electron microscopical and biochemical characterization of infectious pancreatic necrosis virus
TRANSCRIPT

JOURNAL OF VIROLOGY, OCt. 1972, p. 824-834 Vol. 10, No. 4Copyright ( 1972 American Society for Microbiology Printed in U.S.A.
Electron Microscopical and BiochemicalCharacterization of Infectious
Pancreatic Necrosis VirusRUSSELL K. KELLY AND PHILIP C. LOH
Virus Laboratory, Department of Microbiology, University of Hawaii, Honolulu, Hawaii 96822
Received for publication 31 May 1972
An electron microscopical and biochemical examination of the properties ofinfectious pancreatic necrosis virus (IPN) and of its ribonucleic acid (RNA) wasmade. The buoyant density of IPN in CsCI was found to be 1.33 g/cm3. Electronmicroscopical examination of the banded virus revealed structures similar in size(74 nm) and shape to reoviruses but lacking a characteristic inner capsid structure.Polyacrylamide gel electrophoretic analysis of IPN-RNA revealed a single non-segmented component of molecular weight 3.2 X 106. Its susceptibility to ribo-nuclease, base composition, and resistance to thermal denaturation indicated asingle-stranded RNA structure. However, its sedimentation behavior (16S) in-dependent of ionic strength in sucrose gradients, partial solubility in 2 M LiCl, andribonuclease resistance in the presence of Mg2+ suggest an unusual secondarystructure of unknown nature. The accumulated data indicate that IPN virus doesnot belong to either the picornavirus or reovirus groups and may represent a newgroup of viruses.
Infectious pancreatic necrosis (IPN) virus, anagent which causes a high-mortality disease oftrout fingerlings, was tentatively classified as amember of the picornavirus group (1, 21, 39).The classification was based upon its morphologyand size (18 nm), cytoplasmic site of synthesis,ether stability, and lack of inhibition of its syn-thesis by the halogenated pyrimidines (21, 22).However, two recent studies on the basis ofelectron microscopical and limited biochemicaland cytochemical analyses have reported that thevirus is similar in size and morphology to thereovirus (25), and may have for its genome adouble-stranded RNA (dsRNA) component(J. E. Argot, Ph.D. thesis, Lehigh Univ., Bethle-hem, Pa., 1969, microfilms at Ann Arbor, Mich.).In contrast to these findings, Nicholson (26), onthe basis of cytochemical and autoradiographicstudies of infected cells, suggested that thegenome of IPN virus is most probably a single-stranded (ssRNA) structure. To resolve thesedifferences we have undertaken an electron micro-scopical and biochemical characterization of theIPN agent. The results obtained strongly suggestthat the IPN virus does not belong to either thereovirus or the picomavirus group.
MATERIALS AND METHODSVirus. The IPN virus (ATCC VR299) was kindly
supplied by Kenneth E. Wolf of the Eastern Fish
Disease Laboratory, Kearneysville, W. Va. Stockcultures of the virus were prepared in RTG-2 cells(rainbow trout gonads). Virus infectivity titrationswere done in tube cultures of RTG-2 cells at 22 C,and the 50% end point was calculated according tothe method of Reed and Muench (27). For the prepa-ration of radioactively labeled viral nucleic acid, theIPN virus was grown in RTG-2 cells maintained ineither a low-phosphate Eagle basal medium-0.2%fetal calf serum (EBMO *2FC) containing carrier-free 3'P-orthophosphate (10,gCi/ml) or EBMO-2FCcontaining 3H-uridine (3 ,Ci/ml) (20).
Virus purification. For the preparation of purifiedIPN virus, infected RTG-2 cells, exhibiting 4+cytopathic effects after 3 to 4 days at 22 C, wereharvested, and the virus-cell mixture was brieflysonically treated (30 sec at 20 kc). After slow centrif-ugation (850 X g, 10 min) to remove cellular debris,the virus was concentrated by centrifugation at 78,000X g for 2 hr. The pellet was resuspended in Dul-becco's phosphate-buffered saline (D-PBS), pH 7.2,without Ca2+ and treated with the following enzymes:ribonuclease at 20 gg/ml and deoxyribonuclease(containing 0.003 M Mg2+) at 60 ,ug/ml for 1 hr at37 C. The virus was recentrifuged at 78,000 X g for 2hr, and the pellet, after brief sonic treatment todisperse aggregates, was isopycnically banded twotimes in CsCl (500 mg/ml) at 92,000 X g for 24 hr(SW39 rotor). Following centrifugation, fractionswere collected from the bottom of the centrifuge tube,and either the radioactivity or the infectivity of eachfraction was determined. The buoyant densities of
824
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 29
Dec
embe
r 20
21 b
y 60
.112
.194
.132
.
CHARACTERIZATION OF IPN VIRUS
representative fractions were calculated from theirrefractive index measurements (13).
Extraction of viral RNA. The IPN-RNA was ex-tracted from purified virus by one of the two fol-lowing methods, which yielded similar results: (i)extraction by hot phenol-sodium dodecyl sulfate(SDS) (0.5%; reference 28), as modified for reovirus(30); (ii) extraction by 1% SDS and 4 M urea at37 C for 10 min. In some experiments the latter pro-cedure was preceded by treatment with Pronase for 1hr at 37 C. After extraction the viral RNA was pre-cipitated overnight with two volumes of cold ethanolat -20 C and recovered by centrifugation. In certainexperiments when the extraction was by the SDS-urea method, the viral RNA was applied directlyon to polyacrylamide gels and subjected to electro-phoresis.
Based composition analysis. The base compositionof viral RNA was analyzed by chromatography onthe formate form of diethylaminoethyl-cellulose(DEAE-cellulose) paper strips, using formic acidas the irrigation solvent (14).
Polyacrylamide gel electrophoresis. The procedurefor polyacrylamide gel electrophoresis of viral RNAhas been described (32). After electrophoresis, thegels were either frozen, sliced, or solubilized in H202,and the radioactivity was counted or scanned at 260nm in a Gilford spectrophotometer with a lineartransport attachment.
Sucrose gradient analysis. For sucrose gradientanalysis the 32P-labeled IPN-RNA was overlayedonto a preformed 5 to 20% (w/v) sucrose (ribonu-clease-free) gradient and centrifuged in a SW39swinging bucket rotor at 31,000 rev/min for 5 to 12hr at 4 C. In some experiments, either rat or HeLacell 28S ribosomal RNA served as marker. Aftercentrifugation the gradients were fractionated, andthe location of the marker RNA was determined withthe aid of an ISCO density gradient fractionator withan ultraviolet monitor.
Negative staining. For negative stains, one drop ofpurified virus suspension was placed on a carbonizedFormvar-coated grid and allowed to stand for 1 min(8). The excess fluid was drained off, and the grid wasstained with 2% phosphotungstic acid containing0.4% sucrose at pH 6.0 for 1 min. After removal ofthe excess fluid, the grids were air-dried at room tem-perature and examined in a Hitachi HU-11 electronmicroscope.
Standard buffers. The following buffers were used.Acetate buffer, pH 5.1, consisted of 0.01 M Na acetate,0.05 M NaCl, 0.0001 M MgCl2. Tris, NaCI, EDTAbuffer, pH 7.4 (1 X TNE), consisted of 0.01 Mtris(hydroxymethyl)aminomethane (Tris)-hydrochlo-ride (pH 7.4), 0.1 M NaCl, 0.001 M ethylenediamine-tetracetate (EDTA). Standard saline-citrate buffer(1 X SSC) consisted of 0.15 M NaCl, 0.015 M Nacitrate. Dulbecco's phosphate-buffered saline minusCa2+, pH 7.2 (D-PBS), consisted of 0.137 M NaCl,0.003 M KCl, 0.009 M Na2HPO4, 0.0015 M KH2PO4,0.0005 M MgCl2 6H20. Phosphate-buffered saline,pH 7.2, was the same as D-PBS but without Mg2+ andCa2+.
Enzymes and radioisotopes. Pancreatic ribonu-clease and deoxyribonuclease were products of the
Worthington Biochemical Corp., Freehold, N. J.Carrier-free 3P-orthophosphate was obtained fromNew England Nuclear Corp., Boston, Mass. Tri-tiated uridine (specific activity 20 Ci/mM) was pur-chased from Schwarz BioResearch, Van Nuys, Calif.Membrane filters of pore sizes 0.05 ,um and 0.2,um (Triacetate Metrical GA-8 and GA-10) wereobtained from Gelman Instrument Co., Ann Arbor,Mich.
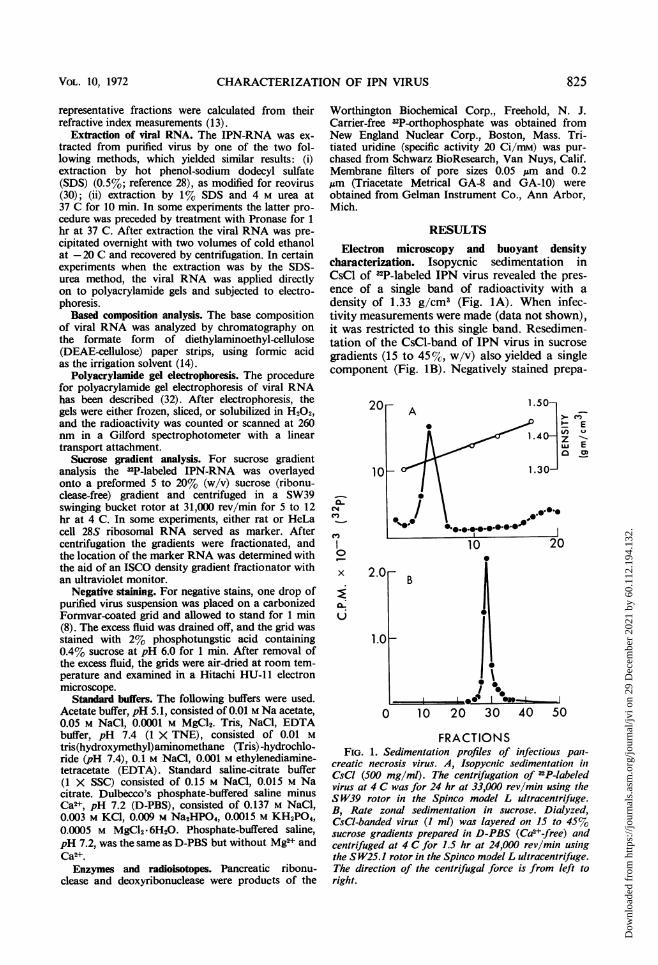
RESULTSElectron microscopy and buoyant density
characterization. Isopycnic sedimentation inCsCl of 32P-labeled IPN virus revealed the pres-ence of a single band of radioactivity with adensity of 1.33 g/cm3 (Fig. lA). When infec-tivity measurements were made (data not shown),it was restricted to this single band. Resedimen-tation of the CsCl-band of IPN virus in sucrosegradients (15 to 45%, w/v) also yielded a singlecomponent (Fig. iB). Negatively stained prepa-
a-CL.C14C.,
C.,)I0
x
a-'UL
2.
' Ee) U
XU EaOO
0 10 20 30 40 50
FRACTIONSFIG. 1. Sedimentation profiles of infectious pan-
creatic necrosis virus. A, Isopycnic sedimentation inCsCI (500 mg/ml). The centrifugation of 32P-labeledvirus at 4 C was for 24 hr at 33,000 rev/min using theS W39 rotor in the Spinco model L ultracentrifuge.B, Rate zonal sedimentation in sucrose. Dialyzed,CsCI-banded virus (I ml) was layered on 15 to 45%sucrose gradients prepared in D-PBS (Ca2+-free) andcentrifuged at 4 C for 1.5 hr at 24,000 rev/min usingthe S W25.1 rotor in the Spinco model L ultracentrifuge.The direction of the centrifugal force is from left toright.
VOL. lo) 1972 825
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 29
Dec
embe
r 20
21 b
y 60
.112
.194
.132
.
KELLY AND LOH
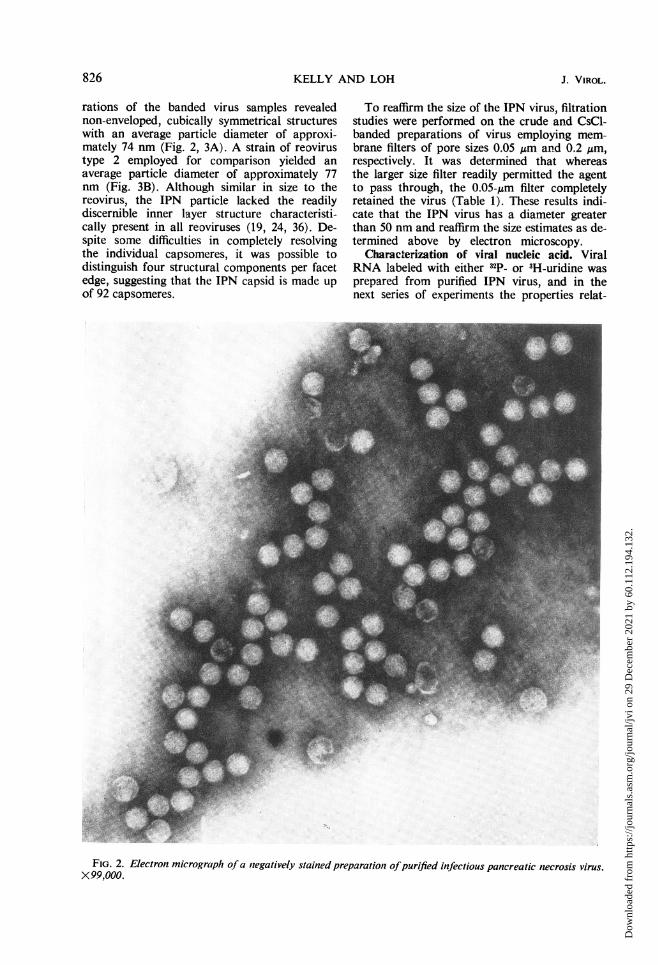
rations of the banded virus samples revealednon-enveloped, cubically symmetrical structureswith an average particle diameter of approxi-mately 74 nm (Fig. 2, 3A). A strain of reovirustype 2 employed for comparison yielded anaverage particle diameter of approximately 77nm (Fig. 3B). Although similar in size to thereovirus, the IPN particle lacked the readilydiscernible inner layer structure characteristi-cally present in all reoviruses (19, 24, 36). De-spite some difficulties in completely resolvingthe individual capsomeres, it was possible todistinguish four structural components per facetedge, suggesting that the IPN capsid is made upof 92 capsomeres.
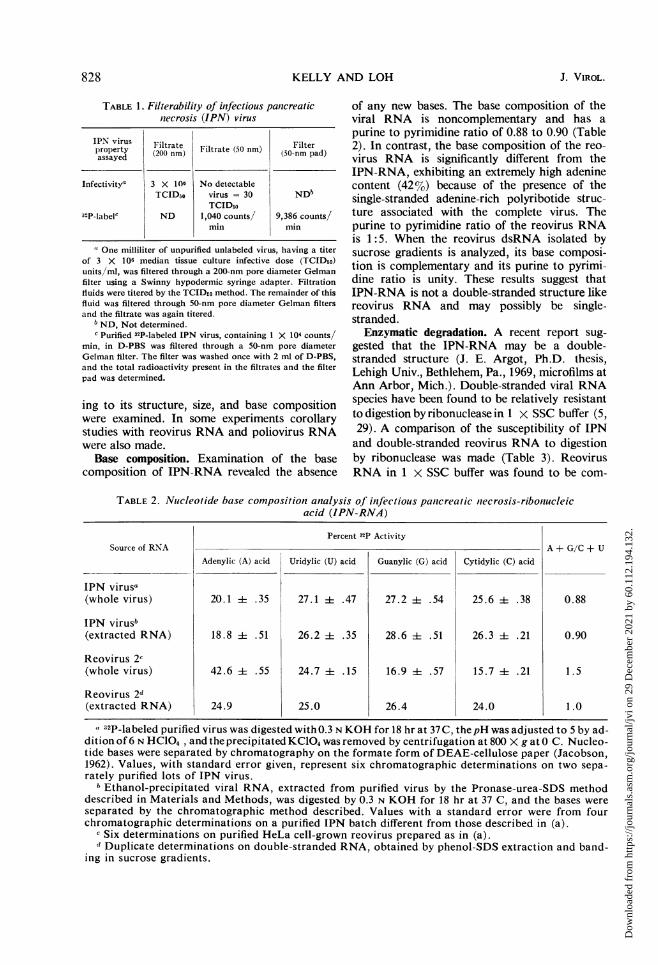
To reaffirm the size of the IPN virus, filtrationstudies were performed on the crude and CsCl-banded preparations of virus employing mem-brane filters of pore sizes 0.05 ,um and 0.2 Mm,respectively. It was determined that whereasthe larger size filter readily permitted the agentto pass through, the 0.05-Aim filter completelyretained the virus (Table 1). These results indi-cate that the IPN virus has a diameter greaterthan 50 nm and reaffirm the size estimates as de-termined above by electron microscopy.
Characterization of viral nucleic acid. ViralRNA labeled with either 32p_ or 3H-uridine wasprepared from purified IPN virus, and in thenext series of experiments the properties relat-
FIG. 2. Electron micrograph of a negatively stained preparation ofpurified infectious pancreatic necrosis virus.X99,000.
826 J. VIROL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 29
Dec
embe
r 20
21 b
y 60
.112
.194
.132
.
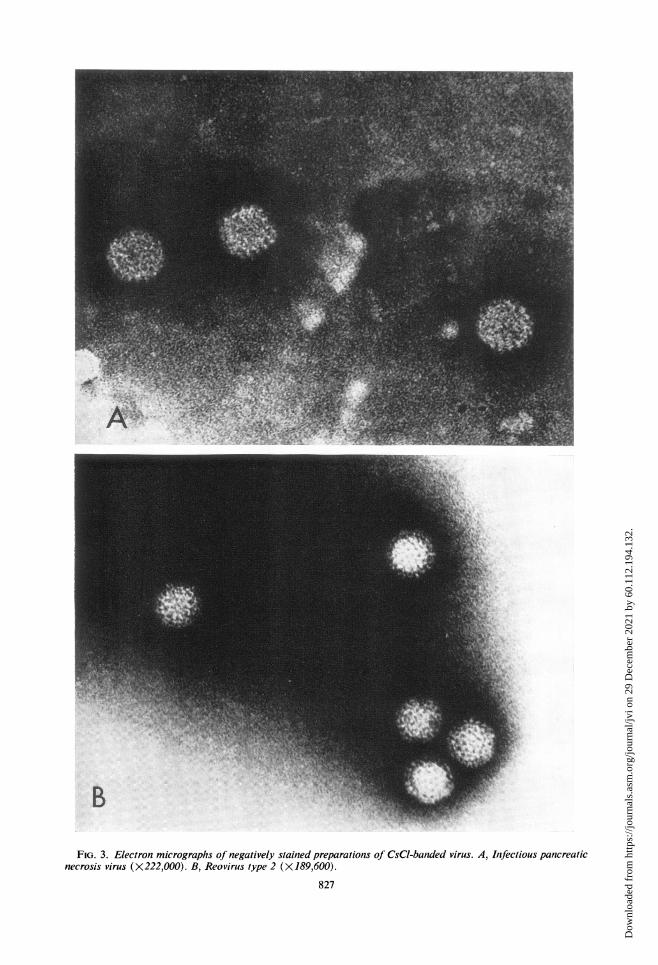
FIG. 3. Electron micrographs of negatively stained preparations of CsCl-banded virus. A, Infectious pancreaticnecrosis virus (X 222,000). B, Reovirus type 2 (X 189,600).
827
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 29
Dec
embe
r 20
21 b
y 60
.112
.194
.132
.
KELLY AND LOH
TABLE 1. Filterability of infectious panicreaticniecrosis (IPN) virus
IPN virus Filtrate Filterl)roperty (200rnm) Filtrate (50 nm) (50-nm pad)assayed
Infectivity" 3 X 106 No detectableTCID5o virus = 30 ND6
TCID5032P-labelc ND 1,040 counts/ 9,386 counts/
min min
aOne milliliter of unpurified unlabeled virus, having a titerof 3 X 106 median tissue culture infective dose (TCIDso)units/ml, was filtered through a 200-nm pore diameter Gelmanfilter using a Swinny hypodermic syringe adapter. Filtrationfluids were titered by the TCIDeo method. The remainder of thisfluid was filtered through 50-nm pore diameter Gelman filtersand the filtrate was again titered.
ND, Not determined.c Purified 32P-labeled IPN virus, containing I X 104 counts/
min, in D-PBS was filtered through a 50-nm pore diameterGelman filter. The filter was washed once with 2 ml of D-PBS,and the total radioactivity present in the filtrates and the filterpad was determined.
ing to its structure, size, and base compositionwere examined. In some experiments corollarystudies with reovirus RNA and poliovirus RNAwere also made.
Base composition. Examination of the basecomposition of IPN-RNA revealed the absence
of any new bases. The base composition of theviral RNA is noncomplementary and has apurine to pyrimidine ratio of 0.88 to 0.90 (Table2). In contrast, the base composition of the reo-virus RNA is significantly different from theIPN-RNA, exhibiting an extremely high adeninecontent (42%7,) because of the presence of thesingle-stranded adenine-rich polyribotide struc-ture associated with the complete virus. Thepurine to pyrimidine ratio of the reovirus RNAis 1:5. When the reovirus dsRNA isolated bysucrose gradients is analyzed, its base composi-tion is complementary and its purine to pyrimi-dine ratio is unity. These results suggest thatIPN-RNA is not a double-stranded structure likereovirus RNA and may possibly be single-stranded.
Enzymatic degradation. A recent report sug-gested that the IPN-RNA may be a double-stranded structure (J. E. Argot, Ph.D. thesis,Lehigh Univ., Bethlehem, Pa., 1969, microfilms atAnn Arbor, Mich.). Double-stranded viral RNAspecies have been found to be relatively resistantto digestion by ribonuclease in 1 x SSC buffer (5,29). A comparison of the susceptibility of IPNand double-stranded reovirus RNA to digestionby ribonuclease was made (Table 3). ReovirusRNA in 1 x SSC buffer was found to be com-
TABLE 2. Nucleotide base composition analysis of inJectious pantcreatic necrosis-ribonucleicacid (IPN-RNA)
Source of RNA
IPN virusa(whole virus)
IPN virusb(extracted RNA)
Reovirus 2c(whole virus)
Reovirus 2d(extracted RNA)
Percent 32P Activity
Adenylic (A) acid
20.1 ± .35
18.8 i .51
42.6 i .55
24.9
Uridylic (U) acid
27.1 i .47
26.2 4 .35
24.7 i .15
25.0
Guanylic (G) acid
27.2 i .54
28.6 ±4 .51
16.9 4 .57
26.4
Cytidylic (C) acid
25.6 i .38
26.3 i .21
15.7 i .21
24.0
A+ G/C+ U
0.88
0.90
1.5
1.0
a 32P-labeled purified virus was digested with O.3 N KOH for 18 hr at 37C, the pH was adjusted to 5 by ad-dition of 6 N HC104, and the precipitated KCIO4 was removed by centrifugation at 800 X g at O C. Nucleo-tide bases were separated by chromatography on the formate form of DEAE-cellulose paper (Jacobson,1962). Values, with standard error given, represent six chromatographic determinations on two sepa-rately purified lots of IPN virus.
I Ethanol-precipitated viral RNA, extracted from purified virus by the Pronase-urea-SDS methoddescribed in Materials and Methods, was digested by 0.3 N KOH for 18 hr at 37 C, and the bases wereseparated by the chromatographic method described. Values with a standard error were from fourchromatographic determinations on a purified IPN batch different from those described in (a).
c Six determinations on purified HeLa cell-grown reovirus prepared as in (a).d Duplicate determinations on double-stranded RNA, obtained by phenol-SDS extraction and band-
ing in sucrose gradients.
828 J. VIROL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 29
Dec
embe
r 20
21 b
y 60
.112
.194
.132
.
CHARACTERIZATION OF IPN VIRUS
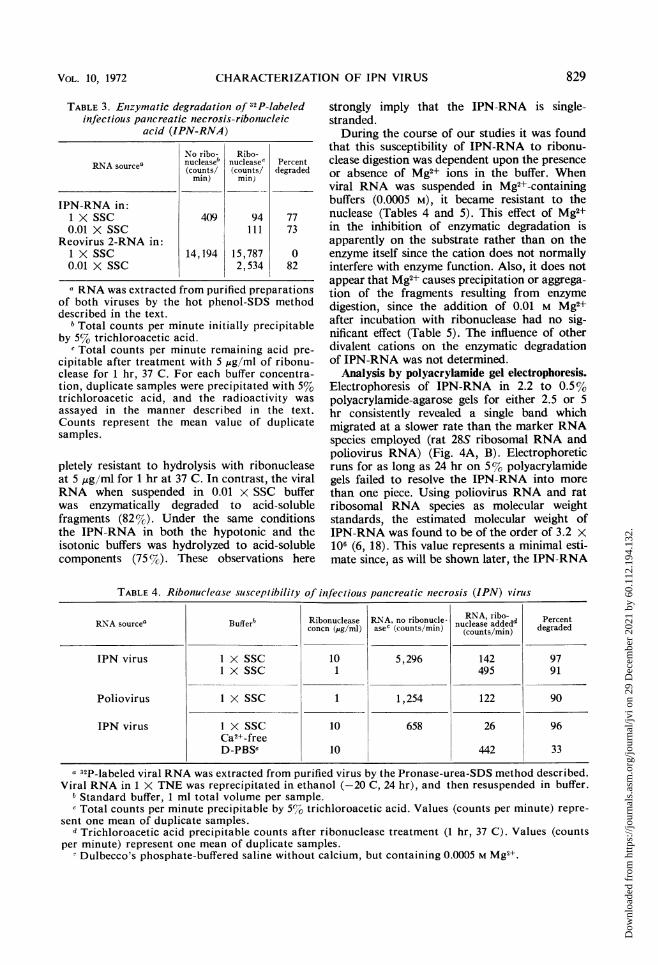
TABLE 3. Enzymatic degradationi of 32P-labeledinfectious pancreatic necrosis-ribonucleic
acid (IPN-RNA)
No ribo- Ribo-RNA sourcea nucleaseb nuclease' Percent(counts/ (counts/ degraded
min) min)
IPN-RNA in:1 X SSC 409 94 770.01 X SSC Ill 73
Reovirus 2-RNA in:1 X SSC 14,194 15,787 00.01 X SSC 2,534 82
a RNA was extracted from purified preparationsof both viruses by the hot phenol-SDS methoddescribed in the text.
I Total counts per minute initially precipitableby 5% trichloroacetic acid.
c Total counts per minute remaining acid pre-cipitable after treatment with 5 ,ug/ml of ribonu-clease for 1 hr, 37 C. For each buffer concentra-tion, duplicate samples were precipitated with 5%trichloroacetic acid, and the radioactivity wasassayed in the manner described in the text.Counts represent the mean value of duplicatesamples.
pletely resistant to hydrolysis with ribonucleaseat 5 ,Ag/ml for 1 hr at 37 C. In contrast, the viralRNA when suspended in 0.01 x SSC bufferwas enzymatically degraded to acid-solublefragments (82%). Under the same conditionsthe IPN-RNA in both the hypotonic and theisotonic buffers was hydrolyzed to acid-solublecomponents (75 %G). These observations here
strongly imply that the IPN-RNA is single-stranded.During the course of our studies it was found
that this susceptibility of IPN-RNA to ribonu-clease digestion was dependent upon the presenceor absence of Mg2+ ions in the buffer. Whenviral RNA was suspended in Mg2+-containingbuffers (0.0005 M), it became resistant to thenuclease (Tables 4 and 5). This effect of Mg2+in the inhibition of enzymatic degradation isapparently on the substrate rather than on theenzyme itself since the cation does not normallyinterfere with enzyme function. Also, it does notappear that Mg2+ causes precipitation or aggrega-tion of the fragments resulting from enzymedigestion, since the addition of 0.01 M Mg2+after incubation with ribonuclease had no sig-nificant effect (Table 5). The influence of otherdivalent cations on the enzymatic degradationof IPN-RNA was not determined.
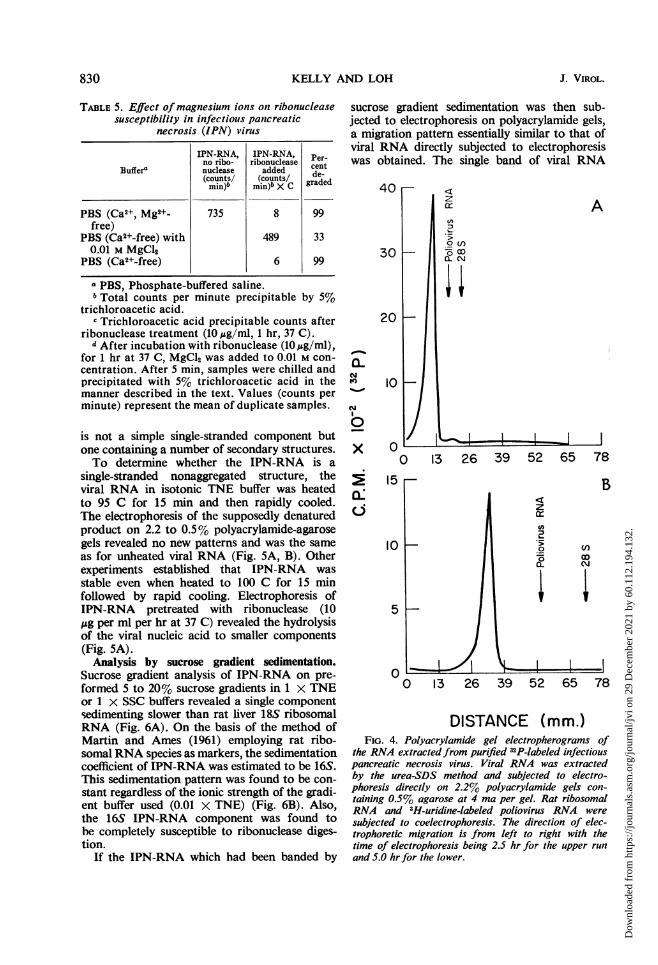
Analysis by polyacrylamide gel electrophoresis.Electrophoresis of IPN-RNA in 2.2 to 0.5%polyacrylamide-agarose gels for either 2.5 or 5hr consistently revealed a single band whichmigrated at a slower rate than the marker RNAspecies employed (rat 28S ribosomal RNA andpoliovirus RNA) (Fig. 4A, B). Electrophoreticruns for as long as 24 hr on 5% polyacrylamidegels failed to resolve the IPN-RNA into more
than one piece. Using poliovirus RNA and ratribosomal RNA species as molecular weightstandards, the estimated molecular weight ofIPN-RNA was found to be of the order of 3.2 X106 (6, 18). This value represents a minimal esti-mate since, as will be shown later, the IPN-RNA
TABLE 4. Ribonuclease suscep-tibility of infectious pancreatic necrosis (IPN) virusI
RN-A sourcea Buffer Ribonuclease RNA, no ribonucle- NIA, addedd Percentconcn (jug/ml) aseC (counts/min) nucoease ae) degraded
IPN virus 1 X SSC 10 5,296 142 971 X SSC 1 495 91
Poliovirus 1 X SSC 1 1,254 122 90
IPN virus 1 X SSC 10 658 26 96Ca2+-freeD-PBSe 10 442 33
a 32P-labeled viral RNA was extracted from purified virus by the Pronase-urea-SDS method described.Viral RNA in 1 X TNE was reprecipitated in ethanol (-20 C, 24 hr), and then resuspended in buffer.
b Standard buffer, 1 ml total volume per sample.c Total counts per minute precipitable by 5%7 trichloroacetic acid. Values (counts per minute) repre-
sent one mean of duplicate samples.d Trichloroacetic acid precipitable counts after ribonuclease treatment (1 hr, 37 C). Values (counts
per minute) represent one mean of duplicate samples.- Dulbecco's phosphate-buffered saline without calcium, but containing 0.0005 M Mg'+.
829VOL. 10, 1972
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 29
Dec
embe
r 20
21 b
y 60
.112
.194
.132
.
KELLY AND LOH
TABLE 5. Effect of magnesium ions on ribonucleasesusceptibility in infectious pancreatic
necrosis (IPN) virus
IPN-RNA, IPN-RNA, Per-no ribo- ribonuclease cBuffera nuclease added cent(counts/ (counts/ gradedmin)b min)b X Cgae
PBS (Ca2+, Mg2+- 735 8 99free)
PBS (Ca2+-free) with 489 330.01 M MgCl2
PBS (Ca2+-free) 6 99
a PBS, Phosphate-buffered saline.b Total counts per minute precipitable by 5%
trichloroacetic acid.c Trichloroacetic acid precipitable counts after
ribonuclease treatment (10,ug/ml, 1 hr, 37 C).d After incubation with ribonuclease (10 ,g/ml),
for 1 hr at 37 C, MgCl2 was added to 0.01 M con-centration. After 5 min, samples were chilled andprecipitated with 5% trichloroacetic acid in themanner described in the text. Values (counts perminute) represent the mean of duplicate samples.
is not a simple single-stranded component butone containing a number of secondary structures.To determine whether the IPN-RNA is a
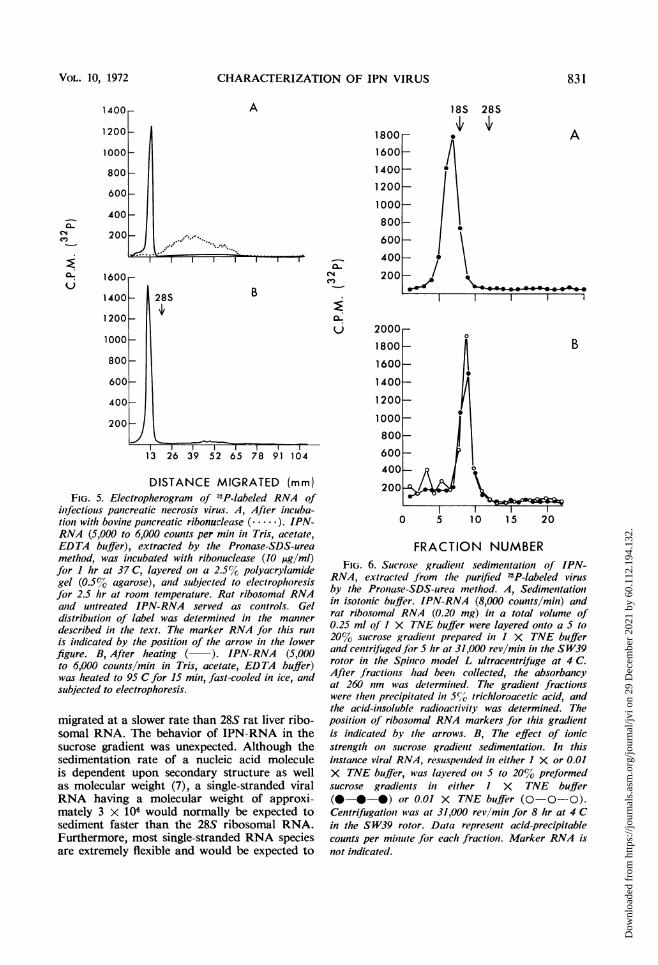
single-stranded nonaggregated structure, theviral RNA in isotonic TNE buffer was heatedto 95 C for 15 min and then rapidly cooled.The electrophoresis of the supposedly denaturedproduct on 2.2 to 0.5% polyacrylamide-agarosegels revealed no new patterns and was the sameas for unheated viral RNA (Fig. 5A, B). Otherexperiments established that IPN-RNA wasstable even when heated to 100 C for 15 minfollowed by rapid cooling. Electrophoresis ofIPN-RNA pretreated with ribonuclease (10,ug per ml per hr at 37 C) revealed the hydrolysisof the viral nucleic acid to smaller components(Fig. 5A).
Analysis by sucrose gradient sedimentation.Sucrose gradient analysis of IPN-RNA on pre-formed 5 to 20% sucrose gradients in 1 x TNEor 1 x SSC buffers revealed a single componentsedimenting slower than rat liver 18S ribosomalRNA (Fig. 6A). On the basis of the method ofMartin and Ames (1961) employing rat ribo-somal RNA species as markers, the sedimentationcoefficient of IPN-RNA was estimated to be 16S.This sedimentation pattern was found to be con-stant regardless of the ionic strength of the gradi-ent buffer used (0.01 X TNE) (Fig. 6B). Also,the 16S IPN-RNA component was found tohe completely susceptible to ribonuclease diges-tion.
If the IPN-RNA which had been banded by
sucrose gradient sedimentation was then sub-jected to electrophoresis on polyacrylamide gels,a migration pattern essentially similar to that ofviral RNA directly subjected to electrophoresiswas obtained. The single band of viral RNA
AO(IzIcE
30 030~~~~Q_ \,;
20
a-NFe)
x
0
10
0
10
5
0
A
0 13 26 39 52 65 78.F B
z
.2OD
CL Ci0 13 26 39 52 65 78
DISTANCE (mm.)FIG. 4. Polyacrylamide gel electropherograms of
the RNA extractedfrom purified 32P-labeled infectiouspancreatic necrosis virus. Viral RNA was extractedby the urea-SDS method and subjected to electro-phoresis directly on 2.2% polyacrylamide gels con-taining 0.5% agarose at 4 ma per gel. Rat ribosomalRNA and 3H-uridine-labeled poliovirus RNA weresubjected to coelectrophoresis. The direction of elec-trophoretic migration is from left to right with thetime of electrophoresis being 2.5 hr for the upper runand 5.0 hr for the lower.
830 J. VIROL.
15
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 29
Dec
embe
r 20
21 b
y 60
.112
.194
.132
.
CHARACTERIZATION OF IPN VIRUS
A
I I I
28S B
E
X
13 26 39 52 65 78 91 104
DISTANCE MIGRATED (mm)FIG. 5. Electropherogram of 32P-labeled RNA of
infectious pancreatic necrosis virus. A, After incuba-tion with bovine pancreatic ribonuclease (......). IPN-RNA (5,000 to 6,000 counts per mini in Tris, acetate,EDTA buffer), extracted by the Pronase-SDS-ureamethlod, was incubated with ribonuclease (10 ,g/ml)for I hr at 37 C, layered on a 2.5% polyacrylamidegel (0.5% agarose), and subjected to electrophoresisfor 2.5 hr at room temperature. Rat ribosomal RNAand untreated IPN-RNA served as controls. Geldistribution of label was determined in the mannerdescribed in thle text. The marker RNA for this run
is indicated by the position of the arrow in the lowerfigure. B, After heating ( ). IPN-RNA (5,000to 6,000 counts/min in Tris, acetate, EDTA buffer)was heated to 95 C for 15 min, fast-cooled in ice, andsubjected to electrophoresis.
migrated at a slower rate than 28S rat liver ribo-somal RNA. The behavior of IPN-RNA in thesucrose gradient was unexpected. Although thesedimentation rate of a nucleic acid moleculeis dependent upon secondary structure as wellas molecular weight (7), a single-stranded viralRNA having a molecular weight of approxi-mately 3 X 106 would normally be expected tosediment faster than the 28S ribosomal RNA.Furthermore, most single-stranded RNA speciesare extremely flexible and would be expected to
18S 28S
1800 . A1600 -
1400 .1200
1000 _800 - I!
600 /400 0
200 /vfr1
l
B
0 5 10 15 20
FRACTION NUMBERFIG. 6. Sucrose gradient sedimentatioti of IPN-
RNA, extracted from the purified n2P-labeled virusby the Pronase-SDS-urea method. A, Sedimentationin isotonic buffer. IPN-RNA (8,000 counts/min) andrat ribosomal RNA (0.20 mg) in a total volume of0.25 ml of I X TNE buffer were layered onto a 5 to20% sucrose gradient prepared in I X TNE bufferand centrifuged for 5 hr at 31,000 rev/min in the SW39rotor in the Spinico model L ultracentrifuge at 4 C.After fractions had bee,i collected, the absorbancyat 260( nm was determined. The gradient fractionswere then precipitated in 5%/O trichloroacetic acid, andthe acid-insoluble radioactivity was determined. Theposition of ribosomal RNA markers for this gradientis indicated by the arrows. B, The effect of ionicstrength on sucrose gradient sedimentation. In thisinstance viral RNA, resuspended in either 1 X or 0.01X TNE buffer, was layered on 5 to 20% preformedsucrose gradieiits in either I X TNE buffer(0--* *) or 0.01 X TNE buffer (0-0-0).Centrifugation was at 31,000 rev/mimz for 8 hr at 4 Cin the SW39 rotor. Data represent acid-precipitablecounts per miniute for each fraction. Marker RNA isnot indicated.
1400r-
0'C., --:-t . ..
1200
1000
800
600
400
200
1600
1400
1200
1000
800
600
400
200
VOL. 10, 1972 831
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 29
Dec
embe
r 20
21 b
y 60
.112
.194
.132
.

KELLY AND LOH
exhibit altered sedimentation patterns on sucrosegradients prepared in buffers of different ionicstrengths (33). However, in the present studythe IPN-RNA behaves as a relatively rigid mole-cule, almost similar to double-stranded RNAin its hydrodynamic properties. In other experi-ments employing sucrose gradients prepared in1 x TNE buffer containing either 8 M urea or1% SDS, the IPN-RNA sedimented in essen-tially the same manner relative to marker RNA.
Precipitation in high salt. Precipitation with2 M LiCl has been used to discriminate betweensingle-stranded and double-stranded RNA spe-cies, with the latter remaining in solution (3).Employing the method of Baltimore (3), theethanol-precipitated 32P-labeled IPN-RNA wasfound to be soluble in 2 M LiCl (93%) (Table6). These results suggest that the structure ofthe viral nucleic acid is unusual and may be coiledand folded back upon itself, assuming a tertiarystructure which exhibits properties akin todouble-stranded RNA.
DISCUSSION
The present evidence indicates that IPN virusdoes not belong to either the reovirus or picor-navirus group. Although similar in size, shape,and symmetry to the mammalian reoviruses, aswas previously reported by Moss and Gravell(25), the present studies indicate that IPN virusis different from the reovirus in several importantproperties. The IPN virus lacks a readily dis-cernible inner capsid layer which is present in allreovirus (19, 24, 36). Furthermore, IPN virus hasa buoyant density of 1.33 g/cm3 in CsCl,whereas reoviruses band at 1.37 to 1.38 g/cm3(11, 20). The IPN-RNA migrates on polyacryla-
TABLE 6. Precipitability of infectious pancreaticnecrosis (IPN)-RNA by 2 M LiCI
LiCl (2 m) precipitation" Total counts/min Percent
Supernatant 32,550 93Precipitate 2,600 7
a Ethanol-precipitated IPN-RNA extracted bythe Pronase-SDS-urea method was resuspended in1 ml of 1 X TNE buffer and LiCl was added to 2 Mconcentration. After 36 hr at 0 C, the sample wascentrifuged at 16,000 X g for 20 min and thesupernatant fluid was decanted. After suspendingthe pellet in 1 ml of 1 X TNE buffer, 0.02-mlduplicate samples of both fractions were spottedon 2.54 by 5.08 cm filter paper strips and assayedin the liquid scintillation spectrometer in themanner described in the text.
mide gels as a single component with a molecularweight of approximately 3 x 106 with no evidenceof additional components such as the poly A,which has been reported for reoviruses (4, 31).On the other hand, the extracted dsRNA genomeof reovirus has yielded multiple segments with atotal molecular weight of approximately 15 X 106(11, 12, 32, 35, 38). Similar properties have beendescribed for other dsRNA-containing viruses(15, 37).In a recent report, the structure of IPN-RNA
has been interpreted to be double-stranded on thebasis of only thermal denaturation studies (J. E.Argot, Ph.D. thesis, Lehigh Univ., Bethlehem,Pa., 1969, microfilms at Ann Arbor, Mich.).From the present study it can be concluded thatthe viral RNA is very likely single-stranded,because of its susceptibility to digestion bypancreatic ribonuclease, its noncomplementarybase composition, and its resistance to denatura-tion upon heating to 95 to 100 C (15 min) andthen rapidly cooling. These properties are in-consistent with a truly double-stranded helicalstructure held together by hydrogen bonding ofcomplementary base pairs of adjacent poly-ribonucleotide chains, as reported for the RNA ofreoviruses (2, 11). Nevertheless, the IPN-RNAis unique, for in sucrose gradients it behaveshydrodynamically as an inflexible rod (likedouble-stranded RNA). The structural configura-tion of single-stranded RNA species, such asTMV-RNA, has been reported to be influencedby the ionic strength of the suspending buffer(17, 33). Thus, TMV-RNA in 0.1 M buffer ascontrasted to stretched out rods in 0.001 M buffer,appears as oblatespherical, tangled, coiled struc-tures which sediment with increased speed in theultracentrifuge (17). However, in the presentstudy the structure of IPN-RNA does not appearto be influenced by ionic strength, since the sedi-mentation rate of the viral RNA remained con-stant in sucrose gradients prepared either in0.01 M or in 0.1 M buffer, respectively.Not all single-stranded viral RNA have been
found to behave in solution like TMV-RNA. Forexample, bacteriophage MS2-RNA when com-pared with TMV-RNA exhibited relatively littlechange in sedimentation behavior when thebuffer concentration was changed from 0.02 Mto 0.2 M NaCl (34). In addition, MS2-RNA wassoluble in 1 M NaCl and had a relatively high Tmof 76 C in isotonic saline.The extremely low sedimentation rate (in-
dependent of ionic strength) on zonal sucrosegradients of IPN-RNA as compared with itsrelatively slow migration on polyacrylamide gels
832 J. VIROL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 29
Dec
embe
r 20
21 b
y 60
.112
.194
.132
.

CHARACTERIZATION OF IPN VIRUS
is unusual for single-stranded RNA species. Itsuggests that the hydrodynamic behavior ofIPN-RNA is very likely due to a highly orderedsecondary and possibly tertiary structure possess-ing a stable configuration analogous to transferRNA (10). Although Mg2+ appears to stabilizethe viral RNA to digestion by ribonuclease, it isnot essential for the continued maintenance ofRNA structure, since the sedimentation patternof IPN-RNA remains the same in spite of thepresence of the chelating agent EDTA (0.001 M)in sucrose gradients. In a somewhat analogoussituation to IPN-RNA, the 19S and 25S single-stranded RNA forms of incomplete Sendai virus,for as yet unexplainable reasons, have been foundto migrate at a slower rate upon polyacrylamidegel electrophoresis than 28S ribosomal RNA(16).Other explanations for the unusual properties
of IPN-RNA are possible, but additional studies,such as examination of the viral RNA throughelectron microscopical methods, should beundertaken to resolve its structure.Based on careful cytochemical and autoradio-
graphic studies of IPN-infected cells, Nicholson(26) has recently concluded that IPN virus con-tains ssRNA. Our cytochemical and biochemicalstudies of the IPN-infected cell (Kelly and Loh,in preparation) are in complete agreement withthis conclusion. All of these studies taken togetherindicate that IPN virus is distinct from the reo-virus, diplornavirus, and picornavirus groups andshould be placed in a new classification group.
ACKNOWLEDGMENTS
We thank Kenneth P. Camyre for assistance in performing thebase ratio analyses and R. M. T. Ratnayake for assistance inelectron microscopy.
LITERATURE CITED
1. Andrews, C. H., and H. G. Pereira. 1967. Viruses of verte-brates. Williams & Wilkins Co., Baltimore.
2. Arnott, J., F. Httchinson, M. Spencer, M. H. F. Wilkins,and W. Fuller. 1966. X-Ray diffraction studies of double-helical ribonucleic acid. Nature (London) 211:227-232.
3. Baltimore, D. 1966. Purification and properties of poliovirusdouble-stranded ribonucleic acid. J. Mol. Biol. 18:421-428.
4. Bellamy, A. R., and W. K. Joklik. 1967. Studies on the A-richRNA of reovirus. Proc. Nat. Acad. Sci. U.S.A. 58:1389-1395.
5. Billeter, M. A., and C. Weissman. 1966. Double-strandedMS2 RNA from 2-infected Escherichia coli, p. 498-512. InG. L. Cantoni and D. R. Davies (ed.), Procedures in nucleicacid research. Harper & Row, New York.
6. Bishop, D. H. L., J. L. Claybrook, and S. Spiegelman. 1967.Electrophoretic separation of viral-nucleic acids on poly-acrylamide gels. J. Mol. Biol. 26:373 387.
7. Boedeker, H. 1968. Molecular weight and conformation ofRNA, p. 429-458. In L. Grossman and K. Moldave (ed.),
Methods in enzymology, vol. 12. Academic Press Inc., NewYork.
8. Brenner, S., and R. W. Horne. 1959. A negative stainingmethod for high resolution electron microscopy of viruses.Biochim. Biophys. Acta 34:103-1 10.
9. Cerini, C. P., and R. G. Malsberger. 1965. Morphology ofinfectious pancreatic necrosis virus. Ann. N.Y. Acad. Sci126:315-319.
10. Cramer, F. 1971. Three-dimensional structure of RNA. Prog.Nucl. Acid Res. Mol. Biol. 11:391-421.
11. Gomatos, P. J., and I. Tamm. 1963. The seconidary structureof reovirus RNA. Proc. Nat. Acad. Sci. U.S.A. 49:707-714.
12. Granboulan, N., and A. Niveleau. 1967. Etude au microscopyElectronique du RNA du reovirus. J. Microsc. 6:23-30.
13. Ifft, J. B., D. H. Voet, and J. Vinograd. 1961. The determina-tion of density distribution and density gradients in binarysolutions at equilibrium in the centrifuge. J. Phys. Chem.65:1138-1145.
14. Jacobson, K. B. 1962. Ribonucleotides of DNA: separationby chromatography on sheets of diethylaminoethyl-cellu-lose. Science 138:515-516.
15. Kalmakoff, J., L. J. Lewandowski, and D. R. Black. 1969.Comparison of the ribonucleic acid subunits of reovirus,cytoplasmic polyhedrosis virus, and wound tumiior virus. J.Virol. 4:851-856.
16. Kingsbury, D. W., A. Portner, and R. W. Darliilgton. 1970.Properties of incomplete sendai virions and subgenomicviral RNAs. Virology 42:857-871.
17. Kiselev, N. A., L. P. Gavrilova, and A. S. Spirin. 1961. Onconfigurations of high-polymer ribonucleic acid macro-molecules as revealed by electron microscopy. J. Mol. Biol3:778-783.
18. Loening, U. E. 1969. The determination of the muolecularweight of ribonucleic acid by polyacr-ylamide gel electro-phoresis. Biochem. J. 113:131-138.
19. Loh, P. C., H. R. Hohl, and M. Soergel. 1965. Finie structureof reovirus type 2. J. Bacteriol. 89:1140-1144.
20. Loh, P. C., and H. K. Oie. 1969. Role of lysine in the replica-tion of reovirus. I. Synthesis of complete and empty virions.J. Virol. 4:890-895.
21. Malsberger, R. G., and C. P. Cerini. 1963. Characteristics ofinfectious pancreatic necrosis virus. J. Bacteriol. 86:1283-1287.
22. Malsberger, R. G., and C. P. Cerini. 1965. Multiplication ofinfectious pancreatic necrosis virus. Ann. N. Y. Acad. Sci126:320-327.
23. Martin, R. G., and B. N. Ames. 1961. A method for deter-mining the sedimentation behavior of enzymes: applicationto protein mixtures. J. Biol. Chem. 236:1372-1379.
24. Mayor, H. D., R. M. Jamison, L. E. Jordon, atnd M. V.Mitchell. 1965. Reoviruses. II. Structure and comiipositionof the virion. J. Bacteriol. 89:1548-1 556.
25. Moss, L. H., and M. Gravell. 1969. Ultrastructure antid sequen-tial development of infectious pancreatic necrosis virus. J.Virol. 3:52-58.
26. Nicholson, B. L. 1971. Macromolecular synthesis in RTG-2cells following infection with infectious pancreatic necrosis(IPN) virus. J. Gen. Virol. 13:359-372.
27. Reed, L. J., and H. Muench. 1938. A simple method of esti-mating fifty per cent and points. Amer. J. Hyg. 27:493-497.
28. Scherrer, K., and J. E. Darnell. 1962. Sedimentationi charac-teristics of rapidly labeled RNA fronm HeLa cells. Biochem.Biophys. Res. Commun. 7:486-496.
29. Shatkin, A. J. 1965. Inactivity of purified reovirus RNA as a
template for Eschserichia coli polymerases in vitro. Proc. Nat.Acad. Sci. U.S.A. 59:1721-1728.
30. Shatkin, A. J., and B. Rada. 1967. Reovirus-directed ribo-nucleic acid synthesis in infected L. cells. J. Virol. 1:24-35.
31. Shatkin, A. J., and J. D. Sipe. 1968. Single-stranded adenine
VOL. 10, 1972 833
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 29
Dec
embe
r 20
21 b
y 60
.112
.194
.132
.

KELLY AND LOH
rich RNA from purified reoviruses. Proc. Nat. Acad. Sci.U.S.A. 59:246-253.
32. Shatkin, A. J., J. D. Sipe, and P. C. Loh. 1968. Separation often rcovirus genomne segments by polyacrylamiiide gel elec-trophoresis. J. Virol. 2:986-991.
33. Spirin, A. S. 1964. Macromiolecul3ar stlrLcture of ribonucleicacids. Reinhold Corp., New York.
34. Strauss, J. H., and R. L. Sinsheimer. 1963. Purification amnproperties of baicteriophage MS2 and its ribonucleic acid.J. Mol. Biol. 7:43-54.
35. VaIsqUez, C., and A. K. Kleinschlmlidt. 1968. Electron micros-
copy of RNA strands released from individual reovirLspatrticles. J. Mol. Biol. 34:137-147.
36. Vasqutez, C., and P. Tournier. 1962. The morphology ofreovirus. Virology 17:503-510.
37. Verwoerd, D. W. 1970. Diplornaviruses: A newly recognizedgroup of double-stranded RNA viruses. Progr. Med. Virol.12:192-210.
38. Watanabv, Y., S. Millward, and A. F. Graham. 1968. Regula-tion of transcription of reovirus genome. J. Mol. Biol.36:107-128.
39. Wolf, K. 1966. The fish viruses. Advan. Virus Res. 12:35-101
834 J. VIROL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 29
Dec
embe
r 20
21 b
y 60
.112
.194
.132
.