enzymes structure and kinetics - medcol.mwcms.medcol.mw/cms_uploaded_resources/20235_1.pdf · for...
TRANSCRIPT


1. Define Enzymes
2. Know enzyme models
3. Understand principles of enzyme kinetics
4. Know how enzymes are inhibited
5. Understand the Michaelis-Menten principle and the Linewaiver-Burk Plots
References:1. Lenhinger chapter 8: page 198 - 236
Learning Objectives.......

1. A catalyst lowers energy of activation by providing a different mechanism for the reaction. Both the rates of forward and backward reaction are enhanced.
GENERAL PRINCIPLES OF CATALYSIS

2. A catalyst forms an intermediate with the reactant(s) in the initial step of the mechanism and is released in the product forming step.
3. A catalyst does not affect the enthalpies or free energies of reactants and products.

1. Homogeneous Catalysis – reactants and catalysts are in the same phase (e.g. solid rxnts, solid catalyst)
2. Heterogeneous Catalysis – reactants and catalysts are in different phases (liquid rxnts, solid catalyst)
3. Enzyme Catalysis – also homogeneous catalysis but catalysts are biological in origin. More complex.
Three Types of Catalysis

Consider the reaction:
Relative rateUncatalyzed: 1
Pt Black (inorganic catalyst): 10,000
catalase (enzyme): 300,000,000,000
Catalysts, in particular, enzymes are capable of astonishing rate enhancements
What sort of acceleration can catalysts provide?

• Enzymes form complex 3D structures that present an “active site” surface to which reactants bind in a chemical reaction.
• Active sites position amino acid R-groups and/or reaction co-factors (such as metals) or prosthetic groups at the appropriate positions to aid in catalysis.
• Two major models for how this might work on the structural level.
How do enzymes work?

TWO MODELS FOR THE ES COMPLEX
Lock - and- Key Model
Induced Fit Model

Mg(2+)Mg(2+)
ATPExample of a real Active Site

[P]
time
[S]
time
Accumulation of product over time (∆[P]/∆t) Loss of substrate over time (∆[S]/∆t)
ENZYME ACTIVITY MEASUREMENT

How does [enzyme] influence observed reaction velocity?
[P]
time
Assumes that [E] is limiting and that the uncatalyzed reaction rate is ~0
1 x [enzyme]∆[P]/∆t = 1
2 x [enzyme]∆[P]/∆t = 2
0.5 x [enzyme]∆[P]/∆t = 0.5

How specific are enzymes for a given substrate?
• Most enzymes, such as arginase, are highly specific, acting on only a small number of substrates that are highly similar in structure. Others, such as alkaline phosphatase, are less specific.
• Specificity arises from structural and chemical complementarity between the substrate and its enzyme (active site).
ENZYME SPECIFICITY

Specificity of enzymes (an example)
Asp with Mg(2+),Lys with Phosphates
Mg (2+)
HydrogenBonds
Gln withAdenine
IonicBonds

Metals, coenzymes, and prosthetics groups
Many enzymes bind non-protein cellular components that are used as key factors in the enzyme activity. These fall into three basic categories:
1.Metals: Metals like Mg, Ca, Zn, Fe etc.) are thought to be bound to ~1/3 of all proteins and can play key roles in activity. An example is the Mg(2+) in the ATPase on the previous slide. These ions can confer a wider array of chemical properties to proteins over those of the 20 natural amino acids.

Metals, cofactors, and prosthetics groups
(2 & 3) Coenzymes and prosthetic groups: Low-molecular organic compounds that bind either weakly (coenzymes) or tightly (prosthetic groups) to the protein. Examples that you will see in this course include, for example, iron-sulfur clusters, heme, and coenzyme A.

Formula for a simple enzyme-catalyzedreaction
E + S ES P + E
E - free enzymeS - SubstrateES - Enzyme-Substrate complexP - product

What are we measuring?
Incr
easi
ng [
S]

Initial Velocity
Measured at thevery beginning ofa reaction whenvery little P hasbeen made.

FOR ENZYME-CATALYZED REACTION
E + S ES P + E
k1 is rate constant for formation of ESk-1 is rate constant for conversion of ES to E+Sk2 is rate constant for product formation. Forthis reaction, k2= kcat
Initial velocity assumption: measure activity beforeappreciable P accumulates: v0 = k2 [ES]

ENZYME-CATALYZED REACTION EXHIBIT SATURATION KINETICS
E + S ES P + E
At high [S], theenzyme is said tobe saturated withrespect tosubstrate
[ES]
[S]

STEADY STATE
The more ES present, the faster ES will dissociate into E + P or E + S.Therefore, when the reaction isstarted by mixing enzymes andsubstrates, the [ES] builds up atfirst, but quickly reaches aSTEADY STATE, in which [ES]remains constant. This steady state will persist until almost all of the substrate has been consumed.

THE MICHAELIS-MENTEN EQUATION
E + S ES P + E
If you assume that the formation of ES equals its breakdown, making [ES] constant (steady state), then:
k1 [E][S] = k-1 [ES] + k2 [ES]
Vmax = maximum velocity where [ES] = [Et]

Important Conclusions of Michaels - Menten Kinetics
• when [S]= KM, the equation reduces to
• when [S] >> KM, the equation reduces to
• when [S] << KM, the equation reduces to

Important Conclusions of Michaels - Menten Kinetics

Bi-substrate Reactions
• The Michaelis –Menten model of enzyme kinetics was derived for single substrate reactions
• The majority of enzymatic reactions have multiple substrates and products
• Bi-substrate reactions account for ~ 60% of the known enzymatic reactions.

Substrate Addition / Product Release
• The order of substrate addition and product release in most enzymatic reactions follow two reaction mechanism
– Sequential reaction - all substrates must bind to the enzyme before the reaction occurs and products are released
• Ordered sequential • Random sequential – Ping-pong reaction - one or more products are released before all
substrates have been added and an alternate stable enzyme form, F, is produced in the half reaction

1) Sequential Reaction• Ordered sequential
• Random sequential

2) Ping-pong Reaction

Lineweaver-Burk Plot
Also known as the double-reciprocol plot, it provides a useful graphical method for analysis of the Michaelis–Menten equation:
where V is the reaction velocity (the reaction rate), Km is the Michaelis–Menten constant, Vmax is the maximum reaction velocity, and [S] is the substrate concentration.

Use of the Lineweaver-Burk Plot
1. The Lineweaver–Burk plot was used to determine terms in enzyme kinetics such as Km and Vmax .
2. The y-intercept of such a graph is equivalent to the inverse of Vmax ;
3. the x-intercept represents −1/Km.
4. It also gives a quick, visual impression of the different forms of enzyme inhibition.
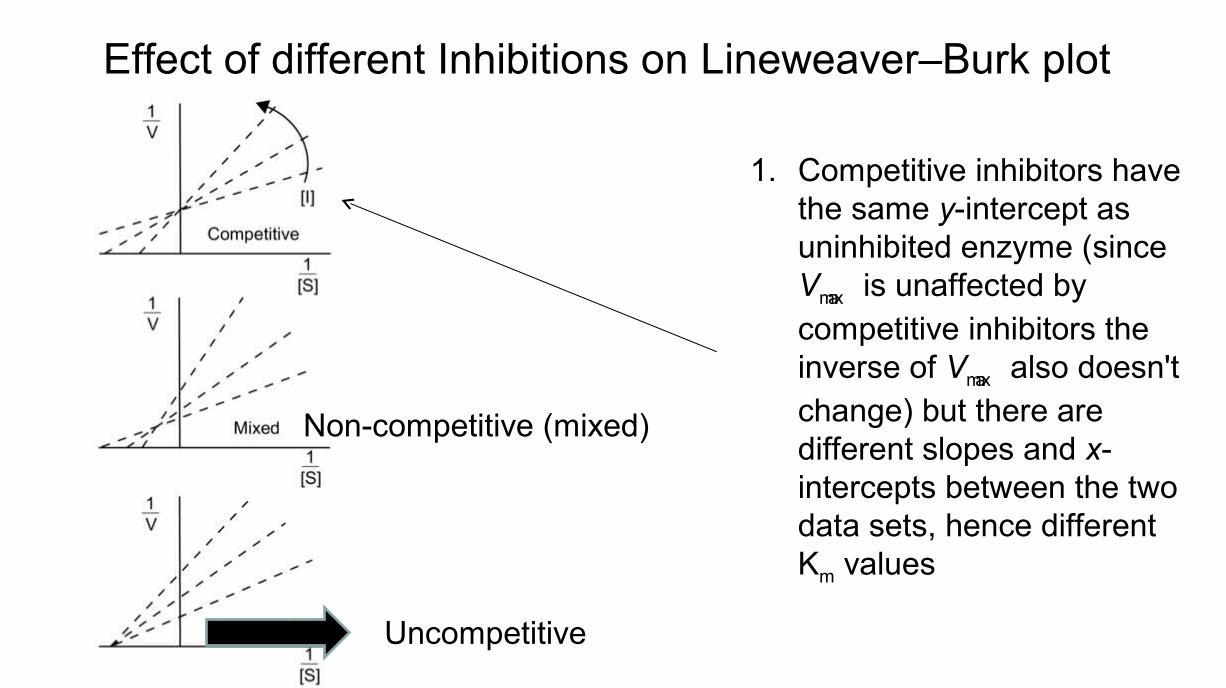
Effect of different Inhibitions on Lineweaver–Burk plot
1. Competitive inhibitors have the same y-intercept as uninhibited enzyme (since Vmax is unaffected by competitive inhibitors the inverse of Vmax also doesn't change) but there are different slopes and x-intercepts between the two data sets, hence different Km values
Non-competitive (mixed)
Uncompetitive

Effect of different Inhibitions on Lineweaver–Burk plot
• Mixed (Non-competitive) inhibition has different x- and y-intercepts, i.e. both Vmax and Km are different
• Uncompetitive inhibition produces plots with the same x-intercept as uninhibited enzyme (Km is unaffected) but different slopes and y-intercepts.
Uncompetitive
Non-competitive (mixed)

Sequential vs Ping-pong Reactions• sequential reaction exhibits an intersecting pattern of lines Order and random substrate additions cannot be distinguished in this type of plot (like uncompetitive inhibitors)
•Ping-pong reaction shows parallel or non- intersecting lines different intercepts on both the y- and x-axes but the same slope..

Influence of enzyme concentration
When [E] >>[S]
As the [E] increases, Vmax also increases, V0 also increases

Influence of temperature
1. Enzymes are proteins and denature at higher temperatures
2. Each enzyme has an optimum temperature,most of them are in the range from 35 to 45 ℃.

Influence of pH
Optimum pH
1. Enzymes are proteins and denature at extreme ends of the pH scale
2. Each enzyme has an optimal pH range within which its catalytic properties are good.,

Enzyme Inhibition
Enzyme inhibitors are important for a variety of reasons
1) they can be used to gain information about the shape on the enzyme active site and the amino acid residues in the active site.
2) they can be used to gain information about the chemical mechanism.
3) they can be used to gain information about the regulation or control of a metabolic pathway.
4) they can be very important in drug design.

Enzyme Inhibition1. Reversible inhibitor: a substance that binds to an enzyme to
inhibit it, but can be released – usually involves formation of non-covalent bonds
– Generally two types
• Dead end
• Product
2. Irreversible inhibitor: a substance that causes inhibition that cannot be reversed
– usually involves formation or breaking of covalent
bonds to or on the enzyme

Inhibitors
1. Irreversible inhibition
2. Reversible inhibition
2.1 competitive inhibition
2.2 non-competitive (mixed) inhibition
2.3 uncompetitive inhibition

Irreversible inhibition
• Irreversible inhibition: The inhibitor combine with essential group of enzyme
active center by covalent bond, resulting in enzymatic activity loss.
Active Enzyme Inhibited Enzyme after inhibitor covalent bond

Reversible Inhibition
• Inhibitors act in a variety of mechanisms
1.An inhibitor may bind at the same site as one of the substrates
2. – these inhibitors structurally resemble the substrate
3.An inhibitor may bind at an alternate site affecting catalytic activity without affecting substrate binding
4.Many inhibitors do both
5.Most common types 5.1 Competitive
5.2 Uncompetitive
5.3 Mixed or Non-competitive

Competitive Inhibition
• Competitive inhibitor competes with a substrate for the enzyme - substrate binding site
Malonate is a
competitive
inhibitor of
succinate for
succinate
dehydrogenase

• A competitive inhibitor reduces the amount of free enzyme available for substrate binding thus increasing the Km for the substrate
• The effect of a competitive inhibitor can be overcome with high concentrations of the substrate
Competitive Inhibition

Competitive Inhibition

• Unimolecular
Reaction
• Bimolecular
Reaction
Competitive Inhibition

Uncompetitive Inhibition
• An uncompetitive inhibitor binds to the enzyme substrate complex but not to free enzyme
• The result is a decrease in Vmax and Km
• The effect of an uncompetitive inhibitor can not be overcome by high concentrations of the substrate

Uncompetitive Inhibition

Uncompetitive Inhibition
• Uncompetitive inhibition produces plots with the same x-intercept as uninhibited enzyme (Km is unaffected) but different slopes and y-intercepts.
Uncompetitive
Non-competitive (mixed)

Mixed or Non-Competitive Inhibition• The inhibitor can bind to both free enzyme and the ES complex
• The affinity of the inhibitor to the two complexes might be different
– If binding of inhibitor changes the affinity for the substrate, Km will be changed
– Since the Vmax is also affected the inhibitor is then termed Non-competitive inhibitor

Mixed Inhibition

• The result will be decrease in Vmax and either an increase or decrease in Km
• The effect of an non-competitive inhibitor can only be partially overcome by high concentrations of the substrate
Mixed Inhibition

Mixed (Non-Competitive Inhibition)
• Mixed (Non-competitive) inhibition has different x- and y-intercepts, i.e. both Vmax and Km are different
Uncompetitive
Non-competitive (mixed)