enzymes - univerzita karlovavyuka-data.lf3.cuni.cz/cvse1m0001/enzymes 21 10 2011...enzyme structure...
TRANSCRIPT

Enzymes
Josef Fontana
EC - 40

Overview of the seminar• Introduction to the topic: Enzymes
– Function of enzymes– Enzyme structure– Cofactors of enzyme groups– Izoenzymes and multienzyme complexes
• Nomenclature of enzymes• Regulation on the cell level
– 1) Compartmentalization of metabolic pathways– 2) Change of enzyme concentration (on the level of synthesis
of new enzyme)– 3) Change of enzyme activity (an existing enzyme is activated
or inactivated)– A) In relation to an enzyme kinetics– B) Activation or inactivation of the enzyme
• Enzymes in a medicine

Introduction to the topic: Enzymes
Function of enzymes
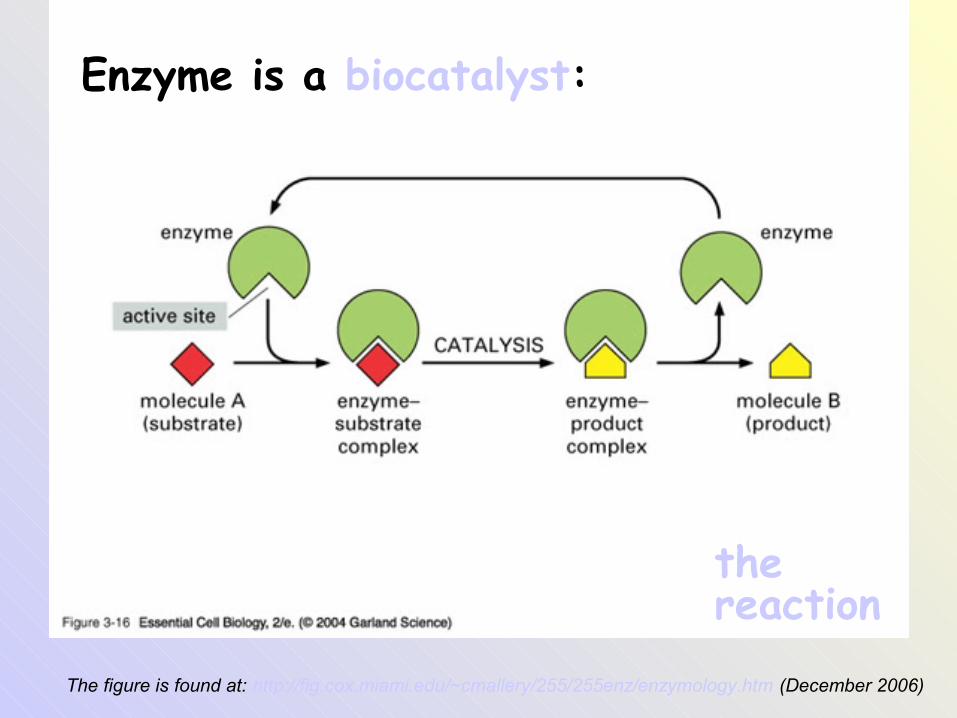
The figure is found at: http://fig.cox.miami.edu/~cmallery/255/255enz/enzymology.htm (December 2006)
Enzyme is a biocatalyst:
the reaction
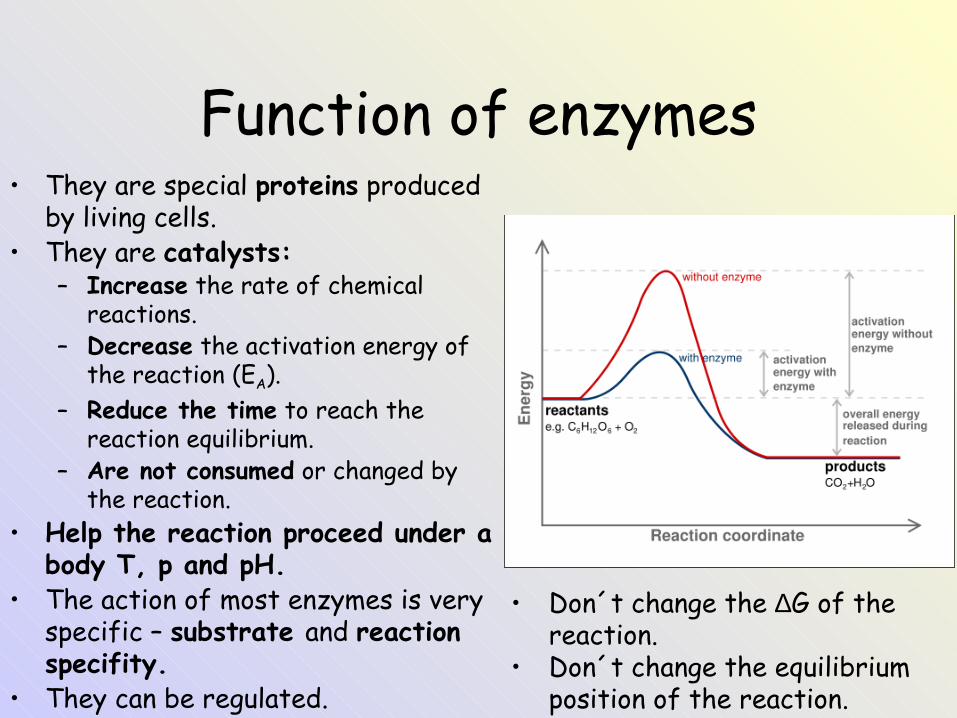
Function of enzymes• They are special proteins produced
by living cells.• They are catalysts:
– Increase the rate of chemical reactions.
– Decrease the activation energy of the reaction (EA).
– Reduce the time to reach the reaction equilibrium.
– Are not consumed or changed by the reaction.
• Help the reaction proceed under a body T, p and pH.
• The action of most enzymes is very specific – substrate and reaction specifity.
• They can be regulated.
• Don´t change the ∆G of the reaction.
• Don´t change the equilibrium position of the reaction.
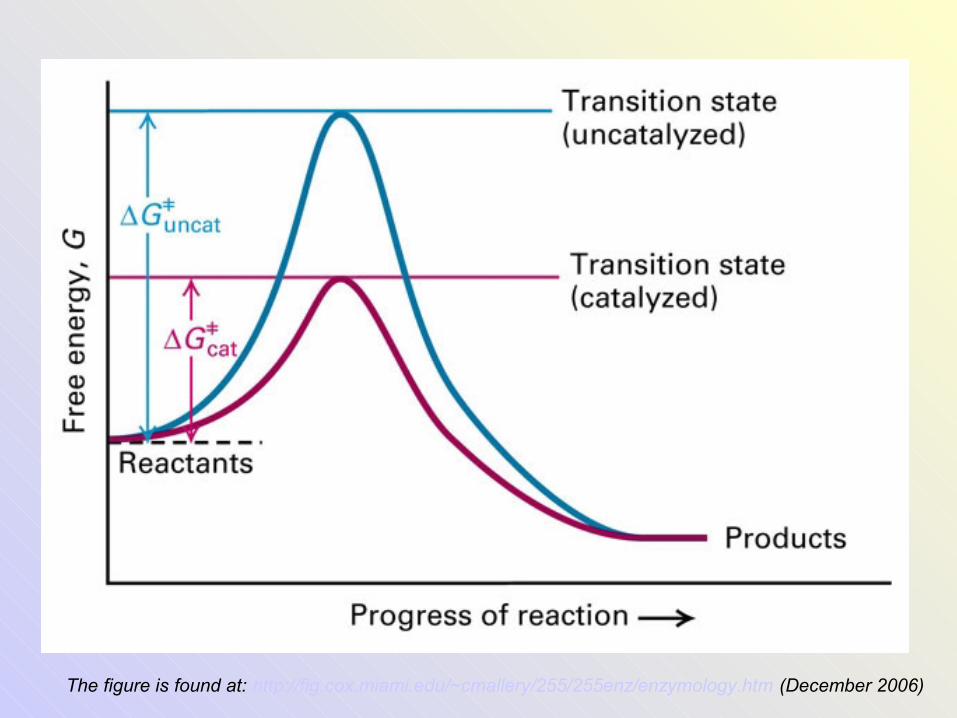
The figure is found at: http://fig.cox.miami.edu/~cmallery/255/255enz/enzymology.htm (December 2006)
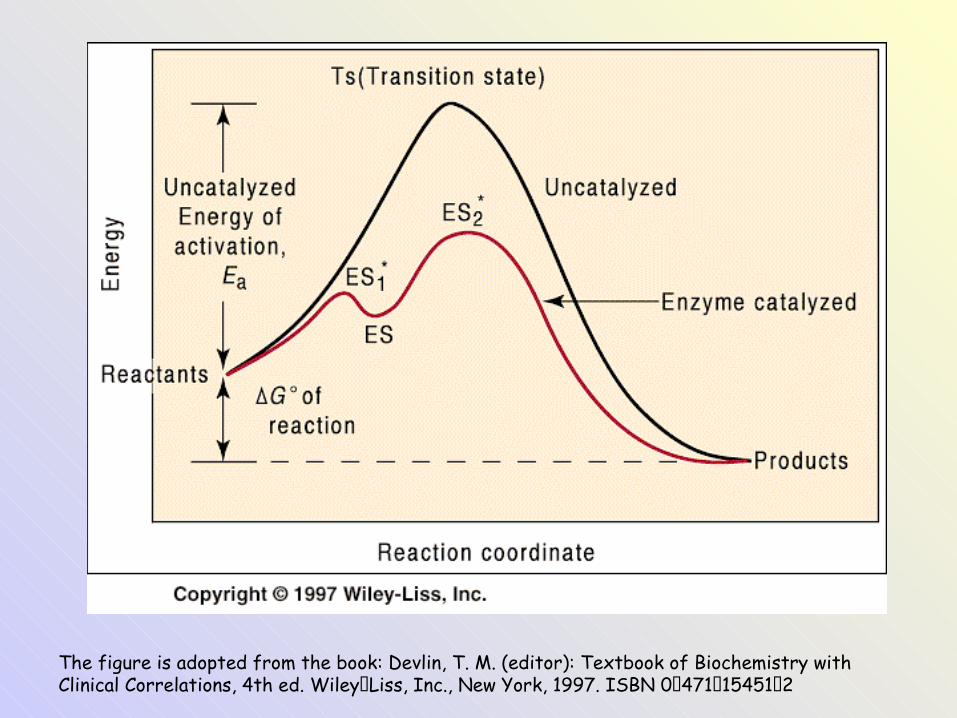
The figure is adopted from the book: Devlin, T. M. (editor): Textbook of Biochemistry with Clinical Correlations, 4th ed. Wiley‑Liss, Inc., New York, 1997. ISBN 0‑471‑15451‑2
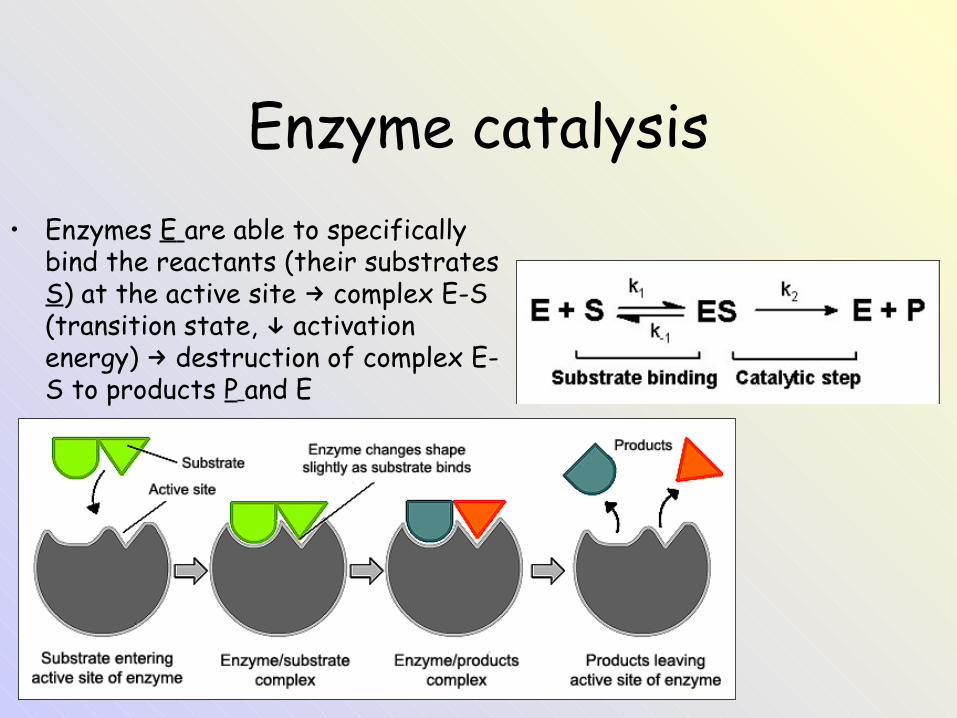
Enzyme catalysis• Enzymes E are able to specifically
bind the reactants (their substrates S) at the active site complex E-S →(transition state, activation ↓energy) destruction of complex E-→S to products P and E
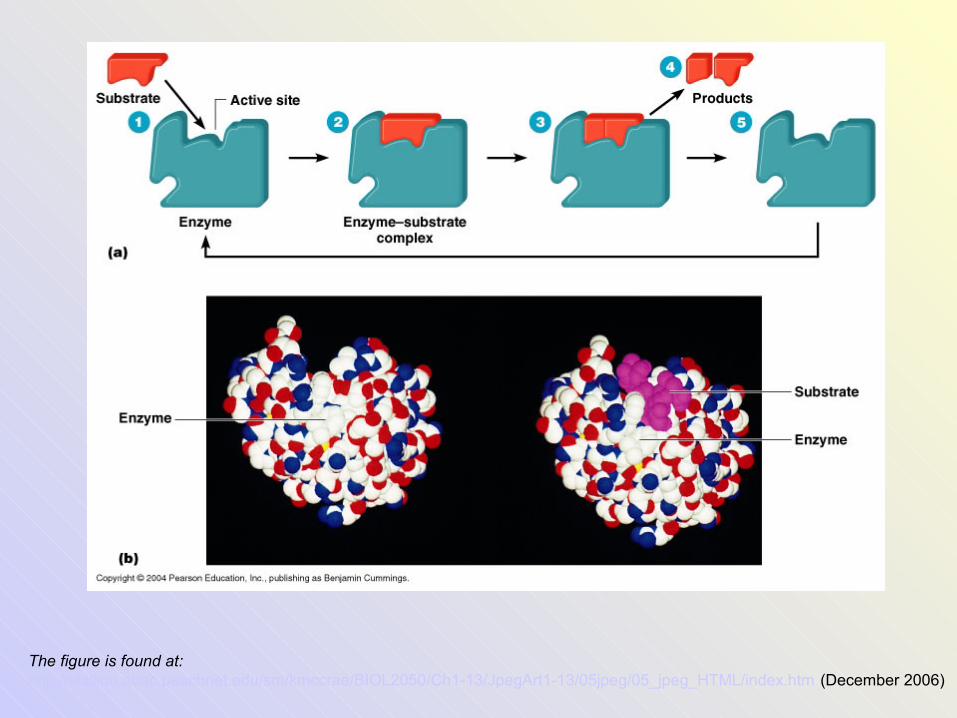
The figure is found at: http://stallion.abac.peachnet.edu/sm/kmccrae/BIOL2050/Ch1-13/JpegArt1-13/05jpeg/05_jpeg_HTML/index.htm (December 2006)

Introduction to the topic: Enzymes
Enzyme structure
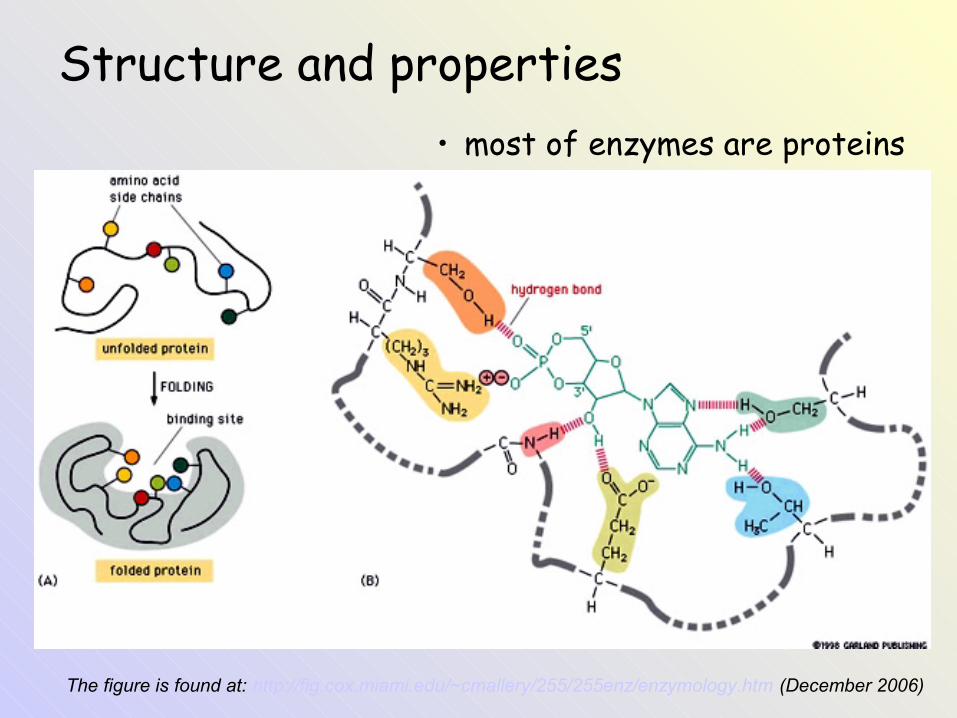
The figure is found at: http://fig.cox.miami.edu/~cmallery/255/255enz/enzymology.htm (December 2006)
Structure and properties• most of enzymes are proteins
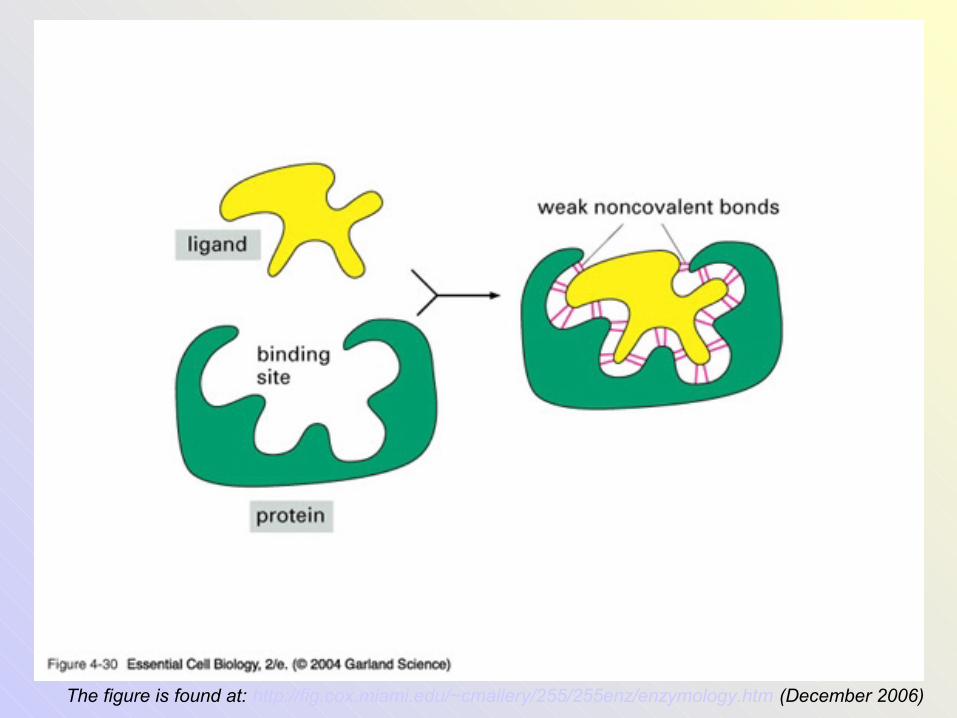
The figure is found at: http://fig.cox.miami.edu/~cmallery/255/255enz/enzymology.htm (December 2006)

Enzyme structure• Enzymes are mostly proteins (exception: ribozyme -
catalytic active RNA).• Some enzymes in addition to protein component
contain also non-protein component. According to this we can divide enzymes into:– Simple enzymes contain only protein (pepsin, trypsin,
ribonuclease).– Complex enzymes contain protein and non-protein
component = cofactor.• Cofactor is the non-protein part of the enzyme
molecule. It is necessary for its catalytic function. Our body can not synthesize them often - therefore, we eat their precursors – e.g. vitamins.

Cofactor can be• 1) metal ion: Zn2+, Mn2+, Mg2+, Fe2+, Cu2+ (trace
elements)• 2) organic molecule:
– coenzymes are slightly bound to the enzyme, undergo a chemical change and are released from the enzyme molecule, they are derivates of vitamins very often: NAD(P)+, FAD, coenzyme Q, ...
– prosthetic groups are tightly bound to the enzyme and remain associated with enzyme during the whole reaction: heme, FAD,…
• Coenzyme + apoenzyme (inactive protein) →holoenzyme (active enzyme)
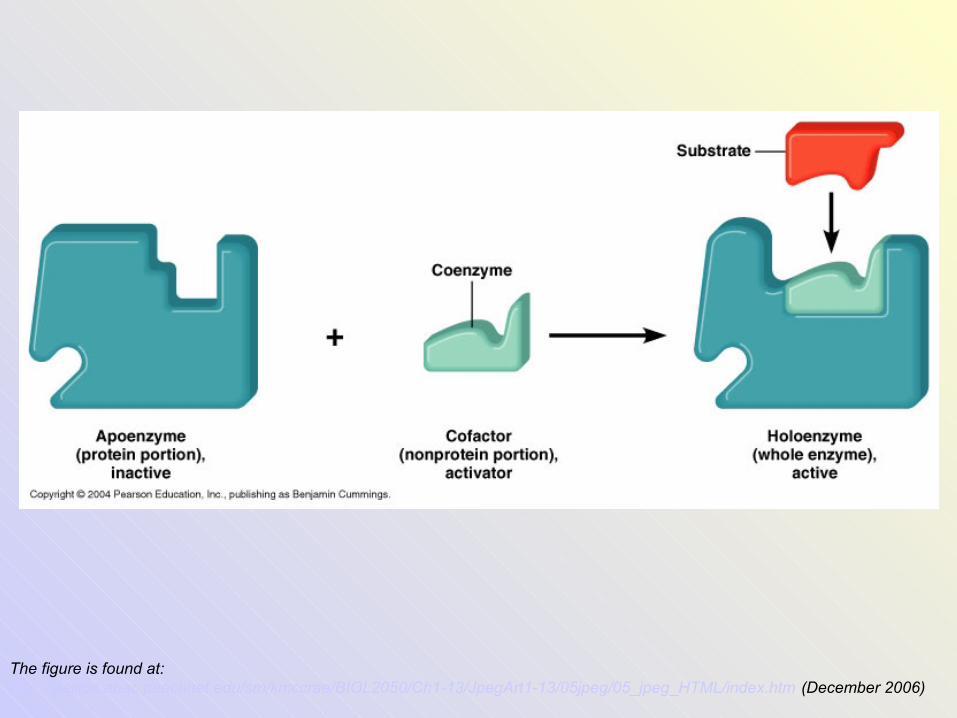
The figure is found at: http://stallion.abac.peachnet.edu/sm/kmccrae/BIOL2050/Ch1-13/JpegArt1-13/05jpeg/05_jpeg_HTML/index.htm (December 2006)

Introduction to the topic: Enzymes
Cofactors of enzyme groups

Cofactors help to catalyze many reactions
• Cofactors of oxidoreductases: NAD(P)+, FAD, cytochromes (contain heme), Fe-S complexes
• Coenzymes carrying C1 radicals: tetrahydrofolate, vitamin B12, S-adenosylmethionine, biotin (cofactor of carboxylases)
• Cofactors carrying acyl: lipoic acid (PDH prosthetic group, α-KGDH) HSCoA, pyridoxal phosphate (transaminases)

Cofactors of oxidoreductases
NAD+ nicotinamide adenine dinucleotideNADP+ nicotinamide aden. dinucl. phosphate(precursor: niacin = nicotinic acid) H+
FAD flavin adenine dinucleotideFMN flavin mononucleotide(precurzor: riboflavin = vitamin B2) 2 H+
heme Fe3+ + e- ↔ Fe2+ ⇒ e-

Cofactors of transferasesATP adenosine triphosphate / phosphateGTP guanosine triphosphate / phosphateTDP thiamine diphosphate / C-fragment(prekurzor: thiamine = vitamin B1)
PALP pyridoxal phosphate / -NH2
(prekurzor: pyridoxine = vitamin B6)
THF tetrahydrofolate / C1-fragment(prekurzor: folic acid)
CoA coenzyme A (HS-Co-A) / acylPAPS phosphoadenosine phosphosulfate / sulfate

Cofactors ofLyases:PALP pyridoxal phosphate (decarboxylases)
Ligases:ATP adenosine triphosphate
→ acyl-CoA-synthetases→ aminoacyl-tRNA-synthetases
biotin = vitamin H (carboxylases)
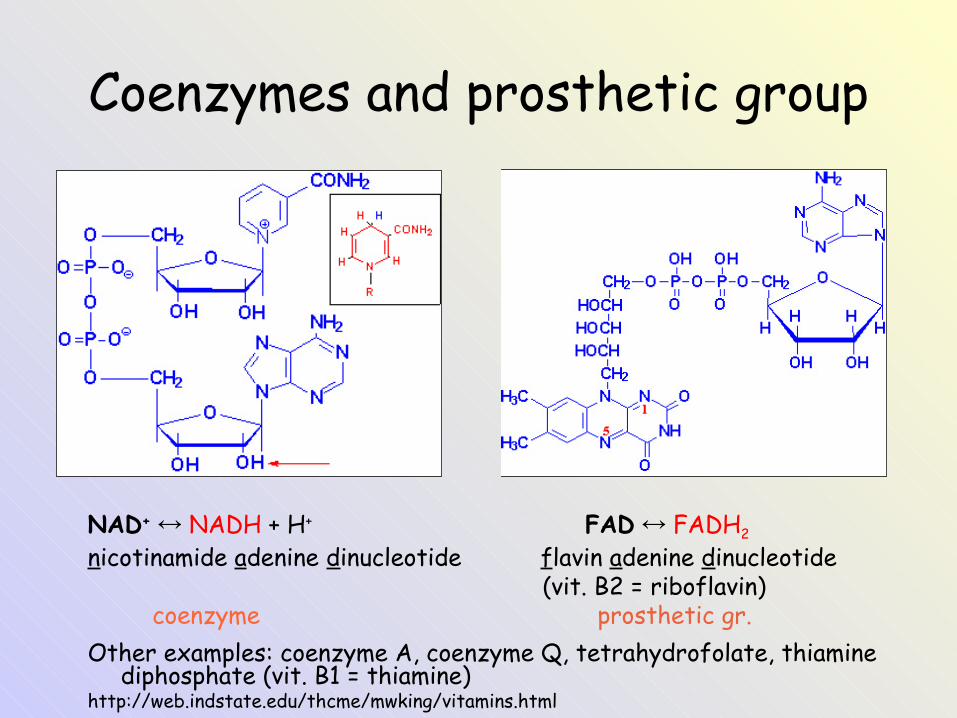
Coenzymes and prosthetic group
NAD+ ↔ NADH + H+ FAD ↔ FADH2
nicotinamide adenine dinucleotide flavin adenine dinucleotide (vit. B2 = riboflavin) coenzyme prosthetic gr.Other examples: coenzyme A, coenzyme Q, tetrahydrofolate, thiamine
diphosphate (vit. B1 = thiamine)http://web.indstate.edu/thcme/mwking/vitamins.html
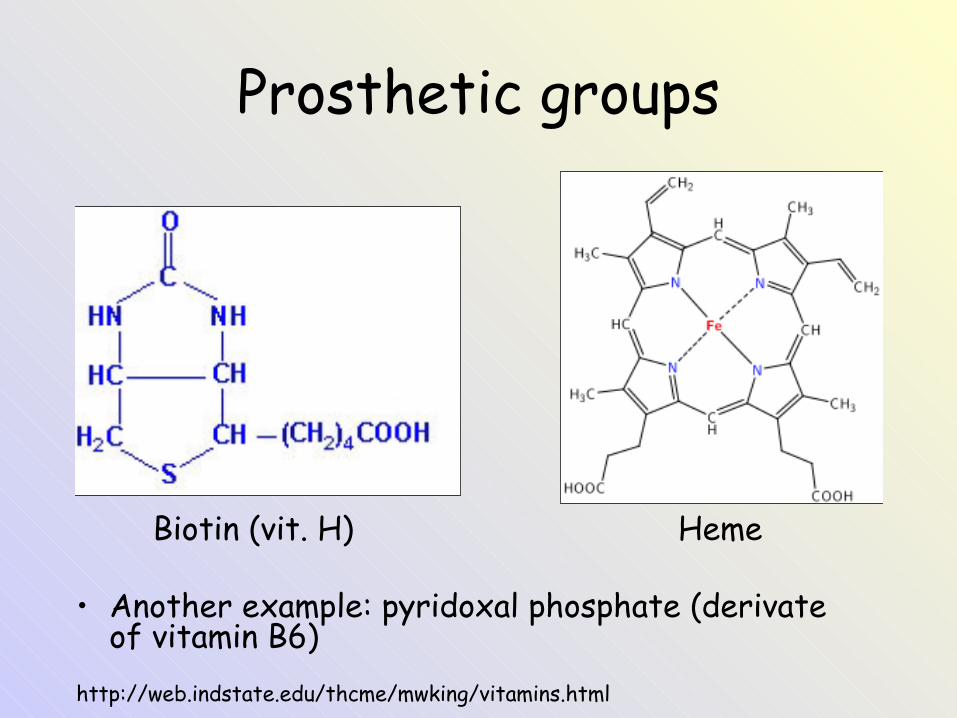
Prosthetic groups
Biotin (vit. H) Heme
• Another example: pyridoxal phosphate (derivate of vitamin B6)
http://web.indstate.edu/thcme/mwking/vitamins.html

Introduction to the topic: Enzymes
Izoenzymes and multienzyme complexes

Isoenzymes• Some enzymes have variants called isoenzymes. • They catalyze the same chemical reaction, but differ in
their primary structure and physico-chemical properties.• Isoenzymes are:
– Produced by different genes (= true isozymes) or– produced by different posttranslational modification (=
isoforms).– Found in different compartments of a cell.– Found in different tissues of an organism.
• For example lactate dehydrogenase (LD) has 5 isoenzymes: LD1 – LD5.
• Isoenzymes are present in skeletal muscle, liver, heart, kidney, erytrocytes. Isoenzymes can be separated by electrophoresis.
– Can be oligomers of various subunits (monomers).

The figure is found at: http://wine1.sb.fsu.edu/bch4053/Lecture26/isozymes.jpg (December 2006)
5 isozymes (various monomer ratio)
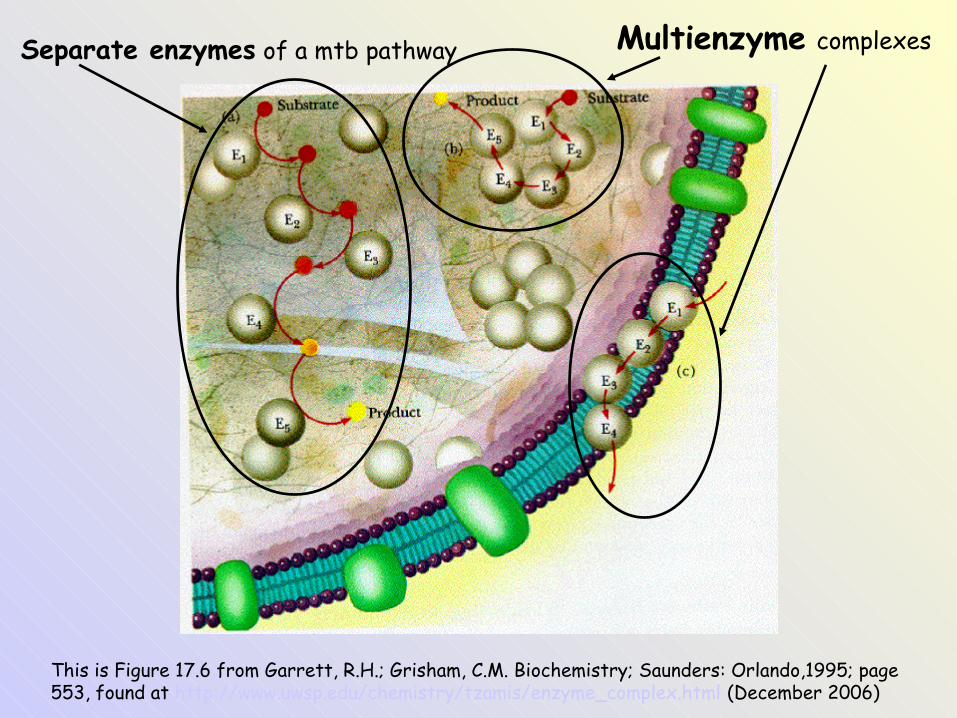
This is Figure 17.6 from Garrett, R.H.; Grisham, C.M. Biochemistry; Saunders: Orlando,1995; page 553, found at http://www.uwsp.edu/chemistry/tzamis/enzyme_complex.html (December 2006)
Multienzyme complexesSeparate enzymes of a mtb pathway
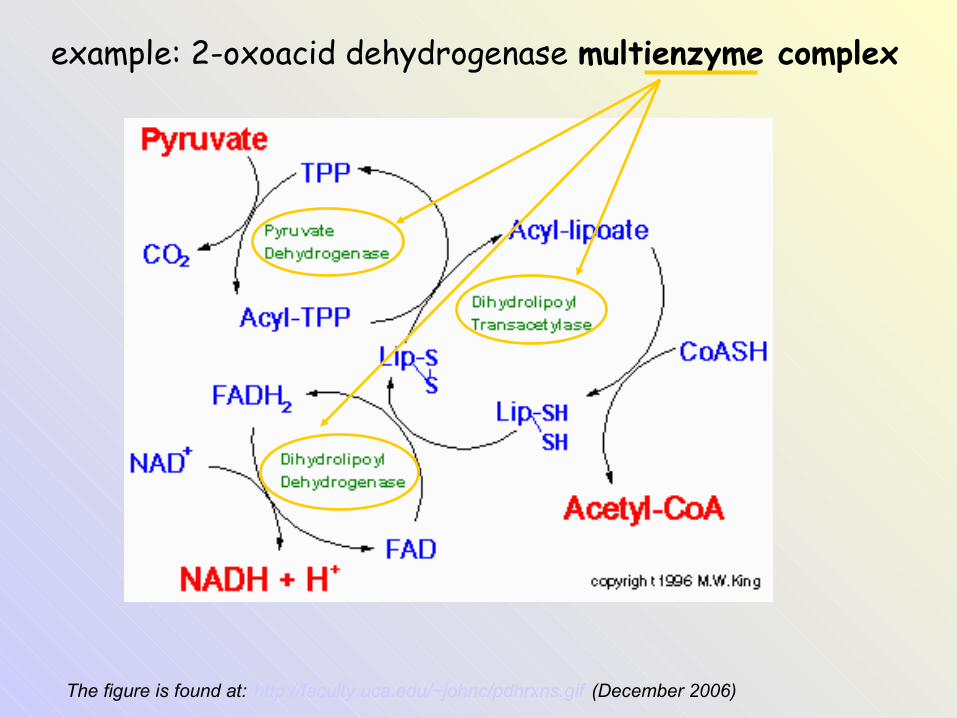
The figure is found at: http://faculty.uca.edu/~johnc/pdhrxns.gif (December 2006)
example: 2-oxoacid dehydrogenase multienzyme complex

Nomenclature of enzymes

Nomenclature of enzymes
• 1) The first discovered enzymes were named according to their source: Name of enzyme + suffix -in– Pepsin is found in the gastric juice (Greek
pepsis = digestion). • 2) Enzymes were named according to
their substrate: Name of substrate + specific suffix of enzymes –ase– Lipase catalyzes the hydrolysis of lipids.– Urease catalyzes the hydrolysis of urea.

Nomenclature of enzymes
• 3) In 1961 International Union of Biochemistry recommended that enzymes be systematically classified according to the general type of reaction they catalyze → 6 major classes.
• Abbreviations of enzymes e.g. LD, ALT, ALP

EC nomenclature
• Each enzyme has a EC number (four-digit number)• We have 6 main classes:• EC 1.x.x.x oxidoreductases• EC 2.x.x.x transferases• EC 3.x.x.x hydrolases• EC 4.x.x.x lyases• EC 5.x.x.x isomerases• EC 6.x.x.x ligases (synthetases)• Classification by a reaction catalyzed by the
enzyme• E.g. Lactate dehydrogenase has the EC number
1.1.1.27

Systematic names
• Are made according to a special rules, they specify a reaction catalyzed by the enzyme.
ATP : D-glucose phosphotransferase (EC 2.7.1.2)
→ transfers (2) phosphate (7) to an alcohol group (1)
ATP + D-Glc → ADP + D-Glc-6-phosphate (Glc-6-P)
Common name - glucokinase

EC nomenclature
• Oxidoreductases catalyze redox reactions:• dehydrogenase (H- or H)• reductase• oxidase• peroxidase (various peroxides)• oxygenase (O2)• hydroxylase (= monoxygenase; -OH)• desaturase (-CH2CH2- → -CH=CH-)• alcohol dehydrogenase, catalase

EC nomenclature
• Transferases catalyze the transfer of functional groups between donors and acceptors• grouptransferase (e.g. aminotransferase)• kinase (= phosphotransferase)• Phosphorylase• Transketolase, transaldolase

EC nomenclature
• Hydrolases catalyze the hydrolytic cleavage of substrates:• Esterase (R1-CO-O-R2)• Phosphatase (phosphate-O-R) → Pi !!!• phosphodiesterase (R1-O-phosphate-O-R2)• nuclease, peptidase, protease, glycosidase,
lipase, α-amylase

EC nomenclature
• Lyases (synthases) catalyze non-hydrolytic and non-oxidation cleavage or synthesis of molecules (removing/addition of the small molecule from/to substrate)– decarboxylase (→ CO2)– dehydratase (→ H2O)– hydratase (-CH=CH- + H2O →
-CH(OH)CH2-)– synthase

EC nomenclature
• Isomerases catalyze intramolecular changes in substrate molecules– epimerase (monosacharide → its epimer)– mutase (rearangement of a phosphate
group)

EC nomenclature
• Ligases (synthetases) catalyze synthetic reactions where 2 molecules are joined at 1 molecule, synthesis requires an energy (ATP)
• A + B + ATP → A-B + ADP + Pi
– Polymerases– synthetase– carboxylase

Regulation on the cell level

Regulation on the cell level
• 1) Compartmentalization of metabolic pathways
• 2) Change of enzyme concentration (on the level of synthesis of new enzyme)
• 3) Change of enzyme activity (an existing enzyme is activated or inactivated)

Regulation on the cell level
1) Compartmentalization of metabolic pathways

Compartmentalization of metabolic patways
• Transport processes between compartments• Various enzyme distribution• Various distribution of substrates and
products (∼ transport)• Transport of coenzymes• Subsequent processes are close to each other

Regulation on the cell level
2) Change of enzyme concentration (on the level of
synthesis of new enzyme)

Synthesis of new enzyme molecule
• Enzyme concentration is much lower than the concentration of substrate. The rate of an enzyme-catalyzed reaction is directly dependent upon the enzyme concentration.
• Induction by substrate or repression by product (on the level of transcription)– xenobiotics → induction of cyt P450– heme → repression of delta-aminolevulate synthase
• Change in the rate of synthesis or degradation of the enzyme - hormonal and nutritional factors– Well-fed state: the liver improves its capacity to
synthesize fat.– Fasting: ↓ in quantity of lipogenetic enzymes; enzymes of
gluconeogenesis are induced (↑synthesis).

Regulation on the cell level
3) Change of enzyme activity (an existing enzyme is
activated or inactivated)A) In relation to an enzyme kinetics
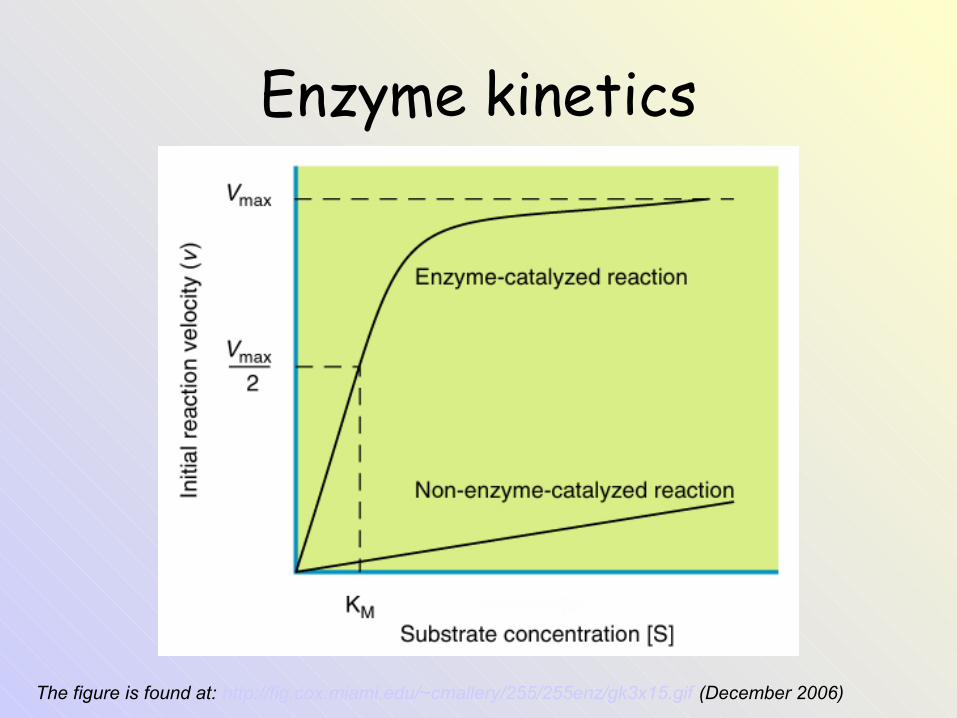
Enzyme kinetics
The figure is found at: http://fig.cox.miami.edu/~cmallery/255/255enz/gk3x15.gif (December 2006)

Enzyme kinetics - the curve can be described by the equation:
• Michaelis constant KM corresponds to the substrate concentration [S] at which velocity V is half of the maximum velocity Vmax (when v = ½ Vmax). An enzyme with a high affinity for its substrate has a low KM value.
• KM = mol/L
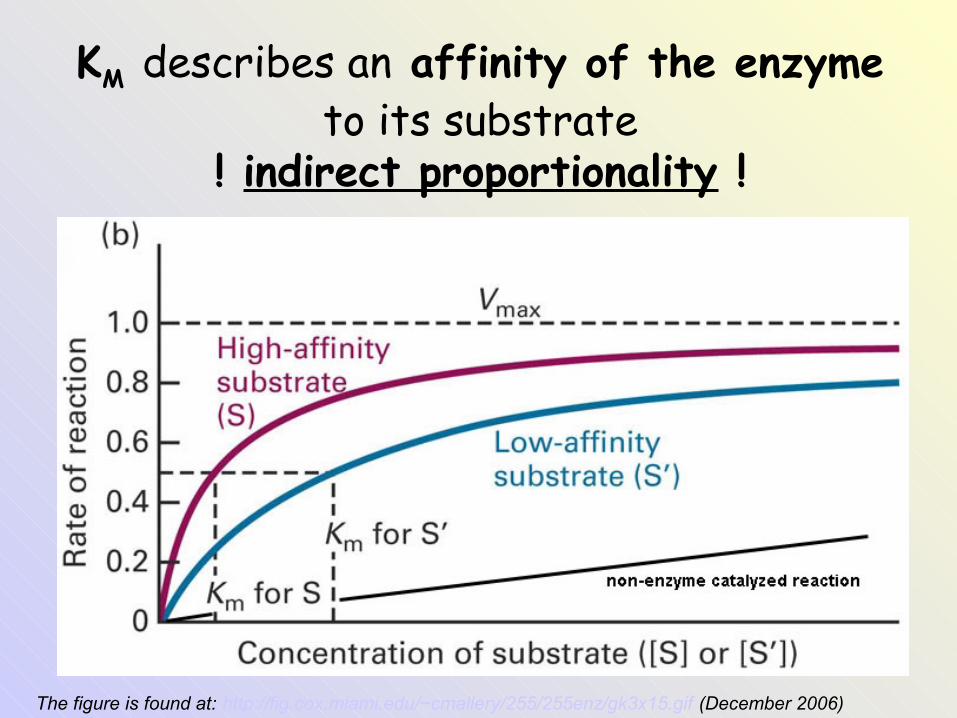
KM describes an affinity of the enzyme to its substrate
! indirect proportionality !
The figure is found at: http://fig.cox.miami.edu/~cmallery/255/255enz/gk3x15.gif (December 2006)

Change of activity of an existing enzyme
• A) In relation to an enzyme kinetics:– concentration of substrates (< Km)– pH and temperature changes– availability of coenzymes– consumption of products– substrate specificity - different Km
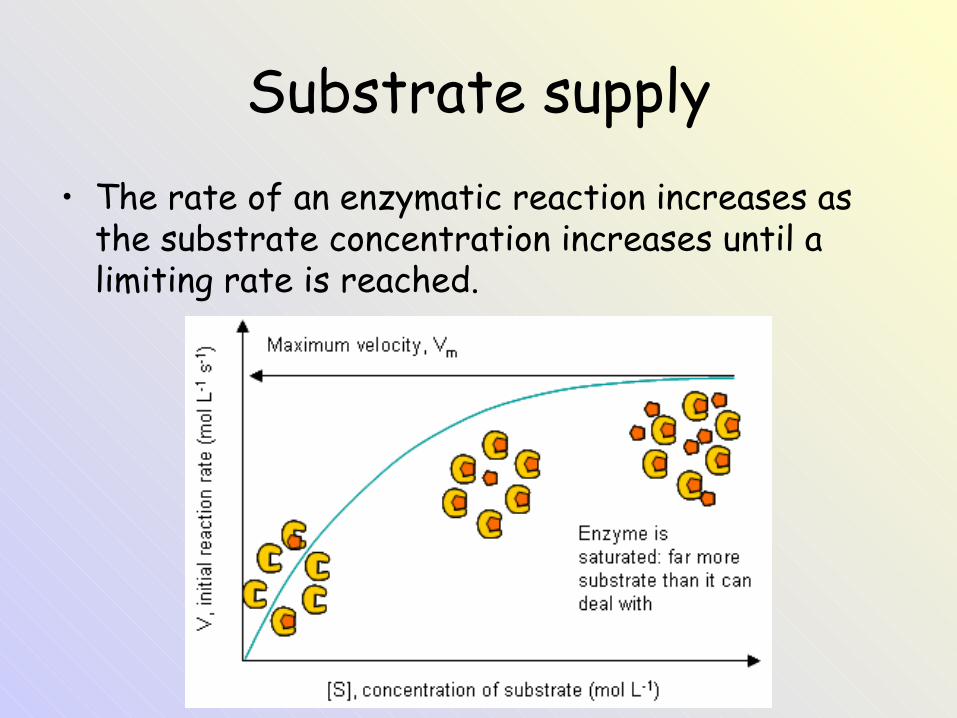
Substrate supply• The rate of an enzymatic reaction increases as
the substrate concentration increases until a limiting rate is reached.

Substrate supply• The rate of an enzymatic reaction increases as
the substrate concentration increases until a limiting rate is reached.
• Major determinant of the rate at which every metabolic processes of the body operates:
– blood fatty acids concentration → ketogenesis in the liver
– excessive amounts of substrates → synthesis of excess fat
– gluconeogenic substrates → rate of gluconeogenesis
– ↑ Gln → ↑citrulline → ↑urea synthesis
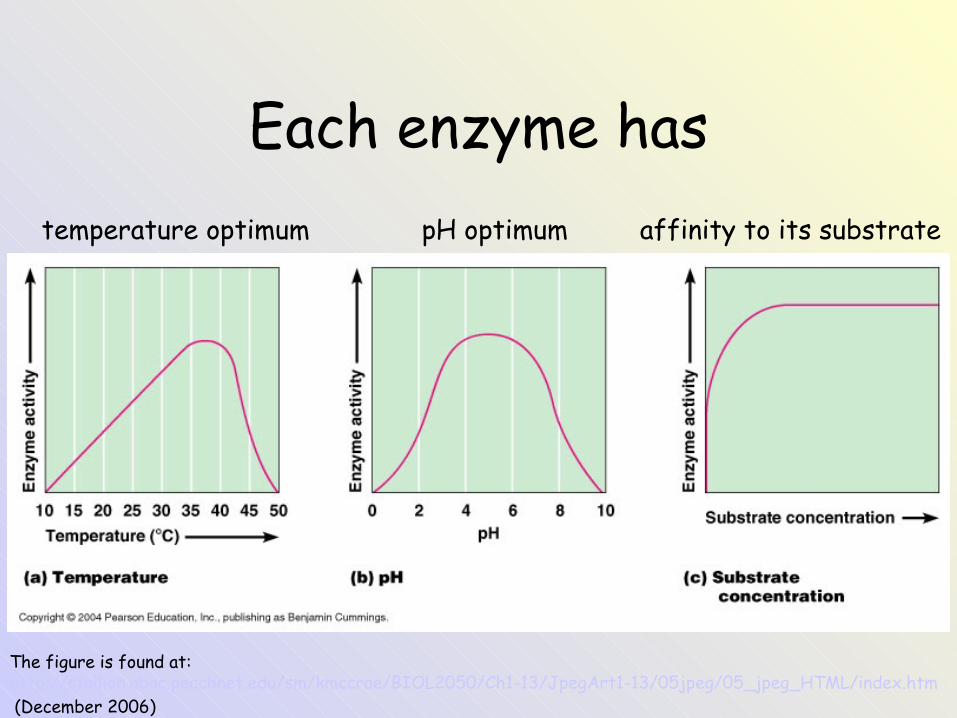
Each enzyme has temperature optimum pH optimum affinity to its substrate
The figure is found at: http://stallion.abac.peachnet.edu/sm/kmccrae/BIOL2050/Ch1-13/JpegArt1-13/05jpeg/05_jpeg_HTML/index.htm (December 2006)
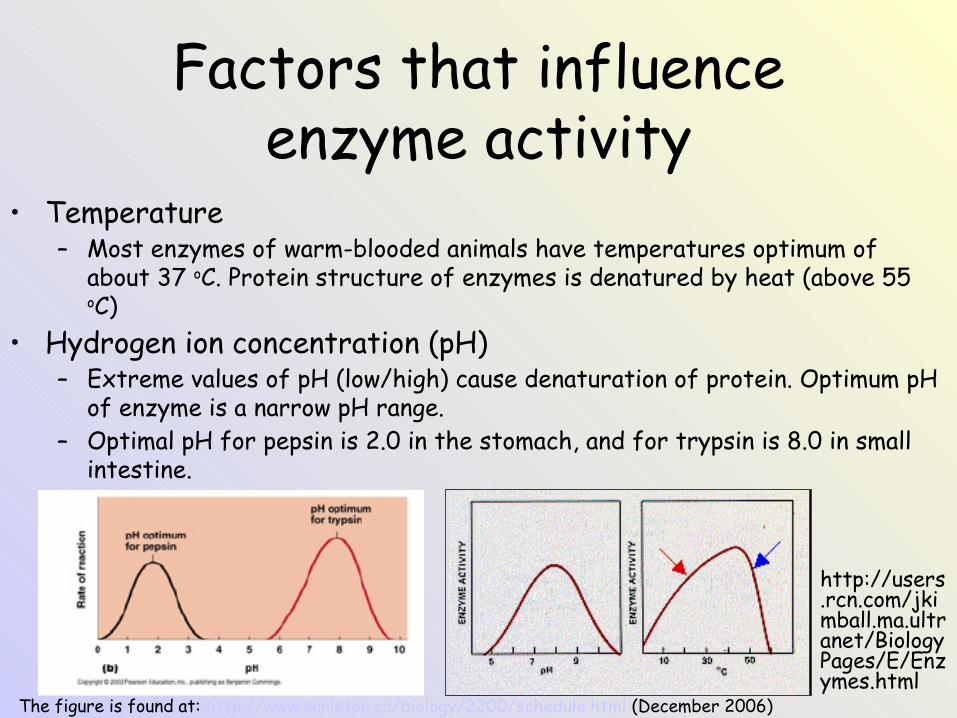
Factors that influence enzyme activity
• Temperature– Most enzymes of warm-blooded animals have temperatures optimum of
about 37 oC. Protein structure of enzymes is denatured by heat (above 55 oC)
• Hydrogen ion concentration (pH)– Extreme values of pH (low/high) cause denaturation of protein. Optimum pH
of enzyme is a narrow pH range.– Optimal pH for pepsin is 2.0 in the stomach, and for trypsin is 8.0 in small
intestine.
http://users.rcn.com/jkimball.ma.ultranet/BiologyPages/E/Enzymes.html
The figure is found at: http://www.carleton.ca/biology/2200/schedule.html (December 2006)

Regulation on the cell level
3) Change of enzyme activity (an existing enzyme is
activated or inactivated)B) Activation or inactivation of the enzyme

Change of activity of an existing enzyme
• B) Activation or inactivation of the enzyme:• Covalent modification of the enzymes
– cleavage of an precursore (proenzyme, zymogen)– reversible phosphorylation and dephosphorylation
(interconversion of enzymes by protein kinase or protein phosphatase respectively)
• Modulation of activity by modulators (ligands):– feed back inhibition– cross regulation– feed forward activation
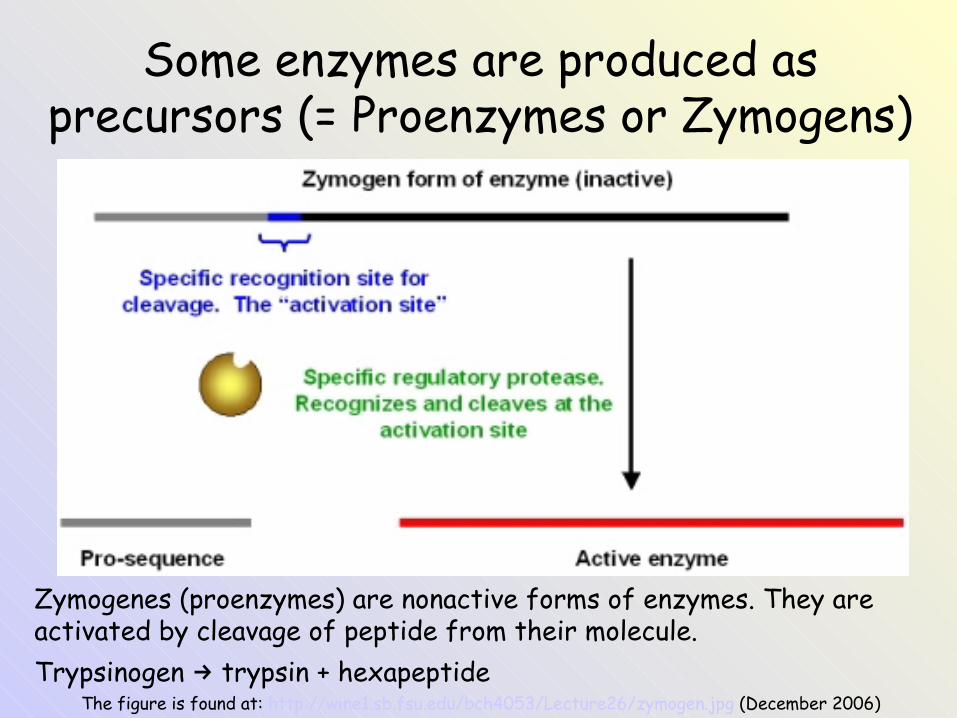
Some enzymes are produced as precursors (= Proenzymes or Zymogens)
The figure is found at: http://wine1.sb.fsu.edu/bch4053/Lecture26/zymogen.jpg (December 2006)
Zymogenes (proenzymes) are nonactive forms of enzymes. They are activated by cleavage of peptide from their molecule.Trypsinogen trypsin + hexapeptide→
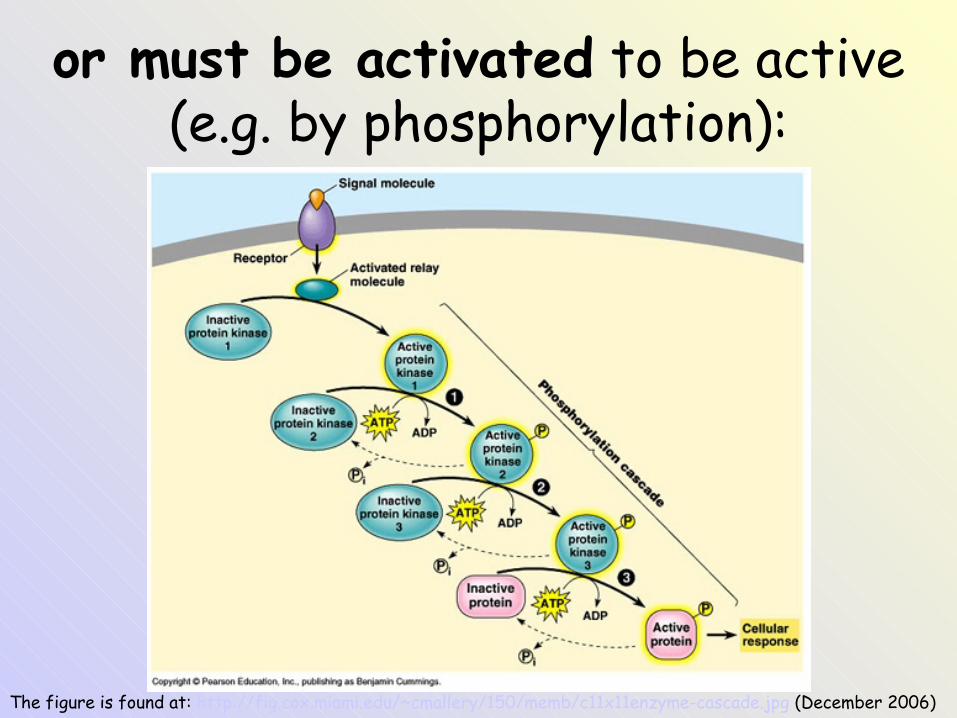
The figure is found at: http://fig.cox.miami.edu/~cmallery/150/memb/c11x11enzyme-cascade.jpg (December 2006)
or must be activated to be active (e.g. by phosphorylation):

Phosphorylation / dephosphorylation
• Some enzymes are active in a phosphorylated form, some are inactive.
• Phosphorylation– protein kinases– macroergic phosphate as a donor of the
phosphate (ATP !)
• Dephosphorylation– protein phosphatase– inorganic phosphate is the product !
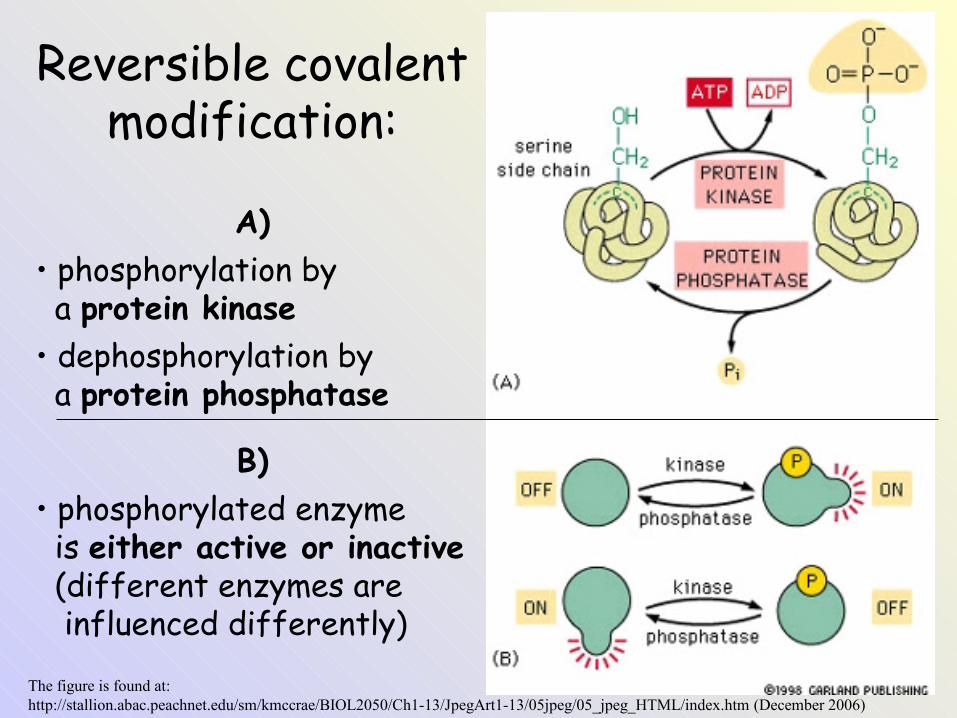
The figure is found at:http://stallion.abac.peachnet.edu/sm/kmccrae/BIOL2050/Ch1-13/JpegArt1-13/05jpeg/05_jpeg_HTML/index.htm (December 2006)
Reversible covalent modification:
A)• phosphorylation by a protein kinase• dephosphorylation by a protein phosphatase
B)• phosphorylated enzyme is either active or inactive (different enzymes are influenced differently)

Modulators of enzyme activity(activators, inhibitors)
• Isosteric modulation: competitive inhibition
• Allosteric modulation:– change of Km or Vmax
– T-form (less active) or R-form (more active)
• Important modulators: ATP / ADP, NAD+ / NADH + H+

Inhibitors• Some chemical compounds can act as enzyme
inhibitors. Enzyme inhibition:– a) irreversible– b) reversible
• Irreversible inhibition– Irreversible inhibitors react with enzyme and form a
covalent adduct with protein or metal ion.– HCN inactives iron-containing enzymes because it
binds to Fe2+ in heme. HCN blocks cellular respiration (cytochrome c oxidase).
– The nerve gases inhibit transmission in nerve system because they block specific enzymes (tabun, sarin).
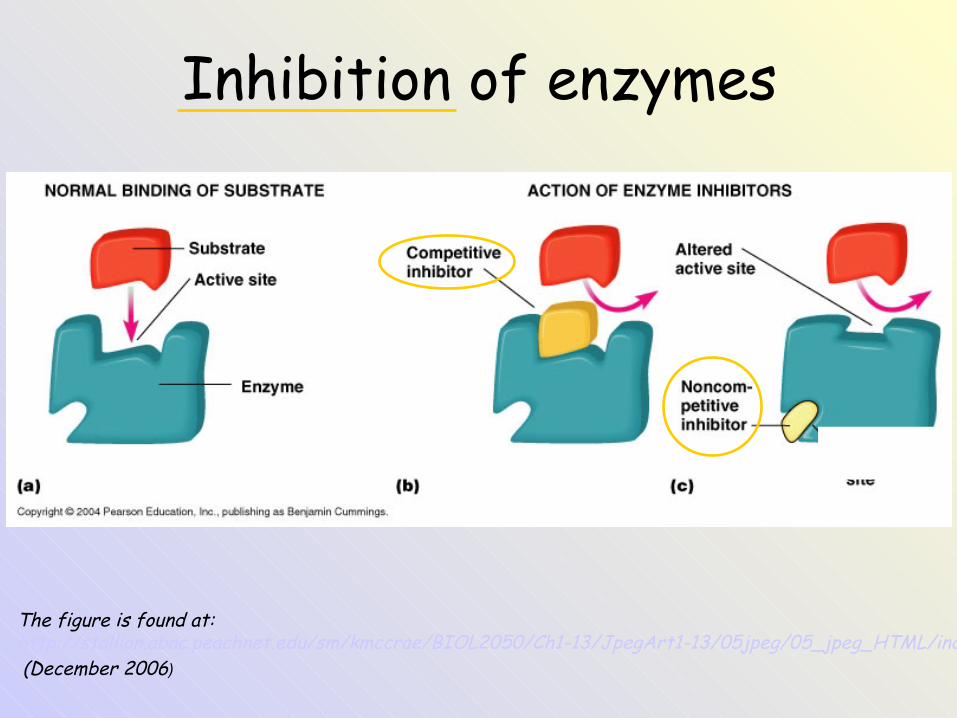
The figure is found at: http://stallion.abac.peachnet.edu/sm/kmccrae/BIOL2050/Ch1-13/JpegArt1-13/05jpeg/05_jpeg_HTML/index.htm (December 2006)
Inhibition of enzymes
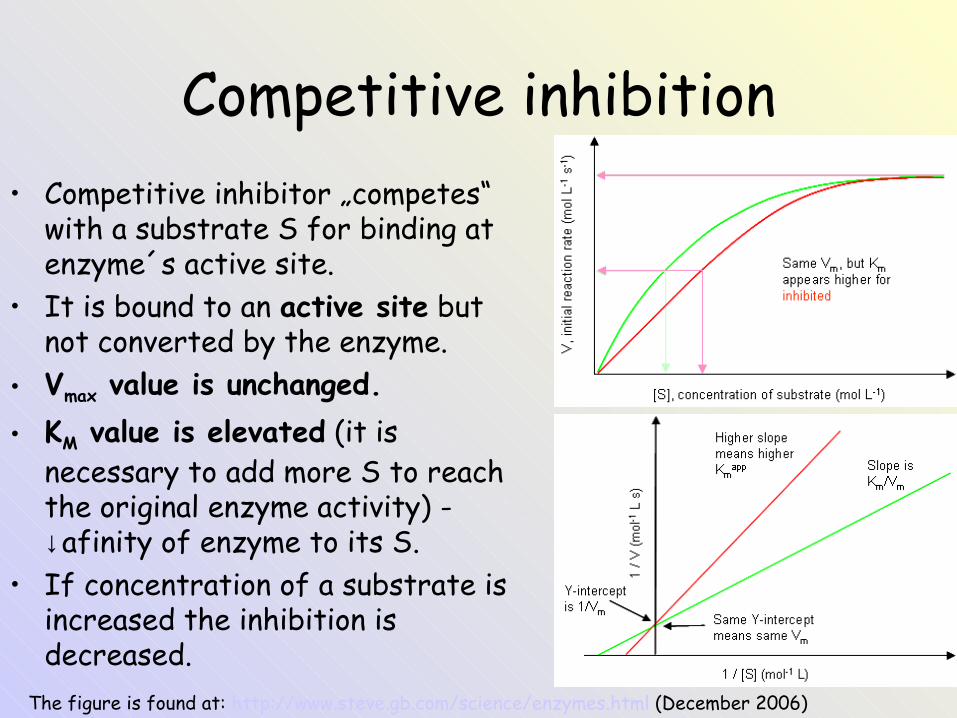
Competitive inhibition• Competitive inhibitor „competes“
with a substrate S for binding at enzyme´s active site.
• It is bound to an active site but not converted by the enzyme.
• Vmax value is unchanged.• KM value is elevated (it is
necessary to add more S to reach the original enzyme activity) - ↓afinity of enzyme to its S.
• If concentration of a substrate is increased the inhibition is decreased.
The figure is found at: http://www.steve.gb.com/science/enzymes.html (December 2006)
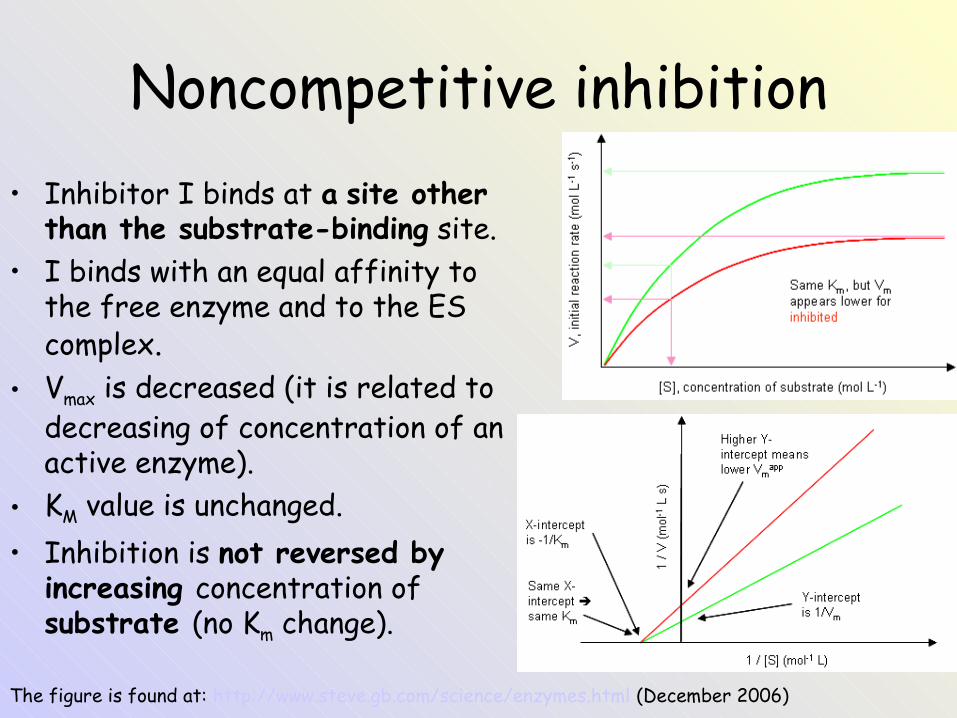
Noncompetitive inhibition• Inhibitor I binds at a site other
than the substrate-binding site.• I binds with an equal affinity to
the free enzyme and to the ES complex.
• Vmax is decreased (it is related to decreasing of concentration of an active enzyme).
• KM value is unchanged.• Inhibition is not reversed by
increasing concentration of substrate (no Km change).
The figure is found at: http://www.steve.gb.com/science/enzymes.html (December 2006)
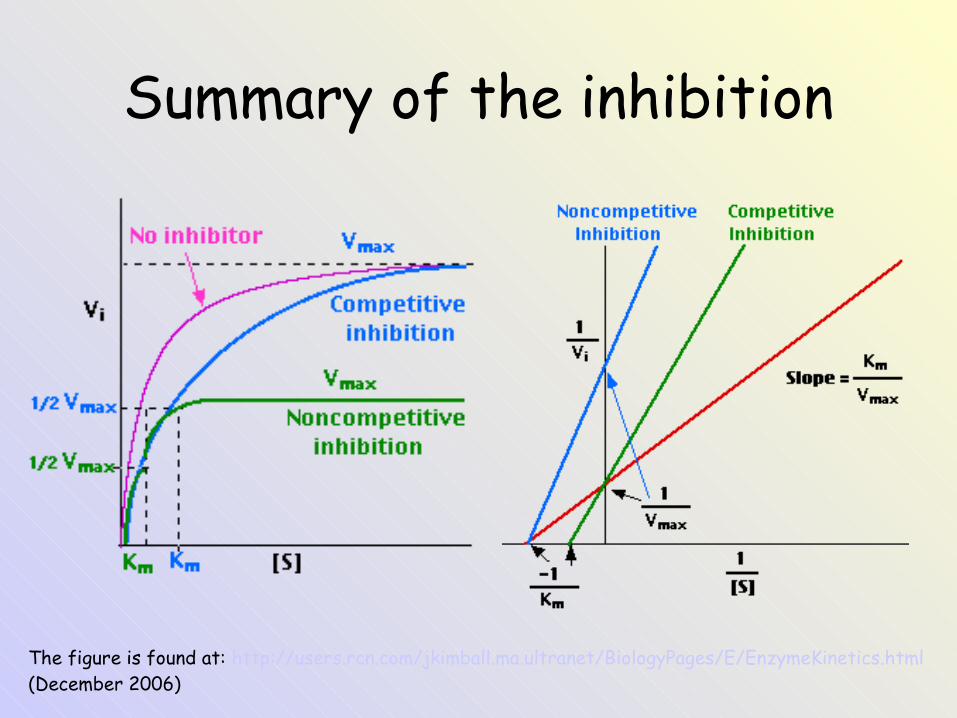
Summary of the inhibition
The figure is found at: http://users.rcn.com/jkimball.ma.ultranet/BiologyPages/E/EnzymeKinetics.html (December 2006)

Allosteric enzyme: a) monomeric, b) oligomeric
The figure is adopted from the book: Devlin, T. M. (editor): Textbook of Biochemistry with Clinical Correlations, 4th ed. Wiley‑Liss, Inc., New York, 1997. ISBN 0‑471‑15451‑2
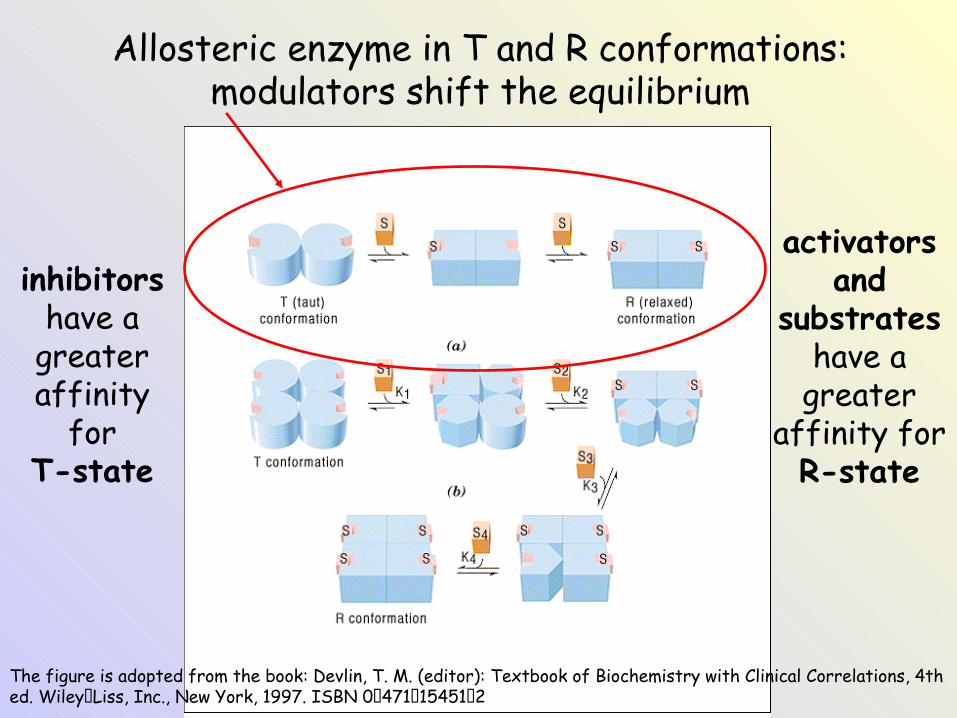
Allosteric enzyme in T and R conformations: modulators shift the equilibrium
inhibitors have a
greater affinity
forT-state
activators and
substrates have a
greater affinity forR-state
The figure is adopted from the book: Devlin, T. M. (editor): Textbook of Biochemistry with Clinical Correlations, 4th ed. Wiley‑Liss, Inc., New York, 1997. ISBN 0‑471‑15451‑2
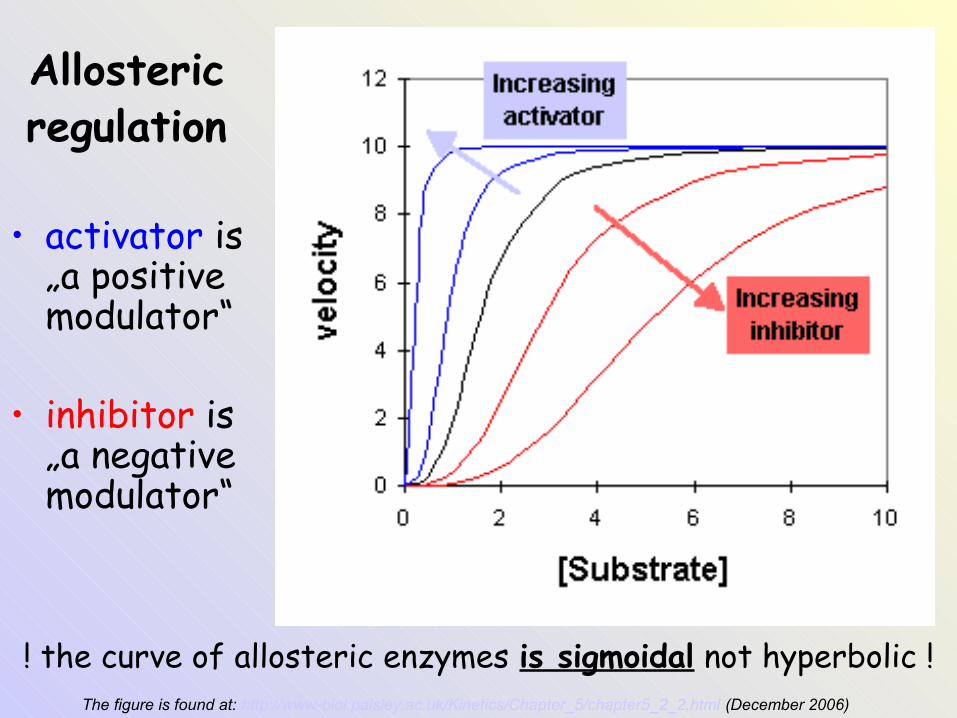
Allostericregulation
• activator is „a positive modulator“
• inhibitor is „a negative modulator“
The figure is found at: http://www-biol.paisley.ac.uk/Kinetics/Chapter_5/chapter5_2_2.html (December 2006)
! the curve of allosteric enzymes is sigmoidal not hyperbolic !

Allosteric effectors (negative or positive)
• ↑glucose: inhibits glycogen phosphorylase, activates glycogen synthase (= mtb of glycogen)
• ↑fructose-2,6-bisphosphate (↑ if insulin is ↑): inhibits fru-1,6-bisphosphatase (= gluconeogenesis), activates 6-PFK-1 (= glycolysis)
• ↑citrate: inhibits 6-PFK-1 (= glycolysis), activates acetyl-CoA carboxylase (= fatty acid synthesis)
• ↑acetyl-CoA: inhibits pyruvate dehydrogenase, activates pyruvate carboxylase (= activation of gluconeogenesis)
• ↑malonyl-CoA inhibits carnitine palmitoyl transferase I (= β-oxidation)
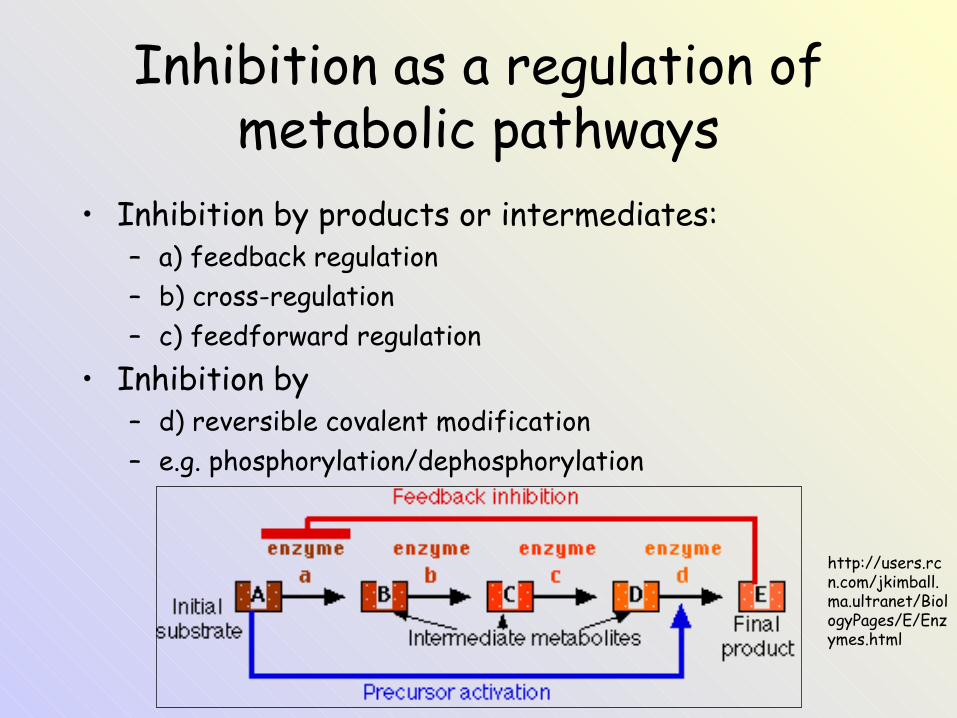
Inhibition as a regulation of metabolic pathways
• Inhibition by products or intermediates:– a) feedback regulation– b) cross-regulation – c) feedforward regulation
• Inhibition by– d) reversible covalent modification – e.g. phosphorylation/dephosphorylation
http://users.rcn.com/jkimball.ma.ultranet/BiologyPages/E/Enzymes.html

Enzymes in a medicine

Enzymes in a medicine
• Determination of enzyme activity in blood.
• Enzymatic analytical methods.• Enzyme therapy.

Diagnostic applications of enzymes
• The measurement of enzyme activity in body fluids (plasma, serum) has become an important tool in medical diagnosis. Under normal conditions the concentrations of enzymes is low in blood. An abnormally high level of a particular enzyme in the blood often indicates specific tissue damage (hepatitis, myocardial infarction,....). Some important enzymes for clinical diagnosis:
Enzyme assayed Organ or tissue damaged
α-amylase (AMS) pancreas
alkaline phosphatase (ALP) bone, liver
creatine kinase (CK) muscle, heart
lactate dehydrogenase (LD) heart, liver
alanine aminotransferase (ALT) liver
aspartate aminotransferase (AST) heart, liver

Enzymatic activity
• Units– 1 katal = 1 mole of a substrate transformed per 1 sec
– 1 IU = 1 μmole of a substrate transformed per 1 minute
• 1 katal = 1 mole / 1 sec• = 106 μmole /1 sec• = 60 x 106 μmole /1 min (= 60 sec)• 1 katal = 6 x 107 IU

Diagnostic applications of enzymes
• Normal (physiological) activity of ALT in blood is up to 0.73 µkat/L.
• Activity of ALT in serum during acute virus hepatitis is 50x higher than normal activity!
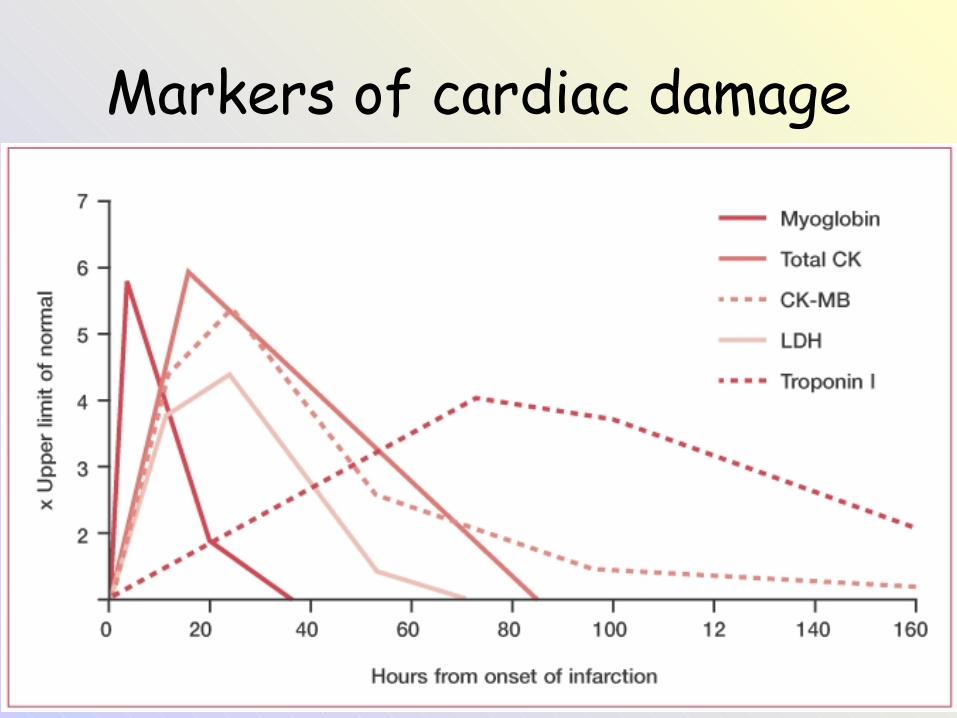
Markers of cardiac damage
• Při poškození buňěk myokardu se původně intracelulární látky dostávají do oběhu a my je tak můžeme stanovit při odběru krve.
• Stanovují se následující látky:• Kreatinkináza (CK) a její izoenzym CK-MB• Troponin T nebo troponin I = výborné markery
poškození myokardu, jsou totiž odlišné od troponinů v kosterní svalovině (>99% specifita pro myokard)
• Dnes už jen historický význam mají AST a myoglobin.

SummaryRegulation of enzyme activity
• availability of a substrate and its concentration
• induction of synthesis of a regulatory enzyme
• activation of enzyme precursors• covalent modification of enzymes• competitive inhibition• allosteric regulation