effects of serum on membrane transport ii. serum and the
TRANSCRIPT

EFFECTS OF SERUM ON MEMBRANE TRANSPORT
II. Serum and the Stimulation of Adenosine Transport,
a Possible Mechanism
PHYLLIS R. STRAUSS
From the Physiology Department, Harvard Medical School, Boston, Massachusetts 02115
ABSTRACT
When monolayers of freshly obtained rabbit lung macrophages are exposed to the nucleo-side analogue, showdomycin (sho), adenosine transport, measured over a 45 s interval, isirreversibly inhibited . Low doses of the drug or short periods of exposure, however, donot result in decreased transport, while higher concentrations or longer exposures result inexponential decline . The initial lag is not due to a long reaction time of sho with the trans-port carrier or to nonspecific sites absorbing the drug .
Previously it was shown that preincubation of monolayers with normal rabbit serum(NRS) results in increased adenosine transport . When monolayers are first exposed to shoso as to inhibit transport to varying degrees and then incubated with NRS, transport isincreased over the inhibited level . Several experiments make it unlikely that serum removesthe drug from the cell surface in a nonspecific fashion . Moreover, serum given before,during, or after sho alters the dose response curve so that no shoulder is seen . One way toexplain these results makes use of target theory : the adenosine transport system could becomprised mainly of "coupled" or "clustered" sites of which only one is active at anytime as well as "hidden" sites which are inactive . When a site in a group is irreversiblyinactivated by sho, another in the group becomes activated . Serum might activate or un-couple all sites and also cause the appearance of hidden sites, which previously neithertransported nor bound sho .
INTRODUCTION
Numerous authors have characterized transportof electrolytes and nonelectrolytes in a variety ofmammalian cells in terms of kinetic parameters,sodium requirements, and specificity (4, 5, 7, 8,15, 26, 33, 34) . In addition, the activities of mem-brane transport systems are altered by pH,' tem-perature (1, 27), internal concentrations of var-ious metabolites (15, 27), and the external medium
' Taube, R., and R. D. Berlin. Manuscript in prep-aration .
bathing the cell (7, 13, 22, 32) . Still there is littleinformation regarding detailed regulatory mech-anisms .
Recently we reported the effects of normalrabbit serum (NRS) on two transport systemsin rabbit lung macrophages (32) . The rate oflysine transport was depressed from the controland that of adenosine was enhanced by prein-cubating cells with NRS for 15-30 min . Thesubstance(s) responsible for the depression was
THE JOURNAL OF CELL BIOLOGY . VOLUME 60, 1974 • pages 571-585
571
on January 8, 2019jcb.rupress.org Downloaded from http://doi.org/10.1083/jcb.60.3.571Published Online: 1 March, 1974 | Supp Info:

(were) small and heat stable, while that (those)responsible for the enhancement was (were)larger and heat labile. In this study we haveexamined in greater detail the serum-inducedstimulation of adenosine transport . In the pre-vious publication, the kinetic data were insufficientto determine whether the number of transportsites, the rapidity of transport, or the affinity ofthe carrier for its substrate had been altered byprevious exposure to serum . In this series of ex-periments the nucleoside analogue showdomycin(sho), originally described by Nishimura et al .(25), was used to irreversibly inhibit the nucleo-side transport system . Both time and concen-tration curves had an interval at early times orlow concentrations where transport did not de-cline . Thereafter, transport fell in an exponentialfashion . Several experiments ruled out the pos-sibility that lag was due to the reaction time ofsho with the transport carrier or to nonspecificsites absorbing the drug . Serum treatment, whilestimulating transport, abolished the interval overwhich transport rates were unaffected by sho .It also caused increased transport in cells alreadyinhibited irreversibly with sho. One way to ex-plain these results makes use of target theory. Theassumptions required for application of this theoryto the data are shown to be valid . From targettheory it is inferred that the adenosine transportsystem might be composed of "coupled" sites ofwhich only one is active at any time and "hidden"sites which are inactive . When a site in a group isirreversibly inactivated by sho, another becomesactivated. Serum might activate all sites and alsocause the appearance of previously hidden sites .
Coupling is consistent with certain models ofenzyme activity where only one site acts on sub-strate at a time, even though all subunits areidentical (21) . Hidden sites are consistent withpublished accounts of regulation of surface ele-ments in fused cells (11, 12) and in lymphocytes(37) . Implicit in the model is the assumption ofmobility of transport carriers within the plasmamembrane as suggested by Singer and Nicolson(31) . The assumption is supported by experi-ments dealing with the effect of NRS on prefixedcells . If the hypothesis is valid, this is the firstdemonstration of a functional constitutent of thecell surface that is regulated in this dual fashion .
MATERIALS AND METHODS
Previous publications have described how rabbitlung macrophages (34) and sera of plasma (32)
572 THE JOURNAL OF CELL BIOLOGY • VOLUME 60, 1974
were obtained . Cells were tested for their ability totransport adenosine by the cover slip technique de-veloped by Hawkins and Berlin (14) . Briefly, mono-layers were formed from freshly obtained lung mac-rophages on circular 22-mm cover slips . Each mono-layer contained 3 X 10 5 cells . The monolayers wereexposed to the materials to be tested for the desiredlength of time at 37 °C unless otherwise stated . Be-tween exposures to different solutions, monolayerswere rinsed through four changes of modified Hanks'solution (MH) (33) at room temperature unless arinse at 4 ° C is indicated . Finally they were tested forthe ability to transport adenosine (0 .1 mM, 2-4sCi/ml) at 37 ° C during a 45 s incubation period inMH. Every determination was run in triplicate orquadruplicate . The concentration of adenosinechosen for testing transport was its Km value (32,35) . Isotopes were obtained from New EnglandNuclear, Boston, Mass., or from Schwarz-MannDiv., Becton, Dickinson and Co ., Orangeburg, N . Y.No serum was present during the transport assayexcept presumed substances remaining adherent tothe macrophage monolayer . Sho was also obtainedfrom Schwarz-Mann.
Metabolism of [ 3H]adenosine taken up during a45 s incubation was determined by analysis of per-chloric acid (PCA) extracts of cells by means ofascending chromatography. After the monolayerswere exposed to isotope, they were frozen and thawedthree times in 5% iced PCA which was then neutral-ized with KOH . Precipitated salt was removed with aSwinnex filter, Millipore Corp ., Bedford, Mass ., porediameter 0.45 µm, and the extracts were stored frozenat -20 °C. Ascending chromatography using What-man paper # 1 was performed in isobutyric acid : am-monium hydroxide : water, 66 :1 :33 with standards ofadenosine, AMP, ADP, ATP, and inosine. The areaof the unknown corresponding to each standard wascut and eluted with 0 .05 N NaOH, and radioactivityin each eluate was determined . As inosine and AMPtended to run together, half the eluate from that frac-tion was rerun by ascending chromatography in theupper phase from a mixture of water :sec-butylalcohol : tert-butyl alcohol, 48 .4 :43 :8 .6, as describedby Fink et al . (10) . Strips corresponding to the stand-ards of inosine and AMP were eluted and counted asabove. In one experiment [ 3H]inulin was included inorder to correct for the amount of fluid trapped in theextracellular space. Isotope trapped in the extracellu-lar space would not be metabolized, and thereforethat amount of [3H]adenosine corresponding to theamount of trapped fluid was subtracted from countsrecovered as adenosine . In normal cells this amountedto 5% total counts and in sho-inhibited cells to 25% .Recovery of total counts from the paper was 100% inall cases . In order to determine the total amounts ofadenine nucleotides in buffer and serum-treatedcells, high pressure liquid chromatography wasperformed on PCA extracts prepared as above with
liquid chromatograph, Varian Aerograph, LCS1000 (Varian Associates, Palo Alto, Calif) (2) .For fluorescence microscopy a Zeiss microscope
equipped with FLIC primary filters manufacturedby Optisk Laboratorium, Lyngby, Denmark wasused. Fluorescein-conjugated concanavalin A waskindly supplied by Dr . Emil Unanue .
RESULTS
Sho Irreversibly InhibitsAdenosine Transport
Komatsu (17) reported that sho inhibitednucleoside transport in Escherichia coli if the ana-logue was present during the exposure to nucleo-side. In rabbit lung macrophages, sho also in-hibited adenosine transport when it was presentduring the 45 s exposure to [3H-]adenosine. Fig . 1is a Lineweaver-Burk plot showing that the in-hibition was competitive in nature, as the twocurves intersected at the ordinate . From thisexperiment it could be calculated that the Ki forhalf-maximal inhibition was 1 .2 mM sho (seereference 33 for calculation). In contrast to thefindings of Komatsu, however, after a 30 minpreincubation with 0 .1 mM sho, transport wasinhibited irreversibly . Reversal in the absence ofsho could not be demonstrated for at least 60 min .Under these conditions the kinetic data indicatednoncompetitive kinetics for the effect of sho onadenosine transport . In the experiment shown inFig. 2 the Vmax for adenosine transport, deter-mined from the intercepts at the ordinate, was
-30 -20 30 40 50 60-10
a 20ADENOSINE (mM - ' )
FIGURE 1 Lineweaver-Burk plot of [ 3H]adenosinetransport in the presence (o) or absence (0) of 0.5 mMsho. Transport was tested over a 45 s interval . Inhibi-tion is competitive . The calculated K ; is 1 .2 mM. Linesdrawn by the method of least squares .
reduced to 570 of control value by ;2 h exposureto 0.4 mM sho, while the Km, calculated fromthe respective slopes, was unaffected . Thymidine,which has a high affinity for the adenosine carrier,(K; = 0.18 mM [1]), completely prevented in-hibition (Table I) .Komatsu and Tanaka (19) reported that in
E. coli sho inhibits the metabolism of nucleosides .In contrast, using the time and concentrationsindicated, this did not seem to occur to [3H]-adenosine taken up during a 45 s pulse in lung mac-rophages . Adenosine transported during the testinterval was metabolized by normal and drug-treated monolayers in a similar fashion in anexperiment where transport was inhibited to12 .7% of control by ;2 h preincubation with0.3 mM sho (Table II) . That 807, of adenosinetaken up was metabolized to nucleotides in bothtreated and control monolayers provided evi-dence that membrane transport was rate limitingafter exposure to sho as it was in the untreatedcells . Also, when transport was inhibited to 870of control by a 30 min exposure to 0 .4 mM sho,transport was still linear for more than 1 min .Thus, sho irreversibly inhibited the transport ofadenosine without interfering with its metabolismover the transport time and drug concentrationranges employed .
Time-Course of Inactivation ofNucleoside Transport
When cells were incubated with sho for varyingperiods of time, rinsed, and then tested for nucleo-side transport, little or no decrease occurreduntil after a lag of several minutes . After this in-terval the transport rate declined exponentially(Fig . 3) . Even at sho concentrations as high as2.0 mM, well above the K i concentration, therewas a distinct lag before inactivation began . Thelag could correspond to saturation by sho ofcellular sites unrelated to the nucleoside transportsystem. However, when monolayers were pre-incubated for 30 min with 0 .2 mM sho plus 0.4mM adenosine so as to saturate such reactivegroups while protecting nucleoside transportsites, then rinsed and incubated with 0 .2 mMsho for varying lengths of time before assaying fortransport, the time-course of inactivation showedthe usual lag time .
The possibility that the lag was due to slowreaction leading to irreversible combination ofsho with the nucleoside carrier was ruled out by
PHYLLIS R . STRAUSS Effects of Serum on Membrane Transport . II 573

aQ
40
xÔ 30
Ia
20
Io
50
PRE14CUBATION - MH
100
500
000
10
50
V/concn
V/concn
FIGURE 2 Woolf plot of [3H]adenosine transport by monolayers preincubated for 80 min with or without0.4 mM sho. Transport measured over a 45 s interval . Velocity is plotted against velocity divided by theconcentration of adenosine . As the slopes are parallel but the intercepts differ, inhibition is noncompeti-tive. Lines drawn by the method of least squares .
TABLE I
a shoulder at low concentration (Fig . 5) . ThisProtection of Adenosine Transport
was made especially clear when contrasted withCarrier against Sho
the dose dependence for monolayers treated withserum, as will be discussed in detail below .
TABLE II
Metabolism of [ 3H] Adenosine
5 7 4
THE JOURNAL OF CELL BIOLOGY • VOLUME 60, 1974
4
2
PRENCUBATION , 0 .4mM SHO
% DPM recovered*
Monolayers were preincubated with or without shoin the presence or absence of thymidine .
an experiment in which cells were incubated withsho until inactivation had begun, rinsed, andthen reincubated with sho . The second incubationled immediately to further inactivation (Fig . 4) .
Dose Dependency of Inactivation ofNucleoside Transport
When monolayers were incubated with varyingconcentrations of sho for a uniform time period(30 min) and a plot of percent remaining transportvs . concentration prepared, this curve also showed
*PCA extracts were prepared after monolayerswere preincubated for 30 min under the variousconditions above and tested for the transport ofadenosine (0.04 mM, 10 µCi/m1) for 45 s .Metabolites of [ 3 H] adenosine were separated byascending paper chromatography as described inMethods. The strips corresponding to standardsrun simultaneously were eluted and counted .Control monolayers transported 17 .8 pmol/45 s/3 X10 5 cells .
ThymidineSho (0.2 mM)
(0.4 mM)
pmol adenosinetransported/3 X 10 5/
cells/min
25 .815 .3
+ 22 .4+
+ 21 .8 Preincubation(mM Sho) Nucleotides Inosine Adenosine
0 79 .7 5.6 14 .10 .05 85.1 6 .8 8 .00 .30 78.3 12 .2 9 .4

Serum Alters the Response to Sho
Previously we (32) reported that when mono-layers were exposed to NRS, adenosine transportwas stimulated by 35 f 2 .3% (SE, 60 observa-tions) . In order to determine whether serum wasrecruiting new transport sites or affecting oldones, monolayers were preincubated for 30 minwith different concentrations of sho to irreversibly
I
0.1
02
0.3SHOWDOMYCIN (mM)
FIGURE 5 The effect of sho on adenosine transportwith (s) or without (O) serum (NRS) preincubation .Monolayers were preincubated with MH or 5% NRSfor 30 min, washed, and incubated with MH or Shoat the concentration indicated for 80 min . They werethen washed and tested for the ability to transport[3H]adenosine over a 45 s interval. Typical experimentof more than 12 .
mM SHOx-0.05-0.20o-0.40a-2.00
10
20
30 40
50TIME OF EXPOSURE (min)
FIGURE 3 The effect of sho on adenosine transport,time dependence. A short lag precedes decline of trans-port . Dashed lines are the extrapolation to 0 min expo-sure to sho at each concentration of drug .
10
20
30
40TIME OF SECOND EXPOSURE min
(0.25mM SHO)
FIGURE 4 Time dependence of loss of adenosine trans-port due to sho when monolayers have previously beenexposed to sho. Monolayers were treated with 0 .25 mMsho or MH for 15 min, washed through four changesMH, and then further incubated with 0 .25 mM sho orMH for the indicated period . No lag is seen beforetransport declines .
I
inactivate transport to varying degrees and thenpostincubated with MH or 5 % NRS. TableIII lists the data for the eight experiments graphedin Fig. 6 . From the table one can see that treatingmonolayers with NRS rather than MH aftertransport has been inhibited by sho resulted inenhanced levels of adenosine uptake. The in-crement observed when monolayers were treatedfirst with MH and then with NRS is designatedaMH. This stimulation ranged from 18% to 178%(49 f 25%, SE, eight observations) of controlstreated in both intervals by MH . The incrementobserved when monolayers were treated firstwith various concentrations of sho and then withNRS is designated °Sho . This stimulation rangedfrom 15% to 840% (227 f 99%, SE, eight ob-servations) of controls treated first with sho andthen with MH. Were the serum-induced incre-ments dependent on the level of transport remain-ing when serum was applied, °Sho would haveshowed the same percentage increase of its controlnot treated with serum as °MH . The last columnin Table III details the increase for that hy-pothetical case in each experiment . One can seethat the observed values do not follow this predic-tion. Therefore, it was unlikely that the incre-ment induced by NRS after sho was due to con-formational change or to more rapid activity oftransport sites that were left after inhibition .
PHYLLIS R. STRAUSS Effects of Serum on Membrane Transport . II 575
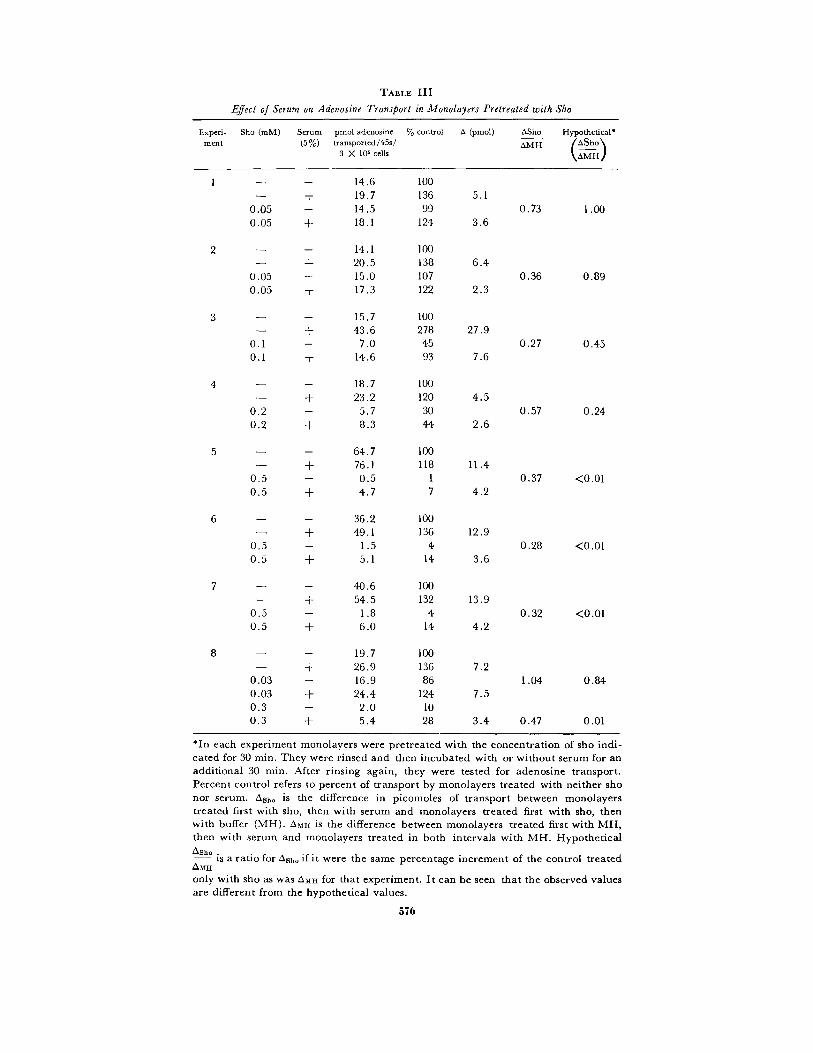
TABLE III
Effect of Serum on Adenosine Transport in Monolayers Pretreated with Sho
*In each experiment monolayers were pretreated with the concentration of sho indi-cated for 30 min . They were rinsed and then incubated with or without serum for anadditional 30 min . After rinsing again, they were tested for adenosine transport .Percent control refers to percent of transport by monolayers treated with neither shonor serum. Asho is the difference in picomoles of transport between monolayerstreated first with sho, then with serum and monolayers treated first with sho, thenwith buffer (MH) . A mil is the difference between monolayers treated first with MH,then with serum and monolayers treated in both intervals with MH . HypotheticalAsh. .is a ratio for ash_ if it were the same percentage increment of the control treatedAMHonly with sho as was OMH for that experiment . It can be seen that the observed valuesare different from the hypothetical values .
5 7 6
Experi- Sho (mM)
Serum pmol adenosine % control a (pmol) tsSho Hypothetical*ment
(5%) transported/45s/3 X 105 cells
OMH AShoAMH
1
-
- 14 .6 100+ 19 .7 136 5.1
0.05
- 14 .5 99 0 .73 1 .000.05
+ 18 .1 124 3.6
2
-
- 14.1 100+ 20.5 138 6 .4
0.05
- 15 .0 107 0.36 0 .890.05
+ 17 .3 122 2 .3
3
-
- 15 .7 100-
+ 43 .6 278 27 .90 .1
- 7 .0 45 0.27 0 .450 .1
+ 14 .6 93 7 .6
4
- 18 .7 100- + 23 .2 120 4 .5
0 .2
- 5 .7 30 0.57 0 .240 .2
+ 8 .3 44 2 .6
5
-
- 64.7 100+ 76 .1 118 11 .4
0 .5
- 0 .5 1 0 .37 <0.010 .5
+ 4 .7 7 4 .2
6
-
- 36 .2 100+ 49 .1 136 12 .9
0 .5
- 1 .5 4 0 .28 <0.010 .5
+ 5 .1 14 3 .6
7
-
- 40 .6 100+ 54 .5 132 13 .9
0 .5
- 1 .8 4 0 .32 <0 .010 .5
+ 6 .0 14 4 .2
8
-
- 19.7 100+ 26 .9 136 7 .2
0 .03
- 16 .9 86 1,04 0.840 .03
+ 24 .4 124 7 .50 .3
- 2 .0 100.3
+ 5 .4 28 3 .4 0.47 0.01
o4sali 0.6 O.-3b o.fo o.4bSHOWDOMYCIN (mM)
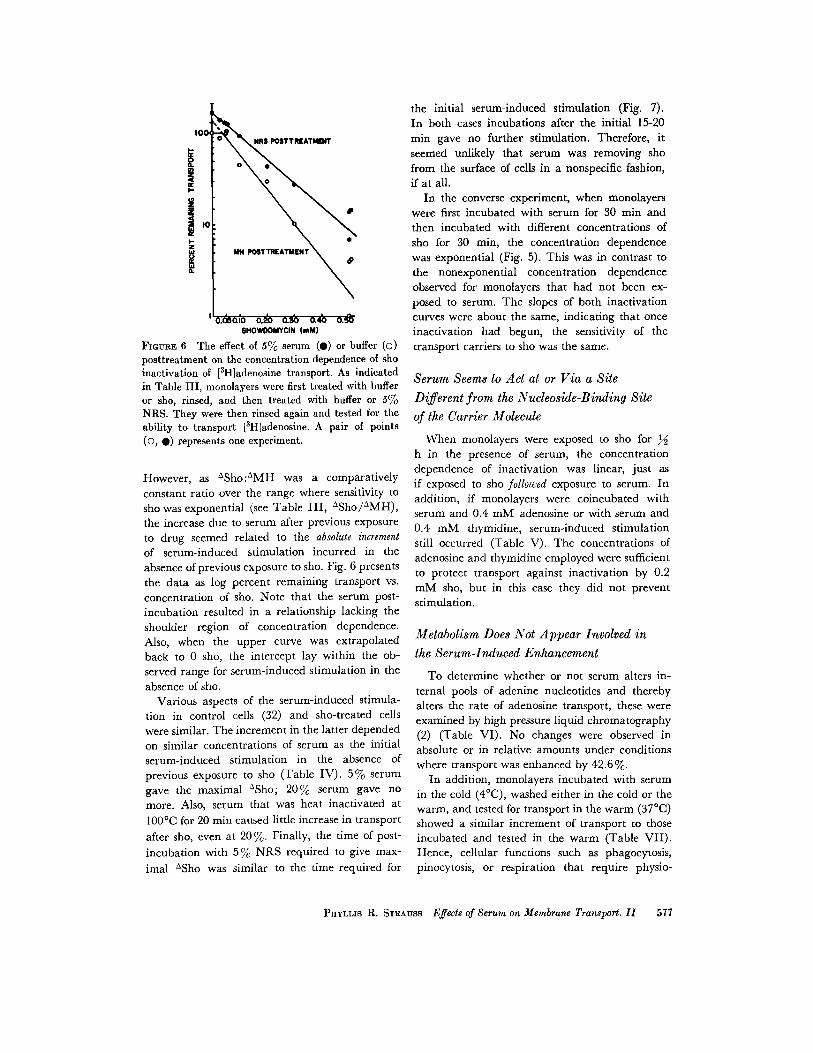
FIGURE 6 The effect of 5% serum (0) or buffer (o)posttreatment on the concentration dependence of shoinactivation of [ 3H]adenosine transport. As indicatedin Table III, monolayers were first treated with bufferor sho, rinsed, and then treated with buffer or SoloNRS. They were then rinsed again and tested for theability to transport [3 H]adenosine . A pair of points(O, •) represents one experiment .
However, as °Sho :°MH was a comparativelyconstant ratio over the range where sensitivity tosho was exponential (see Table III, °Sho/°MH),the increase due to serum after previous exposureto drug seemed related to the absolute incrementof serum-induced stimulation incurred in theabsence of previous exposure to sho . Fig. 6 presentsthe data as log percent remaining transport vs .concentration of sho. Note that the serum post-incubation resulted in a relationship lacking theshoulder region of concentration dependence .Also, when the upper curve was extrapolatedback to 0 sho, the intercept lay within the ob-served range for serum-induced stimulation in theabsence of sho .
Various aspects of the serum-induced stimula-tion in control cells (32) and sho-treated cellswere similar. The increment in the latter dependedon similar concentrations of serum as the initialserum-induced stimulation in the absence ofprevious exposure to sho (Table IV) . 5% serumgave the maximal °Sho ; 20% serum gave nomore. Also, serum that was heat inactivated at100°C for 20 min caused little increase in transportafter sho, even at 20 % . Finally, the time of post-incubation with 5 % NRS required to give max-imal °Sho was similar to the time required for
the initial serum-induced stimulation (Fig . 7) .In both cases incubations after the initial 15-20min gave no further stimulation . Therefore, itseemed unlikely that serum was removing shofrom the surface of cells in a nonspecific fashion,if at all .
In the converse experiment, when monolayerswere first incubated with serum for 30 min andthen incubated with different concentrations ofsho for 30 min, the concentration dependencewas exponential (Fig . 5) . This was in contrast tothe nonexponential concentration dependenceobserved for monolayers that had not been ex-posed to serum . The slopes of both inactivationcurves were about the same, indicating that onceinactivation had begun, the sensitivity of thetransport carriers to sho was the same .
Serum Seems to Act at or Via a Site
Different from the Nucleoside-Binding Siteof the Carrier Molecule
When monolayers were exposed to sho forh in the presence of serum, the concentrationdependence of inactivation was linear, just asif exposed to sho followed exposure to serum . Inaddition, if monolayers were coincubated withserum and 0 .4 mM adenosine or with serum and0.4 HIM thymidine, serum-induced stimulationstill occurred (Table V). The concentrations ofadenosine and thymidine employed were sufficientto protect transport against inactivation by 0 .2HIM sho, but in this case they did not preventstimulation .
Metabolism Does Not Appear Involved inthe Serum-Induced Enhancement
To determine whether or not serum alters in-ternal pools of adenine nucleotides and therebyalters the rate of adenosine transport, these wereexamined by high pressure liquid chromatography(2) (Table VI) . No changes were observed inabsolute or in relative amounts under conditionswhere transport was enhanced by 42 .6% .
In addition, monolayers incubated with serumin the cold (4 °C), washed either in the cold or thewarm, and tested for transport in the warm (37 °C)showed a similar increment of transport to thoseincubated and tested in the warm (Table VII) .Hence, cellular functions such as phagocytosis,pinocytosis, or respiration that require physio-
PHYLLIS R. STRAUSS Effects of Serum on Membrane Transport . II
577
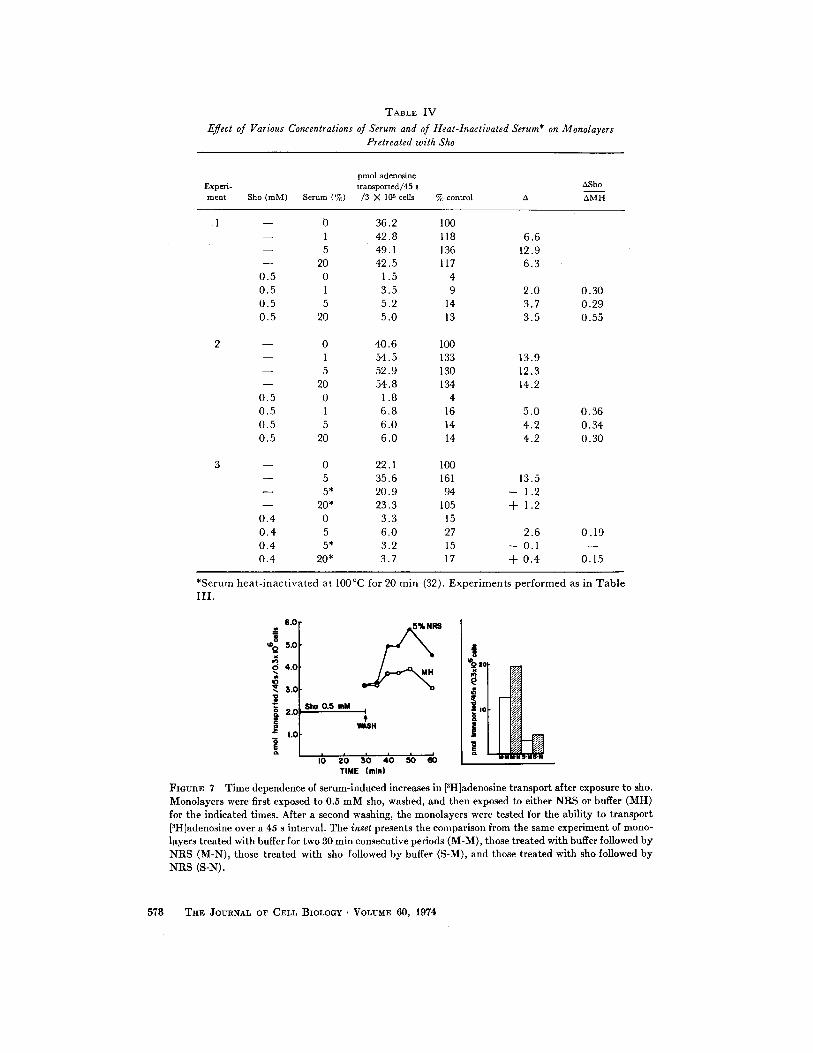
TABLE IV
Effect of Various Concentrations of Serum and of Heat-Inactivated Serum* on MonolayersPretreated with Sho
*Serum heat-inactivated at 100 ° C for 20 min (32) . Experiments performed as in TableIII .
6 .0
•
5 .0kMd 4.0
a 3 .030 2 .0S2
1 .00En
Sho 0.5 mm i
fWASH
578
THE JOURNAL OF CELL BIOLOGY . VOLUME 60, 1974
10 20• 30 40 50 60TIME (min)
FIGURE 7 Time dependence of serum-induced increases in [3H]adenosine transport after exposure to sho .Monolayers were first exposed to 0.5 mM sho, washed, and then exposed to either NRS or buffer (MH)for the indicated times . After a second washing, the monolayers were tested for the ability to transport[3H]adenosine over a 45 s interval . The inset presents the comparison from the same experiment of mono-layers treated with buffer for two 30 min consecutive periods (M-M), those treated with buffer followed byNRS (M-N), those treated with sho followed by buffer (S-M), and those treated with sho followed byNRS (S-N) .
Expert-pmol adenosinetransported/45 s ASho
ment Sho (mM) Serum (%) /3 X 105 cells % control A OMH
1 0 36 .2 1001 42 .8 118 6 .65 49 .1 136 12 .9
- 20 42 .5 117 6 .30.5 0 1 .5 40 .5 1 3 .5 9 2 .0 0 .300 .5 5 5 .2 14 3 .7 0 .290.5 20 5 .0 13 3 .5 0 .55
2 0 40 .6 1001 54 .5 133 13 .95 52 .9 130 12 .3
- 20 54 .8 134 14 .20.5 0 1 .8 40 .5 1 6 .8 16 5 .0 0.360.5 5 6 .0 14 4 .2 0 .340.5 20 6 .0 14 4 .2 0 .30
3 0 22 .1 1005 35 .6 161 13 .5
- 5* 20 .9 94 - 1 .2- 20* 23 .3 105 + 1 .20.4 0 3 .3 150.4 5 6 .0 27 2 .6 0 .190.4 5* 3 .2 15 - 0 .1 -0.4 20* 3 .7 17 + 0 .4 0 .15
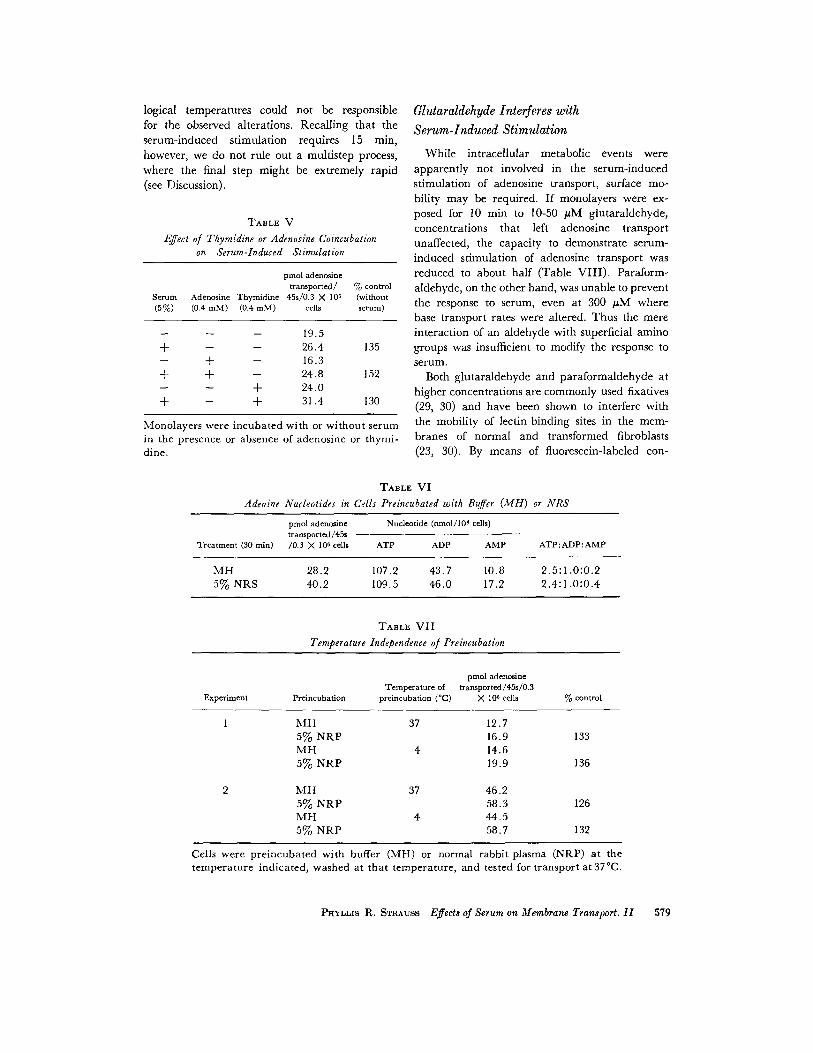
logical temperatures could not be responsiblefor the observed alterations. Recalling that theserum-induced stimulation requires 15 min,however, we do not rule out a multistep process,where the final step might be extremely rapid(see Discussion) .
TABLE V
Effect of Thymidine or Adenosine Coincubationon Serum-Induced Stimulation
Monolayers were incubated with or without serumin the presence or absence of adenosine or thymi-dine .
TABLE VI
Adenine Nucleotides in Cells Preincubated with Buffer (MH) or NRS
pmol adenosine
Nucleotide (nmol/l06 cells)transported/45s
Treatment (30 min) /0.3 X 106 cells
ATP
ADP
AMP
ATP :ADP:AMP
Glutaraldehyde Interferes with
Serum-Induced Stimulation
While intracellular metabolic events wereapparently not involved in the serum-inducedstimulation of adenosine transport, surface mo-bility may be required . If monolayers were ex-posed for 10 min to 10-50 µM glutaraldehyde,concentrations that left adenosine transportunaffected, the capacity to demonstrate serum-induced stimulation of adenosine transport wasreduced to about half (Table VIII) . Paraform-aldehyde, on the other hand, was unable to preventthe response to serum, even at 300 AM wherebase transport rates were altered . Thus the mereinteraction of an aldehyde with superficial aminogroups was insufficient to modify the response toserum .
Both glutaraldehyde and paraformaldehyde athigher concentrations are commonly used fixatives(29, 30) and have been shown to interfere withthe mobility of lectin-binding sites in the mem-branes of normal and transformed fibroblasts(23, 30) . By means of fluorescein-labeled con-
Cells were preincubated with buffer (MH) or normal rabbit plasma (NRP) at thetemperature indicated, washed at that temperature, and tested for transport at 37 °C .
PHYLLIS R . STRAUSS Effects of Serum on Membrane Transport . II
579
MH5% NRS
28.2
107 .2
43 .740.2
109.5
46 .010.817 .2
2 .5 :1 .0 :0 .22 .4 :1 .0 :0 .4
TABLE VII
Temperature Independence of Preincubation
Experiment PreincubationTemperature of
preincubation (°C)
pmol adenosinetransported /45s/0.3
X 106 cells % control
1 MH 37 12 .75% NRP 16 .9 133MH 4 14 .65% NRP 19 .9 136
2 MH 37 46 .25% NRP 58 .3 126MH 4 44 .55% NRP 58 .7 132
Serum Adenosine Thymidine(5%)
(0.4 mM) (0 .4 mM)
pmol adenosinetransported/45s/0.3 X 106
cells
% control(withoutserum)
- 19 .5- 26.4 135
+
- 16 .3+
- 24 .8 152+ 24.0+ 31 .4 130

TABLE VIII
Effects of Aldehydes on Serum-Induced Stimulation of Adenosine Transport
canavalin A (FCCA), it was possible to demon-strate in experiments parallel to those describedabove that exposure of monolayers to 20 AMglutaraldehyde but not to 300 AM paraformalde-hyde was sufficient to interfere with the overallredistribution of fluorescence . Monolayers weretreated with the appropriate fixative or with MHfor 10 min at 37 °C, washed, exposed to FCCAat 4°C for 10 min, rinsed, wet mounted at roomtemperature, and examined at various intervalsup to 1 h. Within 5 min, fluorescence due toFCCA in unfixed cells appeared in a ring in theperinuclear region and as clumps or patches
580
THE JOURNAL OF CELL BIOLOGY • VOLUME 60, 1974
Monolayers were treated with buffer (NIH), paraformaldehyde (PFA), or glutaralde-hyde (GLUT), rinsed, and then treated with either MH or 570 NRS .
over the nucleus. Cells fixed with I c/ (100 mM)glutaraldehyde before the application of FCCAretained a diffuse distribution over cytoplasmand nucleus, though occasional patches of dyewere seen over cytoplasm and nucleus . Cellsfixed with 300 µM paraformaldehyde and thenexposed to FCCA resembled the unfixed controls :fluorescence rapidly disappeared from the cyto-plasm of most cells and appeared as a perinuclearring and/or stippled over the nucleus . Cells ex-posed first to 20 µM glutaraldehyde and then toFCCA retained much diffuse cytoplasmic flu-orescence, even 1 h after FCCA was removed .
Experiment Treatment I (10 min)Treatment II
(30 min)% control (no
aldehyde, no serum)
% control (ifaldehyde, no
serum)
1
µM
MH MH 100MH NRS 164
PFA, 300 MH 89 100PFA, 300 NRS 141 158
GLUT, 5 MH 102 100GLUT, 5 NRS 136 133
2 MH MH 100MH NRS 169
GLUT, 20 MH 101 100GLUT, 20 NRS 134 133
3 MH MH 100MH NRS 180
GLUT, 10 MH 103 100GLUT, 10 NRS 157 153
GLUT, 100 MH 54 100GLUT, 100 NRS 73 124
4 MH MH 100MH NRS 170
GLUT, 10 MH 101 100GLUT, 10 NRS 155 153
GLUT, 50 MH 114 100GLUT, 50 NRS 174 152

However, some cells acquired a perinuclear ringand some patching occurred . Thus, the low con-centration of glutaraldehyde that reduced thecapacity of cells to respond to serum was sufficientto alter the gross mobility characteristics of thecell surface, while high concentrations of para-formaldehyde did not affect the serum responseand also had no effect on the mobility of FCCA .
DISCUSSION
Preincubation of rabbit lung macrophages withNRS for 15 min stimulates adenosine transport(32) . Here it is shown that serum does not seemto act at or via the nucleoside-binding site of thecarrier molecule, as inclusion of either thymidineor adenosine with NRS during preincubationdoes not alter the induced stimulation . Serumdoes not alter adenine nucleotide pools . As itseffect is observed even if cells are preincubatedand washed at 4 °C, temperature-sensitive cel-lular functions such as phagocytosis and pino-cytosis are apparently not responsible . Cyto-plasmic and mitochondrial protein syntheses areprobably not involved in the response, as ex-posure to 5 µg/ml cycloheximide and/or up to500 µg/ml chloramphenicol do not alter theserum-induced stimulation (unpublished results) .
In order to determine whether the activity ofold transport sites was altered or if new sites wererecruited in response to serum, an inhibitor ofadenosine transport, sho, was used . When rabbitlung macrophages are exposed to sho, using thetime and concentrations described, sho acts as anirreversible inhibitor of adenosine transport butdoes not alter the metabolism of adenosine takenup during the 45 s transport period . This observationis in contrast with the drug's effects on E . coliwhere sho inhibits nucleoside metabolism (17, 18,19) . Our evidence indicates that sho acts on theadenosine transport system at or via the transportsite, as coincubation with thymidine or adenosineprevents the inhibition of transport . Even so,concentration (Figs. 5, 6) and time (Fig . 3) de-pendence curves show shoulders at early timesand at low concentrations .
Serum can induce an increment in adenosinetransport after adenosine transport has beenreduced by sho. The increment does not seem tobe due to nonspecific removal of sho from thecell surface as the optimal serum concentration(Table IV) and incubation time (Fig . 7) are thesame for the serum-induced stimulation regardless
of whether sho has been applied . In both casesexposure to higher concentrations of serum, heat-inactivated serum, or longer times does not alterthe response . The percentage increase when sho-treated cells are then treated with serum is muchgreater than the percentage increase due to serumin control cells . In fact, the absolute amount ofthe increment is independent of transport remain-ing after sho inhibition . It can, however, becorrelated with the magnitude of serum stimula-tion in control cells (Table III and Fig . 6) . As shounder control conditions irreversibly inhibitsadenosine transport and as the serum-inducedincrement is independent of transport remainingwhen cells are pretreated with sho, whateverchanges that serum causes are unlikely to involvethe transport sites remaining after sho . In otherwords, conformational change in the remainingtransport carriers or more rapid movement bythe carriers is unlikely to be primarily responsiblefor serum-induced changes . Moreover, exposureto serum alters the relationship of sho inhibitionof adenosine transport to sho concentration sothat the nonresponsive portion, as indicated bythe shoulder, is not seen .
The irreversible inhibitor need not be a nucleo-side analogue . Preincubation with p-chloro-mercuribenzosulfonic acid, a sulfhydryl reagentknown to inactive adenosine transport (34), fol-lowed by exposure to serum, also leads to an in-crease over controls treated with the sulfhydrylreagent followed by buffer. Again, the time andconcentration requirements for the increment ininhibitor-treated cells are the same as for controls(unpublished data) .
The data pertaining to sho sensitivity and theeffect of serum thereon are presented as log per-cent remaining transport vs . concentration of shoor vs . time of exposure to sho. From the plots onecan see that transport diminishes in an exponentialfashion except in the case of cells not exposed toNRS, where a shoulder is seen at early times andat low concentrations. Data which assume suchan appearance can be interpreted on the basis oftarget theory so long as they satisfy the require-ments of randomness of hit and discreteness oftarget (9) . One might consider individual trans-port carriers as individual targets, thus satisfyingthe discreteness requirement. A plot of data fromvarious experiments (Fig . 8) of the time requiredto inactivate 37 jo of transport vs . the i • iprocalof the dose of sho (D 37) shows that the time-dose
PHYLLIs R . STRAUSS Effects of Serum on Membrane Transport . II
581

relationship is constant . That is, the interactionof sho with the transport carrier can be con-sidered a random event . Under these conditionsone can consider the probability P of a givendrug molecule interacting with a given transportsite, as in footnote 2 .
Presented in this form, log percent-remainingtransport appears to be a function of concen-
2 The probability that after n hits any target willhave been hit m times and not hit (n-m) times willbe
n!P{M}= ml(n- m)1pmgn-m .
(1)
For p very small, such that np is small, this can beapproximated by the Poisson distribution :
ÂmeP{M} = , where X = np . (2)
m!
Relating target theory to the deterministic descriptionof exponential events, then, if a process exhibits ex-ponential reduction, the incremental change intransport, dN, can be described mathematically fora single variable by
dN = -cladt (Eq. 3)
which, when integrated, can be expressed as
N = Noe-coat (Eq . 4),
where c1 , is the initial concentration of drug, a isthe sensitivy of transport to drug, N is the trans-port remaining at time t, and No is initial transport .This can be related to unhit targets by setting m = 0in the Poisson description above, then,
tration of drug (c), time of exposure (t), andsensitivity of transport sites to sho (a) . When ashoulder is observed as well, as is the case here,target theory offers two models : a single-hit,multitarget model and a multihit, single-targetmodel (9) . 3
Single-hit, multitarget models involve a cluster-ing or coupling of targets . In terms of these data,then, some adenosine transport sites would existin the plasma membrane coupled and some un-coupled. The percentage coupled must be small,as the extrapolation number for the inactivationcurves (and the amount of serum-induced stimula-tion seen in the absence of sho) is in the neighbor-
3 The following models have been considered and rejected for the reasons given .
NP{0) =
= e- c ae .No
Model(a) Serum-induced increment due entirely to
covered sites in the membrane which serumuncovers or causes to be uncovered . Coveredsites cannot transport, nor can they bind sho .
(b) Affinity changes of the carrier for adenosineaccount for the serum-induced stimulation .
(c) Serum-induced changes due to unclusteringonly .
(d) Serum-induced changes due to unclustering .Uncoupled sites have a higher affinity foradenosine .
(5)
5 82 THE JOURNAL OF CELL BIOLOGY • VOLUME 60, 1974
N2arcF, 30K
W
20
g
rc 10
aWCW
F00
5
10
15
( 037) -1
FIGURE 8 The relationship between time required toi iactivate 37% of transport and dose of sho (D37) .Closed circles are points taken from Fig . 3 . Open circlesare points taken from five different experiments similarto the one presented in Fig 5 . The X represents D37from Fig 6 . Data from serum-treated monolayers arenot included .
20
Reason for rejecting
No shoulder should be seen on the concentrationvs . log per cent remaining transport or on the
time of exposure to drug at constant concentra-tion vs . log per cent remaining transport.
No shoulder should be apparent, as for a . Alsoexperiments where cells are treated first with shoand then with serum should show increment de-pending on remaining transport, and do not .
No increment should be seen in the ranges whereinactivation is exponential .
Same objection as for c .

hood of 1 .5 . Only one transport site per grouptransports at any time ; the others are inactive buthave the potential ability to transport. There is
precedence for this type of behavior with the en-zyme alkaline phosphatase in E . coli (21) .
However, all sites in a group can bind sho,and when one site is inactivated, another is re-
cruited. Hence, at low concentrations of sho or
at early times, sho will appear to have no or littleeffect on transport . At somewhat higher con-centrations of sho, when all sites save the last inthe group are inactivated, the curve takes on anexponential downward appearance. Serum un-
couples or unclusters sites so that each transportsindependently of the others . Thus, every sho
molecule hits a functioning target so that everyhit is registered as a loss in transport . Therefore,
there is no shoulder on the dose response curve .Experiments dealing with cells which are first
exposed to sho and then to serum can be inter-preted either as rescue of certain sites or as ex-pression of a class of sites which never interactedwith the drug. We do not favor a rescue hypothesisbecause serum should then rescue more carriers,
perhaps by competition-displacement, whenmonolayers treated with sho are then treatedwith very high concentrations of serum (20%),and this does not occur. In support of expressionof sites that never saw drug, there are severalreported instances of activities present at the
surface of cells which are uncovered by enzy-matic treatment (11, 12, 24) . Treatment of rabbitlung macrophages with proteolytic enzymes doesnot alter the rate of adenosine transport (35) .However, some other enzymatic activity of serummay be involved, especially as the substance(s)
in serum responsible for the enhancement ofadenosine transport is (are) nondialyzedableand inactivated by heating at 100°C (32) .
The second class of models predicted by targettheory, the multihit, single target, would entail
the following . As these data have a shoulder when
plotted as log percent control vs . sho or vs . time of
exposure to sho, they can be interpreted (fromthe intercept of the exponential region of thecurves with the ordinate) such that each transportsite requires more than one sho molecule to ir-reversibly inactive it . The intercept on the ordinatewould indicate that more than 1 but less than 1 .5sho molecules are needed per transport carrier
for inactivation . Serum pretreatment would alter
that requirement so that only one sho molecule isrequired . The experiments where cells are first
treated with sho and then with serum would beinterpreted as above, that is, as either rescued ornew sites .
It is impossible from the data at hand to dis-tinguish unequivocally which model is the moreappropriate . However, the ratio of shoulder totransport levels at various times and concen-trations, e .g ., the narrow shoulder, would strongly
suggest the former (9) . Moreover, a tentativechoice in favor of the first can be made based onthe aldehyde experiments . Exposure to low con-centrations of glutaraldehyde (10-50 ,LM) but
not formaldehyde (300 uM) for 10 min reducesthe capacity of these cells to demonstrate serum-induced stimulation of adenosine transport withoutaffecting base transport rates. Moreover, 20 µMglutaraldehyde like 100 HIM (I %) glutaraldehyde
but not 300 µM paraformaldehyde interfereswith the mobility of fluorescein-conjugated con-canavalin A over th ° surface of rabbit lung mac-rophages. Thus, these data would tend to supporta model involving serum-induced movement ofproteins on the cell surface . That the serum-induced changes also occur when the preincuba-
tion temperature is 4 °C does not exclude a multi-step process involving rotation or movementthrough very small distances : macrophage mono-layers are rapidly warmed to 37 °C when transportis tested . Rotation of rhodopsin in frog retinaoccurs within microseconds and is also prevented
by cold or by previous exposure to glutaraldehyde(3, 6) .This model, involving both coupled and hidden
sites, demonstrates a means by which cells canrespond to environmental stimuli rapidly andeffectively . Indeed, other activities that go on atthe plasma membrane such as the accumulation
of potassium in phytohemagglutinin-treated lym-phocytes (28, 37), the insulin-induced stimula-tion of glucose transport (36), and the serum-induced changes in sodium pump density of Helacells (20) might be explained in a similar fashionto those presented here . In line with suggestionsby Holley (16), one might further speculate thatmalignant cells which do not demonstrate con-tact or density-dependent inhibition might bepermanently uncoupled so that transport occurs
maximally at all times. Indeed, concanavalinA binding sites seem to move more freely in the
membranes of transformed as opposed to normal
cells (29) .
I thank Dr . Richard D . Berlin for his valuable sug-
PRYLLIS R . STRAUSS Effects of Serum on Membrane Transport . II 583

gestions and criticisms throughout this work, Dr .
12.Jerry R . Williams for stimulating discussions ontarget theory, and Dr . Phyllis Brown for performingthe high pressure liquid chromatography.
This work was supported by National Institutes of 13 .Health, AM 15361, and National Science Founda-tion, BG-30496X. During its tenure the author wasinitially a fellow of the Massachusetts Heart Associa-tion and then the recipient of National Institutes of
14.Health Fellowship Award GM 5432-01 .
Received for publication 25 July 1973, and in revisedform 12 October 1973.
REFERENCES
15 .
1 . BERLIN, R . D. 1973. Temperature dependence
16.
of nucleoside membrane transport in rabbitalveolar macrophages and polymorphonuclearleucocytes . J. Biol . Chem . 248 :4724 .
2. Brown, P. 1970. Rapid separation of nucleotidesin cell extracts using high pressure liquidchromatography . J. Chromatogr . 52 :257 .
3. BROWN, P. K. 1972 . Rhodopsin rotates in thevisual receptor membrane . Nat . New Biol.236 :35 .
4. CHRISTENSEN, H . N. 1969 . Some special kineticproblems of transport . Adv. Enzymol . Relat .Areas Mol. Biol . 32 :1 .
5 . CHRISTENSEN, H . N ., and J . A. ANTONIOLI. 1969 .Cationic amino acid transport in the rabbitreticulocyte. Na+-dependent inhibition ofNa+-independent transport. J . Biol. Chem .244 :1497 .
6. CONE, R. A. 1972 . Rotational diffusion of rho-dopsin in the visual receptor membrane . Nat .New Biol . 236 :39 .
7. CUNNINGHAM, D ., and A . PARDEE . 1969 . Trans-port changes rapidly initiated by serum addi-tion to "contact-inhibited" 3T3 cells. Proc .Nat! . Acad. Sci. U. S. A . 64:1049 .
8. CURRAN, P. F ., and S . G . SCHULTZ . 1968 . Trans-port across membranes : general principles .in Handbook of Physiology. III . AlimentaryCanal. C. F. Code and W. Heidel, editors .American Physiology Society, Washington,D. C. 1217-1244.
9. ELKIND, M. M., and G . F. WHITMORE. 1967 .The Radiobiology of Cultured Mammalian 23 .Cells . Gordon and Breach, Science Publishers,Inc. New York.
10. FINK, K., R. E. CLINE, and R . M. FINK . 1963 .Paper chromatography of several classes ofcompounds : correlated RI values in a varietyof solvent systems . Anal. Chem . 35 :389 .
11 . FRYE, L . D., and M . EDIDIN . 1970. The rapidintermixing of cell surface antigens after for-mation of mouse-human heterokaryons . J .Cell &i . 7 :319 .
17 .
18 .
19 .
20 .
21 .
22 .
24 .
25 .
5 84
THE JOURNAL OF CELL BIOLOGY . VOLUME 60, 1974
GORDON, S ., and Z . COHN . 1971 . Macrophage-melanoma cell heterokaryons. IV. Unmaskingthe macrophage-specific membrane receptor .J. Exp. Med. 134:947 .
HARE, J . D. 1972 . Studies on the mechanism ofserum stimulation of uridine uptake in serum-less mouse cells. Biochim. Biophys . Acta . 255 :905.
HAWKINS, R . A., and R . D. BERLIN . 1969 . Purinetransport in polymorphonuclear leukocytes .Biochim . Biophys . Acta . 173 :324.
HEINZ, E ., and P. M. WALSH . 1958. Exchangediffusion, transport and intracellular levelsof amino acids in Ehrlich carcinoma cells .J. Biol . Chem . 233 :1488 .
HOLLEY, R. W. 1972 . A unifying hypothesisconcerning the nature of malignant growth .Proc . Natl . Acad. Sci . U. S. A . 69 :2840 .
KOMATSU, Y. 1971 . Mechanism of action ofshowdomycin. IV. Interactions between themechanisms for transport of showdomycinand of various nucleosides in Escherichia coli .Agric . Biol. Chem . 35 :1328 .
KOMATSU, Y., and K. TANAKA. 1968. Mech-anism of action of showdomycin . I . Effect ofshowdomycin on the synthesis of nucleic acidsand proteins in Escherichia coli K-12. Agric.Biol . Chem. 32 :1021 .
KOMATSU, Y., and K. TANAKA. 1971 . Mechanismof action of showdomycin . III . Effect ofshowdomycin on the synthesis of deoxyribonu-cleotides in cell-free extracts of Escherichia coli .Agric . Biol . Chem . 35 :526 .
LAMB, J . F., L . J . BOARDMAN, J. P . NEWTON, andJ . F. AITON . 1973. Effect of calf serum onsodium pump density in HeLa cells . Nat. NewBiol . 242 :115 .
LAZDUNSKI, M ., C. PETITCLERC, D. CHAPPELET,and C. LAZDUNSKI. 1971 . Flip-flop mech-anisms in enzymology . A model : the alkalinephosphatase of Escherichia coli . Eur. J. Bio-chem . 20 :124.
MARTINEAU, R., M. KOHLBACHER, S. SHAW, andH. AMos . 1972. Enhancement of hexose entryinto chick fibroblasts by starvation : differentialeffect on galactose and glucose . Proc. Natl.Acad. Sci . U. S. A . 69:3407 .
NICOLSON, G . 1973 . Temperature-dependentmobility of concanavalin A sites on tumour cellsurfaces . Nat. New Biol . 243 :218 .
NIcoLSON, G. L. 1973. Neuraminidase "un-masking" and failure of trypsin to "unmask"ß-D-galactose sites on erythrocytes, lymphoma,
and normal and virus-transformed fibroblastmembranes . J. Nat! . Cancer Inst. 50:1443 .
NISHIMURA, H ., M. MAYAMA, Y. KOMATSU, H .
KATO, N. SHIMAOKA, and K. TANAKA. 1964.

Showdomycin, a new antibiotic from a Strep-tomyces sp . J. Antibiot . (Tokyo) Ser. A . 17 :148 .
26. PLAGEMANN, P. G . W. 1971 . Nucleoside transportby Novikoff rat hepatoma cells growing insuspension culture . Biochim. Biophys . Acta .233 :688 .
27 . PLAGEMANN, P . G. W., and J . ERBE . 1973. Nu-cleotide pools of Novikoff rat hepatoma cellsgrowing in suspension culture. IV. Nucleosidetransport in cells depleted of nucleotides bytreatment with KCN . J. Cell Physiol . 81 :101 .
28 . QUASTEL, M. R ., and J . G . KAPLAN . 1970 . Earlystimulation of potassium uptake in lympho-cytes stimulated with phytohemagglutinin .Ex p . Cell . Res . 63 :230 .
29 . ROSENBLITH, J. Z ., T . E . UKENA, H. H . YIN, R. D .BERLIN, and M. J . KARNOVSKY . 1973. A com-parative evaluation of the distribution ofconcanavalin A-binding sites on the surfacesof normal, virally-transformed, and protease-treated fibroblasts . Proc . Natl . Acad. Sci . U. S . A .70:1625.
30 . SABATINI, D. D ., K . BENSCH, and R . J . BARRNETT .
1963. The preservation of cellular ultrastruc-ture and enzymatic activity by aldehydefixation . J. Cell Biol. 17 :19.
31 . SINGER, S. J ., and G. N . NICOLSON . 1972. Thefluid mosaic model of the structure of cellmembranes . Science (Wash . D. C .) . 175 :720 .
32 . STRAUSS, P. R ., and R . D . BERLIN . 1973 . Effectsof serum on membrane transport . I . Separationand preliminary characterization of factorswhich depress lysine or stimulate adenosinetransport in rabbit lung macrophages. J. Exp .Med. 137 :359 .
33 . TSAN, M. F., and R. D . BERLIN . 1971a. Mem-brane transport in the rabbit alveolar mac-rophage. The specificity and characteristics ofamino acid transport systems. Biochim . Bio-phys. Acta . 241 :155 .
34 . TSAN, M. F ., and R. D . BERLIN . 1971 b . Effect ofphagocytosis on membrane transport of non-electrolytes . J. Exp . Med. 134:1016.
35 . TSAN, M. F ., R . TAUBE, and R. D . BERLIN . 1973 .The effect of trypsin on membrane transport ofnon-electrolytes . J. Cell Physiol . 81 :251 .
36 . WEISS, L. S ., and H. T. NARAHARA . 1969 . Regula-tion of cell membrane permeability in skeletalmuscle. I . Action of insulin and trypsin on thetransport system for sugar. J. Biol . Chem . 244 :
3084 .37 . WRIGHT, P., M . R . QUASTEL, and J. G. KAPLAN .
1973. Potassium binding sites in lymphocytetransformation and the effect of their pro-longed inactivation by ouabain . In SeventhLeucocyte Culture Conference . F . Daguillard,editor . Academic Press Inc ., New York .
PHYLLIS R . STRAUSS Effects of Serum on Membrane Transport . II 585