effects of solar radiation on the photosynthetic activity of the red alga corallina elongata ellis...
TRANSCRIPT

Jom'aal of
AND ~ o ~
B:BIOLOGY E L S E V I E R Journal of Photochemistry and Photobiology B: Biology 37 (1997) 196-202
Effects of solar radiation on the photosynthetic activity of the red alga Corallina elongata Ellis et Soland
Donat-P. H~ider a,., Michael Lebert a, Antonio Flores-Moya b, Carlos Jimdnez c, Jesfis Mercado c, Soluna Salles c, Jos6 Aguilera c, Fdlix L. Figueroa c
a lnstitutfiir Botanik und Pharmazeutische Biologie. Friedrich-Alexander-Universiti~'t, Staudtstr. 5, D-91058 Erlangen, Germany Departamento de Biologfa Vegetal, Facultad de Ciencias, Universidad de M6laga, Campus Universitario de Teatinos s/n, E-29071 Mcilaga, Spain
c Departamento de Ecologfa, Facultad de Ciencias, Universidad de Mdlaga, Campus Universitario de Teatinos s/n, E-29071 Mdlaga, Spain
Received 8 January 1996; accepted 10 May 1996
Abstract
The photoinhibition of photosynthesis was investigated in two morphotypes of the red alga Corallina elongata, i.e. sun and shade types, using pulse amplitude modulated (PAM) chlorophyll fluorescence and oxygen evolution. The contents of chlorophyll, phycoerythrin, phycocyanin and soluble protein were analysed. Exposure to solar radiation caused a strong decrease in the effective photosynthetic quantum yield. This decline was more pronounced in the shade than in the sun morphotype. The plants partially recovered from this effect, indicating that it was mainly due to reversible photoinhibition and, to a smaller extent, non-reversible photodamage. Photoinhibition occurred in this alga even in its natural habitat when the sun was at high angles. Recovery from photoinhibition was higher in algae grown in the sun than in the shade. The net photosynthetic oxygen production was higher in the shade than in the sun morphotype. A decrease in oxygen evolution was observed at about noon, recovering at dusk. The concentrations of chlorophyll a (Chl a) and phycoerythrin were higher in the shade than in the sun morphotype. The Chl a and phycocyanin contents did not change significantly throughout the day. However, a clear daily variation in phycoerythrin content was observed with a strong decrease around noon. The soluble protein concentration decreased around noon, but only in the sun morphotype. The relationship between the photosynthetic quantum yield, oxygen evolution and pigment concentration is discussed, together with photoinhibition. © 1997 Elsevier Science S.A. All fights reserved.
Keywords: Corallina elongata; Oxygen evolution; PAM fluorescence; Photoinhibition; Pigment content; Rhodophyta; Solar radiation
1. I n t r o d u c t i o n
Most ecophysiological work on photosynthetic processes has concentrated on higher plants [ 1,2], despite the fact that about 50% of the primary biomass production on our planet is based on aquatic ecosystems [3,4]. While most of the
productivity of aquatic habitats is due to phytoplankton, macroalgae are important biomass producers. In contrast with phytoplankton, most macroalgae are sessile and consequently
Abbreviations: Chl a, chlorophyll a; Fo, initial fluorescence in the dark- adapted state; all reaction centres are open (oxidized); Fro, maximal fluo- rescence in the dark-adapted state; all reaction centres are closed (reduced); Fv, variable fluorescence (F m - Fo); F'o, F'm and F'v, the same for the light- adapted state; Ft, current steady state fluorescence; PAM fluorescence, pulse amplitude modulated fluorescence; PAR, photosynthetically active radia- tion; PC, phycocyanin; PE, phycoerythrin; qp, photochemical quenching of chlorophyll fluorescence determined as qp= (F'm- Ft)/(F'm - F'o) ; qN, non-photochemical quenching calculated as qN = 1 - (F'm - F'o) / ( Fm - Fo); SP, soluble protein
* Corresponding author. Tel.: + 49 9131 858216; fax: + 49 9131 858215.
1011-1344/97/$17.00 © 1997 Elsevier Science S.A. All rights reserved PHS101 1-1344(96)07402-7
have to cope with the changing irradiance regime of solar
radiation. As they are adapted to a lower irradiance than that of solar radiation at the water surface, macroalgae face a
serious light stress when exposed to higher irradiances [ 5,6 ]. Excess irradiation causes reversible photoinhibition [7-11 ]
or even irreversible photodamage. The mechanism of pho- toinhibition is still under debate, but it can be interpreted as
an active protective mechanism against overexposure to solar
radiation by a reversible interruption of the photosynthetic
electron transport chain [ 2,12]. In the past, ecophysiological investigations of photosyn-
thesis have been limited to gas exchange measurements [ 13 ] ; however, in recent years, the technique of pulse amplitude
modulated (PAM) fluorescence has been introduced [ 2,14- 16]. PAM fluorescence allows information to be obtained on
the physiological status and the regulatory processes of pho- tosynthetic energy allocation in vivo. Initial measurements
were restricted to the laboratory, but recent miniaturization
and computer control have facilitated operation of the equip-

D.-P. HiMer et al. / Journal of Phowchemistry and Photobiology B: Biology 37 (1997) 196-202 197
ment in the field [17]. For macroalgae, in particular, it is preferable to perform measurements in the field, since the transport of the specimens to the laboratory introduces severe stress factors, including changes in temperature, irradiance and salinity.
The method of quenching analysis is based on the meas- urement of fluorescence parameters in response to saturating light pulses in dark- or light-adapted specimens [18,19]. PAM fluorescence measurements usually start with the meas- urement of the initial fluorescence Fo induced by low irradi- ation of a dark-adapted sample. After a single saturating flash, the maximal fluorescence Fm is detected. The difference between Fo and F~ is called the variable fluorescence Fv. The optimal quantum yield can be calculated from the ratio Fv/ Fm [20]. Light adaptation of the sample usually leads to a decrease in F m (then called F'm) and an increase or decrease in Fo (leading to F'o). The overall effective quantum yield of photochemical energy conversion can be calculated from (F'm-Ft) /F'm [21].
The aim of this paper is to describe the effects of solar radiation in the field on the photosynthetic fluorescence induction, oxygen production and pigmentation in the red macroalga Corallina elongata.
filled with seawater with the water maintained at the temper- ature at which the thalli had grown. The Oz concentration was determined using a Crison OXI 92 oxygen electrode before and after incubation.
2.3. Photosynthetic pigments and soluble protein determination
Algal thalli (20-30 mg) were collected every 2 h and deep frozen in liquid nitrogen until the determination of the pho- tosynthetic pigment and soluble protein (SP) concentrations. Chl a was extracted in 90% neutralized (with sodium car- bonate) acetone, and its concentration was determined with an SPG-550 PYE UNICAM spectrophotometer using the equation of Jeffrey and Humphrey [23]. PE, phycocyanin (PC) and SP were extracted at 4 °C in phosphate buffer (0.1 M, pH 6.5) containing 10 mM EDTA-Naz and 4 mM phen- ylmethylsulphonyl fluoride (PMSF). The biliprotein concen- tration was determined spectrophotometrically using the equations of Beer and Eshel [ 24 ]. The SP concentration was estimated by the Coomassie Blue G-250 method [25].
2.4. Measurements of fluorescence induction
2. Materials and methods
2.1. Plant material
Thalli of the common red alga Corallina elongata (Cor- allinaceae) Ellis et Soland were collected in early September 1995 from a depth range of 0-5 m by diving on a south- exposed rocky shore at Punta Carnero (37°58'N, 23°0'W, southern Spain). The algae growing at the surface are char- acterized by the sun morphotype and the algae growing at 5 m and in caves by the shade morphotype according to Algarra et al. [22]. The shade type has a higher assimilative surface and higher chlorophyll a (Chl a) and phycoerythrin (PE) contents, but a lower carotenoid content. The sun type is further characterized by a high canopy density. Both mor- photypes are easily interconvertible on replantation [22]. The water temperature was around 17 °C and did not differ much between the surface and 5 m depth because of mixing by strong wind and waves. The harvested thalli were pro- tected from direct radiation and immediately subjected to measurements.
2.2. Oxygen evolution measurements
Net oxygen production and respiration rates were deter- mined every 2 h under solar irradiance. For this purpose, 100-200 mg fresh weight of epiphyte-free algae was incubated in cylindrical glass bottles (250 ml) for 15-20 min. Sodium bicarbonate (5 mM) was added to the seawater at the beginning of the incubations to avoid possible carbon limitation. The bottles were placed at the surface of aquaria
In vivo induced chlorophyll fluorescence was measured on site with two portable PAM fluorometers (PAM 2000, Waltz, Effeltrich, Germany) as described by Schreiber and Bilger [ 26 ]. The instrument provides preprogrammed experimental runs, which allow the determination of the dependence of the fluorescence parameters on the actinic irradiance. Before the run, Fm and Fo were determined in dark-adapted specimens. The samples were then exposed for 10 min to an intermediate irradiance (23 W m -2) of red light for light adaptation and activation of the Calvin enzymes. Subsequently, the irradi- ance was increased in 11 steps, each lasting 6.5 min. At the end of each irradiation period, a saturating white light pulse was given to measure the fluorescence parameters Ft and F' m and to calculate the effective quantum yield.
To determine the photoinhibition and subsequent recovery in the field, freshly harvested thalli were mounted in UVB translucent Plexiglas frames (GS 2458, R6hm and Haas, Darmstadt, Germany) and kept in shallow water. After dark adaptation for at least 30 rain, the fluorescence parameters were determined. From these values, the optimal quantum yield was calculated using the Fv/Fm ratio. Subsequently, the algae were exposed to solar radiation and the fluorescence parameters were measured again; from these values, the effective quantum yield was calculated using ( F ' m - F t ) /
F'm. After this treatment, the samples were covered again, and recovery of the photosynthetic yield was determined at predefined intervals during the following hours. The control values were measured in samples which had been treated exactly as the other specimens except for the exposure to solar radiation. In another type of experiment, the effective quantum yield at the natural growth site was determined over the course of a full day. For this purpose, thalli were collected

198 D.-P. Hiider et al. / Journal of Photochemistry and Photobiology B: Biology 37 (1997) 196-202
every hour from sunrise to sunset, and the fluorescence par- ameters were estimated immediately after harvesting.
2.5. Statistics
During the PAM measurements in the field, at least eight individual measurements were taken from which the mean values and standard deviations were calculated (except for the automatic runs in Fig. 6). The photosynthetic oxygen exchange was measured at least three times for each irradia- tion. Pigment and SP determinations were done in triplicate in all cases. All experimental runs were repeated several times.
2.6. Measurement o f solar radiation
Solar radiation was measured in three wavelength bands (UVB, 280-315 nm; UVA, 315-400 nm; PAR, 400-700 nm) using a filter radiometer developed recently (ELDO- NET, Real Time Computer, Erlangen, Germany). The instru- ment is based on waterproof sensors (Grt~bel, Ettlingen, Germany) and takes readings at 1 s intervals which are aver- aged over 1 min. After amplification and analogue/digital conversion, the data are graphically displayed and stored in a computer. Hourly and daily doses were calculated for each wavelength band.
3. Results
3.1. lrradiation measurements
Fig. 1 shows the daily profile of the irradiance data for a typical day during the measurement period. All days showed a clear blue sky with symmetrical irradiance data centred around local noon. The temperature data measured on the shaded side of the instrument are included.
3.2. Oxygen exchange measurements
In general, net photosynthesis (not corrected for dark res- piration) was higher in the shade than in the sun morphotype,
500- 40
45O - 35
400-
3O
3oE ! 2~, 2oI •
~ 2 ~ - 15
- - ~ 5 0 -
10
5 t l - 5
0 . . . . . 0
07:00 08:00 I~:00 10:00 11:00 12:1~ 13:1X) 14:00 15:00 16:00 17:00 18:00 19:00 20:00 21:00
Local t l l / le
Fig. 1. lrradiance in the UVB, UVA and PAR wavelength bands measured with an ELDONET instrument one day (20 September 1995) during the measurement period.
t ~m
o
S
1,0 I 120
(a)
lO:15 12:15 1(]:15 18:15
LocalUme
E
o I
2~1 (b)
10:15 12:15 16:15 18:15
Local t ime
Fig. 2. Daily variation of the photosynthetic oxygen evolution rate in the sun (open columns) and shade (filled columns) morphotypes of the red alga Corallina elongata, expressed in terms of the dry weight (a) and Chl a content (b).
both in terms of the dry weight and chlorophyll content ( Fig. 2). The photosynthesis rate increased from early morn- ing to noon. However, a strong decrease was observed in the afternoon ( 16:15, local time; 14:15, solar time) mainly in the shade type; late in the afternoon ( 18:15), a recovery of the rate of oxygen production was detected. The differences in the rate of net photosynthesis between sun and shade mor- photypes decreased when the data were expressed in terms of Chl a content (Fig. 2(b) ). Thus a clear photoinhibition under natural conditions was observed in Corallina elongata during the hours of maximal irradiance. The dry weight (dw) is correlated with the fresh weight (fw) by the following equation for the shade type
dw = 0.57fw + 0.002 r = 0.98
and for the sun type
dw = 0.43fw + 0.001 r = 0.97
Respiration was also generally higher in the shade than in the sun morphotype of Corallina (Fig. 3). In the former, a decrease from early morning to afternoon was observed, but the reverse occurred in the sun type. At dusk the respiration rate was similar to that observed in the morning hours.
3.3. Photosynthetic pigments and soluble proteins
The Chl a and PE concentrations were higher in the shade than in the sun type algae (Fig. 4), but the two pigments did

D.-P. Hiider et al. / Journal of Photochemistry and Photobiology B: Biology 37 (1997) 196-202 199
o
i
10:15 12:15 16:15 18:15
Local time
Fig. 3. Daily variation of the dark respiration rate in the sun and shade morphotypes of Corallina elongata (symbols as in Fig. 2).
m
F
10:15 12:15 18:t6 18:15
Local time
(b)
not show the same variation throughout the day. While the Chl a content did not change significantly in either of the morphotypes, the concentration of PE clearly decreased from its morning value through noon and only recovered again at dusk (Fig. 4 (b) ) . Both the depletion and recovery of this pigment were higher in the shade morphotype. The concen- tration of PC in Corallina was very low compared with PE, and only in the sun morphotype was a significant depletion of this pigment detected during the high irradiance hours (Fig. 4(c) ).
The concentration of SP was similar in the sun and shade type algae during most of the day; only late in the afternoon ( 16:15, local time) was a significant reduction in concentra- tion detected in the sun type algae (Fig. 5 (a ) ) , coinciding with the depletion of PE, PC and oxygen production. The ratio of PE to SP was higher in the shade than in the sun type algae, and a clear daily cycle of this ratio was observed with higher values during the morning and close to dusk (Fig. 5(b) ).
3.4. PAM fluorescence measurements
In order to determine the dependence of the fluorescence parameters on the actinic light, an automatic run was per- formed on light-exposed plants freshly harvested at the sur- face (Fig. 6(a) ) and at 5 m depth (Fig. 6 (b) ) . Before the automatic run, the samples were dark adapted, and Fo and Fm
F
g F
10:15
015 ]
12:15 16:15 18:15
LOCal t i m
(c)
10:15 12:15 16:15 18:15
LOCal time
(b)
10:15 12:15 16:15 15:15
Loctl t l r ~
Fig. 4. Daily variation of Chl a (a), PE (b) and PC (c) concentration, expressed as milligrams per gram of algal dry weight, in the sun and shade morphotypes of Corallina elongata (symbols as in Fig. 2).
10:15 12:15 16:15 18:15
Local time
Fig. 5. Daily variation of the concentration of SP (a) and percentage of PE with respect to SP (b) in the sun and shade morphotypes of Corallina elongata (symbols as in Fig. 2).

200 D.-P. Hinder et al. / Journal of Photochemistry and Photobiology B: Biology 37 (1997) 196-202
1.60
1.2S : ~
1.00- ~ -
0.75
i u. 0.60
0.25
1.00
(a)
:- z2
10 100
Irradlance [W m'2l
~ (b)
0.26
0"00 . . . . . . . . . . . . . . . . . .
10 100 Irradlance [IN m "a]
Fig. 6. Fluorescence parameters measured in CoraUina elongata, harvested from the surface (a) and at 5 m depth (b), as a function of the irradiance of actinic red light. Before the experiments, the thalli were adapted to an intermediate fluence rate of 23 W m -2 for 10 rain and then exposed to increasing irradiances for periods of 6.5 min each. At the end of each period, the fluorescence parameters were determined: diamonds, Ft; squares, pho- tosynthetic quantum yield; triangles, photochemical quenching; open circles, non-photochemical quenching; asterisks, F'm; filled circles, F'o.
were measured. The thalli were then allowed to adapt to an irradiance of 23 W m - 2 produced from the built-in red light- emitting diodes (LEDs); subsequently, the irradiance was increased in 11 steps from 1 to 79 W m - 2. The steady state fluorescence Ft first decreased and then increased slightly to an optimum at an irradiance of 23-36 W m-2. F'o followed a similar pattern. In contrast, F'm decreased from a maximum of 1.3 at 1 W m - 2 to a value of about 0.8 at 79 W m - 2. The photosynthetic quantum yield had an optimum at about 1.6 W m - 2 and declined to a value near zero at high irradiances. The photochemical quenching started with values close to unity (indicating optimal energy exploitation for photosyn- thesis) and decreased to values near 0.3 at high irradiances. In contrast, non-photochemical quenching increased from zero to about 0.8. Specimens ~aarvested from greater depths had a different morphology than the light-adapted surface algae, characterized by a higher pigment content, especially of PE. They also showed a different behaviour in terms of their fluorescence parameters (Fig. 6(b) ). While Ft and F'o followed similar kinetics to the light-adapted specimens, F ' m
decreased from a significantly lower initial value of 0.93. The
yield was slightly lower than in sun-adapted plants and the non-photochemical quenching increased at higher actinic irradiation than in the surface specimens.
In order to determine the photoinhibition and recovery, thalli were harvested from 0 m and 5 m depth and kept in shallow water suitable for on-site measurements with the PAM instrument (Fig. 7). Immediately after dark exposure for 1 h, the optimal quantum yield was determined. The thalli were then exposed to direct solar radiation for 1 h, and finally shaded for recovery, which was estimated at predefined times for up to 6 h. Immediately after solar exposure, the yield of sun type plants had decreased to less than 35% of the initial value, and this recovered to 75% of the original value after 6 h in the shade. After that time, a specimen which had been subjected to the same treatment except for solar exposure was also measured to determine whether experimental handling caused any stress in addition to solar irradiation (controls). These dark-adapted samples showed the same yield as the initial measurements immediately after harvesting. Speci- mens harvested from a 5 m depth were treated in the same way and showed an even higher inhibition of the photosyn- thetic yield after 1 h solar exposure. Their recovery was also slower and to a lower final value.
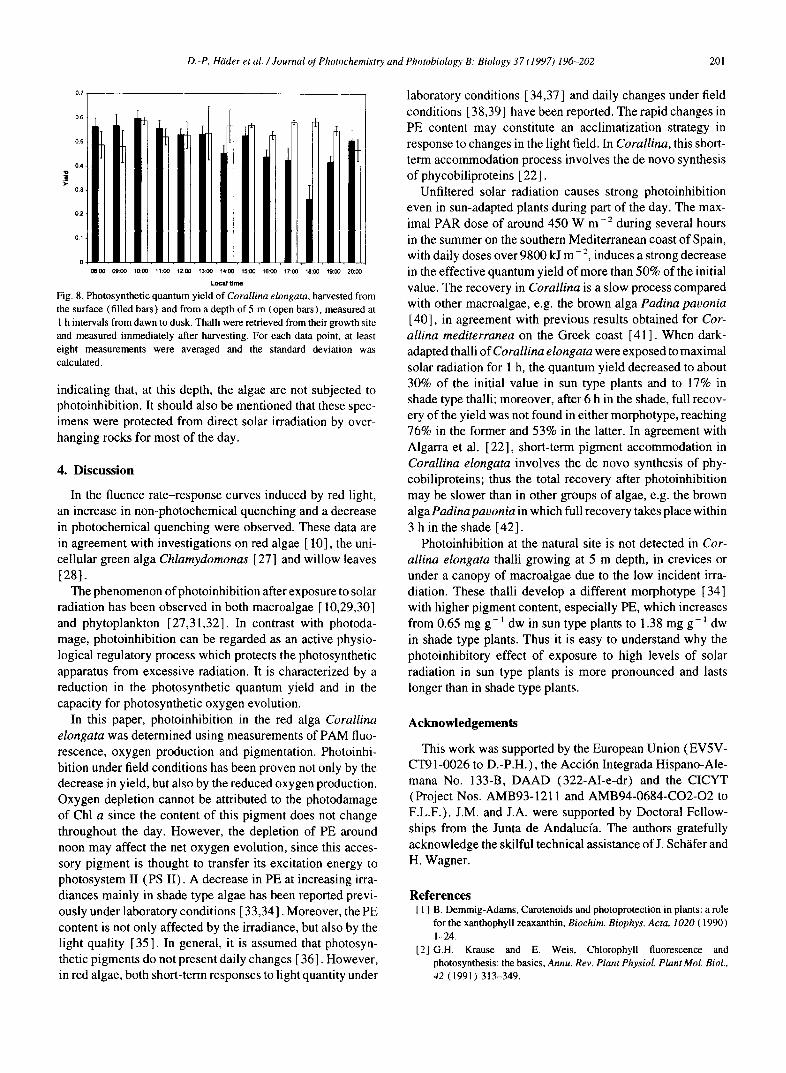
In the previous experiments, the thalli were exposed to solar irradiation in shallow water and kept in the same posi- tion so that the same part of the thallus was exposed all the time. In order to determine whether photoinhibition occurs at their natural site, where the thalli are agitated by the waves, algae were harvested again from the surface and at 5 m depth from dawn to dusk at 1 h intervals, and the yield was deter- mined immediately after harvesting (Fig. 8). The samples from the surface showed a maximal quantum yield of close to 0.6 for the first few hours; the yield then gradually decreased. The yield in the specimens harvested from a 5 m depth started at a slightly (but statistically not significant) lower level and did not change significantly during the day,
O.7
0.6
0.5
0.4
°i 111(111111 l h ~ o.sh~ lh~ 2h~ 3h~ 4h rll£ ml~ t:ohlrol
Fig. 7. Photosynthetic quantum yield of Corallina elongata, harvested from the surface (filled bars) and from a depth of 5 m (open bars), measured after dark adaptation, after exposure to solar radiation in a rock pool and during recovery (in the shade). The controls were treated in exactly the same manner as the other samples except for the exposure to solar radiation. For each data point, at least eight measurements were averaged and the standard deviation was calculated.
D.-P. Hinder et al. / Journal of Photochemistry and Photobiology B: Biology 37 (1997) 196--202 201
. . . . . i
0.6
0.5
0.4-
0.3- 0.2- 0.1-
0 . . . . .
0@:00 09:00 10:0~ 11:00 12:D0 13:00 140(I 1500 17: 1B: 19 :
Local t ime
Fig. 8. Photosynthetic quantum yield of Corallina elongata, harvested from the surface (filled bars) and from a depth of 5 m (open bars), measured at 1 h intervals from dawn to dusk. Thalli were retrieved from their growth site and measured immediately after harvesting. For each data point, at least eight measurements were averaged and the standard deviation was calculated.
indicating that, at this depth, the algae are not subjected to photoinhibition. It should also be mentioned that these spec- imens were protected from direct solar irradiation by over- hanging rocks for most of the day.
4. Discussion
In the fluence rate-response curves induced by red light, an increase in non-photochemical quenching and a decrease in photochemical quenching were observed. These data are in agreement with investigations on red algae [ 10], the uni- cellular green alga Chlamydomonas [27] and willow leaves [28].
The phenomenon of photoinhibition after exposure to solar radiation has been observed in both macroalgae [ 10,29,30] and phytoplankton [27,31,32]. In contrast with photoda- mage, photoinhibition can be regarded as an active physio- logical regulatory process which protects the photosynthetic apparatus from excessive radiation. It is characterized by a reduction in the photosynthetic quantum yield and in the capacity for photosynthetic oxygen evolution.
In this paper, photoinhibition in the red alga Corallina elongata was determined using measurements of PAM fluo- rescence, oxygen production and pigmentation. Photoinhi- bition under field conditions has been proven not only by the decrease in yield, but also by the reduced oxygen production. Oxygen depletion cannot be attributed to the photodamage of Chl a since the content of this pigment does not change throughout the day. However, the depletion of PE around noon may affect the net oxygen evolution, since this acces- sory pigment is thought to transfer its excitation energy to photosystem II (PS II). A decrease in PE at increasing irra- diances mainly in shade type algae has been reported previ- ously under laboratory conditions [ 33,34 ]. Moreover, the PE content is not only affected by the irradiance, but also by the light quality [35]. In general, it is assumed that photosyn- thetic pigments do not present daily changes [ 36 ]. However, in red algae, both short-term responses to light quantity under
laboratory conditions [ 34,37] and daily changes under field conditions [ 38,39] have been reported. The rapid changes in PE content may constitute an acclimatization strategy in response to changes in the light field. In Corallina, this short- term accommodation process involves the de novo synthesis of phycobiliproteins [ 22].
Unfiltered solar radiation causes strong photoinhibition even in sun-adapted plants during part of the day. The max- imal PAR dose of around 450 W m-2 during several hours in the summer on the southern Mediterranean coast of Spain, with daily doses over 9800 kJ m z, induces a strong decrease in the effective quantum yield of more than 50% of the initial value. The recovery in Corallina is a slow process compared with other macroalgae, e.g. the brown alga Padina paoonia [40], in agreement with previous results obtained for Cor- allina mediterranea on the Greek coast [41]. When dark- adapted thalli of Corallina elongata were exposed to maximal solar radiation for 1 h, the quantum yield decreased to about 30% of the initial value in sun type plants and to 17% in shade type thalli; moreover, after 6 h in the shade, full recov- ery of the yield was not found in either morphotype, reaching 76% in the former and 53% in the latter. In agreement with Algarra et al. [22], short-term pigment accommodation in CoraUina elongata involves the de novo synthesis of phy- cobiliproteins; thus the total recovery after photoinhibition may be slower than in other groups of algae, e.g. the brown algaPadinapavonia in which full recovery takes place within 3 h in the shade [42].
Photoinhibition at the natural site is not detected in Cor- allina elongata thalli growing at 5 m depth, in crevices or under a canopy of macroalgae due to the low incident irra- diation. These thalli develop a different morphotype [ 34] with higher pigment content, especially PE, which increases from 0.65 mg g- 1 dw in sun type plants to 1.38 mg g- 1 dw in shade type plants. Thus it is easy to understand why the photoinhibitory effect of exposure to high levels of solar radiation in sun type plants is more pronounced and lasts longer than in shade type plants.
Acknowledgements
This work was supported by the European Union (EV5V- CT91-0026 to D.-P.H.), the Acci6n Integrada Hispano-Ale- mana No. 133-B, DAAD (322-AI-e-dr) and the CICYT (Project Nos. AMB93-1211 and AMB94-0684-CO2-O2 to F.L.F.). J.M. and J.A. were supported by Doctoral Fellow- ships from the Junta de Andalucfa. The authors gratefully acknowledge the skilful technical assistance ofJ. Schiifer and H. Wagner.
References [ I ] B. Demmig-Adams, Carotenoids and photoprotection in plants: a role
for the xanthophyll zeaxanthin, Biochim. Biophys. Acta, 1020 (1990) 1-24.
[2] G.H. Krause and E. Weis, Chlorophyll fluorescence and photosynthesis: the basics, Annu. Rev. Plant Physiol. Plant Mol. Biol., 42 (1991) 313-349.

202 D.-P. Hiider et al. / Journal of Photochemistry and Photobiology B: Biology 37 (1997) 196-202
[ 3 ] R.A. Houghton and G.M. Woodwell, Global climatic change, Sci. Am.. 260 (1989) 18-26.
[4] U. Siegenthaler and J.L. Sarmiento, Atmospheric carbon dioxide and the ocean, Nature, 365 (1993) 119-125.
[5] D.-P. H~der, R.C. Worrest and H.D. Kumar, Aquatic ecosystems, UNEP Environmental Effects Panel Report, United Nations Environmental Programme, 1991, pp. 3340.
[6] D.-P. H~ider, R.C. Worrest and H.D. Kumar, Effects of increased solar ultraviolet radiation on aquatic ecosystems, UNEP Environmental Effects Panel Report, United Nations Environmental Programme, Nairobi, Kenya, 1994, pp. 71-84.
[7] R. Coutinho and R. Zingmark, Diurnal photosynthetic responses to light by macroalgae, J. Phycol., 23 (1987) 336-343.
[8] K. Gao, Diurnal photosynthetic performance of Sargassum horneri, Jpn. J. Phycol., 38 (1990) 163-165.
[9] D.R. du Preez, E.E. Campell and G.C. Bate, Photoinhibition of photosynthesis in the surf diatom, Anaulus australis Drebes et Schulz, Bot. Mar., 33 (1990) 539-543.
[10] D. Hanelt, K. Hupperts and W. Nultsch, Photoinhibition of photosynthesis and its recovery in red algae, Bot. Acta, 105 (1992) 278-284.
[ 11 ] D. Hanelt, K. Huppertz and W. Nultsch, Daily course of photosynthesis and photoinhibition in marine macroalgae investigated in the laboratory and in the field, Mar. Ecol. Prog. Ser., 97 (1993) 31-37.
[ 12] A. Trebst, A contact site between the two reaction center polypeptides of photosystem II is involved in photoinhibition, Z Naturforsch., 46 ( 1991 ) 557-562.
[ 13] C.B. Field, J.T. Ball and J.A. Berry, Photosynthesis: principles and field techniques, in R.W. Pearcy, J. Ehleringer, H.A. Mooney and P.W. Rundel (eds.), Plant Physiological Ecology, Chapman and Hall, London, 1989, pp. 209-253.
[ 14] J.M. Briantais, C. Vernotte, G.H. Krause and E. Weis, Chlorophyll a fluorescence of higher plants: chloroplasts and leaves, in Govindjee, J. Amesz and C.D. Fork (eds.), Light Emission by Plants and Bacteria, Academic Press, New York, 1986, pp. 539-583.
[ 15 ] U. Schreiber and W. Bilger, Rapid assessment of stress effects on plant leaves by chlorophyll fluorescence measurements, in J.D. Tenhunen, F.M. Catarino, O.L. Lange and W.D. Oechel (eds.), Plant Response to Stress, Springer Verlag, Berlin, Heidelberg, 1987, pp. 27-53.
[ 16] U. Schreiber and W. Bilger, Progress in chlorophyll fluorescence research: major developments during the last years in retrospect, Prog. Bot., 54 (1992) 151-173.
[17] U. Schreiber, U. Schliwa and W. Bilger, Continuous recording of photochemical and non-photochemical chlorophyll fluorescence quenching with a new type of modulation fluorometer, Photosynth. Res., 10 (1986) 51~i2.
[ 18 ] J. Kolbowski, H. Reising and U. Schreiber, Computer-controlled pulse modulation system for analysis of photoacoustic signals in the time domain, Photosynth. Res., 25 (1990) 309-316.
[ 19] C. Biichel and C. Wilhelm, In vivo analysis of slow chlorophyll fluorescence induction kinetics in algae: progress, problems and perspective, Photochem. Photobiol,, 58 (1993) 137-148.
[20] O. Bj6rkman and B. Demmig, Photon yield of O2-evolution and chlorophyll fluorescence characteristics at 77 K among vascular plants of diverse origins, Planta, 170 (1987) 489-504.
[21] B.E. Genty, J.M. Briantais and N.R. Baker, Relative quantum efficiencies of the two photosystems of leaves in photorespiratory and non-photorespiratory conditions, Plant Physiol. Biochem., 28 (1989) 1-10.
[22] P. Algarra, G. de la Vifia and F.X. Niell, Effects of light quality and irradiance level interactions on short-term pigment response of the red alga Corallina elongata, Mar. Ecol. Prog. Ser., 74 ( 1991 ) 27-32.
[23] S.W. Jeffrey and G.F. Humphrey, New spectrophotometric equations for determining chlorophylls a, b, cl and c2 in algae, phytoplankton and higher plants, Biochem. Physiol. Pflanz., 167 (1975) 191-194.
[24] S. Beer and A. Eshel, Determining phycoerythrin and phycocyanin concentrations in aqueous crude extracts of red algae, Aust. Z Mar. Freshwater Res., 36 (1985) 785-792.
[25] MM. Bradford, A rapid and sensitive method for quantitation of microgram quantities of protein utilizing the principle of protein--dye binding, Anal Biochem., 72 (1976) 248-254.
[26] U. Schreiber and W. Bilger, Progress in chlorophyll fluorescence research: major developments during the past years in retrospect, in U. Liittge and H. Ziegler (eds.), Progress in Botany, Vol. 54, Springer Verlag, Berlin, 1993, pp. 151-153.
[ 27 ] J.W. Leverenz, S. Falk, C.-M. Pilstrfm and G. Samuelsson, The effects of photoinhibition on the photosynthetic light-response curve of green plant cells (Chlamydomonas reinhardtii), Planta, 182 (1990) 161- 168.
[28] E. Ogren and M. Sj6stri3m, Estimation of the effect of photoinhibition on the carbon gain in leaves of a willow canopy, Planta, 181 (1990) 560-567.
[29] D. Hanelt, Photoinhibition of photosynthesis in marine macrophytes of the South China sea, Mar. Ecol. Prog. Sen, 82 (1992) 199-206.
[30] L.A. Franklin, G. Levavasseur, C.B. Osmond, W.J. Henley and J. Ramus, Two components of onset and recovery during photoinhibition of Ulva rotundata, Planta, 186 (1992) 399--408.
[ 31 ] E.W. Helbling, V. Villafafie, M. Ferrario and O. Holm-Hansen, Impact of natural ultraviolet radiation on rates of photosynthesis and on specific marine phytoplankton species, Mar. Ecol. Prog. Ser., 80 (1992) 89-100.
[32] H. Herrmann, F. Ghetti, R. Scheuerlein and D.-P. H~der, Photosynthetic oxygen and fluorescence measurements in Ulva laetevirens affected by solar irradiation, J. Plant Physiol., 145 ( 1995 ) 221-227.
[33] P. Algarra and F.X. Niell, Structural adaptations to light reception in two morphotypes of Corallina elongata Ellis et Soland, Mar. Ecol., 8 (1987) 253-287.
[ 34] P. Algarra and F.X. Niell, Short-term pigment response of Corallina elongata Ellis et Soland to light intensity, Aquat. Bot., 36 (1990) 127- 138.
[ 35 ] F. L6pez-Figueroa and F.X. Niell, Photocontrol of chlorophyll and biliprotein synthesis in seaweeds: possible photoreceptors and ecological considerations, Sci. Mar., 55 ( 1991 ) 519-527.
[36] M.J. Dring, Photocontrol of development in algae, Annu. Rev. Plant Physiol. Plant Mol. Biol., 39 (1988) 157-174.
[37] F. L6pez-Figueroa, Red, green and blue light photoreceptors controlling chlorophyll a, biliprotein and total protein synthesis in the red alga Chondrus crispus, Br PhycoL J., 26 ( 1991 ) 383-393.
[38] F. L6pez-Figueroa, Diurnal variation in pigment content in Porphyra laciniata and Chondrus crispus and its relation to the diurnal changes of underwater light quality and quantity, P.S.Z.N.L Mar. Ecol,, 13 (1992) 285-305.
[39] W. Rtidiger and F. L6pez-Figueroa, Yearly review. Photoreceptors in algae, Photochem. Photobiol., 55 (1992) 949-954.
[40] D.-P. H~ider, H. Herrmann and S. Regas, Effects of solar radiation and solar radiation deprived of UV-B and total UV on photosynthetic oxygen production and pulse amplitude modulated fluorescence in the brown alga Padina pavonia, FEMS MicrobioL Ecol., 19 (1996) 53- 61.
[41] D.-P. H~ider, H. Herrmann, J. Sch~fer and S. Regas, Photosynthetic fluorescence induction and oxygen production in Corallinacean algae measured on site, Bot. Acta, 109 (1996) 285-291.
[42] D.-P. Hader, M. Lebert, J. Mercado, J. Aguilera, S. Salles, A. Flores, C. Jim6nez and F. Figueroa, Photosynthetic oxygen production and PAM fluorescence in the brown alga Padina pavonia measured in the field under solar radiation, Mar. Biol., in press.