“estima de la abundancia y distribución de la tarabilla … · ... (saxicola dacotiae) en la...
TRANSCRIPT

Tesis de Maestría Alexandra Kouri
“Estima de la abundancia y distribución de la
tarabilla canaria (Saxicola dacotiae) en la isla
de Fuerteventura (Islas Canarias)”
Profesor responsable: Javier Seoane Pinilla
Master en Ecología 2007/2009

ÍNDICE
1. INTRODUCCION .......................................................................................... 1
2. MATERIAL Y MÉTODOS ............................................................................... 5
2.1. ÁREA DE ESTUDIO ..................................................................................... 5
2.2. TRABAJO DE CAMPO................................................................................... 6
2.3. ESTIMA DE LA POBLACIÓN Y SU DISTRIBUCIÓN .............................................. 7
2.3.1. Estratificación ................................................................................ 7
2.3.2. Abundancia y Densidad.................................................................. 8
2.3.3. Construcción y Evaluación del mapa de Distribución .................. 11
3. RESULTADOS............................................................................................. 14
3.1. DETECTABILIDAD .................................................................................... 14
3.2. ABUNDANCIA Y DENSIDAD........................................................................ 15
3.3. DISTRIBUCIÓN Y VALIDACIÓN .................................................................. 17
3.4. NIVEL DE PROTECCIÓN DEL ÁREA DE DISTRIBUCIÓN DE LA ESPECIE................. 20
4. DISCUSION................................................................................................ 22
4.1. ABUNDANCIA Y DENSIDAD........................................................................ 22
4.2. DISTRIBUCIÓN Y VALIDACIÓN .................................................................. 23
4.3. ESTADO DE CONSERVACIÓN ...................................................................... 24
5. CONCLUSIONES......................................................................................... 27
ANEXO I. CARTOGRAFICO…………………………………………………………………I
ANEXO II. TABLAS…………………………………………………………………………IV
ANEXO I. METODOLOGIA GIS…….……………………………………………………..V

1. INTRODUCCION El conocimiento de la distribución y abundancia de una especie es un
requerimiento básico para muchos estudios ecológicos e imprescindible a la hora de
evaluar el estado de conservación de la especie o desarrollar un plan de gestión.
Sin embargo, es difícil realizar estudios de la abundancia y distribución de las
especies a escala regional y además, los resultados de censos a esa escala raras
veces son analizados en el ámbito de la conservación (Seoane et al., 2003).
Estas estimas de abundancia deberían tener bajo sesgo (menor sobre- o
subestimación sistemática) y alta precisión (bajo nivel de incertidumbre). Sin
embargo, en la práctica eso puede ser difícil y caro de conseguir. Como
consecuencia, se utiliza como índice de abundancia el número de individuos
observados durante un trabajo de campo estandarizado, con el argumento que las
estimas de abundancia absoluta no son necesarias. Pero, en el caso que la
inferencia sobre tendencias o diferencias en la abundancia es imprecisa puede que
los recursos de la conservación acaben mal aplicados. Las dos causas de
imprecisión más frecuentes son (i) que la proporción de individuos presentes en el
área de estudio que han sido detectados varía en el tiempo y el espacio, es decir la
detectabilidad es variable y (ii) que los muestreos realizados no son representativos
del área total de estudio a la que se extrapolan las conclusiones (Buckland et al.,
2008).
La disminución durante el muestreo de campo de la incertidumbre
provocada por la variabilidad de la detectabilidad de las especies es difícil de
conseguir. Por lo tanto, se han desarrollado métodos alternativos de campo, como
el método del “doble observador” (Nichols et al., 2000) y el método de “distance
sampling” propuesto por Buckland et al. (2001), que proporcionan una estima de la
detectabilidad a posteriori.
Aunque la significación de la variabilidad en la detectabilidad ha sido
señalada en muchos estudios durante las últimas dos décadas o más (p.ej. Bibby &
Buckland, 1987; Nichols et al., 2000), sigue siendo ampliamente aceptado que los
índices de abundancia relativa son comparables entre distintos estudios de una
especie considerando como el único control necesario de la variabilidad, la
estandarización de la metodología. Los métodos alternativos, entre ellos “distance
sampling”, se utilizan con menos frecuencia a causa de la percepción que son
métodos más costosos en tiempo y esfuerzo (Norvell et al., 2003). La abundancia
relativa es muy popular y se considera comparable entre estudios basándose en la
supuesta insignificancia de la variabilidad en detectabilidad, a pesar de las
evidencias de lo contrario. Según el estudio de comparación entre el método
1

tradicional de estima de abundancia relativa y el método “distance sampling”
realizado por Norvell et al. (2003), las comparaciones entre distintos estudios o aún
en el mismo estudio utilizando valores de abundancia relativa se basan en una
suposición/condición poco fundada. En estos estudios de abundancia relativa se
acumula información que no es comparable entre hábitats, especies o momentos
temporales debido a la variabilidad de la detectabilidad. Mientras tanto, las
robustas estimas derivadas de “distance sampling” durante ese estudio justifican la
creciente necesidad de modelar las probabilidades de detección.
La distribución de una especie puede estudiarse con varios métodos basados
en modelos predictivos o directamente en datos de campo. El más utilizado para
cartografiar la distribución de una especie consiste en la generación de modelos
predictivos (Guisan & Zimmerman, 2000; Osborne, 2001; Brotons et al., 2004;
Seoane et al., 2003, 2006) basados en datos de presencia-ausencia de la especie.
A la hora de generar estos modelos hay dos enfoques básicos para conseguir el
mejor uso de los datos disponibles. El método de interpolación espacial, oscilando
entre la interpolación linear simple y el “kriging”, estima la presencia de una
especie entre puntos de muestreo basado en su distribución espacial (Osborne,
2001). El enfoque alternativo relaciona la presencia de una especie en los puntos
con unas variables disponibles para toda el área de estudio (Osborne & Tigar, 1992;
Buckland & Elston, 1993). Para la aplicación de este último se necesitan datos
ambientales de todo el área de distribución de la especie (Osborne, 2001; Conlisk
et al., 2009).
La distribución de especies se predice basándose en parámetros ecológicos
que se consideran como la fuerza motriz de su distribución y abundancia. Factores
ecológicos como estos están generalmente extraídos por mapas digitales con el uso
de sistemas de información geográfica (SIG), ya que normalmente es difícil o
costoso medirlos. Los Modelos Digitales de Elevación (DEM) son una herramienta
muy exacta para conseguir esto, como constituyen la base para la generación de
nuevos mapas con variables ambientales (Guisan & Zimmermann, 2000).
El método de estima de la abundancia y distribución que se utilizó en el
presente estudio consiste de un modelo de campo basado únicamente en el diseño
de muestreo realizado y la cartografía existente. Los datos de campo se
recolectaron mediante transectos lineales, procurando evitar las dos fuentes de
imprecisión más frecuentes, que son la variabilidad en detectabilidad (que se
controló utilizando el método de “distance sampling”) y la realización de un
muestreo representativo de todo el área (Bibby et al., 2000; Buckland et al.,
2001); y estratificando posteriormente por los distintos hábitat presentes en la isla,
con el uso de la cartografía digital disponible. El método utilizado consiste en la
2

estima de la abundancia de la especie utilizando los datos de censos y calculando
mediante el método de “distance sampling” y el software Distance 5.0 (Thomas et
al, 2006) los valores absolutos de densidad y abundancia.
Mediante el uso de la cartografía digital se pueden ilustrar los resultados de
manera más eficaz y estimar la distribución geográfica de la especie. La cartografía
digital, las imágenes de satélite y el uso de sistemas de información geográfica
(SIG) pueden resultar herramientas útiles para la modelización geográfica y la
aplicación de estos estudios en el ámbito de la conservación (Osborne et al., 2001;
Guisan and Zimmermann, 2000; Seoane et al., 2003), así como a la evaluación de
planes y estrategias de conservación ya existentes.
La tarabilla canaria (Saxicola dacotiae) es una especie de ave paseriforme
endémica de Fuerteventura (Imagen 1a.) que está catalogada como especie “En
Peligro” (EN) por la UICN (UICN, 2007) y como “Vulnerable” en el Catálogo
Nacional de Especies Amenazadas (Illera, 2004). Se trata de una especie muy
territorial que está principalmente amenazada por la alteración o destrucción de su
hábitat debidas a la rápida expansión urbanística que se está produciendo en la isla
de Fuerteventura (Illera, 2004). Su población fue estimada en 1.300-1.700
individuos (750 ± 100 parejas) a finales de los años ochenta (Bibby & Hill, 1987).
Un estudio más reciente estima la abundancia de la tarabilla canaria en 1.035
individuos (832-1.287) (García del Rey, 2009). Por otro lado, en la revisión de
Áreas Importantes para las Aves (IBA), publicada por SEO/Birdlife en 1998, se
identificaron nueve áreas de especial importancia para esta especie (Viada, 1998;
Illera, 2004). Sin embargo, no se ha realizado aún un análisis que compruebe la
congruencia de estas IBAs con los núcleos poblacionales más importantes ni con la
red de espacios protegidos de las islas Canarias.
Imagen 1. (a)Individuo joven de tarabilla canaria, (b) el barranco de la Torre. Zonarodeada por hábitat adecuado para la especie y su nidificación debido a la presenciade matorral, la pendiente empinada y la accesibilidad al agua.
3

La distribución de la tarabilla canaria en Fuerteventura se explica por una
combinación de características tanto a nivel de microhabitat (en un radio de 25m)
como de paisaje (radio de 1Km.). A nivel de paisaje selecciona medios terroso-
pedregosos y barrancos, con pendientes que presentan una alta (>15%) cobertura
arbustiva de medio y gran porte (>0,25m de altura). A una escala de microhábitat,
prefiere zonas de pendiente de hábitat terroso-pedregosos y barrancos, con
presencia de piedras grandes (>0,25m de altura) y pocas piedras de menos de
0,25m de altura (Illera, 2001). Según García del Rey (2009) la especie parece ser
más abundante en laderas empinadas que en barrancos. Estos medios no sólo
mantienen las abundancias de invertebrados más altas, sino que además ofrecen
lugares para ubicar sus nidos y abundantes posaderos desde donde buscar
artrópodos de los que se alimenta (Imágenes 1b y 2). Los adultos muestran alta
fidelidad por el territorio durante todo el año. Ubican sus nidos en el suelo, debajo
de rocas y arbustos, o también, en el interior de grietas en muros, por lo que es
muy vulnerable a depredadores introducidos, fundamentalmente gatos y ratas
(Illera, 2003). La alta fidelidad al territorio reportada para esta especie se
interpreta como una adaptación a la baja variabilidad espacial de los factores
(bióticos y abióticos) en la isla de Fuerteventura (Illera & Díaz, en revisión en Illera,
2007). Su fenología reproductora está estrechamente relacionada con la lluvias
otoñales e invernales y con la disponibilidad de invertebrados relacionada con estas
(Illera & Díaz, 2006).
El principal objetivo del presente estudio es obtener una estima del tamaño
poblacional de la tarabilla canaria y su distribución en Fuerteventura para contribuir
Imagen 2. Punta de la Entallada. Paisaje típico de la isla de Fuerteventura que constituyeun hábitat idóneo para la especie
4

a la evaluación de su estado de conservación. Con este objetivo, se elaboró un
mapa de distribución de la especie, a una resolución espacial fina, en función de la
densidad que alcanza en sus hábitats óptimos. Este mapa permite, por un lado,
obtener una estima de su tamaño poblacional (mediante la agregación de las
estimas por superficie) y, por otro, evaluar el nivel de solapamiento de sus
principales zonas de concentración y las zonas protegidas (Espacios Naturales
Protegidos y Red Natura 2000). Estos dos aspectos servirán para evaluar el estado
de conservación de la tarabilla canaria.
La red Canaria de Espacios Naturales
protegidos incluye en la isla de
Fuerteventura los Parques Naturales de
Jandia y de Corralejo (Fig.1, zonas de color
amarillo), el Parque Rural de Betancuria
(Fig.1, zona de color rojo), varios
Monumentos Naturales (Fig.1, zonas de
color azul oscuro) y los Paisajes Protegidos
de Malpaís Grande y de Vallebrón (Fig.1,
zonas de color azul claro). Los dos Parques
Naturales, el Parque Rural de Betancuria y el
Paisaje Protegido de Malpaís Grande han
sido también declarados como Zonas de
Especial Protección para las Aves (ZEPA)
según lo que establece la Directiva
79/409/CEE relativa a la Conservación de
las Aves Silvestres (Gobierno de Canarias,
2008).
Imagen 3. La red de Espacios Naturales Protegidos en la isla de Fuerteventura. En color amarillo se ilustran los Parques Naturales, en rojo el Parque rural, en azul oscuro los Monumentos Naturales y en azul claro los Paisajes Protegidos (fuente: Gobierno de Canarias, 2008)
2. MATERIAL Y MÉTODOS 2.1. Área de estudio
El archipiélago de las islas Canarias pertenece a la región biogeográfica de
las islas del Atlántico, también llamada Macaronesia. Está formado por siete islas
volcánicas localizadas en el noreste del océano atlántico, a 100km al oeste de África
continental. El clima de las islas está determinado por la proximidad de ellos a las
altas presiones atmosféricas de las Azores y al desierto de Sahara, la corriente fría
de Canarias, y la diferente topografía de cada isla. Fuerteventura (28º46’Ν,
14º31’W) es la segunda más grande de las islas Canarias (1655 km2) y la más
próxima a África continental. Su topografía es principalmente baja y llana,
5

alcanzando la máxima elevación a los 807m.s.n.m. Las condiciones climáticas de la
isla (áridas y semiáridas) reflejan la escasez de agua. La precipitación promedia es
de 143mm/año, concentrada durante los meses de otoño / invierno, mientras que
la temperatura mensual varía entre 19ºC en Enero y 21ºC en Agosto. La vegetación
es xerofítica y está dominada por pastizal ralo y matorral (Illera & Díaz, 2006)
Imagen 4. Área de estudio. Los puntos negros separan la localización del punto central de cada transecto
2.2. Trabajo de campo
El presente estudio usará los muestreos realizados a lo largo de la isla
durante los periodos reproductores de los años 2005 y 2006, que ya han sido
analizados para estudiar el tamaño poblacional y la selección de hábitat de otras
aves de las islas canarias orientales (Carrascal et al., 2006, 2008; Palomino et al.,
2008; Fig. 2). Los muestreos se realizaron mediante 1471 transectos lineares de
500m, sin limite de banda, dispuestos aleatoriamente y recorridos a pie campo a
través, a una velocidad entre 2 y 3 Km/h. Se anotaron todos los contactos que se
registraron (oídos o vistos), el número de individuos por contacto y la distancia
perpendicular al recorrido. Los recorridos se emplazaron procurando cubrir (i) toda
la variedad de medios en relación con la vegetación, la pendiente y el tipo de suelo,
y (ii) toda la isla. Puesto que se pretendía cuantificar la población reproductora, se
descartaron de los análisis los escasos individuos juveniles que se encontraron.
6

2.3. Estima de la población y su distribución
La estima de la abundancia y distribución de la tarabilla canaria se realizó
mediante una inferencia basada en muestreos de campo. Por lo tanto se basó en
análisis del muestreo para la extrapolación de la información recogida en el campo
a toda la superficie de la isla. Para alcanzar este objetivo teniendo en cuenta la
variabilidad de condiciones en toda la isla, se decidió estratificar la superficie por
los distintos hábitat presentes según las preferencias de la especie, lo cual facilitó la
extrapolación de la información y la elaboración del mapa final de distribución de la
especie, además de la evaluación del resultado mediante un método de validación
del mapa final.
2.3.1. Estratificación
La estratificación de la isla se realizó con la elaboración de un mapa de
hábitat. Con el término hábitat se describe la combinación de categorías de
estructura de vegetación y de pendiente, previamente definidas e ilustradas en dos
mapas distintos. Se combinó la información de la estructura de vegetación con la
de pendiente porque estas parecen ser las variables que condicionan a la especie
durante la estación reproductora en el proceso de selección de hábitat (Illera,
2001), además de la granulometría del terreno que no se incorporó en el mapa por
falta de cartografía relacionada.
El mapa de vegetación actual de Fuerteventura se elaboró con el programa
ArcGis 9.0 en formato vectorial utilizando como fuente principal el mapa de
vegetación canaria (Del Arco et al., 2003) de escala 1:25.000 (metodología
detallada en AnexoIII, apartado 1.) y como método de control del resultado la
ortofotografía de la isla de Fuerteventura. El mapa base contiene información de las
comunidades vegetales, definidas por su composición florística. Utilizando esta
información proporcionada por el mapa de vegetación canaria y según el
conocimiento propio de campo se identificaron nueve categorías de vegetación
según su estructura: matorral alto, matorral medio, matorral bajo, bosque, medio
rural, pastizal, acuática, sin vegetación y se ignora (se refiere a la isla de Lobos)
(AnexoI Mapa 1.). A continuación el mapa se convirtió en formato raster para hacer
posible su combinación con el mapa de pendientes.
El mapa de pendientes se elaboró con el programa IDRISI Kilimanjaro a
partir de un modelo digital de elevación (DEM) de 90m de resolución espacial
(metodología detallada en Anexo III, apartado 2.). A la hora de decidir umbrales de
pendiente para la clasificación del mapa en distintas categorías se consideró
oportuno utilizar umbrales con sentido ecológico por conocerse que son limitantes
7

para otras especies. Por lo tanto, la reclasificación del mapa se hizo basándose en
umbrales de pendiente que son limitantes para especies corredoras y sensibles a
pendientes muy pronunciadas como el corredor sahariano (Cursorius cursor) y la
alondra de dupont (Chersophilus duponti); y después de explorar el número de
transectos realizados en cada combinación de las categorías de pendiente con las
categorías de vegetación para establecer la presencia de suficiente número de
transectos en cada una de las combinaciones (Anexo II., tabla 1.). Las
probabilidades de ocurrencia del corredor sahariano (Cursorius cursor) es mayor en
áreas relativamente llanas soportando como máximo un valor de pendiente del
11% (Palomino et al., 2007), mientras que la alondra de Dupont (Chersophilus
duponti) evita las áreas con pendiente mayor que 5,1% (Seoane et al., 2006). Por
lo tanto, el mapa de pendientes se reclasificó en tres categorías: ≤5%, 6%-11%,
≥12%).
Finalmente, el mapa de hábitat se elaboró combinando los dos mapas
anteriormente mencionados y reclasificándolo (herramientas utilizadas en AnexoIII,
apartado 3.). La reclasificación se hizo manteniendo las categorías de vegetación
“medio rural”, “acuática” y “se ignora” independientes de la pendiente debido a la
presencia limitada o falta absoluta de transectos de muestreo en ellas, además de
la pequeña superficie de este tipo de vegetación en el caso de la categoría
“acuática” y la ausencia de la especie en el caso de la isla de Lobos (categoría de
vegetación “se ignora”). Las categorías de vegetación “pastizal” y “sin vegetar” se
dividieron en dos subcategorías en vez de tres según la pendiente para mantener el
número de transectos por categoría por encima de los 10 (Anexo II, tabla 1.). Así el
mapa final de hábitat consiste de 19 categorías de distintas combinaciones de
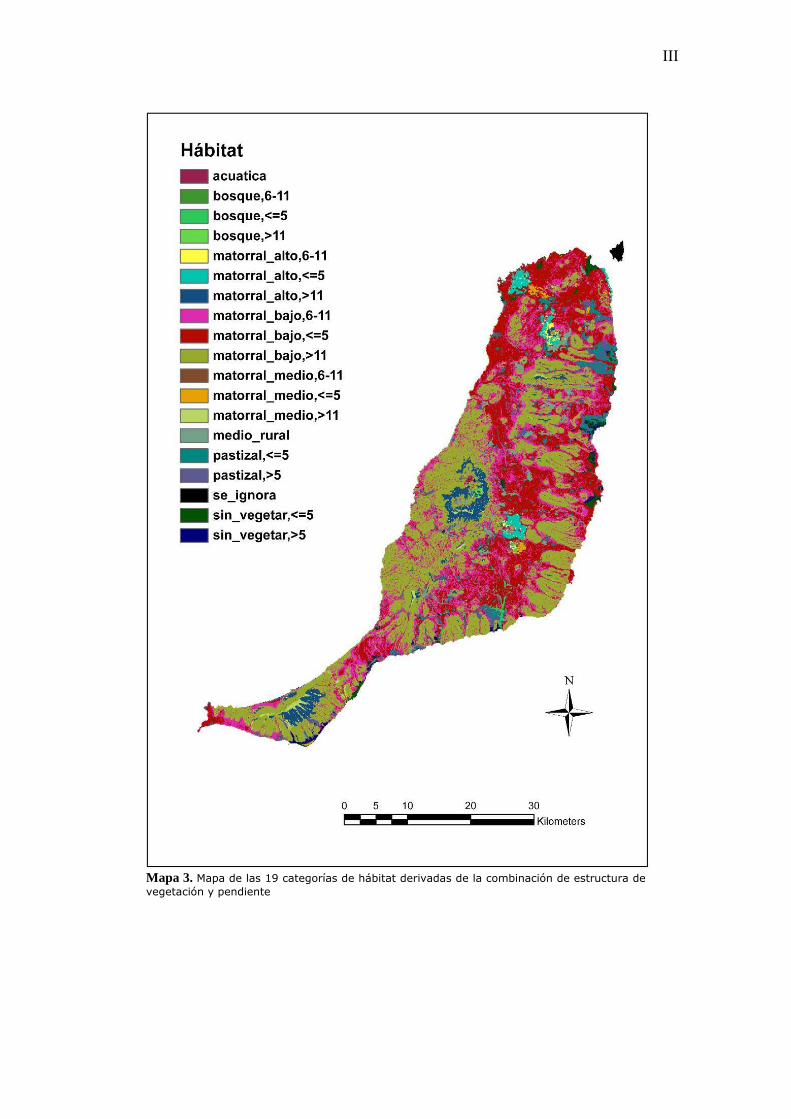
vegetación y pendiente (AnexoI, Mapa 3.).
2.3.2. Abundancia y Densidad
Teniendo el mapa de hábitat de Fuerteventura se pudo proseguir con la
estima de abundancia de la tarabilla canaria y la elaboración de un mapa de
distribución. Para conseguir esto fue necesaria la estima de la detectabilidad de la
especie en cada uno de los hábitat determinados mediante el método de “distance
sampling” (muestreo con distancias o análisis de distancias de encuentro, Buckland
et al. 2004) utilizando el software gratuito Distance 5.0 (Thomas et al., 2006) para
poder extrapolar los valores obtenidos en el campo a toda la superficie ocupada por
cada hábitat.
El “distance sampling” es un grupo de métodos ampliamente utilizados para
estimar la densidad y/o la abundancia de poblaciones biológicas. Los métodos
principales son: transectos lineales y transectos puntuales. A la hora de realizar un
8

transecto no se detectan todos los individuos de la especie en cuestión en el área
de muestreo, pero la suposición fundamental de estos métodos es que se detectan
todos los individuos presentes en ese momento en la línea del transecto y que
cuanto más lejos están los individuos de la línea del transecto más difícil es
detectarlos, resultando en un menor número de detecciones a mayores distancias
del trayecto. La clave en el análisis de “distance sampling” es ajustar una función
de detección a las distancias observadas, que describe la pérdida de detectabilidad
con la distancia de muestreo al observador, y usar esta función para estimar la
proporción de individuos que no han sido detectados durante el muestreo (Thomas
et al., 2002). La diferencia entre este método y el método tradicional de muestreo
en transectos lineales de banda de ancho fijo es que en el segundo (i) se considera
que en el espacio delimitado por las bandas se detectan todos los individuos algo
que en muchos casos no es cierto, dependiendo principalmente de la especie
estudiada; y (ii) que los individuos que se detectan fuera de esta banda están
ignorados. Si el ancho de banda es lo suficientemente pequeño como para cumplir
con la suposición de que se detectan todos los individuos, es probable que gran
parte de los individuos presentes quede fuera del área determinado por las bandas
y por tanto ignorado; y además el área cubierta por cada transecto disminuye y por
lo tanto se reduce el área total censada (Thomas et al., 2002). El método de
“distance sampling” no considera que todos los individuos dentro de un área
previamente determinada estén detectados sino estima la probabilidad de detección
dentro del área de muestreo.
Teniendo los valores de la distancia perpendicular (x) de cada uno de los
individuos observados podemos establecer un valor w (distancia de truncamiento o
“truncation distance”) de distancia a la línea del transecto a partir de la cual
ignoramos las observaciones considerándolas poco fiables, pues las estimas de
distancia para contactos lejanos se hacen con menos precisión (Buckland et al.,
2008; Thomas et al., 2002). Si todos los individuos en un transecto de ancho de
banda w son detectados, la densidad será igual que el número de individuos
observados (n) dividido por el área total estudiada a=2wL, donde L es la longitud
total de los transectos realizados y multiplicar por dos da cuenta de que la
observación se hace a ambos lados del recorrido. Si Pa es la probabilidad de que un
individuo elegido al azar dentro del área muestreada sea detectado entonces la
densidad sería igual a:
9

Para estimar Pa se define una función de detección f(x) que es la probabilidad de
que un individuo a distancia x del transecto sea detectado, con 0≤x≤w, asumiendo
que f(0)=1. Si las distancias perpendiculares medidas se representan gráficamente
en un histograma se puede establecer un parámetro µ o ESW, como se referirá a
este a partir de ahora, que es el ancho de banda efectivo (“effective strip width”),
lo cual es la distancia a la línea del transecto a partir de la cual se han detectado
tantos individuos como los que se han dejado de detectar dentro del área que
define (Graf.1, Thomas et al., 2002).
El área definido por µ
(ESW) para la función de
detectabilidad f(x) cuando es
expresado como una
proporción del área definido
por w para f(0)=1 es la
probabilidad de que un
individuo dentro del área de
estudio sea detectado
(Buckland et al., 2008,
Thomas et al., 2002).
Entonces Pa= µ/w y la
densidad es igual a :
Graf. 1. Representación gráfica de la función de la probabilidad de detección. El área definido por µ≤x≤w (sombreado debajo de la curva)es igual que el área sombreado encima de la curva que representa los individuos no detectados por 0≤x≤µ.El software Distance 5.0 realiza todos los cálculos anteriormente
mencionados según la información y el modelo introducidos.
La información que se introdujo en el Distance para cada uno de los 1471
transectos realizados consiste de: el hábitat y el área que ocupa en la isla de
Fuerteventura, el identificador de cada transecto y su longitud; y el número de
observaciones y su distancia perpendicular al transecto para cada contacto
establecido a lo largo de cada transecto (Buckland et al., 2001).
En principio se realizó un análisis exploratorio para establecer el valor w
adecuado utilizando un modelo seminormal (“Half-normal”), con términos de ajuste
de coseno (HNc). El mismo modelo se repitió incorporando información sobre el
volumen de la vegetación para cada transecto con valores de volumen y volumen
elevado al cubo para comprobar si influye en la probabilidad de detección de la
tarabilla canaria en Fuerteventura.
10

Los análisis finales se realizaron con w=80 y sin considerar el volumen de la
vegetación para cuatro familias de modelos que sugieren Buckland et al., 2001
como los más robustos para calcular la función de detectabilidad. Los modelos
utilizados son: seminormal (“Half-normal”) con terminos de ajuste de coseno
(HNc), seminormal (“Half-normal”) con términos de ajuste polinomiales (HNp),
exponencial negativo (“Negative exponential”) con terminos de ajuste de coseno
(NEc), exponencial negativo (“Negative exponential”) con términos de ajuste
polinomiales (NEp), uniforme (“Uniform”) con terminos de ajuste de coseno (UNc),
uniforme (“Uniform”) con términos de ajuste polinomiales (UNp), tasa de riesgo
(“Hazard-rate”) con terminos de ajuste de coseno (HRc) y tasa de riesgo (“Hazard-
rate”) con términos de ajuste polinomiales (HRp). En todos los modelos se utilizó el
análisis convencional (Conventional “distance sampling”, CDS); los términos de
ajuste se seleccionaron con el método secuencial, con criterio de selección el valor
de AIC y un máximo de 5 términos de ajuste. La estima se realizó estratificando
posteriormente (post-stratify) por la variable de hábitat; calculando la densidad
global y por estrato (hábitat), la tasa de encuentro por estrato (hábitat), la función
de detección global para cada modelo y el tamaño de cluster por estrato. La
densidad global (abundancia) se calculó como la media de las densidades de cada
estrato (hábitat) ponderando por el área de cada uno de ellos. Para calcular la
varianza de la abundancia total de la especie se incorporó en el análisis una estima
bootstrap no parametrica de 999 remuestreos dentro de cada estrato (hábitat). La
estima de la varianza de la densidad de cada estrato (hábitat) se calculó con el
método empírico.
Los valores de abundancia y densidad se calcularon con la media ponderada
de los mejores modelos según el criterio de información de Akaike corregido para
muestras pequeñas (AICc). Los modelos con ∆AICc>2 no se incluyeron en los
cálculos. El peso (b) de cada modelo en la ponderación se calculó utilizando la
función (Burnham & Anderson, 2002):
bi =exp (-1/2*∆i)
exp (-1/2*∆r)Σr=1
Rbi =exp (-1/2*∆i)
exp (-1/2*∆r)Σr=1
R
exp (-1/2*∆i)
exp (-1/2*∆r)Σr=1
R
exp (-1/2*∆r)Σr=1
R
Σr=1
R
donde: ∆i- ∆r es ∆AICc de i=1 a r
2.3.3. Construcción y Evaluación del mapa de Distribución
Después de haber alcanzado el primer objetivo del trabajo que es la estima
de la abundancia de la tarabilla canaria en Fuerteventura y teniendo calculado el
valor de densidad por hábitat (nº de aves/ha) se pudo cruzar con el mapa de
11

hábitat previamente elaborado y obtener un mapa de densidad por hábitat para el
conjunto de la isla.
Como la densidad se calculó teniendo en cuenta la totalidad de la superficie
ocupada por cada hábitat el mapa resulta tener una precisión alta, pero poco fiable
en casos de poligonos de hábitat de muy pequeña superficie. Por lo tanto, se
consideró que al transformar el mapa de densidad en un mapa de cuadriculas UTM
este efecto se suaviza, quedando mejor ilustrada la distribución de la especie.
Para crear el mapa de distribución de la especie se superpuso una malla de
cuadriculas UTM de 1Km2 al mapa de densidad por hábitat y se calculó la densidad
en cada cuadricula multiplicando el área que ocupe cada hábitat dentro de la
cuadricula con su respectivo valor de densidad. Se eligió la malla de 1Km2 y no de
0.5Km2 para conseguir cuadriculas mejor censadas, ya que con transectos de
0,5Km de longitud sería imposible tener más que un transecto en una cuadricula de
0,5Km2.
Para comprobar la fiabilidad del mapa de distribución se decidió efectuar una
evaluación del mismo utilizando un método de validación interna. La validación
consiste en la comparación de las densidades por Km2 calculadas con el método de
“distance sampling” con las observadas en el campo y se denomina interna porque
los valores comparados están calculados basándose en los mismos datos
principales. Como no se trata de una estima de densidad basada en modelos
predictivos sino en datos de campo no se pudo emplear el método óptimo que sería
excluir una serie de transectos del análisis y utilizarlos como control para evaluar
los resultados, porque este método comprometería la exactitud de los resultados.
Por lo tanto se eligió el método de validación interna que no es muy preciso pero
nos puede mostrar si hay grandes inconcordancias en la estima final y las
cuadriculas en las que se observan. El método de validación óptimo en este caso
sería la repetición del muestreo y la comparación de la estima con los nuevos datos
de campo. La comparación no se hizo con los valores del mapa sino que se repitió
el proceso para disminuir el efecto de la aleatoriedad.
La validación se realizó utilizando de nuevo el software Distance 5.0 esta vez
empleando un remuetreo mediante el método bootstrapping de 1000 repeticiones
para cada uno de los estratos (hábitat) para luego calcular 1000 valores de
densidad por Km2 y comparar el resultado con los datos de campo para poder
determinar su concordancia.
Los análisis se realizaron con los mismos modelos y características
previamente descritas para obtener finalmente 1000 valores de densidad para cada
uno de los 19 estratos (hábitat) de los cuatro modelos más significativos (según el
AICc global de los modelos como anteriormente utilizado). Se calculó la media
12

Dob =nob
2*L*ESWDob =
nob
2*L*ESW
nob
2*L*ESW
ponderada de los cuatro modelos para cada uno de los valores y se repitió la
metodología anteriormente descrita para obtener el valor de densidad por
cuadricula, resultando en 1000 valores de densidad para cada una de las
cuadriculas de la isla. Además, se calculó el valor de la densidad observada en cada
una de las cuadriculas basándose en el número de transectos realizados y el
número de observaciones en ellas. Por lo tanto, la densidad observada es igual a:
donde: nob es el número de aves observadas dentro de la cuadricula
L es la longitud total de los transectos realizados dentro de la cuadricula
y ESW es el ancho de banda efectivo calculado como la media ponderada de
los cuatro modelos
La comparación se realizó con una correlación de Pearson entre el valor de la
densidad observada extrapolada a la superficie de cuadricula (1Km2) y de la
densidad predicha para cada uno de los remuestreos en las cuadriculas que
contienen transectos. A continuación se agruparon dichas cuadriculas según el valor
de densidad predicha en siete categorías (0-5, 5-10, 10-20, 20-30, 30-40, 40-60,
>60). Se compararon cualitativamente la mediana de cada categoría con la
mediana de la densidad observada mediante un gráfico de cajas y bigotes. El
procedimiento se repitió para las cuadriculas que sólo contienen un número de
transectos mayor que dos, considerando que el valor de la densidad observada es
más fiable en ellas. Todo lo arriba mencionado se realizó con el programa S-Plus
8.0.
Finalmente, se comparó visualmente la distribución de la tarabilla canaria
con la superficie protegida de la isla superponiendo el mapa de las Zonas de
Especial Protección para las Aves (ZEPAs, EUROPARC-España, 2007) al mapa de
distribución creado.
13

3. RESULTADOS
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
0 10 20 30 40 50 60 70 800.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
0 10 20 30 40 50 60 70 800.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
0 10 20 30 40 50 60 70 80
Distancia perpendicular en metros
Prob
abili
dad
de d
etec
ción
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
0 10 20 30 40 50 60 70 800.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
0 10 20 30 40 50 60 70 800.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
0 10 20 30 40 50 60 70 80
Distancia perpendicular en metros
Prob
abili
dad
de d
etec
ción
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
0 10 20 30 40 50 60 70 800.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
0 10 20 30 40 50 60 70 800.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
0 10 20 30 40 50 60 70 80
Distancia perpendicular en metros
Prob
abili
dad
de d
etec
ción
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
0 20 40 60 80 100 120 1400.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
0 20 40 60 80 100 120 140
Distancia perpendicular en metros
Pro
babi
lidad
de
dete
cció
n
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
0 20 40 60 80 100 120 1400.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
0 20 40 60 80 100 120 140
Distancia perpendicular en metros
Pro
babi
lidad
de
dete
cció
n
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
0 20 40 60 80 100 120 1400.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
0 20 40 60 80 100 120 140
Distancia perpendicular en metros
Pro
babi
lidad
de
dete
cció
n
Graf. 2. La probabilidad de detección según el modelo HNc para todos los contactos observados(izquierda) y para los contactos después de emplear el filtro de w=80m (derecha).
3.1. Detectabilidad
Los resultados del primer análisis exploratorio con el programa Distance
indican que la distancia de truncamiento w=80m es una distancia al trayecto
adecuada por el escaso número de contactos que existen a partir de este punto
(Gráf.2). Buckland et al. (2001) recomiendan elegir la distancia de truncamiento
eliminando un 5-10% de valores de distancia extremos.
Los análisis realizados incluyendo los valores de volumen de vegetación y
volumen elevado al cubo indican que no existe diferencia en la función de
detectabilidad para distintos valores de volumen (Gráf. 3), por lo tanto no influyen
en la detectabilidad de la especie en el entorno estudiado. Esto se debe,
probablemente, a la homogeneidad del estado de vegetación en las partes de la isla
donde se han detectado individuos de la especie.
0
0.2
0.4
0.6
0.8
1
0 10 20 30 40 50 60 70 80
VOL=1.3,2.7,5.3
0
0.2
0.4
0.6
0.8
1
0 10 20 30 40 50 60 700
0.2
0.4
0.6
0.8
1
0 10 20 30 40 50 60 70 80
VOL=1.3,2.7,5.3
Distancia perpendicular en metros
Pro
babi
lidad
de
dete
cció
1.32.75.3
n
0
0.2
0.4
0.6
0.8
1
0 10 20 30 40 50 60 70 80
VOL=1.3,2.7,5.3
0
0.2
0.4
0.6
0.8
1
0 10 20 30 40 50 60 700
0.2
0.4
0.6
0.8
1
0 10 20 30 40 50 60 70 80
VOL=1.3,2.7,5.3
Distancia perpendicular en metros
Pro
babi
lidad
de
dete
cció
1.32.75.3
n
0
0.2
0.4
0.6
0.8
1
0 10 20 30 40 50 60 70 80
VOL=1.3,2.7,5.3
0
0.2
0.4
0.6
0.8
1
0 10 20 30 40 50 60 700
0.2
0.4
0.6
0.8
1
0 10 20 30 40 50 60 70 80
VOL=1.3,2.7,5.3
Distancia perpendicular en metros
Pro
babi
lidad
de
dete
cció
1.32.75.3
n
0
0.2
0.4
0.6
0.8
1
0 10 20 30 40 50 60 70 80
VOL=5.8,24.,74.
0
0.2
0.4
0.6
0.8
1
0 10 20 30 40 50 60 700
0.2
0.4
0.6
0.8
1
0 10 20 30 40 50 60 70 80
VOL=5.8,24.,74.
Distancia perpendicular en metros
Pro
babi
lidad
de
dete
cció
n
5.82474
0
0.2
0.4
0.6
0.8
1
0 10 20 30 40 50 60 70 80
VOL=5.8,24.,74.
0
0.2
0.4
0.6
0.8
1
0 10 20 30 40 50 60 700
0.2
0.4
0.6
0.8
1
0 10 20 30 40 50 60 70 80
VOL=5.8,24.,74.
Distancia perpendicular en metros
Pro
babi
lidad
de
dete
cció
n
5.82474
0
0.2
0.4
0.6
0.8
1
0 10 20 30 40 50 60 70 80
VOL=5.8,24.,74.
0
0.2
0.4
0.6
0.8
1
0 10 20 30 40 50 60 700
0.2
0.4
0.6
0.8
1
0 10 20 30 40 50 60 70 80
VOL=5.8,24.,74.
Distancia perpendicular en metros
Pro
babi
lidad
de
dete
cció
n
5.82474
Graf. 3. La probabilidad de detección calculada con covariable (i) el volumen de la vegetación(izquierda) con valores representados de 1.3, 2.7 y 5.3, (ii) el volumen de la vegetación elevado al cubo(derecha) con valores representados de 5.8, 24 y 74. En los dos casos se observa que la función novaría según el valor de volumen o volumen elevado al cubo.
14

Tabla 1. Los cuatro mejores modelos según el criterio deAICc y el peso (b) de cada modelo.
Modelo Delta AICc AICc bHNp80 0.00 3280.091 0.394
c80 .08 281.167 .230UNc80 1.47 3281.563 0.189NEp80 1.48 3281.569 0.188
NE 1 3 0
3.2. Abundancia y Densidad Del análisis definitivo destacaron cuatro modelos con ∆AICc>2: HNp/HNc
(que resultaron ser iguales al no incluirse ningún termino de ajuste), NEc, UNc y
NEp (Anexo II, tabla 2.). Se calculó el peso de ponderación de cada uno de los
cuatro modelos (Tabla 1.) así como la media ponderada de abundancia y densidad
por estrato (hábitat).
La abundancia (N) de la tarabilla canaria en Fuerteventura es de 20.504
individuos (CI 95%: 16.217-25.973) (Anexo II, tabla 3.). La densidad calculada por
hábitat se puede ver en la tabla 2. (cálculos en Anexo II, tablas 4 y 5).
Como se puede observar los mejores hábitats son los de matorral alto,
medio y bajo con pendientes elevadas (>11). El hábitat de matorral medio con
pendientes medias (6-11) es medianamente bueno. Los hábitats de bosque de todo
el rango de pendientes parecen ser medianamente buenos. En estas tres últimas
categorías de bosque están incluidos parches de vegetación que consisten
principalmente en tabaibales, la aptitud de los cuales para la tarabilla canaria
D1 se_ignora 02 sin_vegetar,>5 03 pastizal,>5 0,03654 matorral_bajo,6-11 0,05235 matorral_bajo,<=5 0,02306 sin_vegetar,<=5 07 matorral_medio,<=5 0,06388 pastizal,<=5 0,01319 matorral_medio,6-11 0,119110 medio_rural 011 matorral_bajo,>11 0,251312 matorral_alto,<=5 013 matorral_alto,6-11 014 matorral_medio,>11 0,432015 matorral_alto,>11 0,312016 bosque,<=5 0,119117 bosque,6-11 0,186118 bosque,>11 0,145919 acuatica 0
HABITAT
Tabla 2. El valor de la media ponderada de loscuatro modelos de la densidad(n/ha) por cada hábitat
15

depende también de otros factores no incluidos en la asignación de las categorías
de vegetación como la densidad de la vegetación; además del área cubierta por un
bosque de pinar situado en el centro de la isla, lo cual constituye un hábitat
bastante malo para la especie. Por lo tanto, estos tres tipos de hábitat pueden
tener asignados valores de densidad que no corresponden a la totalidad de los
medios incluidos en estas categorías y pueden resultar poco fiables.
Los resultados de densidad se pueden ver incorporados en el mapa de
hábitat en el Mapa.1.
Mapa 1. Mapa de densidad en cinco categorías basado en el mapa de hábitat y el valorde densidad por hábitat (n/ha) previamente calculado.
16

El mapa de hábitat resultante muestra dos grandes zonas muy buenas para
la especie, una situada en la península de Jandía y otra en Betancuria en el centro-
oeste de la isla (Mapa 1., zonas de color azul oscuro). Gran parte de la isla resulta
ser hábitat bueno o medianamente bueno para la tarabilla canaria (Mapa 1., zonas
de color azul claro), mientras que zonas regulares hay bastante pocas (Mapa 1.,
color verde). Las zonas malas se sitúan en el centro y norte de la isla, en la costa y
en el istmo de Jandía (Mapa 1., colores rojo y amarillo).
Mapa 2. Mapa de cuadriculas UTM de 1x1Km de la densidad (n/Km2) y distribución dela tarabilla canaria en Fuerteventura.
3.3. Distribución y Validación
Superponiendo la malla de cuadriculas UTM de 1Km2 al Mapa 1. se pudo
extraer la información necesaria sobre el área que ocupa cada hábitat en las
cuadriculas y calcular el valor de densidad por cuadricula, lo cual resulta a un mapa
de distribución en cuadriculas UTM 1XKm (Mapa.2).
17

Después de calcular el valor de densidad por cuadrícula para cada
remuestreo bootstrap y el valor de la densidad observada, se pudo continuar con la
validación de los resultados de distribución. La correlación de Pearson se realizó
entre la densidad observada y la predicción de densidad por cuadricula para cada
uno de los 1000 remuestreos. Los valores se pueden observar en el Gráfico 4.
Del valor de la mediana de las 1000 correlaciones de Pearson se puede
deducir que hay una correlación positiva entre la densidad observada y la densidad
predicha. Con un test de permutaciones se estableció el p-valor, p<0.001.
Un gráfico de cajas nos puede proporcionar con una evaluación cualitativa
de las cuadriculas. El gráfico se construyó con siete grupos de valores de densidad
con el último grupo construido a partir de los valores máximos de densidad
observada (Gráf. 5). Las cuadrículas se dividieron según su valor predicho en estos
siete grupos y se ilustró con cajas el rango de valores de densidad observada de las
cuadrículas de cada grupo. Además, se incorporaron los valores medios de cada
grupo para facilitar la evaluación del gráfico (Gráf. 5).
R2
Correlación de Pearson
0.48
50.
490
0.49
50.
500
0.50
50.
510
0.51
5
R2
Correlación de Pearson
0.48
50.
490
0.49
50.
500
0.50
50.
510
0.51
5
Graf. 4. Mediana de los resultados de las 1000 correlaciones realizadas entre la densidad observada y la predicha
18

020
4060
8010
012
014
0
[0,5) [5,10) [10,20) [20,30) [30,40) [40,60) [60,228]
GRUPOS DE CUADRICULAS SEGÚN D
DEN
SID
AD
(n/K
m2)
Nb=1000 Nb=1000 Nb=1000 Nb=1000 Nb=991 Nb=466 Nb=0
No=514 No=16
No=37
No=25
No=11
No=24
No=34
Densidad observada
Densidad media de grupo de cuadriculas
020
4060
8010
012
014
0
[0,5) [5,10) [10,20) [20,30) [30,40) [40,60) [60,228]
GRUPOS DE CUADRICULAS SEGÚN D
DEN
SID
AD
(n/K
m2)
Nb=1000 Nb=1000 Nb=1000 Nb=1000 Nb=991 Nb=466 Nb=0
No=514 No=16
No=37
No=25
No=11
No=24
No=34
Densidad observada
Densidad media de grupo de cuadriculas
020
4060
8010
012
014
0
[0,5) [5,10) [10,20) [20,30) [30,40) [40,60) [60,228]
GRUPOS DE CUADRICULAS SEGÚN D
DEN
SID
AD
(n/K
m2)
Nb=1000 Nb=1000 Nb=1000 Nb=1000 Nb=991 Nb=466 Nb=0
No=514 No=16
No=37
No=25
No=11
No=24
No=34
020
4060
8010
012
014
0
[0,5) [5,10) [10,20) [20,30) [30,40) [40,60) [60,228]
GRUPOS DE CUADRICULAS SEGÚN D
DEN
SID
AD
(n/K
m2)
Nb=1000 Nb=1000 Nb=1000 Nb=1000 Nb=991 Nb=466 Nb=0
No=514 No=16
No=37
No=25
No=11
No=24
No=34
020
4060
8010
012
014
0
[0,5) [5,10) [10,20) [20,30) [30,40) [40,60) [60,228]
GRUPOS DE CUADRICULAS SEGÚN D
DEN
SID
AD
(n/K
m2)
Nb=1000 Nb=1000 Nb=1000 Nb=1000 Nb=991 Nb=466 Nb=0
No=514 No=16
No=37
No=25
No=11
No=24
No=34
Densidad observada
Densidad media de grupo de cuadriculas
Densidad observadaDensidad observada
Densidad media de grupo de cuadriculasDensidad media de grupo de cuadriculas
Graf. 5. La mediana de los valores de densidad observada según los grupos de valorespredichos por el remuestreo bootstrap para las 661 cuadriculas que contienen transectos.Debajo de la representación grαfica se presenta el número de remuestreos bootstrap a losque aparecen cuadriculas que pertenecen a ese grupo de valores (Nb); y encima de larepresentación grafica el numero de cuadriculas de cada grupo que tiene un valor dedensidad observada asignado (No).
Según el Gráfico 5 parece que las cuadriculas de mejor calidad, con un valor
medio de densidad predicha mayor que 20 n/Km2, han sido subestimadas por la
predicción ya que tienen asignado un valor de densidad bastante inferior a la
densidad observada en esas mismas cuadriculas. Además, no hay cuadrículas en
las que se haya predicho un valor de densidad tan elevado como el observado en
algunas cuadrículas (Graf. 1, último grupo de valores de densidad). Esto puede
deberse a que realmente las cuadriculas han sido subestimadas o a que son
cuadriculas con pocos transectos con altas densidades locales que sobrestiman la
densidad para toda la superficie de la cuadricula. La extrapolación de la densidad
observada por superficie censada a toda la cuadricula supone un grado de error ya
que se considera que las condiciones son iguales en toda la superficie extrapolada.
Por esta razón, en cuadriculas que contienen una proporción muy alta de contactos
por transecto el valor de densidad observada se dispara a la hora de extrapolarlo a
la totalidad de la superficie de la cuadricula y llega a tomar valores muy altos que
no han sido predichos para ninguna cuadricula como se observa en el último grupo
(60-228). Mientras tanto, la predicción de la densidad, al ser estimada basándose
en el hábitat presente y las observaciones en toda la superficie de este hábitat está
más suavizada.
La predicción de la densidad para cuadriculas de peor calidad parece haber
sido más precisa, aunque ligeramente sobreestimada. Esto puede deberse a que los
transectos a los que se basa el calculo de la densidad observada no se ubican en el
19

hábitat más representativo de la cuadricula, obteniendo un número de
observaciones menor de las predichas por la proporción de hábitat.
Para intentar comparar las densidades minimizando estos efectos se repitió
el gráfico utilizando solamente cuadriculas en las que se han realizado más que dos
transectos, es decir, donde se hizo un mayor esfuerzo de muestreo (Gráf.6).
010
2030
4050
60
[0,5) [5,10) [10,20) [20,30) [30,40) [40,60) [60,118]
GRUPOS DE CUADRICULAS SEGÚN D
DEN
SID
AD
(n/K
m2)
Nb=1000 Nb=1000 Nb=1000 Nb=1000 Nb=738 Nb=43 Nb=0
No=152No=16
No=22
No=3
No=6
No=8
No=5
Densidad observada
Densidad media de grupo de cuadriculas
010
2030
4050
60
[0,5) [5,10) [10,20) [20,30) [30,40) [40,60) [60,118]
GRUPOS DE CUADRICULAS SEGÚN D
DEN
SID
AD
(n/K
m2)
Nb=1000 Nb=1000 Nb=1000 Nb=1000 Nb=738 Nb=43 Nb=0
No=152No=16
No=22
No=3
No=6
No=8
No=5
010
2030
4050
60
[0,5) [5,10) [10,20) [20,30) [30,40) [40,60) [60,118]
GRUPOS DE CUADRICULAS SEGÚN D
DEN
SID
AD
(n/K
m2)
Nb=1000 Nb=1000 Nb=1000 Nb=1000 Nb=738 Nb=43 Nb=0
No=152No=16
No=22
No=3
No=6
No=8
No=5
Densidad observada
Densidad media de grupo de cuadriculas
Densidad observadaDensidad observada
Densidad media de grupo de cuadriculasDensidad media de grupo de cuadriculas
Graf. 6. La mediana de los valores de densidad observada según los grupos de valorespredichos por el remuestreo bootstrap para las 212 cuadriculas que contienen un númerode transectos mayor que dos. Debajo de la representación grαfica se presenta el númerode remuestreos bootstrap a los que aparecen cuadriculas que pertenecen a ese grupo devalores (Nb); y encima de la representación grαfica el número de cuadriculas de cadagrupo que tiene un valor de densidad observada asignado (No).
En el Gráfico 6 se observa mayor concordancia entre los valores de densidad
observada y predicha para la mayoría de los grupos. En el grupo de las mejores
cuadriculas según los valores de densidad predicha (40-60) sigue habiendo una
discordancia significativa, aunque esta vez lo que se observa es una sobreestima
del valor predicho en comparación con el valor observado.
3.4. Nivel de protección del área de distribución de la especie
Incorporando un mapa de las Zonas de Especial Protección para las Aves
(ZEPAs, EUROPARC-España, 2009) en la isla de Fuerteventura al mapa de
distribución elaborado se puede hacer una estima visual del estado de protección
del área de distribución de la tarabilla canaria según el resultado de este estudio
(Mapa 3.).
20

Mapa 3. Comparación de las ZEPAs en la isla de Fuerteventura con la distribución dela tarabilla canaria
Como se observa en el mapa la mayor parte del área de altas densidades de
tarabillas canarias (cuadriculas de color azul oscuro) está incluida en las Zonas de
Especial Protección para las Aves, excluyendo dos áreas; una en el Centro-
Norte/Noroeste de la isla y otra al Norte de la península de Jandia.
21

4. DISCUSION 4.1. Abundancia y Densidad
El tamaño de la población de la tarabilla canaria estimado por este estudio
(20.504 individuos) resulta superior con diferencia a lo hasta ahora estimado. Bibby
& Hill (1987) en el primer intento de calcular la abundancia de la especie en 1987,
estimaron la población en 1.500 individuos, mientras que García del Rey (2009) en
un estudio más reciente estimó 1.035 individuos. Esta discordancia entre los
valores estimados podría deberse a diferencias en los métodos de muestreo o de
extrapolación de los valores al área total de distribución.
Aunque los resultados obtenidos no son directamente comparables con la
estima de Bibby & Hill (1987) ya que es una estima de la abundancia relativa
(Norvell, 2003), lo son con el estudio de García del Rey (2009), que utilizó el
mismo método de “distance sampling”. Entre los tres trabajos hay diferencias en el
diseño de muestreo y el área total de extrapolación, por lo tanto se puede intentar
una comparación entre los métodos utilizados.
Bibby & Hill (1987) basaron su estudio en un muestreo de 21 bloques de 12
cuadriculas de 1x1Km seleccionados al azar entre todas las cuadriculas de la isla. El
censo se realizó cubriendo toda la superficie de las cuadriculas, excluyendo
posteriormente del análisis las que no han podido ser recorridas enteras por falta
de tiempo. La densidad se calculó por bloque de cuadriculas y utilizando el método
“jack-knife” se estimó el tamaño poblacional por parejas y por número total de
machos. La abundancia total se estimó considerando que se ha detectado el 90%
de las parejas presentes en la isla. Para validar esta suposición debería quedar
claro cual fue el método de censo utilizado para llegar a la misma. Carrascal et al.
(2008) demostraron que en ambientes semi-desérticos se puede detectar más que
el 85% de los individuos presentes, pero utilizando un ancho de detección de 25m
por cada lado del transecto. Por lo tanto, con los métodos de censo de transectos
de ancho de banda fijo, utilizados hasta ahora, se subestima ligeramente la
densidad de la especie censada en medios como los de las Islas Canarias. Bibby &
Hill (1987) no especifican el ancho de detección que consideraron durante los
censos, pero es probable que fuera mayor que 25m.
Por otro lado, García del Rey (2009) utiliza un método de extrapolación
mucho más robusto, pero ni el muestreo realizado ni el área a la que extrapola son
representativos del área total de distribución de la especie. Durante el diseño de
muestreo García del Rey excluyó del estudio gran parte de la isla basándose en la
distribución de la especie que estimaron Bibby & Hill (1987), en la bibliografía
disponible sobre el hábitat de la tarabilla (Illera, 2001) y en conocimiento propio
22

de la isla. Finalmente, redujo el universo de muestreo a 1.953 ha (19,53 Km2) de
las cuales parte es de laderas y parte de barrancos. Dentro de esta área muestreó
140 cuadriculas de 500x500m al azar estratificando por laderas y barrancos. La
estima de la abundancia se realizó con el programa Distance 4.1 utilizando el
mismo método que se aplicó en el presente trabajo, pero con diferentes estratos y
extrapolando los resultados solamente a la superficie predeterminada por él. García
del Rey subestimó el tamaño poblacional de la tarabilla canaria excluyendo áreas
donde, por propio conocimiento, está presente la especie e ignorando un factor
muy importante que es la estructura de la vegetación.
4.2. Distribución y Validación
En cuanto a la distribución de la especie, parece ser que las estimas del
presente trabajo coinciden en gran parte con las de Bibby & Hill (1987). Como se
puede observar en la imagen 5., el mapa de distribución resultante del presente
trabajo concuerda con la distribución de la especie propuesta por Bibby & Hill
(1987).
La validación de los resultados mediante el remuestreo bootstrap señaló la
robustez del modelo de campo aplicado, indicando posibles errores en la estima que
por lo demás parece ser bastante precisa. Los errores se deben al método de
validación en si, además de la estima realizada. El método empírico de asignación
de clases de vegetación ha sido poco preciso en varias partes de la isla, debido a la
Imagen 5. (a) el mapa de distribución resultante del presente estudio, (b) la distribución de la especiepropuesta por Bibby & Hill (1987).
23

falta de información de estructura vegetal. En muchos casos la altura de la
vegetación se asignó basándose solamente en la información taxonómica
proporcionada por el mapa sin tener en cuenta las condiciones presentes (p.ej.
carga de pastoreo). Por otro lado, la densidad de la vegetación no se ha tenido en
cuenta por falta de información, a pesar de que parece que influye bastante en la
selección de hábitat de la tarabilla canaria (Illera, 2001). Estas faltas en el método
de asignación de tipos de vegetación resultaron en la posible unificación de áreas
diferentes en cuanto a los factores que influyen la presencia de la tarabilla canaria.
Además, la disminución de la superficie en cuadriculas de 1x1Km dificultó la estima
de la densidad observada, resultando a la presencia de pocos transectos por
cuadricula o la concentración de ellos en tipos de hábitat que no son
representativos de toda la cuadricula provocando el elevado desajuste entre los
valores observados y predichos en varias cuadriculas.
El método idóneo para evaluar las predicciones estimadas por este estudio
sería utilizando datos independientes. Esto se lograría con una repetición del
muestreo de campo y la comparación de los datos obtenidos con la predicción. Por
desgracia, pocos estudios han intentado validar modelos de distribución y hábitat
utilizando datos independientes, principalmente porque no existen datos disponibles
y resulta costoso en tiempo y esfuerzo recogerlos (Jewell et al., 2007).
El procedimiento de validación interna seguido en el presente trabajo
proporciona una estima muy precisa de la exactitud de clasificación,
particularmente cuando se trata de evaluar modelos predictivos (Verbyla & Litvaitis,
1989; Jewell et al., 2007). Otros métodos ampliamente utilizados, como la
validación cruzada (cross-validation) o la validación en 10 partes (10-fold
validation), han sido descartados por tener que omitir parte de los datos del análisis
principal para realizar este tipo de evaluación. Omitiendo una parte de los datos del
análisis cambiaría los resultados de distribución iniciales, ya que estos resultados se
basan exclusivamente a las observaciones de campo. Por lo tanto, utilizando el
método de remuestreo con bootstrap obtenemos una evaluación bastante precisa
de la robustez de los resultados, aunque esto no excluye la necesidad de validar las
predicciones utilizando datos independientes.
4.3. Estado de Conservación
Actualmente, la tarabilla canaria está catalogada como especie en peligro
según los criterios B1a-b(ii,iii,iv,v) y C2a(ii) de la UICN (Illera, 2004). Según, el
tamaño poblacional estimado en este estudio el criterio C2a (ii) no se cumple, ya
que se refiere a especies con población pequeña, menor que 2.500 individuos
maduros, y en declive continuado (observado, proyectado o inferido) en el número
24

de individuos maduros (C2), con estructura de la población con al menos 95% de
los individuos maduros en una subpoblación (a (ii)) (Madroño et al., 2004). El
criterio B1a-b(ii,iii,iv,v) se refiere a especies con área de distribución/ ocupación
pequeña, <5.000 Km2 (B1), severamente fragmentada ó que solo se conoce en 5
localidades (a) y con declive continuo en: el área de ocupación (b (ii)); el área,
extensión y/o calidad del hábitat (b (iii)), el número de localidades o
subpoblaciones (b (iv)) y el número de individuos maduros (b (v)) (Madroño et al.,
2004). Este criterio sigue siendo valido, aunque no está comprobado el último
apartado (b(v)).
Aún teniendo un tamaño de población mayor con diferencia de lo esperado,
la especie sigue siendo amenazada por la rápida expansión urbanística en la isla de
Fuerteventura y la pérdida de su hábitat. Según Viada (1998), las mayores
amenazas en gran parte de las áreas identificadas como importantes para la
especie son el turismo y el sobre-pastoreo.
Las principales fuentes de ingreso en las islas Canarias son el sector
primario (ganadería, pesca etc.), el turismo y la construcción (fuente: Gobierno de
Canarias, 2009). La ganadería, principalmente de cabras, constituye una amenaza
importante para la tarabilla, ya que sus preferencias de hábitat incluyen un cierto
volumen de vegetación. Sin embargo, la amenaza principal es la construcción y
consecuentemente el turismo (Illera, 2001). La rápida expansión urbanística,
incluyendo el aumento en viviendas, hoteles y campos de golf, que se ha observada
durante las últimas décadas en Fuerteventura debido a la industria de recreo tiene
como consecuencia la perdida y partición del hábitat de la especie.
La tarabilla canaria es una especie extremadamente filopátrica (Illera, 2001;
2006). La partición de su hábitat debida a la expansión urbanística, así como el
istmo de Jandía, la parte desértica que conecta la península de Jandía con el resto
de la isla, pueden separar los distintos núcleos poblacionales.
Las islas Canarias son una de las tres Áreas de Aves Endémicas (EBAs)
identificadas en Europa por Birdlife International. El Área de Aves Endémicas de
Madeira y las islas Canarias albergan 10 especies endémicas, de las cuales 9 están
presentes en Canarias, siendo 5 de ellas exclusivas de este archipiélago. Tenerife
tiene el mayor número de especies de distribución restringida y Fuerteventura
cuenta con la Tarabilla Canaria.
En la revisión de las Áreas Importantes para las Aves en España publicada
por la SEO/Birdlife (Viada, 1998) se identifican nueve áreas importantes para la
tarabilla canaria (EBAs). De estas nueve áreas sólo cinco cuentan con un régimen
de protección para las aves. Las áreas incluidas en las ZEPAs de Fuerteventura son:
el macizo de Pozo Negro-Vigán, la península de Jandía, el barranco de Ajuí-
25

Betancuria, el barranco de los Molinos-llano de la Laguna y el morro Tabaiba-morro
de los Rincones- Vallebrón.
Las cuatro áreas restantes se pueden ver en la Imagen 6. De estas cuatro,
dos (el barranco del río Cabras y la costa de Corralejo-Tostón) no albergan
densidades grandes de la especie según este estudio. Sin embargo, el Cuchillete de
Buenavista-Barranco de la Torre-Los Alares y el Macizo de Tarajalejo parecen ser
áreas importantes para la especie y no están bajo ninguna figura de protección.
Existe un área extenso entre Betancuria y la Península de Jandía, señalado
en la Imagen 6., que según este estudio es un área de altas densidades de la
especie. Sin embargo no pertenece en ninguna de las ZEPAs de la isla y tampoco
ha sido identificada como EBA por Viada (1998).
Costa de Corralejo-Tostón
Mazico de Tarajalejo
Cuchillete de Buenavista-Barranco de la Torre-Los Alares
Barranco del río Cabras
Costa de Corralejo-Tostón
Mazico de Tarajalejo
Cuchillete de Buenavista-Barranco de la Torre-Los Alares
Barranco del río Cabras
Costa de Corralejo-Tostón
Mazico de Tarajalejo
Cuchillete de Buenavista-Barranco de la Torre-Los Alares
Barranco del río Cabras
Costa de Corralejo-Tostón
Costa de Corralejo-Tostón
Costa de Corralejo-Tostón
Costa de Corralejo-Tostón
Mazico de TarajalejoMazico de Tarajalejo
Cuchillete de Buenavista-Barranco de la Torre-Los Alares
Cuchillete de Buenavista-Barranco de la Torre-Los Alares
Barranco del río Cabras
Barranco del río Cabras
Imagen 6. Las cuatro IBAs propuestas por Viada (1998) que no han sido incluidas enlas ZEPAs de la isla. En azul y señalada por la flecha roja está el área entre Betancuriay la península de Jandía que no ha sido identificada como zona de importancia para laespecie ni pertenece en ninguna de las ZEPAs.
26

5. CONCLUSIONES El tamaño poblacional de la tarabilla canaria es bastante más alto de lo que
se estimaba hasta ahora. Aunque haya habido cambios en el tamaño poblacional de
la especie durante los últimos 20 años, una diferencia tan marcada probablemente
no se debe a las fluctuaciones poblacionales sino a los métodos utilizados para su
estima.
La especie está catalogada como “En Peligro” (EN) principalmente por su
tamaño poblacional estimado hasta ahora y su limitada área de distribución y
endemismo. A pesar de la nueva información aportada por este trabajo sobre su
tamaño poblacional, la tarabilla canaria sigue siendo amenazada por la perdida de
su hábitat mediante la urbanización, el sobre-pastoreo etc.
Para completar este estudio habría que conseguir el plan urbanístico de
Fuerteventura y averiguar como influirían las construcciones previstas para el
futuro próximo a los núcleos poblacionales de la especie detectados por el mapa de
distribución elaborado. Así como, intentar una validación más precisa de la
distribución de la tarabilla, quizá calculando la densidad observada dividiendo la isla
en áreas geográficas más grandes y comparándola con la predicción. Es
imprescindible, además validar los resultados con nuevos datos de campo
independientes de los ya utilizados para estimar la abundancia y distribución de la
especie.
Dado que los resultados de este trabajo se alejan mucho de previas estimas
sería conveniente intentar de nuevo una estima, esta vez con un modelo estadístico
de predicción utilizando variables ambientales de vegetación y pendiente o un
índice de vegetación más preciso (p.ej. NDVI) y comprobar su concordancia con los
resultados de este trabajo.
Finalmente, sería útil estudiar en el futuro el efecto de aislamiento que
puede haber entre el núcleo poblacional de la península de Jandía y el resto de la
población, debido al istmo de Jandía.
27

REFERENCIAS Bibby, C.J., Burgess, N.D., Hill, D.A. & Mustoe, S.H., 2000. Bird Census Techniques. Academic Press. London Bibby, C.J. & Hill, D.A., 1987. Status of the Fuerteventura Stonechat Saxicola dacotiae. IBIS 129: 491-498 Bibby, C.J. & Buckland, S.T., 1987. Bias of bird census results due to detectability varying with habitat. Acta Ecologica 8: 103-112 Buckland, S.T., Anderson, D.R., Burnham, K.P., Laake, J.L., Borchers, D.L., & Thomas, L., 2001. Introduction to Distance Sampling: Estimating abundance of biological populations. (1 ed.). Oxford: Oxford University Press. Buckland, S.T. & Elston, D.A., 1993. Empirical models for the spatial distribution of wildlife. Journal of Applied Ecology 30: 478-495 Buckland, S.T., Marsden, S.J. & Green, R.E., 2008. Estimating bird abundance: making methods work. Bird Conservation International 18: 91-108 Burnham, K.P. & Anderson, D.R., 2002. Model selection and multimodel inference: a practical information-theoretic approach. Springer-Verlag New York, Inc.2nd edition. Carrascal L.M., Seoane J., Palomino D. y Alonso C.L. (2006) Preferencias de hábitat, estima y tendencias poblacionales de la avutarda hubara Chlamydotis undulata en Lanzarote y la Graciosa (Islas Canarias). Ardeola 53: 251-269 Carrascal L.M., Palomino D., Seoane J. y Alonso C.L. 2008a Habitat use and population density of the houbara bustard Chlamydotis undulata in Fuerteventura (Canary Islands). African Journal of Ecology 46: 291-302 Carrascal, L.M., Seoane, J. & Palomino, D., 2008b. Bias in the density estimations using strip transects in dry open-country environments in the Canary Islands. Animal Biodiversity and Conservation 31.2: 45-50 Conlisk, E., Conlisk, J., Enquist, B., Thompson, J. & Harte, J., 2009. Improved abundance prediction from presence-adsence data. Global Ecol. Biogeogr. 18: 1-10 Del Arco, M., Wildpret, W., Pérez de Paz, P.L., Rodriguez, O., Acebes, J.R., García, A., Martín, V.E., Reyes, J.A., Salas, M., Díaz, M.A., Bermejo, J.A., González, R., Cabrera, M.V. & García, S., 2003. Cartografía 1:25.000 de la Vegetación Canaria. GRAFCAN S.A. Santa Cruz de Tenerife. Accesible en: http://idecan2.grafcan.es/ServicioWMS/Vegetacion? EUROPARC-España, 2009. Visualizador Cartográfico, última actualización en 2007. http://www.europarc-es.org/intranet/EUROPARC/preview/descargas_vis.html García del Rey, E., 2009. The status of the Fuerteventura Stonechat Saxicola dacotiae: a new evaluation using distance sampling and GIS. Bird Conservation International 19: 1-9 Gobierno de Canarias, 2008. Red Canaria de Espacios Naturales Protegidos –Fuerteventura-http://www.gobiernodecanarias.org/cmayot/espaciosnaturales/informacion/fuertev_todo.html Gobierno de Canarias, 2009. Estadísticas de la Comunidad Autónoma de Canarias. http://www.gobiernodecanarias.org/istac/estadisticas.html Guisan, A. & Zimmermann, N.E., 2000. Predictive habitat distribution models in ecology. Ecological Modelling 135: 147-186. Illera, J.C., 2001. Habitat selection by the Canary Island stonechat (Saxicola dacotiae) (Meade-Waldo, 1889) in Fuerteventura Island: a two-tier habitat approach with implications for its conservation. Biological Conservation 97:339-345
28

Illera, J.C., 2003. Tarabilla Canaria, Saxicola dacotiae. En, Martí, R. & Del Moral, J.C. (Eds.): Atlas de las Aves Reproductoras de España, pp.428-429. Dirección General de Conservación de la Naturaleza-Sociedad Española de Ornitología.Madrid. Illera, J.C., 2004. Tarabilla Canaria, Saxicola dacotiae. En Madroño, A., González, C. & Atienza, J.C. (Eds.): Libro Rojo de las Aves de España, pp. 327-328, 382. Dirección General para la Biodiversidad-SEO/ Birdlife. Madrid. Illera, J.C. & Díaz, M., 2006. Reproduction in an endemic bird of a semiarid island: a food-mediated process. Journal of Avian Biology 37 (5): 447-456 Illera, J.C., 2007. Tarabilla Canaria, Saxicola dacotiae. En Lorenzo, J.A. (Ed.): Atlas de las aves nidificantes en el archipiélago canario (1997-2003), pp.356-357. Dirección General de Conservación de la Naturaleza-Sociedad Española de Ornitología. Madrid IUCN 2007. 2007 IUCN Red List of Threatened Species. <www.iucnredlist.org>. Último acceso en 12 Febrero 2008. Jewell, K.J., Arcese, P. & Gergel, S.E., 2007. Robust predictions of species distribution: Spatial habitat models for a brood parasite. Biological Conservation 140: 259-272 Madroño, A., González, C. & Atienza, J.C. (Eds.), 2004. Libro Rojo de las Aves de España, pp. 22-23. Dirección General para la Biodiversidad-SEO/ Birdlife. Madrid. Nichols, J.D., Hines, J.E., Sauer, J.R., Fallon, F.W., Fallon, J.E. & Heglund, P.T., 2000. A double-observe approach for estimating detection probability and abundance from point counts. Auk 117: 393-408 Norvell, R.E., Howe, F.P. & Parrish, J.R., 2003. A seven-year comparison of Relative-Abundance and Distance-Sampling methods. Auk 120: 1013-1028 Osborne, P.E, Alonso, J.C., Bryant, R.G., 2001. Modelling ladscape-scale habitat use using GIS and remote sensing: a case study with great bustard. Journal of Applied Ecology 38: 458-471 Osborne, P.E & Tigar, P.J, 1992. Interpreting bird atlas data using logistic models: an example from Lesotho, Southern Africa. Journal of Applied Ecology 29: 55-62 Palomino D., Seoane J., Carrascal L.M. y Alonso C.L. (2008) Competing effects of topographic, lithological, vegetation structure and human impact in the habitat preferences of the Cream-coloured Courser. Journal of Arid Environments 72: 401-410 Seoane, J., Justribó, J.H., García, F., Retamar, J., Rabadán, C., Atienza, J.C., 2006. Habitat-suitability modelling to assess the effects of land-use changes on Dupont’s lark Chersophilus duponti: A case study in the Layna Important Bird Area. Biological Conservation 128:241-252 Seoane, J., Viñuela, J., Díaz-Delgado, R., Bustamante, J., 2003. The effects of land use on red kite distribution in the Iberian peninsula. Biological Conservation 111: 401-414 Thomas, L., Buckland, S.T., Burnham, K.P., Anderson, D.R., Laake, J.L., Borchers, D.L. and Strindberg, S., 2002. Distance sampling. Encyclopedia of Environmetrics 1: 544-552 Thomas, L., Laake, J.L., Strindberg, S., Marques, F.F.C., Buckland, S.T., Borchers, D.L., Anderson, D.R., Burnham, K.P., Hedlev, S.L., Pollard, J.H., Bishop, J.R.B. and Marques, T.A., 2006. Distance 5.0. University of St. Andrews, UK: Research Unit for Wildlife Population Assessment. http://www.ruwpa.st-and.ac.uk/distance/ Verbyla, D.L. & Litvaitis, J.A., 1989. Resampling Methods for Evaluating Classification Accuracy of Wildlife Habitat Models. Environmental Management 13.6: 783-787 Viada, C. (ed.), 1998. Áreas Importantes para las Aves en España. 2ª edición revisada y ampliada. Monografía Nº5 pp. 80-84, 324-330, 369-370. SEO/ Birdlife. Madrid.
29

I
ANEXO I. CARTOGRÁFICO
Mapa 1. Mapa de las nueve categorías de vegetación según su estructura

II
Mapa 2. Mapa de las tres categorías de pendiente elegidas
III
Mapa 3. Mapa de las 19 categorías de hábitat derivadas de la combinación de estructura devegetación y pendiente

IV
ANEXO II. TABLAS Y GRÁFICOS
1. Asignación de categorías de pendientes Las categorías de pendiente se eligieron procurando obtener un número de
transectos mayor que 10 por categoría de hábitat (combinación de categorías de
vegetación y pendiente). Las categorías que se unificaron para conseguir esto están
señaladas en color amarillo en la tabla 1.
slope bosque mat_alto mat_bajo mat_medio medio_rural pastizal sin_vegetar Total general0-5 18 28 584 15 8 38 19 7106-11 13 11 375 13 4 15 9 440>11 11 36 252 16 1 3 2 321Total general 42 75 1211 44 13 56 30 1471
Tabla 1. El número de transectos realizados según la combinación de categorías de pendiente yvegetación. Las categorías de vegetación “acuática” y “se ignora” no están incluidas ya que no se hanmuestreado.
2. Resultados de los modelos
Según el test de Kolmogorov-Smirnov y el grupo de tests de Cramer-von Mises
realizados para los cuatro modelos con el programa Distance, el desajuste de la
distribución no resulta significativo en ninguno de los casos.
Tabla 2. Resultados de las pruebas de normalidad realizadas con el programa Distance 5.0
Modelo Kolmogorov-Smirnov Cramer-von Mises (uniforme) Cramer-von Mises (cosino) HNp80 p=0.1615 0.4<p≤0.5 0.3<p≤0.4 NEc80 p=0.3473 0.4<p≤0.5 0.3<p≤0.4 UNc80 p=0.0773 0.2<p≤0.3 0.15<p≤0.2 NEp80 p=0.2009 0.4<p≤0.5 0.3<p≤0.4
En las tablas siguientes se presentan los cálculos realizados a partir de los
resultados del programa Distance 5.0. para obtener los valores medios, ponderados
según el peso de cada modelo, de la abundancia (tabla 3.), la densidad por
categoría de hábitat (tabla 5.) y el ancho de banda efectivo (ESW, tabla 6.).
Tabla3. Calculos de la media ponderada de abundancia
Modelo bIC
extremo inferior
NIC
extremo superior
b*IC extremo inferior
b*Nb*IC
extremo superior
80 0.394 15602 19366 23732 6141.15 7622.71 9341.23c80 0.230 16376 21800 29444 3763.94 5010.62 6767.5580 0.189 15433 19161 24057 2910.02 3612.96 4536.14
NEp80 0.188 18095 22648 28346 3401.56 4257.45 5328.59TOTAL 16216.68 20503.75 25973.51
HNpNEUNc
Tabla 3. Cálculos para obtener el valor medio de abundancia ponderado por el peso (b) de cada uno delos cuatro modelos

V
Tabla 4. Los resultados de densidad por hábitat para los cuatro mejores modelos obtenidos mediante el análisis con el programa Distance 5.0. Los hábitats a los que no se han detectado individuos de la especie estudiada no están incluidos en la tabla.
Modelo D_3 D_4 D_5 D_7 D_8 D_9 D_11 D_14 D_15 D_16 D_17 D_18HNp80 0.035 0.050 0.016 0.061 0.013 0.114 0.240 0.413 0.298 0.114 0.195 0.139NEc80 0.038 0.054 0.018 0.066 0.014 0.124 0.262 0.450 0.325 0.124 0.213 0.152UNc80 0.033 0.048 0.048 0.058 0.012 0.108 0.229 0.393 0.284 0.108 0.186 0.133NEp80 0.041 0.059 0.019 0.072 0.015 0.135 0.285 0.491 0.354 0.135 0.135 0.166
Tabla 5. Cálculos realizados para obtener el valor medio de densidad por categorías de hábitat, ponderado por el peso (b) de los cuatro modelos. Igual que en la previa tabla, en los hábitat ausentes de la tabla no se detectaron individuos de la especie.
Modelo b*D3 b*D4 b*D5 b*D7 b*D8 b*D9 b*D11 b*D14 b*D15 b*D16 b*D17 b*D18HNp80 0.014 0.020 0.006 0.024 0.005 0.045 0.094 0.162 0.117 0.045 0.077 0.055NEc80 0.009 0.013 0.004 0.015 0.003 0.028 0.060 0.103 0.075 0.028 0.049 0.035UNc80 0.006 0.009 0.009 0.011 0.002 0.020 0.043 0.074 0.054 0.020 0.035 0.025NEp80 0.008 0.011 0.004 0.014 0.003 0.025 0.054 0.092 0.067 0.025 0.025 0.031TOTAL 0.036 0.052 0.023 0.064 0.013 0.119 0.251 0.432 0.312 0.119 0.186 0.146
Tabla 6. Cálculos realizados para obtener elvalor medio ponderado del ancho de bandaefectivo (ESW).
Modelo b ESW b*ESWHNp80 0.394 41.023 16.147NEc80 0.230 37.643 8.652UNc80 0.189 43.058 8.119NEp80 0.188 34.499 6.485
TOTAL 39.403
ANEXO III. METODOLOGÍA GIS DETALLADA
1. Creación del mapa de vegetación
El mapa de vegetación se creó generando un nuevo campo en el mapa base
(mapa de vegetación canaria 1:25.000) y añadiendo las categorías de estructura de
vegetación previamente mencionadas. Las categorías se asignaron a cada polígono
según la información de vegetación principal, secundaria, terciaria y potencial
proporcionada por el mapa base, información de hábitat recolectada durante la
realización de los censos, conocimiento propio del medio e información
complementaria de la ortofotografía de Fuerteventura.
2. Elaboración del mapa de pendientes
El mapa de pendientes se elaboró a partir de un Modelo Digital de Elevación
(DEM) que se consiguió. Las pendientes se calcularon a través del programa Idrisi
Kilimanjaro con la herramienta adecuada.

VI
3. Creación del mapa de hábitat
El mapa de hábitat final se creó uniendo el mapa de pendientes (raster) con
el mapa de vegetación (previamente convertido en raster con pixeles del mismo
tamaño, 90x90). La unión se realizó con la herramienta “Combine” de ArcGis.
Posteriormente se reclasificó el mapa, con la herramienta “reclassify” del “Spatial
Analyst” de ArcGis, para obtener las 19 categorías finales.
4. Asignación de densidad por hábitat
El mapa de densidades se elaboró uniendo la tabla de atributos del mapa de
hábitat con la hoja de calculos que contenía los valores de densidad por hábitat con
la herrmienta “join” del ArcGis.
5. Mapa de distribución (cuadriculas UTM)
Para crear el mapa de distribución en cuadriculas UTM de 1Km2 hizo falta
extraer la información sobre el área de hábitat que contiene cada cuadricula.
Previamente a esto se convirtió el mapa de hábitat a vectorial aplicando la
herramienta adecuada (herramientas de conversión de raster a shapefile) y
utilizando la herramienta “Dissolve” para unificar los polígonos de cada hábitat. Al
mapa vectorial se superpuso la malla de cuadriculas UTM 1Km2 y se aplicó la
herramienta “tabulate areas” (con tamaño de celda 10) para extraer los valores de
área necesarios. Entonces, se calculó la densidad por cuadricula multiplicando los
valores de densidades por hábitat con el área de hábitat por cuadricula y se adjuntó
a la malla de cuadriculas UTM utilizando de nuevo la herramienta “join”.