo, ya que el vinagre siempre ha sido
considerado como un subproducto del vino. Tradicionalmente la producción de estos
vinagres no se realiza ni en las barricas ni en las maderas adecuadas, pues la mayoría de
ellas son barricas muy viejas, donde la porosidad es prácticamente nula, con lo que hay baja
difusión de oxigeno. Las barricas utilizadas en vino están diseñadas para minimizar la
presencia de oxígeno y así evitar el desarrollo de las bacterias acéticas, exactamente al
contrario de aquello que es necesario para la producción de vinagre. Asimismo, la madera
más habitualmente utilizada es el roble, adecuada para el envejecimiento de los vinos, pero
no necesariamente para la producción de vinagres.
Las bacterias acéticas son los principales microorganismos implicados en la transformación
de etanol a ácido acético. Pocos son los estudios ecológicos realizados sobre bacterias
acéticas, la mayoría de ellos se han centrado en el vino (Du Toit and Lambrechts, 2002;
Bartowsky et al., 2003; González et al., 2004; 2005) en vinagres industriales (Sokollek et

5
al., 1998; Schüller et al., 2000), u otros tipos de vinagre: vinagre de caqui (Jin-Nam et al.,
2005) o vinagre de arroz (Nanda et al., 2001, Haruta et al., 2006). Ahora bien, ningún
estudio ha sido realizado en vinagre de vino obtenido por el método tradicional salvo el
realizado por Gullo et al., 2006 en vinagre balsámico tradicional.
La limitación fundamental para la realización de este tipo de estudios se debe al hecho de
que las bacterias acéticas son consideradas “fastidiosas” por su difícil cultivabilidad y
recuperación en medios de cultivo de laboratorio. Los problemas de cultivabilidad pueden
ser debidos a dos causas: Por un lado la ausencia de medios adecuados para su cultivo, pero
por otro la existencia del estado “viable pero no cultivable” (VBNC) (Millet and Lonvaud,
2000). Aunque no se puede asegurar que los medios de cultivo sean siempre los adecuados,
la recuperación de cepas de bacterias acéticas se ha venido realizando en medios de cultivo
óptimos, donde suelen demostrar un buen crecimiento. No obstante, es difícil asignar un
medio de cultivo “adecuado” para células provenientes de un medio extremo, como podría
ser el vinagre. Un problema adicional que justifica diferencias entre el crecimiento en placa
y los contajes en el microscopio es la asociación que tienen las bacterias acéticas, ya que es
frecuente observarlas en parejas, cadenas de longitud variable o grumos, lo que
generalmente desarrollaría una sola colonia.
Hasta ahora, los cultivos de bacterias acéticas utilizados en la producción de vinagres son
cultivos poco definidos y presumiblemente mixtos. Esto es debido, a que la identificación
de bacterias acéticas tradicionalmente se ha realizado mediante una serie de pruebas
fisiológicas y bioquímicas (De Ley et al., 1984), lo que ha impedido una buena selección y
mantenimiento de posibles cultivos iniciadores. Recientemente se han descrito técnicas de
biología molecular rápidas, fiables y de fácil manejo para su identificación, así tenemos,
para la identificación a nivel de género y especie la PCR-RFLPs (Restriction Fragment

6
Length Polymorphisms) del 16S rDNA y de los ITS 16S-23S rDNA (Ruiz et al., 2000); la
DGGE (Denaturing gradient gel electrophoresis) (López et al., 2003; De Vero et al., 2004);
FISH (Fluorescence in situ hybridisation) (Baena-Ruano et al., 2006). La tipificación a
nivel de cepa se realiza mediante técnicas como ERIC-PCR (Enterobacterial Repetitive
Intergenic Consensus) y REP-PCR (Repetitive Extragenic Palindromic) (González et al.,
2004); (GTG)5-PCR (Vuyst et al., 2006); RAPD-PCR (Random amplified polymorphic
DNA-PCR) (Nanda et al., 2001).
El presente estudio tiene como objetivo hacer un estudio ecológico de diferentes
acetificaciones realizadas por el método tradicional en dos vinagrerías que utilizan vinos de
la DOC Priorat (España) y la AOC Banyuls (Francia). Para la realización del estudio se han
empleado barricas nuevas de diferentes maderas y grosores. El uso de estas barricas tiene
como finalidad estudiar el efecto de estas dos variables (porosidad y grosor de la madera)
sobre la velocidad de acetificación y la evolución de la microbiota a lo largo de estos
procesos.

7
MATERIALES Y MÉTODOS
Muestreo y condiciones de acetificación
Las acetificaciones se llevaron a cabo en dos vinagrerías: Mas d’en Gil (Priorat, Tarragona)
y La Guinelle (Banyuls, Francia) durante los años 2005-2006. El estudio se realizó por
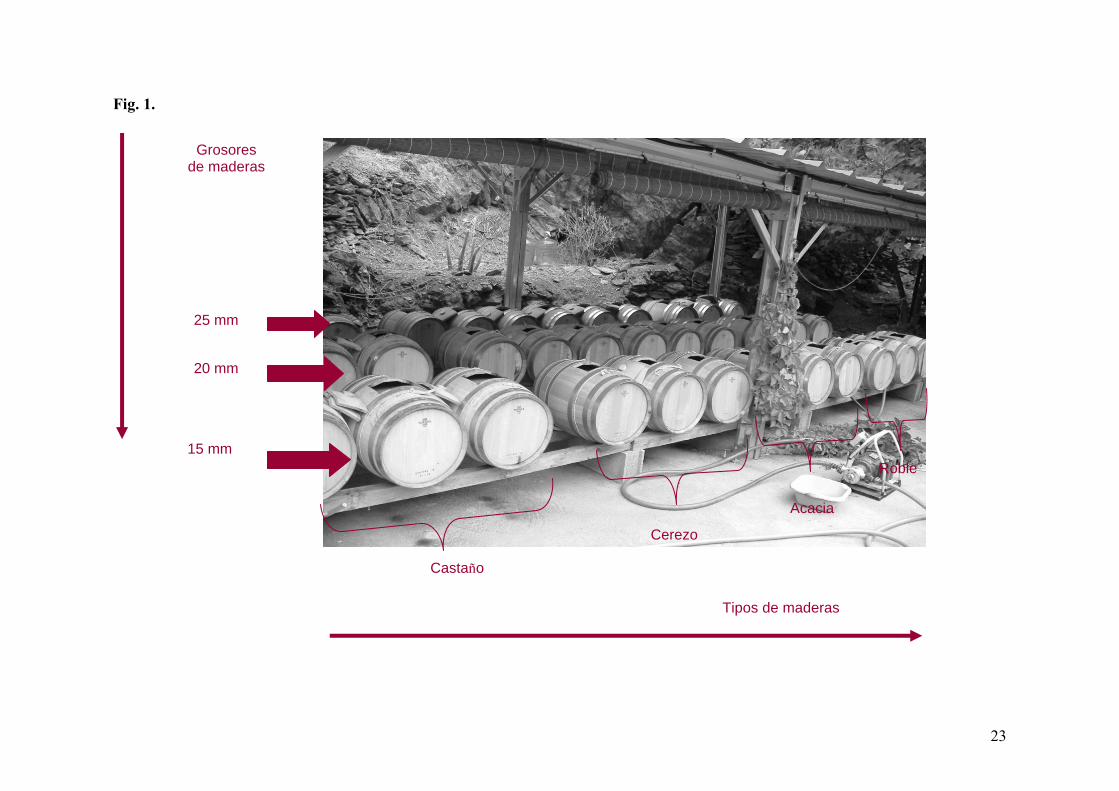
triplicado en barricas de 60 litros de diferentes maderas y grosores (Fig. 1). Las maderas
utilizadas fueron la acacia, el cerezo, el castaño y el roble americano, siendo éste último el
más utilizado en la producción de vinagres. Por lo que respecta a los grosores, se fabricaron
barricas con tres grosores diferentes, 15, 20 y 25 mm.
Para las acetificaciones se utilizó tanto el vino habitual en cada una de las vinagrerías como
los cultivos superficiales desarrollados en cada vinagrería (madres del vinagre), que
correspondían en todos los casos a cultivos mixtos.
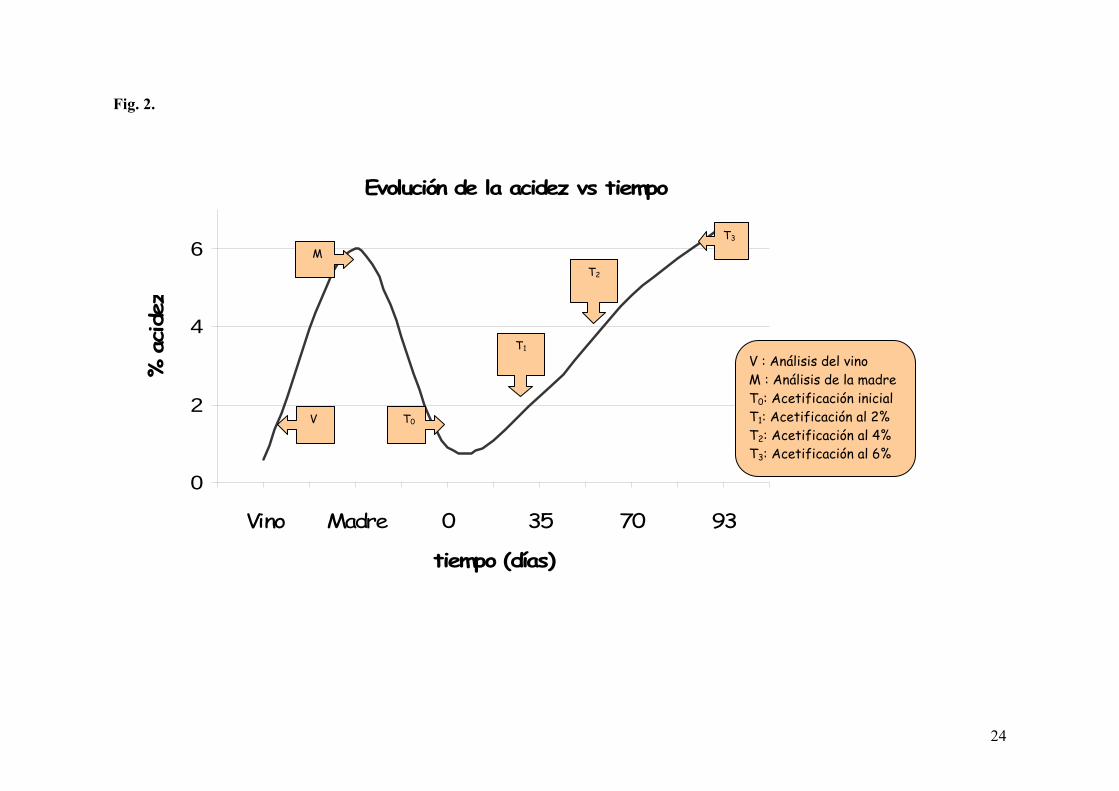
Se tomaron muestras en diferentes puntos de la acetificación: T0 (inicio), T1 (2% acidez), T2
(4% acidez), T3 (6% acidez) (Fig. 2). En dichos puntos, se realizaron controles de
temperatura, oxígeno disuelto, etanol, pH y recuento de BA por microscopía y siembra en
placa. Dicha siembra se realizó en un sembrador automático en espiral WASP II (Don
Whitley Scientific Limited, England) en medio agar GY (1% de extracto de levadura
(Cultimed, Barcelona, España); 5% de glucosa (Cultimed); 1,5% de Agar (Cultimed) (g/l),
suplementado con natamicina E-325/Delvocid (100 mg/l) (DSM; Delft; The Netherlands)
para inhibir el crecimiento de levaduras y hongos. Todas las muestras fueron incubadas a
28ºC durante 2-4 días. Se seleccionaron 10 colonias aleatoriamente y fueron replicadas
nuevamente en placas del mismo medio de cultivo con 3% de CaCO3 con el propósito de
asegurar que las colonias aisladas producían ácido acético.

8
La determinación del ácido acético (acidez total) se efectúa mediante una valoración ácido-
base en presencia de fenolftaleína como indicador. La cantidad de etanol y de azúcares
residuales presentes en la muestra se realizó mediante un kit enzimático comercial (Roche
Diagnostics, Mannheim, Germany), según las especificaciones del fabricante.
La velocidad máxima de acetificación es la máxima pendiente obtenida al representar la
producción de ácido acético frente al tiempo. Este valor se expresa en mg de ácido acético
obtenidos por litro en un día (mg l-1 día-1). Se considera pérdida de etanol la cantidad de
etanol consumida que no ha sido transformada en ácido acético. Para realizar este cálculo
se ha tenido en cuenta el inicio y el final de cada acetificación en las diferentes maderas y
grosores de las barricas.
Análisis de RFLPs-PCR del 16S rDNA y (GTG)5-PCR
La extracción del DNA se realizó mediante el método CTAB de Ausubel et al., (1992).
Para la identificación a nivel de especie, se utilizó la técnica de los RFLPs del 16S rDNA
descrita por Ruiz et al., (2000). El producto amplificado del gen del 16S rDNA fue digerido
con dos enzimas de restricción: TaqI y AluI. Adicionalmente, se utilizó el enzima BccI para
diferenciar las especies Ga. hansenii, Ga. europaeus y Ga. xylinus que presentan el mismo
perfil con los dos enzimas anteriores. Para la tipificación de las cepas se utilizó la técnica
del (GTG)5-PCR (Vuyst et al., 2006). De forma rutinaria las muestras se analizaron en
geles de agarosa y sólo aquellos perfiles diferentes se pasaron por el Bioanalizador Agilent
2100 (Agilent Technologies, Böblingen, Germany) utilizando el microchip 7500 de DNA
para determinar de manera precisa el peso molecular de cada una de sus bandas.

9
La diversidad se calcula como el porcentaje de cepas diferentes (sin importar la especie)
entre el total de colonias analizadas.
RESULTADOS
Las acetificaciones en ambas vinagrerías se comenzaron en agosto del 2005. En cada una
de ellas se pusieron en marcha un total de 36 barricas, repartidas en 4 maderas y 3 grosores
de cada una. Para este estudio se hicieron análisis físico-químicos y microbiológicos del
vino, la madre y el vinagre durante los diferentes puntos del proceso.
En la Tabla 1 se muestran los resultados obtenidos para el vino, la madre y el punto de
inicio de la acetificación. En ella se pueden observar claras diferencias entre los vinos
utilizados. Cabe destacar la alta concentración de azúcares en el vino de Banyuls que se ha
utilizado en la vinagrería La Guinelle.
A lo largo de las acetificaciones los parámetros de temperatura y oxígeno no mostraron
diferencias ni entre maderas ni entre grosores. La concentración de oxígeno se mantuvo en
niveles muy bajos (aprox. 5 mg/ml) en todos los puntos estudiados, salvo en los casos en
los que se produjeron paradas de fermentación, en los cuales, la concentración de éste
aumentó.
Cinética de la acetificación
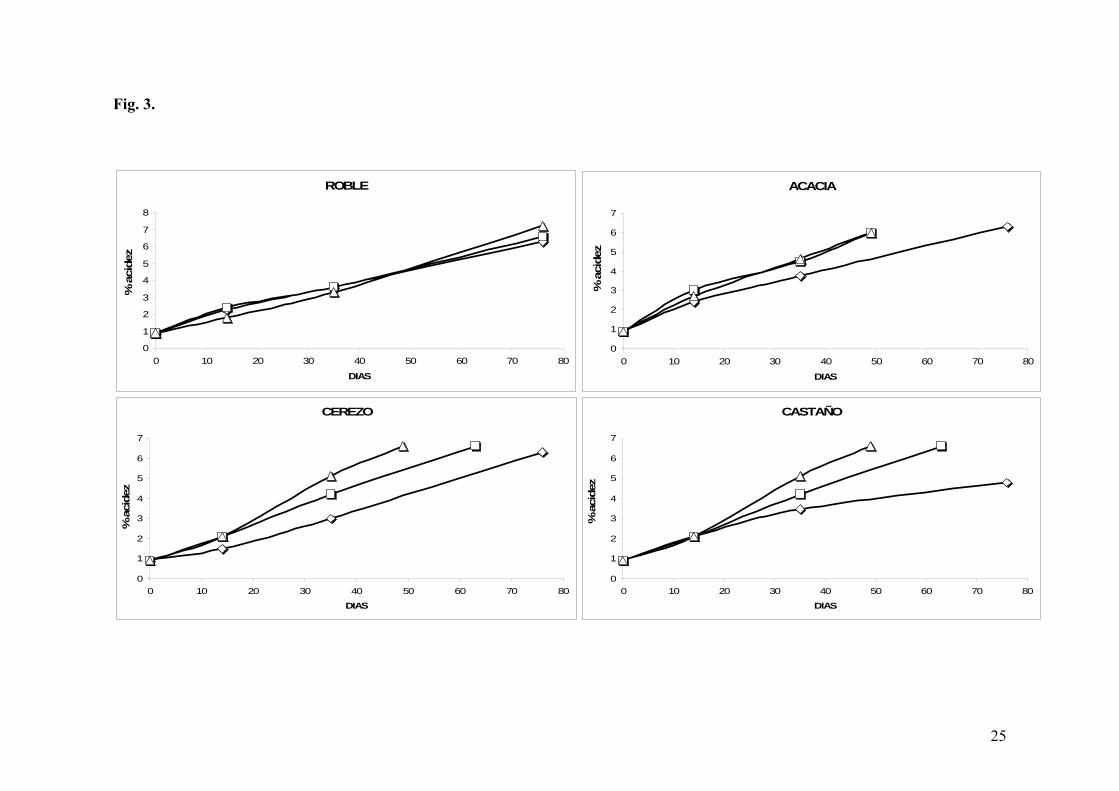
En la vinagrería La Guinelle, todas las acetificaciones tardaron en finalizar entre 49 y 76
días. Con respecto a los grosores, los resultados fueron diferentes a lo postulado. Así, se
observó que el grosor de 15 mm presentaba una ralentización frente a 20 mm y 25 mm,
siendo éste último el que presentó una mayor reducción en el tiempo de acetificación (Fig.

10
3). Excepcionalmente, este comportamiento no se cumplió para el caso del roble, donde los
tres grosores presentaron cinéticas muy similares. Estas diferencias pudieron ser explicadas
posteriormente con los resultados microbiológicos.
Con respecto a las maderas, la acacia resultó ser la mejor a nivel cinético ya que en todos
los grosores, las velocidades de arranque fueron superiores a las otras maderas (resultados
no mostrados).
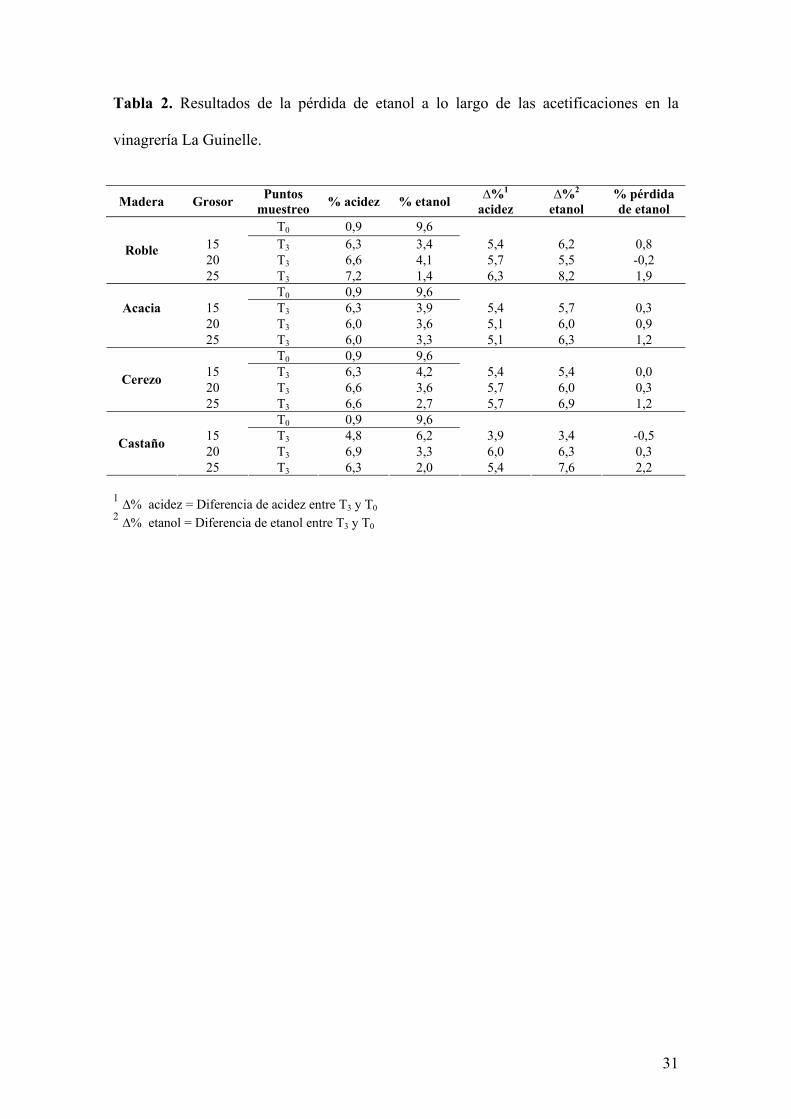
En la tabla 2 se presentan los resultados de la pérdida de etanol a lo largo de las
acetificaciones. El resultado más relevante es que los valores de pérdida de etanol son
inversamente proporcionales al grosor de la barrica y directamente proporcionales a la
velocidad de la acetificación. Por tanto, la mayor pérdida se produce en las acetificaciones
realizadas con barricas de 25 mm. En el caso de la acacia, en la que las acetificaciones de
20 mm y 25 mm se comportaron de manera similar, se observa también una pérdida de
etanol semejante. Esta mayor velocidad parece favorecer una posible pérdida por
evaporación.
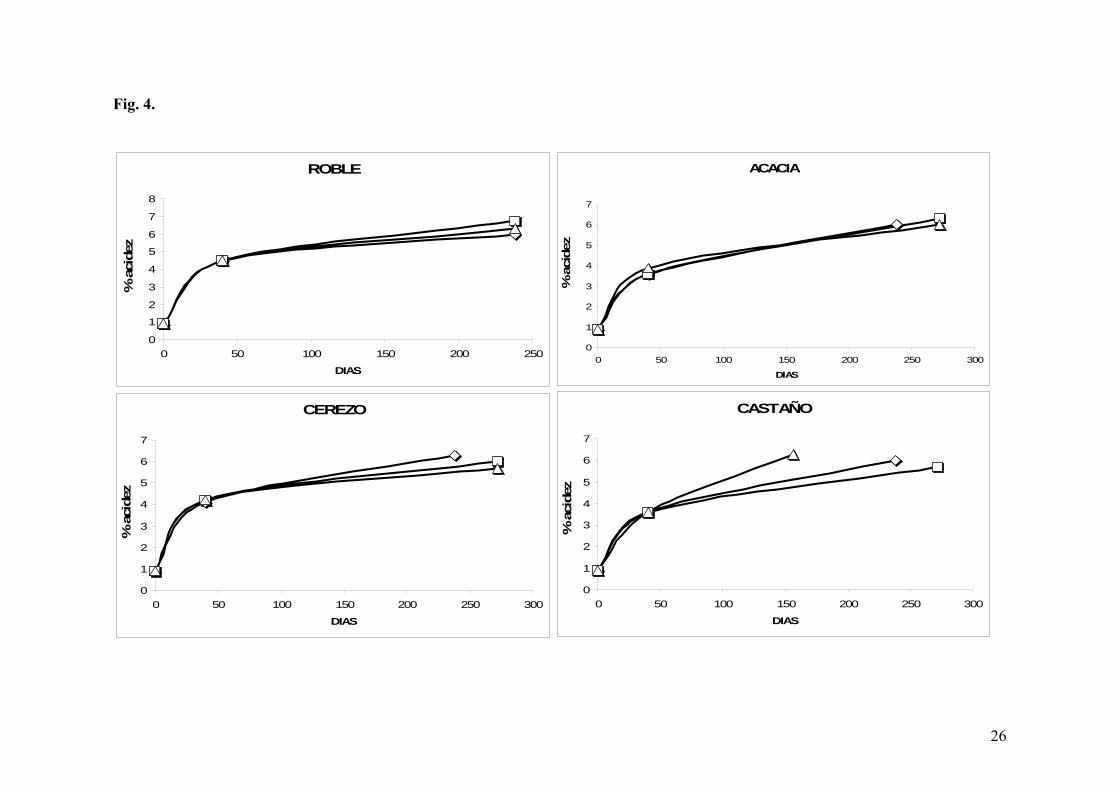
En la vinagrería Mas d’en Gil, todas las acetificaciones tardaron en finalizar entre 156 y
272 días. Este largo período de acetificación se debió a una ralentización al inicio del
proceso ligada a un drástico descenso de las temperaturas con la llegada del invierno,
teniendo como consecuencia una parada de la acetificación con una acidez aproximada del
5% (Fig. 4).
Con respecto tanto a los grosores como a las maderas, no se observaron diferencias
significativas excepto en el caso del castaño, donde de nuevo, la barrica de grosor 25 mm
presentó una acetificación claramente más rápida que los otros dos grosores.
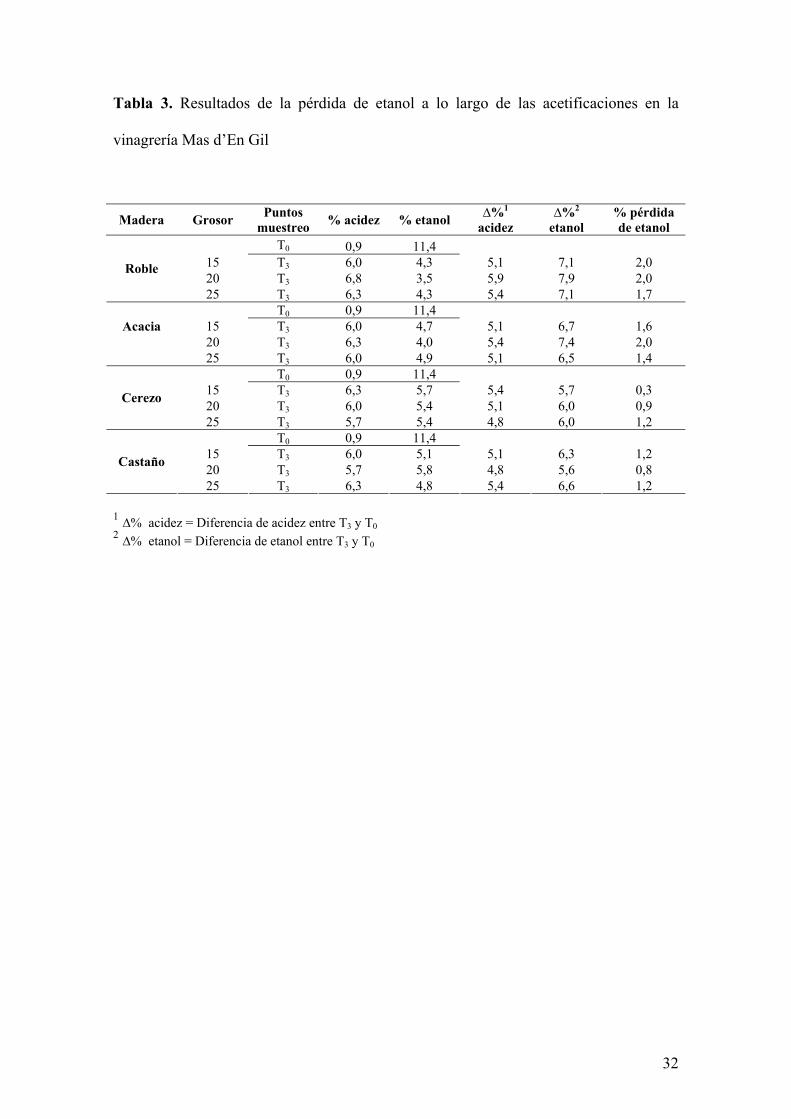
A diferencia de lo que sucedía en la vinagrería La Guinelle, en este caso, los resultados de
la pérdida de etanol no presentaron correlación ni con los grosores de las barricas ni con la

11
velocidad de acetificación (Tabla 3). A pesar de ello, las pérdidas en esta vinagrería fueron
importantes, alcanzando valores hasta del 2%. Esto podría relacionarse con la larga
duración de estas acetificaciones.
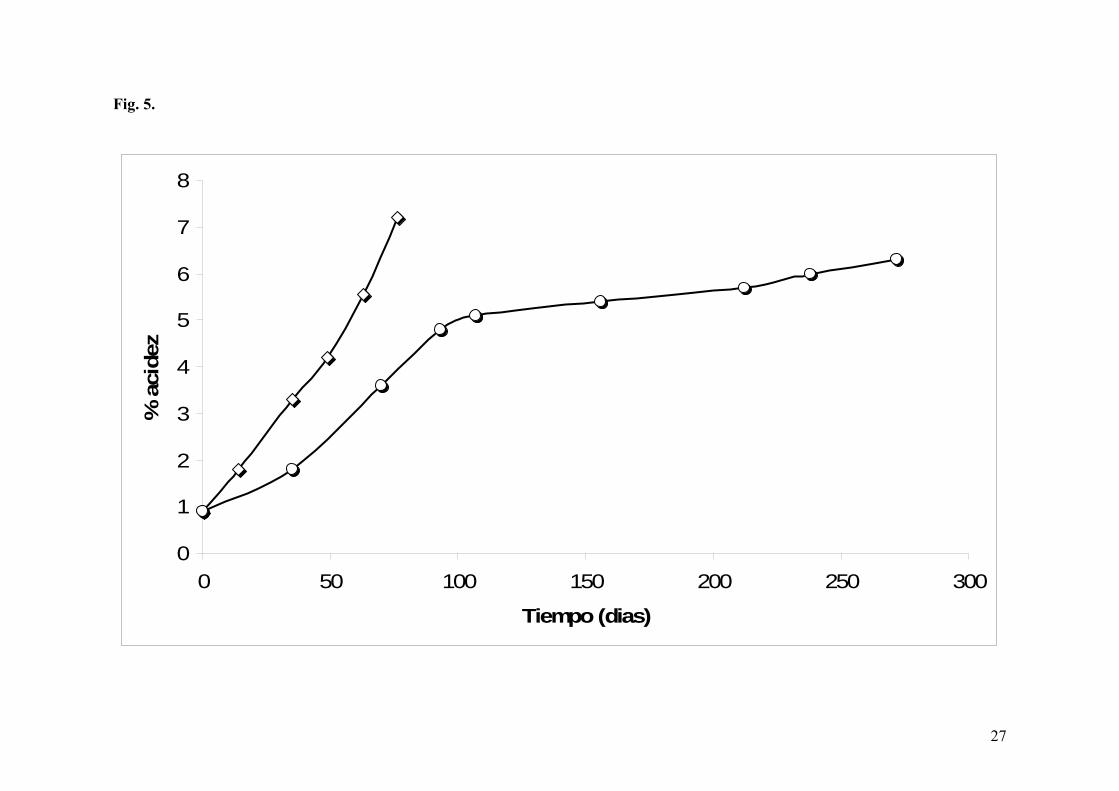
En la Fig. 5 se comparan las cinéticas de las acetificaciones llevadas a cabo en ambas
vinagrerías. Estas acetificaciones tuvieron un comportamiento desigual debido
principalmente a la diferente velocidad de arranque. En Banyuls, ésta fue mayor que en el
Priorat, probablemente por una mejor adaptación de las bacterias acéticas al medio. Esta
mayor velocidad ha permitido un desarrollo más rápido de la acetificación. Como ya hemos
comentado anteriormente, en el Priorat además del retardo en el inicio de la acetificación se
produjo una parada. De todas maneras, este retardo también podría ser debido al alto grado
alcohólico de la mezcla inicial (T0: 11,4% vs. 9,6% en Banyuls).
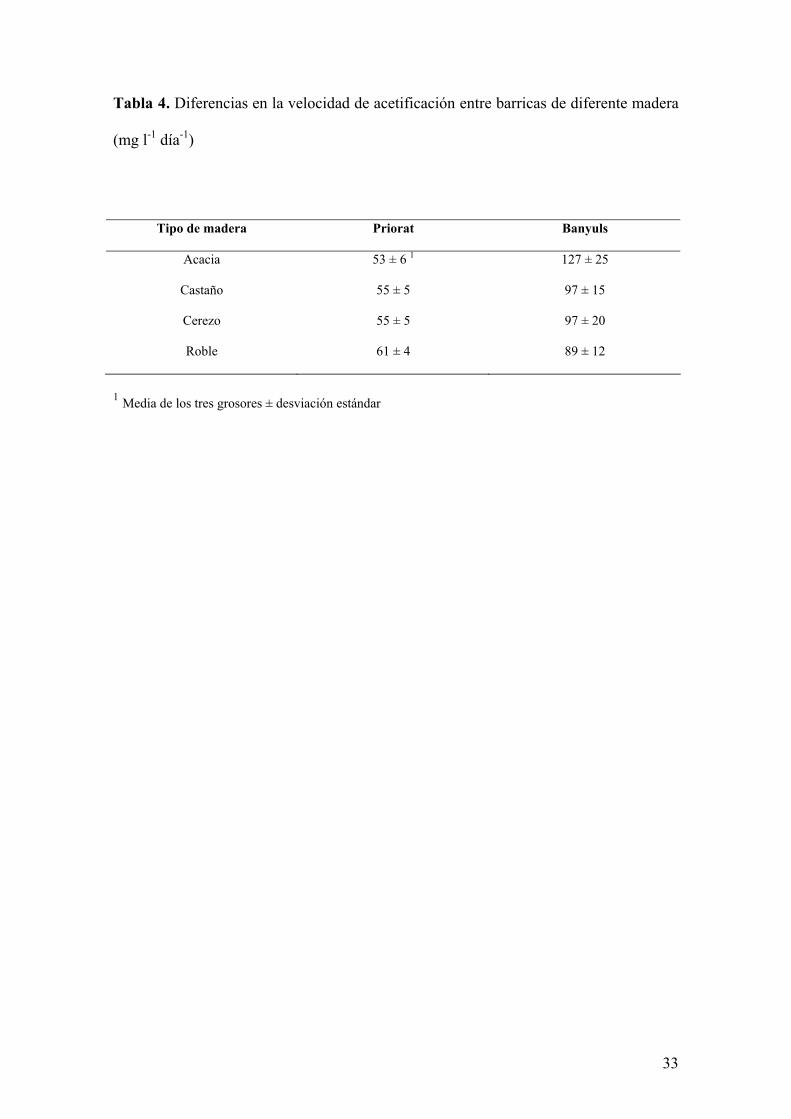
En la Tabla 4 se muestran las diferencias en la velocidad máxima de acetificación entre los
diferentes tipos de maderas en las dos vinagrerías. En la vinagrería de Priorat, las
velocidades en todos los casos son muy similares, mientras que en la vinagrería de Banyuls,
como ya comentamos anteriormente, la acacia destaca sobre el resto de maderas.
Parámetros microbiológicos
A nivel poblacional no se observaron grandes diferencias entre las dos vinagrerías. En
ambos casos, los contajes totales fueron de aproximadamente 108 cél/ml manteniéndose
estable a lo largo de toda la acetificación. Por lo que respecta al contaje en placa, los
valores obtenidos en ambos casos fueron de 104-105 ufc/ml. Esta diferencia entre ambos
contajes podría ser debida a una posible asociación celular o al estado de viabilidad pero no
cultivabilidad descrito anteriormente para bacterias acéticas. Para minimizar este estado de
“no cultivables”, se probaron diferentes medios de cultivos (GY, medio manitol, medio AE,

12
medio RAE, GY: vino filtrado (50:50)), siendo el más efectivo el medio GY (resultados no
mostrados). Esto implica que la identificación se ha reducido a las bacterias cultivables.
Identificación a nivel de especie y cepa
El análisis microbiológico se realizó a un total de 310 colonias en el caso de La Guinelle y
274 en el Mas d’en Gil. Sólo se estudiaron los aislamientos que formaron halo en un medio
GY con carbonato de calcio. A pesar de las variaciones encontradas a nivel cinético entre
grosores de barrica, principalmente en la vinagrería de Banyuls, estas diferencias no fueron
corroboradas con los análisis microbiológicos. Por tanto, los resultados de los grosores se
agruparan dentro de la misma madera, considerándose como réplicas. Para este estudio se
analizaron las muestras de vino, madre, T0, T1, T2 y T3.
En Banyuls, la cepa mayoritaria tanto en la madre como en T0 fue la cepa AVF2
perteneciente a la especie A. pasteurianus (92% en la madre y 95% en T0). Las otras cepas
minoritarias identificadas en estos puntos corresponden, en el caso de la madre, a una cepa
(AVF3, 8%) perteneciente a la especie de Ga. europaeus, y en el punto T0, a otra cepa
(AVF4, 5%) perteneciente a la especie A. pasteurianus. En el vino, no se encontró ninguna
bacteria acética.
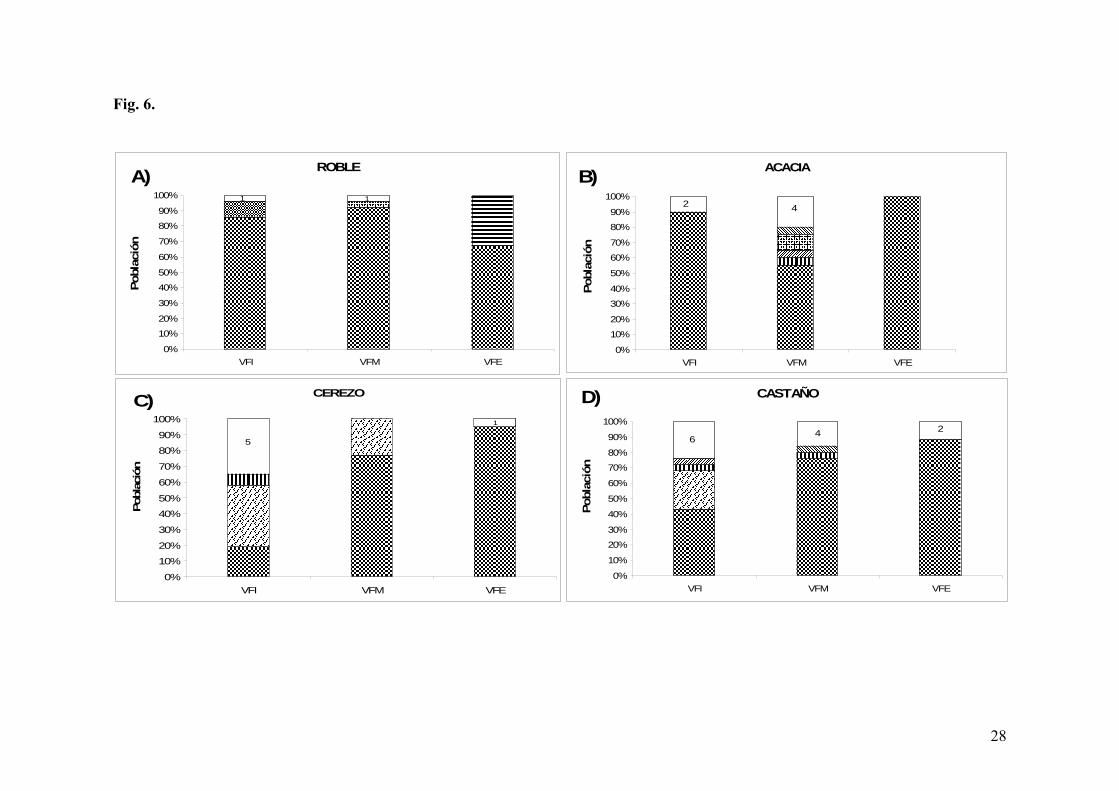
En cuanto al análisis en los diferentes puntos de acetificación, en el punto T1, inicio de
acetificación, se mantuvo la clara imposición de la cepa AVF2 tanto en el roble como en la
acacia (Fig. 6, a y b). En las acetificaciones realizadas en cerezo y castaño (Fig. 6, c y d),
además de la cepa AVF2, se observó una mayor diversidad de cepas, entre las cuales
destaca la cepa AVF10 (Tabla 5).

13
Por lo que respecta al punto T2, mitad de acetificación, la cepa AVF2 fue la cepa
mayoritaria en todos los casos, siendo en la acacia donde presentó una menor imposición
debido a la mayor diversidad detectada (Fig. 6b y Tabla 5).
Por último en el punto T3, final de acetificación, además de la clara imposición de la cepa
AVF2, lo más relevante es el alto porcentaje de una cepa de la especie Ga. europaeus
(AVF34, 33%) en la acetificación realizada en roble, concretamente en el grosor de 25 mm.
Este hecho explicaría la ralentización observada en la cinética del roble 25 mm respecto al
resto de maderas de grosor 25 mm.
En la tabla 5 se presentan los resultados de identificación a nivel de especie de las cepas
analizadas durante las acetificaciones. En todos los casos, las cepas aisladas pertenecían a
la especie A. pasteurianus, a excepción de la cepa AVF34 identificada como Ga.
europaeus.
En la vinagrería Mas d’en Gil, tanto en la madre como en el punto T0 se aislaron 3 cepas,
únicamente una cepa (AVT2) fue coincidente en ambas muestras. Esta cepa identificada
como A. pasteurianus fue la mayoritaria en ambos casos (55% en la madre y 50% en T0).
El resto de cepas apareció en un porcentaje menor: en la madre AVT3 (27%) y AVT4
(18%) y en T0, AVT5 (42%) y AVT6 (8%). Cada una de ellas perteneciente a una especie
diferente: A. pasteurianus, Ga xylinus, G. oxydans y Ga. europaeus, respectivamente. En el
caso del vino, no se identificó ninguna bacteria acética.
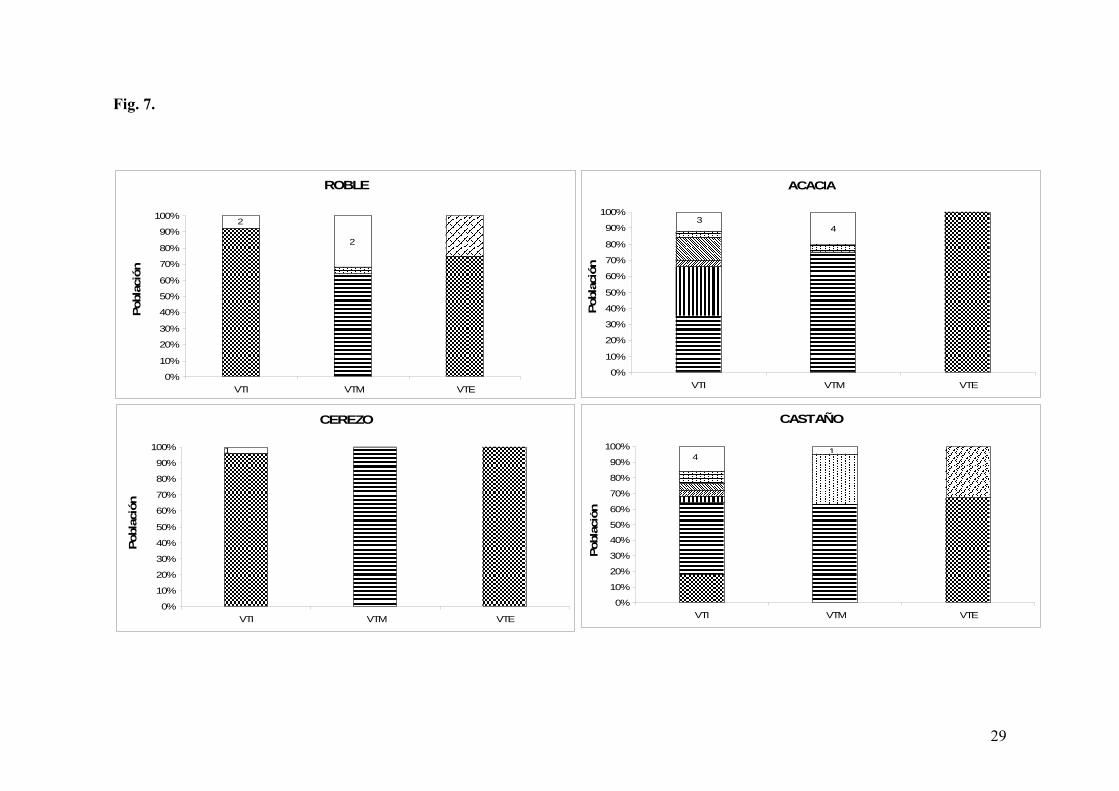
En el punto T1, inicio de acetificación, la cepa AVT2 se impuso tanto en el roble como en
el cerezo (>90%; Fig. 7, a y c). Por el contrario, en el castaño y en la acacia, la presencia de
esta cepa fue mínima o nula. En estas dos maderas, se observó un mayor número de cepas
diferentes (Tabla 6), entre las que destacan las cepas AVT9 (45 % en castaño y 35% en
cerezo) y AVT10 (30% acacia) (Fig. 7, b y d).

14
En el punto T2, mitad de acetificación, la cepa AVT9 es la cepa mayoritaria en todas las
maderas, siendo incluso de un 100% en cerezo. Cabe destacar que la cepa AVT2 no fue
detectada en ninguna de las maderas.
Por último, en el punto T3, final de acetificación, se impone de nuevo la cepa AVT2 en un
100% en cerezo y castaño, y en más de un 70% en las otras dos maderas, habiendo un claro
descenso de la diversidad de cepas (Tabla 6).
Respecto a la identificación a nivel de especie, todas las cepas aisladas pertenecían a la
especie A. pasteurianus (Tabla 6).
DISCUSIÓN
En los últimos años se han realizado diferentes estudios sobre bacterias acéticas en vino
(Du Toit and Lambrechts, 2002; Bartowsky et al., 2003; González et al., 2004; 2005), así
como, en vinagres industriales (Sokollek et al., 1998; Schüller et al., 2000), y otros tipos de
vinagre: vinagre de caqui (Jin-Nam et al., 2005) o vinagre de arroz (Nanda et al., 2001,
Haruta et al., 2006), pero prácticamente nada se conoce sobre las bacterias responsables de
la producción de vinagres de vino por el método tradicional (Gullo et al., 2006) tanto a
nivel poblacional como ecológico.
Además, el proceso de producción de vinagres por este método se encuentra en peligro
debido a los elevados riesgos inherentes al tiempo excesivo de producción. Una reducción
efectiva de este tiempo de acetificación podría conseguirse mediante un aumento de la
aireación. Por ello, en este trabajo se lleva a cabo un estudio del uso de barricas de
diferentes maderas y grosores como alternativa para mejorar la difusión de oxígeno y por
tanto, acelerar el proceso de acetificación.

15
De los tres grosores ensayados, la duración de las acetificaciones en las barricas de 25 mm
fue la menor, seguida de las de 20 mm y 15 mm. Estos resultados se contradicen con la
hipótesis formulada, ya que era de esperar una mayor difusión de oxígeno en las barricas de
15 mm. En cuanto a maderas, tampoco se obtuvieron resultados concluyentes. Esto parece
indicar que la mayor difusión de oxígeno no esté tan relacionada con el grosor o madera de
la barrica como con la superficie de contacto con el aire. Por tanto, quizás sería de interés
centrarse en un aumento de dicha superficie de contacto, variando por ejemplo, la forma de
la barrica. De todas maneras, otro parámetro a tener en cuenta es la posible pérdida de
etanol por evaporación. Dicha pérdida está en función de diferentes parámetros como el
tamaño de la barrica, la temperatura, la humedad relativa o la circulación del aire alrededor
del barril (Guymon, 1972; Guymon y Crowell, 1970). Se conoce que la falta de etanol en el
medio provoca que algunas bacterias acéticas sean capaces de oxidar el ácido acético a
productos no deseados como son el agua y dióxido de carbono. Por ello, la elección de una
buena barrica para este tipo de procesos será un compromiso entre una rápida acetificación
y una baja pérdida de etanol por evaporación.
Las acetificaciones en las dos vinagrerías estudiadas tuvieron un comportamiento diferente
debido principalmente a la velocidad de arranque. El hecho de que esta velocidad sea
menor en Mas d’en Gil puede deberse a que se trata de una vinagrería nueva frente a una
vinagrería de una cierta tradición, como puede ser la vinagrería de La Guinelle. Por otro
lado, cabe señalar que los vinos utilizados fueron muy diferentes, en Banyuls se utilizan
vinos con residuos de azúcares y poco sulfitados, mientras que en Priorat son vinos secos
(sin azúcar) y con niveles de sulfitación elevados. A esto hay que añadir el alto grado
alcohólico del T0 observado en el caso del Priorat. La consecuencia más importante de esta
menor velocidad de arranque fue que estas acetificaciones se alargaron, en alguno de los

16
casos, hasta más de 8 meses. Esto también se debió al descenso de las temperaturas
provocado por la entrada del invierno, dando lugar a una parada que duró hasta la llegada
de la primavera con el consecuente aumento de la temperatura.
En cuanto a lo que al aspecto microbiológico se refiere, parece claro que, en nuestro estudio
la especie responsable de la producción de los vinagres es Acetobacter pasteurianus, a
diferencia de lo que sucede en el estudio de Gullo et al., 2006 donde en la producción de
vinagre balsámico por el método tradicional la especie predominante es
Gluconoacetobacter xylinus. De todas maneras, se trata de productos de características muy
diferentes y por tanto, ya era de esperar esta diferencia a nivel microbiológico. Lo mismo
ocurre en la producción de vinagres por cultivos sumergidos donde las especies
mayoritarias también pertenecen al género Gluconoacetobacter como pueden ser Ga.
europaeus (Sievers et al., 1992, Trcek et al., 2000), Ga entanii (Schüller et al., 2000), Ga.
obodiens (Sokollek et al., 1998), Ga intermedius (Boesch et al., 1998, Trcek et al., 2000).
A nivel de cepa, en ambas vinagrerías hubo una imposición de una sola cepa en todas las
acetificaciones. En el caso de La Guinelle, la cepa AVF2 fue la predominante a lo largo de
toda la acetificación, encontrándose tanto en la madre como en el resto de puntos
analizados. Por su parte, en la vinagrería Mas d’en Gil, a pesar de ser la cepa AVT2 la
mayoritaria en la madre y la que finalizó la acetificación, ésta no fue detectada en el punto
medio de la acetificación. En este punto, la cepa AVT9 fue la cepa predominante. Esta
alternancia entre ambas cepas puede explicarse porque entre la toma de muestra del punto
T2 y T3 hubo una parada de la acetificación de 6 a 8 meses. Durante este período se
desconoce que pudo suceder a nivel microbiológico en las diferentes barricas pero el hecho
de que la cepa AVT2, aislada en la madre y T0, T1, reapareciese a final de la acetificación
(T3) hace pensar que se trate de una cepa que resiste la combinación de alto grado acético

17
(5%) y bajas temperaturas. Por el contrario, la cepa AVT9 no fue detectada al final del
proceso, indicando que se trata de una cepa que no fue capaz de resistir estas condiciones.
Como resultado de este trabajo, las dos cepas mayoritarias encontradas en estas vinagrerías,
AVF2 en La Guinelle y AVT2 en Mas d’en Gil, parecen ser buenas candidatas para su
utilización como iniciadores con alta capacidad acetificadora en la producción de vinagres
de vino por el método tradicional.
AGRADECIMIENTOS
Este trabajo se ha realizado gracias a la financiación de los proyectos: CRAFT nº 017269
del 6º Programa Marco de Investigación de la Unión Europea y nº AGL2004-07494-C02-
02 del Ministerio de Educación y Ciencia.

18
BIBLIOGRAFIA
Ausubel FM., Brent R., Kingston RE., Moore DD., Seidman JG., Smith JA., Struhl K.
(Eds). (1992). Short protocols in molecular biology. 2nd ed. London. Jonh Willey & Sons
Inc.
Baena-Ruano S., Jiménez-Ot C., Santos-Dueñas I.M., Cantero-Moreno D. (2006).
Rapid method for total, viable and non-viable acetic acid bacteria determination during
acetification process. Process Bioch. 41, 1160-1164
Bartowsky, E.J., Xia, D., Gibson, R.L., Fleet, G.H. & Henschke, P.A. (2003). Spoilage
of bottled red wine by acetic acid bacteria. Lett Appl Microbiol 36, 307-314
Boesch, C., Trcek, J., Sievers, M. & Teuber, M., (1998). Acetobacter intermedius, sp.
nov. Syst Appl Microbiol 21, 220-229
De Ley J., Gilli M., Swings J. (1984). Family VI. Acetobacteraceae. In: Krieg, N.R, Holt,
J. G. (Eds). Bergey’s manual of systematic bacteriology, vol. 1. Williams and Wilkins,
Baltimore. Pp. 267-278.
De Vero, L., Gullo, M., Solieri, L., Landi, S. & Giudici, P. (2004). A new approach to
study the acetic acid bacteria: the DGGE. P. 27-37. In I microorganismi dell’aceto
balsamico. Modena. Italy.
De Vuyst L, Camu N., De Winter T., Van de Perre V., Swings J., Cleenwerck I.,
Vancanneyt M. (2006). Development of a high throughput (GTG)5-PCR fingerprint
method for the identification of acetic acid bacteria. Segundas Jornadas de I+D+I en la
Elaboración de Vinagres. Córdoba-España
Du Toit, W.J. & Lamberchts, M.G. (2002). The enumeration and identification of acetic
acid bacteria from South African red wine fermentations. Int J Food Microbiol 74, 57-64

19
González A., Hierro N., Guillamón J.M., Mas A., Poblet M. (2006). Enumeration and
detection of acetic acid bacteria by real-time PCR and nested-PCR. FEMS Microbiol. Let.
254, 123-128
González A., Hierro N., Poblet M., Mas A., Guillamón JM. (2005). Application of
molecular methods to demonstrate species and strain evolution of acetic acid bacteria
population during wine production. Int. J. Food. Microbiol. 102, 295-304.
González A., Hierro N., Poblet M., Rozés N., Mas A., Guillamon JM. (2004).
Application of molecular methods for the differentiation of acetic acid bacteria in a red
wine fermentation. J. Appl. Microbiol. 96, 853-860.
Gullo M, Caggia C., De Vero, L., Guidici P. (2006). Characterization of acetic acid
bacteria in “traditional balsamic Vinegar”. Int. J. Food Microbiol. 106, 209-212
Guymon J.F. (1972). Influence of warehouse temperatures on the aging of California
brandy. Wines & Vines, 54(1), 36-38.
Guymon J.F. y Crowell E.A. (1970). Brandy aging, some comparisons of american and
french oak cooperage. Wines & Vines, 51(1), 23-25.
Haruta S, Ueno S, Egawa I, Hashiguchi K, Fujii A, Nagano M, Ishii M, Igarashi Y.
(2006). Succession of bacterial and fungal communities during a traditional pot
fermentation of rice vinegar assessed by PCR-mediated denaturing gradient gel
electrophoresis. Int. J. Food Microbiol. 109, 79-87
Jin-Nam K., Jong-Sok Ch., Young-Jung W., Jong-Sun Y., Hwa-Won R.
(2005). Culture medium optimization for acetic acid production by a persimmon vinegar-
derived bacterium. Appl. Bioch. and Biotech. 123, 1-3
Lopez, I., Ruiz-Larrea, F., Cocolin, L., Orr, E., Phister, T., Marshall, M.,
VanderGheynst, J. & Mills, D. A. (2003). Design and evaluation of PCR primers for

20
analysis of bacterial populations in wine by denaturing gradient gel electrophoresis. Appl
Environ Microbiol 69, 6801-6807
Millet, V. & Lonvaud-Funel, A. (2000). The viable but non-culturable state of wine
microorganisms during storage. Lett Appl Microbiol 30, 126-141
Nanda, K., Taniguchi, M., Ujike, S., Ishihara, N., Mori, H., Ono, H. & Murooka, Y.
(2001). Characterization of acetic acid bacteria in traditional acetic acid fermentation of
rice Vinegar (komesu) and unpolished rice vinegar (kurosu) produced in Japan. Appl
Environ Microbiol 67, 986-990
Ruiz, A., Poblet, M., Más, A. & Guillamón, JM. (2000). Identification of acetic acid
bacteria by RFLP of PCR-amplified 16S rDNA and 16S-23S rDNA intergenic spacer. Int. J
Syst Evol Microbiol 50, 1981-1987
Sievers, M., Sellmer, S. & Teuber, M. (1992). Acetobacter europaeus sp. nov., a main
component of industrial vinegar fermenters in Central Europe. Syst Appl Microbiol 15, 386-
392.
Sokollek, SJ., Hertel, C. & Hammes, WP., (1998). Description of Acetobacter oboediens
sp. nov. and Acetobacter pomorum sp. nov., two new species isolated from industrial
vinegar fermentations. Int J Syst Bacteriol 48, 935-940
Schüller G., Hertel C., Hammes W.P. (2000). Gluconacetobacter entanii sp. nov., isolated
from submerged high-acid industrial vinegar fermentations. Int. J. Syst. Evol. Microbiol.,
50, 2013-2020.
Tesfaye W., Morales M.L., García-Parrilla M.C., Troncoso A.M. (2002). Wine
Vinegar: technology, authenticity and quality evaluation. Trends Food Sc. & Tech. 13, 12-
21

21
Trcek J., Raspor P., Teuber M. (2000). Molecular identification of Acetobacter isolates
from submerged vinegar production, sequence analysis of plasmid pJK2-1 and application
in the development of a cloning vector. Appl Microbiol Biotechnol 53: 289-295

22
LEYENDA DE FIGURAS
Fig. 1. Disposición de las barricas en la vinagrería de Banyuls.
Fig. 2. Evolución de la acetificación, acidez vs. tiempo.
Fig. 3. Evolución de la acidez vs. tiempo en La Guinelle, en función de los grosores:
15mm (◊), 20mm (□) y 25mm(∆)
Fig. 4. Evolución de la acidez vs. tiempo en el Priorat, en función de los grosores:
15mm (◊), 20mm (□) y 25mm(∆)
Fig. 5. Evolución de la acidez vs. tiempo en ambas vinagrerías. La Guinelle (◊◊), Mas
d’en Gil (○○)
Fig. 6. Porcentajes de las diferentes cepas de bacterias acéticas aisladas en las diferentes
maderas a lo largo de las acetificaciones en la vinagrería La Guinelle (Banyuls):
AVF2, AVF6, AVF10, AVF13, AVF19, AVF23, AVF28,
AVF34, OTROS (los números indican el número de diferentes cepas
minoritarias <10%)
Fig. 7. Porcentajes de las diferentes cepas de bacterias acéticas aisladas en las diferentes
maderas a lo largo de las acetificaciones en la vinagrería Mas d’en Gil (Priorat):
AVT2, AVT9, AVT10, AVT13, AVT14, AVT16,
AVT34, AVT35, AVT36, OTROS (El número dentro de las barras
corresponde al número de cepas minoritarias <10%)
23
Fig. 1.
15 mm
20 mm
25 mm
Roble
Acacia
Cerezo
Castaño
Tipos de maderas
Grosores de maderas
24
Fig. 2.
Evolución de la acidez vs tiempo
0
2
4
6
Vino Madre 0 35 70 93
tiempo (días)
% acide
z
T1
T2
T0
T3
V
M
V : Análisis del vino M : Análisis de la madre T0: Acetificación inicial T1: Acetificación al 2% T2: Acetificación al 4% T3: Acetificación al 6%
25
Fig. 3.
ROBLE
0
1
2
3
4
5
6
7
8
0 10 20 30 40 50 60 70 80
DIAS
% a
cide
z
ACACIA
0
1
2
3
4
5
6
7
0 10 20 30 40 50 60 70 80
DIAS
% a
cide
z
CEREZO
0
1
2
3
4
5
6
7
0 10 20 30 40 50 60 70 80
DIAS
% a
cide
z
CASTAÑO
0
1
2
3
4
5
6
7
0 10 20 30 40 50 60 70 80
DIAS%
aci
dez
26
Fig. 4.
CEREZO
0
1
2
3
4
5
6
7
0 50 100 150 200 250 300
DIAS
% a
cide
z
ACACIA
0
1
2
3
4
5
6
7
0 50 100 150 200 250 300
DIAS
% a
cide
z
ROBLE
0
1
2
3
4
5
6
7
8
0 50 100 150 200 250
DIAS
% a
cide
z
CASTAÑO
0
1
2
3
4
5
6
7
0 50 100 150 200 250 300
DIAS
% a
cide
z
27
Fig. 5.
0
1
2
3
4
5
6
7
8
0 50 100 150 200 250 300
Tiempo (dias)
% a
cide
z
28
Fig. 6.
ROBLE
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
VFI VFM VFE
1 1
A)
Pobl
ació
n
ACACIA
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
VFI VFM VFE
2 4
B)
Pobl
ació
n
CEREZO
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
VFI VFM VFE
5
1
C)
Pobl
ació
n
CASTAÑO
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
VFI VFM VFE
64 2
D)
Pobl
ació
n
29
Fig. 7.
ROBLE
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
VTI VTM VTE
2
2
Pobl
ació
n
ACACIA
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
VTI VTM VTE
34
Pobl
ació
n
CEREZO
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
VTI VTM VTE
1
Pobl
ació
n
CASTAÑO
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
VTI VTM VTE
41
Pobl
ació
n
30
LEYENDA DE TABLAS
Tabla 1. Análisis físico-químicos iniciales realizados en las dos vinagrerías
Azúcares residuales (g/l) SO2 (mg/l) Muestras % acidez Glucosa Fructosa % etanol pH SO2 libre SO2 total Mas d’en Gil: Vino Madre T0
0.45 3.30 0.90
0.34 0.46 0.35
0.75 0.92 0.78
13.30 4.20 11.34
3.22 3.08 3.26
9.0 --- 6.0
18.0 ---
12.0 La Guinelle: Vino Madre T0
0.60 6.00 0.90
27.21 -----
20.90
57.90 -----
43.38
14.86 2.56 9.62
3.56 2.64 3.37
13.0 ---- 5.0
35.0 ---- 9.0
31
Tabla 2. Resultados de la pérdida de etanol a lo largo de las acetificaciones en la
vinagrería La Guinelle.
Madera Grosor Puntos
muestreo % acidez % etanol ∆%1 acidez
∆%2 etanol
% pérdida de etanol
T0 0,9 9,6 15 T3 6,3 3,4 5,4 6,2 0,8 20 T3 6,6 4,1 5,7 5,5 -0,2
Roble
25 T3 7,2 1,4 6,3 8,2 1,9 T0 0,9 9,6
15 T3 6,3 3,9 5,4 5,7 0,3 20 T3 6,0 3,6 5,1 6,0 0,9
Acacia
25 T3 6,0 3,3 5,1 6,3 1,2 T0 0,9 9,6
15 T3 6,3 4,2 5,4 5,4 0,0 20 T3 6,6 3,6 5,7 6,0 0,3 Cerezo
25 T3 6,6 2,7 5,7 6,9 1,2 T0 0,9 9,6
15 T3 4,8 6,2 3,9 3,4 -0,5 20 T3 6,9 3,3 6,0 6,3 0,3 Castaño
25 T3 6,3 2,0 5,4 7,6 2,2
1 ∆% acidez = Diferencia de acidez entre T3 y T0 2 ∆% etanol = Diferencia de etanol entre T3 y T0
32
Tabla 3. Resultados de la pérdida de etanol a lo largo de las acetificaciones en la
vinagrería Mas d’En Gil
Madera Grosor Puntos muestreo % acidez % etanol ∆%1
acidez ∆%2
etanol % pérdida de etanol
T0 0,9 11,4 15 T3 6,0 4,3 5,1 7,1 2,0 20 T3 6,8 3,5 5,9 7,9 2,0
Roble
25 T3 6,3 4,3 5,4 7,1 1,7 T0 0,9 11,4
15 T3 6,0 4,7 5,1 6,7 1,6 20 T3 6,3 4,0 5,4 7,4 2,0
Acacia
25 T3 6,0 4,9 5,1 6,5 1,4 T0 0,9 11,4
15 T3 6,3 5,7 5,4 5,7 0,3 20 T3 6,0 5,4 5,1 6,0 0,9 Cerezo
25 T3 5,7 5,4 4,8 6,0 1,2 T0 0,9 11,4
15 T3 6,0 5,1 5,1 6,3 1,2 20 T3 5,7 5,8 4,8 5,6 0,8 Castaño
25 T3 6,3 4,8 5,4 6,6 1,2
1 ∆% acidez = Diferencia de acidez entre T3 y T0 2 ∆% etanol = Diferencia de etanol entre T3 y T0
33
Tabla 4. Diferencias en la velocidad de acetificación entre barricas de diferente madera
(mg l-1 día-1)
Tipo de madera Priorat Banyuls
Acacia 53 ± 6 1 127 ± 25
Castaño 55 ± 5 97 ± 15
Cerezo 55 ± 5 97 ± 20
Roble 61 ± 4 89 ± 12
1 Media de los tres grosores ± desviación estándar

34
Tabla 5. Identificación a nivel de especie y diversidad de cepas en las diferentes
maderas a lo largo de las acetificaciones en la vinagrería La Guinelle.
MUESTRAS Nº COLONIAS % ESPECIES DIVERSIDAD DE CEPAS
T1 29 100% A. pasteurianus 10% (3)1
T2 27 100% A. pasteurianus 11% (3) 67% A. pasteurianus
ROBLE
T3 30 33% Ga. europaeus
7% (2)
T1 21 100% A. pasteurianus 14% (3)
T2 21 100% A. pasteurianus 43% (9) ACACIA
T3 19 100% A. pasteurianus 5% (1) T1 15 100% A. pasteurianus 53% (8)
T2 22 100% A. pasteurianus 9% (2) CEREZO
T3 26 100% A. pasteurianus 8% (2)
T1 29 100% A. pasteurianus 35% (10)
T2 24 100% A. pasteurianus 29% (7) CASTAÑO
T3 16 100% A. pasteurianus 19% (3)
1 Número de cepas diferentes identificadas

35
Tabla 6. Identificación a nivel de especie y diversidad de cepas en las diferentes
maderas a lo largo de las acetificaciones en la vinagrería Mas d’en Gil.
MUESTRAS Nº COLONIAS % ESPECIES DIVERSIDAD DE CEPAS T1 25 100% A. pasteurianus 12% (3)1
T2 29 100% A. pasteurianus 31% (9) ROBLE
T3 4 100% A. pasteurianus 50% (2)
T1 26 100% A. pasteurianus 31% (8)
T2 21 100% A. pasteurianus 29% (6) ACACIA
T3 22 100% A. pasteurianus 5% (1)
T1 28 100% A. pasteurianus 7% (2) T2 21 100% A. pasteurianus 5% (1) CEREZO T3 20 100% A. pasteurianus 5% (1)
T1 24 100% A. pasteurianus 42% (10)
T2 19 100% A. pasteurianus 16% (3) CASTAÑO
T3 12 100% A. pasteurianus 17% (2)
1 Número de cepas diferentes identificadas

36
Proyecto de Tesis para optar al título de Doctor en Enología
Carlos A. Vegas
Universitat Rovira i Virgili
Directores: Montse Poblet y Maria Jesús Torija
Título provisional:
Estudio de las poblaciones de bacterias acéticas implicadas en la elaboración de
vinagres artesanales e industriales: Análisis de la capacidad oxidativa de cepas
seleccionadas
Observaciones previas:
En el mundo de producción de vinagres el inicio de las acetificaciones se suele realizar
mediante la adición del vino a vinagres ya en proceso de acetificación o bien mediante
el mantenimiento de madres de vinagre. No obstante, dichos procesos se realizan de
forma poco controlada y sin verdaderos cultivos iniciadores, ya que tanto los vinagres
como las madres corresponden a cultivos mixtos. En dichas condiciones es difícil
controlar la calidad final del producto, sometida a variaciones en el proceso
fermentativo debido a la imposición de diferentes poblaciones microbianas durante el
proceso de acetificación.
Actualmente, pocos estudios se han realizado sobre bacterias acéticas en la producción
del vinagre de vino, así tenemos: en vinagres industriales por cultivo sumergido
(Sokollek et al., 1998; Schüller et al., 2000) y el vinagre Tradicionale de Módena por
cultivo superficial (De Vero et al., 2004); por lo que el estudio de la población y

37
ecología presente en el vinagre y su posterior selección de bacterias acéticas como
cultivos iniciadores permitiría un mayor control del proceso de acetificación ante
posibles contaminaciones por otros microorganismos y obtener un producto de una
calidad más uniforme.
Hipótesis:
La selección de cepas de bacterias acéticas autóctonas permite un mejor control del
proceso de acetificación y, como resultado, del mantenimiento de características
organolépticas del producto final típica de la zona. Además favorece a un inicio más
rápido de la acetificación (reducción de la fase de latencia) contribuyendo así, la
disminución del tiempo de acetificación.
Objetivo general:
Analizar las poblaciones de bacterias acéticas obtenidas en diferentes vinagres,
identificarlas, seleccionarlas y probar su potencial biotecnológico para la producción del
vinagre.
Objetivos específicos:
- Identificar las diferentes cepas y especies presentes en vinagres de diferentes
acetificaciones (vinagres procedentes del método por cultivo superficial y sumergido).
- Analizar la capacidad oxidativa de dichas cepas realizando acetificaciones en
laboratorio (análisis de acidez, etanol, producción de ácido glucónico, pH, etc.)
- Selección de cepas para su posterior utilización como cultivos iniciadores realizando
acetificaciones en vinagrerías empleando barricas de diferentes maderas, volúmenes,
formas, etc.

38
- Establecer las condiciones óptimas de almacenamiento a corto y largo plazo mediante
análisis de los nutrientes requeridos por las bacterias acéticas para su recuperación en
medios de cultivos que aseguren el mantenimiento de la capacidad oxidativa.
- Estudiar los mecanismos que permitan una rápida recuperación y producción de
biomasa.
Metodología y Plan de trabajo
1. Identificación de diferentes cepas y especies aisladas de vinagres obtenidos en
vinagrerías comerciales y experimentales. Se utilizarán técnicas de biología
molecular ya desarrolladas en el grupo de investigación (RFLP-PCR 16S, (GTG)5-
PCR, ERIC-PCR, RT-PCR). Primer semestre de 2008 y a lo largo del período
mientras haya nuevas muestras.
2. Análisis de la capacidad oxidativa de las cepas mediante seguimiento rápido de
producción de acético en medios con etanol. Se analizará la velocidad de producción
de ácido acético así como su relación con la expresión génica de genes directamente
implicados en el proceso de acetificación (expresión génica de la alcohol
deshidrogenasa) y/o crecimiento en medios con etanol como fuente de carbono.
Segundo semestre de 2008.
3. Análisis de los medios de mantenimiento de cepas de bacterias acéticas: se
compararán diferentes medios y condiciones de mantenimiento y la pérdida de
capacidad oxidativa en cada medio. Se realizará en paralelo al punto anterior y se
complementará con éste. Segundo semestre de 2008
4. Optimización de sistemas de aislamiento, mantenimiento y recuperación de cepas de
bacterias acéticas. Como conclusión de los puntos anteriores se propondrán sistemas

39
adecuados para el mantenimiento y recuperación de bacterias acéticas y los métodos
de seguimiento adecuados. 2009.
Bibliografía
De Vero, L., Gullo, M., Solieri, L., Landi, S. & Giudici, P. (2004). A new approach
to study the acetic acid bacteria: the DGGE. P. 27-37. In I microorganismi dell’aceto
balsamico. Modena. Italy.
Sokollek, S. J., Hertel, C. & Hammes, W. P. (1998). Description of Acetobacter
oboediens sp. nov. and Acetobacter pomorum sp. nov., two new species isolated from
industrial vinegar fermentations. Int J Syst Bacteriol 48, 935-940.
Schüller G., Hertel C., Hammes W.P. (2000). Gluconacetobacter entanii sp. nov.,
isolated from submerged high-acid industrial vinegar fermentations. Int. J. Syst. Evol.
Microbiol., 50, 2013-2020.